國 立 交 通 大 學
生 物 科 技 學 院
生 物 科 技 學 系
碩 士 論 文
微藻養殖生產油脂並利用微藻油脂產製生質
柴油之研究
Study on the Lipid Production from Microalgal Cultures and
Producing Biodiesel from the Microalgal Lipid
研 究 生:蔡 明 達
指導教授:林 志 生 博士
微藻養殖生產油脂並利用微藻油脂產製生質柴油之研究
Study on the lipid production from microalgal cultures and producing
biodiesel from the microalgal lipid
研 究 生:蔡明達 Student:Ming-Ta Tsai 指導教授:林志生 Advisor:Chih-Sheng Lin Ph.D. 國 立 交 通 大 學 生 物 科 技 學 院 生 物 科 技 學 系 碩 士 論 文 A Thesis
Submitted to Department of Biological Science and Technology
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master in Biological Science and Technology
July 2009
Hsinchu, Taiwan, Republic of China 中 華 民 國 九 十 八 年 七 月
致 謝
兩年的碩士班生活在實驗、課業與疑惑三部曲的交錯唱和下謝幕了,摻雜其中的 汗水、淚水與歡笑如劃開暗夜的流星般在驚嘆中一瞬即逝,但此情此景卻如滾燙的烙 印默默燒存在我腦海深處,不會忘記的是那一份份的感動與感激。最要感謝的是我的 指導教授林志生老師,宛如嚴父的外表下我卻能體會那如慈母般柔軟的心,再多的教 誨與叮嚀皆因恨鐵不成鋼,不僅教導我們實驗的技巧與思考能力,更身教言教的告訴 我們面對人生應有的態度與精神,讓我更能了解翩然到來成功的勝利女神背後是奮力 不懈與永不放棄的執著。雖然愚生駑頓與粗心,但我相信在社會上遭遇困難之時,猶 言在耳的字句珠磯定能給我不同於此時的深層體會。 時光的短暫與漫長總是困擾著凡人,而與實驗室夥伴相處的快樂時光總是令人覺得 須臾瞬間。感謝有藻類組老大聖壹學長降臨人間,有如久旱逢甘霖般解決我疑惑的心 與殘破的知識,更教導我許多實驗的技巧與小撇步,沒有他的協助,我就如鯉魚離開 了水一般痛苦不能呼吸,有了他,不再唇亡齒寒,實驗室生活是彩色的。而千雅同學 (偽學姊),教導了我許多的實驗技術外,更讓我學會怎麼運用「霸氣」,讓我在擊倒 敵人時不再猶豫。藻類組的另一個大姊筱晶,感謝你陪我度過Subculture 的每一天, 戶外的驕陽如炙也讓你我擁有用之不竭的維他命D與健康的古銅色皮膚。此外還要感 謝思豪學長的慷慨解囊與疼惜學弟,感謝柔情鐵漢的棠青學長為實驗室奉獻無私的 愛,感謝高科技農夫曜禎學長的甜美果實呵護著我的腸胃。感謝証皓同學秀麗的臉龐 與榕均同學挺拔的英姿陪伴我一起度過實驗室艱苦難熬的日子。感謝學妹四朵花:子 慧、小媽、小韓、小魚的加入,讓實驗室不再陽氣鼎盛而柔美增色不少。也感謝畢業 的建龍學長、俊旭學長與宜貞學姊,在我孤立無援的孱弱背影前不吝伸出熱切溫暖的 援手。 最後,對於撥冗前來參加口試的委員曾慶平教授、李唐教授與林昀輝教授獻上萬分的 敬意與感恩,感謝您們的建議與提點。在此也祝福各位不管在研究、學業或工作上都 能得心應手、無往不利而步步高昇! 蔡明達 謹致 于 2009 年交通大學微藻養殖生產油脂並利用微藻油脂產製生質柴油之研究
研究生:蔡明達 指導教授:林志生 博士 國 立 交 通 大 學 生 物 科 技 學 院 生 物 科 技 學 系 碩 士 論 文中 文 摘 要
近年來,全球面臨地球暖化危機與石化能源耗竭兩大危機。由於人類大量開採石化 能源及發展工業化社會導致溫室氣體CO2大量累積與石化燃料枯竭之嚴重問題。因故低 污染的再生能源與CO2減量之策略發展為世界各國所積極研究的議題。然而以海洋微藻 培養之利用正是一項高效益的綠色能源開發方法。微藻為能行使光合作用之單細胞植 物,能快速且大量生產植物生物質與累積大量油脂於微藻細胞中。有鑑於此,本研究之 目的為以高脂海洋微藻利用廢氣CO2培養以進行大量油脂生產並藉由轉酯化反應生成 生質柴油之研究。在最佳培養狀況下所篩選海洋微藻生長效率可達一天微藻增生2 至 3 倍。微藻培養時 Chlorella sp.與 N. oculata 之微藻內油脂從生長對數期至生長穩定期,培 養狀態進行至氮源缺乏時,可由12%與 21%分別提升至 21%與 50%。然而以半連續式 微藻培養於光生物反應器,通入2% CO2之半連續式微藻光生物反應器中,其生長能力 與產脂率表現最佳,氮在不同濃度CO2培養下(2 至 15%),高濃度 CO2對於微藻的生 長與產脂量能保持穩定並不受高濃度CO2的抑制。然而微藻也能利用不同的有機碳源在 混營或異營脂培養狀態下進行生長與油脂堆積。本研究指出,以蔗糖行異營培養之 N. oculata 其油脂累積雖可達 54%,但其生物質產量有降低的情形,而在以蔗糖進行混營之 培養下油脂產量可提昇至每公升 0.284 公克。因此微藻培養時可分為兩個階段:增值階 段及肥育階段,先利用最適生長環境來快速培養增加微藻的細胞濃度,再將其轉入低硝 酸鹽或以蔗糖混營培養環境中肥育,即可有效率的大量產脂。本研究中,微藻油脂轉酯化生成生質柴油因其轉化方式可分為化學製程以及生物製程 兩種,以最佳油醇莫耳比下,酸化學催化反應與微生物脂解酶催化反應可分別達到88% 與82%。酵素催化之轉化率雖佳,但其成本高且反應時間過長,在工業上多以快速簡便 之化學催化轉酯化反應達到快速、低成本且高產率的目的。本研究成果證明由藻類油脂 製成生質柴油作為再生燃料具可行性與未來性,且在藻類培養過程中能有效的降低環境 中的溫室氣體之危害並能提供快速且大量生物油脂之生產以提供生成生質柴油。 關鍵字: 微藻、小球藻、擬球藻、微藻油脂、生質柴油、轉酯化反應
Study on the lipid production from microalgal cultures and producing
biodiesel from the microalgal lipid
Graduate Student: Ming-Ta Tsai Advisor: Chih-Sheng Lin Ph.D. Department of Biological Science and Technology
National Chiao Tung University
Abstract
In recent years, people over the world face some acute problem with regard to global warming and energy crisis. Humans exploited the fossil energy and developed the industry and civilization well in past hundred years to result in the environmental problems,
green-house gas emission rising, and petrochemical fuel exhausting. Photosynthetic
organism, microalgae, can use solar energy efficiently to combine water with CO2 to produce
biomass. Microalgae can not only produce biomass but accumulate lipid in microalgal cells. Lipids from microalgae can be extracted and converted to biodiesel fuel by transesterification.
In the study, the biomass and lipid productivity of Chlorella sp. and Nannochloropsis
oculata were evaluated in the different conditions of culture in the closed photobioreactors.
Results showed that the lipid accumulation of Chlorella sp. and N. oculata from logarithmic phase to stationary phase were significantly increased from 12% to 24% and 21% to 50%, respectively. In the semiconscious culture of Chlorella sp. and N. oculata, the total lipid productivity was 0.143 and 0.142 (g/L/d), respectively although the cultures were daily replaced half of broth. The results showed that Chlorella sp. and N. oculata were potential candidates for biomass and lipid production by semicontinuous cultures. The comparison of lipid productivities in the semicontinuous systems in which the culture broth were replaced at an interval of 24 h (one-day replacement) or 72 h (three-day replacement) was performed.
The result indicated the total lipid yield in the semicontinuous culture operated by one-day replacement was more efficient.
Moreover, different carbon sources supplied in the culture could make distinct growing ability and lipid accumulation. The results showed the cultivation of N. oculata using
sucrose as carbon source in mixotrophic growth gave the highest maximum biomass (0.80 g/L) and the lipid productivity was up to 0.284 (g/L). Although the lipid contents from the
heterotrophic cultivations with sucrose could increase to 54%, the biomass productivity decreased in the heterotrophic cultures. Therefore, the recovery of microalgal cells from semicontinuous cultures can transfer to mixotrophic cultivation for higher lipid production. The fast analysis of lipid contents of live microalgal cells by Nile red staining under
fluorescence was established. The linear regression of fluoresce intensity of lipid content was established to measure the lipid contents of microalgae.
The typical transesterification method used in this study was chemical and enzymatic processes. The results indicated the efficiency of transesterification by acid catalyst could approach 88% and the efficiency by lipase-catalyzed transesterification could reach 82% by the optimal oil/methanol molar ratio. The high cost and long reaction time of enzymatic processes may be not convenient for usage. In industrial and commercial application, the chemical catalyst is common for transesterification to reach the purposes of low-cost and fast reaction rate. Therefore, the fast and suitable transesterification method in this study is acid-catalytic process.
Keywords: Microalgae, Chlorella sp., N. oculata, Microalgal lipid, Biodiesel, Transesterification
Content
Acknowledgment ……….………... i
Chinese Abstract ……….………… ii
English Abstract ……….……... iv
Content ……….…………. vi
List of Tables ………..……….…………... viii
List of Figures ………...………….……… ix
I. Literature Review ……….……… 1
1-1. Global warming and energy shortage ……….………. 1
1-2. Microalgae ………...……….. 4
1-3. Microalgae for reducing pollution and renewable fuels ……….………….... 5
1-4. Microalgae cultivation ………..……….……….…...… 6 1-4-1. Growth medium ……… 6 1-4-2. Nitrogen contents ……… 7 1-4-3. Light ……… 8 1-4-4. Temperature ……… 9 1-4-5. Carbon dioxide ……… 10 1-4-6. pH value ………. 11 1-4-7. Growth phase ……….. 12
1-5. Microalgae cultures techniques ………...……….………... 14
1-6. Mixotrophic and heterotrophic culture of microalgae ………....…….. ….…. 16
1-7. Acetyl-coenzyme A carboxylase of microalgae ……….……….…... 18
1-8. Extraction method of microalgae ……….……….…… 19
1-9. Biodiesel ……….…... 20
1-10. Biomass for biodiesel ……….…….……… 21
1-11. Biodiesel characteristic ……….…….……. 22
1-12. Method of biodiesel production ……….…….…… 24
1-12-2. Alcohol type and effect of alcohol molar ratio in transesterification …… 27
1-13. Biodiesel source and fatty acid composition ……….……….. 28
1-14. Biodiesel from microalgae ……….…….…… 29
1-15. CO2 Fixation and producing biodiesel using microalgal cultures ………….……. 31
II. Materials and Methods ………..………... 33
2-1. Microalgal strains …………..……….….……… 33
2-2. Microalgae inoculums …...……..………. .….…….... 33
2-3. Culture medium and nutrients .……….…………..…………....……... 34
2-4. Microalgae cryopreservation ………..……... 34
2-5. Experimental system with photobioreactors ……….……...… 34
2-6. Microalgal cell counting and biomass ………...………….…... 35
2-7. Growth rate of Microalgae ………...………...…...….……...….. 36
2-8. Measurement of nitrogen concentration in culture ……….…….. 36
2-9. Lipid analysis ………..……….…………. 37
2-9-1. Measurement of lipid content by lipid extraction ………. 37
2-9-2. Saponification and esterification for GC analysis ………. 37
2-9-3. GC analyzer ………... 37
2-9-4 Measurement of lipid content by fluorescent spectrometry ……… 38
2-10. Setup of semicontinuous culture system ………..……..……...…. 38
2-11. Autotrophic, heterotrophic, and mixotrophic growth in microalgal cultures ... 39
2-12. Effect of organic carbon on the growth ………..………...…. 40
2-13. Transesterification of microalgal oil ……….…... 40
2-14. RNA extraction of microalgae ……….…..…... 40
2-15. ACCase expression ……… 41
2-16. Parameter analyses ……….…..………. 42
2-17. Statistics ………..……….. 42
III. Results and Discussion ………...……. 43
3-1. Growth ability and lipid content of four potential strains ………...…….. 43
3-2. Cultivation with CO2 and lipid composition …..….……….…....…... 43
3-4. Effect of CO2 on biomass and lipid production …..…………..………... 44
3-4-1. Growth of microalgae aerated with different CO2 Concentration ………… 44
3-4-2. Effect of CO2 on cell growth in microalgal semicontinuous cultivation …. 46
3-4-3. Effect of CO2 on lipid and biomass production in semicontinuous cultures 47
3-5. Lipid content of microalgae at different growth phases and nitrate limitation ….. 49 3-6. Comparison of productive efficiencies in semicontinuous system with different
culture approaches ……….… 50 3-7. Biomass and lipid production of microalgae in mixotrophic and heterotrophic
cultivation with various carbon sources ..………..……. 51 3-7-1. Effects of organic carbons on the growth of N. oculata ……….. 51 3-7-2. Effects of inorganic and organic carbons on the growth of N. oculata …… 53 3-7-3. Lipid contents and production in N. oculata cultures with different carbon
sources ………. 53
3-8. Measurement of microalgae by fluorescence …..……….……….….. 54 3-9. Comparison between ACCase expression and lipid contents ………... 56 3-10 Transesterification by different catalyst types and methanol molar ratio ……….. 56 3-11 Biodiesel applied to the biodiesel car ……….... 57 IV. Conclusions ……… 58 V. References ……….…. 61
List of Tables
Table 1-1. Comparison between alkali-catalysis and lipase-catalysis methods for
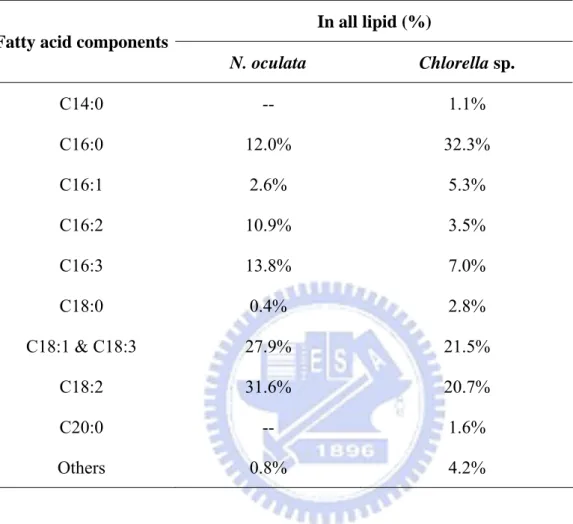
biodiesel fuel production ………...……….. 74 Table 1-2. Production of Biodiesel in Different Countries ………... 75 Table 1-3. Fatty acid composition of biodiesel obtained from different sources …….... 76 Table 1-4. Oil productivity of different energy plants …..………..… 77 Table 2-1. Table of trace elemental solution of f/2 culture medium ………..…. 78 Table 3-1. Lipid components of Chlorella sp. and Nannochloropsis oculata ……..….. 79 Table 3-2. Recovery of lipid and biomass productivity of the Chlorella sp. as waste
broth in the semicontinuous photobioreactor under different concentrations of CO2 aeration ……….. 80
Table 3-3. Daily recovery of biomass and lipid of N. oculata NCTU-3 cultured in the semicontinuous system aerated with different concentrations of CO2 ….…. 81
Table 3-4. Biomass and lipid productivity of N. oculata NCTU-3 cultured in the semicontinuous system aerated with 2% CO2 under the treatment of
one-day and three-day replacement ……….. 82 Table 3-5. Biomass and lipid production of the N. oculata cultutre supplemented with
different organic carbons under mixotrophic condition and photoautrophic culture with 2% CO2 aeration ……… 83
List of Figures
Figure 1-1. The relationship of carbon dioxide consecration elevating and global
temperature ……….... 84 Figure 1-2. A model for CO2 concentration in eukaryotic microalgae ……….... 85
Figure 1-3. Growth phases of microalgae cultures ……….……….. 86 Figure 1-4. Average changes in mass emissions from diesel engines using the
biodiesel mixtures relative to the standard diesel fuel (%) …….………... 87 Figure 1-5. Transesterification reaction of general equation and mechanism ………. 88 Figure 1-6. Flow diagrams comparing biodiesel production using the alkali- and
lipase-catalysis processes ……….. 89 Figure 1-7. The strategy of culturing microalgae in the photobioreactors for the
application to mitigate greenhouse gas and develop substitute energy …. 90 Figure 2-1. The photograph of five different microalgae stains ………... 91 Figure 3-1. The comparison with different microalgal strains cultures in growth …... 92 Figure 3-2. The comparison with different microalgal strains cultures in lipid
contents ……….. 93
Figure 3-3. The comparison with Chlorella sp. and N. oculata cultures by aeration
of CO2 in growth ………..….. 94
Figure 3-4. The gas chromatography patent of Chlorella sp. and Nannochloropsis
oculata ……….….. 95
Figure 3-5. Calibration curves and equations of optical density at A682 to the cell
density and biomass concentration in Chlorella sp. cultures ……….…... 96 Figure 3-6. Calibration curves and equations of optical density at A682 to the cell
density and biomass concentration in Nannochloropsis oculata
cultures ……….….. 97
Figure 3-7. Effects of different concentrations of CO2 aeration on the growth of
Figure 3-8. Calibration curves and equations of optical density at A220 to the nitrate
concentration in microalgal cultures ……….………. 99 Figure 3-9. Growth and lipid content of Chlorella sp. culture with nitrate
consumption ……….…….. 100 Figure 3-10. Growth and lipid content of Nannochloropsis oculata culture with
nitrate consumption ……….…….. 101 Figure 3-11. Growth profiles of N. oculata NCTU-3 cultured in the semicontinuous
system with 2% CO2 aeration and operated by one-day and three-day
replacements ……….…. 102 Figure 3-12. The growth curves of the N. oculata cultures with aeration of the
ambient air supplemented with different organic carbon sources in
mixotrophic, heterotrophic cultivations ……...……….………. 103 Figure 3-13. The growth curves of the N. oculata cultures with aeration of the
ambient air supplemented with different organic carbon sources in
mixotrophic condition ……….……... 104 Figure 3-14. Effects of different carbon sources on lipid content of N. oculata
compared with the control cultures of mixotrophic and heterotrophic
cultivations ……….……… 105 Figure 3-15. Images of Nile Red stained N. oculata NCTU-3 cells and the calibration
curve of Nile Red fluorescent intensity vs. lipid content ………….……. 106 Figure 3-16. ACC expression compare with lipid content ……….…… 107 Figure 3-17. The reaction of transesterification by three types of catalysts: acid, base,
and enzyme (lipase) ……….…….. 108 Figure 3-18. Figure of microalgal oil and biodiesel car ……….…… 109
I.
Literature Review
1-1 Global warming and energy shortage
Past years, the industrial revolution brought with it rapid economical development and a great improvement in our standard of living. As a result, it has also become an enormous burden on nature. The problem of global warming contributed by greenhouse gas is receiving great attention by the worldwide people.
Greenhouse gases are those gaseous constituents of the atmosphere, both natural and anthropogenic, those absorb and emit radiation at specific wavelengths within the spectrum of thermal infrared radiation emitted by the Earth’s surface, and then this property causes the greenhouse effect. Greenhouse gases are essential to maintaining the current temperature of the Earth. Planet Earth is habitable because of its location relative to the sun and because of the natural greenhouse effect of its atmosphere [Karl, 2003]. Earth's most abundant
greenhouse gases are water vapor, carbon dioxide (CO2), methane (CH4), nitrous oxide (NOx),
ozone (O3), and chlorofluorocarbons. The most important greenhouse gases are water vapor
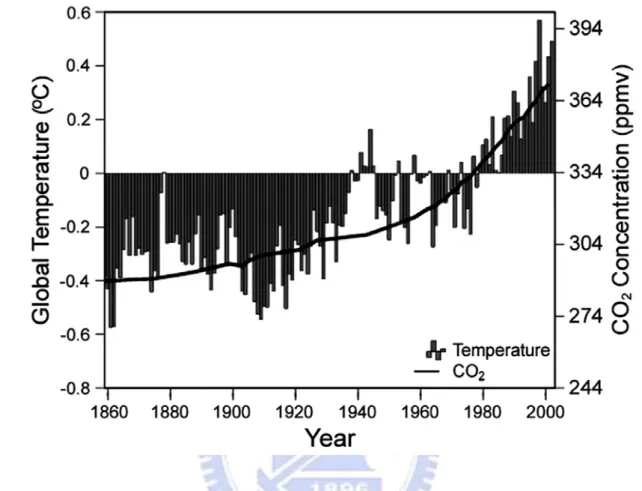
and carbon dioxide [Treut et al., 2007]. Human activities intensify the blanketing effect through the release of greenhouse gases. For instance, the amount of carbon dioxide in the atmosphere has increased by about 31% from the 1961 to 1990, and this increase is known to be due to human activities, primarily the combustion of fossil fuels and removal of forests. In the researches, the level of carbon dioxide in pre-industrial period is 280 parts per million by volume (ppmv), and the current level of carbon dioxide increased to more than370 ppmv today. Thus, humankind has dramatically altered the chemical composition of the global atmosphere with substantial implications for climate. Figure 1-1 shows the amount of CO2
is still elevating as an average increasing rate in these years and the global average surface heating approximates that of the increases of carbon dioxide [Houghton et al., 2001; Karl and
Trenberth, 2003]. The greenhouse gases trap outgoing radiation fromthe Earth to space, creating a warming of the planet.
The burning of fossil fuels has especially emitted large amounts of CO2, into the
atmosphere, which is causing a problem known as greenhouse effect and global warming [Michiki, 1995]. Over ten times more CO2 is fixed by plants into biomass, and annually
released by decomposers and food chains, than is emitted to the atmosphere due to the burning of fossil fuels. Human activity is already directly and indirectly affecting almost half of the terrestrial biological C cycle. Management of even a small fraction of the biological C cycle would make a major contribution to mitigation of this greenhouse gas [Hughes and Benemann, 1997].
CO2 is responsible for well over half of the total warming potential of all greenhouse gases.
Globally about 20 billion tons of fossil CO2 are emitted each year from the burning of fossil
fuels, and another 2 to 8 billion tons are released through human-mediated oxidation of the biosphere: intensive agriculture, deforestation and other unsustainable practices.
Cumulatively, net biosphere CO2 emissions due to human activities over the past century
have rivaled emissions from fossil CO2 sources. Indeed, human activities are currently
impacting a large fraction, approaching half, of the total annual terrestrial primary biological productivity of our planet, estimated at 500 billion tons of CO2 fixed annually [Vitousek,
1994; Hughes and Benemann, 1997].
To solve this international problem, some researches of biological CO2 fixation and
utilization have been carried out to develop those technologies in which CO2 is fixed by
utilizing microorganisms such as bacteria and microalgae and converting it into useful substances [Michiki, 1995]. Electric power generation is responsible for roughly one third of fossil CO2 emissions. Direct CO2 mitigation processes are those that reduce fossil CO2
emissions from specific power plants. Direct biological CO2 mitigation processes include
the cultivation of microalgae on flue-gas or captured CO2, and the co-firing of wood with
fossil fuels.
Another international problem is energy crisis which was also caused may be
over-consumption of fossil energy. Majority of the world energy needs are supplied through petrochemical sources, coal and natural gases, with the exception of hydroelectricity and nuclear energy, of all, these sources are finite and at current usage rates will be consumed shortly [Srivastava and Prasad, 2000; Meher, 2006]. Diesel fuels have an essential function in the industrial economy of a developing country and used for transport of industrial and agricultural goods and operation of diesel tractor and pump sets in agricultural sector. Economic growth is always accompanied by commensurate increase in the transport. The high energy demand in the industrialized world as well as in the domestic sector and pollution problems caused due to the widespread use of fossil fuels make it increasingly necessary to
develop the renewable energy sources of limitless duration and smaller environmental impact
than the traditional one. Global consumption of petrodiesel raised in an average of 16.1 million tons/year among 1990 and 2003 [IEA, 2006]. If the tendency keeps on, the worldwide demand for petrodiesel will increase to 1383 million tons/year by 2050. The global supply of petroleum is finite and expected to peak between 5 and 30 years from 2008 [Pahl, 2005; Pin Koh, 2007], after which demand will inevitably outstrip production. This has stimulated recent interest in alternative sources for petroleum-based fuels. The
alternative fuel must be technically feasible, economically competitive, environmentally acceptable, and readily available. One possible alternative to fossil fuel is the use of oils of plant origin like vegetable oils, tree borne oil seeds and microalgal oils for biodiesel
1-2 Microalgae
Microalgae are diverse group of prokaryotic and eukaryotic photosynthetic microorganisms that grow rapidly due to their simple structure. It is estimated that the biomass productivity of microalgae could be 50 times more than that of switch grass, which is the fastest growing terrestrial plant [Demirbas, 2006; Nakamura, 2006]. They are single- cellular photosynthetic microorganisms that convert sunlight, water and carbon dioxide to microalgal biomass.
The primary producers of oxygen and consumer of CO2 in aquatic environments are algae,
especially planktonic microalgae. Microalgae are microscopic in size and grow in liquid culture, nutrients can be maintained at or near optimal conditions potentially providing the benefits of well-controlling. Microalgae can perform continuous productivity similar to microbial fermentation. From these points of view, the study aims to search and screen microalgae from the natural sources like oceans and lakes, and to establish the most
appropriate culture conditions and then evaluate the photosynthesizing capabilities thereof. However, many microalgae are exceedingly rich in oil [Chisti, 2007; Banerjee, 2002], which can be converted to many products such as renewable fuels using existing technology. The utilization fields of microalgae can be categorized as follows [Michiki, 1995].
1. Energy-generating substances such as hydrocarbons, hydrogen, and methanol. 2. Foods and chemicals such as proteins, oils and fats, sterols, carbohydrates, sugar,
alcohols.
3. Other chemicals colorants, perfumes, vitamins, physiologically-active substances. Elementary techniques including separation and purification have been studied and evaluated.
system in which lipids and biodiesel from microalgae are separated and purified.
1-3 Microalgae for reducing pollution and renewable fuels
Photosynthetic microorganisms, microalgae, hold the key to realize the effective system of CO2 fixation and utilization. Particularly, about one half of global photosynthesis and
oxygen production is accomplished by marine microalgae. They play an important role in CO2 recycling through photosynthesis, which is similar to higher plants in O2-evolved
systems (PSI and PSII system).
Furthermore, technology development of microalgae culture that makes it possible to convert the formations resulting from photosynthesis into energy substances such as fuel oil. The study also seeks to develop technologies that separate and refine a variety of useful substances [Michiki, 1995]. Microalgae are sunlight-driven cell factories that convert carbon dioxide to potential biofuels, foods, feeds and high-value bioactives [Metting and Pyne,1986; Schwartz, 1990; Kay, 1991; Shimizu, 1996,2003; Borowitzka, 1999; Ghirardi et al., 2000; Akkerman et al., 2002; Banerjee et al., 2002; Melis, 2002; Lorenz and Cysewski, 2003; Metzger and Largeau, 2005; Singh et al., 2005; Spolaore et al., 2006; Walter et al., 2005; Chisti, 2007].
Notwithstanding, microalgae can provide several different types of renewable biofuels. These include methane produced by anaerobic digestion of the algal biomass [Spolaore et al., 2006; Chisti, 2007]; biodiesel derived from microalgal oil [Roessler et al., 1994; Sawayama et al., 1995; Dunahay et al., 1996; Sheehan et al., 1998; Banerjee et al., 2002; Gavrilescu and Chisti, 2005; Chisti, 2007] and photo-biologically produced biohydrogen [Ghirardi et al., 2000; Akkerman et al., 2002; Melis, 2002; Fedorov et al., 2005; Kapdan and Kargi, 2006; Chisti, 2007]. Using microalgae as a source of fuel is not a new idea [Chisti, 1980-1981;
Nagle and Lemke,1990; Sawayama et al., 1995; Chisti, 2007], but it is now being paid much attention because of the escalating price and exhausting of petroleum and, more significantly, the emerging concern about global warming that is associated with burning fossil fuels [Gavrilescu and Chisti, 2005].
1-4 Microalgae cultivation
Grow ability, chemical composition of microalgae, lipid content and lipid composition of microalgae are influenced by environmental conditions, such as culture medium, light,
temperature, carbon dioxide and so on [Richmond, 1986; Tomaselli et al., 1988, James et al., 1989; Oliveira et al., 1999; Renaud et al., 2002]. Producing microalgal biomass is generally more expensive than growing crops. Photosynthetic growth requires light, carbon source, water and inorganic salts. Temperature must remain generally within 20 to 30°C. To minimize expense, biodiesel production must rely on freely available sunlight, despite daily and seasonal variations in light levels.
1-4-1 Growth medium
Growth medium must provide the inorganic elements that constitute the algal cell. Essential elements include nitrogen (N), phosphorus (P), iron and in some cases silicon. Minimal nutritional requirements can be estimated using the approximate molecular formula of the microalgal biomass, that is CO0.48H1.83N0.11P0.01. This formula is based on data
presented by Grobbelaar [2004]. Sea water supplemented with commercial nitrate and phosphate fertilizers and a few other micronutrients is commonly used for growing marine microalgae [Molina Grima et al., 1999]. Growth media are generally inexpensive. Nutrients such as phosphorus must be supplied in significant excess because the phosphates added complex with metal ions, therefore, not all the added phosphorus is bio-available.
Silicon is specifically used for the growth of diatoms which utilize this compound for production of an external shell. Micronutrients consist of various trace metals and the vitamins thiamin (B1), cyanocobalamin (B12) and sometimes biotin. Two enrichment media
that have been used extensively and are suitable for the growth of most algae are the Walne medium and the Guillard’s f/2 medium. Commercially available nutrient solutions may reduce preparation labor. The complexity and cost of the above culture media often excludes their use for large-scale culture operations. And there are alternative suitable enrichment medium for mass production of microalgae in large-scale extensive systems contain only the most essential nutrients and are composed of agriculture-grade rather than laboratory-grade fertilizers.
1-4-2 Nitrogen contents
Organisms use carbon and nitrogen for the important nutrient source of energy and
construction of cell structure. Other than carbon, nitrogen is quantitatively the most element in algal nutrition. Nitrogen supply is essential for preparation of algal culture medium for most microalgae. Microalgae are usually able to use nitrate, nitrite, ammonia, or other organic nitrogen sources such as urea. In practical, the preferred nitrogen source is ammonia or urea, either of which is economically more favorable than nitrate or nitrite, which is more expensive.
The growth capacity is usually the same with these nitrogen sources. The average nitrogen requirement for many green algae is approximately 5-10% of the dry weight or 5-50mM [Becker, 1994]. Nevertheless, nitrogen amounts are considerably variable, since the nitrogen content in the culture medium can be operated to produce nitrogen-limitation algal cells. Nitrogen limitation affects photosynthesis by reducing the efficiency of energy collection due to loss of chlorophyll a and increases in non-photochemically active carotenoid
but may cause accumulation of numerous carbon compounds, i.e. polysaccharides or lipids. A low supply of nitrogen may result in a low respiration rate and an increase in the lipid reserves in most microalgae [Becker, 1994].
The major nonpolar lipids reported in algae are the triglycerides and hydrocarbons, and the major polar lipids classes include phospholipid, cardiolipin, diphosphatidylglycerol and glycolipids [Volkman et al., 1989 Dembittsky et al., 1991, Behrens and Kyle, 1996]. It is widely known that the main storage lipid class, triglycerides, accumulates in response to exhaust of nitrogen supply and during the stationary phase of a batch culture in most species [Richardson et al., 1969; Gordillo et al., 1998]. At the same time, the amounts of nitrogen supply may make difference in the fatty acid compositions of total lipid production and the length of fatty acid chains [Makrides et al., 1995; Regnault et al., 1995].
1-4-3 Light
Solar light is the energy source of the algal culture system for photosynthetic metabolism and is one of the major factors to determine the efficiency of the whole system. There are some environmental factors affect the lipid composition and growth of algae such as temperature, irradiation, and nutrient status [Thompson et al.., 1996]. Light drives
photosynthesis reaction and in this regard intensity, spectral quality and photoperiod need to be considered. Light intensity plays an important role, but the requirements vary greatly with the culture depth and density of the microalgal culture. However, light intensities that were adequate or optimal for growth in the log phase to declining growth phase can become stressful in stationary phase and lead to a condition known as photo-inhibition. It is
important that while the measured light intensity within the culture will decrease with increasing biomass if the incident illumination is maintained relatively high and then a large proportion of cells may become stressed, and the photo-inhibited culture can be pushed into
preferable for many species to halve or further reduce the incident light intensity when cultures enter stationary phase to avoid photo-inhibition.
At higher depths and cell concentrations, the light intensity must be rose up to penetrate through the culture medium. For example, 1000 lux is suitable for Erlenmeyer flasks, and 5000-10000 lux light is necessary for larger volumes. On the one hand, there are several studies of culturing Botryococcus brauni as well as Chlorella sp. by continuous light, and the other hand some studies are applying dark and light photoperiod for growing Chlorella sp. [Hogetsu and Miyachi, 1970; Maeda et al, 1995; Sawayama et al., 1995; Allard and Templier, 2000; Sato et al., 2003; Achitouv et al., 2004; Miao and Wu, 2006]. In order to complement and utilize the solar light at the maximum efficiency, many important studies of algae culture aim to develop a highly-efficient light collection device that minimizes the light loss due to reflection and absorption in the visible light area directly influencing the CO2 fixation
capability of microorganisms [Michiki, 1995]. 1-4-4 Temperature
The optimal growth temperature for phytoplankton cultures is generally between 20 and 24°C, although this may vary with the elements of the culture medium, the species and strain diversity. Microalgae do not have the ability to regulate their internal temperature, so the temperature may in turn affect the microalgae growth and substrate utilization rate [Esener et al., 1981]. Most species of cultured microalgae generally grow in appropriate temperatures between 16-27°C. Temperatures lower than 16°C will decrease the growth rate, whereas those higher than 35°C are lethal for a number of species. If necessary, microalgal cultures can be cooled by a water flow of cooling system over the surface on the culture vessel or by controlling the air temperature with refrigerated air - conditioning units.
metabolism and the metabolic reaction rate were well-known [Pirt, 1971; Novak, 1974; Mayo and Noike, 1996]. High growth temperature leads to different production of lipids and carbohydrates [Tomaselli et al., 1988; Oliveira et al., 1999; Renaud et al., 2002]. However, other studies have found that the response of microalgal composition to high and low growth temperatures varies from species to species. High growth temperature has been associated with increases in protein content and decreases in carbohydrate, and lipid in some
species[Thompson et al., 1992; Renaud et al.,1995], but these workers found no overall trend in gross biochemical composition for all species under study. We known temperature is a major effect on the types of fatty acids produced by microalgae. In some microalgal species, decreasing of growth temperature can bring about increasing of the unsaturated to saturated fatty acids [Ackman et al.,1968; Satu and Murata, 1980; Mortensen et al.,1988; Thompson et al., 1992; Oliveira et al., 1999; Renaud et al., 2002]. However, the response to growth temperature varies from species to species, with no overall consistent relationship between temperatures and chemical composition.
However, for the large scale of outdoor culturing system, microalgal culture could be heated over 40°C by sunshine in some extremely hot places. Microalgal growth is highly inhibited at high temperature, so there are many studies for searching the microalgae good in thermo-stability and productivity at high temperature for special cultivation.
1-4-5 Carbon dioxide
Microalgae need inorganic carbons to perform photosynthesis. Both CO2 and HCO3- are
potential sources of carbon for photosynthesis in microalgae. While CO2 can diffuse
through the microalgal cell membrane, HCO3- needs specific transport mechanisms to enter
the algal cell [Raven and Johnston, 1991; Beer, 1994; Moroney and Somanchi, 1999]. Figure 1-2 shows the model for CO2 concentration in eukaryotic microalgae. Microalgal
diffusion from the air. The natural CO2 concentration in air (0.03%) is too low to sustain
optimal growth and high productivity. Microalgae growing in fresh water with low salinity and at nearly neutral pH must be supplied with additional carbon to ensure satisfactory growth. Although intensive mixing may increase the entrance coefficient for CO2 from the air into the
culture, in most microalgal cultures, additional carbon mainly in the form of CO2-enriched air
is supplied [Becker, 1994]. There is also studies showing the utilization of organic carbon for heterotrophic or mixotrophic cultures.
CO2 addition furthermore buffers the water against pH changes as a result of the
CO2/HCO3- balance. High CO2 aeration will result in pH decreasing of culture solution.
High CO2 concentration aeration may be a harmful effect on the microalgal cells growth, but
increasing of microalgal cell density could promote the growth capacity of microalgae in the cultures aerated with higher CO2 concentrations [Chiu et al., 2008]. Therefore, the adequate
amount of CO2 supplied must not be inhibitory effects on microalgal cell growth inhibition;
otherwise lipid content will decrease. It was reported that the carbon assimilation of lipid synthesis was decreased with decrease of pH [Yung and Mudd, 1966, Chiu et al., 2008]. Higher CO2 aeration may cause the decrease of lipid content, but the increase of biomass may
improve the lipid productivity. 1-4-6 pH value
The pH value in the medium in algal system is known to influence the biomass regulation, ion transport system and metabolic rate [Guffanti et al., 1984; Mayo and Noike, 1994]. So controlling pH in culture medium will be one of the most critical factors affecting biomass and lipid production of microalgae. The pH value of the culture medium may be a simpler, indirect method for determining the degree of the cell growth of microalgae because the pH gradually rises as bicarbonate added to the culture medium is dissolved to produce CO2,
However, pH of algal cultures can be affected by several factors such as medium composition, buffering capacity of the medium, CO2 dissolution efficiency, temperature
(influences solubility of chemical compounds) and metabolic activity of the algal cells. Almost pH value of the medium in algal culture is usually neutral or slightly acidic values. The main reason is to avoid precipitation of microalgal cells and several major elements. Furthermore, different microalgal species vary greatly in their response to the pH and different microalgal strains show their own clear dependency on the pH of the growth in culture medium [Becker, 1994].
Increase of pH value during cultivation mainly caused by the depletion of the anions NO3+
and the formation of CO2 as well as releasing of OH- ions in the medium. When the pH
value increases above 9, it may make the precipitation of several calcium salts, i.e. carbonates, phosphates and sulfates, following deficiencies of nutrients, growth retardations and even algal flocculation which is induced by the precipitating elements[Becker, 1994]. Therefore, controlling stable and suitable pH value in the broth must be necessary for healthy cultures and optimal productivity of microalgal cultivation.
1-4-7 Growth phase
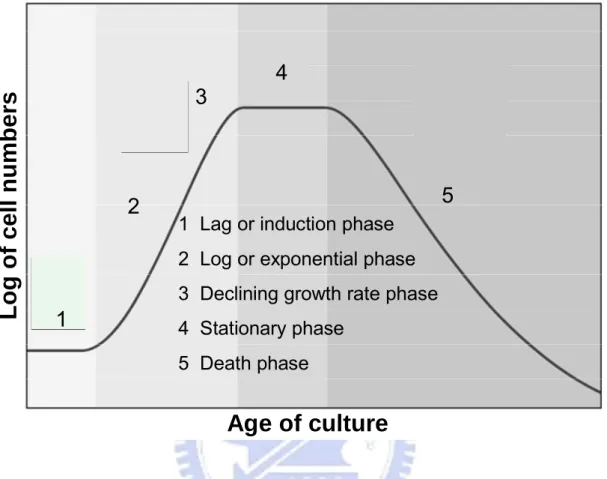
There are 5 reasonably well defined phases of algal growth in batch cultures. The characterized phases are including lag phase, exponential phase, phase of declining growth rate, stationary phase, and death phase [Fogg and Thake, 1987]. Each phase is described in Figure 1-3.
Lag phase, the condition of the inoculums has a strong bearing on the duration of this period [Spencer, 1954]. A lag phase may also occur if the inoculum is transferred from one set of growth conditions to another. An inoculum taken from a healthy exponentially growing culture is unlikely to have any lag phase when transferred to fresh medium under
phase will be proportional to the length of time the inoculum has been in phases 3-5. Log phase, the cell density of culture during which numbers increase exponentially and which is represented by a part of the growth curve that is a straight line segment if the logarithm of numbers is plotted against time. The growth rate of a microalgal population is a measure of the increase in biomass over time and it is determined from the log phase. The duration of log (exponential) phase in cultures depends upon the size of the inoculum, the growth rate and the capacity of the medium and culturing conditions to support algal growth. Declining of growth phase normally occurs in cultures when either a specific requirement for cell division is limiting or something else is inhibiting reproduction. In the phase of growth biomass is often very high and exhaustion of a nutrient, limiting carbon dioxide or light limitation becomes the primary causes of declining growth. Light limitation at high density cultures occurs when the cells absorb most of the incoming irradiation and individual cells shade each other called self-shading effect. In stationary phase, cultures reach a relatively constant cell density, and cells may undergo dramatic biochemical changes within a matter of hours. The nature of the changes depends upon the growth limiting factor. Nitrogen limitation may result in the reduction in protein content and relative or absolute changes in lipid and carbohydrate content. Light limitation will result in increasing pigment content of most species and shifts in fatty acid composition. In the final phase, death phase, water quality deteriorates and nutrients are exhaust to a level incapable of sustaining growth and survival. Cell density decreases rapidly and the culture eventually collapses.
In the life cycle of algae, growth (in cell number or biomass) continues slowly for some time in a medium completely depleted of nitrogen, producing cells of high lipid content. When nitrogen depletion progresses, chlorophyll and protein contents decrease and carbohydrate content increases, followed by increases in lipid content [Iwamoto and Sugimoto, 1958].
The gross biochemical composition of microalgae was affected more by the growth phase than by the nitrogen source. Protein was higher during exponential growth (about 37-45% AFDW, ash free dry weight) but the pattern changed as the cultures aged, and lipids were the main algal constituents on all nitrogen sources in the late stationary phase. In all cultures, the relative abundance of neutral lipid increased in the late stationary phase along with a proportional reduction in phospholipids, whereas galactolipids only slightly changed during the growth of the cultures. Total fatty acid content was affected by nitrogen source and growth phase. Maximal PUFA (polyunsaturated fatty acid) values were obtained at the early stationary phase and decreased throughout the stationary phase [Fidalgo et al.,1998].
1-5 Microalgae cultures techniques
Photosynthetic microorganisms play an influential role in the conversion of solar energy into chemical energy. Photosynthetic conversion is an efficient and alternative process used in several industrial fields. This use of algal biomass is an important consideration for industrial applications of microalgal cultures. With advances in processing technology, algal biomass has come to be a possible source of fuels, fine chemicals, and pharmaceuticals. So, developing a large-scale method for the cultivation of microalgae have been performed using many different kinds of cultivation systems for providing alternatives to fermentation,
agriculture, fuel products [Matsunaga et al., 2005; Chisti, 2007]. The microalgal culture systems have been constructed well in many different systems. The development of efficient large-scale culture systems is necessary for algal mass production and the industrial
applications of microalgae. These open culture systems are the simplest method of algal cultivation and offer advantages in low construction cost and ease of operation [Torzillo et al., 2003].
Large-scale culture have been constructed and classified as open and closed systems. Open culture systems are uncovered cultures such as unstirred open ponds, raceway ponds, circular ponds, paddle-wheel pond and so on [Richmond, 1992; Chaumont, 1993]. The open culture systems require large surface areas and shallow depth (12-15 cm) to improve light penetration efficiency [Matsunaga et al., 2005]. In open pond systems, any cooling is
achieved only by evaporation. Swung temperature is dependent on a diurnal cycle and seasons. Evaporative culture medium loss can be significant because of uncovered cultures and
significant water losses to atmosphere, open pond systems low utilization efficiently of carbon dioxide than closed photobioreactors. The biomass productivity remains low because open pond systems can’t be sustained in a stable condition. Furthermore, open ponds are poorly mixed and cannot keep an optically dark zone. However, the biggest problem in open culture systems is contamination by unwanted algal species and other microorganisms. Production of microalgal biomass for making biodiesel fuels has been extensively evaluated in raceway ponds in studies sponsored by the United States Department of Energy. Open ponds are considered to be less expensive than photobioreactors, because they cost less to build and operate. Although raceways are low-cost, they have low biomass productivity compared with photobioreactors [Sheehan et al., 1998; Chisti, 2007].
Closed systems, photobioreactors have been expected to overcome the problems of open culture systems, and also several types of photobioreactors have been created such as helix tubular reactors, plane tubular reactors, laminar reactors, column reactors and so on. Photobioreactors permit essentially single-species culture of microalgae for prolonged durations of growth. Photobioreactors have been successfully used for producing large quantities of microalgal biomass until now [Molina Grima et al., 1999; Pulz, 2001; Carvalho et al., 2006]. The advantages of closed photobioreactors are (1) facilitating maintenance of mono-algal cultures by preventing them from contamination; (2) reducing water evaporation
and the subsequent increase of salinity in the culture medium; (3) resulting in higher productivity with greater cell densities, reducing harvesting costs; and (4) applicable to various microalgal species under favorable and controllable culture conditions. The production yield of algal biomass cultures depends on the light path length to each cell, and therefore the increase of surface area-to-volume ratio (SA: VOL) is an important factor for efficient light utilization in photobioreactors. Consequently, light is a limitation factor for mass culture productivity, and high productivity of cell biomass at harvest potentially can elicit a low production cost. However, the closed systems do have some disadvantages. Oxygen concentrations resulting from active photosynthesis can be raised because of the high biomass, and therefore high concentration of dissolved oxygen in combination with intense sunlight by the high SA: VOL result in photo-inhibition and photo-oxidative damage to algal cells [Brown, 2002; Matsunaga et al., 2005; Chisti, 2007]. So degasser systems sometimes are required. Photobioreactors also require cooling systems during daylight hours and temperature control at night is also useful.
Open pond systems have lower productivity of algal biomass, require larger land areas for cultivation, and involve higher land costs. By contrast, closed culture systems can reach high-density culture and decrease large land costs. However, high surface area-to-volume ratio required for collecting a sufficient amount of solar energy, and therefore operating costs are higher than for open systems. Furthermore, suitable culture systems should be chosen according to the target products and available environmental conditions.
1-6 Mixotrophic and heterotrophic culture of microalgae
Autotrophs are organisms that produce complex organic compounds from simple inorganic molecules and external sources of energy, such as light or chemical reactions of inorganic
compounds. Most microalgae are autotroph organisms and also called photoautotrophs because they can perform photosynthesis by utilizing inorganic carbon, CO2. However,
heterotrophs are organisms that require organic substrates to get carbon for growth and development and mixotrophs are organisms obtain electrons from inorganic electron sources, but uses organic matters as carbon sources. So mixotrophic organisms can derive metabolic energy both from photosynthesis and from external energy sources. Some microalgae can be cultivated in heterotrophic or mixotrophic cultures.
A sufficient supply of carbon is vital important for successful cultivation since about 50% of the algal biomass consists of carbon [Sánchez Mirón et al., 2003]. Carbon can be supplied as an inorganic substrate in the form of CO2, as is the case for most photoautotrophic
forms. However, microalgae take up organic molecules as a primary source of nutrition in heterotrophic growth and may also obtain nutrition by combining photoautotrophic and
heterotrophic mechanisms in mixotrophic growth. In heterotrophic and mixotrophic cultures, organic carbon sources are mainly sugars or acetate [Laliberté and de la Noüe, 1993; Becker, 1994]. Some algal species are applied to heterotrophic growth culture using organic carbon as their energy and carbon source. These species can be grown in conventional fermenters, and heterotrophs can be grown to much higher cell densities than phototrophs, resulting in high productivity because light is not required [Behrens and Kyle, 1996].
The economic feasibility of microalgal mass culture for biodiesel production greatly depends on the high biomass productivity and considerable lipid yields. However, the high cost of oil feedstock is the main reason to the high cost of biodiesel and makes the obstacle to its broader commercialization [Wright, 2006; Li et al., 2007]. There are some researches make efforts on minimizing oil feedstock costs [Al-widyan and Al-shyoukh, 2002; Antolin et al., 2002; Zhang et al., 2003; Noureddini et al., 2005; Dhaermadi et al., 2006; Li et al., 2007]. Heterotrophic culture may not only overcome nearly all of the limitations of the phototrophic
growth systems but also provide a cost-effective, large-scale alternative method of cultivation for some microalgae that utilize organic carbon substances as their sole carbon and energy sources. This mode of growth can apply to low cost culture which eliminates the
requirement for light and offers the possibility of greatly increasing microalgal cell
concentration and volumetric productivity in batch and fed-batch systems [Anderson et al., 1978; Boswell et al., 1992]. Li et al. [2007] reported that heterotrophic Chlorella grown in commercial large-scale bioreactors could reach a high biomass concentration and high lipid content. However, the heterotrophic or mixotrophic growth of microalgae may change the fatty acid composition significantly depending on the growth mode [Constantopoulus, 1970; Regnault et al., 1995; Behrens and Kyle, 1996]. For example, Regnaul et al. [1995] reported the photosynthetic culture of Euglenophyceue contain mostly C14-C22 with C16 and C18 dominating , while grown organisms in heterotrophic cultures contain largely 14:0, 16:0 and 18:l.
1-7 Acetyl-coenzyme A carboxylase of microalgae
Acetyl-coenzyme A carboxylase (ACCase) is a biotin-containing enzyme that catalyzes the the ATP-dependent carboxylation of acetyl-CoA to form malonyl-CoA. This reaction is believed to be a key regulatory step in fatty acid biosynthesis in animals, bacteria, and plants, especially including microalgae [Roessler and Ohlrogge, 1993]. Two partial reactions are involved in this process. First is carboxylation of the enzyme-bound biotin molecule and second is transfer of the carboxyl group to acetyl-CoA.
The rate of fatty acid synthesis in plants appears to be dependent on the activity of ACCase. ACCase enzyme has been receiving an increasing amount of attention as a target for
capabilities of various organisms because ACCase plays an important role in controlling the rate of fatty acid biosynthesis.
1-8 Extraction method of microalgae
High oil-contents microalgae can be used to produce biodiesel, which is chemically modified natural lipid and biodiesel form microalgae is emerging as an exciting new option for diesel engines. The actual production of the biofuel from the microalgae requires the extraction of the natural microalgal oils in the first, and then the formation of useable fuel from oil. Apart from culture affecting factors, ex. nitrogen source, light, and temperature, lipid extraction method can also significantly affect lipid yield [Grima et al., 1994; Lee et al., 1998; Rodriguez-Ruiz et al., 1998; Shen et al., 2009].
Typical microalgae lipid extraction usually follows two steps: cell disruption and solvent extraction. Several lipid extraction methods work differently on different algae species because various microalgae species have large variations in algal cell shape, cell size, cell wall structure and characteristics of algal lipids. Lee et al. [1998] reported the effects of five cell disruption methods along with five solvent systems on lipid extraction of Botryococcus brauni. The five cell disruption methods are French press, sonication, homogenization, bead-beater and lyophilization. For examples of press and sonication methods, expression press method is the simplest way to extract oil but works best based on algal species. Expression or expeller press method is essentially a large press used to compress the algae bodies making the natural oils to seep out and the microalgae must first be dried. And
further, the other method, ultra-sonication, is the use of an ultrasonic reactor to extract the
natural algal oils. Ultrasonic reactor sends ultrasonic waves through the microalgae which create cavitation bubbles. When cavitation bubbles break around and destroy the cell walls
of the microalgae, the shock waves produced collapse the cell wall releasing the intracellular contents. After all, the use of cell disruption usually couples with chemical extraction to reach optimum extraction efficiency.
Besides, the five disruption methods connect with five solvent systems, i.e.
chloroform/methanol, hexane/isopropanol, dichloroethane/methanol, dichloroethane/ethnol and acetone/dichloromethane. However, different algal strains are suitable for own specific cell disruption/ solvent system. Some researches showed that bead-beater followed by
solvent system of chloroform/methanol (2:1, v/v) was the most effective method for
Botryococcus brauni and the dichloroethane/methanol and dichloroethane/ethanol systems
were successful on lipid extraction of green alga Cladofora [Damyanova et al., 1992; Shen et al., 2009]. Solvent extraction is easy-manipulation and low capital investment for
equipment, but the major disadvantages are that many of the chemicals have the potential to be harmful, non-continuous process with waiting periods and requirement of more manual labor. Therefore, the various lipid extraction of microalgae are developing for discover more effective method for optimal extraction technique of microalgal lipid. The different
common lipid extraction methods applying to microalgae contain soxhlet extraction, supercritical fluid extraction, enzymatic extraction, osmotic shock extraction and so on.
1-9 Biodiesel
Because the prices for crude oil rose up dramatically and the concerns of global warming
grew, people are searching for alternatives to petroleum-based energy sources actively. [Pacala and Socolow, 2004; Ragauskas et al., 2006; Tilman et al., 2006; Pin Koh , 2007]. Biodiesel fuel is a diesel-equivalent, renewable fuel derived from biological feedstocks (i.e., usually agricultural crops used as raw materials for biodiesel production). Biodiesel is
gaining more and more importance as an attractive fuel due to the depleting fossil fuel
resources. A global shift in consumption trends from petroleum-based diesel (petrodiesel) to biodiesel could reduce the world’s dependence on a finite supply of fossil fuels.
Presently, Western Europe (Germany, France, Italy, Austria, and Spain) makes efforts in
biodiesel and contributes over 90% of global biodiesel production (global production: 2.8 million tons/year). Meeting the estimated global biodiesel demand in 2050 would require an
almost 100-fold increase from current production capacities [Pahl, 2005; Pin Koh, 2007].
The world’s largest consumer of petrodiesel-United States [IEA 2006]-strives for reducing its dependence on petroleum through its Energy Conservation Reauthorization Act of 1998, in which federal agencies are mandated to meet fuel credit requirements by either purchasing alternative-fueled vehicles or using motor fuel containing at least 20% biodiesel by volume [U.S.Department of Energy, 2001].
1-10 Biomass for biodiesel
Biodiesel generally falls under the category of biomass which refers to renewable organic
matter such as energy crops, crop residues, wood municipal and animal wastes that are used
to produce energy [Pahl, 2005]. More specifically, biofuels, a subcategory of biomass includes three energy-crop-derived liquid fuels: ethanol (grain alcohol), methanol (wood alcohol), and biodiesel. The plant oils usually contain free fatty acids, phospholipids, sterols, water, odorants and other impurities. Because of these, the oil cannot be used as fuel
directly. To overcome these problems the oil requires slight chemical modification mainly
transesterification,pyrolysis and emulsification. Among these, the transesterification is the
key and foremost important step to produce the cleaner and environmentally safe fuel from vegetable oils. Pure biodiesel does not contain any petroleum diesel or fissile duel of any
sort.
Technically, biodiesel is fatty acid alkyl ester which can be easily made through a simple chemical process form virtually any vegetable oil, such as soy, corn, rapeseed, cottonseed, peanut, sunflower, avocado, mustard seed and so on. Actually, biodiesel can also be made from recycling cooking oil and animal fats. However, there have been more and more promising experiments with the use of microalgae as a biodiesel feedstock. And the chemical process of making biodiesel is so simple that biodiesel can be made by virtually anyone, although the chemicals required (usually lye and methanol) are hazardous and need to be handled with extreme caution.
1-11 Biodiesel characteristic
Biodiesel is an alternative source of energy that is replacing the use of gasoline and diesel. The emergence of biodiesel fuel substitutes has led to several studies on their properties. Biodiesel is similar to petroleum diesel fuel in many chemical and physical properties. Engine performance and the fuel consumption were favorable that make it better substitute to diesel fuel [Bajpai and Tyagi, 2006]. Biodiesel is 100% renewable and being plant based. The advantages of biodiesel as diesel fuel are liquid nature portability, ready availability, renewability, sustainable, higher biodegradable, higher combustion efficiency, higher cetane number, non-toxic, lower sulfur and aromatic content, non-polluting and environmentally safe [Ma and Hanna, 1999;Mudge and Pereira, 1999; Speidel et al., 2000; Zhang et al., 2003; Knothe et al., 2006]. Main advantages of biodiesel given in the literature include domestic origin, reducing the dependency on imported petroleum, biodegradability, high flash point and inherent lubricity in the neat form [Mittelbach and Remschmidt,2004; Knothe et al., 2005]. By the by, the higher oxygen content of biodiesel than diesel improves the
combustion efficiency due to the increase of the homogeneity of oxygen with the fuel during combustion and decreases its oxidation potential.
Biodiesel is the only alternative fuel so that low concentration biodiesel-diesel blends run on conventional unmodified engines. It can be stored anywhere where petroleum diesel fuel is stored. The risks of handling, transporting and storing biodiesel are much lower than petrodiesel. Biodiesel is safe and easy to handle and transport because it is as biodegradable and has a high flash point compared to petroleum diesel fuel. Biodiesel can be used alone or mixed in any ratio with petroleum diesel fuel. The most common blend is a mix of 20% biodiesel with 80% petroleum diesel, or B20 under recent scientific investigations.
In view of environmental considerations, biodiesel is considered as a “carbon neutral” fuel because all the carbon dioxide released during biodiesel consumption had been sequestered from the atmosphere for the growth of vegetable oil crops [Barnwal et al., 2005]. Studies have shown that the combustion of 1 liter of diesel fuel leads to the emission of about 2.6 kg of CO2 against 1 kg of CO2/liter of biodiesel [Tickell, 1999]. The combustion of biodiesel
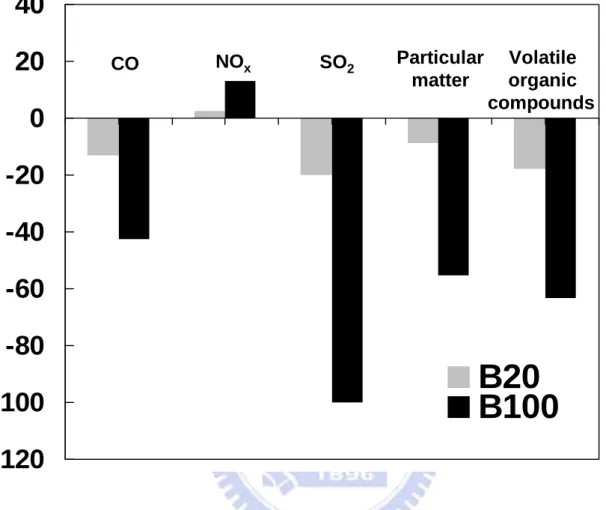
compared to diesel has been reported to emit lesser pollutants, such as SO2, soot, CO,
hydrocarbons (HC) and polyaromatic hydrocarbons (PAHs), and aromatics. Combustion of biodiesel alone provides over a 90% reduction in total unburned hydrocarbons, and a 75-90% reduction in polycyclic aromatic hydrocarbons (PAHs). Biodiesel provides a slight increase or decrease in nitrogen oxides depending on engine family and testing procedures. The average of pollutants emission from diesel engines using the biodiesel mixtures relative to the
standard diesel is shown in Figure 1-4 [Morris et al., 2003]. Biodiesel contains virtually
trace amount of sulfur and the sulfur content of petrodiesel is 20-50 times those of biodiesels,
so SO2 emissions are reduced in direct proportion to the petrodiesel replacement. Emissions
of all pollutants except NOx appear to decrease when biodiesel is used. The fact that NOx emissions increase with increasing biodiesel concentration could be a detriment in areas that
are out of attainment for ozone. The use of blends of biodiesel and diesel oil is preferred in
engines, in order to avoid some problems related to the decrease of power and to the increase
of NOx emissions (a contributing factor in the localized formation of smog and ozone) with increasing content of pure biodiesel in the blend [Schumacher et al., 1996].
1-12 Method of biodiesel production
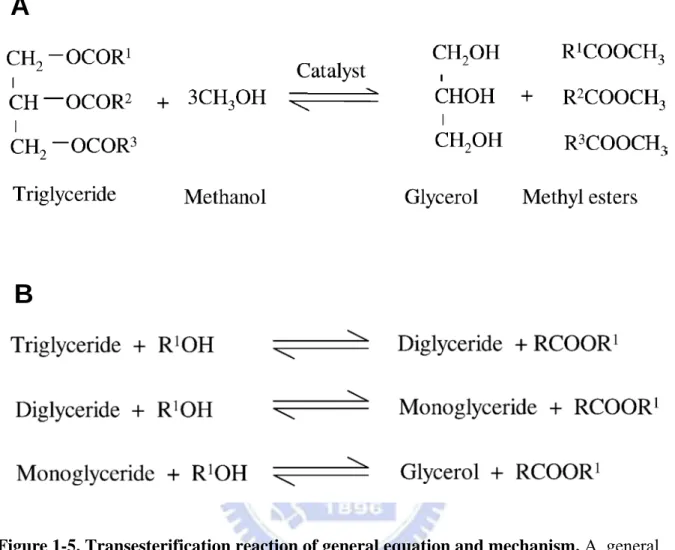
Current oil used in producing biodiesel almost consists of triglycerides which contain three fatty acid molecules and a glycerol skeleton. Three molecules of fatty acid are esterified with a molecule of glycerol to form a triglyceride (TG) molecule [Chisti, 2007]. For the purpose of making biodiesel, the algal or vegetable oil is reacted with methanol and catalysts (alkaline, acid or enzyme). This reaction is known as transesterification or alcoholysis, and the products in this reaction are glycerol and fatty acid methyl esters, which is namely biodiesel [Srivastava and Prasad, 2000; Fukuda et al., 2001; Chisti, 2007].
Transesterification reaction requires 3 mol of alcohol for each mole of triglyceride to produce 1 mol of glycerol and 3 mol of methyl ester (Figure 1-5A). There are three steps in this reaction, the first one is diglycerides formation, and then diglycerides are converted to monoglycerides and eventually to glycerol (Figure 1-5B) [Meher, 2006]. The glycerol layer settles down at the bottom of the reaction vessel. Diglycerides and monoglycerides are the intermediates in this process.
The step wise reactions are reversible and a little excess of alcohol is used to shift the equilibrium towards the formation of esters. In presence of excess alcohol, the foreword reaction is pseudo-first order and the reverse reaction is found to be second order. It was also observed that transesterification is faster when catalyzed by alkali [Freedman et al., 1986].
1-12-1 Catalyst type and concentration in the transesterification
Catalysts used for the transesterification of triglycerides are classified as alkali, acid, enzyme or heterogeneous catalysts. The alkali catalysts are sodium hydroxide (NaOH), sodium methoxide (NaOMe), potassium hydroxide (KOH), potassium methoxide (KOMe) are more effective and usual for use [Ma and Hanna, 1999; Meher et al., 2006]. Some
researches have investigated the inhibitory effect of base-catalyzed transesterification of triglycerides containing substantial amount of free fatty acid. Free fatty acids will react with the alkaline catalyst which is added for the reaction and give rise to soap, as a result of which, one part of the catalyst is neutralized and is therefore no longer enough for transesterification reaction. The soap consumes the catalyst and reduces the catalytic efficiency, as well as causing an increase in viscosity, the formation of gels, and difficulty in achieving separation of glycerol [Ma and Hanna, 1999].These high FFA content oils/fats are processed with an immiscible basic glycerol phase so as to neutralize the free fatty acids and cause them to pass over into the glycerol phase by means of monovalent alcohols [Turck, 2002; Meher et al.,
2006]. Many studies stressed on the importance of oils being dry and free fatty acids and also suggested that the free fatty acid content of the refined oil should be as low as possible below 0.5% and the ester yields were significantly reduced if the reactants did not meet these requirements; sodium hydroxide or sodium methoxide reacted with moisture and carbon dioxide in the air which are diminishing their effectiveness[Freedman et al., 1984; Ma and Hanna, 1999; Feuge and Grose, 1949].
So when the oil has high free fatty acid content, high acid value and more water content, acid catalyzed transesterification is suitable to solve these problems even though the transesterification by acid catalysis is much slower than that by alkali catalysis [Ma and Hanna, 1999; Srivastava and Prasad, 2000; Fukuda, 2001; Freedman, 1984]. The most common acids to use are sulfuric acid (H SO ), phosphoric acid (H PO ), hydrochloric acid



![Figure 1-2. A model for CO 2 concentration in eukaryotic microalgae. The font sizes of CO 2 and HCO 3 - indicate the relative concentrations of these inorganic carbon species [Moroney and Somanchi, 1999]](https://thumb-ap.123doks.com/thumbv2/9libinfo/8449387.182389/97.892.107.766.204.731/concentration-eukaryotic-microalgae-indicate-concentrations-inorganic-moroney-somanchi.webp)