國立臺灣大學生物資源暨農學院昆蟲學系 碩士論文
Department of Entomology
College of Bioresources and Agriculture National Taiwan University
Master Thesis
重組鞘蛋白抑制粉蝨傳播番茄黃化捲葉泰國病毒
Recombinant Coat Protein Suppress Whitefly Transmission of Tomato yellow leaf curl Thailand virus
曾建豪
Chien-Hao Tseng
指導教授:蔡志偉 博士 Advisor: Chi-Wei Tsai, Ph.D.
中華民國 104 年 7 月
July 2015
誌謝
時光荏苒,六年來於臺大昆蟲系的生活點滴歷歷在目。碩士班修業期間承蒙 蔡志偉老師的指導,得以順利完成論文。其次,非常感謝在研究過程中提供專業 協助的黃莉欣博士、吳岳隆老師及張慧羽老師。感謝黃博士提供實驗所需的病毒,
以及幫我寫出國開會的推薦信,讓我可以順利拿到補助參加國際研討會;感謝吳 岳隆老師在專題討論課的指導,以及提供實驗儀器及耗材讓我能順利完成實驗;
感謝張慧羽老師幫我修改許多上台報告的講稿,使我獲益良多。此外,非常感謝 兩位口試委員陳宗祺老師及張雅君老師對我論文的指導及修改。
感謝病媒研究室的敏廷學姊、佩芯學姊、崧夏學長、宣任學長、鑫餘學長、
仁弗、芊桂、英迪、芸嘉的陪伴與鼓勵,讓我在研究期間備感溫暖。特別感謝佩 芯學姊在實驗技巧上的教導,每次實驗遇到任何困難,佩芯學姊總是能在第一時 間給予建議與幫助。此外,更要感謝昆蟲所同學一路以來的陪伴與支援,每次實 驗耗材用罄時,總是能鼎力相助。求學生涯中曾經幫助我的你們都是我的貴人,
因為你們的幫助,這篇論文才能順利完成。
最後要特別我的家人,他們總是無條件支持我的決定,在我開心時一同分享
喜悅,在我低潮時為我加油打氣,你們的鼓勵是我研究生涯中最堅強的後盾。「莫
忘初衷」是送給自己的話,期許未來能成為更好的人,往夢想大步前進。
中文摘要
粉蝨不僅藉由取食造成植株的危害,更做為媒介昆蟲傳播植物病毒。菸草粉 蝨傳播的 begomoviruses 威脅世界上許多作物的栽培。番茄黃化捲葉泰國病毒 (Tomato yellow leaf curl Thailand virus, TYLCTHV) 及番茄捲葉台灣病毒 (Tomato leaf curl Taiwan virus, ToLCTWV) 是臺灣番茄田間最常見的兩種病毒。鞘蛋白 (coat protein, CP) 是 begomoviruses 唯一的結構蛋白,CP 被推測與菸草粉蝨中腸 推定的受體 (receptor) 有交互作用。In vivo binding assay 結果顯示 recombinant TYLCTHV CP 與菸草粉蝨 (Bemisia tabaci) 的中腸細胞有專一性的結合。本研究 的目的是探討菸草粉蝨獲得病毒前先餵食 recombinant TYLCTHV CP 是否會競爭 粉蝨中腸推定的受體,進而降低其獲得 TYLCTHV 病毒量與粉蝨傳播 TYLCTHV 的能力。實驗結果顯示先餵食 recombinant TYLCTHV CP 會降低 TYLCTHV 在 粉蝨中腸細胞內的累積程度,但是卻沒有降低粉蝨體內的 TYLCTHV 病毒量。此 外,先餵食 recombinant TYLCTHV CP 會抑制粉蝨傳播 TYLCTHV。本研究也利 用相似的實驗來探討先餵食 recombinant TYLCTHV CP (homologous to ToLCTWV CP) 是否也會降低粉蝨中腸細胞內 ToLCTWV 的累積程度與粉蝨體內的病毒量,
以及抑制粉蝨傳播 ToLCTWV。實驗結果指出先餵食 recombinant TYLCTHV CP 不 會 降 低 ToLCTWV 在 菸 草 粉 蝨 中 腸 細 胞 內 的 累 積 程 度 與 粉 蝨 體 內 的 ToLCTWV 病毒量。此外,先餵食 recombinant TYLCTHV CP 也不會抑制菸草粉 蝨傳播 ToLCTWV。本研究證實 TYLCTHV CP 做為 attachment protein 的功能並 且能抑制菸草粉蝨傳播 TYLCTHV。此外,餵食 recombinant TYLCTHV CP 無法 有效降低菸草粉蝨傳播 ToLCTWV 的結果也顯示 TYLCTHV CP 與菸草粉蝨中腸 的 receptor 具有專一性。了解 CP 與菸草粉蝨中腸的交互作用不僅可以揭開菸草 粉蝨傳播 begomoviruses 的機制,日後進一步的研究更能幫助我們發展對抗病害 的防治策略。
關鍵詞:begomoviruses、菸草粉蝨、重組病毒蛋白、病媒傳播、免疫螢光分析
ABSTRACT
Whiteflies not only damage plants through feeding but also serve as vectors transmitting plant viruses. Whitefly-transmitted begomoviruses threaten a wide range of crops worldwide. Tomato yellow leaf curl Thailand virus (TYLCTHV) and Tomato leaf curl Taiwan virus (ToLCTWV) are two most prevalent viruses in tomato fields in
Taiwan. Coat protein (CP), the only structural protein of begomoviruses, is hypothesized to interact with a putative receptor in the midgut of Bemisia tabaci. In vivo binding assay demonstrated recombinant TYLCTHV CP specifically bound to the midgut cells of B. tabaci. The objective of this study was to determine if preacquisition of TYLCTHV CP competes the putative receptor in the midgut of B. tabaci hence suppresses the acquisition and vector transmission of TYLCTHV. Preacquisition of recombinant TYLCTHV CP decreased the accumulation level of TYLCTHV in the midgut cells of B. tabaci but did not reduce the acquisition of TYLCTHV in B. tabaci.
Further, preacquisition of recombinant TYLCTHV CP suppressed the whitefly transmission of TYLCTHV. Similar experiments were conducted to investigate if preacquisition of TYLCTHV CP (homologous to ToLCTWV CP) would reduce virus accumulation and acquisition in B. tabaci, and the whitefly transmission of ToLCTWV.
The results demonstrated that preacquisition of recombinant TYLCTHV CP did not influence the accumulation and acquisition of ToLCTWV in B. tabaci and did neither reduce the whitefly transmission of ToLCTWV. In conclusion, TYLCTHV CP functioned as an attachment protein and preacquisition of TYLCTHV CP reduced the whitefly transmission of TYLCTHV. The failure that using TYLCTHV CP to inhibit the whitefly transmission of ToLCTWV suggested that the interaction between TYLCTHV CP and the putative receptor in the midgut of B. tabaci is specific. Understanding the
CP-midgut interaction would unveil the mechanism of whitefly transmission of begomoviruses and warrant further study to develop strategies for disease control.
Key words: begomoviruses, Bemisia tabaci, recombinant viral protein, vector transmission, immunofluorescence assay
CONTENTS
誌謝 ... ii
中文摘要 ... iii
ABSTRACT ...iv
CONTENTS ...vi
LIST OF FIGURES ... viii
Chapter 1 Introduction ... 1
Chapter 2 Materials and methods ... 4
2.1 Insects, viruses, and plants ... 4
2.2 Construction of expression plasmids of viral proteins ... 4
2.3 Expression and purification of recombinant proteins ... 5
2.4 SDS-PAGE... 6
2.5 In vivo binding assay ... 6
2.6 Competition for the virus binding site in midguts ... 7
2.7 Inhibition of TYLCTHV acquisition ... 8
2.8 Inhibition of TYLCTHV transmission ... 9
2.9 Inhibition of ToLCTWV acquisition and transmission ... 9
2.10 Statistics ... 11
Chapter 3 Results ... 12
3.1 Expression and purification of recombinant proteins ... 12
3.2 TYLCTHV CP bound to the midgut cells of B. tabaci... 12
3.3 TYLCTHV CP competed with TYLCTHV for the midgut binding site in B. tabaci ... 13 3.4 Influence of TYLCTHV CP on the acquisition of TYLCTHV by B. tabaci14
3.5 Influence of TYLCTHV CP on whitefly transmission of TYLCTHV ... 14 3.6 TYLCTHV CP did not compete with ToLCTWV for the midgut binding site in B. tabaci ... 15 3.7 Influence of TYLCTHV CP on the acquisition of ToLCTWV by B. tabaci16 3.8 Influence of TYLCTHV CP on whitefly transmission of ToLCTWV ... 16 3.9 Inhibition efficiency of TYLCTHV CP on whitefly transmission of TYLCTHV and ToLCTWV ... 17 Chapter 4 Discussion... 18 REFERENCES ... 22
LIST OF FIGURES
Figure 1. Analysis of recombinat viral proteins by SDS-PAGE. ... 28 Figure 2. In vivo binding assay. ... 29 Figure 3. Recombinant TYLCTHV CP competed with TYLCTHV for the midgut binding site in Bemisia tabaci... 30 Figure 4. Relative abundance of TYLCTHV in Bemisia tabaci after feeding recombinant viral proteins and the virus sequentially. ... 31 Figure 5. Transmission rate of TYLCTHV after Bemisia tabaci feeding recombinant viral proteins and the virus sequentially. ... 32 Figure 6. Recombinant TYLCTHV CP competed with ToLCTWV for the midgut binding site in Bemisia tabaci... 33 Figure 7. Relative abundance of ToLCTWV in Bemisia tabaci after feeding recombinant TYLCTHV CP and the virus sequentially. ... 34 Figure 8. Transmission rate of ToLCTWV after Bemisia tabaci feeding recombinant TYLCTHV CP and the virus sequentially. ... 35 Figure 9. Inhibition efficiency of TYLCTHV CP on whitefly transmission of TYLCTHV and ToLCTWV. ... 36
Chapter 1 Introduction
Tomato yellow leaf curl disease associated with a group of closely related tomato yellow leaf curl viruses threatens the tomato production in tropical and subtropical areas (Czosnek and Ghanim, 2011). Typical foliar symptoms of infected tomato plants include yellowing leaflets, upward curling leaf edges, and leaf mottling. Severe growth reductions such as reduced size of leaves, stunting, and decreased tomato yields are also found on infected tomato plants (Navas-Castillo et al., 2011). The group of tomato yellow leaf curl viruses belongs to the genus Begomovirus of the family Geminiviridae (Brown et al., 2012). Predominant tomato-infecting begomoviruses detected in Taiwan are Tomato yellow curl Thailand virus (TYLCTHV) and Tomato leaf curl Taiwan virus (ToLCTWV). Most infected tomato plants in Taiwan were either infected with
TYLCTHV or mix-infected with TYLCTHV and ToLCTWV (Tsai et al., 2011).
Begomoviruses can be divided into two groups, monopartite and bipartite, based on their genomic structures (Brown et al., 2012). Bipartite begomoviruses contain two circular, single-stranded DNA segments (referred to as DNA-A and DNA-B) with similar size; monopartite begomoviruses contain single DNA segment homologous to DNA-A. In bipartite begomoviruses, there are six genes in DNA-A. AV1 and AV2 genes are on the viral sense strand. AV1 gene encodes the coat protein (CP) which is the only structural protein of begomoviruses (Wartig et al., 1997). CP is responsible for not only viral encapsulation but also virus movement and vector transmission (Czosnek, 2008;
Yaakov et al., 2011). The exact function of AV2 gene is uncertain (Fondong, 2013). AC1, AC2, AC3, and AC4 genes on the complementary sense strand encode replication associated protein, transcription activator protein, replication enhancer protein, and RNA silencing suppressor, respectively (Fondong, 2013). The BC1 and BV1 genes on
DNA-B encode the movement protein (MP) and nuclear shuttle protein (NSP), respectively. Both MP and NSP play vital roles in the pathogenesis of bipartite begomoviruses in host plants. NSP binds to viral DNA and transport it from the nucleus to the cytoplasm (Gafni and Bernard, 2002). MP alters the structure of plasmodesmata and thereby facilitates the viral DNA transport to adjacent cells (Lazarowitz and Beachy, 1999). Both structural and non-structural proteins are important for morphogenesis and pathogenesis of viruses.
Begomoviruses are transmitted by Bemisia tabaci and barely by mechanical inoculation but not transmitted through seeds (Ghanim et al., 1998; Brown and Czosnek, 2002). B. tabaci belongs to the family Aleyrodidae of the order Hemiptera. Recently, phylogenetic analyses indicated that B. tabaci is composed of at least 28 morphologically indistinguishable species (Dinsdale et al., 2010; Hu et al., 2011).
Middle East Asia Minor 1 (MEAM 1), previously referred to as B. tabaci B biotype, is one of the most destructive whitefly species in the fields worldwide (Polston et al., 2014). The expansion of whitefly population to a new area always accompanies with the invasion of tomato begomoviruses.
B. tabaci complex is one of the most important agricultural pests because it causes
feeding damages and transmits viral pathogens. Tomato yellow leaf curl virus (TYLCV) is transmitted by B. tabaci in a persistent-circulative manner (Hogenhout et al., 2008;
Ghanim, 2014). TYLCV is ingested through whitefly feeding on virus-infected plants, and the virions enter the midgut lumen. CP is the only protein composing the TYLCV virion, and it interacts with a putative receptor in the midguts of B. tabaci (Ghanim, 2014). Afterwards, TYLCV enters the midgut cells probably by the endocytosis (Czosnek et al., 2002). The virions then translocate from the midgut tissue to salivary glands across hemolymph (Ghanim et al., 2001a). Once virus-infected whiteflies feed
on healthy plants, the virus will be transmitted to the plants through the salivary secretions during feeding.
Understanding the mechanism of vector transmission of plant pathogens is the key to develop effective control strategies to plant diseases. Wang et al. (2014) reported that TYLCV CP directly binds to the midgut cells when recombinant CP was fed to B.
tabaci. It suggests that CP serves as an attachment protein mediating the specific
binding between CP and the midgut receptor of B. tabaci. Tomato yellow leaf curl Sardinia virus (TYLCSV) possessing CP mutants was found nontransmissible (Caciagli
et al., 2009). A specific interaction between TYLCV CP and Heat shock protein 70 (HSP70) of B. tabaci inhibits the whitefly transmission of TYLCV (Gotz et al., 2012).
Since the role of CP is convinced to be an attachment protein, whether CP can be directly used to saturate the virus binding site in the midgut of B. tabaci remains unknown.
The objective of this study was to determine whether TYLCTHV CP saturates the virus binding site in the midgut of B. tabaci hence suppress the whitefly transmission of the virus. Since sequence similarity of TYLCTHV CP and ToLCTWV CP is 74.3%, we further examined whether the homologous CP (TYLCTHV CP) acts as the same function on the suppression of whitefly transmission of ToLCTWV. Understanding the CP-midgut interaction unveils the mechanism of whitefly transmission of begomoviruses and warrants further study to develop strategies for disease control.
Chapter 2 Materials and methods
2.1 Insects, viruses, and plants
B. tabaci MEAM 1, previously referred to as B biotype, was initially collected
from a tomato plant (Solanum lycopersicum) in Tainan, Taiwan. The colony was maintained on Chinese kale (Brassicae oleracea) in whitefly-proof cages (150 meshes, 30×30×30 cm3) under the condition of 28°C, 70% relative humidity, and a 16:8 h (light:dark) photoperiod. A polymerase chain reaction (PCR) assay (Ko et al., 2007) was employed to examine the purity of the colony every three months.
The TYLCTHV- and ToLCTWV-infected tomato plants were provided by AVRDC—The World Vegetable Center and maintained in tomato plants cv. ANT22 by whitefly-mediated passage. All plants used in the experiments (tomato and Chinese kale) were grown from seeds under the same aforementioned condition. Tomato seedlings with three to five leaves were used as test plants for transmission assays.
2.2 Construction of expression plasmids of viral proteins
The open reading frames (ORFs) of TYLCTHV CP, TYLCTHV MP and Banana bunchy top virus (BBTV) CP (GenBank accession numbers: GU723742, GU723754,
and AY534140, respectively) were amplified from total DNA of virus-infected plants by PCR. DNA extraction and PCR were performed using DNeasy Blood & Tissue kit (Qiagen) and KOD plus DNA polymerase (Toyobo) following the manufacturers’
instructions. BBTV is transmitted by aphids in a persistent-circulative manner (Watanabe and Bressan, 2013), so BBTV CP was used as a negative control. The primer set for cloning TYLCTHV CP was 5’-GGGAATTCATGTCGAAGCGTCC
AGCAGA-3’ and 5’-GGCTCGAGATTCGTCACTGAGTCATAGAAA-3’. The primer set for cloning TYLCTHV MP was 5’-GGGAATTCATGGAGTCCAG AACTAACAATA-3’ and 5’-GGCTCGAGTATTTTCTTTGCATTAGAAGAGAC-3’.
The primer set for cloning BBTV CP was 5’-GGGAATTCATGGCTAGGTATCC GAAGAA-3’ and 5’-GGCTCGAGAACATGATATGTAATTCTGTTCTG-3’. EcoRI and XhoI recognition sites underlined were incorporated into the 5’ ends of the forward and reverse primers, respectively, to facilitate directional cloning. The PCR products were ligated into the pET32a vector (Novagen) and then transformed into E. coli strain BL21 (RBC Bioscience). pET32a vector was chosen because of its His tag and S tag sequence for purification and detection. Successful transformations were examined by restriction enzyme digestion and DNA sequencing.
2.3 Expression and purification of recombinant proteins
The transformed E. coli cells were cultured in LB broth (tryptone 10 g/L, yeast extract 5 g/L, NaCl 10 g/L) containing ampicillin (100 μg/mL) at 37°C until the OD600
reached 0.6-0.8. Protein expression was induced by 0.1 mM isopropyl beta-D-thiogalactopyranoside (IPTG). Affinity purification was used to purify the 6xHis-tagged recombinant proteins using Ni Sepharose High Performance (GE Healthcare). Bacterial cells were pelleted and lysed with binding buffer (20 mM sodium phosphate, 20 mM imidazole, 8 M urea, pH 7.4) by ultrasonication. The supernatant were collected after centrifugation (10000 g, 30 min) and applied to the column packed with Ni sepharose. The columns were then washed with binding buffer until the absorbance reached the baseline. The 6xHis-tagged proteins were then eluted with elution buffer (20 mM sodium phosphate, 500 mM imidazole, 8 M urea, pH 7.4). The
eluted proteins were re-purified using the same column and subsequently dialyzed against phosphate buffered saline (PBS) using Slide-A-Lyzer G2 Dialysis Cassette (Thermo Scientific) following the manufacturer’s instructions. Some precipitates occurred after dialysis, and soluble proteins in the supernatant were collected. The concentration of the recombinant proteins was quantified by Pierce 660 nm Protein Assay (Thermo Scientific) following the manufacturer’s instructions. The protein solutions were stored at -80°C for later usage.
2.4 SDS-PAGE
The purity of the recombinant proteins was analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The eluted proteins were separated by electrophoresis in 10% polyacrylamide gels, and stained with Bio-Safe Coomassie (Bio-Rad) following the manufacturer’s instructions.
2.5 In vivo binding assay
To determine if TYLCTHV CP binds to the midgut cells of B. tabaci, an insect feeding assay and an immunofluorescence assay were employed. Adult whiteflies were fed purified recombinant protein (1.8 ng/µl) in artificial feeding medium (PBS, 1 mM EDTA, 15% sucrose, 1% bovine serum albumin [BSA]) through a layer of stretched Parafilm for 24 h. A green food dye (McCormick) was used to ensure that the whiteflies have ingested the medium. Four treatments were as followed: (1) TYLCTHV CP, (2) TYLCTHV MP, (3) BBTV CP, and (4) medium only. BBTV CP was included to test the specificity of viral proteins binding to the midgut cells of B. tabaci. The whiteflies were then dissected in PBS, and the midguts were fixed in 4% paraformaldehyde in PBS for
30 min. After fixation, the fixative was removed by washing three times with PBS. The samples were then permeabilized in PT buffer (PBS, 0.1% Triton X-100) for 30 min.
The midguts were subsequently incubated in blocking buffer (PBS, 1% BSA) for 1 h.
After which, the midguts were incubated in a mouse anti-S tag antiserum (Bioman) at a 1:200 dilution in PBT buffer (PBS, 0.1 % BSA, 0.1% Triton X-100) for 1.5 h. After washed three times with PT buffer, the samples were incubated in a goat anti-mouse IgG antiserum conjugated with Alexa fluor 633 (Molecular Probes) at a 1:200 dilution in PBT buffer for 1 h. The samples were washed again with PT buffer three times. The dissected midguts were mounted in SlowFade Gold antifade reagent with DAPI (Molecular Probes) and examined with a Leica TCS SP5 II confocal laser scanning microscope. Images were collected using the same laser power and gain. The in vivo binding assay was repeated three times, and three replicate insects were examined each time.
2.6 Competition for the virus binding site in midguts
Adult whiteflies were fed recombinant TYLCTHV CP, TYLCTHV MP, or BBTV CP in artificial feeding medium or medium only through membrane feeding for 24 h as described above for the in vivo binding assay. The whiteflies were then transferred to TYLCTHV-infected tomato plants to acquire the virus for another 24 h. Subsequently, the whiteflies were dissected, and the midguts were fixed and permeabilized according to the abovementioned protocols. The midguts were then incubated in a mixture of a mouse anti-S tag antiserum (Bioman) and a rabbit anti-TYLCTHV antiserum (courtesy of Dr. Stephan Winter from Leibniz-Institut DSMZ) at a 1:200 dilution in PBT buffer for 1.5 h. After washed three times with PT buffer, the samples were incubated in a
mixture of a goat anti-mouse IgG antiserum conjugated with Alexa fluor 633 (Molecular Probes) and a goat anti-rabbit IgG antiserum conjugated with Alexa fluor 555 (Molecular Probes) at a 1:200 dilution in PBT buffer for 1 h. The samples were washed again with PT buffer three times. The dissected midguts were mounted in SlowFade Gold antifade reagent with DAPI (Molecular Probes) and examined with a Leica TCS SP5 II laser scanning confocal microscope. Images were collected using the same laser power and gain. The experiments were repeated three times, and three replicate insects were examined each time.
2.7 Inhibition of TYLCTHV acquisition
To access if preacquisition of TYLCTHV CP by whiteflies would inhibit the acquisition of TYLCTHV, a virus quantification assay was employed. Adult whiteflies were fed recombinant TYLCTHV CP, TYLCTHV MP, or BBTV CP in artificial feeding medium or medium only through membrane feeding for 24 h as described above for the in vivo binding assay. The whiteflies were then transferred to TYLCTHV-infected
tomato plants to acquire the virus for another 24 h and subsequently fed a 15% sucrose solution for 6 h to clear the viruses remaining in the gut lumen. For each treatment, 20 adult whiteflies were collected, and their DNA was extracted using DNeasy Blood &
Tissue kit (Qiagen) following the manufacturer’s instructions. TYLCTHV in the tested whiteflies was quantified by real-time PCR. The primer set for TYLCTHV
quantification was 5’-CACGCCCGTCTCGAAAGT-3’ and
5’-GCTGTTGTATGGGCTGTCGAA-3’. The primer set for the quantification of endogenous Heat shock protein 90 (HSP90) gene was 5’-ATCGCCAAATCTGG AACTAAAGC-3’ and 5’-GTGTTTTGAGACGACTGTGACGGTG-3’ (Li et al., 2013).
The reaction was carried out using Fast SYBR Green Master Mix (Applied Biosystems) following the manufacturer’s instructions. The relative abundance of TYLCTHV DNA in the tested whiteflies was normalized to HSP90 gene. The experiments were repeated four times.
2.8 Inhibition of TYLCTHV transmission
To access if preacquisition of TYLCTHV CP by whiteflies would inhibit the whitefly transmission of TYLCTHV, a transmission experiment was employed. Adult whiteflies were fed recombinant TYLCTHV CP, TYLCTHV MP, or BBTV CP in artificial feeding medium or medium only through membrane feeding for 24 h as described above for the in vivo binding assay. The whiteflies were then transferred to TYLCTHV-infected
tomato plants to acquire the virus for another 24 h. After which, five whiteflies were used to inoculate each healthy test plant for a 24 h inoculation access period (IAP). The whiteflies were removed from the test plants after the IAP, and then a systemic insecticide (acetamiprid, diluted 1:1500) was used to kill all remaining whiteflies on the plants. Whitefly-inoculated plants were maintained in insect-proof Bugdorms (47.5 x 47.5 x 93 cm3) (Megaview) within a growth chamber under the same abovementioned conditions for disease development. A PCR assay (Weng et al., 2015) was employed to detect the virus infection in the test plants two weeks after inoculation. Ten replicate test plants were inoculated for each treatment, and the experiments were performed five times.
2.9 Inhibition of ToLCTWV acquisition and transmission
To determine if preacquisition of TYLCTHV CP by whiteflies would reduce the
virus binding to midgut cells, virus acquisition, and whitefly transmission of ToLCTWV, similar experiments were conducted as described above. To examine the competition between TYLCTHV CP and ToLCTWV for the virus binding site in the midgut cells of B. tabaci, adult whiteflies were fed recombinant TYLCTHV CP and ToLCTWV-infected tomato plants successively, and the whiteflies were dissected, fixed, and processed to immunofluorescence assays. The samples were treated with a mixture of a mouse anti-S tag antiserum (Bioman) and a rabbit anti-ToLCTWV antiserum at a 1:200 dilution. Primary antibody binding was detected with a mixture of a goat anti-mouse IgG conjugated with Alexa fluor 633 (Molecular Probes) and a goat anti-rabbit IgG conjugated with Alexa fluor 555 (Molecular Probes) at a 1:200 dilution.
The dissected midguts were examined with confocal microscopy. The experiments were repeated three times, and three replicate inscets were examined each time. ToLCTWV in the tested whiteflies was quantified by real-time PCR. The primer set for ToLCTWV
quantification was 5’-GCGACCCGCCGATATAGTC-3’ and
5’-CAGACGGCGACGAACCTT-3’. The relative abundance of ToLCTWV DNA in the tested whiteflies was normalized to HSP90 gene. The experiments were repeated three times.
To access if preacquisition of TYLCTHV CP would inhibit whitefly transmission of ToLCTWV, adult whiteflies were fed recombinant TYLCTHV CP and ToLCTWV-infected tomato plants successively, and five whiteflies were used to inoculate each healthy test plant. Disease development and PCR assay were conducted as described above. Ten replicate test plants were inoculated for each treatment, and the experiments were performed five times.
2.10 Statistics
Data obtained by real-time PCR were normalized and analyzed by one-way ANOVA, using the differences between the Ct value of TYLCTHV/ToLCTWV and HSP90 gene. Data from the transmission assays were analyzed by χ2 test.
Chapter 3 Results
3.1 Expression and purification of recombinant proteins
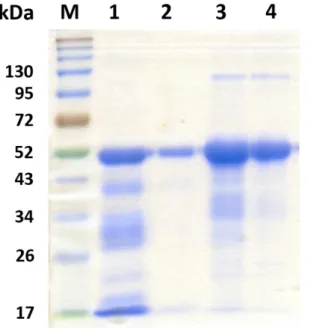
A time course experiment was conducted to determine the period for maximal yield of recombinant viral proteins. Maximal yield was occurred at about 3 h after induction by IPTG, and the recombinant proteins were accumulated in cell pellets (data not shown). After affinity purification of 6xHis-tagged proteins, the purified proteins were examined by SDS-PAGE. The elution of recombinant TYLCTHV CP and BBTV CP showed multiple bands on the polyacrylamide gel (Fig. 1, lanes 1, 3), and a rebinding using the same columns was performed. After the rebinding, the purified protein showed single band on the gel (Fig. 1, lanes 2, 4). The predicted molecular weights of the recombinant TYLCTHV CP and BBTV CP were 50 kDa, containing CP (30 kDa), a His tag (1 kDa), an S tag (1.7 kDa), and Trx tag (12 kDa). The predicted molecular weights of the recombinant TYLCTHV MP was 33 kDa, containing MP (13 kDa), a His tag (1 kDa), an S tag (1.7 kDa), and Trx tag (12 kDa) The respective protein bands matched the predicted sizes (Fig. 1, lanes 2, 4, 5).
3.2 TYLCTHV CP bound to the midgut cells of B. tabaci
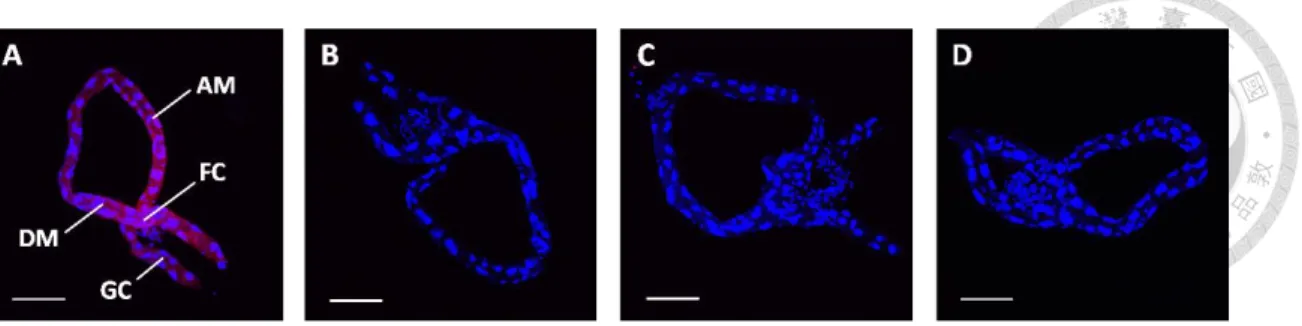
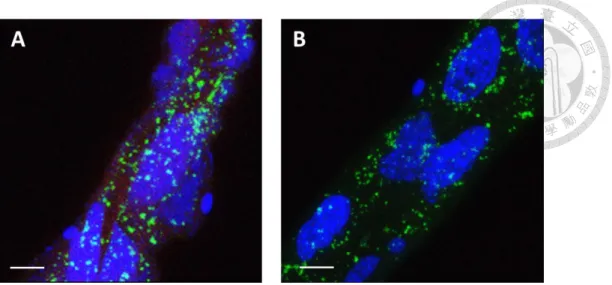
TYLCV CP has been proved to bind to the midgut cells of B. tabaci (Wang et al., 2014), so we conducted an in vivo binding assay to determine if recombinant TYLCTHV CP also binds to the midgut cells of B. tabaci. The midgut and filter chamber were examined in this study because they are the organs that were heavily infected (Ghanim et al., 2001a). Whiteflies fed recombinant TYLCTHV CP, TYLCTHV MP, or BBTV CP were dissected and examined by indirect immunofluorescence assay.
Recombinant TYLCTHV CP bound to the cells of midgut, filter chamber, and gastric
caeca (Fig. 2A). To examine the specificity of TYLCTHV CP-midgut binding, whiteflies were fed recombinant TYLCTHV MP. Recombinant TYLCTHV MP did not bind to the cells of midgut, filter chamber, and gastric caeca (Fig. 2B). Another recombinant protein, BBTV CP, was fed to whiteflies to examine the nonspecific binding of viral CP to the midgut cells of B. tabaci. Recombinant BBTV CP did neither bind to the cells of midgut, filter chamber, and gastric caeca (Fig. 2C). The artificial feeding medium containing no recombinant protein in it was fed to whiteflies to confirm that no self-fluorescence was detected in the midgut of B. tabaci (Fig. 2D). Altogether, the results showed that recombinant TYLCTHV CP specifically bound to the midgut cells of B. tabaci. The finding implied that there was a specific cellular interaction between TYLCTHV CP and a midgut molecule of B. tabaci.
3.3 TYLCTHV CP competed with TYLCTHV for the midgut binding site in B. tabaci
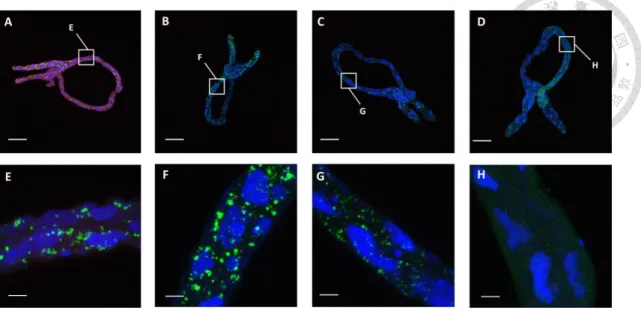
An in vivo binding assay was conducted to determine the ability of TYLCTHV CP to block the midgut binding sites of TYLCTHV in B. tabaci. Recombinant TYLCTHV CP bound to the midgut cells of B. tabaci, and the accumulation of TYLCTHV was also detected in the midguts (Fig. 3A and 3E). To compare TYLCTHV accumulation level in B. tabaci fed recombinant TYLCTHV CP with that in B. tabaci fed other proteins,
whiteflies were fed recombinant TYLCTHV MP or BBTV CP before they fed on TYLCTHV-infected tomato plants. The accumulation of TYLCTHV was also detected in the midguts of B. tabaci, but the fluorescent signals of recombinant TYLCTHV MP (Fig. 3B and 3F) and BBTV CP (Fig. 3C and 3G) were not detected in the midguts.
However, less TYLCTHV accumulated in the midguts when the whiteflies were fed
recombinant TYLCTHV CP prior to the TYLCTHV acquisition compared to those fed recombinant TYLCTHV MP or BBTV CP (Fig. 3E, F, G). Whiteflies fed artificial medium were served as control (Fig. 3D and 3H). The results indicated that preacquisition of TYLCTHV CP blocked the midgut binding sites of TYLCTHV in B.
tabaci hence reduced the binding of TYLCTHV to the midgut cells of B. tabaci.
3.4 Influence of TYLCTHV CP on the acquisition of TYLCTHV by B. tabaci
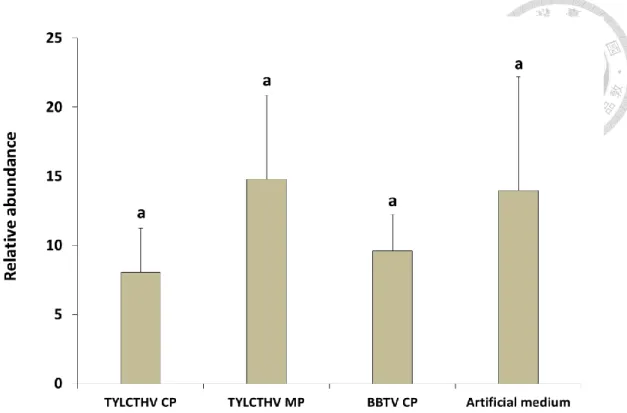
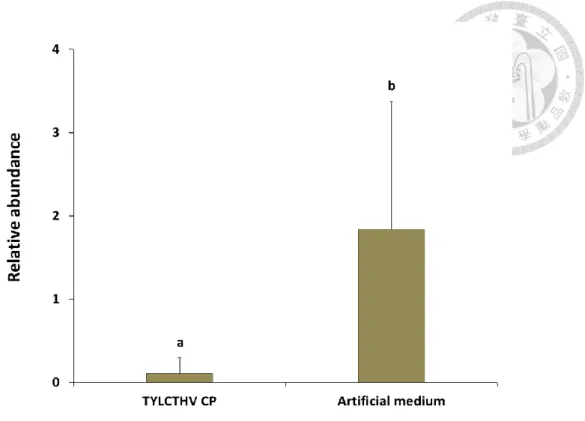
To access if preacquisition of TYLCTHV CP would reduce the acquisition of TYLCTHV by B. tabaci, we employed a virus quantification assay. The relative abundance of TYLCTHV DNA in the tested whiteflies fed recombinant TYLCTHV CP or artificial feeding medium alone prior to TYLCTHV acquisition were not significantly different (ANOVA, P>0.05) (Fig. 4). There were also no significant differences in the relative abundance of TYLCTHV DNA in the tested whiteflies fed recombinant TYLCTHV MP or BBTV CP prior to TYLCTHV acquisition compared to those were without preacquisition of recombinant proteins (ANOVA, P>0.05) (Fig. 4). The results indicated that preacquisition of TYLCTHV CP did not affect the amount of TYLCTHV that the tested whiteflies acquired.
3.5 Influence of TYLCTHV CP on whitefly transmission of TYLCTHV
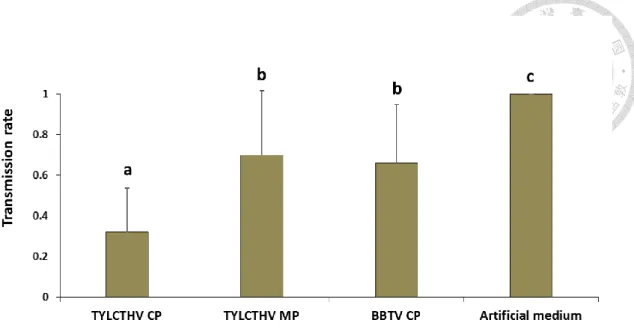
We further conducted transmission experiments to access if preacquisition of TYLCTHV CP would reduce the whitefly transmission of TYLCTHV. The results of
transmission experiments are shown in Fig. 5. The transmission rate of TYLCTHV inoculated by the whiteflies fed artificial feeding medium alone prior to TYLCTHV acquisition was 100%. The transmission rate of TYLCTHV inoculated by the whiteflies fed recombinant TYLCTHV CP prior to TYLCTHV acquisition reduced to 32%
compared to whiteflies fed artificial feeding medium alone (χ2 test, P<0.001). The preacquisition of recombinant TYLCTHV MP and BBTV CP reduced the whitefly transmission rates of TYLCTHV to 52% and 66%, respectively (χ2 test, P<0.001). The results suggested that the preacquisition of TYLCTHV CP would reduce the whitefly transmission rate of TYLCTHV, and the transmission rate of the whiteflies fed either TYLCTHV MP or BBTV CP did not greatly reduced.
3.6 TYLCTHV CP did not compete with ToLCTWV for the midgut binding site in B. tabaci
To determine if preacquisition of TYLCTHV CP could block the midgut binding sites of ToLCTWV in B. tabaci, similar in vivo binding assays were conducted.
Recombinant TYLCTHV CP bound to the midgut cells of B. tabaci, and the accumulation of ToLCTWV was also detected in the midguts (Fig. 6A). The accumulation level of ToLCTWV in the midgut cells of B. tabaci fed recombinant TYLCTHV CP prior to ToLCTWV acquisition was similar to those fed artificial feeding medium alone prior to ToLCTWV acquisition (Fig. 6). The results indicated that the feeding of TYLCTHV CP prior to ToLCTWV acquisition did not block the midgut binding sites of ToLCTWV in B. tabaci.
3.7 Influence of TYLCTHV CP on the acquisition of ToLCTWV by B. tabaci
To access if preacquisition of TYLCTHV CP would reduce the acquisition of B.
tabaci by B. tabaci, we employed a virus quantification assay. The relative abundance
of ToLCTWV DNA in the tested whiteflies fed recombinant TYLCTHV CP or artificial feeding medium alone prior to ToLCTWV acquisition were significantly different (Mann-Whitney test, P<0.05) (Fig. 7). The results indicated that preacquisition of TYLCTHV CP affected the amount of ToLCTWV that the tested whiteflies acquired.
3.8 Influence of TYLCTHV CP on whitefly transmission of ToLCTWV
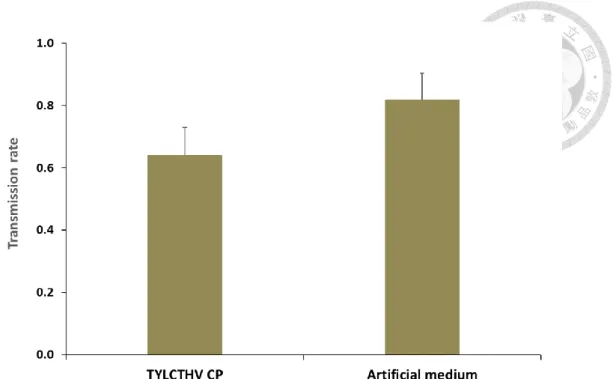
We further conducted transmission experiments to access if preacquisition of TYLCTHV CP would reduce the whitefly transmission of ToLCTWV. The transmission rate of ToLCTWV inoculated by the whiteflies fed artificial feeding medium alone prior to ToLCTWV acquisition was 82% (Fig. 8). The transmission rate of ToLCTWV inoculated by the whiteflies fed recombinant TYLCTHV CP prior to ToLCTWV acquisition reduced to 64% compared to the whiteflies fed artificial feeding medium alone (χ2 test, P<0.05) (Fig. 8). Despite the fact that the transmission rate of ToLCTWV by B. tabaci fed recombinant TYLCTHV CP or artificial feeding medium alone was significantly different, the preacquisition of recombinant TYLCTHV CP did not greatly reduce the whitefly transmission of ToLCTWV. The results suggested that the preacquisition of TYLCTHV CP cannot sufficiently suppress the transmission of ToLCTWV by B. tabaci.
3.9 Inhibition efficiency of TYLCTHV CP on whitefly transmission of TYLCTHV and ToLCTWV
We further collected the transmission data from previous transmission experiments to determine the inhibition efficiency of TYLCTHV CP on whitefly transmission of TYLCTHV and ToLCTWV. The inhibition rate of using TYLCTHV CP to suppress whitefly transmission of TYLCTHV was 68 %±22 %. The inhibition rate of using TYLCTHV CP to suppress whitefly transmission of ToLCTWV was 22 %±11 % (Fig.
9). Significant difference of the inhibition efficiency between TYLCTHV and ToLCTWV by using TYLCTHV CP was observed (student t test, P=0.002). The result indicated that using TYLCTHV CP to suppress whitefly transmission of TYLCTHV was more effective than whitefly transmission of ToLCTWV.
Chapter 4 Discussion
Whitefly-transmitted begomoviruses threaten a wide range of crops worldwide (Czosnek et al, 2002). The studies on virus translocation have confirmed that the midgut of whitefly is the major organ where the majority of virus reservoirs and absorbs into hemolymph (Ghanim et al., 2001b; Ghanim et al., 2009). The CP of begomoviruses is not only responsible for encapsulating viral genomes, but also involved in vector transmission (Azzam et al., 1994; Noris et al., 1998) and vector specificity (Briddon et al., 1990). In this study, we investigated the role of TYLCTHV CP in the whitefly transmission of begomoviruses.
We discovered that TYLCTHV CP bound to the midgut cells of B. tabaci which provided strong evidence that TYLCTHV CP served as an attachment protein in the transmission of TYLCTHV by B. tabaci. Furthermore, the failure of TYLCTHV MP and BBTV CP binding to the midgut cells of B. tabaci implied that the interaction between CP and a putative receptor in midgut was specific. Our result is consistent with a previous study reporting the CP of TYLCV, a monopartite begomovirus, bound to the midgut cells of B. tabaci (Wang et al., 2014). Höfer et al. (1997) reported that the exchange of CP gene between whitefly transmissible virus and non-transmissible virus altered the whitefly transmissibility. A small change of the amino acids of the begomovirus CP resulted in the failure of transmission by B. tabaci (Noris et al., 1998;
Höhnle et al., 2001). Thus, the structure of CP determined by amino acid sequence may be essential for specific recognition by a putative receptor in B. tabaci.
Since TYLCTHV CP has been proved to be an attachment protein, we further examined the ability of TYLCTHV CP to block the putative receptor of TYLCTHV in the midguts of B. tabaci. The feeding of TYLCTHV CP prior to TYLCTHV acquisition
decreased the accumulation of TYLCTHV in the midguts of B. tabaci. We hypothesized that TYLCTHV CP competed the midgut binding sites of TYLCTHV hence reduced the acquisition of TYLCTHV by B. tabaci. Recently, TYLCV CP was proved to be an attachment protein binding to the midgut cells and decreased TYLCV acquisition in B.
tabaci (Wang et al., 2014). However, our real-time PCR assay showed that
preacquisition of TYLCTHV CP did not reduce TYLCTHV acquisition by B. tabaci.
Our result of real-time PCR on TYLCTHV acquisition by B. tabaci was not consistent with previous study which may result from the TYLCTHV source plants. Different infection level in TYLCTHV source plants for virus acquisition may affect the result of virus quantification. Other than acquiring TYLCTHV from TYLCTHV source plant, feeding whiteflies with purified TYLCTHV may be more accurate. In addition, we must not exclude the possibility that TYLCV replication occurs in the whitefly. A time course experiment should be taken to analyze the replication of TYLCTHV.
Transmission experiments demonstrated that the feeding of TYLCTHV CP prior to TYLCTHV acquisition decreased the transmission rate of TYLCTHV by B. tabaci. Less reduction of TYLCTHV transmission by preacquisition of TYLCTHV MP and BBTV CP was also observed in the transmission assays. The surprising results might due to the non-specific protein-protein interaction between TYLCTHV MP or BBTV CP with the putative receptor of TYLCTHV in B. tabaci. Besides, the different infection level in TYLCTHV source plants for virus acquisition in each trial might influence the outcomes of transmission assays. Many studies have demonstrated that preacquisition of structural viral proteins reduced vector transmission of the viruses. The ability of Nilaparvata lugens transmitting Rice ragged stunt oryzavirus (RRSV) was inhibited after the feeding of the spike protein of RRSV (Zhou et al., 1999). Similar result was also observed in the research on Frankliniella occidentalis transmitting Tomato spotted
wilt virus (TSWV). The transmission rate of TSWV was reduced after feeding soluble
glycoprotein GN prior to virus acquisition (Whitfield et al., 2008).
The disease control of begomovirus would be accelerated if the CP of begomovirus serves as an inhibitory agent to reduce the vector transmission of closely relative viruses. We investigated the potential of TYLCTHV CP to decrease the vector transmission of ToLCTWV. ToLCTWV was chosen since the tomato plants in Taiwan were either infected with TYLCTHV or mix-infected with TYLCTHV and ToLCTWV (Tsai et al., 2011). In vivo binding assays and real-time PCR revealed that preacquisition of TYLCTHV CP can suppress ToLCTWV accumulation in the midgut cells of B. tabaci. However, transmission assays suggested that the preacquisition of TYLCTHV CP could not sufficiently reduce vector transmission of ToLCTWV. The results indicated that the amount of ToLCTWV acquired by whiteflies did not correlate to the transmissibility of ToLCTWV by whiteflies. We inferred that the failure of using TYLCTHV CP to inhibit the vector transmission of ToLCTWV might result from the different receptors that mediate the virus recognition in the midguts of B. tabaci. The initial interaction between virus particles and the cell surface mediates virus entry and retention in their vectors. Therefore, transmissibility of a vector insect is characterized by the level of vector specificity.
The circulative transmission pathway for begomoviruses transmitted by B. tabaci has been well studied. (Ghanim, 2014). The uptake sites for TYLCV were the filter chamber and midgut of B. tabaci (Czosnek et al., 2002; Ghanim and Medina, 2007).
Our results demonstrated that TYLCTHV CP bound to the midgut cells of B. tabaci, and the interaction was stable and specific. A putative receptor on the surfaces of midgut cells may mediate the interaction between the virus and midgut. Various receptors may exist given the circumstances that a selective barrier was found in a
nonvector whitefly to TYLCV (Ohnishi et al., 2009). It might explain why the preacquisition of TYLCTHV CP did not reduce the vector transmission of ToLCTWV.
Although the potential cellular receptors for Barley yellow dwarf virus have been identified in its vector aphid Sitobion avenae (Li et al., 2001), the potential receptors for begomoviruses in B. tabaci still remains unknown. In order to develop an effective and specific control strategy, further investigations are needed to identify the receptors for begomoviruses. Also, the salivary gland is the key organ that determines vector specificity and transmission (Wei et al., 2014). Other than the midgut, it is likely that there are receptors in salivary gland of the whitefly. The research on begomoviruses addressing the interaction with salivary gland is also important.
Previous studies have shown that various endosymbionts were detected inside alimentary ducts of whiteflies (Gottlieb et al., 2008). The GroEL protein produced by the endosymbionts in B. tabaci played the role in the transmission of TYLCV (Morin et al., 1999; Morin et al., 2000; Gottlieb et al., 2010). The mechanism of attachment and circulation of begomoviruses inside whiteflies are key factors that affect whitefly transmission efficiency. It is crucial to provide information regarding whitefly transmission to prevent the spread of tomato yellow leaf curl diseases.
In this study, we have investigated the role of CP in the transmission of begomoviruses by B. tabaci. The function of TYLCTHV CP as an attachment protein was confirmed. The preacquisition of TYLCTHV CP competed the putative receptor in the midguts of B. tabaci hence suppressed the virus acquisition and whitefly transmission of TYLCTHV.
REFERENCES
Azzam O, Frazer J, De La Rosa D, Beaver JS, Ahlquist P, Maxwell DP. 1994. Whitefly transmission and efficient ssDNA accumulation of Bean golden mosaic geminivirus require functional coat protein. Virology 204: 289-296.
Briddon RW, Pinner MS, Stanley J, Markham PG. 1990. Geminivirus coat protein gene replacement alters insect specificity. Virology 177: 85-94.
Brown JK, Czosnek H. 2002. Whitefly transmitted viruses. Adv Bot Res 36: 65-100.
Brown JK, Fauquet CM, Briddon RW, Zerbini M, Moriones E, Navas-Castillo J. 2012.
Geminiviridae. pp 351-373. In: King AMQ, Adams MJ, Carstens EB, Lefkowitz
EJ (eds). Virus taxonomy-Ninth report of the International Committee on Taxonomy of Viruses. Elsevier Academic Press, London.
Caciagli P, Medina Piles VM, Marian D, Vecchiati M, Masenga V, Mason G, Falcioni T, Noris E. 2009. Virion stability is important for the circulative transmission of Tomato yellow leaf curl Sardinia virus by Bemisia tabaci, but virion access to
salivary glands does not guarantee transmissibility. J Virol 83: 5784-5795.
Czosnek H. 2008. Tomato yellow leaf curl virus (Geminiviridae). pp 138-145. In: Mahy BWJ, Van Regenmortel MHV. (eds). Encyclopedia of Virology, vol 5, 3rd ed.
Elsevier, Oxford.
Czosnek H, Ghanim M. 2011. Bemisia tabaci - Tomato yellow leaf curl virus interaction causing worldwide epidemics. pp 51-67. In: Thompson WMO (ed). The whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) interaction with Geminivirus-infected
host plants. Springer, Dordrecht.
Czosnek H, Ghanim M, Ghanim M. 2002. The circulative pathway of begomoviruses in the whitefly vector Bemisia tabaci—insights from studies with Tomato yellow leaf
curl virus. Ann Appl Biol 140: 215-231.
Dinsdale A, Cook L, Riginos C, Buckley YM, De Barro P. 2010. Refined global analysis of Bemisia tabaci (Gennadius) (Hemiptera: Sternorrhyncha: Aleyrodoidea) mitochondrial cytochrome oxidase 1 to identify species level genetic boundaries.
Ann Entomol Soc Am 103: 196-208.
Fondong VN. 2013. Geminivirus protein structure and function. Mol Plant Pathol 14:
635-649.
Gafni Y, Epel BL. 2002. The role of host and viral proteins in intra-and inter-cellular trafficking of geminiviruses. Physiol Mol Plant Pathol 60: 231-241.
Gottlieb Y, Ghanim M, Gueguen G, Kontsedalov S, Vavre F, Fleury F, Zchori-Fein E.
2008. Inherited intracellular ecosystem: symbiotic bacteria share bacteriocytes in whiteflies. FASEB J 22: 2591-2599.
Gottlieb Y, Zchori-Fein E, Mozes-Daube N, Kontsedalov S, Skaljac M, Brumin M, Sobol I, Czosnek H, Vavre F, Fleury F, Ghanim M. 2010. The transmission efficiency of Tomato yellow leaf curl virus by the whitefly Bemisia tabaci is correlated with the presence of a specific symbiotic bacterium species. J Virol 84:
9310-9317.
Ghanim M. 2014. A review of the mechanisms and components that determine the transmission efficiency of Tomato yellow leaf curl virus (Geminiviridae;
Begomovirus) by its whitefly vector. Virus Res 186: 47-54.
Ghanim M, Medina V. 2007. Localization of Tomato yellow leaf curl virus in its whitefly vector Bemisia tabaci. pp 171-183. In: Czosnek H (ed). Tomato yellow leaf curl virus disease: Management, molecular Biology, breeding for Resistance.
Springer, Dordrecht.
Ghanim M, Morin S, Zeidan M, Czosnek H. 1998. Evidence for transovarial
transmission of Tomato yellow leaf curl virus by its vector the whitefly Bemisia tabaci. Virology 240: 295-303.
Ghanim M, Morin S, Czosnek H. 2001a. Rate of Tomato yellow leaf curl virus translocation in the circulative transmission pathway of its vector, the whitefly Bemisia tabaci. Phytopathology 91: 188-196.
Ghanim M, Rosell RC, Campbell LR, Czosnek H, Brown JK, Ullman DE. 2001b.
Microscopic analysis of the digestive, salivary and reproductive organs of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) biotype B. J Morphol 248: 22-40.
Ghanim M, Brumin M, Popovski S. 2009. A simple, rapid and inexpensive method for localization of Tomato yellow leaf curl virus and Potato leafroll virus in plant and insect vectors. J Virol Methods 159: 311-314.
Götz M, Popovski S, Kollenberg M, Gorovitz R, Brown JK, Cicero JM, Czosnek H, Winter S, Ghanim M. 2012. Implication of Bemisia tabaci heat shock protein 70 in begomovirus–whitefly interactions. J Virol 86: 13241-13252.
Höfer P, Bedford ID, Markham PG, Jeske H, Frischmuth T. 1997. Coat protein gene replacement results in whitefly transmission of an insect nontransmissible geminivirus isolate. Virology 236: 288-295.
Hogenhout SA, Ammar ED, Whitfield AE, Redinbaugh MG. 2008. Insect-vector interactions with persistently transmitted viruses. Annu Rev Phytopathol 46:
327-359.
Höhnle M, Höfer P, Bedford ID, Briddon RW, Markham PG, Frischmuth T. 2001.
Exchange of three amino acids in the coat protein results in efficient whitefly transmission of a nontransmissible Abutilon mosaic virus isolate. Virology 290:
164-171.
Hu J, De Barro P, Zhao H, Nardi F, Wang J, Liu SS. 2011. An extensive field survey
combined with a phylogenetic analysis reveals rapid and widespread invasion of two alien whiteflies in China. PLoS One 6: e16061.
Ko CC, Hung YC, Wang CH. 2007. Sequence characterized amplified region markers for identifying biotypes of Bemisia tabaci (Hem., Aleyrodidae). J Appl Entomol 131: 542-547.
Lazarowitz SG, Beachy RN. 1999. Viral movement proteins as probes for intracellular and intercellular trafficking in plants. Plant Cell 11: 535-548.
Li CY, Cox-Foster D, Gray SM, Gildow F. 2001. Vector specificity of Barley yellow dwarf virus (BYDV) transmission: Identification of potential cellular receptors
binding BYDV-MAV in the aphid, Sitobion avenae. Virology 286: 125-133.
Li R, Xie W, Wang S, Wu Q, Yang N, Yang X, Pan H, Zhou X, Bai L, Xu B, Zhou X, Zhang Y. 2013. Reference gene selection for qRT-PCR analysis in the sweet potato whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). PLoS ONE 8: e53006.
Morin S, Ghanim M, Zeidan M, Czosnek H, Verbeek M, van den Heuvel JFJM. 1999. A GroEL homologue from endosymbiotic bacteria of the whitefly Bemisia tabaci is implicated in the circulative transmission of Tomato yellow leaf curl virus.
Virology 256: 75-84.
Morin S, Ghanim M, Sobol I, Czosnek H. 2000. The GroEL protein of the whitefly Bemisia tabaci interacts with the coat protein of transmissible and
nontransmissible begomoviruses in the yeast two-hybrid system. Virology 276:
404-416.
Navas-Castillo J, Fiallo-Olivé E, Sánchez-Campos S. 2011. Emerging virus diseases transmitted by whiteflies. Annu Rev Phytopathol 49: 219-248.
Noris E, Vaira AM, Caciagli P, Masenga V, Gronenborn B, Accotto GP. 1998. Amino acids in the capsid protein of Tomato yellow leaf curl virus that are crucial for
systemic infection, particle formation, and insect transmission. J Virol 72:
10050-10057.
Ohnishi J, Kitamura T, Terami F, Honda K. 2009. A selective barrier in the midgut epithelial cell membrane of the nonvector whitefly Trialeurodes vaporariorum to Tomato yellow leaf curl virus uptake. J Gen Plant Pathol 75: 131-139.
Polston JE, De Barro P, Boykin LM. 2014. Transmission specificities of plant viruses with the newly identified species of the Bemisia tabaci species complex. Pest Manag Sci 70: 1547-1552.
Tsai WS, Shih SL, Kenyon L, Green SK, Jan FJ. 2011. Temporal distribution and pathogenicity of the predominant tomato-infecting begomoviruses in Taiwan. Plant Pathol 60: 787-799.
Wang LL, Wei XM, Ye XD, Xu HX, Zhou XP, Liu SS, Wang XW. 2014. Expression and functional characterisation of a soluble form of Tomato yellow leaf curl virus coat protein. Pest Manag Sci 70: 1624-1631.
Wartig L, Kheyr-Pour A, Noris E, De Kouchkovsky F, Jouanneau F, Gronenborn B, Jupin I. 1997. Genetic analysis of the monopartite Tomato yellow leaf curl geminivirus: roles of V1, V2, and C2 ORFs in viral pathogenesis. Virology 228:
132-140.
Watanabe S, Bressan A. 2013. Tropism, compartmentalization and retention of banana bunchy top virus (Nanoviridae) in the aphid vector Pentalonia nigronervosa. J Gen
Virol 94: 209-219.
Wei J, Zhao JJ, Zhang T, Li FF, Ghanim M, Zhou XP, Ye GY, Liu SS, Wang XW. 2014.
Specific cells in the primary salivary glands of the whitefly Bemisia tabaci control retention and transmission of begomoviruses. J Virol 88: 13460-13468.
Weng SH, Tsai WS, Kenyon L, Tsai CW. 2015. Different transmission efficiencies may
drive displacement of tomato begomoviruses in the fields in Taiwan. Ann Appl Biol 166: 321-330.
Whitfield AE, Kumar NKK, Rotenberg D, Ullman DE, Wyman EA, Zietlow C, Willis DK, German TL. 2008. A soluble form of the Tomato spotted wilt virus (TSWV) glycoprotein GN (GN-S) inhibits transmission of TSWV by Frankliniella occidentalis. Phytopathology 98: 45-50.
Yaakov N, Levy Y, Belausov E, Gaba V, Lapidot M, Gafni Y. 2011. Effect of a single amino acid substitution in the NLS domain of Tomato yellow leaf curl virus-Israel (TYLCV-IL) capsid protein (CP) on its activity and on the virus life cycle. Virus Res 158: 8-11.
Zhou G, Lu X, Lu H, Lei J, Chen S, Gong Z. 1999. Rice ragged stunt oryzavirus: role of the viral spike protein in transmission by the insect vector. Ann Appl Biol 135:
573-578.
Figure 1. Analysis of recombinat viral proteins by SDS-PAGE.
Proteins were purified by nickel affinity chromatography, seperated by electrophoresis and stained with Bio-Safe Coomassie. M: Pre-stained sharp protein marker III (Yestern).
Lane 1: recombinant TYLCTHV CP; Lane 2: re-purified recombinant TYLCTHV CP;
Lane 3: recombinant BBTV CP; Lane 4: re-purified recombinant BBTV CP; Lane 5:
recombinant TYLCTHV MP.
Figure 2. In vivo binding assay.
Bemisia tabaci were fed recombinant (A) TYLCTHV CP, (B) TYLCTHV MP, (C)
BBTV CP, or (D) artificial feeding medium only for 24 h. After feeding, the midguts were dissected and immunolabeled with a mouse anti-S tag antiserum and a goat anti-mouse IgG antiserum conjugated with Alexa fluor 633 (red). All the samples were mounted in the SlowFade Gold antifade reagent with DAPI (blue) and examined by confocal microscopy. AM: ascending midgut; DM: descending midgut; FC: filter chambe; GC: gastric caeca. Scale bar: 100 µm.
Figure 3. Recombinant TYLCTHV CP competed with TYLCTHV for the midgut binding site in Bemisia tabaci.
Adult whiteflies were fed recombinant (A) TYLCTHV CP, (B) TYLCTHV MP, (C) BBTV CP, or (D) artificial feeding medium only for 24 h, and they were subsequently fed TYLCTHV-infected tomato plants for another 24 h to acquire the virus. The midguts were dissected and immunolabeled. The recombinant proteins were labelled with a mouse anti-S tag antiserum and a goat anti-mouse IgG antiserum conjugated with Alexa fluor 633 (red). TYLCTHV was labelled with a rabbit anti-TYLCTHV antiserum and a goat anti-rabbit IgG antiserum conjugated with Alexa fluor 555 (green). All the samples were mounted in the SlowFade Gold antifade reagent with DAPI (blue) and examined by confocal microscopy. Scale bars in (A)~(D) were 100 µm; (E)~(H) were 10 µm.
Figure 4. Relative abundance of TYLCTHV in Bemisia tabaci after feeding recombinant viral proteins and the virus sequentially.
Adult whiteflies were fed recombinant TYLCTHV CP, TYLCTHV MP, BBTV CP, or artificial feeding medium only through membrane feeding for 24 h, and they were subsequently fed on TYLCTHV-infected tomato plants for another 24 h to acquire the virus. After which, they were fed a sucrose solution for 6 h to clear the gut lumen.
TYLCTHV DNA was quantified by real-time PCR. Relative abundance of TYLCTHV DNA was normalized to endogenous Heat shock protein 90 gene of B. tabaci. Same letters indicate no statistical differences between means (α=0.05), analyzed using one-way ANOVA. Vertical bars indicate standard deviation. (n=4)
Figure 5. Transmission rate of TYLCTHV after Bemisia tabaci feeding recombinant viral proteins and the virus sequentially.
Adult whiteflies were fed recombinant TYLCTHV CP, TYLCTHV MP, BBTV CP, or artificial feeding medium only for 24 h, and they were subsequently fed on TYLCTHV-infected tomato plants for another 24 h to acquire the virus. After which, five whiteflies were given 24 h to feed on each healthy test plant to inoculate the virus.
Ten test plants were inoculated for each trial. PCR were used to verify the virus infection of test plants two weeks after inoculation. The experiments repeated five times (Ten test plants each time). Different letters indicate statistical differences between means (α=0.05), analyzed using χ2 test. Vertical bars indicate standard deviation. (n=5)
Figure 6. Recombinant TYLCTHV CP competed with ToLCTWV for the midgut binding site in Bemisia tabaci.
Adult whiteflies were fed artificial feeding medium with (A) or without (B) recombinant TYLCTHV CP for 24 h, and they were subsequently fed on ToLCTWV-infected tomato plants for another 24 h to acquire the virus. The midguts were dissected and immunolabeled. Recombinant TYLCTHV CP was labelled with a mouse anti-S tag antiserum and a goat anti-mouse IgG antiserum conjugated with Alexa fluor 633 (red). ToLCTWV was labelled with a rabbit anti-ToLCTWV antiserum and a goat anti-rabbit IgG antiserum conjugated with Alexa fluor 555 (green). All the samples were mounted in the SlowFade Gold antifade reagent with DAPI (blue) and examined by confocal microscopy. Scale bar: 10 µm.
Figure 7. Relative abundance of ToLCTWV in Bemisia tabaci after feeding recombinant TYLCTHV CP and the virus sequentially.
Adult whiteflies were fed artificial feeding medium with or without recombinant TYLCTHV CP for 24 h, and they were subsequently fed on ToLCTWV-infected tomato plants for another 24 h to acquire the virus. After which, they were fed a sucrose solution for 6 h to clear the gut lumen. ToLCTWV DNA was quantified by real-time PCR. Relative abundance of ToLCTWV DNA was normalized to endogenous Heat shock protein 90 gene of B. tabaci. Same letters indicate no statistical differences between means (α=0.05), analyzed using Mann-Whitney test of nonparametric method.
Vertical bars indicate standard deviation. (n=4)
Figure 8. Transmission rate of ToLCTWV after Bemisia tabaci feeding recombinant TYLCTHV CP and the virus sequentially.
Adult whiteflies were fed artificial feeding medium with or without recombinant TYLCTHV CP for 24 h, and they were subsequently fed on ToLCTWV-infected tomato plants for another 24 h to acquire the virus. After which, five whiteflies were given 24 h to feed on each healthy test plant to inoculate the virus. Ten test plants were inoculated for each trial. PCR were used to verify the virus infection of test plants two weeks after inoculation. The experiments repeated five times (Ten test plants each time). Different letters indicate statistical differences between means at (α=0.05), analyzed using χ2 test.
Vertical bars indicate standard deviation.
Figure 9. Inhibition efficiency of TYLCTHV CP on whitefly transmission of TYLCTHV and ToLCTWV.
Transmission data of using TYLCTHV CP to suppress whitefly transmission of TYLCTHV and ToLCTWV from Fig 5 and Fig 8 were collected and analyzed by student t test. Different letters indicate statistical differences between means at (α=0.05).
Vertical bars indicate standard deviation.