國立臺灣大學生物資源暨農學院農業化學所 博士論文
Graduate Institute of Agricultural Chemistry College of Bioresources and Agriculture
National Taiwan University Doctoral Dissertation
大腸桿菌 clpQ
+clpY
+及 gspS
+基因之研究:
基因之調控及其基質辨識
Investigation of clpQ
+clpY
+and gspS
+in Escherichia coli:
gene regulation and substrate recognition
連湘芸
Hsiang-Yun Lien
指導教授:吳蕙芬 博士 Advisor : Whi-Fin Wu, Ph. D.
中華民國九十八年六月
June, 2009
誌謝
從大學、研究所到博士班,在公館學園待了整整十四年,終於要畢業了!論 文能順利完成,要感謝的人真的很多。首先,感謝 吳蕙芬教授對我的悉心指導,
使我這個駑鈍的學生總是感受到老師無限的耐心,以及循循善誘的苦心;而老師 專注嚴謹的實驗態度更是我學習的典範。
口試期間,承蒙 楊盛行教授、陳昭瑩教授、杜宜殷教授、張傳雄教授及張 世宗教授,細心審閱逐字釜正、並提供許多寶貴的意見,使論文內容更趨完備,
學生在此致上最誠摯的謝意。也感謝李佳音教授、賈景山教授、林俊宏教授及林 乃君教授在資格考及預審時,對研究方向及內容給予精闢的見解,使本論文架構 更清楚明白。也謝謝劉瑞芬教授與李佳音教授於實驗期間不吝提供實驗室及儀 器,使論文能順利完成。
此外,感謝所有曾在實驗室一起工作的伙伴們,因為你們使一成不變的生活 變得活潑有趣。也感謝電子大樂隊暨粉絲團的狐群狗黨,你們是讓我心情愉快的 最佳特效藥!
還有我最最親愛的家人,感謝爹娘在精神及金錢上的陪伴與支持,以及弟弟 風雨無阻的接送,讓我能無後顧之憂地完成論文。
最後,謹向這幾年被我殺死以及弄殘的Escherichia coli大軍致敬!
摘要
熱休克蛋白普遍存在於生物體內,作用為幫助生物抵抗環境衝擊。本文針對 大腸桿菌熱休克蛋白ClpYQ (HslUV)的基因調控及基質辨識深入研究。遇到環境 衝擊時,大腸桿菌會發生熱休克反應,此時折疊錯誤的蛋白促使細胞內熱休克基 因表現,這些表現常由調控蛋白sigma factor,如RpoH (或稱σ32) 所調控。本文使 用大腸桿菌熱休克基因clpQ+clpY+的啟動子與報導基因lacZ建構轉錄融合基因 clpQ+::lacZ (op)及轉譯融合基因clpQ+::lacZ (pr),利用噬菌體λRS45將融合基因 clpQ+::lacZ帶入野生株及rpoS -,rpoH -,rpoH -rpoS -突變株後,偵測β-galactosi dase活性以瞭解clpQ+::lacZ的表現。當溫度由30℃升至42℃時,野生株及rpoS - 突變株的β-galactosidase活性上升,但rpoH -及rpoH -rpoS -突變株則未見此現象。
由β-galactosidase 活性分析與北方點墨法分析結果,可知clpQ+::lacZ轉錄的 mRNA訊號強度與β-galactosidase活性成正比,且clpQ+::lacZ與clpQ+clpY+的表現 相似。針對clpQ+clpY+啟動子上σ32 (rpoH) 可辨可辨識識的的保保守守序序列列做C→T點突變,此 突變使融合基因clpQm(c→t)::lacZ無法被σ32活化,β-galactosidase活性下降。經由遺 傳分析結果證實,大腸桿菌clpQ+clpY+的啟動子可被σ32辨識。此外,
clpQ+clpY+operon的五端未轉譯區域 (5’-UTR),亦即轉錄起始處 (transcription start site) 與起始密碼 (start codon) 之間,長度為71 bp,此五端未轉譯區域帶有 一段inverted repeat sequence (IR序列) 5’ CC CCGTAC TTTTGTACGGGG 3’,此IR 序列普遍存在於腸道菌的clpQ+clpY+五端未轉譯區域中。藉由刪除此段IR序列,
並與lacZ融合,分析融合基因clpQm2△40bp::lacZ的β-galactosidase 活性,以及此 段序列缺失對ClpQ與ClpY間交互作用的影響,顯示IR所形成的stem-loop二級結 構在clpQ+clpY+表現時,具有穩定mRNA的效果,本研究為ATP依賴型蛋白酶 (ATP-dependent protease)中,首次發現5’ stem-loop結構具有穩定下游mRNA的功 能。在基質辨識的研究部分,ClpYQ以六元環方式組合,其中ClpY負責基質辨識,
打開基質結構,並傳送至ClpQ進行分解。ClpY可分為三個作用區(domain),
N-terminal domain,I-intermediated domain及C-terminal domain,N domain具有 ATPase的功能,C domain則與self-oligomerization及ClpQ的蛋白酶活性相關。本 文使用酵母菌雙雜交系統,得知ClpY的I domain負責基質辨識,C domain則可與 ClpQ作用,而I domain中的loop L2(175-209 aa)除了與基質結合外,並與後續的基 質傳遞及分解相關。另外,本文亦研究大腸桿菌的酵素glutathionylspermidine synthetase之基因gspS,由於病原蟲Trypanosomatida會造成人類昏睡,發炎及死 亡,而酵素TryS為此種病原蟲所特有,在人類細胞中不存在,所以TryS的研究對 治療Trypanosomatida所造成的疾病極為重要,本文針對大腸桿菌中與TryS構造及 功能皆相似的酵素GspS,其基因表現之調控做一初步的探討。大腸桿菌的gspS 長 1860 bp,產物為glutathionylspermidine synthetase (GspS),共有619個胺基酸,是 一種具有雙重功能的酵素 (bifunctional enzyme),可執行GSH和spermidine之間醯 胺鍵的合成與分解。本研究確認gspS為一單獨的轉錄單位而非以操縱子形式存 在,且GspS起始密碼上游序列具有啟動子功能,而在in vivo情況下,H2O2與BaeR 都可誘導gspS之表現。
關鍵詞:大腸桿菌; 熱休克蛋白; ATP依賴型蛋白酶ClpYQ(HslUV); 啟動子活性;
基因調控; RNA二級結構; mRNA穩定性; 基質辨識; 麩氨基硫─精胺質合成酶。
Abstract
(1) Heat shock responses are typically observed in E. coli. Upon heat shock, protein misfolding leads to a cascade of intracellular protein synthesis, usually dependent on a sigma factor, i.e., σ32, for their gene expression. In this study, the transcriptional (op) or translational (pr) clpQ+::lacZ fusion gene was constructed, with the clpQ+clpY+ promoter fused to a lacZ reporter gene. The clpQ+::lacZ (op or pr) fusion gene was each crossed into lambda phage. The λclpQ+::lac (op), a
transcriptional fusion gene, was used to form lysogens in the wild-type, rpoH - or/and rpoS - mutants. Upon shifting the temperature up from 30 ℃ to 42 ℃, the wild-type λclpQ+::lacZ(op) demonstrates an increased β-galactosidase activity. However, the β-galactosidase activity of clpQ+::lacZ(op) was decreased in the rpoH - and rpoH
-rpoS - mutants but not in the rpoS - mutant. The levels of clpQ+::lacZ mRNA transcripts correlated well to their β-galactosidase activity. Similarly, the expression of the clpQ+::lacZ gene fusion was nearly identical to the clpQ+clpY+ transcript under the in vivo condition. The clpQm(c→t)::lacZ, containing a C to T point mutation in the -10 promoter region for RpoH binding, showed decreased β-galactosidase activity, independent of activation by RpoH. Thus, through a genetic analysis, the clpQ+clpY+ promoter is in vivo recognized by σ32. The transcriptional start point of the
clpQ+clpY+ gene lies 71 bases upstream from the clpQ+ start codon. An untranslated region (UTR) upstream of this mRNA contains a 20 bp inverted repeat (IR) sequence 5’CCCCGTACTTTTGTAC GGGG3’, which is unique for the clpQ+clpY+ operon. In addition, from the wild bacterial genome, the 5’UTR of clpQ+clpY+ also exists in other bacterial species. The clpQ+clpY+ message carries a conserved 71 bp at the 5’
untranslated region (5’UTR) that is predicted to form the stem-loop structure by analysis of its RNA secondary structure. The clpQm2△40bp::lacZ, with a 40 bp
deletion in the 5’UTR, showed a decreased β-galactosidase activity. In addition, from our results, it is suggested that this stem-loop structure is necessary for the stability of the clpQ+clpY+ message. It is noteworthy that this is the first example in the ATP dependent protease to demonstrate that the 5’ stem-loop structure itself participates in the stability of its downstream mRNA. (2) Regarding ClpY substrate recognition study, in the presence of ATP, the ClpYQ complex forms an active protease with an Y6Q6Q6Y6 configuration. ClpY binds, unfolds, and transfers the substrates outside the cylinder into a catalytic core where ClpQ degrades the substrates. The ClpY molecule is divided into three domains: the N-terminal domain, I-intermediate domain and C-terminal domain. The N domain has an ATPase activity, and the C domain is responsible for self-oligomerization of ClpY. Using the yeast two-hybrid system, we show that domain I of ClpY is responsible for recognition of its natural substrates while domain C is necessary for association with ClpQ. The loop 175-209 aa plays a role in substrate tethering. (3) In addition to clpQ+clpY+, gspS+ in Escherichia coli is included in this study. Parasitic Trypanosoma species cause serious tropical diseases such as kala-azar, African sleeping sickness, and Chagas diseases. Trypanothione synthetase (TryS) is a protein unique to Trypanosoma. However, Escherichia coli produce only the metabolic intermediate GspdSH by enzyme GspS, but not
trypanothione. Evolutionary, TryS and GspS share the similarly functional domains.
The gspS of E. coli, encoding a bifunctional enzyme GspS of 619 amino acids, is a gene with 1860 bp. GspS is responsible for the activities of amidase and synthetase between GSH and spermidine.In this study, we showed that gspS in E. coli is an unique transcriptional unit, and the singular promoter was present in the upstream region of the GspS structural gene. In addition, the gspS promoter is in vivo induced by H2O2 and BaeR.
Key Words: Escherichia coli; heat shock; ATP dependent protease ClpYQ(HslUV);
promoter activity; gene regulation; mRNA stability; substrate recognition;
glutathionylspermidine synthetase (GspS).
目錄
誌謝 ... i
摘要 ... ii
Abstract ... iv
目錄 ... vii
表目錄 ... x
圖目錄 ... xi
已發表文章 ... xii
第一章 前言 ... 1
1.1 clpQ+clpY+研究源起 ... 1
1.2 熱休克蛋白 (heat shock proteins) 與 ATP 依賴型蛋白酶 (ATP-dependent protease) 簡介 ... 1
1.3 ClpYQ 蛋白酶之簡介 ... 2
1.3.1 clpQ+clpY+操縱子 (operon) 的發現及命名 ... 2
1.3.2 ClpYQ 為熱休克蛋白的證據 ... 3
1.4 ClpYQ 為 ATP 依賴型蛋白酶之研究 ... 3
1.4.1 ClpY 為 ATPase 之研究 ... 3
1.4.2 ClpY 具 chaperone 功能之研究 ... 4
1.4.3 ClpYQ 蛋白酶為 Threonine 蛋白酶 ... 5
1.4.4 ClpYQ 蛋白酶為 ATP 依賴型蛋白酶 ... 6
1.5 ClpYQ 蛋白複合體的結構 ... 7
1.5.1 ClpQ 及 ClpY 可形成複合體 ... 7
1.5.2 由蛋白分子量推測 ClpYQ 蛋白複合體結構 ... 7
1.5.3 由電顯影像分析 ClpYQ 蛋白複合體結構 ... 8
1.6 ClpYQ 蛋白酶基質辨識區域之相關研究 ... 9
1.7 ClpYQ 蛋白酶基質之研究 ... 10
1.7.1 ClpYQ 蛋白酶可分解異蛋白 (abnormal proteins) ... 10
1.7.2 ClpYQ 蛋白酶可分解 SulA ... 10
1.7.3 ClpYQ 蛋白酶可分解 RcsA ... 12
1.8 clpQ+clpY+實驗目的 ... 12
1.9 gspS+研究源起 ... 13
1.10 GspdSH 簡介 ... 14
1.10.1 GspdSH 組成物質: GSH 及 spermidine ... 14
1.10.2 大腸桿菌中 GspdSH 及其合成酵素 GspS 相關之研究 ... 15
1.11 雙成份控制系統 BaeSR 之研究 ... 17
1.12 gspS+實驗目的 ... 19
第二章 調控蛋白 sigma factors 對 clpQ+clpY+表現的影響 ... 21
2.1 摘要... 21
2.2 材料與方法 ... 21
2.2.1 建立 clpQ+clpY+上游啟動子區域 ( promoter region ) 表現系統 21 2.2.2 β-galactosidase 活性分析 ... 24
2.2.3 北方點墨法 (Northern blotting)... 25
2.2.4 引子延伸實驗 (primer extension) ... 26
2.3 結果... 28
2.3.1 clpQ+::lacZ 轉錄及轉譯融合基因的表現 ... 28
2.3.2 RpoH 對 clpQ+::lacZ 基因表現的影響 ... 28
2.3.3 clpQm1::lacZ 啟動子上的點突變對 clpQm1::lacZ 基因表現的影響31 2.3.4 clpQm(c→t)::lacZ 及 clpQm1::lacZ 啟動子上的點突變不影響轉錄起始 點... 32
2.3.5 clpQ+clpY+ 操縱子 (operon) 的 mRNA 表現與 clpQ+::lacZ 的 β-galactosidase 活性 ... 32
2.3.6 不同蛋白酶缺失對 clpQ+::lacZ 的影響 ... 33
2.4 討論... 34
第三章 clpQ+clpY+ 啟動子五端未轉譯區域 (5’-UTR) 對 ClpQ 表現的影響 ... 36
3.1 摘要... 36
3.2 材料與方法 ... 36
3.2.1 建立 AC3112(cpsB::lacZ)/pBAD33-clpQ/pBAD24-clpY 表現系統 ... 36
3.2.2 AC3112/pBAD33-clpQ/pBAD24-clpY 中,ClpYQ 對兩種基質 RcsA 及SulA 的分解能力測試 ... 38
3.2.3 β-galactosidase 活性分析 ... 39
3.2.4 北方點墨法 (Northern blotting)... 39
3.2.5 mRNA 穩定性測試... 41
3.2.6 西方點墨法 (Western blotting) ... 41
3.2.7 預測 RNA 二級結構所使用的網站 ... 43
3.3 結果... 43
3.3.1 clpQ+clpY+ 啟動子的五端未轉譯區域 (5’-UTR) 對 clpQ+::lacZ 表 現的影響 ... 43
3.3.2 五端未轉譯區域 (5’-UTR) 中的 IR 序列對 clpQ+ clpY+表現的影 響... 44
3.3.3 IR 序列所形成的 stem-loop 結構對 clpQ+ clpY+表現的影響 ... 46
3.4 討論... 47
第四章 ClpY 基質辨識位置之探討 ... 49
4.1 摘要... 49
4.2 材料與方法 ... 49
4.2.1 酵母菌雙雜交系統的菌株、載體及培養基 ... 49
4.2.2 leu2 expression : 生長測試 ... 50
4.2.3 lacZ expression:X-gal 測試 ... 50
4.2.4 lacZ expression:β-galactosidase 活性分析 ... 51
4.2.5 ClpY 及其突變分解基質之偵測 ... 52
4.3 結果... 52
4.3.1 ClpY 基質辨識:ClpY△I+7Gly及ClpY△L1, △L2與SulA 之間沒有交 互作用 ... 52
4.3.2 ClpY 基質辨識: loopL2 上的點突變 ClpYL199Q造成 ClpYL199Q/ClpQ 無法分解基質 ... 53
4.3.3 ClpY 基質辨識:ClpY 的基質辨識與 loopL2 上的疏水性胺基酸密 切相關 ... 53
4.3.4 ClpY 與 ClpQ 之交互作用: ClpY△L1,ClpY△L2,ClpY△L1, △L2及 ClpY△I+7Gly能與ClpQE61C結合,ClpY△c,ClpYX 則否 ... 54
4.4 討論... 55
第五章 大腸桿菌 gspS 基因表現之調控 ... 56
5.1 摘要... 56
5.2 材料與方法 ... 56
5.2.1 建立 gspS 上游啟動子區域表現系統 ... 56
5.2.2 β-galactosidase 活性分析 ... 58
5.2.3 預測啟動子、密碼子及操縱子所使用的網站 ... 58
5.3 結果... 59
5.3.1 gspS 上游序列之探討 ... 59
5.3.2 影響 gspS 表現的因素 ... 64
5.4 討論... 67
參考文獻 ... 69
表目錄
表一、使用於clpQ+clpY+之研究的菌株 ... 76
表二、使用於clpQ+clpY+之研究的載體與噬菌體 ... 77
表三、使用於clpQ+clpY+之研究的引子 ... 78
表四、使用於gspS 之研究的菌株 ... 79
表五、使用於gspS 之研究的載體與噬菌體 ... 80
表六、使用於gspS 之研究的引子 ... 81
表七、操縱子預測結果 ... 82
表八、SG20250/pRS415-gspS-lacZ(op)與 SG20250/λRS45-gspS-lacZ 的 β-galactosidase 活性分析 ... 83
表九、BaeR 對 gspS-lacZ 表現之影響 ... 84
表十、BaeR 與 H2O2對gspS-lacZ 表現的影響 ... 85
圖目錄
圖一、clpQ+ clpY+操縱子的啟動子區域之序列圖示 ... 86
圖二、不同長度的轉錄及轉譯融合基因clpQ+clpY+::lacZ 之活性 ... 87
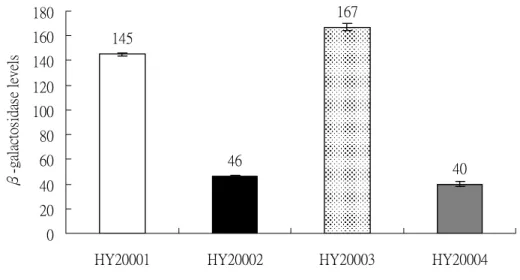
圖三、wt, rpoS -, rpoH -, rpoS –rpoH –四種菌株內,42 ℃對 clpQ+::lacZ 的熱誘導 ... 88
圖四、42 ℃對 clpQ+::lacZ 熱誘導受啟動子點突變(C→T)的影響 ... 89
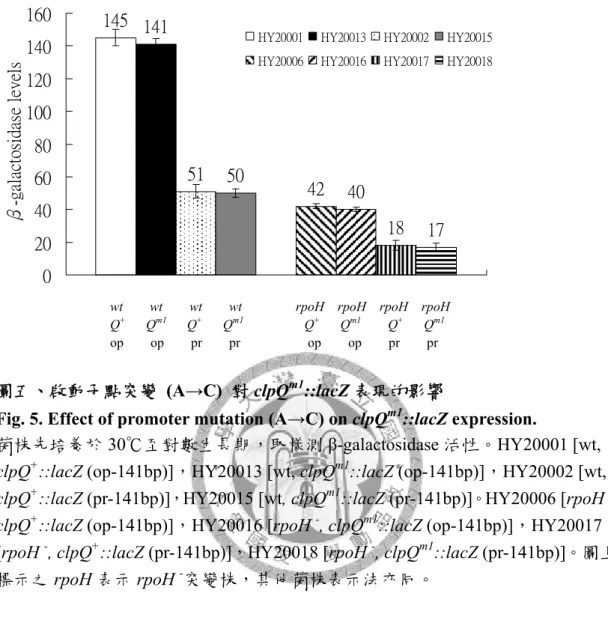
圖五、啟動子點突變 (A→C) 對 clpQm1::lacZ 表現的影響 ... 90
圖六、clpQ+啟動子點突變對轉錄起始點(+1)沒有影響 ... 91
圖七、clpQ+::lacZ 與 clpQ+clpY+在30℃時的 mRNA 表現 ... 92
圖八、蛋白酶缺失對clpQ+::lacZ 表現的影響 ... 93
圖九、去除5’-UTR 對 clpQm2△40bp::lacZ 表現的影響 ... 94
圖十、五端未轉譯區域 (5’-UTR) 中的 IR 序列對 clpQ+ clpY+表現的影 響... 95
圖十一、IR 序列對 ClpQ/ClpY 交互作用之影響 ... 96
圖十二、IR 序列所形成的 stem-loop 結構對 clpQ+ clpY+表現的影響 97 圖十三、不同菌種間clpQ+ clpY+五端未轉譯區域(5’-UTR)保守性序列之 比對... 98
圖十四、ClpY 三個主要作用區示意圖 ... 99
圖十五、ClpY 的 loop L1 與 loop L2 與 SulA 間交互作用之結果。 . 100 圖十六、四組ClpY 點突變 M187I,A188S,L199Q,N205K 與基質結 合及分解基質能力 ... 101
圖十七、三組ClpY 突變 I186N,E193L,E194L,Q198L,Q200L 與基質 結合及分解基質能力 ... 102
圖十八、帶有ClpQE61C/ClpY 融合蛋白的 EGY48/p8op-lacZ,其 Leu2 的表現 ... 103
圖十九、gspS 上游區域之啟動子預測 ... 104
圖二十、gspS 上游區域的編碼區預測 ... 105
圖二十一、啟動子與編碼區預測結果比較 ... 106
圖二十二、編碼區與操縱子預測結果比較 ... 107
圖二十三、gspS-lacZ 融合基因之建構 ... 108
圖二十四、H2O2對gspS-lacZ 表現的影響 ... 109
圖二十五、BaeR 與 H2O2對HY1002(gspS-lacZ)的生長狀況之影響 110 圖二十六、H2O2對HY1002(gspS-lacZ)的生長狀況之影響 ... 111
附圖一、大腸桿菌 ClpYQ 結構圖 ... 112
已發表文章
Lien HY, Yu CH, Liou CM, Wu WF. (2009) Regulation of clpQY (hslVU) Gene Expression in Escherichia coli. Open Microbiol J. 3, 29-39.
Lien HY, Shy RS, Peng SS, Wu YL, Weng YT, Chen HH, Su PC, Ng WF, Chen YC, Chang PY, Wu WF. (2009) Characterization of the Escherichia coli ClpY (HslU) substrate recognition site in the ClpYQ (HslUV) protease using the yeast two-hybrid system. J Bacteriol. 191(13), 4218-31.
第一章 前言
1.1 clpQ+clpY+研究源起
本文針對大腸桿菌熱休克蛋白ClpYQ (HslUV)之基因調控及基質辨識做深 入探討。熱休克蛋白一詞源於果蠅唾腺細胞受熱之研究,將溫度提高於正常生長 溫度,會使細胞快速產生的蛋白質,統稱為熱休克蛋白 (heat shock protein),而 這種現象稱為熱休克反應(heat shock response)。
熱休克蛋白普遍存在於真核與原核細胞內,主要功能為幫助生物抵抗環境衝 擊,細胞在遭遇高溫、重金屬、氧化、缺氧、發炎等環境壓力時,會產熱休克蛋 白,此為細胞因應環境變化以求得生存的反應方式,所以這類蛋白質也稱為壓力 蛋白質(stress protein)。
由於全球溫度逐漸升高,研究熱休克蛋白的基因調控,對於動植物適應高溫 及其他逆境有極高之應用價值,且熱休克蛋白辨識其他蛋白質的功能,目前也已 逐漸應用於癌症治療。本文針對原核生物大腸桿菌熱休克蛋白ClpYQ (HslUV) 之基因調控及基質辨識深入研究,以期增進對熱休克蛋白之瞭解,並有助於未來 在科學及醫學上之應用。
1.2 熱休克蛋白 (heat shock proteins) 與 ATP 依賴型蛋白酶 (ATP-dependent protease) 簡介
當Esherichia coli 受到熱休克衝擊時,細菌會產生一些熱休克蛋白 (heat-shock proteins) 來分解變質的蛋白 (denatured proteins) 或者幫助新合成的蛋白摺疊成正 確的結構。因此,許多熱休克蛋白本身即為chaperones 或蛋白酶。至今,熱休克的 反應受到多種尚未完全了解的機制所調控,其所牽涉之調控基因包括rpoH (Bukau,
1993; Yura et al., 1993)、htrC (Missiakas et al., 1996)等,這些基因轉譯之蛋白所調控 的範圍包含一些ATP 依賴型蛋白酶 (ATP-dependent protease) 如 Lon 及 Clp family 等熱休克蛋白的表現。Clp family 至少被分為 5 個 subfamilies,包括 ClpA、ClpB、
ClpC、ClpX 及 ClpY。其中 ClpA/B/C 擁有兩段具保守性的 ATP binding-sites; ClpX 和ClpY 僅具有一段具保守性的 ATP binding-site。
在E. coli 中,已有許多 ATP 依賴型蛋白酶被發現,這些 ATP 依賴型蛋白酶,
有些存在細胞質中,如Lon 及 Clp 蛋白酶;有些則為膜蛋白,如 FtsH。Lon 蛋 白酶可形成同單元聚合體;FtsH 可形成異單元聚合體,而 Clp 蛋白酶是由兩種 次單元所組成,分子量較小的次單元 (約 19k Da) 為 peptidase,ClpP 及 ClpQ (又 名HslV) ;分子量較大的次單元為 ATPase,計有 ClpA (84 kDa) 、ClpX (46 kDa) 及ClpY (又名 HslU) (49 kDa) 。其中, ClpA 或 ClpX 可和 ClpP 結合形成 ATP 依賴型蛋白酶ClpAP 或 ClpXP;而 ClpY 則和 ClpQ 結合形成另一 ATP 依賴型蛋 白酶ClpYQ。
1.3 ClpYQ 蛋白酶之簡介
1.3.1 clpQ+clpY+操縱子 (operon) 的發現及命名
clpQY/hslVU 基因序列由 Chuang 等人在 E. coli 中以基因選殖的方法發現 (Chuang et al., 1993)。clpQ+clpY+位於E. coli 基因圖譜的 88.9 分鐘處,可同時被 轉錄成mRNA,故為一組操縱子 (operon)。由於 clpQ+clpY+operon 之 mRNA 活 性經由熱誘導後為未熱誘導之1.5 倍,且其上游啟動子 (promoter) 序列與可被 σ32辨識的啟動子保守序列相似,亦與groE 及 dnaK 等熱休克基因 (heat-shock gene) 之啟動子保守序列相符 (Cowing et al., 1985),因此推論 clpQ+clpY+operon 為熱休克基因,故由 “heat shock locus” 的意義而命名為 hslV 及 hslU。若以蛋白 來分類,由於其蛋白層次具下列兩點特徵,因此將之分屬於Clp family : (1)從
胺基酸序列比對的結果發現,ClpQ 與真核生物 20S proteasome 中的 β-type 單體 具相似度,應具蛋白酶的功能,於Clp family 中可扮演蛋白酶的角色; (2)ClpY 在距蛋白N 端 60 個胺基酸有一段可能為 ATP/GTP- binding motif 的序列
“ GPTGVGKT ” ,係與之前被較深入研究的一種 ATP 依賴型蛋白酶
ClpA/ClpX-ClpP complex 中之 ATPase ClpA 相同,此外,ClpY 亦與 ClpX 的胺基 酸序列有50% 的相似度,故 ClpY 應屬於 Clp family 中負責基質辨識及 ATPase 的角色。由於此組操縱子於胺基酸比對上類似於Clp family,因此,又被命名為 ClpYQ (Gottesman et al., 1993)。
1.3.2 ClpYQ 為熱休克蛋白的證據
σ32為一些熱休克蛋白於轉錄階段時之調控蛋白,而clpQ+clpY+ operon 上游具 有一段由σ32辨識的啟動子序列。Chuang 等人經由 primer extention 確認此段序 列為clpQ+clpY+operon 之啟動子(Chuang et al., 1993)。 Rohrwild 等人將 σ32選殖 於具有lac promoter fusion 的質體上,而後以 IPTG 誘導 σ32大量表現,發現ClpYQ 蛋白的表現量增加約10 倍,且於 ATP 存在時,水解 peptide Z-Gly-Gly-Leu-AMC 之能力亦提高約10 倍 (Rohrwild et al., 1996)。以上之證據均暗示 clpQ+Y+受σ32 之調控,而ClpYQ 可能為熱休克蛋白。
1.4 ClpYQ 為 ATP 依賴型蛋白酶之研究
1.4.1 ClpY 為 ATPase 之研究
Missiakas 等人針對 ClpY 中假設性的 ATP-binding site 作研究。實驗係將假 設性的ATP-binding site 中之 Gly59 置換成 Ala;或將 Gly61 置換成 Ala。結果發 現 : (1)免疫沈澱的結果顯示,經置換過的 ClpY 蛋白無法如野生種的 ClpY 蛋白一般,可與ClpQ 蛋白形成複合體而共同沈澱出來; (2)帶有 clpQ+ clpY59
的多倍數質體,並不能如帶clpQ+ clpY+的多倍數質體一般可使熱休克反應下降;
(3)經純化所得的 ClpY59 及 ClpY61 失去胞外水解 ATP 的能力。由結果推論,
存在於E. coli 中之 ClpY 的 ATP-binding site “ GPTGVGKT ” 應與 ClpY 水解 ATP 的活性,及與ClpY 和 ClpQ 蛋白形成複合體有關 (Missiakas et al., 1996)。
Shin 等人亦針對 ClpY 的 ATP-binding site 作研究。利用定點突變的方式,將 ATP-binding site 中之 Lys63 置換成 Thr,而重組出 HslU/K63T 蛋白,並針對 HslU/K63T 與 HslU 蛋白作 ATP 水解能力及特性上的比較 (Shin et al., 1996)。研 究結果發現 : (1)HslU/K63T 失去了水解 ATP 的功能; (2)HslU/K63T 無法 在ATP 存在下協同 ClpQ 分解基質 Z-Gly-Gly-Leu-AMC; (3)HslU/K63T 在純 化過程中,無論ATP 存在與否,均以單體 (50 kDa) 存在,而 HslU 蛋白於 ATP 不存在時以雙體之形式出現 (100 kDa),於 ATP 存在時,則為分子量介於 350 至 450 kDa 之複合體; (4)HslU/K63T 較 HslU (ClpY) 蛋白不穩定,容易在純化 過程中被分解成許多小片段蛋白。HslU/K63T 無論 ATP 或 ADP 是否存在,都極 易被Trypsin 分解,但是 HslU (ClpY) 於 ATP 或 ADP 存在時,可受其保護,因 而不易被Trypsin 分解。由以上的結果推論,位於 ATP-binding site 的 Lys63 對 ClpY 蛋白扮演著十分重要的角色,而 ATP-binding site 至少具有三個重要性 :
(1)水解 ATP; (2)形成 ClpY 複合體的結構; (3)ClpY 與 ATP 結合,可 增加ClpY 的穩定性。
1.4.2 ClpY 具 chaperone 功能之研究
Seong 等人將純化所得的 ClpY (HslU) 和 MBP-SulA 及 FXa 一起置於 Tris-HCl 緩衝液中,測其混濁度,發現無論 ATP 存在與否,ClpY (HslU) 蛋白的 存在可降低混濁度,即ClpY (HslU) 可以阻止 SulA 的聚集 (Seong et al., 2000)。
此外,實驗結果亦發現E. coli. hsl 突變菌株的生長較野生型好,此乃由於在 hsl 突變菌株中缺少ClpY (HslU) 蛋白,故有部分 SulA 蛋白聚集失去活性,因此細 胞生長較有ClpY (HslU) 存在而能降低 SulA 蛋白聚集的野生型細胞好。以上的
實驗結果顯示無論於胞內或胞外,ClpY (HslU) 皆具有 molecular chaperone 的功 能,能降低細胞分裂抑制物SulA 之聚集。Seong 等人亦證實 ClpY 的
oligomerization,而非 ATP 水解,為 ClpY 具有 molecular chaperone 功能所必須 (Seong et al., 2000)。
1.4.3 ClpYQ 蛋白酶為 Threonine 蛋白酶
Rohrwild 等人利用各種蛋白酶的抑制物,進行 ClpYQ 蛋白酶活性抑制實驗,
以分析ClpYQ 蛋白酶之特性。實驗結果發現,ClpYQ 蛋白酶分解 peptide Z-Gly-Gly-Leu-AMC 之能力會受到多種可抑制哺乳類 proteasome 的抑制物之抑 制 (Rohrwild et al., 1996)。這些抑制物包括 MG132 及 calpain inhibitor I、3, 4-dichloroisocoumarin 及 lactacystin,其中 3,4-dichloroisocoumarin 及 lactacystin 會修飾哺乳類及古細菌 (archaebacteria) 之 proteasome 中 N 端的 Threonine。
Rohrwild 等人將純化所得之 ClpQ 蛋白進行 N 端定序,發現 N 端有兩個 Thr 之殘 基,推論此兩個Thr 應與其蛋白酶活性有關。
由於ClpQ 蛋白與真核生物中 20S proteasome 的 β-type subunit 蛋白具有同源 性,且N 端均有兩個連續的 Thr 殘基。 Seemuller 等人對 Thermoplasma
acidophilum 中 20S proteasome 之 β-type subunit 的研究報告,推測 ClpYQ 蛋白酶 屬於Threonine 蛋白酶,且 ClpQ 蛋白中的 Thr1 及 Thr2 與蛋白酶活性有關 (Seemuller et al., 1995)。因此,Missiakas 等人針對 Thr1 及 Thr2 對 ClpQ 的活性 影響作研究,分別將ClpQ 的 Thr1 及 Thr2 置換成 Ala,而得到 HslV1 及 HslV2 兩重組蛋白,結果顯示HslV2 完全失去其蛋白酶 活性,並且於 clpQY 基因缺失 的突變種中大量表現hslV2 hslU 基因時,將無法像大量表現正常 hslV hslU 基因 時,可抑制puromycin,hslV2 hslU::Ω 亦無法像 hslV hslU::Ω 突變株可形成較小 的菌落,這顯示Thr2 與 ClpQ 的活性及 ClpYQ 蛋白酶的功能有關,ClpYQ 應屬 於Threonine 蛋白酶 (Missiakas et al., 1996)。
1.4.4 ClpYQ 蛋白酶為 ATP 依賴型蛋白酶
Rohrwild 等人發現 ClpQ 抗體可降低 ClpYQ 蛋白酶的活性,且活性降低程 度和ClpQ 抗體濃度成正比關係,然而,利用 ClpY、ClpP、Lon 等抗體或是 preimmune serum 不影響 ClpYQ 蛋白酶的活性,由此推論 ClpQ 為 ClpYQ 蛋白酶 水解活性所必須。此外,亦發現純化之ClpYQ 蛋白僅於 ATP 存在時可分解 peptide Z-Gly-Gly-Leu-AMC,而 CTP、GTP、UTP、ADP、AMP 以及 ATP 的類似物 (analogue) AMP-PNP 皆無法取代 ATP 使 ClpYQ 蛋白酶分解 peptide,且當 ATP 存在時,水解Z-Gly-Gly-Leu-AMC 的活性提高 150 倍,因而推論 ATP 水解為 ClpYQ 蛋白酶活性所必須 (Rohrwild et al., 1996)。1996 年,Missiakas 等人亦發 現在ClpY 及 ATP 存在下,會大大增加 ClpQ peptidase 的水解活性 (Missiakas et al., 1996; Rohrwild et al., 1996)。
1996 年 Yoo 等人亦得到相同的實驗結果 (Yoo et al., 1996)。Yoo 等人以多倍 數質體大量表現ClpYQ 以純化其蛋白,並對 ClpYQ 蛋白酶作特性分析。實驗結 果發現: (1)利用膠體過濾法 (gel filtration) 無論 ATP 是否存在的情況下,均可 純化得到250 kDa 的 ClpQ 複合體,推測由 12~14 個 ClpQ 單體組成; (2)在 ATP 存在的情況下可得到450 kDa 的 ClpY 複合體 (8~10 個單體),但 ATP 不存在時 則僅可得到100 kDa 的 ClpY 複合體 (2 個單體),這顯示 ATP 的存在與 ClpY 形 成複合體有關; (3)ClpQ 單獨存在時其水解活性相當低;當 ATP 及 ClpY 同時存 在時,ClpQ 水解 peptide Z- Gly-Gly-Leu-AMC 之能力劇烈提高,這顯示細胞中 ClpY 和 ClpQ 需在 ATP 的存在下協同作用; (4)ClpY 可水解 ATP,是一種 ATPase,而 ClpQ 存在時會刺激 ATPase 的活性達兩到四倍; (5)在 ATP 存在 下,ClpQ 可水解 peptide Z-Gly- Gly-Leu-AMC,於 dATP、CTP、α, β-methylene ATP 存在時,僅有15~40%的 peptide 被分解,此因其水解速度為 ATP 水解速度的 15
~35%,由於 GTP 及 UTP 不被水解,故無法刺激 ClpQ 水解 peptide,而 ADP 及 AMP 並非 ClpY 之基質,因而無法刺激 ClpQ 水解 peptide。無法被水解的 ATP
之類似物β,γ-methylene ATP 亦無法取代 ATP,促使 ClpYQ 蛋白酶分解 peptide,
此結果顯示ATP 水解為 ClpQY 蛋白酶活性所必須。綜合上述結果可知,細胞中 ClpY 和 ClpQ 需在 ATP 存在下協同作用,故 ClpYQ 蛋白酶為一 ATP 依賴型蛋白 酶。
1.5 ClpYQ 蛋白複合體的結構
1.5.1 ClpQ 及 ClpY 可形成複合體
Rohrwild 等人在缺乏 Lon 及 ClpP 兩個 ATP 依賴型蛋白酶之菌株中,將 σ32 選殖於具有lac promoter fusion 的質體上,而後以 IPTG 誘導而大量表現 σ32,進 而誘導ClpYQ 蛋白的表現 (Rohrwild et al., 1996)。於純化 ClpYQ 蛋白的過程中 可得到分子量約650~700 kDa 的部分,此結果顯示 ClpQ 及 ClpY 蛋白可能自己 單獨或彼此相互結合形成複合物;亦可能有其他蛋白共同參與形成複合物。此 外,在同時大量存在ClpQ 及 ClpY 蛋白的情形下,可偵測出絕大部分水解 Z-Gly-Gly-Leu-AMC peptide 的活性。Rohrwild 等人更進一步利用 ClpQ 抗體,將 ClpQ 及 ClpY 蛋白共同沈澱出來,直接證明 ClpQ 及 ClpY 蛋白能形成具有水解 活性之蛋白酶複合體。
1.5.2 由蛋白分子量推測 ClpYQ 蛋白複合體結構
Rohrwild 等人於 ATP 存在時,純化藉由誘導 σ32而增加ClpYQ 表現之菌體 中的ClpYQ 蛋白,可流洗出一分子量介於 650 kDa 至 750 kDa 的蛋白,因 SDS-PAGE 偵測出這些具有活性的部分,至少含有 15 至 20 種其他的蛋白,
Rohrwild 等人推測 ClpYQ 蛋白酶可能由 ClpQ 或 ClpY 蛋白彼此相互結合而形 成,亦可能有其他蛋白參與複合體之形成,但於該菌體中無法純化出更純且帶有 水解活性的ClpYQ 複合體。於是,Rohrwild 等人進一步純化於多倍數質體上大
量表現的ClpYQ 蛋白,結果發現 ClpYQ 可同時複合出現在分子量約 650-700 kDa 的部分,SDS-PAGE 僅偵測出兩個主要的蛋白,經西方雜交法確認為 ClpQ 及 ClpY 兩蛋白,其中大部分的ClpQ 蛋白出現於 280~320 kDa 的部分,由於 ClpQ 的單 體為19k Da,故推測此複合體約含 14-16 個 ClpQ 單體;ClpY 大部分出現於 480
~520 kDa 的部分,因 ClpY 單體的分子量為 50 kDa,故推測複合體至少含 10 個ClpY 單體。
Yoo 等人則不論在 ATP 存在與否,皆可純化出分子量約 250 kDa 之 ClpQ 複 合體,約含12~14 個 ClpQ 單體 (Yoo et al., 1996)。於 ATP 不存在時,純化得到 分子量為100 kDa 之 ClpY,應為 ClpY 之雙體;但在 ATP 存在時,可得到分子 量為450 kDa 之複合體,即包含 8~10 個 ClpY 單體。據此,Yoo 等人推測 ATP 在ClpY 形成複合體之過程中扮演重要的角色。
Kessel 等人純化由帶有 clpQ+clpY+基因之多倍數質體所大量表現的ClpQ 和 ClpY 蛋白時,亦與 Rohrwild 等人的研究有相似的結果 (Kessel et al., 1996)。Kessel 等人在ATP 存在的條件下,經由 Superose 膠體過濾純化之過程,分離出分子量 約220 kDa 的 ClpQ 複合體 (約由 12 個單體組成) 及分子量約 350 kDa 的 ClpY 複合體 (約由 7 個單體組成) ,以及分子量大於 600 kDa 的 ClpQ 和 ClpY 所組成 的ClpYQ 複合體 (推論含有 12 個 ClpQ 單體以及至少一組至兩組的 ClpY 六元 體) 。
1.5.3 由電顯影像分析 ClpYQ 蛋白複合體結構
Kessel 等人將純化之蛋白經負染處理後,以電子顯微鏡觀察,發現 ClpQ 的 複合體是由六個次單體形成的環狀結構,直徑約10~11 nm;ClpY 的複合體於 MgATP 存在時,在電子顯微鏡下呈現圓形,亦由六個次單體形成環狀的結構,
較ClpQ 略大,直徑約 13 nm。
Rohrwild 等人利用 TEM (transmission electron microscopy) 、STEM (scanning
transmission electron microscopy) 以及負染影像處理 (image processing of
negatively staining) 的方法,對純化出的 ClpQ、ClpY 及 ClpYQ 蛋白複合體進行 結構分析 (Rohrwild et al., 1997)。結果發現,ClpQ 複合體為兩個環狀結構所組 成,每個環狀結構由六個ClpQ 單體組成。在 ATP、ADP 及無水解能力之 ATP 類似物AMP-PNP 或是 ATP-γ-S 等核甘酸存在時,ClpY 可形成環狀複合物,直 徑約14 nm;但無上述核甘酸存在時,則無法看見該環狀複合物。因此,Rohrwild 等人推測ATP 之結合而非 ATP 水解為 ClpY 形成複合體所必須。分析負染處理 較佳之ClpY 複合體,發現 ClpY 複合體之歧異度較 ClpQ 複合體為大,經對稱及 數位分析,發現有24%之 ClpY 複合體呈現六元對稱,36%呈現七元對稱,但有 40%之 ClpY 複合體無法歸入上述兩對稱形式中。進一步分析 ClpY 複合體之結 構,不論是利用STEM 測量質量之分析,或利用 BS3(bis(sulfosuccinimidyl) suberate) 進行crosslinker 之實驗,皆可確認 ClpY 可由六或七個單體形成環狀複合結構。
在AMP-PNP 存在時,電子顯微鏡下可觀察到穩定存在之 ClpYQ 之複合體,該 複合體由四個平行排列的環狀構造形成一圓柱體,其中由ClpQ 複合體的兩個環 狀結構組成蛋白酶的催化中心, ClpY 複合體的環狀結構則平貼於 ClpQ 兩個環 狀複合體之外側,形成一YQQY 平行環狀圓柱狀構造。
1.6 ClpYQ 蛋白酶基質辨識區域之相關研究
於1997 及 1999 年時分別由 Levchenko 和 Smith 等人提出 Clp family 蛋白酶 的蛋白C 端具有同源性的基質辨識區域。Levchenko 將此蛋白 C 端的基質辨識區 域命名為PDZ-like domains,初始研究 PDZ domains 與許多真核生物中的 signaling proteins 有關,其利用 C 端區域與其他協同的蛋白產生蛋白間聯結的作用。
Levchenko 推論於 Clp family 中此段 PDZ-like domains 可能與基質辨識的機制有 關 (Levchenko et al., 1997)。Smith 等人提出同為 ATP-dependent protease 的 Lon
以及Clp family protease 及 chaperones 於蛋白 C 端具有同源性的基質辨識區域,
並將之命名為SSD (sensor- or substrate-discrimination) domains (Smith et al., 1999),研究結果推論 Lon 以及 Clp family protease 使用相同的基質辨識機制,推 測這些ATP 依賴型蛋白酶於細菌中係利用此段專一性之區域去辨識基質的某段 區域,然而詳細的辨識機制或基質被辨識之區域的資料則尚無研究報告。
1.7 ClpYQ 蛋白酶基質之研究
1.7.1 ClpYQ 蛋白酶可分解異蛋白 (abnormal proteins)
當E. coli 暴露於高溫時會誘導熱休克蛋白(heat shock proteins)大量生成,
這是由於不穩定的σ32漸趨穩定,而指揮RNA 聚合酶轉錄熱休克基因所致。利 用添加胺基酸類似物或是嘌呤黴素 (puromycin) 而生成異蛋白或外來蛋白的大 量累積,皆會誘導熱休克蛋白的生成,致使菌體內產生類似熱休克反應之現象。
Missiaka 等人發現,過量表現的 clpQ+clpY+ operon 可抵抗造成 E.coli 累積有害蛋 白質的毒素如puromycin。以上結果顯示,ClpYQ 在 E. coli 中扮演分解異蛋白的 角色 (Missiakas et al., 1996)。
Kanemori 等人測量 σ32的半衰期 (Kanemori et al., 1997),結果顯示在 lon – clpP –X – hslV –U -突變株中,σ32半衰期增長,而在Lon 與 ClpXP 蛋白酶缺乏的菌 體中,利用質體大量表現ClpQY 蛋白酶,會降低 σ32的表現,此結果說明,這些 ATP 依賴型蛋白酶之間,彼此偕同作用,藉由分解 σ32及異蛋白來調控細胞內熱 休克反應。
1.7.2 ClpYQ 蛋白酶可分解 SulA
Misuzawa 及 Gottesman 等人於 1983 及 1991 年發表,在 lon-突變株中,施以 MMS (methyl methanesulfonate) 或 UV,因缺乏 Lon protease 使細胞分裂抑制物
SulA 無法被分解,導致細菌死亡,證實 SulA 可被 E. coli 蛋白酶 Lon 所分解,故 SulA 為 Lon 的基質之一(Gottesman & Stout, 1991; Mizusawa & Gottesman, 1983)。
SulA 是由 sfiA 所轉譯出的蛋白質,可於 UV 照射後或以 Nitrofurantion 處理 而被誘導表現,SulA 於細胞中會和 FtsZ 形成複合物,而使 FtsZ 失去蛋白活性因 而抑制細胞分裂,故 SulA 為細胞分裂抑制物。ClpYQ 蛋白酶可分解 SulA 始於 Khattar 等人發表的報告 (Khattar, 1997),Khattar 等人篩選到一質體可在 lon-突變 株中抑制SOS 反應 (SOS response),進一步分析該質體所帶的基因序列,確認能 抑制SOS 反應是由於 clpQ+clpY+ operon 大量表現所致。
Kanemori 等人利用免疫印漬法 (immunoblotting) 發現在△(clpPX-lon)菌株 中增加clpQY 基因突變,可偵測到原本無法偵測的 SulA 蛋白 (Kanemori et al., 1999),而在缺乏 Lon 及 Clp 蛋白酶活性時,誘導 SulA 表現,發現在△clpQY 突 變株中,SulA 的累積遠比 clpQ+菌株快,此外,SulA 的半衰期在△clpQY 突變株 中較長,在野生株clpQ+中較短。胞外實驗亦證實,ATPase 存在時,ClpQ 及 ClpY 蛋白需同時存在才能分解 SulA。 根據以上結果,Kanemori 等人認為,ClpYQ 與Lon 具有相同功能,可參與細胞中 SulA 的分解,且當細胞中 Lon 及 Clp 蛋白 酶活性低於某一程度時,ClpYQ 為生長所必須。
同年,Wu 等人藉由在 lon -突變株中,大量表現ClpYQ,使細菌在 MMS 當 中亦能生存,不致受SulA 累積的影響而死亡 (Wu et al., 1999);且在 lon -突變株 中,大量表現ClpYQ,SulA 也可以被分解;另外,與 lon -突變株相比,SulA 在 lon -clpQ -,lon -clpY -突變株中都比較穩定,半生期變長,此結果再度證明SulA 為ClpYQ 的基質。
由於ClpY 為一 ATPase,在 ClpYQ chaperone 中,負責基質辨識的角色,Lee 等人以yeast two-hybrid system 方式證實,ClpY 與 SulA 之間有蛋白質交互作用
(protein-protein interaction),除說明SulA 為 ClpYQ 的基質之外,更進一步證明 ClpY 與 SulA 在 in vivo 中可互相作用 (Lee et al., 2003b)。
1.7.3 ClpYQ 蛋白酶可分解 RcsA
RcsA 為 capsule synthesis 的基因 cpsB 之活化物,Gottesman 等人確認 RcsA 為蛋白酶Lon 的基質, lon -突變使細菌產生mucoid 的表現型 (Gottesman & Stout, 1991)。
Wu 等人首度提出 RcsA 為 ClpYQ 蛋白酶基質之初步觀察 (Wu et al., 1999),
利用可受RcsA 調控之 cpsB::lacZ 報告基因 (reporter gene),藉由 cpsB::lacZ 表現 β-galactosidase 的活性,來觀察 ClpYQ 蛋白酶對 RcsA 之影響。實驗結果顯示由 多倍數質體大量表現的ClpYQ 蛋白酶會使得 cpsB::lacZ 表現下降為對照組的 20 倍。據此,推論ClpYQ 蛋白酶可分解 RcsA。
Kuo 等人亦針對 RcsA 為 ClpYQ 的基質做更進一步的研究 (Kuo et al., 2004),利用在 lon -突變株中,大量表現ClpYQ,可抑制因 lon -而產生的mucoid 表現型,以及在lon –clpQ –clpY -突變株中,cpsB::lacZ 之 β-galactosidase 活性比 lon –clpQ +clpY +高,證實ClpYQ 可藉由分解 RcsA,調控 cpsB::lacZ 的表現。
1.8 clpQ+clpY+實驗目的
本論文針對clpQ+clpY+operon 在菌體內 (in vivo) 的調控及 ClpY 的基質辨識 功能做探討。利用clpQ+clpY+ 啟動子與報導基因 lacZ 所建構之融合基因
clpQ::lacZ,證明 σ32可直接調控clpQ+clpY+啟動子。
在clpQ+clpY+operon 的五端未轉譯區域 (5’-UTR),亦即轉錄起始處
(transcription start site) 與轉譯起始處 (translation start site) 之間,有一段 inverted
repeat sequence (IR),藉由觀察此段序列缺失對 ClpQ 與 ClpY 間交互作用的影響,
可知IR 所形成的 stem-loop 二級結構為 clpQ+clpY+operon 表現時所必須。
前人研究可知ClpYQ 以六元環方式組合(Kessel et al., 1996; Missiakas et al., 1996; Rohrwild et al., 1996),其中 ClpY 負責基質辨識,打開基質結構,並傳送至 ClpQ 進行分解(Gottesman, 2003),已知的 ClpYQ 基質有 SulA 及 RcsA 等,SulA 為細胞分裂抑制物,若在菌內累積則細菌無法生長,RcsA 為活化物,可活化形 成capsule 的基因 cpsB。
ClpY 可分為三個作用區(domain),N-terminal domain,I-intermediated domain 及C-terminal domain,N domain 具有 ATPase 的功能,C domain 則與
self-oligomerization 及 ClpQ 的蛋白酶活性相關。由 X-ray 結構分析,認為 I domain 能與ClpQ 結合,並負責與基質互相作用(Bochtler et al., 2000)。
ClpY 的 I domain 中,有兩個 loop 結構,分別為 loop L1(137-150 aa)與 loop L2(175-209 aa),已知將 loop L2 移除後,ClpY△(175-209 aa)在ClpQ 存在時仍然無法 分解SulA (in vitro),但其分子機制未明(Song et al., 2000)。利用酵母菌雙雜交系 統(yeast two-hybrid system)分析,可知 ClpY 與 SulA 之間有蛋白質交互作用(Lee et al., 2003a),本文亦藉由酵母菌雙雜交系統,進一步得知 ClpY 的 I domain 負責基 質辨識,C domain 則可與 ClpQ 作用,而 I domain 中的 loop L2(175-209 aa)除了 與基質結合外,並與後續的基質傳遞及分解相關。
1.9 gspS+研究源起
Trypanosomatida 包含 Trypanosoma,Leishmania,Crithidia 等,此類致病性 原蟲,可藉由蚊蟲叮咬,進入血液,中樞神經及淋巴,在非洲及南美,此類原蟲 會引起昏睡,發炎等病症,使許多人死亡。目前已從Trypanosoma 分離出 TSH [trypanothione,N1, N8-bis(glutathionyl)spermidine] (Fairlamb et al., 1985; Fairlamb et al., 1986)。在 Trypanosoma,TSH 取代 glutathione (GSH) 在一般物種(如人類
及E. coli)中維持細胞氧化還原平衡的角色,在人類或 E. coli,藉由還原態的 GSH 使 Glutaredoxin (Gxn) 還原,進而調整其他物質的氧還,但在 Trypanosoma,
則用TSH/ Tryparedoxin 取代 GSH/ Gxn 的作用。例如,Peroxynitrite (ONOO-) 是 巨噬細胞用來對抗致病菌的重要物質,在培養Trypanosoma 時,加入
peroxynitrite,會耗損病原蟲的 TSH;不但如此,在 Trypanosoma 中,GSH 只和 含硫的蛋白質有作用,大部分氧化物的代謝,都是透過TSH (Thomson et al., 2003)。因此與 TSH 相關的酵素,近年來吸引許多人的探討,希望對藥物發展有 所幫助。TSH 的合成,是由 GSH 與 spermidine 間的醯胺鍵結 (amide bond),先 合成GspdSH (glutathionylspermidine),再由 GspdSH 與 GSH 間醯胺鍵生成,產 生TSH。 Trypanosoma 用酵素 TryS (Trypanothione synthetase) 來進行 GSH 與 GspdSH 間醯胺鍵結生成,產生 TSH,此病原蟲缺乏 GspS (glutathionylspermidine synthetase) 的基因, GSH 與 spermidine 間醯胺鍵生成,產生 GspdSH 也是靠 TryS。另一致病原蟲 Leishmania,GspdSH 與 TSH 的合成,和 Trypanosoma 相同,
都是靠TryS 作用,但 Leishmania 有一退化的 GspS 基因 (gspS pseudogene)。此 外,在Crithidia,GspdSH 的合成可用 GspS 或 TryS,TSH 的合成則靠 TryS,但 至目前為止,只知E. coli 以 GspS 合成 GspdSH,尚未在 E. coli 測得 TSH 存在。
TryS 與 GspS 的性質類似,都有合成酶 (synthetase) 和醯胺酶 (amidase) 活性,
兩者的胺基酸序列亦相似 (Ariyanayagam et al., 2005),另外,根據親源分析 (phylogenetic analysis) 結果,E. coli 的 GspS 與 Crithidia 的 GspS 及 Leishmania 的GspS 非常相似,位於樹狀圖中相鄰的位置,GspS 與 TryS 相似度也很高,兩 者在樹狀圖中的位置也很相近 (Comini et al., 2005)。藉由研究 E. coli 基因 gspS,可增進對 tryS 的瞭解也有益於 Trypanosoma 的研究。
1.10 GspdSH 簡介
1.10.1 GspdSH 組成物質: GSH 及 spermidine
GSH 是一種 tripeptide,由 L-γ-glutamyl-L-cysteinyl-glycine 組成,在好氧物 種 (aerobic species) 中是最主要的低分子量硫醇 (low molecular weight thiol),細 胞內濃度約為0.1 至 10 mM。細胞內大部分 GSH 維持在還原態 (reduced ,thiol form),亦即 GSH,其他則以 GS-S-protein、GSSG、thioester 形式存在 (Pai et al., 2006)。藉由將大多數 GSH 維持在還原態,GSH 可維持細胞內氧化還原狀態的平 衡,還原態GSH 可將 H2O2或其他過氧化物還原,本身則變成氧化態的glutathione disulphide (GSSG),GSSG 再經由 glutathione reductase 及 NADPH 的作用,還原 成GSH。
Spermidine [C9H17N3,N-(3-aminopropyl)butane-1,4-diamine] 是細菌細胞中主 要的多胺 (polyamine)。Spermidine 在細菌細胞中作用廣泛,可和磷酯質結合,
達到穩定細胞膜或其他膜結構的效果,或調節膜通透性。另外,多胺類物質亦與 細胞生長及抗氧化有關,其方式為多胺可刺激調控蛋白的生成 (Raj et al., 2001;
Tkachenko & Nesterova, 2003),進而影響許多與生長或抗氧化相關基因的表現,
如多胺可刺激RpoS、OxyR、FecI、Fis 等調控蛋白的生成,OxyR 可誘導抗氧化 相關基因的表現,RpoS 則與生長遲滯期 (stationary phase) 的許多基因表現有關 (Jung & Kim, 2003)。
1.10.2 大腸桿菌中 GspdSH 及其合成酵素 GspS 相關之研究
GSH 的 glycine carboxylate 與 spermidine 之間,可用醯胺鍵結,形成 GspdSH。
在一般培養條件下,E. coli 生長至指數生長期 (log phase) 或遲滯期 (stationary phase),菌體內都無法測得 GspdSH。但若將 E. coli 以 M9 minimal medium 搖動 通氣培養 (shaker) 到遲滯期時停止搖動,可在菌體中偵測到 GspdSH 的存在 (Tabor & Tabor, 1975),Tabor 等人偵測此條件下 (厭氧/生長遲滯期),二十分鐘內 細胞中GspdSH 和 spermidine 濃度不變,但 GspdSH 卻一直進行生成和分解,並 且發現,在指數生長期時,spermidine 沒有顯著代謝,但進入遲滯期後,大部分 的spermidine 改以 GspdSH 形式存在。若將遲滯期的 E. coli 培養液用新鮮培養 基稀釋,會使GspdSH 快速分解成 GSH 及 spermidine,據此推測使 GspdSH 在生
長遲滯期累積的原因可能有二,一是GspdSH 可能在所有生長期皆可生成,但在 指數生長期時,GspdSH 的分解速度快,另一可能性是遲滯期細胞內的酸鹼值或 氧化還原狀態改變,造成某種酵素活性的改變,使GspdSH 累積(此時期尚未發 現合成GspdSH 的酵素 GspS)。De Craecker 等人觀察 E. coli 所有生長期中 GSH 與GspdSH 含量變化 (De Craecker et al., 1997),在 aerobic/ M9 minimal medium 狀態下,整個生長期中的GSH 含量穩定,指數生長期中期 (mid-log phase) 會生 成微量GspdSH,約佔 GSH 總量的 3%,到了遲滯期,GspdSH 生成量增加,佔 GSH 總量的 11%。在 aerobic/M9 minimal medium 狀態下培養 E. coli 至指數期,
再換到厭氧狀態 (anaerobic) 培養 6 小時,測得 GspdSH 逐漸增加,最後 80%以 上的GSH 轉變成 GspdSH;但在全程中都沒有測到 N1, N8-bis (glutathionyl) spermidine,即 TSH 的存在,此外,在 glutathione reductase 中加入不同的基質 (substrate),發現 glutathione reductase 除了可還原 GSH,也可以還原 GspdSH 與 GSH 間的雙硫鍵 (GspdS-SG),所以 De Craecker 等人推測,GspdSH 的作用可能 是調節GSH 或 spermidine 含量,或者 GspdSH 就像 TSH 在 Trypanosoma,是一 種比GSH 更好的抗氧化物,由於 GspdSH 在遲滯期會累積,GspdSH 也可能和壓 力反應 (stress response) 有關。
GSH 和 spermidine 結合,形成 GspdSH,此作用在 E. coli 中由酵素 GspS 進 行。 E. coli 的 gspS 在 1995 年首次由 Bollinger 等人發現,並定出基因及胺基酸 序列 (Bollinger et al., 1995),此基因位於 E. coli 染色體 67.5 分鐘處,全長 1860 bp,其產物 GspS 由 619 amino acids 組成,約 70 kDa。是一種具有雙重功能的酵 素 (bifunctional enzyme),有合成酶 (synthetase)活性,在 ATP 及 Mg2+存在下,
可催化glutathione 與 spermidine 間醯胺鍵的形成,產生 GspdSH。除了合成酶之 外,GspS 也具有醯胺酶 (amidase) 活性,可使 GspdSH 的醯胺鍵水解成 GSH 及 spermidine。之後,更進一步發現,GspS 的 N 端 1 至 195 胺基酸有醯胺酶 (amidase) 功能 (Pai et al., 2006),C 端 206 至 619 胺基酸是合成酶,linker region 為 196 至 205 胺基酸。
1.11 雙成份控制系統 BaeSR 之研究
許多細菌已發展出各種雙成份控制系統 (two component regulatory system),
並利用這些系統適應環境的改變,典型的雙成分控制系統包含兩種蛋白,一為 kinase sensor,位於細胞膜,另一為同源性 response regulator,存於細胞質中。雙 成分控制系統控制許多基因表現,與細菌的營養狀態,致病性,抗藥性等息息相 關。在E. coli 中,已知的雙成分控制系統大約有 30 種,但許多雙成分控制系統 的功能未明
藉由抑制 response regulators EnvZ、PhoR 及 CreC,Nagasawa 等人在 1993 年 時發現一新的雙成分控制系統 BaeSR (Nagasawa et al., 1993)。之後,Raffa 與 Raivio 等人探討 envelope stress 與 BaeSR 的關係時,發現 BaeSR 可以誘導 spy
(spheroplast protein Y,是一種 periplasmic protein,與 envelope 的形成有關)的 表現 (Raffa & Raivio, 2002),他們觀察在 spheroplast formation,indole 等 envelope stress 情況下,BaeSR 和另一組與 envelope stress 相關的雙成分控制系統 CpxAR 都可以誘導spy,在 spheroplast formation 誘導 spy 表現時,CpxAR 和 BaeSR 扮 演同樣重要的角色,無論是cpxR-或是baeR-,都會使spheroplast formation 對 spy 的誘導減低10 倍,但在 indole 造成的 envelope stress response 中,BaeSR 比 CpxAR 重要,因為在cpxR-時,spy 表現降低至三分之二,baeR-時,spy 表現降低至五分 之一。據此,他們認為BaeSR 可調控一組與 envelope stress 相關的基因表現,此 組基因與CpxAR 調控的基因不同,而 BaeSR 和 CpsAR 可能會被相同的訊號 (signal) 誘導,亦即某種 envelope stress (如 spheropast formation 或 indole) 造成 envelope proteins 的錯誤折疊,而不同種類的蛋白質折疊錯誤會分別促使 BaeSR 以及CpxAR 生成,此兩組蛋白質再進一步調控兩組與 envelope stress 相關的基 因表現。他們認為有一BaeSR regulon 存在,其組成基因除了與 envelope stress 相關的蛋白質以外,可能也包含與envelope stress 相關性未明的 cytoplasmic
proteins。
此外,根據Garbe 等人在 2000 年的研究結果顯示,indole 可誘導 superoxide dismutase 及 alkylhydroperoxide 生成,此兩者和氧化毒性 (oxidant toxicity) 相關 (Garbe et al., 2000),因此認為 indole 會破壞細胞膜,造成氧化損害 (oxidative damage),使細胞的氧還狀態 (redox potential) 發生改變,產生過氧化物,誘發 superoxide dismutase 及 alkylhydroperoxide 生成。基於以上理由,Raffa 與 Raivio 等人推論,BaeSR 可能對細胞膜的改變或 envelope 氧還狀態的破壞有反應,進 而引發下游基因表現。
接著,Baranova 等人亦發現,BaeR 大量表現時,E. coli 的抗藥性增加 (Baranova & Nikaido, 2002), 並經由實驗證實 baeSR 位於 multidrug transporter gene cluster mdtABCD 下游,且 BaeR 可直接與 mdtA 的 promoter 結合,至此,
BaeSR 已被證實與 envelope stress 及 multidrug transport 相關。
一般認為不同組雙成分控制系統之間,可能有交互作用 (cross talk),但 Verhamme 等人觀察四組雙成分控制系統 Uhp (與 phosphorylated sugar 相關), Pho (與 phosphate 相關), Ntr (與 nitrogen 相關), Arc (與 oxygen 相關) (Verhamme et al., 2002),認為非同源性 (noncognate),且雙成分控制系統之間的交互作用並非 主要的調控方式,此結果使Raffa 等人解釋 BaeSR 與 CpxAR 皆可誘導 spy 時 (Raffa & Raivio, 2002),傾向於 BaeSR 與 CpxAR 以不同的方式調控 envelope 相 關基因,且兩組基因之間可能有重疊,而非BaeSR 會與 CpxAR 發生交互作用。
Hirakawa 等人發現,大量表現 response regulators BaeR、EvgA、CpxR、
OmpR、RcsB 會誘發許多有助排除藥物的基因 (drug exporter genes) 表現 (Hirakawa et al., 2003a; Hirakawa et al., 2003b)。2005 年時,他們探討 indole 如何 誘導multidrug exporter genes (Hirakawa et al., 2005),除了證明兩種基因 acrD 和 mdtA 的表現受 BaeSR 與 CpxAR 控制,並且發現 BaeSR 所產生的誘導不需 CpxAR 的幫助,但CpxAR 所產生的誘導需要 BaeSR。並且找出兩種 response regulator (BaeR 和 CpxR) 與 acrD 及 mdtA promoter 結合的位置。因此,Hirakawa 等人認
為,indole 可刺激 kinase sensor BaeS 和 CpxA,兩者分別與 BaeR 及 CpxR 作用 之後,BaeR 直接誘導 acrD 及 mdtA,CpxR 則可增強 BaeR 的影響。
2005 年,Nishino 等人欲對 BaeSR 做一全面性的瞭解,他們利用生物晶片分 析,發現將response regulator BaeR 過量表現,可誘發 59 個基因表現 (Nishino et al., 2005),他們將這些基因分為五大類,分別是 two component system、chemotactic response、flagellar biosynthesis、maltose transport 以及 multidrug transport,其中 gspS 的表現增強為原來的 13 倍,但未明確歸類。他們比較野生株 (wild type) 和 baeSR-兩種情況下的基因表現,發現gspS 在兩種菌株中的比值 (wt/baeSR-) 為 1.7,沒有顯著不同。他們也在培養時加入 indole,發現在野生株中,加 indole 和不加indole 兩者的 gspS 表現量比值為 1.2 (indole/no indole =1.2),在 baeSR-情 況下,比值為0.8,兩者差異不大,因此認為 indole 經由 BaeSR 對 gspS 的誘導 並非直接作用,此外,在phoBR-情況下,gspS 被 BaeR 過量表現誘導的倍數為 14 倍(有些基因誘導倍數會下降至 0),因此推論 gspS 受 BaeR 的調控與 PhoBR 與BaeSR 間的交互作用無關。不僅如此,他們更利用三種條件:(1)會被 BaeR 過量表現誘導,(2)野生株和 baeSR-兩種情況下的基因表現比值大於2,(3) 在野 生株中,加indole 和不加 indole 兩狀況的表現量比值大於 2,篩選出直接受 BaeSR 調節的BaeSR regulon,但 gspS 並未包含在此 regulon 當中,最後,他們比對 BaeSR regulon 基因的啟動子區域,歸納出一 BaeR 與啟動子的結合區,但此區的位置,
與前述Hirakawa 等人在 2005 年經實驗所得之 BaeR 與啟動子結合區稍有不同 (Hirakawa et al., 2005)。
1.12 gspS+實驗目的
E. coli 的 gspS 長 1860 bp,為一反向基因,起始密碼子 (start codon) 位 在E. coli 基因組(genome)全序列 3136544 處,產物為 glutathionylspermidine
synthetase (GspS),共有 619 個胺基酸,是一種具有雙重功能的酵素 (bifunctional enzyme),可執行 GSH 和 spermidine 之間醯胺鍵的合成與分解,至目前為止,尚 未有任何與gspS 轉錄調控 (transcription regulation) 相關之研究。本文對大腸桿
菌gspS 基因轉錄做一初步探討,已確認 gspS 為一單獨的轉錄單位而非以操縱子
形式存在,且GspS 起始密碼上游序列具有啟動子功能,而在 in vivo 情況下,H2O2
與BaeR 都可誘導 gspS 之表現。
第二章 調控蛋白 sigma factors 對 clpQ
+clpY
+表現的影響
2.1 摘要
大腸大腸桿桿菌菌的的CCllppYYQQ蛋白蛋白酶屬屬於於熱熱休休克克蛋蛋白白的的一一種種,,而而σ factor 是一些熱休克 蛋白的調控蛋白(regulator),clpQ+clpY+ operon 啟動子(promoter)中的一段序列 CCCCCCCAT CTATAATT,具有具有σ32 (rpoH) 可辨可辨識識的的保保守守序序列列 (c(coonnsseennssuuss bbiinnddiinngg seseqquueennccee)) CXCCCCAT.T (圖一),且熱休克可增加 clpQ+clpY+的表現 (Chuang et al., 1993),但熱休克誘導 clpQ+clpY+的表現是否由σ32 (rpoH)直接造成則尚未證 實。本研究利用clpQ+clpY+ 啟動子與報導基因 lacZ 所建構之融合基因
clpQ+::lacZ,在菌株的培養過程中,提高溫度造成熱休克反應的條件,使 σ32誘 導clpQ+::lacZ 大量表現,但在 rpoH -突變株中則無此誘導現象;並利用
β-galactosidase 活性分析與北方轉印法,證實 clpQ+clpY+operon 的 mRNA 及蛋白 質表現,在 rpoH -突變株中均會降低。另外,針對clpQ+clpY+啟動子上σ32 (rpoH) 可辨可辨識識的的保保守守序序列列做C→T 點突變,造成 σ32對clpQm(c→t)::lacZ 的熱誘導降低,
本文首次證明σ32可直接調控clpQ+clpY+啟動子。
2.2 材料與方法
2.2.1 建立 clpQ+clpY+上游啟動子區域 ( promoter region ) 表現系統 (a) 菌株,載體,培養基和培養條件
本研究選殖基因的載體為 pRS415 與 pRS414 (Simons et al., 1987),兩載體皆 以lacZ 為 reporter gene,pRS415 用以建構不同長度 clpQ+clpY+ promoter region 與lacZ 之轉錄融合基因(operon fusion or transcriptional fusion),包含 clpQ+::lacZ (op-141bp),clpQ+::lacZ (op-478bp),clpQm(c→t)::lacZ (op-141bp),clpQm1::lacZ (op-141bp)及 clpQm2△40bp::lacZ (op),pRS414 用於建構轉譯融合基因(protein