Inhibition of Proinflammatory Tumor Necrosis Factor-
␣-Induced
Inducible Nitric-Oxide Synthase by Xanthine-Based
7-[2-[4-(2-Chlorobenzene)piperazinyl]ethyl]-1,3-dimethylxanthine
(KMUP-1) and 7-[2-[4-(4-Nitrobenzene)piperazinyl]ethyl]-1,
3-dimethylxanthine (KMUP-3) in Rat Trachea: The Involvement
of Soluble Guanylate Cyclase and Protein Kinase G
Bin-Nan Wu, Chien-Wen Chen, Shu-Fen Liou, Jwu-Lai Yeh, Hui-Hsuan Chung,
and Ing-Jun Chen
Department and Graduate Institute of Pharmacology, College of Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan Received March 23, 2006; accepted June 5, 2006
ABSTRACT
In the study of anti-proinflammation by [2-[4-(2-chlorobenze-ne)piperazinyl] ethyl]-1,3-dimethylxanthine (KMUP-1) and 7-[2-[4-(4-nitrobenzene)piperazinyl]ethyl]-1,3-dimethylxanthine (KMUP-3), exposure of rat tracheal smooth muscle cells (TSMCs) to tumor necrosis factor-␣ (TNF-␣), a proinflammatory cytokine, increased the expression of inducible nitric-oxide synthase (iNOS) and NO production and decreased the expres-sion of soluble guanylate cyclase␣1(sGC␣1), soluble guanylate cyclase1(sGC1), protein kinase G (PKG), and the release of cGMP in TSMCs. The cell-permeable cGMP analog 8-Br-cGMP, xanthine-based KMUP-1 and KMUP-3, and the phos-phodiesterase 5 inhibitor zaprinast all inhibited TNF-␣-induced increases of iNOS expression and NO levels and reversed TNF-␣-induced decreases of sGC␣1, sGC1, and PKG expres-sion. These results imply that cGMP enhancers could have anti-proinflammatory potential in TSMCs. TNF-␣ also increased
protein kinase A (PKA) expression and cAMP levels, cyclooxy-genase-2 (COX-2) expression, and activated productions of prostaglandin (PG) E2and 6-keto-PGF1␣ (stable PGI2 metabo-lite). Dexamethasone and N-[2-(cyclohexyloxyl)-4-nitrophenyl]-methane sulfonamide (NS-398; a selective COX-2 inhibitor) attenuated TNF-␣-induced expression of COX-2 and activated productions PGE2and PGI2. However, KMUP-1 and KMUP-3 did not affect COX-2 activities and did not further enhance cAMP levels in the presence of TNF-␣. It is suggested that TNF-␣-induced increases of PKA expression and cAMP levels are mediated by releasing PGE2and PGI2, the activation prod-ucts of COX-2. In conclusion, xanthine-based KMUP-1 and KMUP-3 inhibit TNF-␣-induced expression of iNOS in TSMCs, involving the sGC/cGMP/PKG expression pathway but without the involvement of COX-2.
Pro-inflammatory cytokines, including TNF-␣, play an im-portant role in regulating the tracheal smooth muscle contrac-tility that is found in the asthmatic phenotype. TNF-␣ is
in-creased in the sputa of patients with bronchial asthma and present in the bronchoalveolar lavage fluid of symptomatic asthmatic patients (Renauld, 2001). As a member of these cy-tokines, TNF-␣ attracts and activates nonspecific inflammatory macrophages and neutrophils during infection and hypersensi-tivity induced by the inhalation of organic particles or fumes (Mohr, 2004; Mendez-Samperio et al., 2006).
Likewise, proinflammatory TNF-␣, inducible nitric oxide-This study was supported by grants 2320-B-037-002 and
NSC-94-2323-B-037-005 to I.-J.C. from the National Science Council, Taiwan. Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
doi:10.1124/mol.106.024919.
ABBREVIATIONS: TNF-␣, tumor necrosis factor-␣; TSM, tracheal smooth muscle; COX-2, cyclooxygenase-2; iNOS, inducible nitric-oxide
synthase; PDE, phosphodiesterase; PG, prostaglandin; PKA, protein kinase A; PKG, protein kinase G; KMUP-1, 7-[2-[4-(2-chlorobenzene)pipera-zinyl] ethyl]-1,3-dimethylxanthine; KMUP-3, 7-[2-[4-(4-nitrobenzene)pipera7-[2-[4-(2-chlorobenzene)pipera-zinyl]ethyl]-1,3-dimethylxanthine; sGC, soluble guanylate cyclase; TSMC, tracheal smooth muscle cell; NS-398, N-[2-(cyclohexyloxyl)-4-nitrophenyl]-methane sulfonamide; IL, interleukin; TTBS, Tris-buffered saline/Tween 20; PBS, phosphate-buffered saline; PDEI, phosphodiesterase inhibitor; buffer A, Triton X-100 and bovine serum albumin in phosphate-buffered saline; CPT, chlorophenylthio; YC-1, 3-(5⬘-hydroxymethyl-2⬘-furyl)-1-benzylindazole; Bay-41-2272, 5-cyclopropyl-2-[1-(2-fluorobenzyl)-1H-pyrazolo[3,4-b]pyridin-3-yl]pyrimidin-4-ylamine; Rp-CPT-cAMPs, 8-(4-chlorophenylthio)adenosine-3⬘,5⬘-cyclic monophospho-rothioate, Rp-isomer; Rp-CPT-cGMPs, 8-(4-chlorophenylthio)guanosine-3⬘,5⬘-cyclic monophosphorothioate, Rp-isomer.
MOLECULARPHARMACOLOGY Vol. 70, No. 3
Copyright © 2006 The American Society for Pharmacology and Experimental Therapeutics 24919/3133007
Mol Pharmacol 70:977–985, 2006 Printed in U.S.A.
977
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org
synthase (iNOS), and cyclooxygenase-2 (COX-2) are coex-pressed in pulmonary airway infection. Local production of TNF-␣ has been found to be regulated by iNOS and COX-2 and thus serves to orchestrate the inflammation pathway (Watkins et al., 1999). Enhanced COX-2 and iNOS expres-sion by TNF-␣ can increase the production of cAMP and cGMP as a result of activated adenylate cyclase and guany-late cyclase, respectively. Because high-output cyclic nucleo-tide production in response to inflammation suppresses pro-tein kinase G (PKG) expression, and cAMP analogs are more potent than cGMP analogs in reducing PKG mRNA expres-sion, suggesting that PKA mediated the effects of cAMP and cGMP through cross-activation (Browner et al., 2004).
Xanthine-based phosphodiesterase inhibitors (PDEIs) have been used as bronchodilators because they can nonse-lectively inhibit PDE, resulting in increases of cAMP. There-fore, they have been used to treat bronchospastic diseases associated with immunoresponses (Giembycz, 2000; Car-amori and Adcock, 2003). In contrast, xanthine-based 7-[2-[4-(2-chlorobenzene)piperazinyl] ethyl]-1,3-dimethylxan-thine (KMUP-1), with endothelium- and epithelium-derived NO-enhancing activities, has been found to relax smooth muscle contractions by both activating sGC and inhibiting PDE, leading to the accumulation of cGMP (Wu et al., 2001, 2004; Lin et al., 2002). KMUP-1 has been shown to activate large-conductance Ca2⫹-activated K⫹ channels in basilar artery myocytes via cAMP- and cGMP-dependent protein kinases (Wu et al., 2005). Furthermore, intratracheal 7-[2-[4-(4-nitrobenzene)piperazinyl]ethyl]-1,3-dimethylxanthine (KMUP-3) has been reported to increase the respiratory per-formance and to protect against TNF-␣-induced airway re-sistance, involving the activation of sGC/cGMP/PKG (Lin et al., 2006). Previously, however, TNF-␣ was reported to mod-ulate tracheal responses to G-protein-coupled receptor ago-nist and to act as an inflammatory cytokine in patients with asthma (Chen et al., 2003). Thus, it is still not known whether xanthine-based KMUP-1 and KMUP-3, chemically with imidazole (isoform of indazole) moiety, have anti-proinflammatory actions to modulate or attenuate TNF- ␣-induced iNOS and COX-2 expression, cGMP, and expression of PKG.
A nonxanthine sGC activator, YC-1, chemically with inda-zole moiety, exerts cGMP-dependent and -independent ac-tions. The latter include the inhibition of PDE (Friebe et al., 1998) and untoward COX-2 expression in pulmonary epithe-lial cells (Chang et al., 2004). Thereafter, a more selective sGC activator, indazole-based Bay-41-2272, similar to YC-1, has also been reported to come with cGMP-dependent and -independent actions in vascular system (Teixeira et al., 2006). In addition, KMUP-1 and KMUP-3 were described previously to activate sGC in trachea (Wu et al., 2004; Lin et al., 2006). sGC activation by YC-1 and BAY-41-2272 has thus encouraged us to investigate the involvement of sGC/cGMP/ PKG in associated anti-proinflammation by KMUP-1 and KMUP-3. The difficulty in using sGC activators is that they act nonselectively on several organs (Doggrell, 2005), which might limit their clinical potential, particularly in the pul-monary airway.
PDE5 inhibitors with cGMP-increasing activity have proven to induce tracheal relaxation. One of them, sildenafil, was found to induce endothelial nitric-oxide synthase and delay preconditioning through an iNOS-dependent pathway
(Salloum et al., 2003). However, proinflammatory iNOS is undesirable when researching new and safe tracheal relax-ants. In contrast, classic xanthine-based PDEIs, theophylline and pentoxifylline, reduce TNF-␣ and iNOS expression and improve indomethacin-induced enteropathy, indicating their anti-proinflammatory merits (Saud et al., 2005). Xanthine-based KMUP-1 and KMUP-3 have been found to have cGMP-enhancing activity and thus may provide cGMP-dependent anti-proinflammatory benefits.
There are two classes of compounds: 1) indazole-based YC-1, BAY-41-2272, and sildenafil; and 2) imidazole-based KMUP-1, KMUP-3 (Fig. 1), and 8-Br-cGMP, predominantly mimic the action of cGMP by regulating one to three compo-nents of an endothelial nitric-oxide synthase/sGC/PDE-mixed functional enzyme system. However, in the present study, a cGMP-dependent bronchodilator with anti-inflam-matory activity is desired to suppress TNF-␣-induced bron-chospastic dysfunction. The goal of this study was to develop a xanthine-based sGC activator/cGMP enhancer with tra-cheal relaxant and anti-inflammatory properties but devoid of YC-1’s COX-2 and sildenafil’s iNOS expression activity. So far, in addition to corticosteroids and2-adrenoceptor ago-nists, neither xanthine-based PDEIs nor sGC activators are inhaled to attenuate inflammatory bronchospastic dysfunc-tion.
This study aimed to investigate the anti-proinflammatory effects of xanthine-based KMUP-1 and KMUP-3 on TNF- ␣-induced expression of iNOS/COX-2 and the involvement of sGC/cGMP/PKG in tracheal smooth muscle cells (TSMCs).
Materials and Methods
Animals. Male Wistar rats (250–300 g) were provided by the
National Laboratory Animal Breeding and Research Center (Taipei, Taiwan) and housed under constant temperature and controlled illumination. Food and water were available ad libitum. This study was approved by the Animal Care and Use Committee of the Kaohsi-ung Medical University.
Cell Culture. Rats were injected intraperitoneally with a lethal
dose of pentobarbital. The tracheas were excised and cut longitudi-nally through the cartilage. Using a dissecting microscope, TSM strips were dissected from the surrounding parenchyma. The epithe-lium was removed from the luminal surface, and bands of TSM were gently separated from the underlying connective tissue. The TSM strips were then chopped into small sections (1 mm3) and incubated
in Hanks’ balanced salt solution (138 mM NaCl, 4 mM NaHCO3, 5
mM KCl, 0.3 mM KH2PO4, 0.3 mM Na2HPO4, and 1.0 mM glucose)
Fig. 1. Chemical structures of KMUP-1 and KMUP-3. The shadow ring
indicates imidazole moiety.
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org
with 0.05% elastase type IV and 0.2% collagenase type IV (Invitro-gen, Carlsbad, CA) for 30 min at 37°C with gentle shaking. The solution of dissociated smooth muscle cells was centrifuged (6 min at 500g), and the pellet was resuspended in 1:1 Dulbecco’s modified Eagle’s medium/Ham’s F-12 medium supplemented with 10% fetal bovine serum, 0.244% NaHCO3, and 1% penicillin/streptomycin.
Cells were cultured, with or without KMUP-1, KMUP-3, and other test agents, in 25-cm2flasks at 37°C in humidified air containing 5%
CO2. The medium was changed every 2 to 3 days. Confluent cells
were passaged every third to fifth day onto 100-mm culture dishes after trypsinization. Experiments to measure levels of NO, cGMP, PGE2, and PGI2were performed in cell cultures of 24-well plates
with 10,000 cells per well. Determinations of iNOS, sGC, PKG, PKA, and COX-2 expressions were performed in cell cultures of 100-mm Petri dishes. The cells were used in passages 2 to 6.
Immunofluorescent Stain. According to our previous method
(Wu et al., 2004), TSMCs (5⫻103cells/well) grown in 24-well plates
on sterile 12-mm glass coverslips were rinsed twice with ice-cold phosphate-buffered saline (PBS) and fixed with fixing buffer (3% formaldehyde and 2% sucrose in PBS) for 5 min. Samples were washed with washing buffer (0.1 M glycine in PBS), and the cells were permeabilized for 20 min with permeabilization buffer (10% goat serum and 0.4% Triton X-100 in PBS). Samples were then washed with buffer A (0.2% Triton X-100 and 0.2% bovine serum albumin in PBS) and incubated with fluorescein isothiocyanate-conjugated monoclonal mouse anti-smooth muscle␣-actin antibody for 2 h in a dark room. The resulting samples were washed with buffer A, aqueous fluorescent mounting media were added, and then coverslips were affixed to the slides. Finally, the coverslips were allowed to dry in the dark before viewing.
Protein Extraction and Western Blot Analysis. After
incuba-tion with or without KMUP-1, KMUP-3, and other test agents, the cells were washed rapidly with ice-cold PBS, incubated with protein extraction reagent (Pierce Biotechnology, Inc., Rockford, IL), and then cells were scraped from plates. The samples were sonicated for 10 s three times and centrifuged at 13,000 rpm at 4°C for 30 min. The protein concentrations of supernatants were determined by using bovine serum albumin as the standard. The cell extracts were then boiled in ratio of 4:1 with sample buffer (100 mM Tris, pH 6.8, 20% glycerol, 4% SDS, and 0.2% bromphenol blue). Electrophoresis was performed using 10% SDS-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes (Millipore Corporation, Bil-lerica, MA). The membrane was blocked with Tris-buffered saline (20 mM Tris and 137 mM NaCl, pH 7.6) containing 0.1% Tween 20 (TTBS) and 5% nonfat milk at room temperature for 1 h, washed with TTBS, and then incubated overnight at 4°C in the appropriate primary antibody of COX-2, iNOS, PKARI, PKG1␣1, sGC␣1, and
sGC1. The membranes were washed in TTBS before being
incu-bated with horseradish peroxidase-conjugated antibody against mouse, goat, or rabbit IgG for 1 h. The membrane was then washed in TTBS and developed with the enhanced chemiluminescence for the detection of the specific antigen. The intensity of the bands was quantitated by densitometry.
Cyclic Nucleotide Radioimmunoassay. Intracellular
concen-trations of cAMP and cGMP in TSMC were measured according to our previous reports (Wu et al., 2004; Lin et al., 2006). In brief, cells were grown in 24-well plates (105 cells/well) with or without
KMUP-1, KMUP-3, and other test agents. At confluence, monolayer cells were washed with PBS and then incubated with isoproterenol, KMUP-1, KMUP-3, and zaprinast (10M) for 20 min at 37°C in medium and further incubated in the absence or presence of TNF-␣ (100 ng/ml) for 9 h. The reaction was terminated by replacement of medium with 1 ml of ice-cold 1 N hydrochloric acid. Cell suspensions were sonicated and then centrifuged at 2500 g for 15 min at 4°C. Then, the supernatants were lyophilized, and the concentrations of cAMP and cGMP of each sample were measured using cGMP-125I
and cAMP-125I radioimmunoassay kits (GE Healthcare, Little
Chal-font, Buckinghamshire, UK).
Prostaglandin Radioimmunoreactivity. After treatment with
cGMP-enhancing KMUP-1 and KMUP-3, the culture media were collected. Concentrations of PGE2and 6-keto-PGF1␣ (stable
metab-olite of PGI2) in the culture medium were measured using PGE2and
6-keto-PGF1␣ EIA assay kits (Cayman Chemical Co., Ann Arbor,
MI). All samples were run in duplicate.
Measurement of Nitrite/Nitrate. After treatment with KMUP-1 and KMUP-3, the culture media were collected. Both nitrite and nitrate (stable breakdown product of NO) were measured in cell culture media. All samples were analyzed using nitrite/nitrate col-orimetric assay kits (Cayman Chemical Co.) and run in duplicate.
Drugs and Chemicals. KMUP-1 and KMUP-3 were synthesized
in this laboratory. 8-Br-cGMP, dexamethasone, indomethacin, iso-proterenol, NS-398, Rp-8-CPT-cAMPS, Rp-8-CPT-cGMPS, TNF-␣, and zaprinast were all purchased from Sigma-Aldrich (St. Louis, MO). COX-2, iNOS, sGC␣1, and sGC1 antibodies were obtained
from Santa Cruz Biotechnology (Santa Cruz, CA). PKARI and
PKG1␣1 antibodies were purchased from Calbiochem (San Diego,
CA). All drugs and reagents were dissolved in distilled water unless otherwise noted. KMUP-1 and KMUP-3 (10⫺2M) were dissolved in 10% absolute alcohol, 10% propylene glycol, and 2% 1 N HCl. Dexa-methasone, indomethacin, and zaprinast (10⫺2M) were dissolved in dimethyl sulfoxide. Serial dilutions were made by distilled water.
Statistical Analysis. All data are expressed as the mean⫾ S.E.
Statistical differences were determined by independent and paired Student’s t test in unpaired and paired samples, respectively. When-ever a control group was compared with more than one treated group, the one-way or two-way analysis of variance was used. When the analysis of variance manifested a statistical difference, results were further analyzed with Dunnett’s or Tukey test. A probability value (P value) less than 0.05 was considered to be significant. Analysis of the data and plotting of the figures were done with the aid of SigmaPlot software (version 8.0) and SigmaStat (version 2.03; both from Systat Software, Point Richmond, CA) run on an IBM-compatible computer.
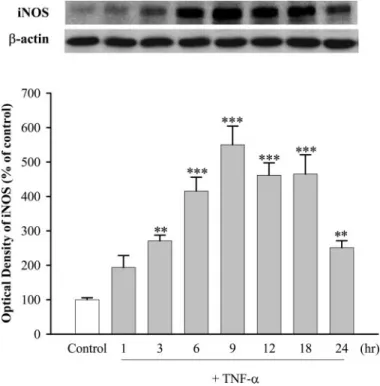
Fig. 2. TNF-␣ modulated the expression of iNOS in cultured TSMCs. Cells treated with TNF-␣ (100 ng/ml) for 1, 3, 6, 9, 12, 18, and 24 h. Values are means⫾ S.E., n ⫽ 6. ⴱⴱ, P ⬍ 0.01, ⴱⴱⴱ, P ⬍ 0.001 versus control.
sGC Activators KMUP-1 and KMUP-3 Reduce TNF-␣-Induced iNOS
979
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org
Results
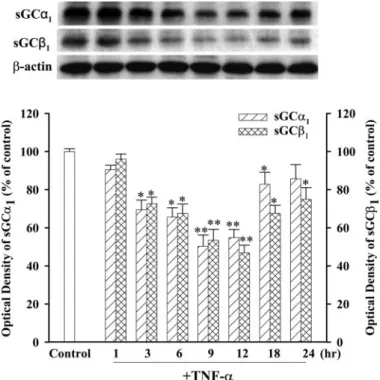
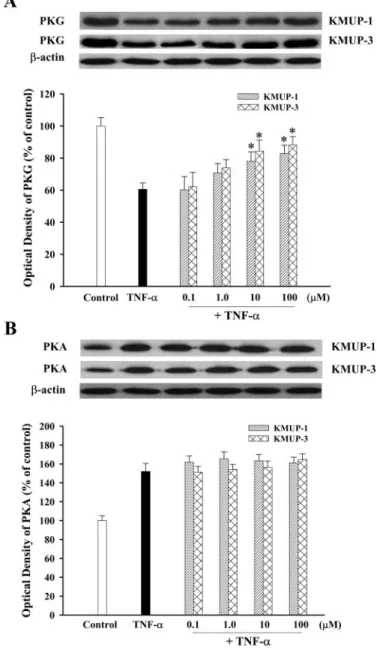
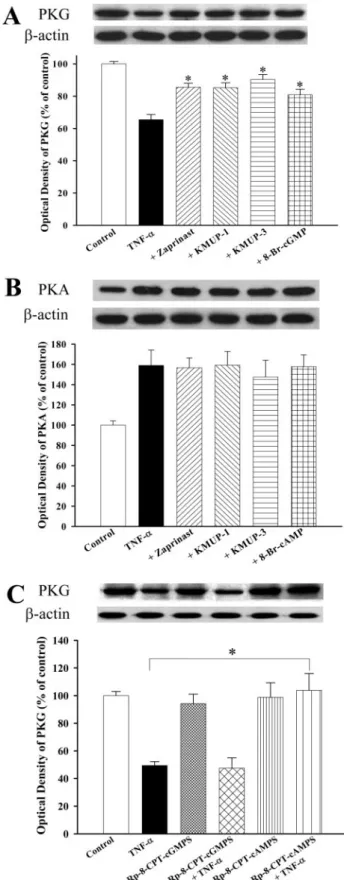
iNOS and sGC Expression. To investigate the effects of TNF-␣ on iNOS and sGC protein, TSMCs were incubated with TNF-␣ (100 ng/ml) for 1, 3, 6, 9, 12, 18, and 24 h, and the levels of iNOS and sGC subunit proteins were measured by immunoblotting. As shown in Fig. 2, exposure of TSMCs to TNF-␣ increased iNOS protein expression in a time-depen-dent manner, with the maximum level evitime-depen-dent at 9 h. KMUP-1 and KMUP-3 (0.01–100M; Fig. 3A) and the PDE5 inhibitor zaprinast and exogenous 8-Br-cGMP (10M; Fig. 3B) inhibited TNF-␣-induced increases of iNOS expression to a similar extent (Fig. 3). In contrast, TNF-␣ decreased the expression of sGC␣1and sGC1proteins in a time-dependent manner, with the maximal inhibition achieved at 9 h (Fig. 4). These inhibitions were reversed by KMUP-1, KMUP-3, zap-rinast, and 8-Br-cGMP at 10M (Fig. 5).
PKG and PKA Expression. TNF-␣ (100 ng/ml)
time-dependently increased PKA within 24 h and significant de-creased the expression of PKG protein between 6 and 9 h
(Fig. 6). The increased expression of PKA protein by TNF-␣ was not further increased by KMUP-1 and KMUP-3 (0.1–100 M); however, decreases of PKG protein expression were reversed by both KMUP-1 and KMUP-3 (Figs. 7 and 8). Zaprinast and 8-Br-cGMP at 10 M also reversed TNF-␣-induced decreases of PKG protein (Fig. 8A), whereas 8-Br-cAMP and zaprinast at 10 M, similar to KMUP-1 and
Fig. 3. Effects of KMUP-1, KMUP-3, and various agents on TNF- ␣-induced iNOS expression in cultured TSMCs. A, concentration-depen-dent effects of KMUP-1 and KMUP-3 (0.01–100 M) on TNF-␣ (100 ng/ml)-induced iNOS expression after 9-h incubation.ⴱ, P ⬍ 0.05; ⴱⴱ, P ⬍ 0.01 versus TNF-␣-treated cells. B, cells pretreated with zaprinast, KMUP-1, KMUP-3, and 8-Br-cGMP (10 M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 9 h. Values are means ⫾ S.E., n ⫽ 6.ⴱⴱ, P ⬍ 0.01 versus TNF-␣-treated cells.
Fig. 4. TNF-␣ modulated the expression of sGC␣1and sGC1in cultured
TSMCs. Cells treated with TNF-␣ (100 ng/ml) for 1, 3, 6, 9, 12, 18, and 24 h. Values are means⫾ S.E., n ⫽ 6. ⴱ, P ⬍ 0.05; ⴱⴱ, P ⬍ 0.01 versus control.
Fig. 5. Effects of KMUP-1, KMUP-3, and various agents on TNF- ␣-inhibited sGC␣1 or sGC1 expression in cultured TSMCs. Cells
pre-treated with zaprinast, KMUP-1, KMUP-3, and 8-Br-cGMP (10M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 9 h. Values are means⫾ S.E., n ⫽ 6. ⴱ, P ⬍ 0.05 versus TNF-␣-treated cells.
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org
KMUP-3, did not affect TNF-␣-induced increases of PKA protein (Fig. 8B). The addition of the PKG inhibitor Rp-8-CPT-cGMPS had no effect on TNF-␣-induced decreases of PKG expression. In contrast, the PKA inhibitor Rp-8-CPT-cAMPS reversed the capacity of TNF-␣ to down-regulate PKG expression (Fig. 8C). These results suggest that the activation of PKA by TNF-␣ down-regulates PKG expression in rat TSMCs.
COX-2 Expression. In the absence of TNF-␣ in rat
TSMCs, the expression of COX-2 showed time-dependent decreases during incubation for 24 h. In the presence of TNF-␣ (100 ng/ml), the expressions of COX-2 were limited to moderate decreases after 6 h and were sustained for 24 h compared with non-TNF-␣ challenge groups (Fig. 9A). It was clear that not KMUP-1 and KMUP-3 (10M) but dexameth-asone (1M) and COX-2 inhibitor NS-398 (10 M) can sig-nificantly inhibit TNF-␣-induced COX-2 expression in TSMCs (Fig. 9B).
Cyclic Nucleotide Levels. Intracellular cGMP produc-tion was decreased, reaching minimal producproduc-tion at 9 h in TSMCs in the presence of TNF-␣ (100 ng/ml). However, in the presence of KMUP-1, KMUP-3, and zaprinast (10M), cGMP was reversed to the basal level (Fig. 10A). In the absence of TNF-␣, the use of KMUP-1, KMUP-3, and isopro-terenol (10 M) in the culture of TSMCs significantly in-creased the production of cAMP compared with the control group. However, in the presence of TNF-␣, KMUP-1, and KMUP-3 could not further increase the production of cAMP compared with the control group (Fig. 10B).
NO Levels Measured by Nitrite/Nitrate. TSMCs were incubated with TNF-␣ for 9 h. Exposure of TSMCs to TNF-␣ (100 ng/ml) led to the accumulation of nitrite and nitrate in the culture medium, indicating the release of NO. 8-Br-cGMP, KMUP-1, KMUP-3, and zaprinast (10M) all
inhib-ited TNF-␣-induced production of nitrite/nitrate, which rep-resented the NO levels (Fig. 11).
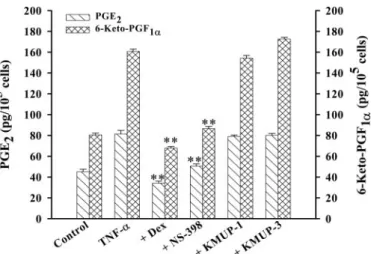
PGs and COX-2 Activities. Incubation of TSMCs with TNF-␣ (100 ng/ml) increased the releases of PGs, PGE2, and 6-keto-PGF1␣, a PGI2 stable metabolite. The greatest in-creases that occurred were measured at 24 h. Dexametha-sone (1M) and NS-398 (10 M), a selective COX-2 inhibitor, significantly inhibited TNF-␣-induced PGE2 and 6-keto-PGF1␣ formation. However, KMUP-1 and KMUP-3 (10 M) did not inhibit the production of PGs, resulting from the activation of COX-2 (Fig. 12).
Discussion
Bronchodilation and immunosuppression or anti-mation activities are important in drug therapy for
inflam-Fig. 7. Effects of KMUP-1 and KMUP-3 on TNF-␣-modulated PKG or
PKA expression in cultured TSMCs. A, concentration-dependent effects of KMUP-1 and KMUP-3 (0.1–100M) on TNF-␣ (100 ng/ml)-regulated PKG expression after 9-h incubation.ⴱ, P ⬍ 0.05 versus TNF-␣-treated cells. B, KMUP-1 and KMUP-3 (0.1–100 M) on TNF-␣ (100 ng/ml)-regulated PKA expression after 24-h incubation. Values are means⫾ S.E., n⫽ 6.
Fig. 6. TNF-␣ modulated the expression of PKG and PKA in cultured
TSMCs. Cells treated with TNF-␣ (100 ng/ml) for 1, 3, 6, 9, 12, 18, and 24 h. Values are means⫾ S.E., n ⫽ 6. ⴱ, P ⬍ 0.05 versus control.
sGC Activators KMUP-1 and KMUP-3 Reduce TNF-␣-Induced iNOS
981
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org
matory bronchospastic dysfunction. Two xanthine- or imida-zole-based derivatives, KMUP-1 and KMUP-3, have been found to increase the expression of cGMP and PKG expres-sion (Wu et al., 2004; Lin et al., 2006), which might make them useful in the treatment of pulmonary airway dysfunc-tions. In comparison, the more previously discovered inda-zole derivatives YC-1 and BAY-41-2272 have been shown to activate sGC by a binding manner, leading to increases of cGMP (Ko et al., 1994; Chang et al., 2004; Balashova et al., 2005). The chemical relationship between KMUP-1/KMUP-3 and YC-1/BAY-41-2272 were thus noted for their sGC acti-vation, PKG expression, and cGMP-enhancing activities.
Exogenous TNF-␣ in vivo has been shown to induce
bron-Fig. 8. Effects of KMUP-1, KMUP-3, and various agents on TNF- ␣-modulated PKG or PKA expression in cultured TSMCs. A, cells pre-treated with zaprinast, KMUP-1, KMUP-3, and 8-Br-cGMP (10M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 9 h. ⴱ, P ⬍ 0.05 versus TNF-␣-treated cells. B, cells pretreated with zaprinast, KMUP-1, KMUP-3, and 8-Br-cAMP (10M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 24 h. C, cells pretreated with Rp-8-CPT-cGMPS and Rp-8-CPT-cAMPS (1M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 9 h. ⴱ, P ⬍ 0.05 versus TNF-␣-treated cells. Values are means⫾ S.E., n ⫽ 6.
Fig. 9. TNF-␣ modulated the expression of COX-2 in cultured TSMCs. A,
cells treated with TNF-␣ (100 ng/ml) for 0, 1, 3, 6, 9, 12, 18, and 24 h. ⴱ, P⬍ 0.05; ⴱⴱ, P ⬍ 0.01 versus respective time without TNF-␣. B, cells pretreated with dexamethasone (Dex, 1M), NS-398 (10 M), KMUP-1 (10M), and KMUP-3 (10 M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 24 h. Values are means ⫾ S.E., n ⫽ 6. ⴱⴱ, P ⬍ 0.01 versus TNF-␣-treated cells.
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org
chial hyper-responsiveness (Hakonarson et al., 1996). In this study, TNF-␣ induced the expression of PKG and cGMP and increased the expression of iNOS and NO. In theory, iNOS would be expected to increase the production of NO and cGMP (Giembycz, 2000; Lincoln et al., 2001). However, cyto-kines have been found in one study to decrease sGC subunit mRNA and protein levels and reduce NO-stimulated sGC enzyme activity (Takata et al., 2001). This reduced genera-tion of cGMP in cytokine-treated cells could be caused by a decrease in sGC, an increase in cGMP phosphodiesterase activity, or an endogenous reaction between NO and O2⫺, leading to the formation of ONOO⫺. This oxidation reaction would affect the activity of sGC (Weber et al., 2001; Yan et al., 2003). In the present study, the time of onset and the duration of the induction of iNOS expression by TNF-␣ were
similar to those of sGC subunit protein levels. Thus, we further investigated the effects of KMUP-1 and KMUP-3 on TNF-␣-induced iNOS and associated sGC expression.
Exogenous cytokine, such as interleukin-1 in pulmonary epithelial cells, has been reported to increase the release of PGE2through the activation of COX-2 (Chang et al., 2004). In this study, exogenous TNF-␣ also caused significant in-creases in the release of PGE2and PGI2, production of cAMP, and expression of PKA in TSMCs (Figs. 8, 10B, and 12). These results seemed to confirm that TNF-␣ is able to elicit the PGE2formation via the activation of COX-2 in TSMCs. KMUP-1 and KMUP-3 cannot further increase PKA and cAMP in the presence of TNF-␣ (Figs. 8B and 10B) and implicate that they might predominantly have the ability to protect against TNF-␣-induced inhibition of PKG (Fig. 13).
Inflammatory response is under the control of cAMP,
Fig. 10. Effects of KMUP-1, KMUP-3, and various agents on TNF- ␣-induced cGMP or cAMP production in cultured TSMCs. A, cells pre-treated with zaprinast, KMUP-1, and KMUP-3 (10M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 9 h. ⴱ, P ⬍ 0.05 versus TNF-␣-treated cells. B, cells pretreated with isoproterenol (Iso), KMUP-1, and KMUP-3 (10M) for 20 min and further incubated with and without TNF-␣ (100 ng/ml) for 24 h. Values are means ⫾ S.E., n ⫽ 3, each conducted in triplicate.ⴱ, P ⬍ 0.05; ⴱⴱ, P ⬍ 0.01 versus control.
Fig. 11. Effects of KMUP-1, KMUP-3, and various agents on TNF- ␣-induced nitrite/nitrate production in cultured TSMCs. Cells pretreated with zaprinast, KMUP-1, KMUP-3, and 8-Br-cGMP (10M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 9 h. Values are means⫾ S.E., n ⫽ 3, each conducted in triplicate. ⴱⴱ, P ⬍ 0.01 versus TNF-␣-treated cells.
Fig. 12. Effects of KMUP-1, KMUP-3, and various agents on TNF-
␣-induced PGE2or 6-keto-PGF1␣ (PGI2stable metabolite) production in
cultured TSMCs. Cells pretreated with dexamethasone (Dex, 1 M), NS-398 (10M), KMUP-1 (10 M), and KMUP-3 (10 M) for 20 min and further incubated with TNF-␣ (100 ng/ml) for 24 h. Values are means ⫾ S.E., n⫽ 3, each conducted in triplicate. ⴱⴱ, P ⬍ 0.01 versus TNF-␣-treated cells.
sGC Activators KMUP-1 and KMUP-3 Reduce TNF-␣-Induced iNOS
983
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org
which is partly regulated by PDE. The role of PDE4 inhibi-tion in anti-inflammainhibi-tion and on COX-2 expression has been described in non-TSMCs (Juergens et al., 1999; Jimenez et al., 2004). PDEIs suppressed IL-1-induced NO release and iNOS mRNA expression (Geng et al., 1998; Beshay and Prud’homme, 2001). Pentoxifylline also affected NO synthe-sis by interferon-␥ (Samardzic et al., 2000). TNF-␣-induced iNOS expressions were reduced by amrinone in cardiomyo-cyte, and such effects were increased by milrinone, a PDE3 and PDE4 inhibitor (Chanani et al., 2002). On the other hand, 8-Br-cGMP augmented NO synthesis in IL-1 -stimu-lated articular chondrocytes and cardiac myocytes (Geng et al., 1998). The increase of cGMP also augmented IL-1-in-duced iNOS expression in vascular smooth muscle cells (In-oue et al., 1995). In addition, 8-Br-cGMP inhibited iNOS expression and NO production in LPS-activated macro-phages (Pang and Hoult, 1997). We observed that 8-Br-cGMP attenuated the expression of iNOS in TSMCs. These evi-dences suggest that the effects of cGMP on iNOS seem to be cell- and probably stimulus-dependent.
We investigated whether the effects of TNF-␣ on TSMCs could be prevented by cGMP enhancers. As shown in Fig. 10A, pretreatment with KMUP-1, KMUP-3 and zaprinast all protected against TNF-␣-induced decreases of cGMP. We further investigated whether exposure of TSMCs to TNF-␣ could affect the expression of PKG protein by cGMP enhanc-ers. As expected, incubation of TSMCs with TNF-␣ decreased the level of PKG, which was reversed by treatment with KMUP-1 and KMUP-3 (Fig. 7A).
In certain physiological and pathophysiological circum-stances, cAMP may act through PKG, showing their cross-interaction. Although PKG is relatively specific for cGMP over cAMP, the basal cAMP concentration in smooth muscle is typically 5 to 6 times higher than the concentration of cGMP, which allows cross-activation when cAMP is elevated. Conversely, in some instances, PKA may be cross-activated by cGMP (Shabb, 2001). In the presence of TNF-␣, the ex-pression of PKG decreased to the minimum at 9 h and then returned to basal level; in contrast, cAMP increased gradu-ally and time-dependently. Thus, we have further confirmed previous findings (Browner et al., 2004) that elevation of cAMP but not cGMP by pro-inflammatory mediators medi-ates the PKG down-regulation in rat TSMCs (Figs. 8C and 13). Here, KMUP-1 and KMUP-3 were suggested to inhibit iNOS and associated ONOO⫺formation, similar to that by aminoguanidine (Ruetten and Thiemermann, 1996), and to increase cGMP by activating sGC and inhibiting PDE (Lin et al., 2006). It is noteworthy that both elevated cAMP- and TNF␣-induced decreases of PKG could be prevented or re-versed by KMUP-1 and KMUP-3 (Fig. 13).
COX-2 is up-regulated by inflammatory stimuli such as cytokines, including TNF-␣, and it contributes to the patho-genesis of inflammation (Laporte et al., 1998). Although COX-2 is generally believed to be inducible, there is evidence that COX-2 is expressed constitutively and the expression is low under physiological conditions (Baber et al., 2003). In this study, the expression of COX-2 protein without TNF-␣ challenge sharply decreased in a time-dependent manner. In contrast, in the presence of TNF-␣, the expression of COX-2 was limited to moderate decreases. However, at a respective incubation time of TSMCs, the expression of COX-2 by TNF-␣ was significantly different from that of nontreatment (Fig. 9A). Accompanied with TNF-␣-induced COX-2 expres-sion in TSMCs, we further investigated whether exposure of TSMCs to TNF-␣ could elicit the production of PGE2 and PGI2. As expected, incubation of TSMCs with TNF-␣ in-creased PGE2 and 6-keto-PGF1␣ (PGI2 stable metabolite). Preincubation of TSMCs with NS-398 (a selective COX-2 inhibitor) caused a marked reduction in TNF-␣-induced PGs, the products of activated COX-2. However, KMUP-1 and KMUP-3 had no significant effects on the expression of COX-2 and the production of PGE2 and PGI2 after TNF-␣ challenge.
In summary, our results indicate that TNF-␣ significantly increases iNOS expression and NO formation in TSMCs. Thus, overproduction of NO and accompanied oxidative re-action were suggested to decrease sGC expression, cGMP production and PKG protein levels (Weber et al., 2001; Mu-nzel et al., 2003; Yan et al., 2003). TNF-␣ also induces ex-pression of COX-2 and productions of PGE2and PGI2, lead-ing to increases in cAMP and PKA. KMUP-1, KMUP-3, and 8-Br-cGMP protect against TNF-␣-induced increases in ex-pression of iNOS and PKA and production of NO. They fur-ther protect against decreases in expression of sGC and PKG in this study. These findings suggest that they might possess iNOS inhibition-associated anti-proinflammatory properties, unrelated to inhibition on COX-2 expression, in TSMCs. KMUP-1 and KMUP-3 did not induce the iNOS expression activity, which was able to be shown by sildenafil (Salloum et al., 2003). Unlike YC-1 in pulmonary epithelial cells (Chang et al., 2004), KMUP-1 and KMUP-3 were not found to induce
Fig. 13. Proposed mechanisms of action of KMUP-1 and KMUP-3 on
TNF-␣-induced inflammatory responses in rat tracheal smooth muscle cells.
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org
the activation of COX-2 in TSMCs. Similar to that with 2-adrenergic agonist, pretreatment of TSMCs with KMUP-1 and KMUP-3 had no significant effects on TNF-␣-induced expression of COX-2 and production of PGE2 and PGI2 (Shore, 2002). However, elevation of cAMP by PGE2 can desensitize the airway response to2-adrenergic agonist by inhibiting cGMP and PKG (Laporte et al., 1998). We thus suggest that KMUP-1 and KMUP-3 might reverse this de-sensitization via enhancing cGMP, sGC activation, and PKG expression.
In conclusion, in the presence of TNF-␣ in TSM, KMUP-1 and KMUP-3 might modulate the cross-action between PKA and PKG by activating sGC/cGMP/PKG pathway without the involvement of COX-2. KMUP-1 and KMUP-3 might reduce cytokine-induced proinflammation and limit the risk of fur-ther worsening of pulmonary dysfunction.
References
Baber SR, Champion HC, Bivalacqua TJ, Hyman AL, and Kadowitz PJ (2003) Role of cyclooxygenase-2 in the generation of vasoactive prostanoids in the rat pulmo-nary and systemic vascular beds. Circulation 108:896 –901.
Balashova N, Chang FJ, Lamothe M, Sun Q, and Beuve A (2005) Characterization of a novel type of endogenous activator of soluble guanylyl cyclase. J Biol Chem
280:2186 –2196.
Beshay E, and Prud’homme GJ (2001) Inhibitors of phosphodiesterase isoforms III or IV suppress islet-cell nitric oxide production. Lab Investig 81:1109 –1117. Browner NC, Sellak H, and Lincoln TM (2004) Downregulation of cyclic
GMP-dependent protein kinase expression by inflammatory cytokines in vascular smooth muscle cells. Am J Physiol 287:C88 –C96.
Caramori G and Adcock I (2003) Pharmacology of airway inflammation in asthma and COPD. Pulm Pharmacol Ther 16:247–277.
Chanani NK, Cowan DB, Takeuchi K, Poutias DN, Garcia LM, del Nido PJ, and McGowan FX Jr (2002) Differential effects of amrinone and milrinone upon myo-cardial inflammatory signaling. Circulation 106:I284 –I289.
Chang MS, Lee WS, Chen BC, Sheu JR, and Lin CH (2004) YC-1-induced cycloox-ygenase-2 expression is mediated by cGMP-dependent activations of Ras, phos-phoinositide-3-OH-kinase, Akt, and nuclear factor-B in human pulmonary epi-thelial cells. Mol Pharmacol 66:561–571.
Chen H, Tliba O, Van Besien CR, Panettieri RA Jr, and Amrani Y (2003) TNF-␣ modulates murine tracheal rings responsiveness to G-protein-coupled receptor agonists and KCl. J Appl Physiol 95:864 – 872.
Doggrell SA (2005) Clinical potential of nitric oxide-independent soluble guanylate cyclase activators. Curr Opin Investig Drugs 6:874 – 878.
Friebe A, Mullershausen F, Smolenski A, Walter U, Schultz G, and Koesling D (1998) YC-1 potentiates nitric oxide- and carbon monoxide-induced cyclic GMP effects in human platelets. Mol Pharmacol 54:962–967.
Geng Y, Zhou L, Thompson WJ, and Lotz M (1998) Cyclic GMP and cGMP-binding phosphodiesterase are required for interleukin-1-induced nitric oxide synthesis in human articular chondrocytes. J Biol Chem 273:27484 –27491.
Giembycz MA (2000) Phosphodiesterase 4 inhibitors and the treatment of asthma: where are we now and where do we go from here? Drugs 59:193–212.
Hakonarson H, Herrick DJ, Serrano PG, and Grunstein MM (1996) Mechanism of cytokine-induced modulation of beta-adrenoceptor responsiveness in airway smooth muscle. J Clin Investig 97:2593–2600.
Inoue T, Fukuo K, Nakahashi T, Hata S, Morimoto S, and Ogihara T (1995) cGMP upregulates nitric oxide synthase expression in vascular smooth muscle cells.
Hypertension 25:711–714.
Jimenez MD, Martin MJ, Alarcon de la Lastra C, Bruseghini L, Esteras A, Herrerias JM, and Motilva V (2004) Role ofL-arginine in ibuprofen-induced oxidative stress and neutrophil infiltration in gastric mucosa. Free Radic Res 38:903–911. Juergens UR, Stober M, and Vetter H (1999) Reproterol—a monomolecular
combi-nation of orciprenaline and theophylline: novel aspects of its mode of action in asthma. Respiration 66:220 –224.
Ko FN, Wu CC, Kuo SC, Lee FY, and Teng CM (1994) YC-1, a novel activator of platelet guanylate cyclase. Blood 84:4226 – 4233.
Laporte JD, Moore PE, Panettieri RA, Moeller W, Heyder J, and Shore SA (1998)
Prostanoids mediate IL-1-induced beta-adrenergic hyporesponsiveness in human airway smooth muscle cells. Am J Physiol 275:L491–L501.
Lin RJ, Wu BN, Lo YC, An LM, Dai ZK, Lin YT, Tang CS, and Chen IJ (2006) A xanthine-based epithelium-dependent airway relaxant KMUP-3 increases respi-ratory performance and protects against TNF-␣-induced tracheal contraction, involving NO release and expression of cGMP and PKG. J Pharmacol Exp Ther
316:709 –717.
Lin RJ, Wu BN, Lo YC, Shen KP, Lin YT, Huang CH, and Chen IJ (2002) KMUP-1 relaxes rabbit corpus cavernosum smooth muscle in vitro and in vivo: involvement of cyclic GMP and K⫹channels. Br J Pharmacol 135:1159 –1166.
Lincoln TM, Dey N, and Sellak H (2001) Invited review: cGMP-dependent protein kinase signaling mechanisms in smooth muscle: from the regulation of tone to gene expression. J Appl Physiol 91:1421–1430.
Mendez-Samperio P, Trejo A, and Miranda E (2006) Activation of ERK1/2 and TNF-alpha production are mediated by calcium/calmodulin, and PKA signaling pathways during Mycobacterium bovis infection. J Infect 52:147–153.
Mohr LC (2004) Hypersensitivity pneumonitis. Curr Opin Pulm Med 10:401– 411. Munzel T, Feil R, Mulsch A, Lohmann SM, Hofmann F, and Walter U (2003)
Physiology and pathophysiology of vascular signaling controlled by guanosine 3⬘,5⬘-cyclic monophosphate-dependent protein kinase. Circulation 108:2172–2183. Pang L and Hoult JR (1997) Repression of inducible nitric oxide synthase and cyclooxygenase-2 by prostaglandin E2 and other cyclic AMP stimulants in J774 macrophages. Biochem Pharmacol 53:493–500.
Renauld JC (2001) New insights into the role of cytokines in asthma. J Clin Pathol
54:577–589.
Ruetten H and Thiemermann C (1996) Prevention of the expression of inducible nitric oxide synthase by aminoguanidine or aminoethyl-isothiourea in macro-phages and in the rat. Biochem Biophys Res Commun 225:525–530.
Salloum F, Yin C, Xi L, and Kukreja RC (2003) Sildenafil induces delayed precon-ditioning through inducible nitric oxide synthase-dependent pathway in mouse heart. Circ Res 92:595–597.
Samardzic T, Stosic-Grujicic S, Maksimovic D, Jankovic V, and Trajkovic V (2000) Differential regulation of nitric oxide production by increase of intracellular cAMP in murine primary fibroblasts and L929 fibrosarcoma cell line. Immunol Lett
71:149 –155.
Saud B, Nandi J, Ong G, Finocchiaro S, and Levine RA (2005) Inhibition of TNF-␣ improves indomethacin-induced enteropathy in rats by modulating iNOS expres-sion. Dig Dis Sci 50:1677–1683.
Shabb JB (2001) Physiological substrates of cAMP-dependent protein kinase. Chem
Rev 101:2381–2411.
Shore SA (2002) Cytokine regulation of-adrenergic responses in airway smooth muscle. J Allergy Clin Immunol 110:S255–S260.
Takata M, Filippov G, Liu H, Ichinose F, Janssens S, Bloch DB, and Bloch KD (2001) Cytokines decrease sGC in pulmonary artery smooth muscle cells via NO-dependent and NO-inNO-dependent mechanisms. Am J Physiol 280:L272–L278. Teixeira CE, Priviero FB, Todd J Jr, and Webb RC (2006) Molecular mechanisms
underlying rat mesenteric artery vasorelaxation induced by the NO-independent soluble guanyl cyclase stimulators BAY 41-2272 and YC-1. J Pharmacol Exp Ther
317:258 –266.
Watkins DN, Peroni DJ, Lenzo JC, Knight DA, Garlepp MJ, and Thompson PJ (1999) Expression and localization of COX-2 in human airways and cultured airway epithelial cells. Eur Respir J 13:999 –1007.
Weber M, Lauer N, Mulsch A, and Kojda G (2001) The effect of peroxynitrite on the catalytic activity of soluble guanylyl cyclase. Free Radic Biol Med 31:1360 –1367. Wu BN, Lin RJ, Lin CY, Shen KP, Chiang LC, and Chen IJ (2001) A xanthine-based KMUP-1 with cyclic GMP enhancing and K⫹channels opening activities in rat aortic smooth muscle. Br J Pharmacol 134:265–274.
Wu BN, Lin RJ, Lo YC, Shen KP, Wang CC, Lin YT, and Chen IJ (2004) KMUP-1, a xanthine derivative, induces relaxation of guinea-pig isolated trachea: the role of the epithelium, cyclic nucleotides and K⫹channels. Br J Pharmacol 142:1105– 1114.
Wu BN, Tu HF, Welsh DG, and Chen IJ (2005) KMUP-1 activates BKCachannels in
basilar artery myocytes via cyclic nucleotide-dependent protein kinases. Br J
Pharmacol 146:862– 871.
Yan C, Kim D, Aizawa T, and Berk BC (2003) Functional interplay between angio-tensin II and nitric oxide: cyclic GMP as a key mediator. Arterioscler Thromb Vasc
Biol 23:26 –36.
Address correspondence to: Dr. Ing-Jun Chen, Department and Graduate Institute of Pharmacology, College of Medicine, Kaohsiung Medical Univer-sity, 100 Shih-Chuan 1st Road, Kaohsiung 807, Taiwan. E-mail: ingjun@ kmu.edu.tw
sGC Activators KMUP-1 and KMUP-3 Reduce TNF-␣-Induced iNOS
985
at Kaohsiung Med Univ Lib on September 27, 2012
molpharm.aspetjournals.org