國 立 台 東 大 學 生 命 科 學 所 碩士論文
苦參對藥物代謝酶之誘導及其與茶鹼之藥 物交互作用
The induction of drug-metabolizing
enzymes and interaction with theophylline by Sophora flavescens
研 究 生:蔡青芩 撰 指導教授:翁芸芳 博士
中華民國九十六年六月
致謝
首先誠摯的感謝指導教授翁芸芳老師於研究上給予我教導,讓我
能一步步朝向研究目標邁進,也不時指點我正確的方向,使我在這一年 多來獲益匪淺,其中要感謝口試委員鄭靜枝老師及廖尉岑老師於百忙中 審閱我的論文。
在近兩年裡的日子,從頭回想起來,一路上有太多人幫助過我,感 謝文驥、敏蕙於實驗上的協助,實驗室的又任學弟及伊安學妹,當然也 不能忘記,此外也感謝惠嵐、玟伶、佩君、郁嫻無論在台東或台北都協 助我生活上的點點滴滴,沒有你們大家的幫忙及搞笑,我可能無法完成 此論文,所以你們的大恩大徳我會銘感在心。
最後,我的親愛家人,是你們提供了我一個不需擔憂生活的環境,
讓我能安心學習,還有我表哥提供我住宿的地方,使我不用露宿街頭。
縮寫字全名對照
AST aspartate aminotransferase ALT alanine aminotransferase BUN blood urea nitrogen CRE creatinine
EROD 7-ethoxyresorufin O-deethylation GST glutathione S-transferase
NQO NAD(P)H-quinone oxidoreductase
NDM N-nitrosodimethylamine N-demethylation NFO nifedipine oxidation
PAH polycyclic aromatic hydrocarbons PROD 7-pentoxyresorufin O-dealkylation P450 cytochrome P450
TOH tolbutamide hydroxylation UGT UDP- glucuronosyltransferase
Abstract
The root of Sophora flavescens (Sf) has been used as a herbal medicine for the treatment of diarrhea, gastrointestinal hemorrhage, and eczema.
Modulation of drug-metabolizing enzymes plays a main role in herb-drug interaction. To assess the possible herb-drug interaction, effects of Sf on drug-metabolizing enzymes including cytochrome P450-dependent monooxygenase, UDP- glucuronosyltransferase (UGT), glutathione
S-transferase (GST), and NAD(P)H-quinone oxidoreductase (NQO) were investigated in mice and rats. In mice, administration of 3 g/kg/day powdered concentrate through agastrogavage for three days resulted in 91%, 281%, 87
%, 52%, and 88% increases of hepatic 7-ethoxyresorufin O-deethylation (EROD), 7-pentoxyresorufin O-dealkylation (PROD), coumarin
hydroxylation, nifedipine oxidation (NFO), and tolbutamide hydroxylation (TOH) activities, respectively. However, N-nitrosodimethylamin
N-demethylation (NDM) activity was not affected by the powdered concentrate. Administration of 1 g/kg/day aqueous extract through agastrogavage for three days resulted in 122%, 221%, 58%, and 43%
increases of hepatic EROD, PROD, coumarin hydroxylation, and NFO activities, respectively. In contrast, microsomal NDM and TOH activities were not affected. Administration of 1 g/kg/day with methanol extract through agastrogavage for three days resulted in 75%, 192%, and 60% increases of hepatic EROD, PROD, and coumarin hydroxylation activities, respectively.
However, microsomal NFO, NDM, and TOH activities were not affected.
Cytosolic GST activity was elevated by Sf all preparation. Methanol extract increased of NQO activity, whereas powdered concentrate and aqueous extract did not. In contrast, hepatic UGT and reanal GST and NQO activities were not affected. Immunoblot analysis of microsomal and cytosolic proteins revealed that Sf powdered concentrate respectively caused 2-fold, 3-fold, 24%, and 28% increases of CYP1A, CYP2B, CYP3A, and GST protein levels. The aqueous extracts respectively caused 12%, 2-fold, 37%, and 47% increases of CYP1A, CYP2B, CYP3A, and GST protein levels. The methanol extract
respectively caused 41%, 2-fold, and 28% increases CYP1A, CYP2B, and GST protein levels. In rats, administration of 3 g/kg/day powdered
concentrate through agastrogavage for three days resulted in 131%, 110%, and 96% increases of hepatic EROD, PROD, and NFO activities,
respectively. In contrast, microsomal NDM and TOH activities were not affected. Cytosolic GST activity was elevated by powdered concentrate.
Immunoblot analysis of microsomal and cytosolic proteins revealed that Sf powdered concentrate respectively caused 35%, 4-fold, 97%, and 43%
increases of CYP1A, CYP2B, CYP3A, and GST protein levels. Pre-treatment of rats with 3g/kg powdered concentrate caused a 31% increase in
theophylline clearance and a 23% and 36% decrease in AUC and AUMC, respectively. These results revealed that powdered concentrate, aqueous- and methanol- extracts of Sf could modulate drug-metabolizing enzymes but showed differential effects on individual enzymes. Attention should be paid to possible drug interaction caused by Sf when patients pretreated with herbal prescription containing Sf.
Keyword: Sophora flavescens; Cytochrome P450; glutathione S-transferase;
liver; kidney
摘要
苦參在中國傳統醫學上主要的功用為清熱燥濕、利尿和抗菌等功 能,可用於治療腸胃道疾病及濕疹等。藥物代謝作用之改變為藥物交互 作用之主要因素之一,為了評估苦參與其他藥物間可能產生之藥物交互 作用,因此研究苦參對鼷鼠及大白鼠肝臟及腎臟中藥物代謝酶,包括:
cytochrome P450 (P450)、UDP- glucuronosyltransferase (UGT)、glutathione S-transferase (GST) 及 NAD(P)H-Quinone oxidoreductase (NQO)之影響。
將鼷鼠連續三天餵食苦參科學中藥3 克/公斤/天,鼠肝 7-ethoxyresorufin O-deethylation (EROD)、7-pentoxyresorufin O-dealkylation (PROD)、
coumarin hydroxylation、tolbutamide hydroxylation (TOH) 及 nifedipine oxidation (NFO)活性分別增加 91%、281%、87%、52% 及 88%,而對 N-nitrosodimethylamin N-demethylation (NDM) 活性無影響。以 1 克/公斤 /天水萃取物處理鼷鼠,連續處理三天,使肝 EROD、PROD、coumarin hydroxylation 及 NFO 活性分別增加 122%、221%、58% 及 43%,但對 TOH 和 NDM 活性無影響。以 1 克/公斤/天甲醇萃取物處理鼷鼠,連續處 理三天,使鼠肝EROD、PROD 及 coumarin hydroxylation 活性分別增加 75%、192% 及 60%,但對 TOH、NDM 及 NFO 活性無影響。科學中藥
及萃取物皆對肝臟及腎臟UGT 活性均無影響;但科學中藥、水萃取物及
甲醇萃取物均可造成肝GST 的活性增加;經甲醇萃取物處理後會增加肝
NQO 活性。以免疫轉印法分析肝臟中的 P450 及 GST 蛋白含量之變化,
結果顯示苦參科學中藥會使CYP1A、CYP2B、CYP3A 及 GST 蛋白含量 分別造成2 倍、3 倍、24%及 28%的增加,水萃取物使 CYP1A、CYP2B、
CYP3A 及 GST 之蛋白含量分別造成 12%、2 倍、37%及 47%的增加,而 甲醇萃取物會使CYP1A、CYP2B 及 GST 的蛋白表現量分別造成 41%、2
倍及28%增加。此外,在大白鼠實驗中,連續三天餵食苦參科學中藥 3 克
/公斤/天,鼠肝 EROD、PROD 及 NFO 活性分別增加 131%、110%及 96
%,而對TOH 及 NDM 活性無影響。而 coumarin hydroxylation 則是測不
到。科學中藥可造成肝GST 的活性增加。在免疫轉印法分析肝臟中的 P450
及GST 蛋白含量之變化,結果顯示苦參科學中藥會使 CYP1A、CYP2B、
CYP3A 及 GST 蛋白含量分別造成 35%、4 倍、97%及 43%的增加。在苦
參科學中藥對茶鹼之交互作用,經科學中藥餵食後能降低茶鹼的AUC 及
AUMC 且增加清除率。綜合上述實驗結果顯示出苦參會增加肝臟藥物代 謝酶之作用,所以對病人在用藥治療時,應注意可能產生之藥物交互作 用的情形。
關鍵詞:苦參、細胞色素P450、穀胱甘肽-硫-轉移酵素、肝、腎
目錄
緒論………1
實驗材料與方法 壹、實驗材料……….6
貳、實驗方法……… 8
實驗結果………..……….16
結論………...22
討論…………..……….24
參考文獻…………...………28
圖表………...………35
圖目錄
Scheme 1.The pictures of kushen………..…………..35 Figure 1. The HPLC chromatograms of matrin in powdered concentrate and methanol extract of kushen………..36 Figure 2. The HPLC chromatograms of sophoraflavanone G and isokurarinone in
powdered concentrate and methanol extract of kushen………37 Figure 3. Dose-respone of effects of powdered concentrate of S. flavenscens on
7-ethoxyresorufin O-deethylation and 7-pentoxyresorufin O-dealkylation activities in mouse liver………..……….39 Figure 4. Time-course of effects of S. flavescens powdered concentrate on
7-ethoxyresorufin O-deethylation and 7-pentoxyresorufin O-dealkyation activities in mouse liver………...40 Figure 5. Immunoblot analysis of liver microsomal cytochrome P450 and cytosol protein levels from control(C) and Sophora flavanscens powdered concentrate(PC)-treated mice……….………..……….………..41 Figure 6. Dose-respone of effects of aqueous extract of S. flavenscens on
7-ethoxyresorufin O-deethylation and 7-pentoxyresorufin O-dealkylation activities in mouse liver……….………..42 Figure 7. Immunoblot analysis of liver microsomal cytochrome P450 and cytosol protein levels from control(C) and S. flavanscens aqueous extract(AE)-treated
mice………..43 Figure 8. Dose-respone of effects of methanol extract of S. flavenscens on
7-ethoxyresorufin O-deethylation and 7-pentoxyresorufin O-dealkylation activities in mouse liver……..……….44 Figure 9. Immunoblot analysis of liver microsomal cytochrome P450 and cytosol protein levels from control(C) and S. flavanscens methanol extract(ME)-treated
mice………..45 Figure 10. Dose-respone of effects of powdered concentrate of S. flavenscens on
7-ethoxyresorufin O-deethylation and 7-pentoxyresorufin O-dealkylation activities in rat liver…….…………..………..………..46 Figure 11. Immunoblot analysis of liver microsomal cytochrome P450 and cytosol protein
levels from control(C) and Sophora flavanscens powdered concentrate(PC)-treated rat……….…...47 Figure 12. The linear relationship between theophylline concentration and the peak area obtained from HPLC analysis………...………...48 Figure 13. Mean unbound theophylline concentrate-time curves in rat blood after
intravenous treatment with 2mg/kg theophylline. (A)Control rat, the solid lines show the result of linear regression; (B) Effects of pre-treatment with S. flavanscens on unbound theophylline concentration. Rats were pre-treated with water (○) or S. flavanscens
powdered concentrate(●) as descried Table 6………..…………...………...49
表目錄
Table 1. Effects of powdered concentrate of S. flavescens on the component conterts and
activites of cytochrome P450-dependent monooxygenase in mouse liver………...50
Table 2. Effects of S. flavescens extracts on cytochrome P450-dependent monooxygenase components and activities in mouse liver……….51
Table 3. Effects of the powdered concentrate and extracts of S. flavescens on UDP-glucuronosyl transferase, glutathione S-transferase, and NAD(P)H-quinone oxidoreductase activities in mouse liver………..52
Table 4. Effects of the powdered concentrate and extracts of S. flavescens on drug-metabolizing enzymes in mouse kidney………..53
Table 5. Effects of powdered concentrate of S. flavescens on the component conterts and activites of cytochrome P450-dependent monooxygenase in rat liver……….54
Table 6. Effects of powdered concentrate of S. flavescens on NAD(P)H-Quinone oxidoreductase, UDP-glucuronosyl transferase, and glutathione S-transferase activites in rat liver……….55
Table 7. Effects of powdered concentrate of S. flavescens on drug-metabolizing enzymes in rat kidney………...56
Table 8. Effects of powdered concentrate of S. flavescens on drug-metabolizing enzymes in rat lung ………...57
Table 9. Effects of powdered concentrate of S. flavescens on serum enzyme……….58
Table 10. Intraday precision and accuracy of theophylline (n=3)………..….59
Table 11. Interday precision and accuracy of theophylline (n=3) ………..….59
Table 12. Pharmacokinetic parameters of interaction between theophylline(2 mg/kg, i.v.) and S. flavescens powdered concentrate in rat blood………...…60
Table 13. Summary of in vivo effects of S. flavescens powdered concentrate, aqueous extract, methanol extract on drug-metabolizing enzymes activities in mouse liver……….61
緒論
在我們日常生活中,都會遇到藥物、天然物、環境汙染物等不同外來化學物質入 侵我們體內,肝臟為代謝外來物最主要的場所,其他組織如腎、肺及小腸等,也都有 進行藥物代謝作用的功能。這些代謝作用在外來化學物質之解毒與毒性活化作用中扮 演重要角色。一般而言,對於異質物會經由藥物代謝酶來代謝成無毒或活性較低的物 質,但也有異質物被藥物代謝酶活化成藥理活性更高的物質,例如:losartan藉由 CYP2C9 和CYP3A4 被氧化成活化型carboxylic acid代謝物,其具藥理活性較低之物質
(Yasar et al., 2001);或轉變成具有毒性或致癌的物質,例如:safrole會經由CYP2C9 及CYP2E1 代謝成具致癌前驅物 1,-hydroxysafrole(Ueng et al., 2004)。此外有少數藥 物(前趨藥物)會在體內轉化為有活性的藥物而起作用,如:cyclophosphamide。
在肝臟中,由酵素參與的代謝作用可分為二階段(Phase I、 Phase II)。Phase I 代 謝主要功能是對於外來物進行氧化(oxidation)、還原(reduction)以及水解(hydrolysis)
等反應來修飾外來物的官能基(functional group)使之攜帶-OH、-COOH、-SH、-O- 和-NH2來增加異質物的水溶性;Phase II則對代謝物進行結合反應(conjugation),來 增加代謝物的親水性。
Phase I 代謝大部份是經由細胞色素 P450(cytochrome P450;簡稱:P450)。由於 此細胞色素處於還原態時,會跟一氧化碳(CO)結合後在光譜 450nm 處附近會有最 大吸光值,所以利用此特性,故命名為細胞色素P450(Omura and Sato, 1964)。在P450 組成的單氧系統中,主要藉由氧化、還原、及水解反應來進行代謝異質物
(xenobiotics),而這些異質物包括藥物、環境汙染、類固醇、化學致癌物……等等,
經由此酵素系統可將異質物變成水溶性較高之代謝產物,以利降低異質物的毒性,但 也有可能因此反而產生高毒性之代謝物。
P450 組成單氧系統中主要存在於細胞上的內質網,目前存在於哺乳類中,至少 有17 個不同 P450 基因族(gene families),大概約有50~60 的同功酵素(Nelson, 1999)。 P450 主要命名的方式以 P450 胺基酸序列的相似度來區別,命名的系統是將
cytochrome P450 以 CYP 作為字頭,若胺基酸序列相似度超過 40%則歸為同一族
(family),以阿拉伯數字 1,2,3,………分類之;在同一族中,若胺基酸序列相似度介 於40~65%,則再分為出一個亞族(subfamily),以英文字母 A, B, C………分類;若 最後在同一個亞族中,每個gene product 為一個成員,其胺基酸序列相似度超過 65%,
以阿拉伯數字來命名。例如:CYP1A1(Nelson et al., 1996)。而主要負責藥物代謝的 是CYP1、CYP2、CYP3。
P450 因具有廣泛的受質特異性(substrate specificity),所以藉由外來化學物質、
內生性荷爾蒙、cytokines 或生長因子來誘導 P450 基因的表現再經由基因轉錄
(transcription)和轉譯(translation)的步驟來產生(Waxman, 1999)。所以我們利用 此特性作為化學指標,來探討P450 同功酶活性的變化。例如:7-ethoxyresorufin O-deethylation、coumarin hydroxylation、7-pentoxyresorufinO-dealkylation、tolbutamide hydroxylation、N-nitrosodimethylamine N-demethylation、nifedipine oxidation 分別來偵 測 CYP1A、CYP2A、CYP2B、CYP2C9、CYP2E 及 CYP3A 的酵素活性。P450 單氧 酶系統除了在肝臟中為主要代謝外來物外,其他器官如:腎、肺和小腸等也都會有單 氧酶系統存在出現。
P450 基因可藉由許多物質(例如:藥物、環境、化學物質或其他異質物等)來 調控誘導出來,此調控對於藥物或化學物質的藥效或毒性有很大的影響。大部分誘導 是經由 receptor-dependent pathway 刺激 CYP 基因而表現出來。其表現機轉是透過異 質物接受器(xenobiotic receptor)之一來誘導一連串作用使基因轉錄。例如:
benzo(α)pyrene(B(α)P)其平面結構為多環芳香族碳氫化合物(polycyclic aromatic hydrocarbons)進入細胞內會與 arylhydocarbon receptor(Ah receptor)結合後誘導 CYP1A 基因轉錄。當 phenobarbital(PB)進入細胞內, constitutive androstane receptor
(CAR)先會與 retinoid X receptor (RXR) 結合成異質二聚體(heterodimer),然後 作用在PB response element 上,而活化 CYP2B 基因轉錄作用(Kakizaki et al., 2003);
在CYP3A 方面,許多異質物(藥物、類固醇和 dexamethasone 等)與 pregnane X receptor
(PXR)結合,來誘導 CYP3A 基因而轉譯為 CYP3A 蛋白表現,有人研究指出從可 治療高血脂的 Commiphora mukul 中萃取出來 gugulipid 和 gugulsterone 皆能在 cell-based assay 及肝臟細胞中活化 PXR,來誘導 CYP3A 蛋白的產生,所以應避免 herb-drug 交互作用(Brobst et al., 2004)。
Phase II主要的代謝酵素為尿苷二磷酸-葡萄糖醛酸轉移酵素(uridine
5,-diphosphate-glucuronosyl transferase,UGT)、穀胱甘肽-硫-轉移酵素(glutathione S-transferase,GST)和NAD(P)H-quinone oxidoreductase(NQO)。UGT和GST分別催 化由phase I產生的代謝物與glucuronic acid和glutathione形成新的官能基(functional group)之結合。而NQO主要是含有quinone成份還原成hydroquinones。
UGT主要是存在於肝細胞的內質網上,也存在腎、胃及小腸等。UDP-
glucuronosyltransferase的命名方式和P450 的命名一樣以UGT作為字頭,若胺基酸序列 相似度超過45%,則視為同一族群(family),以阿拉伯數字 1,2,3,………分類;若在 同ㄧ族群中,胺基酸序列相似度介於45~60%之間,則再分為出一個亞族(subfamily), 以英文字母A, B, C………分類;最後若在同一個亞族中,胺基酸序列相似度超過 60%,則視為同一個成員(gene number),以阿拉伯數字來命名。例如:UGT1A1
(Mackenzie et al., 2000)。UGT是體內對於內生性物及外來物進行葡萄糖醛酸化作用
(glucuronidation)。這些疏水性物包括異質物和內生性物質,例如:藥物、環境汙染 物、膽固醇、類固醇。在細胞中,大部分疏水性物質會先經過phase I作用,進而獲得 一些官能基,再藉由UGT催化,從UDP-glucuronic acid(UDPGA)得到glucuronic acid 結合上去,產生glucuronide代謝物,而完成葡萄糖醛酸化作用。此作用可以將疏水性 物質轉換成水溶性較高的物質進而排出體內,來加速其在腎臟隨著尿液排出體外,以 達到解毒的功能。但有些物質在葡萄糖醛酸化作用之後,產生的代謝物反而會造成毒 理或藥理作用的活化,例如:NSAIDs(non-steroid anti-inflammatory drugs)的葡萄糖 醛酸化代謝物會與host protein作用,造成免疫過敏反應或其他副作用(Kretz-Rommel and Boelsterli, 1993)。UGT亦受到異質物和內生性物質的影響而會有抑制或誘導的現 象,例如:triiodothyronine和 3-methylcholanthrene會誘導UGT的基因進行transcription 而增加其活性(Masmoudi et al., 1997)。而testosterone和ethynyl estradiol會抑制 zidovudine的葡萄糖醛酸化作用,藥物中的naloxone、diazepam和flunitrazepam會抑制 morphine的結合作用,而重金屬中Fe3+, Cr2+, Ni2+, Al3+則會抑制morphine-3- 和 morphine-6-glucuronide的生成(Grancharov et al., 2001)。雖然UGT主要分佈在肝臟 中,但最近幾年有研究指出不管內生性物質或外生性物質,UGT在腎臟中也是扮演一 個重要角色,例如:不飽和脂肪酸(如:arachidonic acid、linoleic acid)及mefenamic acid(為非類固醇消炎止痛藥)會抑制腎臟中 4-methylumbelliferone(4-MU)的葡萄 糖醛酸化作用(Tsoutsikos et al., 2004; Gaganis et al., 2007)。
GST 為一種結合酶,主要是催化 phaseI 產生的物質與 glutathion(GSH)的 SH-group 進行結合反應來提高這些物質的水溶性,使之容易從尿液中排出體外。GST 主要分為三類,為cytosolic、mitochondrial、microsomal GST(Hayes et al., 2005),其 中cytosolic GST 主要代謝藥物及其他異質物;而 microsomal GST 則是代謝內生性物 質。在器官上主要分布在肝臟外,還分布於腎、肺、心及腦中。而cytosolic GST 主
要分為七類(class),分別是Alpha、Mu、Omega、Pi、Sigma、Theta 及 Zeta (Z)(McIlwain et al., 2006)。GST 表現會依照組織和細胞種類而有所不同,例如:hGSTM3 存在於 睪丸中,hGSTM2 則在腦部表現較多;Mu 類 GST 存在於眼球的晶體;Theta 類 GST 主要存在於肝和腎,而有少數存在於肺部裡;肝臟中含有較高的Alpha 類;在 Pi 類 中,proliferating cells 比在 fully differentiated cells 多(van Bladeren, 2000)。現有已 知有許多物質為GST 的受質,有些受質具有很高的選擇性,可做為測定 GST 的酵素 活性或比較不同物種間的GST 活性。但有些受質不具選擇性,例如:
1-chloro-2,4-dinitrobenzene 可偵測總 GST 的活性(van Bladeren, 2000)。
NQO 被當作成 DT-diaphorase 是一種 cytosolic flavoprotein,裡面含有 FAD。在人 體內扮演防止細胞受到氧化傷害的角色,能夠把phaseI 產生的物質含有 quinone 成份 藉由二個電子和二個氫原子還原成hydroquinones 變成沒有毒性的物質。
在藥物代謝中是以肝臟為最主要場所之一,但在1990 年爆發馬兜鈴酸腎病變,
指出中草藥如:關木、廣防己、天仙藤、馬兜鈴等,含有馬兜鈴酸(aristolochic acid)
會造成腎臟病變。Schmeiser 曾在病患腎臟檢體中檢測出 aristolochic acid-DNA 鍵結物
(Schmeiser et al., 1996)。所以腎臟在藥物代謝中也是佔有一席之地。Krajk-Kuzniak
(2005)等人提出原兒茶酸(protocatechuic acid)在肝跟腎中能夠降低 EROD 及 MROD 活性且提高GST 的活性,然而在肝臟中能抑制 NQO 但在腎臟中並不受影響,所以異 質物經由活化或解毒作用途徑,將修飾CYP、GST 及 NQO 來改變代謝活性,其結果 視組織而定。此外,市售葡萄柚汁(grapefruit juice)能與鈣離子阻斷劑(Calcium Channel Blockers)如:felodipine 產生交互作用,是因葡萄柚柚汁抑制小腸中的 CYP3A4 而阻 礙藥物代謝,但對於肝臟中CYP3A4 的影響力較小(Dahan et al., 2004),所以用葡萄 柚汁配藥吃時,藥物則無法正常代謝掉,會增加血液中的藥物濃度,因此容易造成藥 物之副作用的產生。
由於人們日漸使用中草藥普及,天然物對藥物代謝酶之影響產生的藥物之間 交互作用的問題也日漸備受矚目。在文獻當中也有陸續對中草藥萃取物影響單 氧酶系統及其他藥物代謝酶的報導,St. John,s wort萃取物可誘導CYP2D2 和 CYP3A2,但會抑制CYP 2C6(Dostalek et al., 2005)。kava-kava(Piper methysticum)
成分中pipermethystine能增加superoxide dismutase (Cu/Zn SOD)及誘導CYP2E1和 CYP1A2基因表現(Lim et al., 2007)。Rutaecarpine是吳茱萸(Evodia rutaecarpa)中 主要成分成分之一,可誘導CYP1A(Ueng et al., 2001)。
苦參(Sophora flavescens),又名苦骨、地槐、水槐、菟槐、驕槐、野槐、白莖,
為豆科植物苦參的根。苦參性味苦,寒。歸膀胱、心、肝、大腸、小腸經。在中國傳
統醫學上主要的功用為清熱燥濕、利尿和抗菌等功能。自古以來即是常見藥方,《本
草綱目》曰:「治腸風瀉血,並熱痢」。《神農本草經 》曰:「主心腹氣結,癥假積聚,
黃疸,溺有餘瀝,逐水,除癰腫」。《名醫別錄 》曰:「除伏熱腸辟,止渴,醒酒,小
便黃赤,療惡瘡,下部匿」。其功能與黃芩、黃連及龍膽皆相近。苦參中包含許多成
分,例如 alkaloids、 flavonoids 及 isoflavones 等,其中 alkaloids 含最多的成分是 matrine。 對於苦參的藥理活性,有許多相關的研究文獻報導。有人指出從 S. flavescens 甲醇萃取出來的黃酮類能有效抑制醣解酵素(glycosidase)(Kim et al, 2006),利用 S. flavescens 萃取物能刺激 C57BL/6J 鼷鼠毛髮生長,在人類毛乳頭細胞中增加 IGF-1 和KGF 的 mRNA 表現量(Roh et al, 2002)。Oxymatrine 是 matrine 其中一種的成分,
能夠抗發炎抑制發炎反應,可藉由抑制phosphorylated p38 mitogen-activated protein kinase 減少老鼠急性肺損害(Xu et al, 2005)。Lavandulylflavonoids 能抑制 HL-60 細 胞和HepG2 細胞增生和誘導其細胞凋亡,所以 S. flavescens 可能可被使用在癌症化學 療法及化學預防的試劑(Ko et al., 2000)。但苦參對藥物代謝酶的影響尚未被探討。
因此,本篇論文的研究目的如下:
一、研究苦參科學中藥對CYP 的調控作用
中藥的使用方式通常是以口服為主,因此本研究探討口服苦參科學中藥對 C57BL/6J 鼷鼠和 Sprague-Dawley 大鼠之肝、腎藥物代謝酶之影響。測定鼷鼠 和大鼠經餵食苦參科學中藥後之單氧酶組成分子含量及 CYP 活性之變化,並 分析苦參對 UGT、GST 及 NQO 活性之影響。再經由免疫轉印方法分析 CYP 及 GST 的變化情形。
二、研究苦參萃取物對CYP 的調控作用
為了分析苦參中藥物代謝酶誘導物在水萃取物與甲醇萃取物中的分佈,因此本 研究探討口服苦參萃取物對C57BL/6J 鼷鼠之藥物代謝酶之影響。測定鼷鼠經 餵食苦參萃取物後之單氧酶組成分子含量及CYP 活性之變化,並分析苦參對 UGT、GST 及 NQO 活性之影響。再經由免疫轉印方法分析 CYP 及 GST 的變 化情形。
三、研究苦參科學中藥對theophylline 之藥物動力學參數的影響。
測定苦參科學中藥對 theophylline 產生之藥物作用,以評估藥物交互作用。
實驗材料與方法
壹、實驗材料
一、實驗動物
1.雄性 C57BL/6J 鼷鼠(五週齡,體重 16~20 g)購自行政院國家科學委員會國家 動物研究及繁殖中心。
2.雄性 Sprague-Dawley 大鼠(五週齡,體重 180~200 g)購自陽明大學實驗動物 中心及樂斯科生物科技公司。
二、儀器設備
1.高效率液相層析儀(HPLC)
Injector:G1328A(Aglient, Ltd., U.S.A)
Autosample:L-7200(Hitachi, Ltd., Tokyo, Japan)
Pump: L-7100(Hitachi, Ltd., Tokyo, Japan), G1311A(Aglient, Ltd., U.S.A)
UV-visible detector:L-7420(Hitachi, Ltd., Tokyo, Japan), G1315A(Aglient, Ltd., U.S.A)
Fluorescence detector:L-7480(Hitachi, Ltd., Tokyo, Japan)
2.實驗儀器
螢光光譜儀:F4500(Hitachi, Ltd., Tokyo, Japan)
紫外光-可視光光譜儀:U3000(Hitachi, Ltd., Tokyo, Japan)
震盪式恆溫培養水浴槽:Model 903(Hotech, Taiwan), B603D(Firstek Scientific, Taiwan)
細胞研磨機:RW20.n(IKA Labortechnik, Malaysia)
微量離心管離心機:RCM(Sorvall, Inc., U.S.A.)
離心機:TC6(Sorvall, Inc., U.S.A.)
高速離心機:J2-21(Beckman, Inc., California, U.S.A.)
超高速離心機:LE-80, LE-90K(Beckman, Inc., California, U.S.A.)
自動洗片機:X-OMAT M43(Kodak, Japan)
電泳槽:Wide Mini Sub® Cell CT(Bio-Rad Laboratories, Richmond, CA., U.S.A.), HE33-8-1.5(Amersham Pharmacia Biotech, Ltd., U.K.)
電泳電源供應器:PAC300(Bio-Rad Laboratories, Richmond, CA., U.S.A.),
PS500XT(Amersham Pharmacia Biotech, Ltd., U.K.)
血液生化值測試儀:Fuji Dri-Chem 3000(Fuji Photo Film Co., Ltd, Tokyo, Japan)
三、藥品與試劑
1.苦參科學中藥購自順天堂中藥。
2.苦參甲醇萃取物及苦參水萃取物皆由中國醫藥研究所陳建志老師所提供。
3.下列藥品購自 Sigma Chemical Co., St. Louis, M.O., U.S.A:
Glucose-6-phosphate(G6P), NADP, NADH, NADPH, glucose-6-phosphate dehydrogenase(G6PDHase), bovine serum albumin(BSA), cytochrome c, 7-ethoxyresorufin, 7-pentoxyresolufin, N-nitrosodimethylamin, nifedipine, Triton X-100, HEPES, ethylenediaminetetraacetic acid(EDTA), 7-hydroxycoumarin, coumarin, sodium potassium tarteate, tolbutamide, 1-chloro-2,4-dinitrobenzene
(CDNB), N-nitrosodimethylamine, glutathione, UDP-glucuronic acid , p-nitrophenol
4.下列藥品購自 Merck Taiwan Ltd., Taipei, Taiwan:
Acetic acid, acetone, acetonitrile, chloroform, ethanol, methanol, Tween 20, dichlormethan, ortho-phosphorsanre
5.下列藥品購自 J.T. Baker Inc., Phillipsburg N.J., U.S.A:
Potassium phosphate, potassium chloride, maganesium chloride 6-hydrate, sodium phosphate, sodium chloride, trichloroacetic acid
6.下列藥品購自 Bio-Rad Laboratories, Richmond, C.A., U.S.A.:
Acrylamide, N-N,-methylene-bis-acrylamide, TEMED, sodium dodecyl sulfate
(SDS), ammonium persulfate, β-mercaptoethanol, bromophenol blue, blotting grade blocker non-fat dry milk及 glycine
7.單株抗體 Mab 1-7-1(anti-rat CYP1A), 2-13-1(anti-rat CYP3A)獲自 Dr. S. S.
Park(Seoul National University, Korea);goat anti-rat CYP2B 多株抗體購自 Daiichi Pure Chemical Co., Ltd., Tokyo, Japan;goat anti-rat GSTYb 購自 Oxford Biomedical Research, Oxford, MI, USA;rabbit anti-mouse IgG 購自 Pierce Chemical Co., Rockford, IL, U.S.A.; rabbit anti-goat IgG 購自 ICN
pharmaceutical, Inc., Aurora, Ohio, USA
貳、實驗方法
一、苦參萃取物之製備
1.苦參水萃取物(aqueous extract)之製備
將苦參藥材600 克加 4L 的水煮沸萃取,之後過濾,將濾液冷凍乾燥濃縮,可 得苦參水萃取物重116 克。
2.苦參甲醇萃取物(methanol extract)之製備
將苦參1200 克經粉碎後加 8 公升的甲醇,迴流加熱二次,把溶液過濾所得濾 液經濃縮冷凍乾燥,可得苦參甲醇萃取物重224 克。
二、動物處理
1. C57BL/6J 鼷鼠(五週齡,體重 16~20g)於國立中國醫藥研究所動物室,並 給予正常的飼料與水源,有自動控制系統控制12 小時光照/12 小時黑暗,室 溫在20±2。先適應環境一週後,將苦參科學中藥(powdered concentrate)用 研磨器研磨過後溶於水中,依實驗需要分別依不同劑量或不同處理時程來處 理老鼠,用餵食管餵食老鼠,在最後一次處理24 小時後將動物犧牲,犧牲後 取出肝和腎以製備微粒體或細胞質液,建立劑量與酵素活性以及處理時程與 酵素活性之關係圖。而水萃取物及甲醇萃取物分別用研磨器研磨並溶於水 中,依實驗需要分別餵食0.1、0.5、0.75、1、3g/kg,連續餵食三天,將動物 犧牲。
2. Sprague-Dawley 大鼠(五週齡,體重 180~200g)於國立中國醫藥研究所動物 室給予正常的飼料與水源,並有自動控制系統控制12 小時光照/12 小時黑 暗。在適應環境一週後,將苦參科學中藥(powdered concentrate)用研磨器 研磨過後溶於水中,用餵食管餵食老鼠,依實驗需要分別餵食1、3 g/kg,連 續餵食三天,在最後一次處理24 小時後將動物犧牲,犧牲後取出肝和腎以製 備微粒體或細胞質液,依不同劑量與酵素活性分別說明於結果及圖表中。
三、肝微粒體(microsome)及細胞質液(cytosol)之製備
微粒體和細胞質液製備的方法是採分段離心的方法(Alvares and Mannerung, 1970),且製備全程保持於冰浴中。鼷鼠經二氧化碳迷昏後,以斷頸方法犧牲,將鼠 肝和腎於1.15% KCl 中漂洗三次後,清除結締組織、血塊及脂肪,並以吸水紙吸去
多餘的水分,給予秤重。將組織剪碎,加入1.15%KCl 溶液於細胞研磨機(teflon pestle-glass homogenizer)中來回均質研磨六次,倒入高速離心管兩管兩管平衡後在 4℃以下以 9,000×g 離心 20 分鐘後,取得含微粒體沉澱及細胞質液之上清液,將上清 液移置於超高速離心管中兩兩平衡,在4℃以下以 10,000×g 離心 1 小時後,取得微粒 體沉澱及細胞質上清液。收集細胞質液,將剩下微粒體沉澱加1.15% KCl,再均質 一次,倒回超高速離心管後兩兩平衡,在4℃以下以 10,000×g 離心 1 小時後,取得微 粒體(wash microsome),上面覆以 0.1 M 磷酸鉀緩衝溶液(pH 7.4),保存於-75℃冷 凍冰箱,並於二周內完成藥物代謝酶活性測定。
四、蛋白質含量之定量
微粒體及細胞質液蛋白含量之測定,沿用Lowry(1951)等人之方法,以 0.1、
0.2、0.3 及 0.4 mg/ml之牛血清蛋白(BSA)做為標準品,標準品與經水稀釋之微粒體 或細胞質液均加入 2 ml copper溶液(2% Na2CO3/0.1N NaOH:1% CuSO4‧5H2O:
2% Na-K Tartrate=100:1:1),混合均勻置室溫 10 分鐘後,加入 0.2 ml 1N
Folin-Ciocalteu,s phenol 試劑,均勻混合後放置室溫 30 分鐘後,記錄在 750 nm之吸 光值,所有蛋白質含量之測定須在30 分鐘內測定完成。利用標準品濃度與其吸光值 作回歸直線來求得微粒體與細胞質液之蛋白質含量。
五、單氧酶組成分子含量或活性測定 1. P450 含量之測定
沿用 Omura 及 Sato(1964)之一氧化碳差異光譜法。製備 2 ml 約含 1.0~1.25 mg蛋白質之鼠肝及腎微粒體懸浮液,加入約為 1mg之硫代硫酸鈉(Na2S2O4) 予以還原,分別分裝 1 ml樣品至參考比色槽與樣品比色槽,將光譜儀自 350~550
nm作基準線校正,完成後將樣品比色槽取出,緩慢注入一氧化碳 20 秒,放回 樣品比色槽,掃瞄並紀錄波長450 nm附近最大吸收波長與波長 490 nm時之吸 光值差,其吸光係數91 mM-1cm-1,以計算CYP含量。
2.Cytochrome b5含量之測定
沿用 Omura 及 Sato(1964)之方法。將 1 ml 約含 1mg 微粒體蛋白量之鼠 肝微粒體懸浮液,置於參考比色槽與樣品比色槽,將光譜儀自 350~550 nm 作 基準線校正,完成後將樣品比色槽取出,加入 35.3 nM NADH,混合均勻後放
回光譜儀中,掃瞄並紀錄波長最大吸收波長與最小吸收波長時之吸光值差,其 吸光係數 185 mM-1cm-1,以計算cytochrome b5含量。
3.NADPH-P450 reductase 活性之測定
沿用Phillips及Langdon(1962)之方法,使用 cytochrome c當作受質。反 應總體積為 1.2 ml,其中含 0.02~0.04 mg鼠肝與腎之微粒體蛋白,0.6 M磷酸 鉀溶液(pH 7.7),0.2 mM EDTA,5.13 mg/ml cytochrome c,混合均勻後將光 譜儀自動歸零,再分別於參考與樣品比色槽中加入等量之二次去離子水與 0.1mM NADPH 溶液,混合後開始紀錄室溫下 2 分鐘內波長 550 nm之吸光值 變化,其吸光係數21 mM-1cm-1及單位時間內吸光值之變化計算還原cytochrome c之活性。
六、單氧酶活性之測定 1. EROD 之活性測定
沿用 Pohl 及 Fouts(1980)之方法,使用蛋白質含量 0.05~0.1 mg 之肝微 粒體懸浮液或使用 0.2~0.4 mg 腎微粒體懸浮液,反應總體積為 1 ml,反應溶 液含 0.13 M Hepes buffer(pH7.8),6.4 mM G6P,0.14 mM NADP,0.25 單位 G6PDHase,1.8μM 7-ethoxyresorufin。在 37℃恆溫水浴槽中避光反應 10 分鐘 後,加入2.5 ml methanol 終止反應,再以 560×g 離心 15 分鐘去除蛋白質沉澱 後,使用螢光光譜儀讀取上清液,以波長550 nm 為激發波長,585 nm 為放射 波長時之螢光強度,並以0.01μM 及 0.1μM Rhodamine B 的螢光強度來比較,
以求得此酶反應活性。
2. PROD 之活性測定
沿用Lubet(1985)之方法,使用蛋白質含量 0.3~0.6 mg之肝微粒體懸浮 液或使用0.2~0.4 mg腎微粒體懸浮液,反應總體積為 1 ml,反應溶液含 0.1 M MgCl2,0.05 M Tris buffer(pH7.5),6.4 mM G6P,0.14 mM NADP,0.2 單位 G6PDHase,10.6μM 7- pentoxyresorufin。在 37℃恆溫水浴槽中避光反應 10 分 鐘後,加入2.25 ml methanol終止反應,再以 560×g離心 15 分鐘去除蛋白質沉 澱後,使用螢光光譜儀讀取上清液,以波長522 nm為激發波長,586 nm為放 射波長時之螢光強度,並以0.01μM及 0.1μM Rhodamine B的螢光強度來比 較,以求得此酶反應活性。
3. Coumarin hydroxylation 之活性測定
沿用 Souček(1999)之方法,使用蛋白質含量 0.03~0.06 mg之肝微粒體 懸浮液,反應總體積為0.5 ml,反應溶液含 0.05 M磷酸鉀溶液(pH7.4),0.5 mM NADP,5.0 mM G6P,0.1 M MgCl2,0.02 mM coumarin,0.5 單位G6PDHase。
在 37℃恆溫水浴槽中反應 20 分鐘後,加入 0.3 M HClO4終止反應,移置離心 管以8,000×g離心 2 分鐘,離心後取出上清液等待分析。以HPLC的方法分析 coumarin的代謝產物,使用C18 column,以 20% acetonitrile/20 mM NaClO4
(pH2.5),流速 1 ml/min分析代謝產物,紀錄在紫外光波長 272 nm,螢光激 發光(Ex)338 nm和放射光(Em)458 nm波長時之積分面積,使用其 7-OH coumarin 代謝產物之標準品,以計算活性。
4. TOH 之活性測定
沿用 Yamazaki及 Shimada(1997a)與 Knodell(1987)等人之方法,使 用蛋白質含量0.0125 mg之肝微粒體懸浮液,反應總體積為 0.5 ml,反應溶液 含0.13 M磷酸鉀溶液(pH7.4),1.3 mM NADP,25 mM G6P,1 mM tolbutamide,
0.25 單位G6PDHase。在 37℃恆溫水浴槽中反應 15 分鐘後,加入 25μl 1N HCl 和 1.5 ml CH2Cl2終止反應,以560×g離心 5 分鐘,離心後取出 1 ml有機層置 於微量離心管中,以氮氣吹乾,等待分析。以HPLC的方法分析tolbutamide的 代謝產物,使用C18 column,以 36%(V/V) acetonitrile和 60%(V/V)0.04%
H3PO4,流速 1 ml/min分析代謝產物,紀錄在波長 230 nm時之積分面積,使用 其4-OH tolbutamide 代謝產物之標準品,以計算活性。
5. NDM 之活性測定
沿用 Greenlee及 Poland(1978)與 Nash(1953)之方法以呈色反應測定 N-nitrosodimethylamine N-demethylation產生甲醛的活性,使用蛋白質含量 0.8~1.2 mg之肝微粒體懸浮液,反應總體積為 1.25 ml,反應溶液含 0.07M KCl,0.4 mM NADP,12.3 mM G6P,0.5 單位G6PDHase。在 37℃恆溫水浴槽 中反應15 分鐘後,加入 1ml 2.2M ZnSO4,在 37℃恆溫水浴槽中震盪 5 分鐘 後,加入1ml 飽和的Ba(OH)2溶液,再在37℃恆溫水浴槽中震盪 5 分鐘,
將混濁液體以15,000×g離心 10 分鐘後,取出 1 ml加到 0.6 ml Nash試劑中,均 勻混和後在60℃恆溫水浴槽中反應 15 分鐘。反應後冷卻至室溫,使用光譜儀 讀取波長412 nm之吸光值,以formaldehyde 為標準品以計算活性。
6. NFO 之活性測定
沿用 Guengerich等人(1986)之方法,使用蛋白質含量 0.01 mg之肝微粒 體懸浮液,反應總體積為0.5 ml,反應溶液含 0.04 M磷酸鉀溶液(pH7.85),
0.54 mM NADP,0.01M G6P,12.3 mM MgCl2,0.25 單位G6PDHase及 0.5 mM nifedipine。在 37℃恆溫水浴槽中反應 15 分鐘後,加入 0.1 ml 1M Na2CO3/1M NaCl和 1 ml CH2Cl2終止反應,以200×g離心 5 分鐘,離心後取出 0.5 ml有機 層置於棕色微量離心管中,以氮氣吹乾,等待分析。以HPLC的方法分析 nifedipine的代謝產物,使用C18 column,以 55% methanol,流速 1 ml/min分 析代謝產物,紀錄在波長254 nm時之積分面積,使用其pyridine代謝產物之標 準品,以計算活性。
七、結合酶活性之測定 1. GST 活性之測定
沿用Habig(1981)等人之方法,以 1-chloro-2,4-dinitrobenzene為受質來進 行反應。反應溶液含之肝細胞質液蛋白,0.1 M CDNB及 1 M磷酸鉀緩衝溶液
(pH6.5)。在室溫下,將反應液以光譜儀自動歸零,再分別於參考與樣品比色 槽中加入等量之二次去離水與150 mM GSH混和後開始紀錄 2 分鐘內波長 340 nm之吸光值變化,以吸光係數為 9.6 mM-1cm-1,以計算GST活性。
2. NQO 活性之測定
沿用Liu(1989)等人之方法,以menadione為受質,在弱光中進行反應。
反應溶液含0.2 mM NADH,16 mM menadion及 100 mM磷酸鈉緩衝溶液
(pH7.4)。在室溫下,將反應液以光譜儀自動歸零,在樣品比色槽中加入 0.1~0.2 mg之肝細胞質液或 0.06~0.08 mg腎肝細胞質液蛋白混和後開始紀錄 2 分鐘內波長340 nm之吸光值變化,以吸光係數為 6.2 mM-1cm-1,以計算 NAD(P)H-quinone oxidoreductase活性。
3. UGT 活性之測定
沿用Bock(1983)等人之方法,以p-nitrophenol為受質,在弱光中進行反 應。使用蛋白含量0.1 mg之肝微粒體懸浮液或 0.25 mg腎微粒體懸浮液,反應 總體積為0.5 ml,反應溶液含 0.1 M Tris-HCl(pH7.4),5 mM MgCl2,0.01%(V/V) Triton X-100,3 mM UDP-glucuronic acid,0.6 mM p-nitrophenol。在 37℃水浴
槽中反應10 分鐘後,加入 1 ml 5%(W/V) trichloroacetic acid 以終止反應。將 此混合液轉置棕色微量離心管中,以1,500×g離心 15 分鐘已沉澱蛋白質。離心 後取上清液1ml加入 0.25 ml 2N NaOH中,混合均勻後,紀錄空白與樣品組在 波長405 nm的吸光值,得樣品組因結合反應而降低之吸光值,最後以吸光係 數18.1 mM-1cm-1計算活性。
八、免疫轉印分析(Immunoblot)
本實驗沿用 Laemmli(1970)之非連續性電泳(discontinuous electrophoresis)方 法,使用stacking gel 及 separation gel 來進行十二烷基硫酸納(sodium dodecyl sufate, SDS)-聚丙烯醯(polyacrylamide)膠體電泳。將 Tris-HCl 緩衝溶液、acrylamide、
Bis- acrylamide 和 SDS 均勻混和後,以抽氣幫浦去除液體中的氣體,再加入 10%
ammonium persulfate 和 TEMED 催化 acrylamide 的聚合反應。其中 7.5% separation gel 分離CYP2B 及 CYP3A,而 10%和 12% separation gel 則分別分離 CYP1A 及 GST。
樣品處理過程是將微粒體在含 1.25 M Tris-HCl 緩衝溶液(pH6.8)、20% SDS、
0.1% Bromophenol blue、β-Mercaptoethanol 和 glycerol 之樣品製備液於 100℃加熱 3 min 後冷卻進行電泳。電泳過程中,stacking gel 與 separation gel 作用分離時,分別用 15 mA 與 30 mA 之電泳強度。電泳完畢後,沿用 Towbin 等人(1979)的方法,以 transfer buffer(含 25 mM Tris base(pH 8.3, 192 mM glycine, 20%(v/v) methanol)將 nitrocellulose paper(NC)膜濕潤,再將膠體平舖於 NC 膜上,再覆蓋上以 transfer buffer 濕潤之濾 紙,最後置於濕式轉印槽置中,在4℃下以0.1 A 之電流進行轉印隔夜。
轉印完畢後,先將NC膜置於含5﹪脫脂奶粉之 phosphate buffer saline(PBS:含 0.14M NaCl,8.10M Na2HPO4,2.68M KCl,1.47M KH2PO4),於37℃水浴槽中反應 1 小時。反應完畢後,再與稀釋過後的一級抗體於4℃作用隔夜,之後以含 0.5% Tween 20 的PBS清洗四次,每次十五分鐘,以洗去非特異性結合之抗體。接著用稀釋過後的 二級抗體,於室溫下反應1 小時後,再以含 0.5% Tween 20 的PBS清洗四次,每次十 五分鐘。最後以ECL試劑反應 1 分鐘後,將多餘的ECL試劑移去,用NC膜包好,於X 光片上曝光,再放入自動洗片機洗片,最後以數位元影像處理系統 ImageMaster比較 其相對濃度。
九、苦參科學中藥在 Sprague-Dawley 大鼠體內之藥物動力學
使用大鼠為 7~8 週大之雄性 Sprague Dawley Rat,體重介於 290~330 克。實驗組 大鼠先利用胃管口服苦參科學中藥3 g/kg/day 連續餵食三天,第四天進行 theophylline 與科學中藥之交互作用,研究科學中藥對theophylline 藥物動力學之影響。用
pentobarital 麻醉之後,其實驗組及對照組大鼠以股靜脈注射 theophylline 劑量為 2 mg/kg 後 3、5、15、60、90、120、180、240、300、360 分鐘,以 1 ml, 26G 塑膠注 射針以心臟採血方式取血,每次取0.3 ml 置於超微量離心管,採取血液檢品後離心取 上層血清,置於-20℃冰箱儲存,待分析。
1.檢量線之製作
藥物檢量線樣品是利用空白的生物樣品加入以之濃度之藥物標準品來製 備。將配好的不同濃度之樣品注入高效液相層析系統中進行分析,測定藥物的訊 號。利用層析圖記錄藥物波峰的面積數值,將不同濃度所得之面積數值進行回歸 求得檢量線。
2.精確性試驗
為了確認 theophylline 定量分析方法之精確性,因此做同日內(Intraday)及 間日內(Interday)的精確性比較。
(A)同日內(Intraday)試驗
依不同濃度之 theophylline 標準濃度樣品,分別於同一日的早上和晚上 分別個取濃度為1、2 及 3 μg/ml 之 theophylline 標準液(n=3),計算各個校 正濃度之平均值(Mean)、標準偏差(S.D.)及變異係數(S.E.)。
(B)間日內(Interday)試驗
若於不同天以同日內(Intraday)試驗方法操作可得到間日內(Interday)
的精確性比較。
3.血清檢品處理方法
沿用Kim(2003)等人之方法,取 0.05 ml 血清加入 0.3 ml 100% acetonitrile 震盪,以1500×g 離心 10 分鐘,離心後取出 0.3 ml 上清液置於微量離心管中,
以氮氣吹乾,等待分析。以HPLC 的方法分析藥物血中經時間濃度變化,使用 C18 column,以 95%(V/V)10 mM sodium acetate(pH=5)和 5%(V/V) acetonitrile,流速 1 ml/min 分析藥物,紀錄在波長 280 nm 時之積分面積。
十、血清中 ALT、AST、CRE 及 BUN 之分析
使用血液測試組(Fuji Dri-Chem slide),加入10 μg 血清,使用血液中生化值測 試儀來讀取測試結果,測定血液中AST、ALT、CRE 及 BUN 之值。
十一、實驗數據之分析
實驗數據之分析採用 Student’s t-test 和 one-way ANOVA,當 P<0.05 則代表實驗 組與對照組之間,具有統計學上的顯著差異。在測定大鼠體內之藥物動力學的實驗部 份中,使用 WinNonlin 程式,以二室模式(two compartment model)計算 theophylline 在大鼠血液中的藥物動力學參數。
Two-compartment IV-Bolus, macro-constants, no lag time, l st order elimination
K12 K21
Bolus IV K10
1 2
C(t)=A exp(-αt)+B exp(-βt)
實驗結果
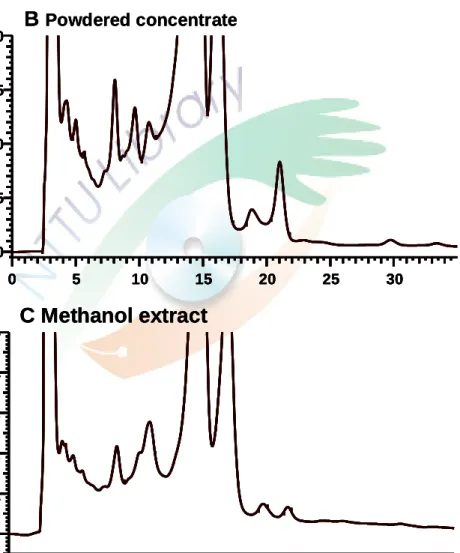
一、苦參科學中藥及其萃取物成分之含量測定
利用 HPLC 分析科學中藥、水萃取物及甲醇萃取物中 matrine、sophoraflavanone G 及isokurarinone 的含量,matrine 標準品的滯留時間(retention time)約為 8.5 分鐘,
Sophoraflavanone G 標準品的滯留時間約為 58 分鐘,isokurarinone 標準品的滯留時間 約為64 分鐘。經分析 HPLC 波峰下面積後結果顯示每克苦參科學中藥及甲醇萃取物 中分別約含有1.24 mg 和 4.64 mg 的 matrin;每克苦參科學中藥及甲醇萃取物中分別 約含有0.17 mg 和 13.6 mg 的 sophoraflavanone G;每克苦參科學中藥及甲醇萃取物中 分別約含有0.05 mg 和 3.12 mg 的 isokurarinone;而在水萃取物中,均未測到 matrine、
sophoraflavanone G 及 isokurarinone 的存在(Figure1、Figure 2)。
二、苦參科學中藥對C57BL/6J 鼷鼠之處理時程和劑量與活性關係
(一)苦參科學中藥之劑量與活性關係
進行苦參科學中藥之劑量與鼠肝 EROD 和 PROD 活性關係之實驗。將 C57BL/6J 鼷鼠分別餵食 0.1、0.5、1、3 g/kg 科學中藥,連續餵食三天後,
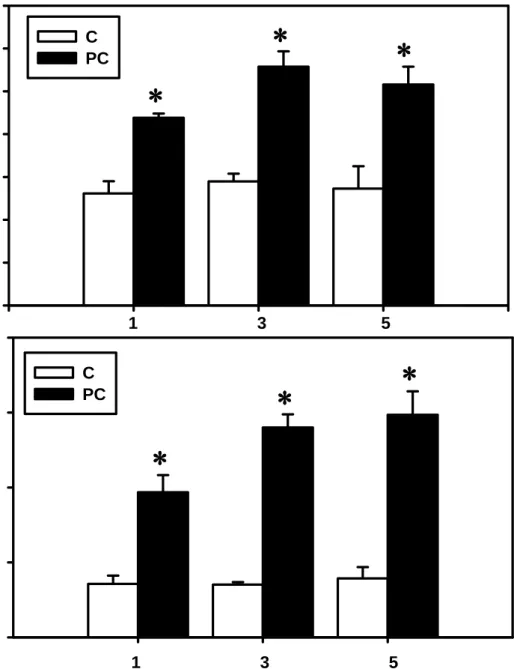
取鼠肝微粒體作EROD 和 PROD 活性測試。結果顯示,在餵食科學中藥 0.5、
1、3 g/kg 造成 EROD 活性分別增加 80%、2 倍、2 倍,經 1、3 g/kg 科學中 藥處理後,PROD 活性分別增加 2 倍、4 倍(Figure 3),而 3 g/kg/day 升高 倍數較高,故接下來的實驗以3 g/kg 劑量的苦參科學中藥來處理 C57BL/6J 鼷鼠,以建立處理時程與藥物代謝酶活性誘導之相關性。
(二)苦參科學中藥之處理時程與藥物代謝酶活性之誘導關係
將 C57BL/6J 鼷鼠餵食 3 g/kg/day 苦參科學中藥,分別餵食一、三、五天 後,取鼠肝微粒體作EROD 及 PROD 活性測試。結果顯示,不論在餵食一、
三、五天後,均可造成EROD 及 PROD 活性增加(Figure 4),其中 EROD 活 性分別造成68%、92%、89%的增加,PROD 活性分別增加 2 倍、3 倍、3 倍,
而其中三天和五天升高倍數相接近,所以接下來的實驗採取處理鼷鼠三天。
(三)苦參科學中藥對鼠的肝臟與腎臟藥物代謝酶活性之影響
為了瞭解科學中藥對肝臟及腎臟單氧酶活性和結合酵素之影響,以 3 g/kg/day科學中藥餵食鼷鼠,連續餵食三天後,取得鼠肝及腎之微粒體測定
單氧酶活性。結果顯示科學中藥處理組,在體重、肝重、微粒體蛋白含量及 cytochrome b5活性與對照組比並無明顯差異,其餘在CYP和NADPH-CYP reductase活性分別上升 42%和 16%。在肝臟單氧酶方面,使EROD、PROD、
coumarin hydroxylation、NFO及TOH的活性分別上升 91%、281%、87%、
52%及 88%; 但對於和NDM活性無影響(Table 1)。在結合酵素測定上,
Table 3 顯示GST的活性上升 55%,而UGT及NQO並無明顯的差異。
在腎臟方面,腎重、微粒體蛋白含量、NADPH-CYP reductase、NQO 及GST 均不受餵食科學中藥的影響與控制組無明顯的差異,其中 EROD、
PROD 及 UGT 均測不到(Table 4)。
綜合上述,可得知在肝臟中 NADPH-CYP reductase 及 GST 的活性皆有 增加,但在腎臟中,二者活性並沒有增加情形;而在NQO 活性測定中,肝 臟和腎臟皆無影響。
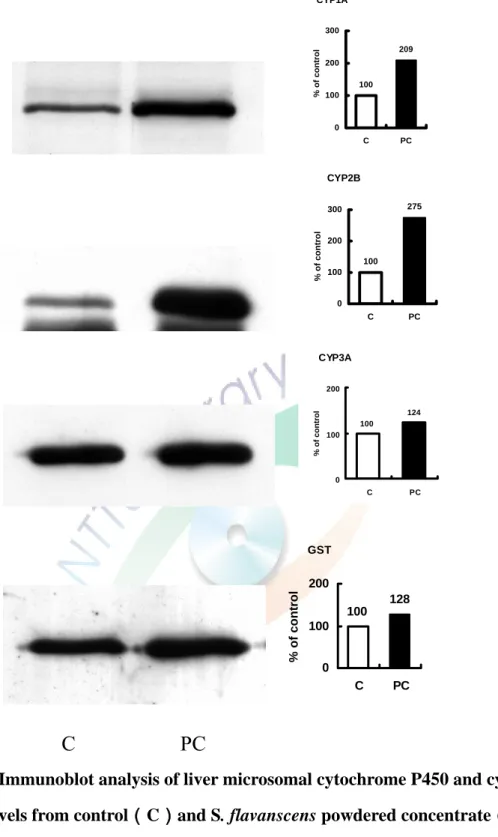
(四)苦參科學中藥對 CYP 蛋白含量的影響
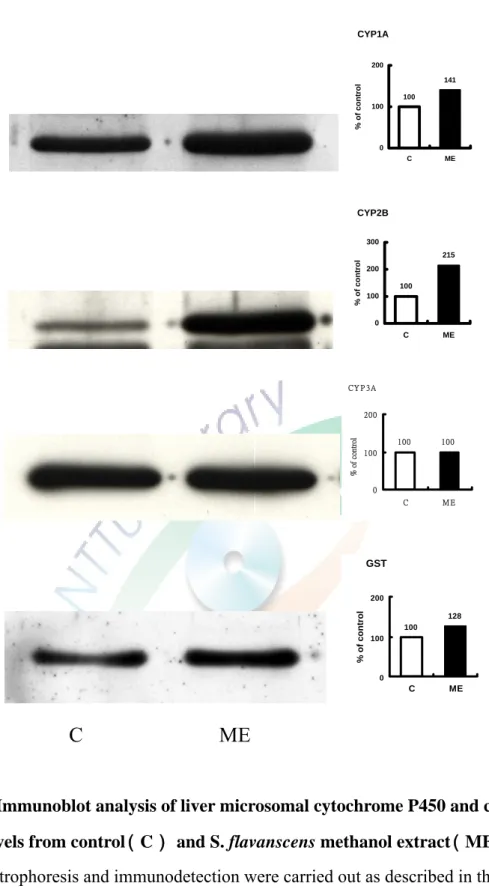
將 C57BL/6J 鼷鼠餵食 3 g/kg/day 苦參科學中藥三天對鼠肝微粒體 CYP 蛋白含量之影響。利用免疫染色方法進行分析,使用鼷鼠單株抗體MAb 1-7-1、goat anti-rat 2B1、MAb 2-13-1 及 goat anti-rat GSTYb 抗體分別分析 CYP1A、 CYP2B、CYP3A 及 GST 蛋白表現量。結果顯示出苦參科學中藥 處理後,使CYP1A、CYP2B、CYP3A 及 GST 免疫相關蛋白之蛋白表現量 分別造成2 倍、3 倍、175%、24%及 28%的增加(Figure 5)。
三、苦參水萃取物對C57BL/6J 鼷鼠之處理劑量與活性關係
(一)苦參水萃取物之劑量與活性關係
進行苦參水萃取物之劑量與鼠肝 EROD 和 PROD 活性關係之實驗。將 C57BL/6J 鼷鼠分別餵食 0.1、0.5、0.75、1、3 g/kg 水萃取物,連續餵食三 天後,但在餵食水萃取物3 g/kg/day 一天後,鼷鼠死亡,所以取 0.1、0.5、
0.75、1 g/kg/day 水萃取物的鼠肝微粒體作 EROD 和 PROD 活性測試。結果 顯示,餵食水萃取物在1 g/kg/day 後,可造成 EROD 和 PROD 活性明顯上 升(Figure 6),而 1 g/kg/day 升高倍數較高,故接下來的實驗以 1 g/kg 劑量 的苦參水萃取物來處理C57BL/6J 鼷鼠,以建立藥物代謝酶活性誘導之相關 性。
(二)苦參水萃取物對鼠的肝臟與腎臟藥物代謝酶活性之影響
為了瞭解水萃取物對肝臟及腎臟單氧酶活性和結合酵素之影響,以 1g/kg/day水萃取物餵食鼷鼠,連續餵食三天後,取得鼠肝及腎之微粒體測定 單氧酶活性。結果顯示水萃取物處理組,在體重、肝重及微粒體蛋白含量與 對照組比並無明顯差異,但在CYP、cytochrome b5和NADPH-CYP reductase 活性分別上升26%、13%和 18%。在肝臟單氧酶方面,使EROD、PROD、
coumarin hydroxylation及NFO的活性分別上升 122%、221%、58%及 43
%; 但對於NDM及TOH活性並無影響(Table 2)。在結合酵素測定上,結果 顯示GST的活性上升 55%,而UGT及NQO的活性並無明顯的差異(Table 3)。 在腎臟方面,腎重、微粒體蛋白含量及 NADPH-CYP reductase、NQO
及GST 的活性均不受餵食水萃取物的影響(Table 4)。而 EROD、PROD 及 UGT 的活性均測不到。
綜合上述,可得知在肝臟中 NADPH-CYP reductase 及 GST 的活性皆有 增加,但在腎臟中,二者活性並沒有增加情形;在NQO 活性測定中,肝臟 及腎臟皆無影響。
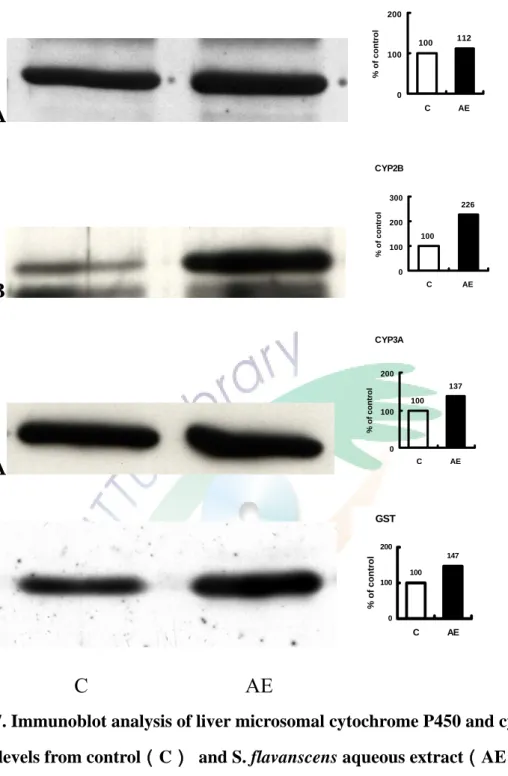
(三)苦參水萃取物對 CYP 蛋白含量的影響
將 C57BL/6J 鼷鼠餵食 1g/kg/day 苦參水萃取物三天對鼠肝微粒體 CYP 蛋白含量之影響。利用免疫染色方法進行分析,使用鼷鼠單株抗體MAb 1-7-1、goat anti-rat 2B1、MAb 2-13-1goat anti-rat GSTYb 抗體分別分析 CYP1A、CYP2B、 CYP3A 及 GST 蛋白表現量。結果顯示出苦參水萃取物 處理後,使CYP1A、 CYP2B、CYP3A 及 GST 免疫相關蛋白之蛋白表現量 分別造成12%、2 倍、37%及 47%的增加(Fighure 7)。
四、苦參甲醇萃取物對C57BL/6J 鼷鼠之處理劑量與活性關係
(一)苦參甲醇萃取物之劑量與活性關係
進行苦參甲醇萃取物之劑量與鼠肝 EROD 和 PROD 活性關係之實驗。
將C57BL/6J 鼷鼠分別餵食 0.1、0.5、0.75、1、3 g/kg 甲醇萃取物,連續餵 食三天後,測定藥物代謝酶之含量或活性。但在餵食甲醇萃取物3 g/kg/day 一天後,鼷鼠死亡,所以取0.1、0.5、0.75、1 g/kg/day 甲醇萃取物的鼠肝微 粒體做EROD 和 PROD 活性測試。Figure 8 顯示,餵食甲醇萃取物在 1
g/kg/day 後,可造成 EROD 和 PROD 活性明顯上升,而 1 g/kg/day 升高倍數 較高,故接下來的實驗以1 g/kg 劑量的苦參甲醇萃取物來處理 C57BL/6J 鼷 鼠,進一步分析其他P450 的活性及 UGT、GST 及 NQO 之活性等。
(二)苦參甲醇萃取物對鼠的肝臟與腎臟藥物代謝酶活性之影響
為了瞭解甲醇萃取物對肝臟及腎臟單氧酶活性和結合酵素之影響,以 1g/kg/day甲醇萃取物餵食鼷鼠,連續餵食三天後,取得鼠肝及腎之微粒體測 定單氧酶活性。結果於Table 2 顯示甲醇萃取物處理組,在體重、肝重、微 粒體蛋白含量及cytochrome b5活性與對照組比並無明顯差異,其餘在CYP和 NADPH-CYP reductase活性分別上升 15%和 26%。在肝臟單氧酶方面,使 EROD、 PROD及 coumarin hydroxylation的活性分別上升 75%、192%及 60
%;但對於NDM、 NFO及TOH活性並無影響。在結合酵素測定上,結果顯 示NQO 及GST的活性分別上升 26%及 43%,而UGT並無明顯的差異(Table 3)。
在腎臟方面,腎重、微粒體蛋白含量、NADPH-CYP reductase、NQO 及GST 均不受餵食甲醇萃取物的影響與對照組無明顯的差異,其中 EROD、
PROD 及 UGT 均測不到(Table 4)。
綜合上述,可得知在肝臟中 NADPH-CYP reductase 、GST 及 NQO 的 活性皆有增加,但在腎臟中,三者活性並沒有增加情形。
(三)苦參甲醇萃取物對 CYP 蛋白含量的影響
將 C57BL/6J 鼷鼠餵食 1 g/kg/day 苦參甲醇萃取物三天對鼠肝微粒體 CYP 蛋白含量之影響。利用免疫染色方法進行分析,使用鼷鼠單株抗體 MAb 1-7-1、goat anti-rat 2B1 及 goat anti-rat GSTYb 抗體分別分析 CYP1A、 CYP2B 及GST 蛋白表現量。Fighure 9 顯示出苦參甲醇萃取物處理後,使 CYP1A、
CYP2B 及 GST 免疫相關蛋白之蛋白表現量升高為 41%、2 倍及 28%。
五、苦參科學中藥對Sprague-Dawley 大鼠之處理劑量與活性關係
(一)苦參科學中藥之劑量與活性關係
進行苦參科學中藥之劑量與鼠肝 EROD 和 PROD 活性關係之實驗。將 Sprague-Dawley 大鼠分別餵食 1、3 g/kg 科學中藥,連續餵食三天後,取鼠 肝微粒體作EROD 和 PROD 活性測試。Figure 10 顯示,餵食科學中藥在大
於3 g/kg/day 後,可造成 EROD 和 PROD 活性明顯上升,而 3 g/kg/day 升高 倍數較高,故接下來的實驗以3 g/kg 劑量的苦參科學中藥來處理
Sprague-Dawley 大鼠,以建立藥物代謝酶活性誘導之相關性。
(二)苦參科學中藥對大鼠的肝臟、腎臟及肺臟藥物代謝酶活性之影響 為了瞭解科學中藥對肝臟及腎臟單氧酶活性和結合酵素之影響,以
3g/kg/day科學中藥餵食大鼠,連續餵食三天後,取得肝、腎及肺之微粒體測 定單氧酶活性。結果顯示科學中藥處理組,在體重、肝重及微粒體蛋白含量 及cytochrome b5、NADPH-CYP reductase活性與對照組比並無明顯差異,但 CYP活性上升 33%。在肝臟單氧酶方面,使EROD、PROD及NFO的活性分 別上升131%、110%及 96%; 但對於NDM 及TOH活性並無影響,而coumarin hydroxylation則是測不到(Table 5)。在結合酵素測定上,結果顯示GST的活 性上升43%,而UGT及NQO並無明顯的差異(Table 6)。
在腎臟及肺臟方面,重量、微粒體蛋白含量、NADPH-CYP reductase、
NQO 及 GST 均不受餵食科學中藥的影響與對照組無明顯的差異,其中 EROD、PROD 及 UGT 均測不到(Table 7、Table 8)。
綜合上述,可得知在肝臟中 GST 的活性有增加,但在腎臟或肺臟中,
則無增加情形;在NADPH-CYP reductase 及 NQO 活性測定中,無論在肝臟、
腎臟或肺臟皆無影響。
(三)苦參科學中藥對CYP 蛋白含量的影響
將 Sprague-Dawley 大鼠餵食 3 g/kg/day 苦參科學中藥三天對鼠肝微粒體 CYP 蛋白含量之影響。利用免疫染色方法進行分析,使用鼷鼠單株抗體 MAb 1-7-1、goat anti-rat 2B1、MAb 2-13-1 及 goat anti-rat GSTYb 抗體分別分析 CYP1A、CYP2B、 CYP3A 及 GST 蛋白表現量。結果顯示出苦參科學中藥 處理後,使CYP1A、 CYP2B、CYP3A 及 GST 免疫相關蛋白之蛋白表現量 分別造成的增加35%、4 倍、97%及 43%(Fighure 11)。
六、苦參科學中藥對鼠肝及腎臟功能之影響
評估大鼠對苦參科學中藥是否對肝及腎臟功能造成的影響,因此以3g/kg 科 學中藥連續餵食三天後,以心臟採血的方式取得大鼠的血清來測試肝臟及腎臟功 能的變化,其測試結果顯示出苦參科學中藥在處理前及處理後之鼠血清肝功能
AST 及 ALT 的值,並無明顯改變;而測試腎功能之 BUN 及 CRE 的值在處理前 及處理後也無明顯差異(Table 9)。
七、苦參科學中藥在大白鼠體內對茶鹼藥物動力學之影響 1.標準檢量線
製作大鼠血液檢品中theophylline 標準檢量線,濃度範圍為 0μg/ml 至 5μg/ml,濃度與積分面積比之回歸,其直線回歸方程式為 y=118.7x+0.08,r
=0.9997,如 Figure12 所示,得之標準檢量線,可用於每回定量大鼠血液檢品 中theophylline 之濃度。
2.同日內及間日內試驗
選取 1、2 及 3 μg/ml 三種濃度製作標準品,於同一日及不同的三日重複分 析各三次,將所得之相對積分面積比數據計算標準偏差,結果如Table 10 及 Table 11 所示,同日內及間日內分析之變異係數均小於 10﹪;顯示本實驗之分 析方法對大白鼠血液檢品中theophylline 之檢測,可達到精確之定量分析。
3. 苦參科學中藥與茶鹼的交互作用
在實驗中苦參科學中藥以預處理的方式,實驗大鼠先利用餵管口服科學中 藥3g/kg/day 連續給予三天,第四天進行 theophylline(2 mg/kg,i.v.)與苦參 科學中藥之交互作用,來研究其theophylline 藥物動力學之影響。結果在 Figure13 顯示對照組與苦參科學中藥處理的血中 theophylline 濃度與時間關係 曲線圖。在A 圖中,顯示具有 fast distribution 及 slower elimination 之二相分佈,
因此採用two-compartment 模式進行動力學分析。在圖 B 中,可以看到處理組 的血中theophylline 濃度在 60 分鐘以後較為低,使用 Win-Nonlin 程式,以 two-compartment model 計算 theophylline 在大鼠血液的藥物動力學參數(Table 12),結果顯現出 AUC 及 AUMC 分別降低 23%及 36%而 Cl 增加 31%。
結論
在苦參科學中藥、水萃取物及甲醇萃取物中,測定matrine、sophoraflavanone G 及isokurarinone 的含量,苦參科學中藥及甲醇萃取物均可測得 matrin、sophoraflavanone G 及 isokurarinone;而在水萃取物中,均未測到 matrine、sophoraflavanone G 及 isokurarinone 的存在。
在鼷鼠實驗中,將鼷鼠連續處理三天餵食苦參科學中藥 3 克/公斤/天,鼠肝 EROD、PROD、coumarin hydroxylation、TOH 及 NFO 活性增加,而對 NDM 之活性 則無影響。若以1 克/公斤/天水萃取物處理鼷鼠,連續處理三天,使肝 EROD、PROD、
coumarin hydroxylation 及 NFO 活性增加,但對 TOH 和 NDM 活性無影響。以 1 克/
公斤/天甲醇萃取物處理鼷鼠,連續處理三天,使鼠肝 EROD、PROD 及 coumarin hydroxylation 活性增加,但對 TOH、NDM 及 NFO 活性無影響。而科學中藥及萃取 物皆對肝臟及腎臟UGT 活性均無影響;但科學中藥、水萃取物及甲醇萃取物均可造 成肝GST 的活性增加;經甲醇萃取物處理後會增加肝 NQO 活性。但不管經由苦參科 學中藥及其萃取物處理後,藥物代謝酶之活性與控制組無明顯差異,表示在不同組織 中會有不同之反應。用免疫轉印法分析肝臟中的P450 及 GST 蛋白含量之變化,結果 顯示苦參科學中藥會使CYP1A、CYP2B、CYP3A 及 GST 蛋白含量的增加,水萃取 物使CYP1A、CYP2B、CYP3A 及 GST 之蛋白含量的增加,而甲醇萃取物會使 CYP1A、CYP2B 及 GST 的蛋白表現量增加。
此外,為了要探討在不同動物中是否有相同的反應,所以在大白鼠實驗中,連續 三天餵食苦參科學中藥3 克/公斤/天,鼠肝 EROD、PROD 及 NFO 活性增加,而對 TOH 及 NDM 活性無影響,而 coumarin hydroxylation 則是測不到。科學中藥可造成 鼠肝GST 的活性增加。分別在鼠腎及肺臟中藥物代謝酶之活性均與控制組無顯著的 差異,亦表示在不同組織中會有不同反應。所以在免疫轉印法分析肝臟中的P450 及 GST 蛋白含量之變化,結果顯示苦參科學中藥會使 CYP1A、CYP2B、CYP3A 及 GST 蛋白含量增加。
而在評估大鼠對苦參科學中藥是否會對肝功能及腎功能兩方面造成的影響,其測 試結果顯示出苦參科學中藥在處理前及處理後之鼠血清肝功能AST 及 ALT 的值,並 無明顯改變;而在腎功能之BUN 及 CRE 的值在處理前及處理後也無明顯差異。
在苦參科學中藥對茶鹼之交互作用情形得知,經由科學中藥餵食後與控制組相比
能降低茶鹼在大白鼠血液中的濃度變化情形。而在two-compartment model 計算 theophylline 在大鼠血液的藥物動力學參數,結果顯現出可降低 AUC 及 AUMC 而增 加其清除率。
討論
近年來不論在東方或西方社會裡都會拿中草藥來治療疾病或長期做保健食品來 強身健體,有時會與西藥併用,因此中草藥與西藥之間產生的藥物交互作用機率也不 可忽視,中草藥與其他中西藥物合併使用後可能會因此產生藥物之間的交互作用使其 療效降低,或產生毒性作用等副作用,所以天然藥物影響藥物代謝酶所造成之藥物交 互作用應值得讓我們重視,例如:St. John,s wort可誘導CYP3A3/4進而促進tretinoin、
anastrozole、doxorubicin及estradiol等的代謝,使其治療效果降低(Block and Gyllenhaal, 2002)而造成對病人不利的影響。因此在評估中草藥藥效的同時,以體內及體外實驗 來探討藥物對於藥物代謝酶之影響,了解可能會因而引發造成藥物之間的交互作用問 題,對於發展中草藥之臨床使用是很重要的。
藥物代謝酶引起之誘導或抑制會與藥物之藥理活性或毒性有種密切關係,若有人 同時使用多種中草藥或其他藥物,則會影響到此藥之毒性與誘導藥物代謝酶的影響,
進而可能產生藥物之交互作用,而影響到民眾用藥之安全性,所以應該研究藥物代謝 酶之作用機轉及其藥物是否會誘導或抑制藥物代謝酶的作用,而避免產生藥物交互作 用,對於藥物開發或應用於臨床上是很重要的指標。
苦參具有清熱燥濕、利尿和抗菌等功能,可用來治療腸胃道疾病及濕疹。大鼠在 肝及腎功能測定中,苦參科學中藥在處理前及處理後之鼠血清肝功能AST及ALT的 值,並無明顯改變(Table 9);而測試腎功能之BUN及CRE的值在處理前及處理後也 無明顯差異(Table 9)。由血清指標測定值的結果來看,顯示出經苦參科學中藥處理 之後,鼠肝及腎功能無顯著的影響。
然而為了瞭解苦參在臨床上的使用情形,我們利用苦參科學中藥,來探討苦參科 學中藥對藥物代謝酶之影響。鼷鼠經苦參科學中藥處理後,不論在EROD或PROD活 性的時程或劑量皆有增加情形,而其中最高的劑量為3g/kg(Figure 3、Figure 4),一 般而言,平均每人(體重為60公斤)建議每日的苦參用量為0.4~1.2克,為鼷鼠的劑量 0.2%~0.7%。而從實驗結果得知(Table 1、Table 3),在單氧酵素測試方面,經科學 中藥處理之CYP的含量以及NADPH-CYP reductase活性均可受苦參的誘導增加顯現 出單氧酵素的組成已有了變化,可能因此會影響到CYP的活性。此外,苦參科學中藥 會使EROD、coumarin hydroxylation、PROD、TOH、NFO及GST的活性都有明顯的上 升情形,分別顯示出CYP1A、CYP2A、CYP2B、CYP2C9、CYP3A及GST的活性受
到誘導影響。由免疫轉印方法分析結果得知CYP1A、CYP2B、CYP3A及GST的蛋白 含量皆有增加,與酵素活性測定的結果一致顯示出其作用機轉發生在蛋白表現的階段 以前。
另外比較分析苦參水萃取物與苦參甲醇萃取物之間的關係,在單氧酵素測定中
(Table 2),可看出在水萃取物和甲醇萃取物處理之CYP的含量以及NADPH-CYP reductase活性都有上升的現象,且使得EROD、PROD及coumarin hydroxylation的活性 都有明顯的上升情形,除此之外經由水萃取物處理之後亦增加cytochromeb5的含量及 NFO的活性。在免疫轉印分析結果中,水萃取物會誘導CYP1A、CYP2B及CYP3A的 蛋白表現量,而甲醇萃取物則會誘導CYP1A及CYP2B的蛋白表現量。在結合酵素方 面(Table 3),水萃取物和甲醇萃取物皆會增加GST的活性,經由免疫轉印方析中結 果發現,上述兩者萃取物皆可增加GST的蛋白表現量。此外,經甲醇萃取物處理後會 增加NQO的活性。雖然不論用苦參科學中藥、水萃取物或甲醇萃取物處理後,coumarin hydroxylation的活性都有明顯上升情形,顯示出CYP2A的活性受到誘導,但由於 CYP2A的抗體尚未取得,故CYP2A的活性結構應進一步探討;另外,此三種苦參製 物對於CYP2E1的活性皆不受影響。由於這三種苦參製劑均可造成藥物代謝酶之誘導 作用,但受誘導的選擇性並不完全相同,如對於CYP2C、CYP3A及NQO之誘導影響
(Table 1-3),除了肝臟是藥物代謝主要場所之外,另外腎臟也是代謝器官重要之一,
無論是經由苦參科學中藥、水萃取物或甲醇萃取物處理後,CYP、UGT、GST及NQO 皆不受影響(Table 4)。綜合上述結果得知,苦參會因不同萃取方式或鼠的組織之不 同對藥物代謝酶造成不同的影響。因此中藥材製程之標準化在臨床應用上是很重要。
根據實驗結果得知,苦參會對CYP1A、CYP2B、CYP3A及GST的誘導影響,在 臨床上若使用苦參時應注意可能產生之藥物交互作用及副作用。CYP1A的藥物受質 包含如:theophylline、caffine及ropivacaine等(Backman et al, 2006),若苦參與這些 藥物同時併用後,則會產生藥物交互作用,使其藥物的療效降低。CYP2B可代謝 phenobarbital、clotrimazole及chlorpromazine等(Kretschmer and Baldwin, 2005)。而超 過50%的藥物是藉由CYP3A4代謝掉,例如:midazolam、budesonide、diltiazem、lidocaine 與nifedipine等(Zuber, 2002)。
在CYP1A誘導方面,鼷鼠與人為同Ah-responsive一類,且鼷鼠和人的CYP1A1及 CYP1A2皆會受到 PAH之誘導,其誘導程度會因種別的特異性或個體之間的差異性而 有所不同,例如:human CYP1A與Ah receptor polymorphisms會造成個體間CYP1A之