國立臺灣大學工學院化學工程學研究所 碩士論文
Department of Chemical Engineering College of Engineering
National Taiwan University Master Thesis
利用 Rhodococcus erythropolis NTU-1 細胞聚集現象 移除正十六烷
Rhodococcus erythropolis NTU-1 Biofloccules for n-Hexadecane Removal
謝惠敏 撰 Hui-Min Hsieh
指導教授: 劉懷勝 博士 Advisor: Hwai-Shen Liu, Ph.D.
中華民國 一百 年 七 月
July, 2011
I
致謝
時光匆匆流逝,碩士生涯不知不覺也到達了尾聲。一路走來,我受到許多人 的支持與鼓勵。這兩年來首先要感謝的是我的指導教授 劉懷勝老師,您讓我們不 斷的去想問題及解決問題,並適時的指點迷津,不時的鼓勵我們。您的教學方式 及處事態度都值得我們學習,未來的生活中我會謹記您給我的建議及啟發的。
感謝工研醋 許駿發博士、台科大 李振綱博士及 王孟菊博士在百忙之中擔任 我的口試委員,給了我許多寶貴的建議,使本論文能夠更加精進。
接下來要感謝的是實驗室的伙伴們。感謝文興學長平常生活中的經驗分享,
在我口試前也提醒了我許多該注意的地方。感謝志文學長在實驗及生活上的提點,
不僅教了我許多實驗上的技巧也提出了許多有趣的想法供我參考。感謝勻錡學姐 在我實驗後期的幫忙,使我的論文更加有說服力。此外,我要感謝玹紳、緯農、
榕庭、煒泰、易修及翔瑜學長,謝謝你們在研究上及生活上給予我的幫助及建議。
感謝一同奮鬥的祐安及詞伊,這兩年不管是在課業上或生活態度上我都從你 們身上學到很多,還記得之前一起在實驗區做實驗時,雖然很辛苦,但是我們都 互相加油,那段日子也成為了我碩士生涯中難忘的回憶。感謝舒婷熱心的幫忙,
在我口試前也分享了許多經驗及該注意的細節。感謝有趣的學弟們鎬綱、政勛及 鈺傑,希望接下來你們的實驗都能順順利利。
我還要感謝一起從中央進入台大就讀的朋友們,瑋菱、佩吟、怡怡、彥廷、
映萱及明哲,謝謝你們這兩年的陪伴,不時的聽我抱怨,為我鼓勵。
最後我要感謝我的家人們,謝謝你們一直支持我,給我依靠,讓我有大步向 前的動力,對你們的感謝是不可言喻的。很慶幸我的生活中有你們,未來我也會 為了你們繼續努力的。
要感謝的人還有很多,在此謹以此論文獻給所有我需要感謝的你們!
謝惠敏,2011 年 7 月
II
中文摘要
本研究目的主要在於探討如何提升及應用 R. erythropolis NTU-1 的細胞聚集現 象,使長鏈烷類在短時間內能大量移除。
研究結果發現,在培養錐形瓶中加入篩網可幫助迴旋式培養下NTU-1 與正十 六烷的接觸機會及形式,因而NTU-1 在 44 小時內即能形成結塊,配合物理撈除的 方式可將2000 ppmv 正十六烷移除掉 95%以上。
利用冷凍乾燥的方式來保存以NB 培養的 NTU-1 細胞,實驗結果發現使用麥 芽糖溶液作為冷凍乾燥保護劑,並存放於-20℃的環境下,乾燥的 NTU-1 在經過 30 天存放後仍具有很高的存活率且其生理特性並不會受到影響。同樣利用冷凍乾 燥的方式來乾燥以正十六烷培養而得的NTU-1 細菌結塊,然而實驗結果顯示乾燥 後的NTU-1 結塊活性很低,無法再降解烷類,但其再形成結塊包覆烷類的能力並 不會受到影響,1 ~ 2 小時內即能再次形成聚集體,第 12 小時以物理撈除的方式可 移除80 ~ 90%濃度為 2000 ppmv 的正十六烷。
接著以烘乾的方式取代成本高及費時的冷凍乾燥法來乾燥NTU-1 結塊。實驗 中選擇烘乾溫度為80℃,其乾燥所需時間約 15 小時且乾燥後的 NTU-1 結塊具有 良好形成聚集包覆烷類的效果。這個部分換算出利用1g/L 的 NTU-1 結塊密度來處 理10000 ppmv 的正十六烷較佳。另外,若以兩段式添加乾燥 NTU-1 結塊的方法,
可使正十六烷的移除效率在12 小時內由 90%提升至將近 100%。
乾燥後NTU-1 結塊再形成聚集將烷類包覆的原因與其細胞表面疏水性及完整 性有關,此兩項特性需同時存在,乾燥NTU-1 結塊才能貼附烷類且以堆疊的方式 形成大顆粒結塊。
由此可知,將NTU-1 結塊冷凍乾燥或烘乾後保存,利用乾燥 NTU-1 結塊的高 度包覆烷類能力,配合物理撈除的方式,使烷類能在12 小時內大量移除,此項成 果無疑的為將來石油污染物移除提供了一個相當具競爭性及發展性的方法。
III
Abstract
Rhodococcus erythropolis NTU-1 is a strain which can not only degrade
hydrocarbons but also traps alkanes in biofloccules during bioremediation process. In this study, we focused on how to accelerate biofloccules formation and to apply NTU-1 biofloccules for n-hexadecane (n-C16) removal.
Results showed that sieves in Erlenmeyer flasks increased the n-C16 agitation efficiency under orbital shaking and assisted NTU-1 to utilize n-C16. NTU-1 formed biofloccules and trapped most residual n-C16 within 44 hr. At this time, more than 95%
n-C16 was removed.
NTU-1 cells (cultured by Nutrient Broth) by freeze-drying preserved most viability with maltose as the protectant. However, freeze-dried NTU-1 biofloccules (cultured by n-C16) resulted in low cell viability. Dried NTU-1 biofloccules did not degrade n-C16 but they still re-formed biofloccules and re-trapped most of n-C16. With these
freeze-dried biofloccules, n-C16 removal efficiency achieved 80 ~ 90% within 12 hr.
Heat-dried biofloccules were further evaluated because of its simplicity in comparison with freeze-drying method. It proved a good method and we found 1g/L dried cell vs. 10000 ppmv n-C16 was an optimal ratio. Moreover, with two-step addition of dried NTU-1 biofloccules, near 100% of n-C16 was removed in 12 hr.
The mechanism of re-forming biofloccules was found to relate to the cell
hydrophobicity and integrity. With these two characteristics, dried NTU-1 biofloccules adhered to n-C16, and n-C16 worked as the linker to form cell pellets in a short time.
These dried biofloccules provided a feasible and potential method for remediation of hydrocarbon pollutants.
IV
目錄
致謝 ... I 中文摘要 ... II Abstract ... III 目錄 ... IV 圖目錄 ... VII 表目錄 ... XIV 照片目錄 ... XV
第一章 緖論 ... 1
1.1 前言 ... 1
1.2 研究目的及論文綱要 ... 2
第二章 文獻回顧 ... 3
2.1 石油碳氫化合物簡介及其對環境和人類之影響 ... 3
2.2 處理石油碳氫化合物污染之方法 ... 5
2.3 生物處理石油碳氫化合物 ... 9
2.3.1 生物復育簡介 ... 9
2.3.2 微生物攝取碳氫化合物之模式 ... 14
2.3.3 微生物分解碳氫化合物之方式 ... 20
2.4 微生物降解碳氫化合物之代謝途徑 ... 23
2.4.1 直鏈烷之氧化機制 ... 23
2.4.2 支鏈烷之氧化機制 ... 26
2.4.3 烯烴類及環烷類之氧化機制 ... 31
2.5 實驗菌株 Rhodococcus erythropolis 之介紹 ... 34
2.5.1 Rhodococcus 菌屬簡介 ... 34
2.5.2 Rhodococcus erythropolis 之特性及應用 ... 37
2.6 微生物之細胞聚集現象 ... 42
2.7 微生物乾燥技術及應用 ... 45
2.7.1 冷凍乾燥 (Freeze-drying) ... 45
2.7.1.1 冷凍乾燥對菌株的影響 ... 46
2.7.1.2 冷凍乾燥時保護劑的添加 ... 48
2.7.1.3 冷凍乾燥後菌株貯存之安定性 ... 51
2.7.2 烘乾之原理及應用 ... 52
V
第三章 實驗材料與方法 ... 53
3.1 實驗菌株 ... 53
3.2 培養基組成與配製 ... 55
3.2.1 液態礦物培養基 ... 55
3.2.2 菌株保存培養基 ... 58
3.2.3 菌株活化培養基 ... 59
3.2.4 計數平板培養基 ... 59
3.3 實驗方法 ... 60
3.3.1 菌株的活化及生長曲線 ... 60
3.3.2 礦物培養基菌液製作 ... 61
3.3.3 正十六烷之生物降解與生物復育實驗 ... 62
3.3.4 以 NB 培養的 NTU-1 進行冷凍乾燥及其存活率測試 ... 65
3.3.5 以正十六烷培養的 NTU-1 細菌結塊進行冷凍乾燥 ... 67
3.3.6 烘乾以正十六烷培養的 NTU-1 細菌結塊 ... 68
3.3.7 乾燥後 NTU-1 細菌結塊對於正十六烷的包覆與移除 ... 69
3.3.8 不同條件乾燥下 NTU-1 細菌結塊之細胞表面疏水性測定 ... 70
3.4 實驗藥品與器材 ... 71
3.4.1 實驗藥品 ... 71
3.4.2 實驗儀器 ... 72
第四章 實驗結果與討論 ... 73
4.1 不同搖晃培養對於 NTU-1 生物降解及包覆正十六烷能力的影響及改進方 法 ... 74
4.1.1 比較往復式及迴旋式培養下 NUT-1 對正十六烷生物復育的效果 . 74 4.1.2 迴旋式培養下錐形瓶中加入篩網時正十六烷生物復育的效果 ... 81
4.1.2.1 篩網孔徑大小為 10 mesh ... 81
4.1.2.2 篩網孔徑大小為 20 mesh ... 89
4.1.2.3 不同孔徑大小但相同折角 (45 度) 篩網之比較 ... 95
4.1.3 討論 ... 98
4.2 以 NB 培養的 NTU-1 冷凍乾燥後細胞存活率及降解包覆能力探討 ... 101
4.2.1 利用 NB 培養的 NTU-1 經過冷凍乾燥後之存活率探討 ... 101
4.2.2 以 NB 培養的 NTU-1 經過冷凍乾燥後對於正十六烷降解及包覆能力 探討 ... 109
4.2.3 討論 ... 113
4.3 利用正十六烷培養的 NTU-1 細菌結塊冷凍乾燥後其存活率及降解包覆能 力探討 ... 114
4.3.1 NTU-1 細菌結塊降解及包覆正十六烷的能力 (未經冷凍乾燥) ... 114
4.3.2 NTU-1 細菌結塊在冷凍乾燥後降解及包覆正十六烷的能力 ... 120
VI
4.3.3 超音波震碎 NTU-1 結塊並進行冷凍乾燥後之應用 ... 144
4.3.4 討論 ... 146
4.4 利用正十六烷培養的 NTU-1 細菌結塊經烘乾後再聚集包覆烷類之能力探 討 ... 149
4.4.1 烘乾 NTU-1 結塊時溫度與時間的選擇 ... 149
4.4.2 不同條件乾燥下 NTU-1 結塊表面疏水性測定 ... 156
4.4.3 烘乾後的 NTU-1 結塊存放溫度與時間之探討 ... 159
4.4.4 烘乾後 NTU-1 結塊於培養過程 12 小時間的細胞聚集情形 ... 162
4.4.5 以不同量之乾燥 NTU-1 結塊處理 2000 ppmv 正十六烷 ... 166
4.4.6 以固定量之乾燥 NTU-1 結塊處理不同濃度的正十六烷 ... 171
4.4.7 提高乾燥 NTU-1 結塊移除正十六烷效率的方法 ... 177
4.4.8 乾燥 NTU-1 結塊再形成聚集體包覆正十六烷之機制 ... 181
4.4.9 討論 ... 184
第五章 結論 ... 187
參考文獻 ... 190
附錄 1 NB 培養的 NTU-1 及正十六烷培養的 NTU-1 結塊經冷凍乾燥後的結合應 用 ... 201
附錄 2 不同條件乾燥後 NTU-1 結塊之細胞粒徑大小 ... 207
VII
圖目錄
第二章
圖2.3.2-1 原油中常見的碳氫化合物化學結構分類 (Alloway and Ayres 1998)。 ... 14 圖2.3.2-2 微生物細胞攝取有生物界面活性劑附著的碳氫化合物示意圖 (Ratledge 1988)。 ... 18 圖2.3.2-3 菌株 Pseudomonas sp.分泌界面活性劑後攝取碳氫化合物示意圖 ... 18 圖2.3.3-1 好氧性微生物降解碳氫化合物的主要流程 (Fritsche and Hofrichter 2000)。
... 21 圖2.3.3-2 長碳鏈烷類的好氧及厭氧性降解途徑 (點狀線的箭頭是指推測的代謝路 徑) (Wentzel et al. 2007)。 ... 22 圖2.4.1-1 微生物代謝分解直鏈烷之途徑 (圖中 1 為 n-alkane 單氧化酶、2 為醇脫
氫酶、3 為醛脫氫酶) (Britton 1984)。 ... 25 圖2.4.2-1 菌株 Brevibacterium erythrogens 降解異十九烷的途徑 (Pirnik et al. 1974)。
... 27 圖2.4.2-2 異十九烷可能的降解途徑。圖中的標記 (R) 及 (M) 分別代表 R. ruber
SBUG 82 及 M. neoaurum SBUG 109 氧化過程中檢測出的產物
( means novel metabolic route; means major metabolic route of β-oxidation) (Nhi-Cong et al. 2009)。 ... 29 圖2.4.2-3 異十九烷次末端氧化過程中以 2, 6, 10, 14-tetramethyl-pentadecan-3-one
作為中間產物,利用ketone monooxygenase 進行的兩種不同單末端氧化 反應。Pathway IIIA 為 R. ruber SBUG 82 及 M. neoaurum SBUG 109 的反 應途徑 (Nhi-Cong et al. 2009)。Pathway IIIB 為 Rontani 等人提出的海洋 混合菌株分解異十九烷途徑 (Rontani et al. 1986)。 ... 30 圖2.4.3-1 烯烴類生物降解可能的代謝途徑 (張蘭英等 2007)。 ... 31 圖2.4.3-2 環己烷的代謝途徑 (Fritsche and Hofrichter 2000)。 ... 32 圖2.4.3-3 不產孢放線菌 (non-sporulating actinomycetes) 降解利用不同碳氫化合物 之途徑及經過β-oxidation 後合成細胞生長所需相關物質 (Alvarez 2003)。
... 33 圖2.5.1-1 不同氧化酶促使 Rhodococcus 菌屬引發不同化學反應並能生長及降解掉 大範圍的有機污染物。(A) 環氧化 (eg. cytochrome P450 enzymes)、(B) 順 式雙羥基化反應 (e.g. by dioxygenase)、(C) 對二級烷基的羥基化、(D) 對 一級烷基的羥基化 (e.g. alkane monooxygenase)、(E) 磺化氧化作用 (sulfoxidation of sulfide to sulfoxide)、(F) 磺化氧化作用 (sulfoxidation of sulfoxide to sulfone) (Larkin et al. 2005)。 ... 37 圖2.7.1.1-1 微生物冷凍乾燥的步驟示意圖 (Morgan et al. 2006)。 ... 47
VIII
第三章
圖3.3.1-1 NTU-1 菌株在 Nutrient Broth (NB) 營養液中的生長曲線。 ... 60 圖3.3.3-1 正十六烷在氣相層析儀中之校正曲線 ... 64 第四章
圖4.1.1-1 培養條件 30℃、100 rpm、初始 pH 值 6.8,NTU-1 處理 2000 ppmv 正十 六烷時,不同搖晃培養方式下的培養基酸鹼值變化。 ... 77 圖4.1.1-2 培養條件 30℃、100 rpm、初始 pH 值 6.8,NTU-1 處理 2000 ppmv 正十 六烷時,不同搖晃培養方式下之細胞生長曲線。 ... 78 圖4.1.1-3 培養條件 30℃、100 rpm、初始 pH 值 6.8,NTU-1 處理 2000 ppmv 正十 六烷時,不同搖晃培養方式下的正十六烷生物降解曲線。 ... 78 圖4.1.1-4 培養條件 30℃、100 rpm、初始 pH 值 6.8,NTU-1 處理 2000 ppmv 正十 六烷時,不同旋轉培養方式下之生物降解量及包覆量。(A) 往復式;(B) 迴 旋式。 ... 79 圖4.1.2.1-1 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,錐形瓶中加入不同折角之 10 mesh 篩網之培養 基酸鹼值的變化。 ... 84 圖4.1.2.1-2 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,錐形瓶加入不同折角之 10 mesh 篩網時細胞生 長曲線。 ... 85 圖4.1.2.1-3 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,錐形瓶中加入不同折角之 10 mesh 篩網,正十 六烷生物降解量。 ... 85 圖4.1.2.1-4 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,於錐形瓶中加入 10 mesh 不同折角篩網之正十 六烷生物降解量及包覆量比較。(A) 沒有加入篩網;(B) 篩網折角為 180 度;(C) 篩網折角為 90 度;(D) 篩網折角為 45 度。 ... 86 圖4.1.2.2-1 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處
理2000 ppmv 正十六烷,於錐形瓶中加入 20 mesh 篩網不同折角時之培 養基酸鹼值變化。 ... 91 圖4.1.2.2-2 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,於錐形瓶中加入 20 mesh 篩網不同折角時之細 胞生長曲線。 ... 91 圖4.1.2.2-3 培養條件 30℃、迴旋式搖晃 100 rpm、初始 pH 值 6.8,NTU-1 處理 2000
ppmv 正十六烷,錐形瓶中加入 20 mesh 篩網時之正十六烷生物降解量。
... 92 圖4.1.2.2-4 培養條件 30℃、100 rpm、初始 pH 值 6.8,NTU-1 處理 2000 ppmv 正 十六烷,錐形瓶中加入20 mesh 篩網之生物降解量及包覆量比較。(A) 沒
IX
有加入篩網;(B) 篩網折角為 180 度;(C) 篩網折角為 90 度;(D) 篩網 折角為45 度。 ... 92 圖4.1.2.3-1 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,於錐形瓶中加入折角 45 度不同孔徑大小篩網之 培養基酸鹼值變化。 ... 96 圖4.1.2.3-2 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,於錐形瓶中加入折角 45 度不同孔徑大小篩網之 細胞生長曲線。 ... 96 圖4.1.2.3-3 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,於錐形瓶中加入折角 45 度不同孔徑大小篩網之 正十六烷生物降解量。 ... 97 圖4.1.2.3-4 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 處 理2000 ppmv 正十六烷,於錐形瓶中加入折角 45 度、6 mesh 篩網之正十 六烷生物降解量及包覆量比較。 ... 97 圖4.2.1-1 不同保護劑下,以 NB 培養的 NTU-1 在-20℃冷凍 24 小時後的存活率。
... 103 圖4.2.1-2 不同保護劑下,以 NB 培養的 NTU-1 經過冷凍乾燥後馬上進行復水的細 胞存活率。 ... 104 圖4.2.1-3 以 NB 培養的 NTU-1,添加不同保護劑並在冷凍乾燥後存放於不同溫度 下0,10 及 30 天後的存活率。 (A) -20℃、(B) 4℃、(C) 30℃。 ... 106 圖4.2.1-4 以 NB 培養的 NTU-1,添加保護劑 1%或 10%麥芽糖溶液並在冷凍乾燥
後存放於不同溫度下0,10 及 30 天後的存活率 (A) -20℃、(B) 4℃、(C) 30℃。 ... 108 圖4.2.2-1 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,培養基中加 入以保護劑1%麥芽糖冷凍乾燥後的 NTU-1,處理 2000 ppmv 正十六烷時 之酸鹼值變化。 ... 111 圖4.2.2-2 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,培養基中加 入以保護劑1%麥芽糖冷凍乾燥後的 NTU-1,處理 2000 ppmv 正十六烷時 之細胞密度變化趨勢。 ... 111 圖4.2.2-3 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,培養基中加 入以保護劑1%麥芽糖冷凍乾燥後的 NTU-1,處理 2000 ppmv 正十六烷時 之正十六烷生物降解量。 ... 112 圖4.2.2-4 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,培養基中加 入以保護劑1%麥芽糖冷凍乾燥後的 NTU-1,處理 2000 ppmv 正十六烷時 之正十六烷生物降解量及包覆量比較。 ... 112 圖4.3.1-1 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 形成 細菌結塊後重新更換MSM 培養基及添加 2000 ppmv 正十六烷,不同時 間下培養基中酸鹼值的變化。(更換 MSM 時間:第 72、102 小時)。 . 118
X
圖4.3.1-2 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 形成 細菌結塊後重新更換MSM 培養基及添加 2000 ppmv 正十六烷,不同時 間下培養基中細胞密度的變化。(更換 MSM 時間:第 72、102 小時)。
... 119 圖4.3.1-3 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1 形成 細菌結塊後重新更換MSM 培養基及添加 2000 ppmv 正十六烷,不同時 間下培養基中正十六烷分布的變化。(更換 MSM 時間:第 72、102 小時)。
... 119 圖4.3.2-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊 (以 10%麥芽糖或 10%甘露糖醇 為保護劑),處理 2000 ppmv 正十六烷時之酸鹼值變化。 ... 124 圖4.3.2-2 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊 (以 10%麥芽糖或 10%甘露糖醇 為保護劑),處理 2000 ppmv 正十六烷時之細胞密度分布及變化趨勢。
... 124 圖4.3.2-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊,(A) 以 10%麥芽糖、(B) 10%
甘露糖醇為保護劑,處理2000 ppmv 正十六烷時之正十六烷初始量及反 應中殘餘在培養基及被包覆量的分布圖。 ... 128 圖4.3.2-4 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊 (以 10%麥芽糖或 10%甘露糖醇 為保護劑),處理 2000 ppmv 正十六烷時之正十六烷總移除百分比。 128 圖4.3.2-5 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷,細 胞密度的變化及分布。(A) 10%麥芽糖為添加劑,不同保存條件:0 天及
-20℃下 10 天; (B) 10%甘露糖醇及 10%乳糖為添加劑,存放在-20℃
下10 天。 ... 134 圖4.3.2-6 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷,正 十六烷初始量及反應中殘餘在培養基及被包覆量的分布圖。 (A) 10%麥 芽糖為添加劑,不同保存條件:0 天及-20℃下 10 天;(B) 10%甘露糖醇 及10%乳糖為添加劑,存放在-20℃下 10 天。 ... 135 圖4.3.2-7 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷,正 十六烷總移除百分比。 ... 135 圖4.3.2-8 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷時之 細胞密度的變化及分布。 (A) 10%麥芽糖為添加劑,不同保存條件:0
XI
天及-20℃下 10 天;(B) 10%甘露糖醇及 10%乳糖為添加劑,相同保存 條件:-20℃下 10 天。 ... 139 圖4.3.2-9 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷時,
正十六烷初始量及反應中殘餘在培養基及被包覆量的分布圖。 (A) 10%
麥芽糖為添加劑,不同保存條件:0 天及-20℃下 10 天;(B) 10%甘露糖 醇及10%乳糖為添加劑,相同保存條件:-20℃下 10 天。 ... 140 圖4.3.2-10 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養中加入以不同添加劑冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正 十六烷時之正十六烷總移除百分比。 ... 140 圖4.3.2-11 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物 培養基中加入冷凍乾燥後的NTU-1 結塊,存放於 30℃下 30 天,處理 2000 ppmv 正十六烷時之細胞密度的變化及分布。... 142 圖4.3.2-12 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 結塊,存放於 30℃下 30 天,處理 2000 ppmv 正十六烷時,正十六烷初始量及反應中殘餘在培養基及被包覆量的 分布圖。 ... 143 圖4.3.2-13 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物 培養基中加入冷凍乾燥後的NTU-1 結塊,存放於 30℃下 30 天,處理 2000 ppmv 正十六烷時之正十六烷總移除百分比。... 143 圖4.4.1-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,培養基中加
入不同烘乾條件乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷,第 12 小時之細胞密度分布。 ... 154 圖4.4.1-2 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,培養基中加 入不同烘乾條件乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷,第 0、
12 小時正十六烷殘餘在培養基及包覆量的分布圖。 ... 155 圖4.4.1-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,培養基中加 入不同烘乾條件乾燥後的的NTU-1 結塊,處理 2000 ppmv 正十六烷,第 12 小時之正十六烷總移除百分比。 ... 155 圖4.4.2-1 不同乾燥條件下 NTU-1 細胞表面疏水性測定 ... 158 圖4.4.3-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基
中加入烘乾後不同存放條件下的NTU-1 結塊,處理 2000 ppmv 正十六烷,
第12 小時的 NTU-1 細胞密度之分布。 ... 160 圖4.4.3-2 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入烘乾後不同存放條件下的NTU-1 結塊,處理 2000 ppmv 正十六烷,
第12 小時正十六烷殘餘在培養基及包覆量的分布圖。 ... 161 圖4.4.3-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入烘乾後不同存放條件下的NTU-1 結塊,處理 2000 ppmv 正十六烷,
XII
第12 小時之正十六烷總移除百分比。 ... 161 圖4.4.4-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷時,隨著時間培 養基酸鹼值的變化。 ... 164 圖4.4.4-2 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基
中加入烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷時,隨著時間 NTU-1 細胞密度之分布。 ... 165 圖4.4.4-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷時,隨著時間正 十六烷殘餘在培養基及包覆量的分布圖。 ... 165 圖4.4.4-4 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷時,隨著時間正 十六烷總移除百分比之變化圖。 ... 166 圖4.4.5-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入不同初始重量烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷第 12 小時 NTU-1 細胞密度之分布。 ... 169 圖4.4.5-2 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入不同初始重量烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷,
第12 小時正十六烷殘餘在培養基及包覆量的分布圖。 ... 170 圖4.4.5-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入不同初始重量烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷,
第12 小時正十六烷總移除百分比。 ... 170 圖4.4.6-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入相同初始重量烘乾後的NTU-1 結塊,處理不同濃度正十六烷,第 12 小時 NTU-1 細胞密度之分布。 ... 175 圖4.4.6-2 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入相同初始重量烘乾後的NTU-1 結塊,處理不同濃度正十六烷,第 12 小時正十六烷殘餘在培養基及包覆量的分布圖。 ... 176 圖4.4.6-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養基 中加入相同初始重量烘乾後的NTU-1 結塊,處理不同濃度正十六烷,第 12 小時正十六烷總移除百分比。 ... 176 圖4.4.7-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,兩段式添加 0.03 g 乾燥 NTU-1 結塊 (第 0 小時及第 6 小時),處理 2000 ppmv 正十六 烷,不同時間點下NTU-1 細胞密度之分布。 ... 179 圖4.4.7-2 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,兩段式添加 0.03 g 乾燥 NTU-1 結塊 (第 0 小時及第 6 小時),處理 2000 ppmv 正十六 烷,不同時間點殘餘在培養基中正十六烷的濃度分布。 ... 180 圖4.4.7-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,兩段式添加
XIII
0.03 g 乾燥 NTU-1 結塊 (第 0 小時及第 6 小時),處理 2000 ppmv 正十六 烷,不同時間點下正十六烷的移除效率百分比。 ... 180 圖4.4.8-1 乾燥後 NTU-1 結塊表面完整時,細胞貼附正十六烷形成大顆粒結塊示
意圖。 ... 183 圖4.4.8-2 乾燥後 NTU-1 結塊表面不完整時 (細胞碎片),細胞貼附正十六烷示意
圖。 ... 183 圖4.4.8-3 NTU-1 結塊經過冷凍乾燥或以 60 及 80℃的條件烘乾後,細胞貼附正
十六烷再形成聚集體之示意圖。 ... 184
附錄
附圖1-1 培養條件 30℃、100 rpm、初始 pH 值 6.8,培養基中加入以不同保護劑冷 凍乾燥後的NTU-1 及不同添加劑冷凍乾燥後的 NTU-1 結塊,處理 2000 ppmv 正十六烷第 24 小時之培養基酸鹼值變化。... 205 附圖1-2 培養條件 30℃、100 rpm、初始 pH 值 6.8,培養基中加入以不同保護劑冷
凍乾燥後的NTU-1 及不同添加劑冷凍乾燥後的 NTU-1 結塊,處理 2000 ppmv 正十六烷第 24 小時之細胞密度分布。... 205 附圖1-3 培養條件 30℃、100 rpm、初始 pH 值 6.8,培養基中加入以不同保護劑冷
凍乾燥後的NTU-1 及不同添加劑冷凍乾燥後的 NTU-1 結塊,處理 2000 ppmv 正十六烷。第 24 小時正十六烷初始量及殘餘在培養基及包覆量的 分布圖。 ... 206 附圖1-4 培養條件 30℃、100 rpm、初始 pH 值 6.8,培養基中加入以不同保護劑冷 凍乾燥後的NTU-1 及不同添加劑冷凍乾燥後的 NTU-1 結塊,處理 2000 ppmv 正十六烷,第 24 小時之正十六烷總移除百分比。... 206 附圖2-1 不同烘乾溫度下,NTU-1 細胞粒徑大小分布 ... 208 附圖2-2 以 120℃烘乾的 NTU-1 結塊在去離子水中所測得的細胞粒徑大小相對於
訊號強度之分布圖。 ... 209 附圖2-3 冷凍乾燥後的 NTU-1 結塊在去離子水中所測得之細胞粒徑大小相對於訊 號強度之分布圖。 ... 210 附圖2-4 先利用超音波將 NTU-1 結塊震碎再進行冷凍乾燥的 NTU-1 在去離子水中 所測得之細胞粒徑大小相對於訊號強度之分布圖。 ... 210
XIV
表目錄
第二章
表2.2-1 地下水污染之各種物理化學處理及生物處理程序之簡介與比較 (Van
Stempvoort and Biggar 2008)。 ... 7
表2.2-2 復育方法適用性與否的影響因素 (Vik et al. 2001)。 ... 8
表2.3.1-1 自然界中能利用石油碳氫化合物之好氧性微生物分類 (Van Beilen et al. 2003)。 ... 10
表2.3.1-2 實際應用的現地生物復育法 (Korda et al. 1997)。 ... 12
表2.3.1-2 (續) ... 13
表2.3.2-1 各種碳氫化合物在水中的溶解度 (Eastcott et al. 1988) ... 15
表2.3.2-2 各種微生物產生的生物界面活性劑分類 (Al-Araji et al. 2007)。 ... 19
表2.5.1-1 1994 ~ 2001 年中 Rhodococcus 菌屬分類的變動 (華及郭 2003)。 ... 35
表2.5.2-1 微生物降解碳氫化合物時之酵素分類 (Van Beilen and Funhoff 2007)。 ... 38
表2.5.2-2 Rhodococcus 菌屬細胞合成的醣脂質類型 (Lang and Philp 1998)。 ... 41
表2.7.1.2-1 使微生物在冷凍乾燥及貯存的過程中仍可維持高存活率的保護劑選擇 (LAB: lactic acid bacteria) (Carvalho et al. 2004)。 ... 49
第三章 表3.2.1-1 液態礦物培養基組成表 ... 57
表3.2.1-2 Trace Salt Solution 組成表 ... 57
表3.2.2-1 菌株保存培養基組成表 ... 58
XV
照片目錄
第三章
照片3.1-1 顯微鏡下的 Rhodococcus erythropolis NTU-1 ... 54 第四章
照片4.1.1-1 培養條件 30℃、往復式搖晃培養,轉速 100 rpm、初始 pH 值 6.8,NTU-1 處理2000 ppmv 正十六烷,不同時間點 NTU-1 細胞的聚集現象。(A) 第 44 小時;(B) 第 68 小時。 ... 80 照片4.1.1-2 培養條件 30℃、迴旋式搖晃培養,轉速 100 rpm、初始 pH 值 6.8,NTU-1 處理2000 ppmv 正十六烷,不同時間點 NTU-1 細胞的聚集現象。(A) 第 44 小時;(B) 第 68 小時。 ... 80 照片4.1.2.1-1 10 mesh 篩網不同折角置放於 250 mL 反應錐形瓶中。(A) 180 °;(B) 90 °;(C) 45 °。 ... 81 照片4.1.2.1-2 培養條件 30℃、迴旋式搖晃培養 100 rpm、初始 pH 值 6.8,NTU-1
處理2000 ppmv 正十六烷,於錐形瓶中加入 10 mesh 篩網不同折角,實 驗至第68 小時的細菌結塊形態。(A) 沒有加入篩網 (俯視);(B) 篩網折 角為180 度 (俯視);(C) 篩網折角為 90 度 (側視);(D) 篩網折角為 45 度 (俯視)。 ... 88 照片4.1.2.2-1 20 mesh 篩網不同折角置放於反應錐形瓶中。(A) 180 °;(B) 90 °;
(C) 45 °。 ... 89 照片4.1.2.2-2 迴旋式培養下 20 mesh 篩網不同折角實驗至第 68 小時的細菌結塊形 態。(A) 沒有加入篩網 (俯視);(B) 篩網折角為 180 度 (俯視);(C) 篩 網折角為90 度 (側視);(D) 篩網折角為 45 度 (俯視)。 ... 94 照片4.1.2.3-1 6 mesh 篩網 45 度折角置放於反應錐形瓶中。 ... 95 照片4.1.2.3-2 實驗條件 6 mesh 篩網 45 度折角在第 68 小時後細菌結塊圖 (開口左,
俯視)。 ... 98 照片4.1.2.4-1 以脂溶性染料 Sudan I (C16H12N2O) 將無色正十六烷染成橘色,加入
含有10 mesh 折角 180 度篩網的錐形瓶中,觀察不同時間 NTU-1 與正十 六烷在錐形瓶中的分布情形。(A) 第 0 小時;(B) 第 24 小時;(C) 第 48 小時;(D) 第 72 小時。 ... 99 照片4.2.2-1 以 1%麥芽糖為保護劑不同條件下的菌落形態。(A) 初始 NTU-1 菌落 形態;(B) 冷凍乾燥的 NTU-1 進行復水後的菌落形態。 ... 109 照片4.3.1-1 NTU-1 細菌結塊在重新更換培養基及添加 2000 ppmv 正十六烷後,
不同時間下,細胞結塊的情形。(A) 反應經過 72 小時後,移除培養基;
(B) 第 77 小時;(C) 第 102 小時;(D) 第 146 小時。 ... 116
XVI
照片4.3.2-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物 培養基中加入冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷時之 NTU-1 細菌結塊形態圖。(A) 10%麥芽糖為保護劑;(B) 10%甘露糖醇為 保護劑;(C) 10%麥芽糖組反應第 12 小時細菌結塊圖;(D) 10%甘露糖醇 組反應第12 小時細菌結塊圖。 ... 127 照片4.3.2-2 NTU-1 細菌結塊加入去離子水冷凍後,真空乾燥 2 天後所呈現的溼
黏狀態 (冷凍乾燥失敗)。 ... 129 照片4.3.2-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物 培養基中加入冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷,第 12 小時之 NTU-1 細菌結塊形態圖。(A) 10%麥芽糖添加劑,存放 0 天;
(B) 10%麥芽糖添加劑,於-20℃存放 10 天;(C) 10%甘露糖醇添加劑,
於-20℃存放 10 天;(D) 10%乳糖添加劑,於-20℃存放 10 天。 .... 133 照片4.3.2-4 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦
物培養基中加入冷凍乾燥後的NTU-1 結塊,處理 2000 ppmv 正十六烷,
第12 小時之 NTU-1 細菌結塊形態圖。(A) 10%麥芽糖添加劑,存放 0 天;
(B) 10%麥芽糖添加劑,於-20℃存放 10;(C) 10%甘露糖醇添加劑,於
-20℃存放 10 天;(D) 10%乳糖添加劑,於-20℃存放 10 天。 ... 138 照片4.3.3-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦物
培養基中加入冷凍乾燥後的NTU-1 碎片 (經超音波震碎),處理 2000 ppmv 正十六烷,第 24 小時之 NTU-1 細胞形態圖。 ... 145 照片4.3.5-1 冷凍乾燥前,以不同碳源培養 NTU-1 產生不同形態的 NTU-1 加入保
護劑 (添加劑) 後 NTU-1 的分布情形。(A) 以 NB 培養後的 NTU-1 均勻 分散在保護劑的情形;(B) 以正十六烷培養後的 NTU-1 結塊懸浮在添加 劑表面的情形。 ... 148 照片4.4.1-1 烘乾過程中,NTU-1 結塊不同乾燥程度之差別。(A) 未乾燥完全;(B)
完全乾燥。 ... 151 照片4.4.1-2 不同溫度烘乾後 NTU-1 結塊乾燥的情形,依左而右分別為 80℃、120
℃及160℃。 ... 151 照片4.4.1-3 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養 基中加入不同溫度烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷,第 12 小時 NTU-1 再形成結塊的情形。(A) 60℃;(B) 80℃;(C) 100℃;(D) 120℃;(E) 160℃。 ... 152 照片4.4.4-1 培養條件 30℃、100 rpm、初始 pH 值 6.8,礦物培養基中加入烘乾後 的NTU-1 結塊,處理 2000 ppmv 正十六烷時,隨著時間 NTU-1 細胞結 塊變化的情形。(A) 2 小時;(B) 4 小時;(C) 6 小時;(D) 9 小時;(E) 12 小時。 ... 164 照片4.4.5-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,培養基中 加入不同初始重量烘乾後的NTU-1 結塊,處理 2000 ppmv 正十六烷第 12
XVII
小時NTU-1 細胞結塊的形態。 ... 169 照片4.4.6-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,礦物培養
基中加入相同初始重量烘乾後的NTU-1 結塊,處理不同濃度正十六烷,
第12 小時 NTU-1 細胞結塊的形態。 ... 173 照片4.4.7-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,兩段式添
加0.03 g 乾燥 NTU-1 結塊 (第 0 小時及第 6 小時),處理 2000 ppmv 正十 六烷,不同時間點下的細胞結塊情形。(A) 第 6 小時;(B) 第 12 小時。
... 179 附錄
附錄照片1-1 培養條件 30℃、往復式震盪培養 100 rpm、初始 pH 值 6.8,MSM 礦 物培養基中加入乾燥後的NTU-1 (以 NB 培養) 及 NTU-1 結塊 (以正十六 烷培養),處理 2000 ppmv 正十六烷,第 20 及 24 小時之 NTU-1 生長及結 塊形態圖。 ... 203
1
第一章 緖論
1.1 前言
石油碳氫化合物廣泛的為人們所利用,與人類的食衣住行皆脫離不了關係。
然而隨著應用範圍越廣,對人類環境造成的污染也日益嚴重。石油的污染主要來 自於油田開採、煉製、運送及使用過程中不慎外漏或是排放到自然環境。2010 年3 月美國發生的墨西哥灣漏油事件更是震驚全球,約 2 億加侖的原油流入墨西 哥灣,造成海洋及沿岸生態環境嚴重影響,勘稱是美國遭遇過的最嚴重的一場環 境災難。除了對自然生態造成的污染可能需很長時間才能復原以外,其對於人體 的影響也是不容忽視的。石油污染物一旦進入人體,可能會影響體內的新陳代謝、
免疫系統、生殖系統甚至產生癌症等可怕的疾病。
為了降低石油碳氫化合物對環境及人類造成影響,各國環保單位對於石化工 業的發展皆採取高標準的規範,以防止其不當外洩或污染。處理石油碳氫化合物 污染的方法也越來越多,一般傳統上常利用物理法及化學法來處理石油污染物,
然而這兩種方法常有使用上的限制,如易造成環境二次污染、處理上較麻煩且成 本昂貴。
因此利用生物復育 (bioremediation) 法來處理石油碳氫化合物逐漸被重視,
生物復育是指利用微生物特有的生理機制來進行石油碳氫化合物污染的代謝及 降解,使這些污染物轉變為分子結構較簡單、毒性較低的物質 (Madsen 1991)。
而微生物在自然界的種類眾多,故能處理種類複雜的污染物,且微生物還有取得 上較容易、使用上較簡單、成本低及不會二次污染環境…等優點。
2
1.2 研究目的及論文綱要
本研究所使用的 R. erythropolis NTU-1 為一株可降解碳數 C6~C32烷類的菌株 (張緯農 2009)。其中,在降解長碳鏈烷類時,NTU-1 能形成特殊的聚集現象將 碳氫化合物包覆於結塊顆粒內,因而我們能在較短時間內配合生物降解及物理撈 除的方法將碳氫化合物污染移除。延續之前實驗室人員的研究,本論文的目的在 於探討如何加速NTU-1 的細胞聚集現象,並將其應用在碳氫化合物的移除上,
使石油碳氫化合物在短時間內即能達到良好的移除效果。接下來論文章節的編排 及內容大致說明如下:
第二章:介紹石油污染復育的相關研究基礎背景及發展,並針對微生物攝取 代謝碳氫化合物的方法途徑及菌株的特性…等相關的文獻作個回顧。
第三章:列出實驗中所使用之材料、儀器、實驗流程及配製方法。
第四章:呈現研究的成果並進行討論。內容包含:不同搖晃培養對於 NTU-1 生物降解及包覆正十六烷能力的影響及改進方法。測試以NB 培養的 NTU-1 經過冷凍乾燥以後,細胞的存活率及降解包覆能力是否受到影響。了解以 NB 培養的 NTU-1 經冷凍乾燥的存活率及降解能力以後,接著利用正十六烷 培養的NTU-1 細菌結塊進行冷凍乾燥,並同樣探討凍乾後的存活率及降解 包覆能力。最後利用烘乾的方式來取代冷凍乾燥,測試烘乾後的NTU-1 結 塊對於正十六烷的包覆移除效果為何。
第五章:將論文內容作個總結
3
第二章 文獻回顧
本研究的目的在於探討生物復育對於石油碳氫化合物污染移除的應用,故在 這一章節中將介紹相關研究的基礎背景及發展。首先會對各種石油碳氫化合物的 移除方法作個簡介與比較。接下來探討生物復育的應用,針對微生物攝取及代謝 碳氫化合物的方法、途徑還有相關的文獻回顧,並對本實驗中所使用的菌株 Rhodococcus erythropolis 的特性及相關研究作個簡介。此外,我們也會探討文獻 中微生物聚集現象發生的原因和冷凍乾燥及烘乾對於微生物造成的影響及其相 關應用。
2.1 石油碳氫化合物簡介及其對環境和人類之影響
石油 (petroleum) 是存在於地底下易燃的一種黏稠、深褐色液體,由一連串 複雜、分子量不一的碳氫化合物及其他元素如硫、氮、氧、微量金屬所組成。石 油碳氫化合物一般以三種形態存在,含三個碳以下的碳氫化合物如甲烷、乙烷、
丙烷以氣態存在,又稱天然氣;液態石油則為含碳量介於4~30 之間的碳氫化合 物;含高碳量的固態石油又以石臘及瀝青為主 (黃武良 1999)。若以石油碳氫化 合物的分子結構來分類,又可將其分成烷烴、烯烴、環烷烴、芳香烴及瀝青稀…
等,其組成隨著不同油田的開採而有不同的分佈。
十九世紀的石油工業發展緩慢,到了二十世紀初內燃機的發明開始產生轉變,
至今石油仍是最重要的內燃機燃料。而石油工業逐的漸蓬勃發展,與人類的生活 緊密的結合,舉凡人類的食衣住行、娛樂…等皆與石油脫離不了關係,例如石化 原料產品、化學肥料、人造纖維、合成樹脂及塑料產品…等。
4
石油帶給人們便利的生活,但也因為不當且過度的利用,造成對環境嚴重的 污染。石油對環境產生的污染主要是來自於油田開採、煉製、運送及使用的過程 中進入土壤、水體或空氣中而造成 (Allard and Neilson 1997, Atlas and Bartha 1992)。儘管目前世界各國的環保意識高漲,對於石油相關開發、工廠的廢水及 廢氣排放,都有嚴格的管制,然而還是會有無法控制、預期外的意外發生。以下 舉出近三年來發生的石油污染事件: 2009 年 8 月,澳洲西艾特拉斯 (West Atlas) 鑽井平台發生大規模的漏油事故,造成120 至 900 萬加侖的石油洩漏,總面積超 過 6000 平方公里。美國最嚴重漏油事件則發生在 2010 年 4 月,英國石油公司 在美國墨西哥灣租用的鑽井平台“深水地平線”發生爆炸,造成 1500 米深海的原 油洩漏,約2 億加侖原油流入墨西哥灣,嚴重破壞沿岸和海洋生態環境 (Camilli et al. 2010)。另外,2010 年 7 月中國大連新港石油管道爆炸,估計有 6 萬至 9 萬 噸原油流入了海洋。這些嚴重的漏油事件造成的污染不僅會影響岸上和海洋生物 的生態系統,並會進入空氣和土壤,需要相當長的時間才能復原。
有鑑於石油污染日益嚴重,造成人類和動物的傷害也不可輕乎。石油燃燒產 生的硫氧化物會嚴重污染大氣且引發人類呼吸道疾病;石油污染土壤後,會導致 石油烴的某些成分在糧食中累積,並通過食物鏈危害人類健康;當石油流入水體 中其可能會黏附在魚類、藻類上造成海洋生物死亡,使水產品品質下降、降低水 體的利用價值。這些石油污染若進入人體中,會影響體內的新陳代謝、免疫系統、
生殖系統甚至產生致命的疾病。因而人們除了積極的開發無污染能源外,也努力 的尋找有效處理石油碳氫化合物污染的方法,目的就是希望維護環境及人們的健 康,使人類能夠永續生存。
5
2.2 處理石油碳氫化合物污染之方法
自然界中,石油碳氫化合物造成的污染主要發生在土壤、海洋或地下水,若 污染濃度較低時,大自然通常具有自淨能力,能透過大氣、水流的擴散,或是光 分解、氧化及微生物的分解作用…等等的物理、化學或生物方法,將污染物轉化 為無害物,使環境中污染濃度降低並回復到原始的狀態 (Golwer 1983)。然而,
如果這些石油污染物濃度過高,超過自然界能負荷的限度時,則會造成環境永久 性的污染。因此,以人工移除石油碳氫化合物的方法漸漸產生且變得重要。
人工復育石油碳氫化合物的方法有很多,若依其處理機制來分大約可分為三 種,物理處理法、化學處理法及生物處理法。以下將為這三種方法作個簡介。
(1) 物理處理:
傳統的物理處理法只是使污染物在氣態、液態及固態相三相間轉換,故難以 將污染物徹底移除。其適用於處理簡單的污染物,耗時較短,主要是藉由高溫或 是電能來破壞污染物的結構,然而其需要大量熱能或電能,使成本也相對較高。
另外,若是對毒性物質以氣化的方式來處理,易形成氣態污染物進入大氣中,造 成空氣的污染,故物理處理法不適用於毒性高的污染物。常見的物理處理法如:
熱處理、熱脫附、焚化、清洗土壤、揮發、固化/安定化、氣提、吸附、結晶、
密度分離、沈降、過濾、透析/電透析、逆滲透…等。
(2) 化學處理:
利用化學反應將污染物的毒性降低或是將污染物轉變成無害的物質,適用於 須立即處理或是迅速降低污染物毒性的情況。然而污染物大多為混合物,故難以 找到能同時處理多元污染物的化學藥劑,使處理上較麻煩且成本高。且在進行化
6
學反應過程中,轉化率很難達到100%,易造成原來污染物的殘留,反應中可能 也會產生其他有毒性的副產物。常見的化學處理法有:化學反應、酸鹼中和、化 學催化、氧化還原、化學萃取、溶劑萃取、蒸氣萃取、化學沈澱…等。
(3) 生物處理:
主要是利用微生物分解轉化污染物,將污染物予以礦化或轉化成危害性較低 的其他物質 (Korda et al. 1997, Vidali 2001)。近年來生物處理法逐漸取代物理、
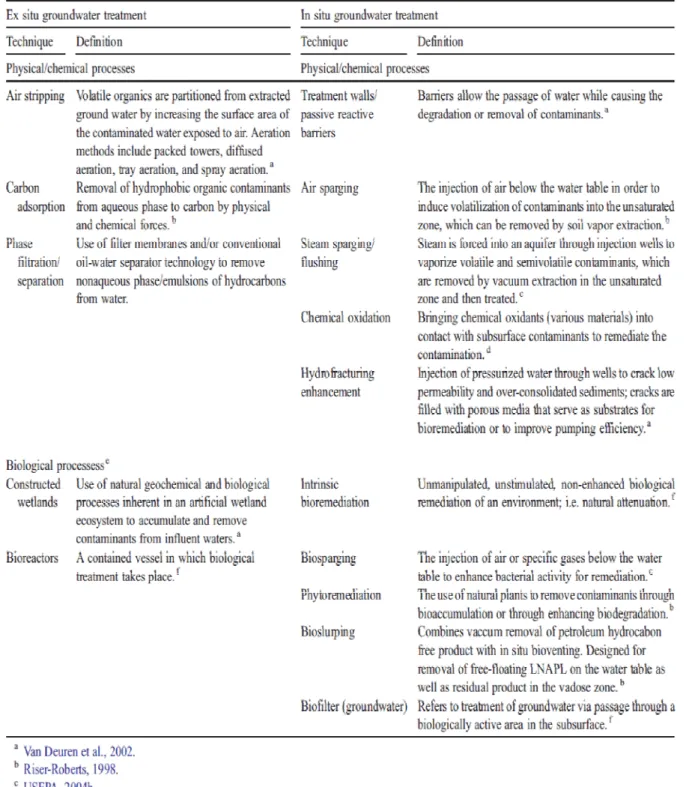
化學處理法成為最常見的石油污染復育法,主要是因為微生物種類眾多,可處理 不同分子結構的碳氫化合物,較不會產生其他有害物質,使用起來較安全,成本 也較其他兩種方法來得低。然而由於微生物培養時間相對較長,所以有耗時的缺 點。表2.2-1 為處理地下水碳氫化合物污染時,各種物理、化學處理及生物復育 技術的簡介及比較 (Van Stempvoort and Biggar 2008)。更詳細的生物處理法將在 2.3 節介紹。
7
表2.2-1 地下水污染之各種物理化學處理及生物處理程序之簡介與比較 (Van Stempvoort and Biggar 2008)。
8
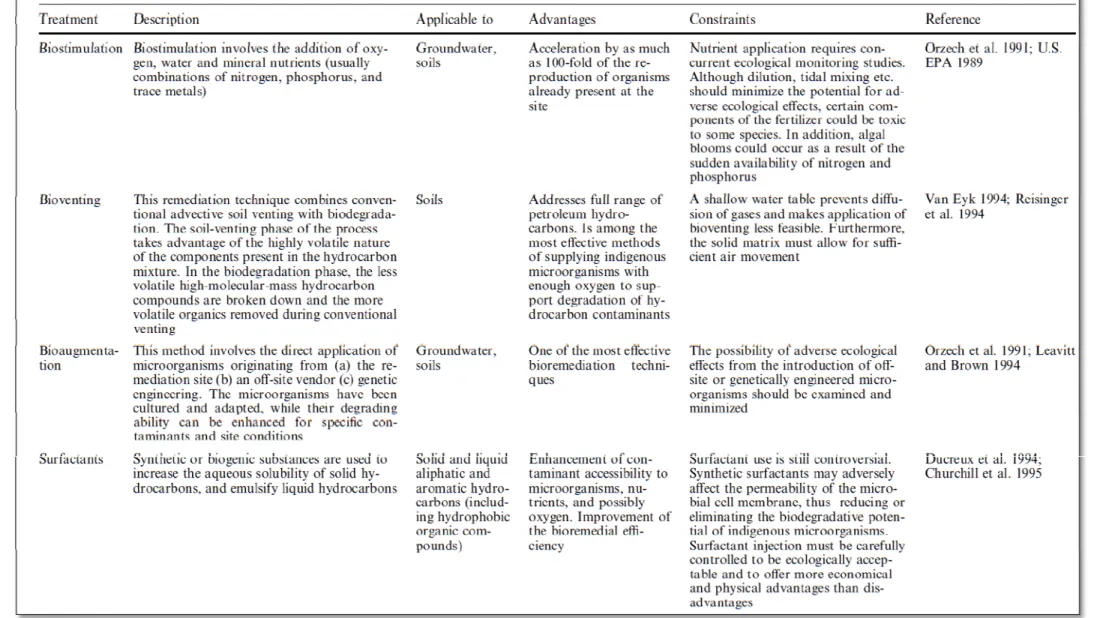
由於每個污染的場址通常會有不同的範圍大小、特性及限制,所以很多時候 會採取混合技術來處理,結合每種方法的長處使復育的效果達到最好。此外,我 們除了了解不同人工復育石油碳氫化合物的方法以外,如何選擇適當的方法來處 理不同情況的污染源也是值得注意的地方,許多因子都是在選擇合適的復育方法 時需考量的。以下列出幾個重要的影響因子 (Vik et al. 2001, Ram et al. 1993):
(1) 污染物及污染場址的特性 (2) 復育工作的驅動因素及目的 (3) 風險管理及環境的永續發展 (4) 投資者的看法及成本效益 (5) 技術上的適用性及可行性
以上幾個影響因子中,最主要考量的還是技術上的適用性及可行性,因為不 同的復育技術在不同污染地區的執行成果也會有所差異。且近年來許多復育的技 術逐漸的被發展出來,如何確認所選擇的復育技術是否合適則是需要經過有系統 及實驗證明的評估程序,表2.2-2 則列出幾個對於復育方法適用性的一般考量因 素 (Vik et al. 2001)。
表2.2-2 復育方法適用性與否的影響因素 (Vik et al. 2001)。
9
2.3 生物處理石油碳氫化合物
2.3.1 生物復育簡介
生物處理法在2.2 節中有稍稍作了簡介,這一節中我們將詳細的介紹生物處 理法中的優缺點、微生物的種類、特性,及常現生物處理的方法。生物復育 (bioremediation) 指的是利用微生物來進行有機污染物的代謝分解,使這些有機 污染物轉變為分子結構較簡單、毒性較低的物質 (Madsen 1991)。
生物復育逐漸取代傳統的物理及化學處理法的原因,主要因為生物復育法有 以下幾個優點 (Van Stempvoort and Biggar 2008):
(1) 相較於物理化學處理法來說,微生物培養容易,成本較低。
(2) 生物復育法使用上來說較安全且對環境影響較少。
(3) 微生物種類眾多,能處理各種石油碳氫化合物。
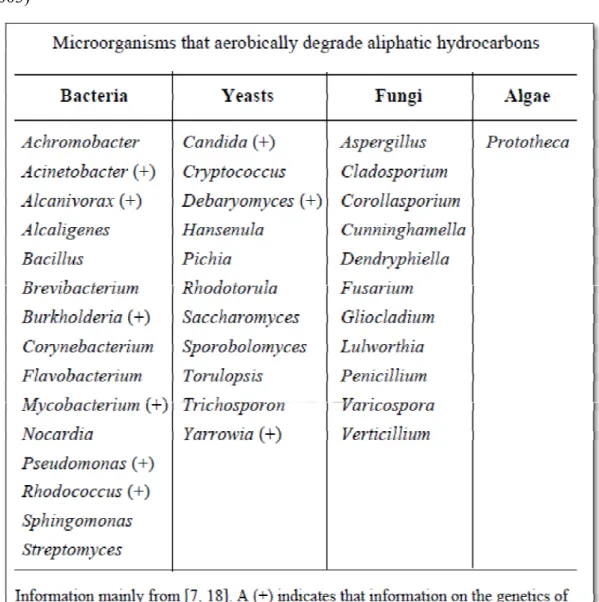
雖然自然界中微生物種類繁多,並不是每種微生物皆具有分解有機污染物的 特性。不過隨著生物科技的進步,微生物復育法已被大量研究及開發,利用生物 復育法來處理石油污染物的成功例子也越來越多,這些能分解石油污染物的微生 物以細菌、放線菌、酵母菌、真菌或藻類為主,表2.3-1 為一些能夠利用石油碳 氫化合物之好氧性微生物 (Van Beilen et al. 2003)。而一項成功的微生物復育通常 都需具備以下條件 (Bouwer and Zehnder 1993):
(1) 能分解污染物,使其失去毒性或使濃度降低。
(2) 能將污染物濃縮,並配合物理法或化學法將污染物完全移除。
(3) 可處理不同種類之混合污染物。
(4) 不會破壞環境平衡或對人體造成傷害。
(5) 具有好的再現性且易於培養、價格低廉。
10
表2.3.1-1 自然界中能利用石油碳氫化合物之好氧性微生物分類 (Van Beilen et al.
2003)。
在實際應用上,生物復育技術又可分成兩大類:現地 (in situ) 生物復育及 離場 (ex situ) 生物復育。接下來為這兩類技術作個簡單的說明及比較。
(1) 現地生物復育:
在受污染的區域直接進行生物復育。處理污染土壤、地下水或海洋時,不需 先將土壤或水質取出移到其他地方再進行生物復育。一般工程化的生物復育技術,
主要會採行下列幾種方式:添加微生物、添加營養鹽、添加電子提供者,如甲烷
11
及丙烷等有機物來進行共代謝作用、添加電子接受者,例如加入氧氣或過氧化氫 以進行好氧分解 (盧至人 2002)。表 2.3.1-2 為一些常見實際應用的現地生物復育 法 (Korda et al. 1997)。
現地生物復育法主要的優點為工程上難度不高、符合自然環境特性、可分解 被吸附的污染物、地表設施較少、設備易護得、相對來說較經濟…等 (盧至人 2002)。然而可能會遭遇到的問題有場址的限制、自然界的因素如天候、植入的 菌株是否經實驗證明其表現會比原始存在於污染區的菌株來得好、是否需要多種 類的菌株才能完全處理這些污染物、處理時間過長及當污染物移除後,這些菌株 將如何處理 (Bartha and Atlas 1977, Boopathy 2000)。
(2) 離場生物復育:
離場生物復育技術是指在污染源擴散前,挖出污染土壤或抽出污染的水體,
帶離所在地做處理 (Riser-Roberts 1998)。依照不同微生物和污染物的交互作用會 有不同的反應器設計,又可分為污泥相的生物復育及固相的生物復育。污泥相的 生物復育主要用來處理以黏粒為主的土壤。而固相生物復育又可分成地耕法、生 物堆土法和堆肥法。現地與離場生物復育法的比較可參照2.2 節中的表 2.2-1。
離場生物復育法的優點為處理較快速、易控制反應的條件且能處理多種污染 物及土壤種類。而其缺點主要為搬運污染物及反應器或其他設備的支出造成投資 成本較高。
12
表2.3.1-2 實際應用的現地生物復育法 (Korda et al. 1997)。
13
表2.3.1-2 (續)
14
2.3.2 微生物攝取碳氫化合物之模式
雖然現今生物復育技術的技術越來越多且更加成熟,但微生物攝取石油碳氫 化合物污染時仍面臨許多的挑戰,主要原因有以下三個 (Abdel-Megeed 2004):
(1) 石油污染物通常都是複雜的混合物。
(2) 較低分子量的碳氫化合物因為溶劑效應 (solvent effects),破壞微生物細胞膜 的調結功能,對細胞膜具有相當的毒性。
(3) 大部分的石油碳氫化合物在常溫下為液體或固體皆屬於難溶於水的物質。
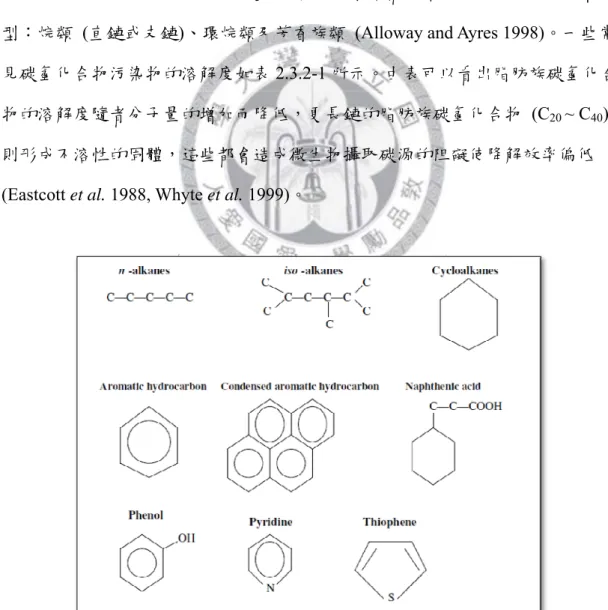
圖2.3.2-1 為原油中常見的碳氫化合物化學結構分類,其大致可分為三種類 型:烷類 (直鏈或支鏈)、環烷類及芳香族類 (Alloway and Ayres 1998)。一些常 見碳氫化合物污染物的溶解度如表2.3.2-1 所示。由表可以看出脂肪族碳氫化合 物的溶解度隨者分子量的增加而降低,更長鏈的脂肪族碳氫化合物 (C20 ~ C40) 則形成不溶性的固體,這些都會造成微生物攝取碳源的阻礙使降解效率偏低 (Eastcott et al. 1988, Whyte et al. 1999)。
圖2.3.2-1 原油中常見的碳氫化合物化學結構分類 (Alloway and Ayres 1998)。
15
表2.3.2-1 各種碳氫化合物在水中的溶解度 (Eastcott et al. 1988)
儘管微生物在利用石油碳氫化合物時,可能會遇到上述三點的挑戰,但已有 許多能利用溶解性低或具毒性碳氫化合物的微生物復育技術被研究出來,主要是 因為這些微生物能夠以不同方式來攝取低溶解性的碳氫化合物,我們可將其歸類 成以下三種模式。這些模式的重點在於低溶解性的碳氫化合物如何由外界傳送及 累積到微生物的表面,接著進入到微生物內部而進行下一步代謝利用的動作 (Hommel 1990)。
(1) 微生物直接攝取溶於水中的碳氫化合物。
Hydrocarbons Class Mol. wt. (g/mol.) Solubility (mg/L) at 25℃
n-Pentane (C5H12) normal alkane 72.2 40.8 n-Hexane (C6H14) n-alkane 86.2 12.3 n-Octane (C8H18) n-alkane 114.2 0.682 n-Dodecane (C12H26) n-alkane 170.3 3.4×10-3 n-Tetradecane (C14H30) n-alkane 198.4 6.55×10-4 n-Hexadecane (C16H34) n-alkane 226.4 5.21×10-5 n-Octadecane (C18H38) n-alkane 254.4 4.05×10-6
Pristane (C19H40) branched-alkane 268 insoluble
n-Tetracosane (C24H50) n-alkane 338 insoluble n-Dotriacontane (C32H66) n-alkane 450 insoluble
Naphthalene 2-ring aromatic 128.2 30.6
Pyrene 4-ring aromatic 202.3 0.135
Chrysene 4-ring aromatic 228.3 <0.01
16
(2) 微生物分泌界面活性劑使碳源乳化成微小油滴(油滴顆粒小於微生物細胞) 進而攝取利用。
(3) 微生物直接接觸大油滴(油滴顆粒大於微生物細胞) 並利用降解。
第一種攝取模式中,主要發生在低分子量的碳氫化合物上。由於低分子量碳 氫化合物的水溶性較高,能確保有一定的質傳速率能傳送到細胞表面讓微生物直 接攝取 (Rojo 2009)。故此模式的降解代謝速率取決於碳氫化合物溶在水中的速 率、濃度及微生物攝取的速率。生物復育發展初期,學者認為只有溶於水中的碳 氫化合物能被細菌所利用 (Gibson 1984)。然而許多學者發現低溶解性碳氫化合 物的降解速率比溶於水相中的碳氫化合物來得高,這樣的結果指出有其他的機制 使微生物能順利攝取低溶解性的碳氫化合物 (Leahy and Colwell 1990)。
第二項機制中主要是與微生物釋放出的生物界面活性劑有關。大部分能降解 石油碳氫化合物的菌株都被研究出其在不同環境下會釋放出生物界面活性劑 (biosurfactant),將疏水性化合物乳化成微小油滴,增加微生物的接觸面積,使降 解速率提高 (Desai and Banat 1997, Rosenberg and Ron 1999)。這個形成微小油滴 的過程又能稱為假溶解 (pseudosolubilizaion),意即形成微胞粒子 (micelles)。界 面活性劑的分子包含親水區及疏水區,當濃度很低時,界面活性劑可完全溶於水 中,隨著濃度上升則會聚集形成微胞粒子。微胞粒子能將疏水的碳氫化合物包覆,
並以親水端將其帶入水相中。
圖2.3.2-2 為碳氫化合物被生物界面活性劑附著後,微生物細胞攝取碳氫化 合物的示意圖 (Ratledge 1988)。圖 2.3.2-3 則為菌株 Pseudomonas sp.以碳氫化合 物為培養碳源時,分泌出生物界面活性劑 (rhamnolipids),與碳氫化合物形成微 胞粒子而被微生物細胞攝取圖 (Fritsche and Hofrichter 2000)。此外,學者發現這
17
些生物界面活性劑還可能會鍵結在微生物表面上,提高微生物表面的疏水性 (Hua et al. 2003, Mulligan 2005)。
目前研究中指出,微生物在碳氫化合物環境下產生的生物界面活性劑主要有 以下幾類 (Niescher et al. 2006):醣脂類 (glycolipids)、脂肽類 (lipopetides)、磷 脂類 (phospholipids)、多醣脂類 (liposaccharides)、脂肪酸類 (fatty acids)、高分 子界面活性劑 (polymeric surfactant) …等。表 2.3.2-2 為各種微生物產生的生物界 面活性劑分類 (Al-Araji et al. 2007)。
最後一種碳氫化合物的攝取方式,微生物能夠貼附於水相與固態或液態疏水 性碳氫化合物的界面上,並將其攝取進入細胞內並加以利用 (Volkering et al.
1997)。這些微生物除了本身表面已經夠疏水外,有些微生物會為了促進與疏水 性化合物的黏附性,在利用碳氫化合物時改變細胞膜表面的成分 (脂質或脂肪酸) 來提高細胞表面的疏水性 (Morgan and Watkinson 1994),例如上述的第二種機制 中,微生物會產生生物界面活性劑鍵結在細胞膜表面。
以上三種攝取機制說明了大多數微生物與碳氫化合物接觸攝取的過程,許多 研究指出這三種機制常常同時存在,或是有兩個同時存在,若單獨只有某一種機 制存在時,我們通常會添加界面活性劑來提高降解的效率。石油碳氫化合物污染 中,大部分的成份為溶解性低的長碳鏈烷類,故微生物多採取第二種和 (或) 第 三種機制來接觸攝取。
18
圖2.3.2-2 微生物細胞攝取有生物界面活性劑附著的碳氫化合物示意圖 (Ratledge 1988)。
圖2.3.2-3 菌株 Pseudomonas sp.分泌界面活性劑後攝取碳氫化合物示意圖 (Fritsche and Hofrichter 2000)。
19
表2.3.2-2 各種微生物產生的生物界面活性劑分類 (Al-Araji et al. 2007)。
20
2.3.3 微生物分解碳氫化合物之方式
自然界中,能夠利用碳氫化合物的微生物種類眾多,也各自擁有獨特降解碳 氫化合物的方式。這些微生物能夠把石油碳氫化合物當作主要碳源,經過上一節 介紹的三種攝取方式將碳源傳送到細胞內並進行代謝降解。在降解過程中,藉由 氧化還原反應的電子轉移來生成能量提供微生物合成蛋白質、脂質或建構新的細 胞…等。接下來我們介紹微生物降解碳氫化合物時較常見的氧化還原方式—好氧 性及厭氧性降解。
(1) 好氧性降解 (aerobic degradation):
好氧性的細菌以氧分子作為最終電子接受者,利用氧化反應將碳氫化合物從 非極性的基質轉化成脂肪酸並進行後續的代謝作用生成細胞生長所需的化合物 及能量 (Wentzel et al. 2007)。圖 2.3.3-1 為好氧性微生物降解碳氫化合物的主要 流程 (Fritsche and Hofrichter 2000)。文獻中也指出,在好氧的情況下,大部分碳 氫化合物能以較快的速率被微生物完全降解 (Riser-Roberts 1998)。不同種類的碳 氫化合物會有不同的代謝途徑,將於2.4 節中詳細介紹。
(2) 厭氧性降解 (anaerobic degradation):
與好氧性降解不同的是,厭氧性降解時電子接受者不是氧分子,而是一些無 機物質如硫酸根、硝酸根、鐵離子、錳離子或二氧化碳…等。近年來,有許多關 於利用硫酸鹽還原菌 (sulfate-reducing bacteria) (Rueter et al. 1994, So et al. 2003, Cravo-Laureau et al. 2005) 及反硝化細菌 (denitrifying bacteria) (Ehrenreich et al.
2000) 在無氧環境下進行長碳鏈烷類氧化反應的研究被報導出來。在這些微生物 系統中,硫酸鹽及硝酸鹽作為最終電子接受者,其角色相當於好氧性分解中的氧 分子。目前碳氫化合物無氧代謝的初始反應現今主要有兩種不同的機制被提出,
21
分別是延胡索酸鹽 (fumarate) 結合反應、羧化反應 (carboxylation) (Wentzel et al.
2007)。好氧性及厭氧性細菌降解長碳鏈碳氫化合物過程及比較如圖 2.3.3-2 所 示。
一般來說,碳氫化合物的厭氧性分解速率較低,主要是因為厭氧菌的生長速 率較好氧菌來得慢。例如,厭氧菌Hxd3 及 Pnd3 在降解正十六烷時,細胞的倍 增時間約為9 天 (Aeckersberg et al. 1998);厭氧菌 AK-01 降解正十六烷時在最佳 化的培養條件下,細胞的倍增時間仍要3 天左右 (So and Young 1999)。
圖2.3.3-1 好氧性微生物降解碳氫化合物的主要流程 (Fritsche and Hofrichter 2000)。
22
圖2.3.3-2 長碳鏈烷類的好氧及厭氧性降解途徑 (點狀線的箭頭是指推測的代謝 路徑) (Wentzel et al. 2007)。
23
2.4 微生物降解碳氫化合物之代謝途徑
微生物利用碳氫化合物時,其降解難易度通常與污染化合物的分子立體結構 有很大的關係,結構越簡單的化合物越容易被分解;相同化學結構的化合物中,
分子量較低的化合物比分子量高的化合物來得容易被微生物利用降解,但碳鏈長 度差異在5 個碳以內則不會有太大的差別 (Huesemann 1995)。石油碳氫化合物 中,微生物利用降解的難易度排序大致如下:直鏈烷類 (normal alkanes)>支鏈 烷類 (branched alkanes)>支鏈烯烴類 (branched alkenes)>分子量較低的芳香族 類 (small aromatics)>環烷類 (cyclic alkanes)>多環芳香族碳氫化合物 (PAHs)
>>瀝青稀類 (asphaltenes) (Huesemann 1995, Ulrici 2000, Atlas and Bragg 2009)。
2.3.3 節中,我們知道了微生物利用碳氫化合物時,主要有好氧性及厭氧性 兩種氧化還原方式。由於本研究中所使用的菌株 Rhodococcus erythropolis NTU-1 為好氧性細菌,故接下來我們主要介紹碳氫化合物的好氧性代謝途徑。
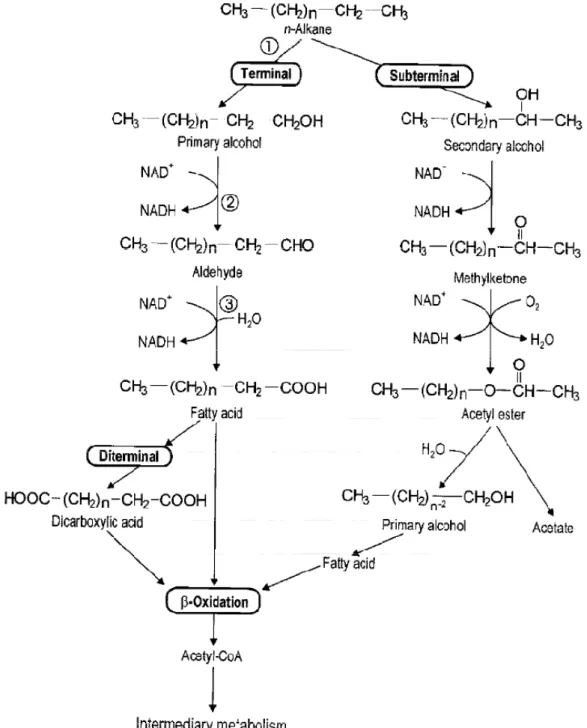
2.4.1 直鏈烷之氧化機制
直鏈烷類 (n-alkanes) 為原油 (crude oil) 污染物中最主要的成分之一
(Harayama et al. 1999)。其中,長碳鏈的直鏈烷類 (C10 ~ C24) 為微生物最常利用 的基質,降解速率最快(Bartha and Atlas 1977);短鏈烷類 (碳數小於 9) 則對於多 種微生物具毒性,會破壞微生物細胞膜的通透及調結功能,但其能以較快的速率 從石油污染物中揮發移除 (Fritsche and Hofrichter 2000)。這一部分我們主要針對 長碳鏈直鏈烷類的代謝途徑來作介紹,直鏈烷類的氧化途徑主要可分為三類:單 末端氧化 (monoterminal oxidation)、雙末端氧化 (diterminal oxidation)、次末端氧 化 (subterminal oxidation),如圖 2.4.1-1 所示 (Gibson 1984)。
24
(1) 單末端氧化 (monoterminal oxidation):
單末端氧化為微生物代謝直鏈烷時最主要的途徑 (Fritsche and Hofrichter 2000)。直鏈烷氧化過程中,微生物釋放出單氧化酶,利用 NADH2作為電子提供 者並結合氧分子作為電子接受者,攻擊直鏈烷末端的碳原子,進而形成一級醇類。
接著再利用醇脫氫酶 (alcohol dehydrogenase) 將醇氧化成醛類,最後微生物釋放 醛脫氫酶 (aldehyde dehydrogenase),利用 NADH2再次結合氧分子,將醛氧化成 長鏈脂肪酸 (fatty acid)。長鏈脂肪酸會結合 CoA 並進入 β-oxidation 進而生成 acetyl-CoA (Watkinson and Morgan 1990, Van Hamme et al. 2003, Wentzel et al.
2007),如圖 2.4.1-1。這些長鏈脂肪酸還有可能會與細胞膜上的脂質 (lipid) 結合,
但主要還是會進入檸檬酸循環 (TCA cycle) (Fritsche and Hofrichter 2000)。
(2) 雙末端氧化 (diterminal oxidation):
直鏈烷氧化過程中,有些微生物除了分泌單氧化酶外,還會分泌出雙氧化酶,
使烷類兩端的甲基同時被氧化 (ω-oxidation) 成雙質子醇,經過一連串的氧化反 應形成長鏈雙質子羧酸 (dicarboxylic acid),最後進入 β-oxidation,此代謝途徑也 可視為單末端氧化過程中的副反應 (Watkinson and Morgan 1990, Coon 2005),如 圖2.4.1-1 所示。
(3) 次末端氧化 (subterminal oxidation):
如圖2.4.1-1 的右半邊圖所示。次末端氧化發生於烷類基質的鏈長至 C16 (Gibson 1984, Whyte et al. 1998, Binazadeh et al. 2009) 或短碳鏈烷類 (C3 ~ C6) 中(Ashraf et al. 1994)。也可稱之為 α-oxidation,主要是氧化直鏈烷中 α 位置的碳,
首先形成二級醇類,接著將其氧化成酮類,最後形成一級醇及醋酸鹽。其中,一 級醇氧化成為脂肪酸後進入β-oxidation,繼續反應成細胞所需的化合物,而醋酸 則進入TCA cycle 產生能量。一些微生物中,單末端及次末端氧化反應能夠同時 存在 (Rojo 2009)。
25
圖2.4.1-1 微生物代謝分解直鏈烷之途徑 (圖中 1 為 n-alkane 單氧化酶、2 為醇 脫氫酶、3 為醛脫氫酶) (Britton 1984)。
26
2.4.2 支鏈烷之氧化機制
支鏈烷類 (branched alkanes) 相對於直鏈烷類較難以被微生物降解,且支鏈 越多的烷類相較於結構較簡單的支鏈烷類更加頑抗 (recalcitrant),特別是化合物 中含3 級或 4 級的碳原子結構,其空間障礙會影響氧化酶的作用,故具有降解支 鏈烷能力的微生物相對較少 (Britton 1984)。
一般來說,異十九烷 (pristane or 2, 6, 10, 14-tetramethylpentadecane) 常被當 作微生物降解支鏈烷類的指標,主要是因為其複雜的支鏈結構往往在原油或石油 污染物的降解過程中呈現穩定的存在而不易被分解 (Morgan and Watkinson 1994)。然而具有降解異十九烷能力的微生物早在許多研究中已被證實,例如 Brevibacterium sp. (Pirnik et al. 1974)、Corynebacterium sp. (McKenna and Kallio 1971a)及 Rhodococcus sp. (McKenna and Kallio 1971b, Fraaije et al. 2002)…等。
圖2.4.2-1 為 Pirnik 學者提出的利用菌株 Brevibacterium erythrogens 降解異十 九烷的單末端氧化及雙末端氧化途徑 (Pirnik et al. 1974)。由於 3 級碳的阻礙,
異十九烷首先會經由α-oxidation 形成異十九酸 (pritanoic acid),接著進行 β-oxidation 形成 trimethyltridecanoic acid、trimethylundodecanoic acid 或是進形 ω-oxidation 形成 trimethyltridecandioic acid 後再進行 β-oxidation,直到異十九烷 被分成成琥珀酸 (succinic acid) 並進入 TCA cycle 為止。
27
圖2.4.2-1 菌株 Brevibacterium erythrogens 降解異十九烷的途徑 (Pirnik et al.
1974)。