國立台灣大學生命科學院微生物與生化學研究所 碩士論文初稿
Institute of Microbiology and Biochemistry College of Life Science
National Taiwan University Master Thesis First Draft
臺灣本土禽流感病毒株血液凝集素重鏈異構物分析及其 醣質表現圖譜
Analysis on multiple isoforms and glycan profile of hemagglutinin 1 from avian influenza viruses isolated in Taiwan.
羅庭芳
Ting-Fang Lo
指導教授:莊榮輝 博士
王金和 博士 Advisor: Rong-Huay Juang, Ph.D.
Ching-Ho Wang, Ph.D
中華民國九十八年六月
June, 2009
謝誌
感謝我的父親羅銀煌、母親蔡雪玉的養育及栽培,從小給予我良好的教育環境,
讓我有機會繼續進修學習,走我想走的路。感謝我的姐姐庭萱和弟弟庭甫,陪我分 享生活的點點滴滴,一起成長。
感謝我的指導教授莊榮輝老師,提供我良好的學習環境、自由的研究空間,讓 我在短短的兩年內,學到了很多很多。也很感謝莊老師在科學精神方面的教導,對 科學要有熱忱,要為了“全世界我第一個知道"的實驗結果而興奮並努力; 「實驗 技術再怎麼發達,科學研究最重要的還是人腦!」,老師的這句話,我會謹記在心,
時時刻刻思考並修正自己的研究方向。
感謝我的共同指導教授王金和老師,教導我禽流感研究所需的知識,一起討論 研究方向及如何撰寫研究論文,並提供我充足的病毒樣本,讓研究得以順利進行。
感謝中研院生化所陳水田老師,從碩二開始,幫助我進行醣質研究。陳老師每 周一次的書報討論及進度報告,讓我學習到很多專業知識,也見識到了老師嚴謹及 腳踏實地的研究態度,而短短一年所受到的訓練,也讓我受益良多。
感謝中國醫藥研究所盧美光老師,幫助我進行全醣分析,熱心教導我如何水解 醣類並操作 HPAEC-PAD。也感謝中研院基因體中心楊文彬老師,提供 HPAEC-PAD 相關之儀器設備。感謝楊健志老師教導我使用 Discovery Studio。
感謝莊老師實驗室的夥伴們: 信傑學長、杰龍學長、皮蛋學姊、玉米學姊、和成 學長、福氣蓋學長、雨亭學姊、之儀、毓庭、惠敏、瑩貞、怡君、迺茵、阿金,給 予我研究及生活上之幫助。感謝王老師實驗室的怡彤,辛苦的進行病毒繁殖,提供 我充足的樣本。感謝陳老師實驗室的夥伴們,尤其是帶著我進行醣質圖譜分析的顯 裕學長,總是忙裡抽空,耐心教導我相關的知識和技能。感謝楊文彬老師實驗室的 瑋婷學姊,幫助我操作使用 HPAEC-PAD。
短短兩年的碩士生涯,感受到了臺大這所學校自由開放的學風,也因此接觸到 廣泛的人群,受到了無數人們的幫助,如今完成碩士論文,我會滿懷感激,謹記各 位師長的教誨,邁向人生的下一個歷程。
目錄
中文摘要………iv
英文摘要………v
第一章 緒論………1
1.1 簡介禽流感病毒………1
1.1.1 歷史背景………1
1.1.2 流感病毒之構造與型態………2
1.1.3 流感病毒之基因體………4
1.1.4 流感病毒蛋白質功能簡介………5
1.1.5 流感病毒生活史………6
1.2 血液凝集素 (hemagglutinin, HA)………9
1.2.1 HA 之結構與功能………9
1.2.2 HA 之受體結合區 (receptor binding sites)………9
1.2.3 HA 與致病力之關係………9
1.2.4 HA 之醣質與致病力之關係………10
1.3 研究動機與目的………13
第二章 材料與方法………14
2.1 禽流感病毒樣本處理方法………14
2.1.1 病毒增殖………14
2.1.2 濃縮與純化………15
2.1.3 病毒蛋白質定量………16
2.2 一般電泳檢定法………16
2.2.1 SDS 膠體電泳………16
2.2.2 膠體染色法 - CBR 染色………18
2.2.3 膠體染色法 - 銀染………19
2.2.4 蛋白質電泳轉印法………20
2.2.5 轉印膜 Ponceau S 染色法………20
2.3 免疫染色法………21
2.4 凝集素染色法 (lectin blot)………22
2.5 醣晶片………23
2.6 二次元膠體電泳………24
2.6.1 脫鹽及溶解………24
2.6.3 第二維 15% SDS-PAGE 膠體電泳………25
2.7 酶切實驗………26
2.7.1 鹼性磷酸酶之去磷酸化………26
2.7.2 神經胺酸水解酶水解唾液酸………26
2.8 使用高效能陰離子交換層析儀 (HPAEC) 配合脈衝式安培法偵測器 (PAD) 分析唾液酸………27
2.8.1 弱酸水解釋放唾液酸………27
2.8.2 使用 HPAEC-PAD 分析唾液酸亞型………27
2.9 蛋白質身份鑑定………28
2.9.1 膠體內蛋白質水解………28
2.9.2 以質譜儀鑑定蛋白質點身分………30
2.10 高效能層析儀 (HPLC) 配合螢光偵測器分析 N-醣質………30
2.10.1 醣質之膠內蛋白質水解釋放及螢光標定………30
2.10.2 使用 HPLC 分析 N-glycans………...………31
2.11 質譜儀醣質定序………31
2.11.1 從蛋白質上釋放並純化 N-glycans………31
2.11.2 N-glycans 之泛甲基化………32
2.11.3 以質譜儀分析 N-glycans………33
第三章 結果………34
3.1 比較非毒性與毒性禽流感病毒株之間蛋白質體的差異………36
3.1.1 兩株 2838 禽流感病毒二次元圖譜之比較………36
3.1.2 血液凝集素重鏈 (HA1) 為兩禽流感病毒株蛋白質圖譜上差異最 大之蛋白質………36
3.1.3 非毒性與毒性 AIV 之 HA1 皆有 6 種不同異構物………...…37
3.2 探究紅血球凝集素 1 (HA1) 有 6 種不同等電點異構物之原因………41
3.2.1 預測血液凝集素 1 (HA1) 之蛋白質轉譯後修飾………41
3.2.2 實驗設計………41
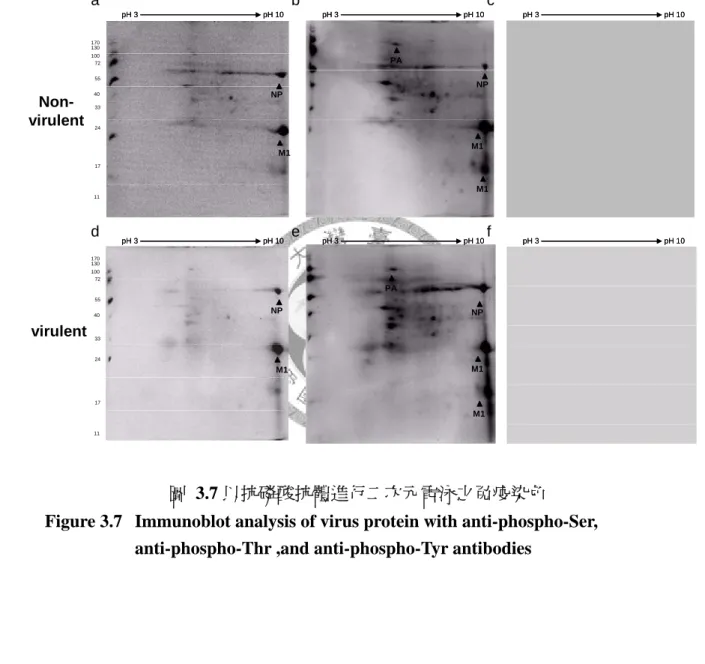
3.3 不同等電點異構物之紅血球凝集素 1 (HA1) 並非蛋白質磷酸化所造 成………44
3.3.1 以鹼性磷酸酶處理禽流感病毒株並進行二次元電泳………44
3.3.2 以抗磷酸抗體進行二次元電泳之免疫染色………44
3.4 不同等電點異構物之紅血球凝集素 1 (HA1) 可能是由其醣質之唾液酸修飾 所造成………47
3.4.1 以神經胺酸水解酶處理禽流感病毒株並進行二次元電泳………47
3.4.2 以凝集素染色法 (lectin blot) 發現 HA1 之醣質可能有 α-2,3 鍵 結之唾液酸………47
3.4.3 以高效能高效能陰離子交換層析儀 (HPAEC) 配合脈衝氏安培法 偵測器 (PAD) 分析 HA1 醣基之唾液酸………48
3.4.4 以凝集素染色法分析經唾液酸酶水解後之 HA1………48
3.5 紅血球凝集素 (HA1) 之醣質表現圖譜………54
3.5.1 使用醣晶片分析 HA1 之醣質………54
3.5.2 使用 HPLC 配合螢光偵測器分析 HA1 之醣質………54
3.5.3 以 MALDI-TOF/TOF MS 及 MS/MS 研究 HA1 之醣質表現圖 譜………54
第四章 討 論 ………66
4.1 探討非毒性及毒性病毒株之間 HA1 等電點之差異,及其各自形成 6 種等 電點異構物之生理意義………66
4.2 HA1 之醣基化位置與其功能探討….………..……66
4.3 除了醣基化外,蛋白質骨架本身及其它蛋白質轉譯後修飾亦可能造成 HA1 異構物………67
4.4 m/z 1867 之 X-glycan 尚未被報導過……….…………..……67
4.5 X-glycan 與 HA1 等電點異構物可能之兩種關係………….…………..……67
第五章 結 論 ………70
第六章 參考文獻………71
中文摘要
許多低病原性禽類流感病毒 (avian influenza viruses, AIV) 已於臺灣本土農場分 離出來,然而這些病毒蛋白質體之改變與毒性之關係尚未被研究清楚。在本研究中,
使用二次元電泳 (2-DE) 並配合專一性之凝集素 (specific lectins) 分析比較,來自同 一 H6N1 族群,但卻表現不同毒性程度之兩株 AIV:非毒性病毒株 (2838N) 及毒 性 病 毒 株 (2838V) 。 比 較 兩 病 毒 株 之 二 次 元 蛋 白 質 圖 譜 發 現 , 血 液 凝 集 素 1 (hemagglutinin, HA) 為差異最大之蛋白質,且兩株 AIV 之 HA1 皆有分子量相近,
但等電點迥異之 6 種異構物。本研究發現,HA1 這些異構物形成之原因,並非因 於蛋白質磷酸化,亦非 HA1 醣質之唾液酸修飾 (sialylation)。另外,我們使用介質 輔助雷射脫附離子化質譜儀 (MALDI MS and MS/MS) 建構 HA1 各異構物之醣質 圖譜。比較 HA1 各異構物之醣質表現圖譜發現,等電點越低之 HA1 異構物其 m/z 1867 之 X-醣質比例越高。而血液凝集素主要功能為辨識並和宿主之唾液酸受體結 合,因此 HA1 本身特別之醣基化可能會影響 AIV 之宿主辨識,亦可能和病毒之組 織趨性及免疫逃脫有關。
英文摘要
Several low pathogenic avian influenza viruses (AIV) had been isolated in local farms. However, the relationship between the protein change and the virulence of these AIV was not clear. In this study, two variants from a single H6N1 population with
different virulence levels, the non-virulent strain (2838N) and the virulent strain (2838V), were analyzed by two-dimensional electrophoresis (2-DE) and then identified with specific lectins. Comparisons of the 2-DE patterns of these two strains showed major difference in hemagglutinin 1 (HA1) which contained six isoforms with similar molecular mass but showing variant isoelectric points (pI). We found that the difference in pI values was not due to either of phosphorylation and sialylation on HA1. In addition, we
performed MALDI mass spectrometry (MS and MS/MS)-based glycomic profile analyses for the N-glycans of the HA1 isoforms. The results showed that the isoform with lower pI contained more m/z 1867 X-glycan. One of the major functions for hemagglutinin is the recognition and binding of sialic acid-containing receptors on the target cell. It is possible that the special glycosylation of HA1 might have effects on host recognition, as well as tissue tropism or immune escape.
第一章 緒論
1.1 簡介禽流感病毒
1.1.1 歷史背景
家禽流行性感冒 (avian influenza) 之歷史,要追溯回 1878 年之北義大利,由 Perroncito 首次報導雞群爆發之嚴重疾病,當時稱為雞瘟 (fowl plague)。最初雞瘟 和家禽霍亂 (fowl cholera) 產生混淆,但很快的於 1880 年 Rivolta 和 Delprato 發 現雞瘟之臨床和病理,都和家禽霍亂有很大之差異,因此重新命名為 Typhus exudatious gallinarum。雖然於 1901 年 Centanni 和 Savonuzzi 已認為此雞瘟是由 濾過性病毒所致,但直到 1955 年才由 Schäfer 研究得知,是由 A 型流行性感冒病 毒 (influenza A viruses) 所致。而 1878 年雞瘟 (fowl plague) 此名稱,也於第一屆 禽流感國際研討會中 (International Symposium on Avian Influenza),正式更名為高病 原性家禽流行性感冒 (highly pathogenic avian influenza, HPAI) 1。
家禽流行性感冒,是由禽流感病毒 (avian influenza virus, AIV) 所引起的動物傳 染病,通常只感染鳥禽類,少見情況會感染豬。禽流感病毒高度針對特定之物種,
但 在 罕 有 之 情 況 下 會 跨 越 種 間 障 礙 感 染 人 類 。 禽 流 感 病 毒 屬 正 黏 液 病 毒 科 (Orthomyxoviridae) 之 A 型流行性感冒病毒,依據血液凝集素 (hemagglutinin, HA) 及神經氨酸酶 (neuraminidase, NA) 二個表面蛋白可區分 16 種 HA 亞型及 9 種 NA 亞型。而 A 型流行性感冒病毒,對於野生鳥類寄主,一般不會出現任何反應,並 能與其寄主達至平衡共生 2,3。而低病原性禽流感病毒 (low pathogenic avian influenza, LPAIV) 從野生雀鳥傳至到家禽雀鳥,可能會出現一些輕微受感染徵兆,
而此現象也輔助了病毒之變異。而病毒遺傳因子變異,除了適應新種寄主外,也會 加速變異為高病原性禽流感病毒。
全球流行之人類流行性感冒,最初為 1918 年爆發之西班牙流行性感冒 (Spanish flu),接著是 1957 年亞洲流行性感冒 (Asian flu) 及 1968 年香港流行性 感冒 (Hong Kong flu) 4。研究顯示 1918 年人類流行性感冒之基因,可能來自於禽 類流感病毒 5,1957 年之流感,懷疑為野鴨病毒之 PB1、HA 和 NA 三段基因與 人類 H2N2 病毒產生基因重組所致;1968 年之香港流感亦發現為野鴨病毒之 PB1 和 HA 與人類 H2N2 病毒產生基因重組所致 6。人類之全球性流感與禽流感之間 緊緊相扣,而禽流感成為全球關注之焦點,是因為 1997 年於香港,出現可感染人
而此 H5N1 病毒跨越剛間傳染屏障,可直接由禽鳥類傳至哺乳類 (如貓、豬及人類)
7,8,因此在病毒持續快速的演化變異之當下,禽流感病毒之基礎科學、公共衛生、
疫苗開發及治療方法成為十分迫切之研究。
1.1.2 流感病毒之構造與型態
禽流感病毒在電子顯微鏡下呈現球狀或絲狀,球狀之直徑在 80 至 120 nm 之 間,絲狀流感病毒的長度可達 400 nm。流感病毒結構自外而內可分為套膜、基質 蛋白以及核心三部分,圖 1.1 9。
套膜 (envelope)
套膜為包裹在病毒顆粒之基質蛋白外之一層磷脂雙分子層膜 (phospholipid bilayer),這層膜來自於宿主的細胞膜,成熟之流感病毒由宿主細胞出芽,將宿主之 細胞膜包裹於病毒本身脫離細胞,接著即可去感染下一個目標。另外,套膜有兩種 嵌入之醣蛋白:血液凝集素和神經胺酸酶,這兩類蛋白質突出病毒體外,長度約為 10 nm 至 40 nm,被稱作棘突蛋白質 (spike protein)。通常一個流感病毒顆粒會有 500 個血液凝集素和 100 個神經胺酸酶棘突,而穿膜之離子通道蛋白質 M2 約有 16 ~ 67 個 10,11。
基質蛋白 (matrix protein)
基質蛋白分為兩種,第一種為構成病毒外殼骨架之基質蛋白 1 (matrix protein 1, M1),M1 蛋白質與細胞之套膜內壁緊密結合,有保護病毒核心及維繫病毒空間結 構之功能;第二種基質蛋白為前段提到之基質蛋白 2 (matrix protein 2, M2),M2 為 穿膜之質子通道蛋白質 (proton channel)。
核心 (core)
病毒之核心包含了存貯病毒遺傳訊息之 8 段單股負鏈 RNA,此 8 段 RNA 會和核殼蛋白 (nucleocapsid protein, NP) 密集纏繞形成 8 個病毒核醣核蛋白 (vRNP)。另外,病毒核心亦包括轉錄這些遺傳訊息所必須之 RNA 聚合酶 (RNA polymerase)。
圖 1.1 A 型流行性感冒病毒圖解 9
Figure 1.1 Structural diagram of the influenza A virus 9
1.1.3 流感病毒之基因體
A 型流感病毒的基因體由 8 段單股負鏈 RNA 所組成,第 1、2、3 個節段編 碼三種 RNA 聚合酶:PB2、PB1 和 PA;第 4 個節段編碼血液凝集素;第 5 節 段編碼核殼蛋白;第 6 個節段編碼神經胺酸酶;第 7 節段編碼基質蛋白 (M1, M2);第 8 個節段編碼非結構性蛋白質 (NS1, NS2) 12。此 8 段 RNA 會和核殼蛋 白密集纏繞,並和 PB2、PB1 和 PA 組成之聚合酶異構三聚體 (heterotrimeric polymerase complex) 結合形成 8 個病毒核醣核蛋白,如圖 1.2 13。
病毒之 8 段基因共轉錄出 10 種蛋白質,其中有 9 種蛋白質包含於病毒顆粒 中,而和 mRNA 剪切 (splicing) 相關之 NS1 蛋白質則不存在於病毒顆粒內。此 外,8 段基因中,有 6 段分別轉譯出一種蛋白質;而第 7,8 兩段基因,因為分 別有兩個開放讀架 (open reading frames),因此各轉錄出兩種蛋白質 M1、M2 和 NS1、NS2 14。近年來新發現第 11 種蛋白質 PB1-F2,此為 PB1 基因之開放讀架 有選擇性讀架 (alternative reading frame) 所致 15。
圖 1.2 流行性感冒病毒核醣核蛋白 (RNP) 組織之圖示模型13 Figure 1.2 Schematic model of RNP organization 13
1.1.4 流感病毒蛋白質功能簡介
(1) RNA 聚合酶 (RNA polymerase)
於 8 段遺傳序列中,最長之三段 RNA 負責編碼三種不同之 RNA 聚合酶,
PB2、PB1 和 PA,而此三種聚合酶會組成異構三聚體,如圖 1.2 13,其功能為在 宿主細胞內轉錄並複製病毒 RNA。此三種聚合酶皆為依賴於 RNA 之 RNA 聚合 酶(RNA-dependent RNA polymerase),其中 PB2 辨識並竊取宿主 mRNA 之 5’ 端 帽 (cap),作為引子 (primer);PB1 負責延長由 PB2 啟始轉錄之 RNA,且 PB1 有 內切酶之功能。而目前已已知 PA 擁有蛋白酶 (protease) 之活性 16-18。另外,近年 來發現之 PB1-F2 則和誘發細胞凋亡 (pro-apoptotic) 相關 15。
(2) 核殼蛋白 (nucleocapsid protein, NP)
核殼蛋白是一種與 RNA 結合之蛋白質 (RNA binding protein),其與 RNA 密 集纏繞形成 8 個病毒核醣核蛋白,並調節 nuclear import 18-20。
(3) 血液凝集素 (hemagglutinin, HA)
血液凝集素為三聚體醣蛋白,能與人、鳥、豬,及豚鼠等動物之紅血球表面的 受體結合,引起凝血,因此被稱為血液凝集素。原態之血液凝集素 (HA0) 經蛋白 質水解切割後,會分為重鏈 (HA1) 和輕鏈 (HA2) 兩部分,HA1 可以與宿主細胞 膜上之唾液酸受體結合,後者則可以協助病毒套膜與宿主細胞膜相互融合。血液凝 集素在病毒進入宿主細胞的過程中扮演了重要角色 21。另外,血液凝集素具有免疫 原性,為流感病毒最主要之抗原。
(4) 神經胺酸酶 (neuraminidase, NA)
神經氨酸酶為四聚體醣蛋白,具有水解唾液酸 (sialic acid) 之活性,當成熟之 流感病毒,以出芽的方式脫離宿主細胞後,病毒表面的血液凝集素 (HA) 依舊會經 由唾液酸與宿主之細胞膜聯繫,因此需要由神經氨酸酶,將唾液酸水解,切斷病毒 與宿主細胞之最後聯繫 22。
(5) 基質蛋白 (matrix protein)
基質蛋白分為兩種,第一種為構成病毒外殼骨架之 M1,其與細胞之套膜內壁 緊密結合,有保護病毒核心及維繫病毒空間結構之功能。此外, M1 亦會和 vRNP 產生交互作用,調控 RNA nuclear export,並會參與病毒之出芽 20。第二種基質蛋 白為 M2,其為穿膜之質子通道蛋白質,和病毒脫去外殼 (uncoating) 及組裝 (assembly) 有關 23。
(6) 非結構性蛋白質 (nonstructural protein, NS)
病毒顆粒中,其主要之功能有調控宿主基因表現,及抑制宿主之干擾素對抗病毒
24。第二種非結構性蛋白質為 NS2 亦稱為 NEP (nuclear export protein),顧名思義,
其主要功能與 RNA 之 nuclear export 相關 20,25。
1.1.5 流感病毒生活史
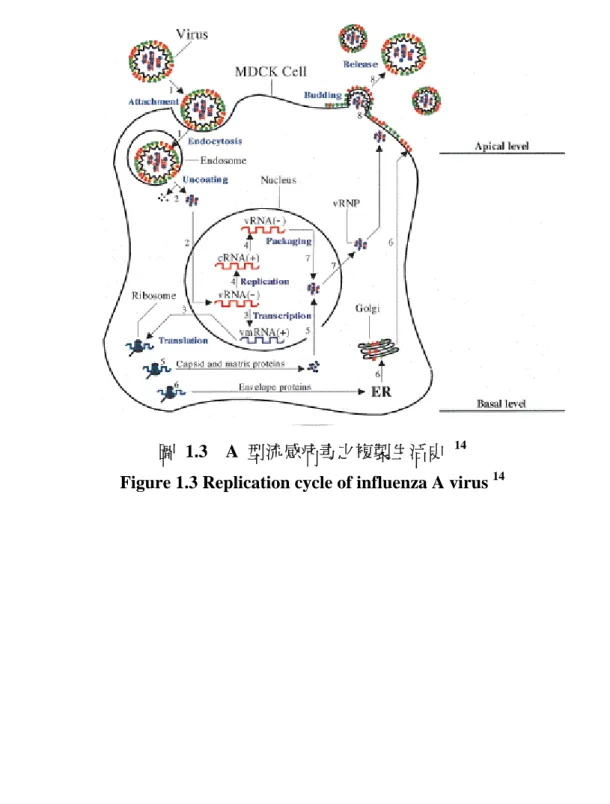
流感病毒之生活史如圖 1.3 所示 14。
(1) 病毒吸附宿主細胞 (virus attachment)
流感病毒之血液凝集素,會辨識宿主細胞上之唾液酸並與之結合吸附宿主細 胞,一般來說,禽流感病毒之 HA 會與唾液酸 α-2,3-半乳糖鍵結之唾液酸結合;
人類流感病毒之 HA 會與唾液酸 α-2,6-半乳糖鍵結之唾液酸結合 26,27。
(2) 病毒進入宿主細胞 (virus entry)
病毒吸附後,主要以依賴 calathrin 型受體媒介內吞作用 (clathrin-dependent receptor-mediated endocytosis) 進入宿主細胞。然而有少許流感病毒利用別種方式進 入細胞 28,29。
(3) 合成病毒 RNA (synthesis of viral RNA) (3a) vRNPs 脫殼並進入宿主細胞核中
當病毒顆粒被吞噬進入吞噬體 (endosome) 時,病毒顆粒上少數之 M2 質子通 道蛋白質,可以使吞噬體內之酸性物質進入病毒顆粒,擾亂 M1 及 vRNP 之交互 作用,並以酸催化的方式使 HA 進行構形重組,幫助 vRNPs 脫殼 (uncoating)。
接著,吞噬體和病毒之膜融合,使 8 組 vRNPs 進入細胞質中。而纏繞住 vRNA 之 NP 帶有細胞核定位訊號 nuclear localization signal (NLS),因此可將 vRNPs 運送 至宿主細胞核內 30。
(3b) 製造病毒之 mRNA (vmRNA Production)
宿主細胞內,主要有三型病毒 RNA,第一為正向之病毒 mRNA (vmRNA);第 二為負向之 genomic RNA (vRNA),第三種為正向之互補 RNA (complementary RNA,cRNA)。vmRNA 有由宿主竊取來之 5'端帽及 3'端多腺嘌呤尾 31-34。而 聚合酶異構三聚體製造完成之 vmRNA 會經由核孔運送出細胞核,進入細胞質中
30。
(3c) 複製病毒基因體 (viral genome replication)
病毒基因體由聚合酶異構三聚體複製,其包含合成全長之 vRNA 及 cRNA。
以 cRNA 作 為 轉 譯 vRNA 之 模 板 , 新 合 成 之 vRNA 則 可 再 轉 譯 出 更 多 之 vmRNA 以及 cRNA。
(4) 合成病毒蛋白質 (synthesis of viral proteins)
(4a) 核殼蛋白 (NP)、非結構性蛋白 (NS)、基質蛋白 (M),與 RNA 聚合酶之合 成
當病毒感染宿主後,宿主細胞之蛋白質合成速度會下降 35,而由細胞核中運送 出之 vmRNA 得以藉由宿主之核醣體 (ribosome) 以最快之速率進行病毒蛋白之合 成。新合成之 NP,M,及 NS 蛋白質被運送回細胞核中後,會參與 M 及 NS 之 mRNA 剪切 (splicing)、轉錄及基因體之複製。
(4b) 合成套膜蛋白質 (envelope protein production)
經核醣體轉譯出之 M2、HA 及 NA 三種套膜蛋白質,會進入內質網 (ER) 進 行醣基化修飾,並進入高基氏體 (Golgi apparatus),接著會移動至宿主細胞膜之病 毒出芽頂端 (apical)。
(5) 包裹 RNA (packaging of RNA) 並運送出宿主細胞核
首先,PB2、PB1、NP 及 NS2 蛋白質結合上 vRNA 並形成 vRNP,接著 M1 結合上 vRNP,形成 M1-vRNP 複合體,並催化 vRNP 出細胞核,進入細胞質內,
而此出核之反應是由 M1、NS2 及 NP 共同完成 36,而和 M1 聯繫在一起之 vRNP,無法再次進入細胞核中。
(6) 病毒出芽與釋出 (virus budding and release)
M2、HA 及 NA 三種套膜蛋白質,移動至宿主細胞膜後,其細胞體漿尾 (cytoplasmic tail) 會和 M1-vRNP 複合體產生交互作用,並於組裝位置 (assembly site) 出芽 (budding)。當病毒出芽完成後,HA 依舊會和宿主細胞膜上之唾液酸受 體結合,而 NA 則會酶切此唾液酸,由宿主細胞釋出病毒顆粒 22。
圖 1.3 A 型流感病毒之複製生活史 14
Figure 1.3 Replication cycle of influenza A virus 14
1.2 血液凝集素 (hemagglutinin, HA)
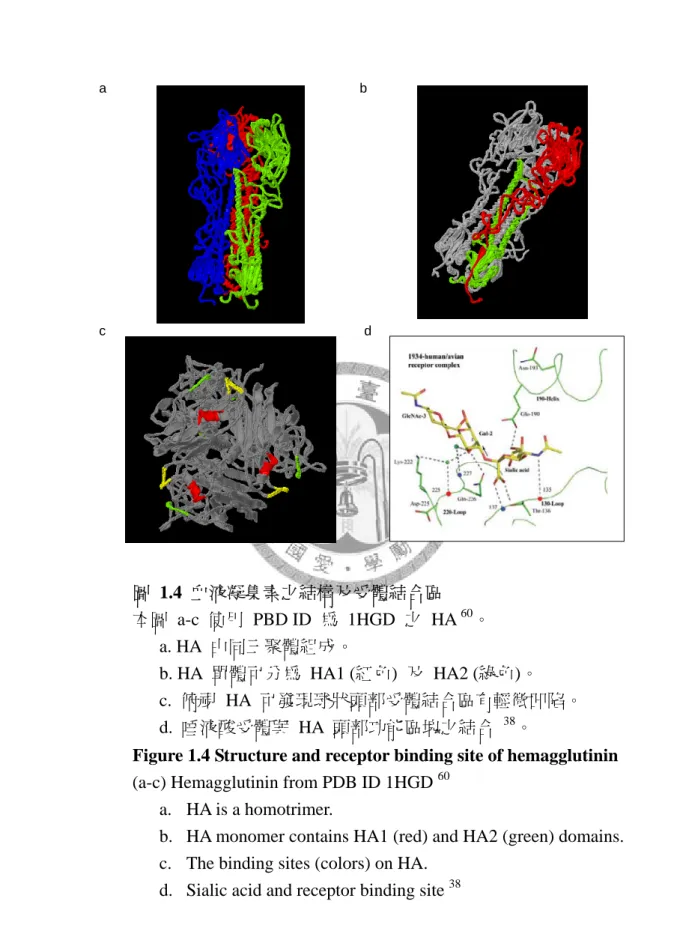
1.2.1 HA 之結構與功能
血液凝集素之結構為同質三聚體 (homotrimer),各單體主要分為兩個功能區 塊:球狀功能區塊 (globular domain) 與莖狀功能區塊 (stem domain),如圖 1.4 a 所 示。原態之血液凝集素 (HA0) 之剪切位 (cleavage site) 經蛋白質水解切割後,會 分為重鏈 (HA1) 和輕鏈 (HA2) 兩部分,其中 HA1 包含頂端球狀頭部及部分莖狀 主幹;HA2 則形成莖狀主幹,如圖 1.4 b 所示,而 HA1 之功能主要為可以與宿 主細胞膜上之唾液酸受體結合 37,吸附宿主細胞;HA2 則可以協助病毒套膜與宿 主細胞膜相互融合。
1.2.2 HA 之受體結合區 (receptor binding sites)
HA 之受體結合區域,位於頂端之球狀功能區塊,而其球狀頭部有輕微凹陷 (圖 1.4 c),使宿主表面醣蛋白 (glycoprotein) 及醣脂質 (glycolipid) 之唾液酸受體,可 進入 HA 之頭部凹陷,形成鎖與鑰匙 (lock and key) 之結合 37,38。而此頭部凹陷是 由構成三角形之 loop-helix-loop 之蛋白質二級結構,與唾液酸受體結合 (圖 1.4 d)
38。各種流行性感冒病毒其 HA 之受體結合區域之不同,會導致病毒感染之特異 性,一般來說,禽流感病毒之 HA1 會與唾液酸 α-2,3-半乳糖鍵結之唾液酸結合;
人類流感病毒之 HA1 則與唾液酸 α-2,6-半乳糖鍵結之唾液酸結合 37。 1.2.3 HA 與致病力之關係
(1) HA 造成宿主專一性
血液凝集素與宿主之唾液酸結合,為流感病毒感染宿主的第一步,而 1.2.2 節 提及之 HA 受體結合區之不同,直接造成了 HA 感染宿主之專一性,也造就了種 間感染屏障 (species barrier)。研究發現,感染禽類之 HA1 會與唾液酸 α-2,3-半乳 糖鍵結之唾液酸結合;感染人類之 HA1 會與唾液酸 α-2,6-半乳糖鍵結之唾液酸結 合 39-41。
流感病毒是經由呼吸道感染人類及豬,但卻以腸道感染禽類。經研究發現位於 人類之深層呼吸道,如肺部,擁有豐富 HA 禽類型受體 (α-2,3-半乳糖鍵結之唾液 酸),而禽類腸道亦擁有 HA 人類型受體 (α-2,6-半乳糖鍵結之唾液酸),而豬之呼 吸道則同時擁有 HA 人類型受體其禽類型受體 42,這些發現意味著新興之流感病 毒很有可能可以跨越感染之種間屏障。然而,人類上呼吸道分泌之黏蛋白擁有大量 HA 禽類型受體 26,可和大部分禽流感病毒結合,預防禽類型流感病毒進入擁有禽 類型受體之肺部,進而感染給人類。
(2) HA 快速突變演化
(2a) 抗原飄變 (antigenic drift)
(anti-HA antibodies) 中和病毒,但流感病毒之 RNA 聚合酶缺乏校正功能,容易造 成點突變 (point mutation),產生抗原飄變之現象,抗原飄變後之 HA 無法被原先 之抗體中和,病毒逃脫宿主免疫系統後,即可感染宿主其他細胞或其他宿主,並有 潛力成為新興病毒造成疫情。研究發現,1968 年香港流感病毒之基因截至 1999 年,每年平均藉由抗原飄變產生 3.5 個胺基酸突變,且其中超過半數為 HA 之抗 原飄變 43。
(2b) 抗原移變 (antigenic shift)
來自不同病毒株之 8 段 RNA,於宿主內基因重組 (gene reassortment) 後,會 形成新的病毒亞型,此種現象稱為抗原移變。人類史上之流行性感冒疫情,多來自 於禽類與人類病毒之抗原移變, 如 1957 年之亞洲流感,可能是野鴨病毒之 PB1、
HA 和 NA 三段基因與人類 H2N2 病毒,於中間宿主 (如豬) 產生基因重組所 致;1968 年之香港流感亦發現為野鴨病毒之 PB1 和 HA 與人類 H2N2,於中間 宿主 (如豬) 產生基因重組所致,而 HA 產生抗原移變後極有可能跨越種間屏障
6,44,45
。
(3) HA 蛋白質水解切位
HA 之剪切活化 (cleavage and activation) 為感染宿主之第一步。HA 藉由宿主 專一性之蛋白酶切割為重鏈 (HA1) 和輕鏈 (HA2) 兩部分,始具有感染能力,而此 切割也造就了病毒感染之組織趨性 (tissue tropism) 及致病力。研究發現高病原性禽 流感病毒 HA 之蛋白質水解切位,具有多個鹼性胺基酸,而低病原性禽流感病毒 較少。亦發現,低病原性禽流感病毒 HA 之蛋白質水解切位突變為鹼性胺基酸後,
病毒之致病力會增加 46。如 1993 年低病原性墨西哥 H5N2 禽流感病毒,其 HA 蛋白質切位為 R-E-T-R,而將其切位插入鹼性胺基酸形成 R-K-R-K-T-R 後,即轉 變為高病原性禽流感病毒 47。而雖然 HA 之切割為流感病毒感染力之主因,但致 病力高低則不完全受此限制 48,49。
1.2.4 HA 之醣質與致病力之關係 (1) HA 醣基化與功能
流感病毒所有之 HA 皆有不同程度之 N-醣基化,而高度保留之醣基於不同病 毒株及不同病毒亞型之間並不常見 50。研究各種病毒株發現,HA 之醣基化位置有 5 至 11 個,且大部分之醣基化位於 HA 之頭部球狀功能區塊 51。而 HA 依賴於 平衡之醣基化 (balance of glycosylation) 進行合適之蛋白質折疊 (folding)、病毒受 體交互作用,和有效之病毒釋放 52。
(2) HA 醣基化影響病毒與宿主結合之能力
研究發現 HA 球狀頭部之醣基化,會減少病毒與宿主之結合力,例如:HA 之 158 號胺基酸若有醣基化修飾,則其與受體之親和力下降 53,54;而於試管實驗增加 H3N2 病毒株 HA 球狀頭部之醣類,亦會減少病毒與受體之結合力,但不會影響 到 HA 之融合能力 55。
相反的,研究發現 HA 位於受體結合位附近之醣基,在病毒之複製與釋放上 扮演了決定性的角色 56,因為這些醣基調控了 HA 與受體結合之親和力 (binding affinity),並控制 HA1 結合受體之專一性 57。
(3) HA 醣基化與宿主之免疫反應
HA 球狀功能區之醣質可遮蔽宿主免疫系統辨識之抗原位置,進而從宿主免疫 系統逃脫。而 HA 亦可藉由抗原移變 (antigenic shift) 產生新的醣基化位置,遮蔽 抗原決定位置,從宿主免疫系統逃脫 55。
(4) HA 醣基化與蛋白質水解切位
HA 藉由宿主專一性之蛋白酶切割為重鏈 (HA1) 和輕鏈 (HA2) 兩部分,始具 有感染能力。研究發現 HA 之醣基化會遮蔽蛋白質水解位置,因此蛋白酶只能在 蛋白質水解切位沒有被醣質遮蔽的狀態下作用 58,59。
a b
d c
圖 1.4 血液凝集素之結構及受體結合區 本圖 a-c 使用 PBD ID 為 1HGD 之 HA 60。
a. HA 由同三聚體組成。
b. HA 單體可分為 HA1 (紅色) 及 HA2 (綠色)。
c. 俯視 HA 可發現球狀頭部受體結合區有輕微凹陷。
d. 唾液酸受體與 HA 頭部功能區塊之結合 38。
Figure 1.4 Structure and receptor binding site of hemagglutinin (a-c) Hemagglutinin from PDB ID 1HGD 60
a. HA is a homotrimer.
b. HA monomer contains HA1 (red) and HA2 (green) domains.
c. The binding sites (colors) on HA.
d. Sialic acid and receptor binding site 38
1.3 研究動機與目的
1986 年至 2003 年間臺灣陸續分離到 H6N1 及 H7N1 等多株弱毒性型家禽流行 性感冒病毒,而在 2004 年間統計有 22 間養禽場之 38 萬隻雞因發現弱毒性 H5N2 禽流感病毒潛藏而遭致撲殺。此表示目前臺灣已有 H5 和 H6 亞型禽流感感染之情 況,為瞭解目前於臺灣廣泛流行之 H6N1 禽流感病毒其蛋白質與毒力之間之關係,本 論文研究 2000 年臺大獸醫系王金和老師實驗室分離出之台灣本土 H6N1 禽流感病毒 株 (A/Chicken/Taiwan/2838/00),而 2838 病毒株經由病毒的毒性強弱,又可分為兩種 亞型,非毒性 (2838N) 及毒性 (2838V) 病毒株 61。
使用二次元電泳,配合本實驗室製備之 HA 單株抗體 62 進行免疫染色,分析比對 兩病毒株之蛋白質體間之差異後,發現兩病毒株之 HA1 皆有 6 種不同等電點的 異構 物 (isoforms)。然而 HA1 有多種等電點異構物於 30 年前就已被發現,但至今尚未研 究出 HA1 形成多種等電點異構物之原因 63。
本論文主要目的是了解 HA1 等電點異構物 (isoforms) 形成之原因,研究 HA1 是 否有被磷酸化,及 HA1 醣質是否有唾液酸化等轉譯後修飾。並利用酶切、凝集素染色 法 (lectin blot) 配合醣晶片、高效能陰離子交換層析儀 (HPAEC)、高效能層析儀配合螢 光偵測器 (HPLC-fluorescence)、 MALDI-Q-TOF MS 以及 MS/MS 建構 HA1 之醣質 表現圖譜。
第二章 材料與方法
本論文使用臺大獸醫系王金和老師,於 2000 年分離出之台灣本土 H6N1 禽 流感病毒株 (A/Chicken/Taiwan/2838/00),此 2838 病毒株經由病毒的毒性強弱又 可分為兩種亞型,非毒性 (2838N) 及毒性 (2838V) 病毒株。非毒性病毒株感染雞 隻後,並不會有任何缺失;但在毒性病毒株感染後,會有腎臟缺失並死亡 61,64。而 在實驗室內以雞胚蛋進行增殖之 2838N 及 2838V 為本實驗主要之研究材料。
2.1 禽流感病毒樣本處理方法
本實驗使用之禽流感病毒,皆由臺大獸醫系王金和老師實驗室之研究生陳怡彤 增殖、純化。使用無特定病源 (specific pathogen free, SPF) 雞胚蛋增殖之病毒,經 BEI (binary ethyleneimine) 將病毒不活化處理後,交由本實驗室進行樣本定量、分 裝並儲存於 -80°C 冰箱。
2.1.1 病毒增殖 儀器設備:
照蛋器
注射器 (Terumo, 1 mL, 25-G 5⁄8-in.) 37°C 孵蛋箱
50 mL 離心管 鑿子
水性填封膠 藥品試劑:
PBS (Phosphate buffer saline):
NaCl (0.13 M) (J.T.Baker) NaH2PO4 (0.01 M) (Wako) pH 7.0,滅菌後冷卻備用。
方法步驟:
1) 選用 9-11 日齡無特定病源 (specific-pathogen-free, SPF) 雞胚蛋,以照蛋器觀 察蛋鈍端,於氣室邊緣選擇避開血管處標記。
2) 以碘液擦拭標記處,使用鑿子鑽一個小洞,以利接種。
3) 將病毒液以滅菌 PBS buffer 稀釋成 106 倍,每個蛋以注射器接種 100 µL 至 尿囊腔,並以水性填封膠將小洞封住。
4) 將雞胚蛋置於 37°C 孵蛋器中培養,每日照蛋觀察,接種 24 小時內死亡之 雞胚視為細菌污染予以丟棄。
5) 接種後 3 天,將雞胚蛋置於 4°C 冷藏 4 hr 後,收集尿囊液進行濃縮與純化。
2.1.2 濃縮與純化 儀器設備:
高速離心機 超高速離心機 藥品試劑:
NET (NaCl-EDTA-Tween) buffer:
NaCl (0.15 M) (J.T.Baker) EDTA.2Na (5 mM) (Merck) Tween 20 (0.05%) (Merck) Tris (50 mM) (Merck) pH 8.0
TEN (Tris-EDTA saline) buffer:
NaCl (0.1 M) (J.T.Baker) EDTA.2Na (0.5 mM) (Merck) Tris (2.5 mM) (Merck) pH 8.4
Sucrose gradient solution
Sucrose (66%) (Sigma) TEN buffer
pH 8.4,配製成 66% (w/w) stock,再依所需配製各蔗糖梯度。
BEI (binary ethyleneimine) 方法步驟:
1) 將 2.1.1 中收集之尿囊液,於 4°C 以 3,000 rpm 離心 15 min。
2) 取上清液,加入 1% BEI,置於 37°°C 隔夜,使病毒不活化。
3) 於 4°C 以 70,000 g 超高速離心 3 hr。
4) 去除上清液後,以原體積 1/100 之 NET buffer 回溶。
5) 配製 10%、20%、30%、40%、50% 之蔗糖梯度。
6) 將回溶液緩緩加入蔗糖梯度中,於 4°C 以 50,000 g 超高速離心 3 hr。
7) 以針吸取可目視之病毒層,加入 4 mL NET buffer 清洗。
8) 於 4°C 以 50,000 g 超高速離心 3 hr 後,視沉澱物多寡以 200 ~ 500 µL NET buffer 回溶,置於 -80°C 保存。
2.1.3 病毒蛋白質定量
本實驗使用均勻懸浮之禽流感病毒,以 Bradford 法進行初步蛋白質定量。
儀器設備:
37°C 恆溫箱 (Memmert)
ELISA 光度計 (Dynatech Laboratories, MRX) 96 孔微量滴定盤 (Nunc, 269620)
藥品試劑:
Bio-Rad Protein assay dye reagent concentrate 5× (Bio-Rad, 500-0006) 牛血清白蛋白標準溶液 (Bio-Rad, 500-0007)
方法步驟:
1) 配製不同濃度蛋白質標準品,濃度範圍在 0~100 µg/mL。
2) 取 50 µL 各濃度標準品及經稀釋之未知濃度樣本,依序加入 96 孔盤。
3) 每孔再加入 200 µL 五倍稀釋之 Protein assay dye,輕拍 96 孔盤一側,使其 混合均勻。
4) 於 37°C 反應 10 min 後,以 ELISA 光度計讀取 570 nm 之吸光值。
5) 將不同濃度蛋白質標準品之 570 nm 吸光值繪成標準曲線,以內插法求得樣 本蛋白質濃度。
2.2 一般電泳檢定法
本論文皆使用 15% SDS 膠體電泳 (sodium dodecyl sulfate-PAGE),以迷你平 板直立式電泳套件進行。
2.2.1 SDS 膠體電泳 儀器設備:
電泳玻璃片及鋁片 (10×8 cm 及 10×10 cm) 間隔條 (spacer, 0.75 mm 及 1 mm)
齒梳 (comb, 10 well 及 15 well) 鑄膠套件 (Hoefer)
平板式垂直迷你電泳槽 (Hoefer, SE-250) 電源供應器 (Pharmacia, EPS 301)
藥品試劑:
A 液:丙烯醯胺溶液:
Acrylamide (Sigma) 29.0 g N,N’-Methylene-bis-acrylamide (Bio-Rad) 0.8 g 加二次水至 100 mL 溶解,置於 4°C 避光儲存。
B 液:分離膠體緩衝液 (Running buffer):
Tris (1.5 M) (Bioman) 45.4 g
TEMED (Sigma) 0.9 mL
以 200 mL 的二次水溶解後,使用 HCl (J.T.Baker) 將 pH 調至 8.8,再定量 至 250 mL。置於 4°C 避光儲存。
◆ TEMED:N,N,N’,N’-Tetramethyl-ethylenediamine C 液:焦集膠體緩衝液 (Stacking buffer):
Tris (0.5 M) (Bioman) 6 g TEMED (Sigma) 0.4 mL
以 80 mL 的二次水溶解後,使用 HCl 將 pH 調至 6.8,再定量至 100 mL。
置於 4°C 避光儲存。
過硫酸銨溶液 (APS):
Ammonium persulfate (10 %) (Bio-Red) 100 mg 取 100 mg ammonium persulfate 溶於 1 mL 二次水中,使用前新鮮配置。
異丙醇 (Isopropanol) (Mallinckrodt chemicals)
SDS 電泳緩衝液 (5×):
Tris (90 mM×5) (Bioman) 54.5 g EDTA-2Na (2.5 mM×5) (J.T.Baker) 4.7 g Boric acid (80 mM×5) (RDH) 24.8 g
加二次水 800 mL 溶解,以 HCl 將 pH 調至 8.4,定量至 1,000 mL。使用 時稀釋 5 倍,加入 10% SDS,使得最終濃度為 0.1% SDS,室溫保存。
SDS 膠體電泳樣品溶液 (SDS-PAGE sample buffer, 2×):
Tris (125 mM×2) (Bioman) EDTA.2Na (2 mM×2) (J.T.Baker)
sodium dodecyl sulfate (2%×2) (J.T.Baker) β-Mercaptoethanol (5%×2) (Sigma)
Bromophenol blue (0.01%×2) (Sigma) Glycerol (20%) (J.T.Baker) pH 6.8
10% SDS 溶液
預先染色之蛋白質分子量標準品 (prestained protein ladder) (Bioman)
1) 利用表 2.1 鑄 15% SDS 電泳膠片
表 2.1 常用的 SDS 膠體電泳溶液之參考濃度配方
分離膠體溶液 焦集膠體
鑄膠溶液
5% 7.5% 10% 12.5% 15% 20% 4%
A 1.65 2.5 3.3 4.15 5.0 6.7 0.66
B 2.5 - C - 1.24
10% SDS 0.1 0.05
H2O 5.7 4.85 4.05 3.2 2.35 0.65 2.95
APS 0.05 0.1 (單位︰mL)
2) 取適量的樣品 (10 µL/well),加入同體積之 SDS 膠體電泳樣品溶液,混合均 勻後於 100°C 中煮沸 10 min,迅速置於冰上。
3) 取 3 µL 蛋白質分子量標準品注入最左側樣本槽中,並取 10 µL 步驟 2 處理 之樣品,注入各樣品槽。
4) 蓋上電泳槽蓋子,確認正負極裝置正確,連接電源供應器,定電壓 160 V 進 行電泳。
5) 待追蹤染料跑出膠片外,即可停止電泳。輕輕撬起玻片切去焦集膠體部份,
在分離膠體右上截角作為正反面之記號,準備進行染色或轉印。
2.2.2 膠體染色法 - CBR染色(Coomassie Brilliant Blue R-250)
儀器設備:
平台震盪器 (TKB, OS701) 染缸
藥品試劑:
CBR 染色液:
CBR (Sigma) 1.5 g
Methanol (Mallinckrodt chemicals) 250 mL
CH3COOH (J.T.Baker) 50 mL
以 Methanol 溶解 CBR 後二次水定量至 250 mL,混勻過濾後使用。
CBR 脫色液:
Methanol (Mallinckrodt chemicals) 200 mL CH3COOH (J.T.Baker) 100 mL 加二次水定量至 1 L。
甲醇溶液 (50%):
Methanol (Mallinckrodt chemicals) 500 mL 加二次水定量至 1 L。
方法步驟:
1) 取下電泳後的膠體置於染缸,CBR 染色液須完全蓋過膠體,置於平台震盪 器上搖盪約 30 min。
2) 倒出染色液回收,並倒入脫色液完全蓋過膠體,於平台震盪器上搖盪 1 hr。
3) 置換新的脫色液 1 至 2 次,直至背景完全透明則脫色完成。
2.2.3 膠體染色法-銀染(Silver staning)
儀器設備:
平台震盪器 (TKB, OS701) 玻璃染缸
藥品試劑:
還原液甲:
Glutaraldehyde (25% GR, Merck) 4 mL Na2S2O3.5H2O (Nakalai) 0.8 g 以二次水定量至 100 mL。
還原液乙:
0.5% Citric acid (USB) 1 mL Formaldehyde (37% GR, Merck) 0.1 mL
Methanol (Mallinckrodt chemicals) 15 mL 以二次水定量至 100 mL。
硝酸銀液:
Silver nitrate (J.T. Baker) 0.8 g 0.36% NaOH (Merck) 21 mL Ammonia solution (28~30% GR, J.T.Baker) 1.4 mL
先將硝酸銀溶於 4 mL 二次水,再緩慢滴入上述二者的混合液中,邊滴邊搖 晃,當全部硝酸銀液滴完時,溶液應呈現澄清。若有混濁現象須以氨水反滴 定至澄清。
反應終止液:
0.5% Citric acid (USB) 10.0 mL Ethylenediamine (Wako) 0.1 mL 以二次水定量至 100 mL。
方法步驟:
1) 取下電泳後的膠體置於玻璃染缸,以 50%甲醇洗三次,每次至少 10 min,或 浸泡隔夜。
2) 倒去甲醇,以二次水洗三次,每次 10 min。
3) 加入還原液甲,反應 1 h。
5) 加入硝酸銀溶液,反應 15 min。
6) 回收硝酸銀液至廢液桶,用二次水洗 3 次,每次 10 min。
7) 加入還原液乙呈色。在背景顏色未加深前倒去還原液乙。
8) 以二次水清洗膠片,加入反應終止液終止呈色,浸泡 1 h 後乾片。
2.2.4 蛋白質電泳轉印法 儀器設備:
電泳轉印槽 (Hoefer, TE 52) 及轉印卡夾、海綿等套件 電源供應器 (Pharmacia, EPS 301)
電磁攪拌器 (Corning PC-420D) 攪拌子
PVDF 轉印膜 (Millipore, Immobilon P) 濾紙 (Whatman 3 mm)
藥品試劑:
轉印緩衝液 (Blotting buffer):
CAPS (10 mM) (J.T.Baker) 2.22 g
加水 600 mL 溶解,以 NaOH 調整 pH 至 11 後,加甲醇 100 mL,再定量 至 1,000 mL;最後含 10% 甲醇。
預先染色之蛋白質分子量標準品 (prestained protein ladder) (Bioman) 方法步驟:
1) 將電泳後之膠片浸於轉印緩衝液平衡 30 min。
2) 將疏水性 PVDF 轉印膜先以 100% 甲醇充分潤溼後,浸入轉印緩衝液中平 衡至少 5 min 後使用。
3) 裝置膠片、轉印膜與卡夾並置入轉印槽中,並開啟攪拌器。
4) 固定電流以 400 mA 轉印 100 min 後終止。
2.2.5 轉印膜 Ponceau S 染色法 儀器設備:
染缸 藥品試劑:
Ponceau S 染色液:
Ponceau S (Sigma) 0.2 g
CH3COOH (J.T.Baker) 10 mL
二次水定量至 250 mL,混合均勻後使用。
方法步驟:
1) 將轉印完成的轉印膜置於染缸,倒入蓋過轉印膜的 Ponceau S 染色液。
2) 輕輕搖晃,使轉印膜被 Ponceau S 染色液均勻浸潤後倒去。
3) 以水漂洗數次,此時可見轉印膜上的蛋白質染成紅色。
4) 再以水清洗至所有紅色染劑消失。
2.3 免疫染色法 儀器設備:
平台震盪器 (TKB OS701)
冷光螢光影像分析系統 (UVP, AutoChemi image system) 藥品試劑:
Urea-PBST:
Urea (6 M) (RDH) 180 g
加入 200 mL PBST 加熱溶解後,以 PBST 定量至 500 mL。
Gelatin-NET (NaCl-EDTA-Tween):
Gelatin (0.25%) (Merck) 5 g NaCl (0.15 M) (Bioshop) 17.5 g EDTA.2Na (5 mM) (J.T.Baker) 3.6 g Tween 20 (0.05%) (Plus one) 1 mL Tris (50 mM) (Bioman) 12.1 g
先以 500 mL 二次水將 gelatin 加熱溶解,然後加其他藥劑補二次水至 1,700 mL,調 Ph 至 8.0,再定量至 2,000 mL。
PBS (Phosphate buffer saline):
詳見 2.1.1 節。
PBST (Phosphate buffer saline & Tween-20):
將 PBS 加入 0.05% (v/v) 的 Tween-20。
二次抗體 (Horse radish peroxidase-goat-anti-mouse, HRP-GAM):
Peroxidase-conjugated goat IgG fraction to mouse immunoglobulins (IgG, IgM,
IgA) (Cappel 5556)
使用前以 Gelatin –NET 稀釋 4000 倍。
ECL 冷光基質:
ECL Western Blotting Substrate (Pierice, 32106)
等量 A 液與 B 液使用前新鮮配製,覆蓋過轉印膜即可。
1) 轉印膜以 Uera-PBST 浸泡 30 min 並倒掉後,以 PBST 洗三次,每次 10 min。
2) 加入 Gelatin-NET 完全蓋過轉印膜,反應 1 hr 後倒掉。
3) 加入以 Gelatin-NET 稀釋 3,000 倍之一次抗體,覆蓋轉印膜,室溫下反應 1 h 後倒掉。並以 PBST 洗 3 次,每次 10 min。
4) 加入以 Gelatin-NET 稀釋 4,000 倍之二次抗體,室溫下反應 1 h,倒掉後以 PBST 洗 3 次,每次 10 min,最後保存在 PBS 中。
5) 加入 ECL 冷光基質完全蓋過轉印膜,以冷光螢光影像分析系統呈相。
2.4 凝集素染色法 (lectin blot)
使用 DIG Glycan Differentiation kit (Roche) 進行實驗。
儀器設備:
平台震盪器 (TKB OS701) 藥品試劑:
TBS buffer (Tris buffered saline)
Tris (0.05 M) (Bioman) NaCl (0.15 M) (Bioshop) pH 7.5
緩衝液 1 (Buffer 1) TBS buffer
MgCl2 (2 mM) (Nacalai) MnCl2 (2 mM) (Merck) CaCl2 (2 mM) (sigma) pH 7.5
緩衝液 2 (Buffer 2)
Tris (0.1 M) (Bioman) MgCl2 (0.05 M) (Merck) NaCl (0.1 M) (Bioshop) pH 9.5
DIG Glycan Differentiation kit (Roche) 1. Blocking Reagent
2. Maackia amurensis agglutinin (MAA) 3. Sambucus nigra agglutinin (SNA) 4. Anti-Digoxigenin-AP
5. NBT/BCIP solution
方法步驟:
1) 轉印膜以 Blocking Reagent 於 4°C 浸泡隔夜,倒掉後以 TBS 洗二次,每 次 10 min,接著以 Buffer 1 洗 10 min。
2) 分別吸取 10 µL 之 MAA 及 50 µL 之 SNA,加入 Buffer 1,使兩種凝集素 溶液體積為 10 mL。
3) 將凝集素溶液覆蓋轉印膜,反應 1 hr。
4) 倒掉凝集素溶液後,以 TBS 洗三次,每次 10 min。
5) 吸取 10 µL 之 Anti-Digoxigenin-AP,加入 Buffer 1,使液體積為 10 mL。
加入並覆蓋轉印膜,反應 1 hr。
6) 倒掉 Anti-Digoxigenin-AP 溶液 後,以 TBS 洗三次,每次 10 min。
7) 吸取 200 µL 之 NBT/BCIP 溶液,加入 Buffer 2,使液體積為 10 mL。
8) 加入 NBT/BCIP 成色液並覆蓋轉印膜,避光反應呈色,並於背景黑掉前,
倒掉成色液,並以二次水清洗。
2.5 醣晶片
使用 Qproteome GlycoArray kit (QIAGEN) 進行實驗。
儀器設備:
平台震盪器 (TKB, OS701) Qproteome GlycoArray kit
晶片掃瞄器 (Axon GenePix 4100A) Qproteome GlycoArray software 藥品試劑:
Qproteome GlycoArray kit (QIAGEN):
blocking solution wash solution
螢光標定之抗小鼠單株抗體 (二次抗體)
血液凝集素單株抗體 (ㄧ次抗體) 方法步驟:
1) 取將框架黏貼至醣晶片之載玻片上,並將載玻片放入培養品中。
2) 加入 blocking solution 搖盪反應 1 hr,到掉後,以 wash solution 洗三次,
每次 5 min。
3) 以拭鏡紙將框架擦乾後,加入 450 µL 之 0.2 mM 蛋白質樣本溶液,搖盪反 應 1 hr ,倒掉後,以 wash solution 洗三次,每次 5 min。
4) 以拭鏡紙將框架擦乾後,加入 450 µL 螢光標定之二次抗體,避光搖盪反應
5) 使用 二次水清洗 1 min 後,將醣晶片風乾後。
6) 使用掃描器掃描晶片,並以軟體分析。
2.6 二次元膠體電泳
2.6.1 脫鹽及溶解 儀器設備:
微量高速離心機 (Beckman, Microfuge) 小型真空離心機 (Thermo, SpeedVac) 超音波震盪器 (Branson, Sonicater) 藥品試劑:
10% TCA/acetone
TCA (10%) (RDH) 50 mL
Acetone (J.T. Baker) 450 mL
保存於 -20°C
100% acetone (J.T. Baker) IPG sample buffer:
Urea (6 M) (RDH) 9 g
Thiourea (2 M) (Amersham) 3.8 g Triton X-100 (0.5%) (Sigma) 0.125 mL 加水至 25 mL,每 mL 分裝一管,置 -20°C 保存,使用前解凍充份回溶,
每管添加 10 mg DTT 及 5 µL IPG buffer (pH 3~10)。
方法步驟:
7) 取 30 ~ 120 µg 病毒樣本,加入 1.5 mL 之 10% TCA/acetone,置於 -20°C 隔 夜,沉澱蛋白質。
8) 於 4°C 以 12,000 rpm 離心 20 min,去除上清液。
9) 加入 1.5 mL 之 100% acetone 震盪,並於 4°C 以 12,000 rpm 離心 20 min,去除上清液。重複此步驟三次。
10) 使用 SpeedVac 將 acetone 抽乾。
11) 加入 125 µL sample buffer 震盪回溶。
2.6.2 第一維等電點聚焦 儀器設備:
EttanTM IPGphorTM isoelectric focusing system (Amersham) Strip (Amersham, 7 cm, pH 3~10) 連續性 strip
Holder (Amersham, 7 cm)
鑷子 藥品試劑:
Dry strip cover oil (Plus one) 方法步驟:
1) 取 125 µL Sample buffer 回溶完全之樣品加入 7 cm holder 中。
2) 放入 pH 3~10 之 IPG strip。
3) 正負二極各加入 0.2 mL cover oil,蓋上蓋子,使蓋子吸住密封 holder。
4) 將 holder 置於 IPGphor system 電極板上,兩端接點確實與電極板接觸,以 30 V 電壓將 strip rehydration 12 hr,使 strip 上膠體膨潤,再進行表 2.2 設 定之 program。
表 2.2 等電點聚焦 program
電壓 (V) Volt-hours (Vhr)
500 250 1,000 500 2,000 1,000 5,000 10,000 8,000 16,000
◆ 系統溫度保持 25°C,每條 strip 電流限制在 50 µA。
2.6.3 第二維 15% SDS-PAGE 膠體電泳 儀器設備:
同 2.2.1 節。
◆ 使用 10×10 cm 電泳玻璃片及鋁片,1 mm 間隔條,不使用齒梳,電泳槽 換 SE-260 底部。
藥品試劑:
SDS-PAGE 電泳試劑同 2.2.1 節
SDS 平衡 buffer:
1.5 M Tris-HCl buffer (50 mM) 10 mL SDS (2%) (J.T.Baker) 4 g Glycerol (30%) (J.T.Baker) 60 mL
Urea (6 M) (RDH) 72 g
Bromophenol blue (0.01%) (sigma) 200 µL 加水至 200 mL。
Agarose:
SDS running buffer 100 mL 將兩者混合後,在微波爐加熱溶解。
DTT (dithiothreitol) (USB)
IAA (iodoacctamide) (Amersham) 方法步驟:
1) 將進行完 IEF 之 strip,以二次水清洗後,放入 5 mL 含 50 mg DTT 的 SDS 平衡緩衝液,平衡 20 min 後,再放入 5 mL 含 125 mg IAA 的 SDS 平衡 緩衝液,平衡 20 min。
2) 以二次水清洗 strip,並將 strip 靠右放入 SDS-PAGE 膠片中。
3) 將滴有蛋白質分子量標準品之 4 mm2 濾紙,以 agarose 封住後,靠左放入 SDS-PAGE 膠片中。
4) 以 agarose 進行封膠。
5) 將上述膠體放入電泳槽,依 2.2.1 節的方法固定電壓 160 V 進行電泳。
6) 電泳完成後,取下膠片去除 strip 及 agarose,於右上角截角標記。
2.7 酶切實驗
2.7.1 鹼性磷酸酶之去磷酸化 儀器用具:
37°C 恆溫箱 (Memmert) 藥品試劑:
Calf intestinal alkaline phosphatase (CIAP) (Promega) 方法步驟:
1) 取 30 µg 病毒蛋白質,加入 3 µl (3 U) 鹼性磷酸酶,於 37°C 下反應 48 hr。
2) 加入 1.5 ml 10% TCA/Acetone 於 -20°C 終止反應並沉澱蛋白質。
3) 參照 2.6 節,以二次元電泳進行分析。
2.7.2 神經胺酸水解酶水解唾液酸 儀器用具:
37°C 恆溫箱 (Memmert)
藥品試劑:
神經胺酸酶反應液:
Neuraminidase (Roach,Cat. No. 11 080 725 001) SDS (0.1%) (J.T.Baker)
Triton X-100 (0.5%) (Sigma)
β- Mercaptoethanal (0.5%) (Sigma)
此品號 Neuraminidase 為 pH 5 之液態溶液,加入以上三種蛋白質變性劑 後,即可使用。
方法步驟:
1) 取 30 µg 病毒蛋白質,加入 40 µl (40 mU) 神經胺酸酶反應液,於 37°C 下 反應 1 ~ 48 hr。加入 1.5 ml 10% TCA/Acetone 於 -20°C 終止反應並沉澱蛋 白質。
2) 參照 2.6 節,以二次元電泳進行分析。
2.8 使用高效能陰離子交換層析儀 (HPAEC) 配合脈衝式安培法偵測器 (PAD) 分析 唾液酸
2.8.1 弱酸水解釋放唾液酸 儀器用具:
小型真空離心機 (Thermo, SpeedVac) 37°C 恆溫箱 (Memmert)
藥品試劑:
Neuraminidase (Roach,Cat. No. 11 080 725 001) 方法步驟:
1) 使用 60 µg 病毒蛋白進行電泳後,將蛋白質轉印到 PVDF 膜上,並剪切下 目標蛋白質。
2) 加入 80 µl 之神經胺酸水解酶,於 37°C 反應 48 hr。
3) 將 PVDF 膜取出後,以 SpeedVac 於 40°C 抽乾。
4) 抽乾後之唾液酸於 -20°C 保存備用。
2.8.2 使用 HPAEC-PAD 分析唾液酸亞型 儀器用具:
ICS 3000 (Dionex)
Chromatography system Gradient Pump
Electrochemical Detector Autosampler
Chromeleon chromatography software (Dionex)
CarboPac PA10 Analytical Column (2 x 250 mm) (Dionex)
Filter (Millex-GV PVDF membrane,0.22 µm、4 mm ) 藥品試劑:
N-Acetylneuraminic acid (Neu5Ac) (Sigma) N-Glycolylneuraminic acid (Neu5Gc) (Sigma)
3-Deoxy-D-glycero-D-galacto-2-nonulosonic acid (KDN) (Toronto Chemicals) Sodium hydroxide, 50% (w/w) (Fisher Scientific)
Sodium Acetate (Merck) 100 mM Sodium Hydroxide
100 mM Sodium Hydroxide/1 M Sodium Acetate 方法步驟:
1) 裝置並以 100 mM NaOH 流洗 CarboPac PA10 及其 guard column。
2) 使用 100 µl 二次水回溶樣本,過濾,裝入樣本瓶後,裝入 autosampler。
3) 使用軟體設定以下程式,並設定流速為 0.25 mL/min
(Dionex Technical note 41) 4) 啟動程式,待程式終止後,以軟體分析。
2.9 蛋白質身分鑑定
2.9.1 膠體內蛋白酶水解 儀器用具:
平台震盪器 (TKB, OS701)
微量高速離心機 (Beckman, Microfuge) 小型真空離心機 (Thermo, SpeedVac) 超音波震盪器 (Branson, Sonicater) 37°C 恆溫箱 (Memmert)
試劑藥品:
Ammonium bicaronate 緩衝溶液
NH4HCO3 (0.25 M) (Sigma) 2.43 g
調 pH 至 8.5,補二次水至 100 mL,使用前加入 DTT 1.54 mg/mL 或 IAA 17.5 mg/mL
蛋白質脫色液:
NH4HCO3 (25 mM) (Sigma) ACN (50%) (Labscan) pH 8.5
酵素緩衝液:
NH4HCO3 (0.2 N) (Sigma) 1.94 g CaCl2 (0.5 mM) (Sigma) 5.5 mg 加二次水至 100 mL。
萃取溶液:
TFA (0.1%) (Merck) 0.1 mL
ACN (60%) (Labscan) 60 g 加二次水至 100 mL。
DTT (dithiothreitol) (USB)
IAA (iodoacctamide) (Amersham) 方法步驟:
1) 膠體經 CBR 染色後,以二次水清洗二次,每次 2 h。
2) 將目標蛋白質點從膠片上割下,放入經甲醇清洗過之微量離心管中。
3) 加入 100 µL 10 mM DDT/25 mM ammonium bicarbonate,pH 8.5 緩衝溶液中,
在 37°C 下反應 1 h。
4) 以 10,000 rpm 離心 1 min 以去除 DTT。
5) 加入 100 µL 100 mM iodoacetamide (IAA)/25 mM ammonium bicaronate,pH 8.5 緩衝溶液,於室溫避光反應 1 h。
6) 以 10,000 rpm 離心 1 min 以去除 IAA。
7) 加入蛋白質脫色液,於 30°C 反應 15 min,重複兩次。
8) 以二次水清洗兩次,每次 10 min,以去除 CBR,最後以 SpeedVac 抽乾。
9) 加入含有 0.1 µg trypsin 之酵素緩衝液 10 µL,水解反應 10 min。
10) 再加入 100 µL 酵素緩衝液,於 37°C 下反應隔夜。
11) 加入 200 µL 萃取溶液。
12) 在 35~40°C 下超音波震盪 30 min,收集萃取液於微量離心管中,重複一次。
13) 用 SpeedVac 抽乾。
2.9.2 以質譜儀鑑定蛋白質點身分 儀器用具:
ESI-Q-TOF (Micromass) 方法步驟:
1) 將經 2.5.1 處理之樣本,送交鐠德科技股份有限公司以 ESI-Q-TOF 質譜儀分 析。
2) 將送回之原始資料傳送至 Mascot 資料庫平台比對蛋白質身分,主要比對資 料庫為 MSDB 或 NCBInr。
2.10 高效能層析儀 (HPLC) 配合螢光偵測器分析 N-醣質
2.10.1 醣質之膠內水解釋放及螢光標定 儀器用具:
65°C 水浴槽 (Kansin instruments) 試劑藥品:
N-Glycosidase F (PNGase F) (BioLabs) N-Glycosidase F 之酵素緩衝液:
25 mM ammonium bicarbonate,pH 8.5 (Sigma) TFA (Merck)
ACN (Labscan)
2-aminobenzamide (Merck) Acetic acid (J.T.Baker) DMSO (Sigma)
Sodium cyanoborohydride (Merck) 冷凍乾燥機
微量高速離心機 (Beckman, Microfuge)
方法步驟:
1) 膠體經 CBR 染色後,參考方法 2.9.1 將 CBR 去除。
2) 加入含有 3 unit N-Glycosidase F 之酵素緩衝液,於 37°C 反應 72 hr。
3) 加入 500 µl 100% ACN/2%TFA,超音波震盪 15 分鐘,離心蒐集上清液。
4) 加入 500 µl 50% ACN/1%TFA,超音波震盪 15 分鐘,離心蒐集上清液。
5) 加入 500 µl 0.1%TFA/ddH20,超音波震盪 15 分鐘,離心蒐集上清液。
6) 重複步驟 5。
7) 加入 500 µl 100% ACN 震盪,離心取上清液。
8) 將前步驟上清液集中,以 10,000 rpm 離心 1 min,取上清液進行冷凍乾燥。
9) 將 1.5 mg 2AB 加入 100 µl 之 30% acetic acid/DMSO 中,混合均勻。
10) 再加入 5.15 mg sodium cyanoborohydride,混合均勻。
11) 吸取 15 µl 步驟 10 配製之試劑,回溶乾燥之醣質,於 65°C 避光反應 3hr。
2.10.2 使用 HPLC 分析 N-glycans 儀器用具:
TSKgel amide-80 column (TOSOH) HITACH L-7100 pump
螢光偵測器 (Waters 2475) 軟體 (SISC 32)
試劑藥品:
20% 50 mM NaOAc/ACN 50 mM NaOAc
方法步驟:
1) 裝置並以 NaOH 20% 50 mM NaOAc 流洗管柱。
2) 注入 10 µl 樣本。
3) 使用軟體設定以下程式,並定流速為 1 mL/min,excitation 330 nm, emission 420 nm
Column: TSKgel amide-80
Detection: fluorescence detector Flow rate: 1 mL/min
Eluent: A. 20% 50 mM NaOAc/ACN B. 50 mM NaOAc
0%
100%
120 min
50%
50%
110 min
50%
50%
100 min
0%
100%
20 min
0%
100%
0 min
B A
0%
100%
120 min
50%
50%
110 min
50%
50%
100 min
0%
100%
20 min
0%
100%
0 min
B A
4) 啟動程式,待程式終止後,以軟體分析。
2.11 質譜儀醣質定序
2.11.1 從醣蛋白質上釋放並純化 N-glycans 儀器用具:
37°C 保溫箱 (Memmert) Sep-Pac C18 column (Waters)
藥品試劑:
N-Glycosidase F 之酵素緩衝液:
25 mM ammonium bicarbonate,pH 8.5 (Sigma) N-Glycosidase F (PNGase F) (BioLabs)
Acetonitrile (ACN) (J.T.Baker) Formic acid (Sigma)
方法步驟:
1) 將方法 2.9.1 萃取乾燥後之 peptide,加入含有 3 unit N-Glycosidase F 之酵 素緩衝液,於 37°C 反應 72 hr。
2) 進行 Sep-Pac C18 column 之再生:
以針筒吸取 5 ml 100% ACN 流洗管柱,於兩分鐘完成。
以針筒吸取 5 ml 50% ACN/0.1% Formic acid 流洗管柱,於兩分鐘完成。
以針筒吸取 5 ml 0.1% Formic acid 流洗管柱,於兩分鐘內完成。
3) 接著注入樣品等待 1 min 後,使用 5 ml 0.1% Formic acid 流洗管柱,並以 玻璃管收集流洗出之 N-glycans。
4) 使用 2 ml 50% ACN/0.1% Formic acid 流洗管柱後,以 3 ml 75% ACN/1%
Formic acid 流洗管柱,並以 15 ml 離心管收集流洗出之 peptides。
5) 將收集之 N-glycans 與 peptides 冷凍乾燥。
2.11.2 N-glycans 之泛甲基化 儀器用具:
玻璃管及玻璃滴管 研缽
冷凍乾燥機 超音波震盪器 藥品試劑:
NaOH (J.T.Baker) DMSO (Sigma)
CH3I (KANIO chemical) CHCl3 (J.T.Baker)
方法步驟:
1) 將 1 g NaOH 加入 DMSO 內,並以研缽研磨至雲狀。
2) 以玻璃滴管吸取 200 µl DMSO/NaOH 加入裝有醣質之玻璃管,於超音波震 盪 30 min。
3) 以玻璃滴管吸取 100 µl CH3I 加入玻璃管,並於超音波震盪 60 min。
4) 重複步驟 2、3。
5) 加入 3 ml 之 CHCl3 後,加入 8 ml H2O 萃取 NaOH,移除水層,重複加入 8 ml H2O 萃取 NaOH 多次,至水層不再呈鹼性為止。
6) 移除水層,並將含有醣質之有機層冷凍乾燥。
2.11.3 以質譜儀分析 N-glycans 儀器用具:
小型真空離心機 (Thermo, SpeedVac) MICROMASS MALDI MicroMX (Waters) 藥品試劑:
70% methanol
2,5-dihydroxybenzoic acid (DHB) 方法步驟:
1) 以 200 µL 70% methanol 回溶醣質,吸取至 1.5 mL 微量離心管,並以 SpeedVac 抽乾。
2) 將醣鏈以 70% methanol 回溶,並以基質溶液 (DHB,20 mM sodium acetate) 稀釋至 3 µL,將樣本點至 MALDI plate 上,真空乾燥後,進行 MALDI TOF/TOF 質譜分析
第三章 結果
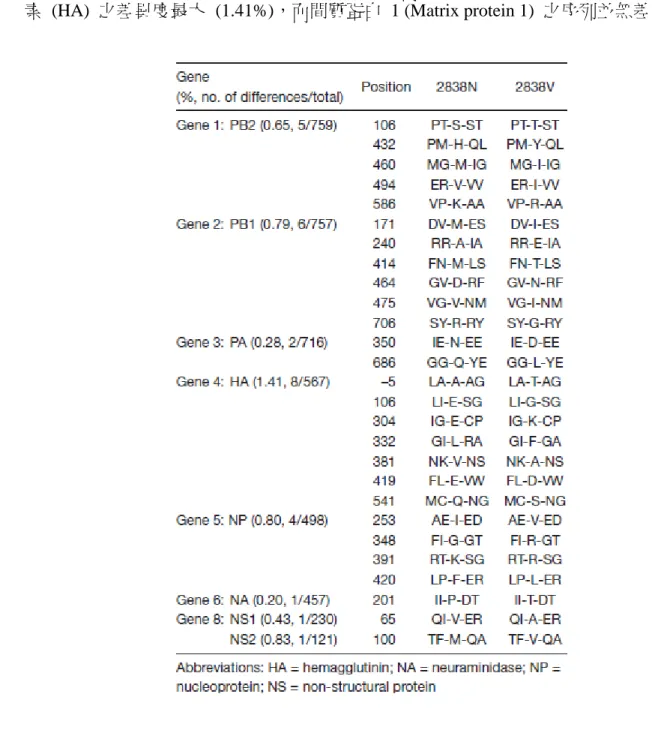
本論文研究源起於 2000 年,王金和老師實驗室分離出台灣本土 H6N1 禽流感病 毒株 (A/Chicken/Taiwan/2838/00),而 2838 病毒株經由病毒的毒性強弱又可分為兩種 亞型,非毒性 (2838N) 及毒性 (2838V) 病毒株 61。毒性及非毒性禽流感病毒株之八段 基因已於 2002 年完成定序工作,而兩病毒株之間胺基酸序列之差異如表 3.1 所示 64。
為了解兩病毒株之蛋白質體間之差異,本論文使用二次元電泳配合本實驗室製備 之 HA 單株抗體 62 進行免疫染色分析比對。發現兩病毒株之血液凝集素 1 (HA1) 皆 有 6 種不同等電點的異構物,為了解異構物形成之原因,實驗轉向蛋白質之轉譯後修 飾。
表 3.1 非毒性與毒性 2838 禽流感病毒株各基因之間胺基酸序列的差異 64
Table 3.1 Comparison of amino acid sequences from the genes of non-virulent and virulent 3838 avian influenza viruses strains 64
經由胺基酸序列比對發現,非毒性及毒性之禽流感病毒株之八段基因中,血液凝集 素 (HA) 之差異度最大 (1.41%),而間質蛋白 1 (Matrix protein 1) 之序列並無差異。
3.1 比較非毒性與毒性禽流感病毒株之間蛋白質體的差異
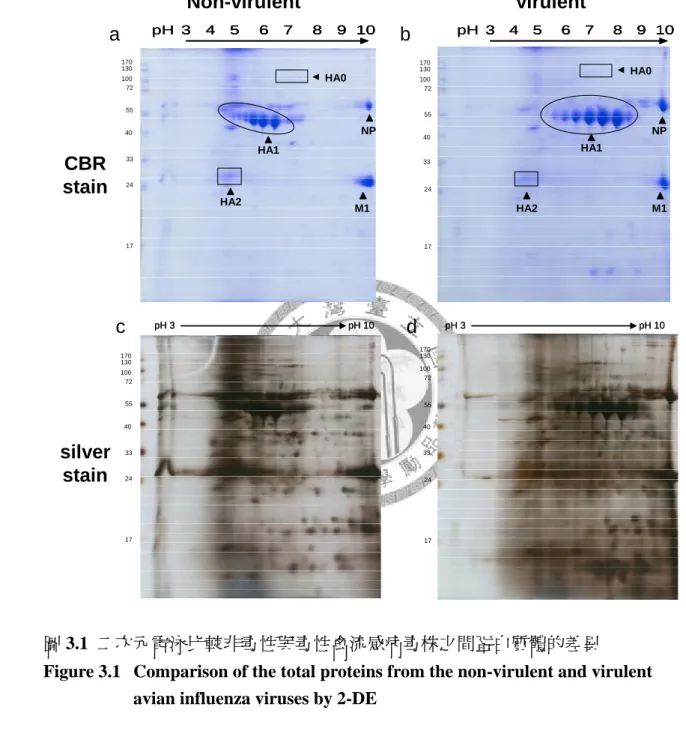
使用由雞胚胎之尿囊液純化出之禽流感病毒,以 10% TCA/Acetone 沉澱其蛋白 質,並進行二維電泳,使用 Coomassie blue 及硝酸銀染色法比對非毒性與毒性 AIV 蛋 白質體之差異。結果發現 HA1 為二維圖譜上差異最大之蛋白質點。接著使用血液凝集 素 (HA) 單株抗體進行免疫染色,發現非毒性及毒性 AIV 之 HA1 皆有 6 種不同等 電點之異構物。
3.1.1 兩株 2838 禽流感病毒二次元電泳圖譜之比較
本實驗使用病毒全蛋白進行二次元電泳分析,比較非毒性 (2838N) 與毒性 (2838V) 禽流感病毒株之蛋白質體。圖 3.1 a,b 使用 Coomassie blue 染色,並以質 譜儀進行蛋白質份鑑定,可在蛋白質圖譜中發現最為豐富的三種蛋白質。第一種為 佔蛋白質總量最大之間質蛋白 1 (M1),M1 分子量約 24 kD 為結合流感病毒 RNA 之蛋白質,因此 M1 富含鹼性胺基酸,其位於圖譜中 pH 10 的位置,但事實上 M1 等電點可能大於 10。第二種為核殼蛋白 (NP),NP 約 60 kD 其本身為包覆 RNA 的 蛋白質,因此其本身等電點亦可能超過 10。第三種為血液凝集素 (HA),圖譜中可 發現分子量約 130 kD 之 HA0,50 kD 之 HA1 及 25 kD 之 HA2。圖 3.1 c,d 為圖 3.1 a,b 之膠片進行硝酸銀染色的結果,可發現更多蛋白質點,這些蛋白質有些來 自病毒本身,有些來自於病毒增殖用之雞胚胎。比較非毒性與毒性禽流感病毒株之 蛋白質體可發現差異最大的為 HA1 之蛋白質群點。
3.1.2 血液凝集素重鏈 (HA1) 為兩禽流感病毒株蛋白質圖譜上差異最大之蛋白質
血液凝集素 (HA) 為非毒性 (2838N) 與毒性 (2838V) 禽流感病毒株之胺基酸 序列差異度 (1.41%) 最大的蛋白質,而在二次元蛋白質圖譜中,血液凝集素重鏈 (HA1) 亦為差異最大的蛋白質,非毒性株 HA1 等電點較毒性株低。圖 3.2 為兩株 病毒 HA 以 BLAST 進行分析之結果,可發現 16 個胺基酸之訊號胜肽中有 1 個 胺基酸不同;而 HA2 的 222 個胺基酸中有 5 個胺基酸不同,但這 5 個胺基酸之 不同,並未使兩株病毒之 HA2 在二次元蛋白質圖譜中產生明顯差異;反觀 HA1,
HA1 在 329 個胺基酸中只有 2 個不同,但此 2 個胺基酸差異卻使非毒性與毒性株 之 HA1 相差 3 個電荷。2838N 中 HA 之第 106 個胺基酸為酸性的麩胺酸 (Glu),但 2838V 為中性的甘胺酸 (Gly);另外,2838N 之第 304 個胺基酸為酸性 的麩胺酸 (Glu),但 2838V 卻為鹼性的離胺酸 (Lys),而 HA 之第 106 及 304 個胺 基酸,就是 HA1 在 329 個胺基酸中僅有的兩個不同,此差異造成兩株病毒之 HA1 相差三個電荷。以 EMBOSS IEP 軟體預測 HA1 之等電點得知,2838N 之 HA1 等 電點為 6.8764;2838V 之 HA1 等電點為 7.9163,此預測結果可和蛋白質二維圖譜 HA1 之蛋白質群點互相呼應,也就是非毒性株 HA1 之等電點較毒性株低,可能源 自於蛋白質一級結構。
3.1.3 非毒性與毒性 AIV 之 HA1 皆有 6 種不同等電點之異構物
雖然兩株病毒之 HA1 等電點差異甚大,但皆有 6 種不同等電點之異構物。圖 3.3 使用 HA 單株抗體進行免疫染色,再次證實。延續 3.1.2 中 HA1 等電點之預 測,2838N 之 HA1 等電點為 6.8764;2838V 之 HA1 等電點為 7.9163,此二預測 之 pI 值,恰好是 HA1 在蛋白質圖譜的 6 個點中,pI 值最大的點,也就是最鹼性 的蛋白質點。因此推測其餘 5 個 pI 值較小,較偏酸性之 HA1 異構物,可能是由不 同程度之蛋白質轉譯後修飾使 HA1 酸化所造成。