行政院國家科學委員會專題研究計畫成果報告
計畫編號:NSC 90-2314-B-006-095
執行期限:90 年 8 月 1 日至 91 年 7 月 31 日 主持人:白明奇 成大醫院神經科
共同主持人:無
計畫參與人員:蕭世朗 中正大學心理系
一、中文摘要
阿茲海默氏症 (Alzheimer’s disease;
AD) 是高齡化社會重要的疾病,其早期的病 變位置侷限於海馬,隨病情進展,逐漸向頂 葉及其他新皮質擴展,由於認路能力與敘述 性記憶皆需海馬功能,因此,推斷認路障礙 應該伴隨失憶症狀的出現,這對照顧者、家 庭及社會之衝擊甚大。依據 Piaget 兒童發 展心理學的理論,人類對認路能力的獲得有 一定的順序,先是地標再認、接著是「自我 中心」的認路、最後是「他中心」的地圖概 念,這種經驗增長與神經網路架構有對應關 係,對應的解剖位置各為下枕顳葉、頂葉及 顳葉海馬區;Reisberg 所提出的「逆生」
(retrogenesis) 概念,若應用於認路行為 及病理變化,恰與 Braak-Braak 的 AD 病理 分期一致。本研究所要驗證的即是隨著 AD 疾病之進展,認路行為首先會出現「他中心」
認路障礙,接著是「自我中心」認路障礙,
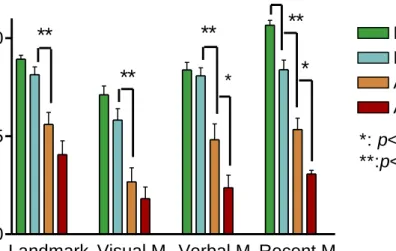
最後是地標再認障礙。本研究利用三個實驗 設計,分別代表前述三種認知功能。共有 81 人完成實驗,包括正常組 21 人、MCI (輕 度認知障礙,AD 的前驅) 組 22 人、及 AD 病人 38 人。AD 患者在測驗均顯示嚴重認知 障礙,地標再認能力仍未達底限;MCI 組的 表現,在「自我中心」認路能力劣於正常組,
「他中心」的認路能力也略劣於正常組,地 標再認則與正常組相同。這些結果與假設預 測一致;「自我中心」的認路測驗具有分出
有所幫忙,同時對困擾照顧者之患者認路問 題提供行為治療的指引。
關鍵詞:阿茲海默氏症、認路症狀、神經 病理、認路測驗、早期診斷
Abstract
Alzheimer’s disease (AD) has become an important and prevalent degenerative disease in the elderly. In order to treat the patients earlier and manage them properly, it is important to detect their cognitive impairment as early as possible. Navigation and declarative memory share same neurosubstrate of hippocampal function. Since declarative amnesia is a major manifestation of preclinical AD, navigation impairment would appear early in AD, although it is overlooked.
From animal and human studies, it has been known that critical components for a successful navigation include landmark recognition, egocentric route learning, and allocentric representation. Their corresponding neuroanatomical structures are lingual gyrus, posterior parietal lobe and hippocampus, respectively. Inferred from the theory of
developmental psychology of Piaget concerning navigation and retrogenesis theory by Reisberg, AD patients would lose their navigation ability firstly in allocentric representation, followed by
egocentric route learning and finally, landmark recognition. The anatomical regions involved in this sequence are compatible with the
Braak-Braak stages for AD.
Real life navigation tasks including the aforementioned 3 components were used to test this hypothesis. A total of 81 participants completed the study, including 21 normal subjects, 22 minimal cognitive impairment (MCI), and 38 early AD patients. MCI patients are supposed to represent a majority of
preclinical AD.
As predicted, early AD performed worst on all tasks. MCI patients performed worse than normal subject on egocentric route learning, which might be a good discriminator for MCI and normal. The manifestations of the three groups on three tasks are compatible with the hypothesis. Moreover, the different
manifestation among group provides advise for early AD patients and their care-givers.
Keywords: topographical orientation, navigation, Alzheimer’s disease, dementia, retrogenesis, early diagnosis, Piaget
二、其餘請見完整報告
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※※
※ ※
※ 早期阿茲海默氏症病患之認路能力研究 ※
※ ※
※※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:■個別型計畫 □整合型計畫 計畫編號:NSC 90-2314-B-006-095 執行期間: 90 年 8 月 1 日至 91 年 7 月 31 日
計畫主持人:白明奇 共同主持人:無
計畫參與人員:蕭世朗 魏淑珍 黃欽威 魏智煌 張雲雯
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位: 國立成功大學附設醫院神經部
中 華 民 國 91 年 10 月 26 日
第一章 緒論
阿茲海默氏症 (Alzheimer’s disease; AD),最常見的原發性皮質失智症 (Mesulam, 2000),無疑地將成為 21 世紀人類重要之疾病,台灣也不例外 (Liu, Lin, Teng et al, 1995;
Liu, Lin, Chen et al, 1996)。
AD 臨床過程長達數十年,發病前的期間可達 25 年 (Bookheimer, Strojwas, Cohen et al, 2000; Braak & Braak, 1996),而發病後病程則依照顧品質,最長可達 20 年
(Mesulam, 2000)。因此,有關早期 AD 的臨床症狀學的研究實屬重要,對有家族遺傳 史或具有危險因子 (如 E4 極脂蛋白等) 的人,更是如此。
早期 AD 病人會出現反覆詢問等失憶症狀,這是因為與記憶功能極為相關的海馬 迴結構產生了病變。海馬迴在鼠類、鳥類及其他動物負責認路的功能,這些動物的海 馬體積相對於全腦的比例,遠大於人類。但已有證據顯示,人類的海馬及相關結構在 認路行為上也扮演著重要角色。因此,在早期 AD 病人出現認路障礙是可以預期的。
所謂認路障礙,是指一個人無法在其周圍的環境中獲得充份之定向感,以致無法 到達想去的地方 (Barrash, 1998)。然而,認路障礙在 AD 病人症狀研究上卻未被重視。
有人認為 AD 全盤的認知功能敗壞便足以解釋此一障礙,而阻礙了此類的研究;同時,
當病人出現認路障礙時,病人本身會避免單獨外出或遠行,家屬也會採取許多對策,
因此,降低了此症狀被旁人觀察到的機會。
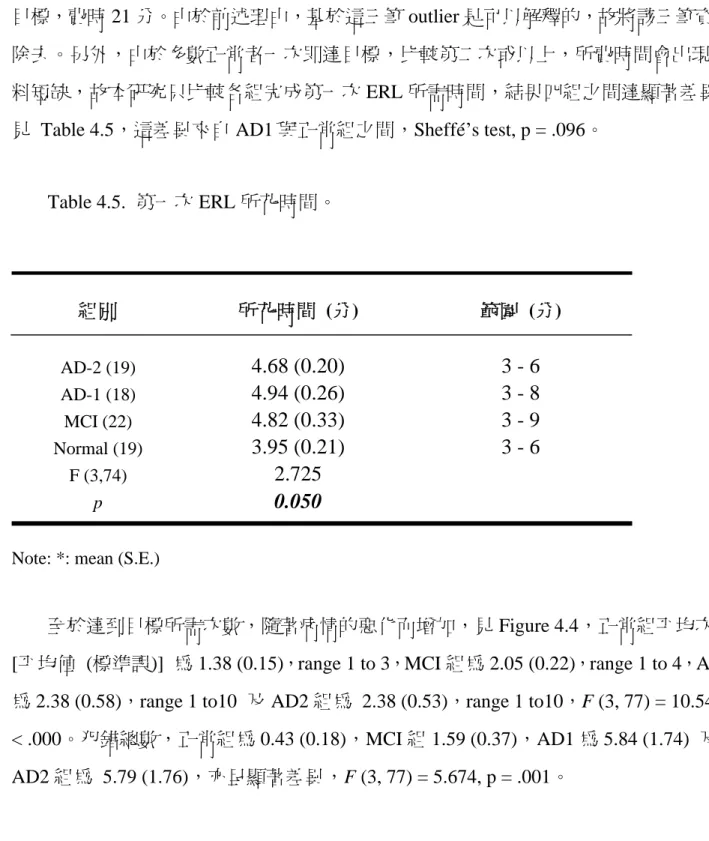
但是失智病人的走失,導致嚴重意外,是一個經常發生的事實。Pai 及 Hsiao (2002a) 曾對南台灣 AD 病患的家屬進行問卷調查,155 位病人中有 11 (7 %) 位病人以「迷路」
為初發症狀,這些病人造成家屬困擾,遠較以其他表現為初發症狀的病人來得大,因 而提早就醫。後來,Pai 及 Hsiao (2002b) 又以結構式問卷詢問 123 位輕度至中度 AD 病人的家屬,其中有 54%的家屬認為病人確實有認路問題,而曾經被他人帶回者達 19%,有 86% 的家屬不放心讓病人單獨外出;16% 的家屬認為迷路是病人的初發症 狀。家屬不放心讓病人單獨外出的比例,遠高於病人有認路障礙的比例,這中間的差 距,值得思考。同時一般神經心理學測驗並無法預測病人迷路與否,這說明認路困難
McShane, Gedling, Keenw, Fairgurn, Jocob 與 Hope (1998) 也曾追蹤 104 位 AD 病 人,每 4 個月詢問一次,這些病人一開始都是住在自宅。5 年之間,有 43 位病人至少 有一次被別人帶回家,有 5 位病人一再迷路;46 位病人曾經被鎖在家裡。曾迷路的病 人比不曾迷路的人後來被送進安養機構的比例高出很多。
認路障礙、甚至迷路,確實是早期 AD 病人的重要症狀,對照顧者、家屬及社會,
衝擊不小。進一步探討 AD 病人的認路行為,相信對 AD 之病理生理機轉會有所助益;
同時,也能對照顧者提供有用的指引。
第二章 文獻探討
人類地域失認症的研究
除了 AD 病人經常迷路外 (McShane, 1998),迷路也會發生於天生認路能力差 (俗 稱路痴) 的人,以及腦部外傷 (Skelton & Gabrieli, 2000; Hecaen & Albert, 1986)、腫瘤 (Suzuki, Yamadori, Hayakawa & Fujii, 1998)、中風 (Pai, 1997; Lin & Pai, 2000)、偏頭痛 發作 (Martins & Sa, 1999) 或癲癇發作的病人。從這些腦傷病人之病灶位置與迷路行為 的關係,可提供研究 AD 症狀學的重要訊息。
因腦傷而導致地域失認 (topographical disorientation, TD) 或迷路之個案,學者稱 呼不一。直到最近,才有結合動物實驗與人類腦傷對認路障礙做有系統分類的文章發 表 (Aguirre & D’Esposito, 1999; Barrash, 1998; Barrash, Damasio, Adolphs, Tranel, 2000),其中 Aguirre (1999) 的綜論最具代表性,他將地域失認分為四類,即 1) 地標 失認 (landmark agnosia);2) 以自我為中心 (egocentric) 的地域失認;3) 頭向 (heading) 地域失認及;4) 前向性 (anterograde) 地域失認。
「地標失認」的病人,對原來被自己用來做地標之建物或特徵不再有反應,這是 屬於一種視覺失認。為了要彌補此一缺失,病人必須依賴細微的環境特徵,如特殊之 大門把手、郵箱或公園坐椅。病人可能有正常的環境知覺,但是其「地理上之意義」
已被剝奪。這類病人之病灶多發生於右側後腦枕葉,更精確地說是右側舌狀腦迴
(lingual gyrus),這與「認臉細胞」(face neuron) 所在之扇形腦迴 (fusiform gyrus) 相近。
這可以解釋地標失認與人臉失認經常並存的現象。(右側)舌狀腦迴與地標之間的關係,
也可以從功能性磁振造影 (functional MRI; f-MRI) 的研究獲得證實,當地標或建築物 出現時,此處之血流上升,代表神經元正興奮,這細胞因而被稱為地標細胞 (landmark cells)。「地標細胞」的發現,以及傷及該區造成地標失認的個案報告,會是一個重要 的契合。
從人類開始築屋至今不過數萬年,有人認為不太可能形成一個專門為了辨認建築 物的神經元;但文字的發明使用也才幾千年,腦中卻有文字的辨認區 (Warrington &
Shallice, 1980; Polk & Farah, 1998)。如果從靈長類的社會行為,乃至於人類的生活型 態,認錯臉或認錯住家往往會導致嚴重的後果,因此發展出專門辨認臉孔與建築物的 神經元,在演化上應屬合理。專門辨認文字的腦區或神經元的形成也可以用後天的制 約或訓練使某些細胞被塑造成某一功能,此與海柏定律 (Hebb’s rule) 或長期增益 (long-term potentiation) (Lømo, 1968) 極有關連。
Levine, Warach, & Farah (1985) 認為地域失認可以是兩個分開的系統之毀損,一個 是前述的地標再認,另一個是空間位置的表徵。
以路為基礎的 (route-based ) 大格局空間的表徵,是在以「自我中心」的空間領域 內形成,一個人可以有正常的視覺再認能力,但卻無法理解「以自我為中心」的空間 關係,這便是「以自我為中心的地域失認」:病人可以認出地標線索,也許也可以描 述它們在空間中之擺設,但是卻無法真正在街道中行走以抵達目的地;這種病人失去 了「以自我為中心」的認路 (egocentric route learning) 能力 (後述),病人的腦傷位置 多位於頂葉後部。
「頭向地域失認」的病人雖然知道目標就在那裡,卻無法走到該處,這種症狀十 分罕見,會被提出是呼應 Taube, Muller & Ranck (1990) 在動物的視丘及前下腳發現的 一群細胞,這些細胞當鼠的頭朝向某一個方向 (相對或絕對) 時會被激發,稱為頭向細 胞 (head direction cells)。不過,人類發生「頭向地域失認」的腦傷病變位置,卻是在 扣帶迴後部。
前向性地域失認及逆向性 (retrograde) 地域失認的分別,類似於敘述性失憶的分 法。前者代表在「熟悉」的環境不會迷路 (Teng & Squire, 1999),但是到了「陌生」的
環境,則毫無學習能力,意即喪失形成學習環境記憶的能力,這類病人的腦傷位置多 包含右側的海馬及副海馬迴 (parahippocamal gyrus; PHG)。相反地,在自宅附近迷路,
則代表一種嚴重的逆向性地域失認。
從以上之個案與腦傷位置,不同認路功能可能與特定腦區有著對應關係,見 Table 2.1。
Table 2.1. 地域失認的重要分類 (Aguirre, 1999)
病變位置 臨床症狀 障礙內容
後頂葉 以自我為中心的地域失認 無法以自我的觀點對物件位置形成表徵
後扣帶迴 頭向地域失認 無法對外在環境形成方向感的表徵
舌狀腦迴 地標失認 無法對地標形成表徵
副海馬 前向性地域失認 無法對環境訊息產生新的表徵
從動物到人類的認路行為
自然界裡,動物的「找路行為」對物種的延續很重要;動物藉此以覓食、避難、
尋偶及安居。有些動物的認路能力,近乎奇蹟,例如北非的雙色箭山蟻 (Cataglyphis bicolor) 在沒有地標的沙漠中,迂迴前進好幾公尺找到食物後,可以筆直地回到洞口;
鼠類也可以在黑暗的環境中,迅速前進,破壞屋宇結構,然後返回窩巢;黑面琵鷺從 西伯利亞、北韓一路南下台灣七股,一旦春回大地,又可安然飛回北陸;鮭魚、信鴿、
大海龜的例子 (Gagliardo, Loale & Bingman, 1999),比比皆是,除了某些動物具有特殊 的感覺受器外 (如侯鳥具有磁場受器) (Finney, 1995),當動物移動時,從視覺、嗅覺、
觸覺、聽覺,甚至前庭及本體感覺等送入大腦的訊息,會不斷地進行登錄、更新、計 算及貯存的過程,隨著物種的進化,這套作業系統有可能愈發複雜。
候鳥、海龜、信鴿等可利用磁場、氣壓、偏光、或紫外線來獲取必要的訊息,以
進行長距離的移行。至於短距離的移行,動物可能採用下列方法來認路 (Pearce, 1997),包括氣味、dead reckoning、單一地標導航 (如蜂及天竺鼠)、多重地標導航以及 地理關係。到底動物是如何運作這些複雜的找路的行為呢?這些行為又有何種神經系 統做支持呢?
認知地圖 (Cognitive map)
二十世紀初的心理學界,由 Thorndik、Watson 及 Skinner 等人主導的行為學派,
將動物之行為看成刺激反應的關係。到了 1940 年代,Tolman, Richie & Kalish (1946) 的 實驗打破了這樣的觀點。Tolman 等人證明大白鼠的認路行為以認地方 (place learning) 是較為接近自然狀況下的行為,而非反應學習 (response learning)。
在一個有名的實驗中,Tolman (1948) 提出了「認知地圖」的概念。雖然「認知地 圖」後來被引申為更廣泛及抽象的說法,但就認路而言,Tolman 認為老鼠在腦中可形 成一個類似地圖的表徵,將在移動過程中經驗到之線索,依出現之順序與相對關係組 合成一個平面擺設,這個概念類似於鳥瞰圖。
依照原文,認知地圖有以下幾種定義:
1) A maplike representation which acts as a framework for organizing its sensory inputs and its perceived as remaining stationary in spite of the movement of the organism (O’Keefe, 1979)
2) A record in the central nervous system of macroscopic geometric relations among surfaces in the environment used to plan movements through the environment (Gallistel, 1990)
3) A global representation of objects within some manifold or coordinate system form which their mutual spatial relationships can be derived but that is, to some extent, independent of the object themselves (Leonard & McNaughton, 1990)
「認知地圖」的形成有助於動物選擇一個全新的路徑,以抵達目的地;或當遇到 阻礙時,能找出替代道路 (detour)。「認知地圖」可使動物定義地方、為自己定位、
記得在什麼地方有什麼東西,並且根據現況所需及動機指出如何從甲地到乙地,同時,
它可讓動物知道物體與事件發生所在的空間情境,這是一種「他中心」(allocentric) 的
看法,也就是說從自身以外的立場來看空間陳設。
在 Tolman 發表「認知地圖」的當時,學者咸認在動物腦中應有處理這些訊息的細 胞。23 年後,O’Keefe 及 Dostrovsky (1971) 利用細胞活動紀錄儀,果然證實這種細 胞之存在,這些細胞被稱為地方細胞 (place cells;簡稱 PC)。O’Keefe (1979) 在一篇 綜論中提到 PC 的三點特點:
1) Any cell that consistently fires when the animal is in a particular part of its environment
2) Location-specific cells whose activity cannot be shown to be dependent on a single specific sensory input
3) Only cells whose location-specific firing can be shown to be controlled by any subset of cues that define the environment
PCs 是大白鼠的海馬最主要的細胞類別,佔了海馬中的金字塔型細胞 (pyramidal cells) 將近 90% (Golledge, 1999)。在 O’Keefe 等人 (1971) 的實驗中發現,當大白鼠走 到環境中的特定地方時 (這個位置稱為地方受野 place field) ,某些 PCs 便被激化,而 另一群 PCs 則對應另一個地方受野,其激化之位置與形態,取決於所有被注意的空間 線索所組成之星座 (constellation;此處將線索喻為星星,而將表徵喻為星座之概念有 助於了解這種關係),而非某單一刺激。線索可以來自迷津外 (extramaze) 或迷津內 (intramaze),也可能是不同類別之感覺輸入。至於 PCs 的激化與動物的頭朝向、或者 動物在做什麼事無關 (Nadel, 1999)。
例如八臂迷津中,朝北末端活化的 PC-n90 (n90 是筆者加上,以作為特定標示),
對朝北末端附近的環境顯然形成情境連結,雖然迷津轉動,朝北的臂端永遠會活化 PC-n90;如果將迷津外環境整個移動,則 PC-n90 也跟著轉動。這很容易讓人聯想 PC 完全受視覺刺激的引導。
但情況並非如此簡單。Hill & Best (1981) 讓大白鼠盲聾,但是老鼠到了特定地方,
PC 依然活化,這使得研究者百思不解與驚訝。他們推斷也許大白鼠可以從出發點,根 據先左轉、再右轉等訊息來組成「認知地圖」,這是透過本體感覺得來的訊息。後來,
將大白鼠用毛巾包起來、然後轉幾圈,再擺進去迷宮,結果 PC 就沒有活化了。另外,
McNaughton, Leonard & Chen (1989) 利用單一細胞紀錄,得到頂葉細胞可以反應大白
鼠的動向,或左或右,趨向目標或遠離目標,而海馬便是利用這些頂葉細胞所提供之 訊息,來追蹤定位大白鼠在黑暗迷宮中的行徑。
除此之外,大白鼠也可能從前庭系統得到訊息。Wiener & Korshunov (1995) 將口 渴的大白鼠放到一個黑暗的正方形空間內,大白鼠只有在中央及東南角可以喝到水,
每隔一段時間,實驗者便轉動空間 90 度、180 度或 270 度,結果,大白鼠總能自動修 正角度,到地理上的東南方喝到水,角度的訊息處理與前庭系統是極相關的,此種類 似的修正也可見於大白鼠海馬的 PC 活化模式;這也支持 Tolman 所說的自然狀況下動 物的找路行為。
這些從視覺、聽覺、本體感覺或前庭系統所得到的空間訊息,可能都是透過內鼻 皮質 (entorhinal cortex; EC) 送入海馬,因為在 EC (Quirk, Muller, Kubie & Ranck, 1992) 及齒狀迴 (dentate gyrus; DG) (Rose, 1983) 都被發現到具有空間受野 (spatial receptive field) 的神經元,但是這些神經元並非一對一的精準配對。EC 受損的大白鼠無法完成 空間測試,同時其海馬的空間受野,也一併瓦解 (Miller & Best, 1980)。
因此,PC 提供了老鼠在環境中所在位置之神經表徵,進而提供定位的足夠訊息 (Wilson & McNaughton, 1993),每個 PC 都有其各自的空間受野,破壞老鼠的海馬,會 導致老鼠移行能力的受損,尤其所用的實驗是需要形成一個空間的內在表徵時,例如 從陌生的起點回到一個沒有標記的定點 (Morris, Garrud, Rawlins, O’Keefe, 1982)。PC 會隨著動物所在位置,而改變其興奮程度,好似某個特定之地理環境可以引起某個或 某群 PC 之興奮;但是證據顯示,這種訊息的來源應是多線索之整合,而非線索與細 胞單一對應關係。
後來,Taube 等人 (1990) 在大白鼠海馬附近的背側前下腳 (presubiculum) 及前視 丘發現了頭向細胞,這些細胞標記著大白鼠頭的方向,不論其身在何方。頭向細胞的 發現,是先前的神經科學家的預料中事。
路徑整合 (path integration)
從環境中的任何一點回到出發點的能力叫做「路徑整合」,有時稱為 dead
reckoning。這種能力可見於天竺鼠、倉鼠、家鼠、鳥、昆蟲、節肢動物、狗、貓以及
人類 (Pearce, 1997)。海馬參與「路徑整合」的過程,扮演極重要的角色 (Maaswinkel, Jarrard & Whishaw, 1999)。前述 PC 之空間表徵形成,至少有兩個訊息的來源,一個是 來自環境外貌的外感 (exteroceptive) 輸入,另一個便是「路徑整合」之 idiothetic 輸入。
即使外感成份消失,「路徑整合」仍然可以維持 PC 製作「認知地圖」(McNaughton et al, 1989)。
Taube (1999) 曾對路徑整合下定義:
Path integration is defined as the process whereby an animal’s orientation and location are updated continuously based on how fast, how long, and in what direction it is moving through the environment. PI most likely involves several process,
including the establishment of an initial reference point, monitoring the appropriate motion cues, computation of the animal’s new position based on its initial position and subsequent movement, a mnemonic component that stores “on-line” the distance and direction of the initial reference point relative to the animal’s current position.
從演化上來看記憶系統之進展及神經結構
記憶對動物來說,最重要的功用應是認路 (Redish, 1999),如果在原來的空間地圖 上加上時間的成份,則這個系統變成了事件性記憶;Squire & Zola (1996) 也認為與認 路有關的「環境記憶」屬於敘述性記憶之一種。
由於大白鼠的視覺系統不發達,與其所處環境有關 PC 的定位計算,端賴嗅覺、觸 覺、平衡及遠處索引,最重要的是動物必須抵達該處,才能知道身處何處;但靈長類 以其發達的視覺及動眼控制系統,可以探索及記得其所在環境之訊息,不必親臨現場,
便可記得於該處發生過的事件,靈長類海馬的空間景觀細胞 (spatial view cells) (Roll, 1999) 取代了 PC。從靈長類再到人類,演化上最重要的進展當屬語言的形成,使大腦 半球的側化更為明顯,是否因此敘述性記憶逐漸轉趨左側海馬功能,而空間記憶則漸 至右腦?
最近,Clark, Broadbent, Zola & Squire (2002) 破壞老鼠海馬與下腳 (subiculum) 的 研究,發現這些結構與動物的非空間性的記憶相關,且出現時間梯度 (temporally graded) 關係。海馬極有可能在人類與動物的事件記憶 (Scoville & Milner, 1957) 與認路兩種重
人類的找路行為
與其他動物一樣,「找路行為」(wayfinding behavior) 對人類也是很重要,從到一 個熟悉的目的地 (工作場所)、或從一個已知的地方 (家) 向外展開探索過程、甚至在 一個全然陌生的地方 (如國外),人類經常也在找路。正常人可能在陌生的地方迷路,
老人或智障者迷路的消息更是經常出現在報章雜誌、新聞報導,AD 病人是這類走失 人口的大宗。
人類生活的環境與野獸所居的原始叢林大不相同,街道規劃完整、類似的屋宇密 密麻麻,間有醒目的招牌或特別的景觀可為地標,但在認路行為上,與動物共有一個 基本的模式可循。
人類認路的第一步通常是地標的確認,這在孩童的發展心理學上也是首先形成的 (Piaget, 1960)。第二步是路徑學習,利用地標出現的先後順序,輔以如左轉或右轉等指 引,逐步從出發點紀錄到目的地,這是路徑學習的第一特點;路徑學習還需要一種所 謂以自我或身體為中心的座標框架,顧名思義,這是「以自我為中心」看向地標所形 成的概念,這是路徑學習的第二特點;但由於路徑只是一系列的線性指引,這種表徵 是不穩定的,只要重要地標改變、消失,或路障出現,所學過的路徑立刻瓦解,這是 路徑學習的第三特點。及長,逐漸學會類似地圖的表徵,這是以「他中心」來看的空 間表徵,涉及物件間的角度與距離關係,符合歐幾里得 (Euclidean) 幾何學概念,這種 類似於空照或鳥瞰的地圖概念,具有恆久性。
一般而言,在大格局的環境中認路,有兩個不同的認知系統,即地標 (包含地方之 外貌) 和俯視 (survey;含地方之相對空間位置) (Aguirre & D’Espositio, 1997)。在充滿 地標的環境中,人類較會採用路徑學習,例如在都會城市之中 (Acredolo & Evans, 1980);反之,若是沒有什麼重要地標者,則較易採用類似地圖的表徵,如沙漠或荒野 之中 (Heft, 1979)。同時,生物體應有能力將自身經過之路徑記憶逐漸轉為較為抽象的 圖樣表徵,以下將逐項詳述。
地標與建物之再認 景觀、地形、地物的學習與再認,是認路重要的第一步。
這種非天然的建物有別於動物在荒野中所見之自然景觀。由臨床案例來看,不論是中 風、腦傷或開刀後病人產生的迷路 (Whiteley & Warrington, 1978; Levine et al, 1985;
Takahashi, Kawamura, Hirayama & Tagawa, 1989; McCarthy, Evans & Hodges, 1996)、或 失智症病人的迷路,能否認出街景是一大關鍵;同時,移行的認知障礙,多可用策略 或線索來獲得改善,唯獨街景再認的問題最大。
除了一般的物件 (object) 辨認外,透過病案研究與功能性造影術,在人類視覺知 覺的領域,某些專一腦區已被發現,如扇形腦迴,可能與「面貌」辨認有關 (Damasio, Damasio & Van Hoesen, 1983; Farah, 1990),可能也有「文字」辨認區的存在 (Warrington et al, 1980; Polk et al, 1998);特別的是,腦中也有「地標」辨認區,即舌狀腦迴 (Aguirre, Zarahn & D’Esposito, 1998)。這推論來自某些病人發生舌狀腦迴腦傷後,僅出現特殊的 地標失認;病人無法利用明顯的地標幫忙找路 (Whiteley et al, 1978; Levine et al, 1985),尤其是街道景觀、地理風貌、紀念碑物、特別是建築物 (Takahashi et al, 1989;
McCarthy et al, 1996)。街景或建物的再認,是可獨立於一般物件再認之外的。
地標的功用便是為地方做標記,其突兀於周遭之環境為其特點,一般人很容易將 具有這種特徵的物體做為地標之用;此外,地標亦具 anchor point 的功能,即有預測的 作用。生活中,建築物確實是最常被用為地標線索的物體。Appleyard (1969) 在一篇標 題為 “Why buildings are known” 的論文中提到:地標通常因為社會文化意義而引起注 意並被記得。Golledge 與 Spector (1978) 對 Columbus, Ohio 的 400 位居民調查,被選 為最有名與最熟悉的地標特徵中,有一半是與「個人活動型態」有關的地方或特徵。
地標及景觀並非全然相等,當一處地方有明顯的地標時,此地標會被用為線索,
但當所有的成份都不構成線索的顯著度 (salience) 時,則整個環境會被視為一個線 索,稱為情境。但若具備形成線索或情境的成份,則不會留下特別印象,例如一片森 林或海洋,難以形成特別專一性。
以自我為中心的認路 空間的框架 (spatial framework) 若隨著物體移動而移動,是謂
「自我中心」;反之,框架若固定於環境中,或不隨著個體移動而移動,叫「他中心」。
很多人有把車停在大型量販店停車場的經驗,請回憶一下購物完要返回停車位的 方法。首先,有一個大略的方位,地標地物之線索 (含有其他感覺輸入,如氣味、光
強刺激、聲音等),走了多久、多遠的感覺,幾個轉彎或角度改變等,這些還牽涉到前 述事件發生之時間順位之「逆轉」。
「自我中心」的表徵,與視覺及平衡感覺相關性特別大,且與頂葉功能有關,人 或動物在環境中移行,會不斷計算、更新目前所在位置,這是一個動態、暫時性的表 徵,指引走向目的地 (Wang & Spelke, 2000),有別於一個穩定的、永久的「他中心」
的「認知地圖」的表徵。
他中心之認路概念 此即前述的俯視知識。人類可將走過的路畫成地圖;也可以根據 地圖到達目的地,真實的地圖在腦中好像有個 mental counterpart,這圖與空間擺設之 間,自由切換,靈活應用。這種「他中心」表徵沒有地圖的指引,而且在新的環境,
必須靠走過的路徑逐漸更新組合而成,與「路徑整合」很有關連,能夠不靠任何語言 索引回到出發點,就是這種能力之最好證明 (Berthoz & Viaud-Delmon, 1995)。這種「他 中心」表徵的形成,與海馬關係密切。
Meguire, Frackowiak & Frith (1996a) 利用 PET,發現 PHG 與處理這種大格局全貌 的以「他中心」表徵有關。「他中心」的地圖並非可以被知覺,而是以不具空間性質 (spaceless) 的訊息型態進入海馬系統後,「認知地圖」於焉形成,海馬是「認知地圖」
的合成場所。
認路行為及神經結構對應
與空間訊息處理與認路有關的神經結構,最常被提及的當屬頂葉及顳葉,尤其是 海馬及其周邊。
Ungerleider 及 Mishkin (1982) 提出了視覺顳葉及視覺頂葉兩個路徑,見 Table 2.2,前者又稱為腹側或顳向流,負責物體之再認,而後者又稱背側或頂向流,負責物 體之位置,以資引導及控制動作,如伸手以抓觸。
Table 2.2. 視覺輸入的背側流與腹測流 (Ungerleider & Mishkin, 1982)。
Dorsal Ventral
Where What
Posterior parietal cortex Inferior extrastriate cortex Parietal or P-T areas Medial occipital gyrus
Fusiform gyrus Position Appearance
Relative spatial (configural) location Visual appearance of familiar places Survey knowledge Landmark knowledge
地圖可經由局部景觀、走過的路、或其他感覺線索而加以分辨,這些線索進入海 馬,先在 DG 與位置訊息以及「路徑整合」混合,接著通往 CA3,這便可代表在某特 定參考框架 (reference frame) 中的某特定點 (Redish, 1999)。局部景觀中之空間成 份,從頂葉經由背側流到後鼻 (postrhinal) 皮質;而非空間成份,則從下顳葉皮質經由 腹側流到鼻周 (perirhinal) 皮質。嗅覺的情報則來自梨狀 (piriform) 皮質,經由側嗅覺 通路到達側 EC。許多較原始之結構則與皮質下 (subcortical) 結構較具相關性,如基底 核 (含尾核、臥核 nucleus accumbens、蒼白核)、穹窿 (fornix)、扣帶迴、上疊體等。
頂葉 長久以來,人類及靈長類的頂葉,便被認為是處理空間行為的重要所在,從人 類腦傷與猴子單一神經元激化的研究,可證實頂葉確與「自我中心」的空間框架中的 知覺 (perception) 及動作 (action) 有關。這種「自我中心」的空間框架由眼睛、頭及 身體的位置所決定。
靈長類的頂葉內溝將頂葉皮質一分為二,前頂皮質含 Brodmann Area (BA) 1, 2, 3, 43,而後頂皮質 (posterior parietal cortex; PPC) 含 BA 5, 7, 39, 40,前頂皮質可視為初 級體感覺區,而 PPC 則為高次多重感覺運動聯合區;其中 BA 5, 7 與視運動 (visuomotor) 控制有關。傷及頂葉內溝及周邊上頂小葉 (superior parietal lobules),會引起 optic ataxia,這是臨床上 Balint’s syndrome 的症狀之一;而傷及右邊的下頂葉,則會引起半
邊忽略。
海馬 透過新路線的選擇、迂迴路線的採取、或者放射迷宮 (radial maze) 的研究觀 察,研究者認為某些動物在腦中可能形成「認知地圖」。自從 O’Keefe 等人(1971) 在 大白鼠的海馬發現 PC 之後,海馬於是被認為是哺乳類動物移行的神經基礎。建立在 這樣基石的研究,逐漸擴展到倉鼠、鳥類,甚至猴子。
在這類研究中最常用的模型首推 Morris milk maze。Morris 等人 (1982) 在一個直 徑 1.3 公尺的圓池中,注入牛奶,其中有一處隱藏的平台,由於水濁的關係,從水面 難以察覺平台所在;大白鼠不喜歡水,一旦下水,便會拼命的找尋登陸的地方,而平 台是唯一可以離開水的地方。海馬被破壞的大白鼠只會漫無目的的找尋,直到碰巧遇 到平台;而正常老鼠或破壞皮質的大白鼠,只要學習幾次,不管從什麼地方下水,都 可以很快地找到平台。顯然海馬是利用圓池外線索形成相關學習 (relational learning) 之關鍵所在。但是,只要將平台露出水面,即使海馬被破壞的大白鼠也能很快游到平 台,這是因為大白鼠仍然可以利用刺激反應學習。值得注意的是,若是每次都將大白 鼠從同一個地方放下水,大白鼠也會學會朝固定方向游去,好像利用圓池外的某一個 地標作為線索似的 (Eichenbaum, Steward, & Morris, 1990)。海馬與空間學習的證據,還 來自破壞海馬的家鴿無法返巢 (Bingman & Bench, 1990);另外,會藏食物的鳥或鼠 (Sherry, Jacob & Gaulin, 1992),或者能力好的家鴿 (Rehkamper, Haase & Frahm, 1988),
都有較大容積比例的海馬。
海馬由兩個外貌不同的細胞層互扣而成,見 Figure 2.1,這兩個細胞層分別是由顆 粒 (granule) 細胞組成之 DG,及金字塔型細胞組成之海馬本體 (或稱太陽神的角,
cornu ammonis ; CA)。DG 的兩臂恰好包住海馬本體的下臂 CA3 所在,兩臂之間叫齒 門 (dentate hilus),而 CA 可再分為 CA1、CA2、CA3 和 CA4 (通常將 CA4 視為 CA3 的一部份),由表層彎入齒門,海馬本體投射到皮質下的路線有二,即由 CA3 到兩側 的 lateral septum 及 CA1 到同側的 lateral septum (經由穹窿) 的路線。海馬有兩大輸入 來源,最大宗為新皮質輸入,匯集於 EC,經由穿通路徑 (perforant path) 纖維束進入 海馬,這些輸入主要傳送高度處理的感覺訊息;另一為皮質下輸入,分別來自視丘、
下視丘、腦幹、中隔 (septum) 及杏仁核,經由穹窿纖維束,這些與覺醒、情緒及自律
功能有關。
EC 的上游之輸入,主要是鼻周皮質及 PHG,而海馬出力為下腳。EC 可分為 lateral 及 medial 兩部份。高度處理的感覺訊息由感覺皮質進入 EC,再經由穿通路徑進入 DG。
在老鼠,嗅覺系統主要進入 lateral EC。而 EC 廣泛地投射到邊緣系、副邊緣系、嗅覺 區,以及到中隔 (尤其是 lateral 中隔) 及紋狀體。下腳複體 (subicular complex; SC) 接 受來自 CA1 之海馬出力,且與 EC 互有往來。SC 可分為三大部份,分別為下腳、前下 腳及副下腳 (parasubiculum)。下腳投射到內側前額葉皮質、中隔複體、臥核、乳狀核、
杏仁核以及 bed nucleus of the stria terminalis。
EC CA1
CA2 CA3
External Afferent Fornix
Mossy fiber
Schaffer Collateral
Perforant path
Sub DG
Figure 2.1 海馬的神經解剖。
在人類的大腦,所謂的海馬形成 (hippocampal formation) 包括 DG、CA fields 及下 腳,甚至還包含 SC;而所謂的靠中線 (medial) 的顳葉,則除了海馬及其周邊外,還 包含了 PHG 及 rhinal 皮質 (包含鼻周及後鼻皮質),有時甚至連杏仁核也算進去。
人類腦傷是容易在海馬發生,例如癲癇、大腦缺氧等。本世紀最重要的疾病之一 的 AD,其病理變化最早發生在海馬。這不免讓人想到「找不到路」或迷路的症狀,
是否也應該是早期出現的症狀之一?
頂葉與海馬的交通 頂葉與海馬的交通,最主要是互動於鼻周皮質 (相當於猴子的 BA 35/36) 及 PHG 區域。鼻周皮質及 PHG 接受來自視覺、聽覺及體感覺聯合區傳入之多 類 (polymodal) 感覺訊息,同時也有直接來自 PPC 和間接來自海馬之訊息。下頂小葉 後三分之一,間接經由 inferior longitudinal fasciculus 傳到 PHG,另路到前下腳。PHG 及前額皮質 (prefrontal cortex) 也互有往來。這些鼻周皮質及 PHG 接著進入 EC,進而 到達海馬。
此外,雖然海馬與找路的關係密切,但左右腦、甚至腦所支配之對側肢體或觸鬚 (whiskers in rats) 在認路上也有功能上之差別,從 LaMendola 與 Bever (1997) 的研究 得知,左腦與右觸鬚與地圖表徵的學習有關,這是以「他中心」之觀點來習得;相反 地,右腦與左觸鬚則與 rote path 之學習有關,這是以「自我中心」之觀點來習得。這 兩種分法,於人類則適用於下顳葉及頂葉,即前述 Ungerleider 及 Mishkin (1982) 提出 的視覺顳葉及視覺頂葉路徑。
功能性造影研究
由動物與人類腦傷可知,某些大腦區域與認路功能中的特定能力有相關。利用功 能性造影術,如正子放射 (positron emission tomography; PET) 或功能性磁振造影術,
可得知正常人在從事環境道路學習與提取的過程中被活化的腦區,各有所屬,與腦傷 研究和動物實驗結果大致吻合。然而受限於造影術的空間與設備,此類神經心理測驗 無法絕對反應實際情境。
目前利用 PET 或 f-MRI 的研究,對移行的解剖定位仍說法不一,但是關鍵的位置 應該包含內壁側 (mesial) 及右下頂葉皮質、後扣帶迴、部份基底核、左側前額葉、兩 側內壁側顳葉區 (含 PHG) 以及海馬本體。其中,最重要的爭議在於內壁側顳葉區。
利用 f-MRI,Aguirre, Detre, Alsop & D’Esposito (1996) 發現在一個虛擬迷津中,只 有 PHG 被活化,但海馬本身卻沒有。然而,在另外一個研究 Maguire, Frackowiak & Frith (1997) 利用倫敦的計程車司機為受測者,發現只有在回憶地標時 PHG 會活化,但是當 回憶複雜路徑 (complex routes) 時卻沒有。另外一個 PET 研究則發現,只有當回憶物
Petrides & Evans, 1996)。另外,被動的觀看景物而不必去記憶,活化的區域亦是兩側 的 PHG 及扇形腦迴 (Epstein & Kanwisher, 1998; Aguirre & D’Esposito, 1997)。這些結 果可以說明 PHG 及後枕顳葉皮質與物件的空間定位 (object-location associations) 有 關,但卻與較複雜之「認知地圖」形成無關。還有一個利用類似迷津的環境的研究,
發現只有當環境中有地標時,PHG 才會被活化 (Maguire, Frith, Burgess, Donnett &
O’Keefe, 1998)。然而,個案報告有兩側局限於海馬的損害,但保留 PHG 的病人除了 嚴重的事件失憶外,也無法找路 (Vargha-Khadem, Gadian, Watkins, Connelly, Paesschen
& Mishkin, 1997),顯然大格局空間需要海馬本身以為表徵,即使非直接,也與它在事 件記憶所扮演角色有關。
如果考慮環境擺設的複雜性,則可以發現簡單的迷津不會活化海馬本體,因為受 測者可以不用「認知地圖」,而用線性 (linear) 或語言表徵去解決問題。同時,病人 沒有身臨其境之感。相反地,當受測者被要求要說出市區路線、或從攝影機鏡頭的拍 攝過程去學習新路、或回憶剛剛在新環境所學的新路時,都會在功能造影術上激化海 馬本體。因此,從右側海馬送出座標直接指向目標的位置,這與大白鼠的海馬功能相 同;而頂葉皮質則與其他變項如「自我中心的軀幹/頭的方位訊息」有關。至於左側海 馬,雖然也參與移行,但是與右側海馬的功能顯然不同。
然而,實驗室破壞某一區域或暫時使之失去功能,只能研究特定區域之特定功能,
未必能適用於疾病;同理,以 f-MRI 或 PET,也只能說明正常的大腦特定區域功能。
此外,虛擬實境無法得到來自前庭的感覺輸入,而前庭是「路徑整合」是重要的訊息 來源。
從以上動物研究、人類腦傷與功能性造影研究 (見附錄 3, 4, 5) 可得到以下的關 係;即地標再認與舌狀腦迴關係密切,且是認路步驟之第一步;以「自我中心」的認 路則與頂葉相關,「他中心」的認知地圖與路徑整合,則與海馬迴及附近組織相關,
見 Table 2.4。
皮亞傑的認路發展理論
皮亞傑 (Piaget, 1960; Piaget & Inhelder, 1967) 研究孩童認路能力的發展,對空間
認知表徵的建立遵循以下三個步驟,首先是地標再認,接著是「自我中心」的路徑學 習,最後才是「他中心」表徵的習得。地標是指外表顯著之物可以用作為線索者,是 最簡單、也最管用的方法。逐漸地,兒童有串起地標的能力,並以自我為中心去看這 個順序,通常這也是我們在報路給陌生人的方法。例如這樣的陳述:
「往前看到一個紅色的郵筒,然後右轉約 100 公尺,看到紅綠燈後左轉,會碰到 一家便利商店,然後再右轉」。
顯然這是一個以自我為中心,靠著重要地標或結點,依時間出現先後順序串起來 的結果。這種找路的方法有幾個特點,主要是 (1) 步驟的順序紀錄;(2) 植基於以自 我為中心的座標框架上;(3) 以及不太具有彈性。因為這是一個紀錄路徑的線性指引,
所以是脆弱的,只要決定性的地標改變、或改道,便會造成這種習得的路徑失敗。
接著,兒童可以從第三者的角度鳥瞰地上之擺設及相關位置,也可以從前述之歷 程組成一個地圖,類似 Tolman 所說之「認知地圖」,即使看不到目標,但可以指出方 向,雖然路徑曲折,也可以指出從何處出發。同時,地理與地圖的互換,也在這階段 形成。有了這三種能力,不只在熟悉的環境,就連陌生的地方,只要走過一次、或者 兩次,就可以掌握大致的情形,人類更可以利用地圖,即使沒有親自走過,也可以介 紹及討論。
後來的研究,也證實兒童對環境的學習確實如此 (Acredolo, 1977; Allen, Kirasic, Siegel & Herman, 1979; Bremner, 1978; Golledge, Smith, Pellegrino, Doherty & marchall, 1985; Herman, Heins & Cohen, 1987; Garino & McKenzie, 1988),與前述之重要步驟相互 呼應。Sigel 與 White (1975) 也提出以下支持的概念:兒童的認知表徵先有地標之再 認,進而出現依地標出現順序組成的一度空間線性表徵,接著才是由多地標組成的歐 氏幾何多度空間表徵。其他的支持,還有來自 Cohen 與 Schuepfer (1980) 及 Cousins, Siegel & Maxwell (1983) 的研究。此外,Ratcliff (1991) 認為自我中心的編碼
(encoding) 是為他中心編碼的先決要件,他甚至認為視運動 (visuomotor) 定位是以自 我為中心,有視運動問題的人,必然有他中心編碼的問題,反之卻不然。不過,Conning 與 Byrne (1984) 及 Spencer 與 Darvizeh (1981) 的研究卻發現,小孩可能很早便有環境 陳設的歐氏幾何學概念。
皮氏的認知能力發展理論,頗能配合神經系統之逐漸成熟及神經網路之搭建,從 初級感覺皮質、次級聯合區皮質,到海馬的整合中心,這種現象也與前述腦傷病灶及 迷路行為的對應,頗有雷同之處,見 Table 2.1 及 Table 2.4。
逆生 (Retrogenesis) 理論與阿氏症病理變化的契合
AD 可以定義為以失憶為主的失智症,它具有許多神經病理標記,特別是類澱粉 斑塊 (amyloid plaque; AP) 及神經纖維糾結 (neurofibrillary tangle, NFT),最終診斷則 需仰賴病理解剖,看到 AP 與 NFT 有一定之密度與分佈情形。1997 年 NIA 及 Reagan Institute of the Alzheimer’s Association 所定的標準,同等強調 AP 與 NFT。根據兩者在 顯微鏡下的密度,可定出高 (20-30 per 10X field,同時出現在海馬、EC 與新皮質),中 (5-10 per 10X field,出現於新皮質與邊緣系統),低 (<5 per 10X field,且只侷限於邊緣 系) 可能率。目前與臨床症狀相關性最高之病理分期首推 Braak-Braak stages (Braak &
Braak, 1996);同時,由智力測驗之表現與病理分期做相關分析,發現 NFT 要比 AP 來 的好。
德國病理學家 Braak 與 Braak (1996) 認為 AD 之病理變化 NFT 最早出現於 EC,
進而到邊緣系統、額葉底部,最後到達新皮質。除非邊緣系統的 NFT 的負荷到達極高,
否則新皮質看不到 NFT;一但新皮質出現 NFT,代表到達臨床失智的階段。這種分期 幾乎適用於所有病人之進展過程。
何以 NFT 會肇發於 EC 呢?為何又會循著前述的順序擴散呢?這可以利用「可塑 性失敗假說」(plasticity failure hypothesis) 來說明 (Mesulam, 2000)。從基因學的觀點,
雖可支持 AP 的病因學說,但是 AP 之分佈和嚴重度與病情無關,AP 與 NFT 之分佈亦 不一致;反而 NFT 之分佈與嚴重度較能與 AD 病人的臨床表現一致,這也是 Braak-Braak 病理分期之根據基準。
「可塑性失敗假說」大旨如下:人類或動物一生之中,由於新經驗、耗損或受傷,
神經重塑 (neuroplasticity) 幾乎隨時都在進行,這過程在成人的大腦包含樹突分支改 變、胞突接合重整、長期增益、軸突發芽、神經膠細胞延伸、胞突接合新生及神經細 胞再生。而邊緣與副邊緣系 (paralimbic) 區域是所有感覺訊息匯集之處,是經驗累積
過程最多負荷的地方,亦即「可塑性負荷」最大之處。AD 的起因,可能是由於某種 原因導致可塑性失敗,增加 tau 蛋白質的磷酸化以及一連串的反應,形成 NFT;支持 的論點來自所有已知的 AD 危險因子 (如類澱粉、早老素、ApoE、與老化) 都與增加
「可塑性負荷」有關,於是邊緣與副邊緣系區域成為 AD 病變肇發處。隨著此處神經 資訊之上游來源 (即輸入) 與下游輸出,展開 NFT 擴散之方向、與拓樸分布,這與病 理學及功能性造影學發現,完全一致,見 Table 2.3。
Table 2.3. AD 之病理分期與 NFT 之分布。
Stage B-B Stages Neurofibrillary tangles
Low limbic I and II Nucleus basalis、entorhinal-transentorhinal cortex of the PHG、海馬、杏仁核、顳極皮質等邊緣系統、
nucleus locus coerulues。
High limbic III and IV (MCI)
幾乎所有的邊緣系統、副邊緣系統,視丘、黑質。
新皮質中會出現 NFT 的區域包含扇形腦迴、下顳 葉與靠中線的顳葉,但為數稀少。前額葉與後頂葉 及枕葉,都還沒有 NFT。
Low neocortical
V (AD) 新皮質扇形腦迴、下及中線側顳迴,開始向中及顳 迴、前額葉與後頂葉皮質聯合區擴展;初級運動及 感覺皮質,只有極少數。
High neocortical
VI (AD) 幾乎所有的皮質聯合區及臥核、olfactory
tubercles。初級運動及感覺皮質,只有少數的 NFT。
Note: BB: Braak-Braak
Table 2.3. 描述 AD 病理變化與病人的臨床表現,詳細的說明如下。
Low limbic stage (相當於 Braak-Braak Stages I and II)
NFT 只零星分布於 nucleus basalis、entorhinal-transentorhinal cortex of the
parahippocampal gyrus、海馬、杏仁核及顳極皮質等邊緣系統。除此之外,在 nucleus locus coerulues 也開始出現 NFT。這期的變化可見於 60 歲以上所有的非失智老人,應當視 為正常老化的結果,神經心理學測驗可出現「因年齡所造成」的較差表現。雖然有人 還是強調 NFT 的出現即屬不正常,只是此期的分佈是局部性 (endemic)。
High limbic stage (相當於 Braak-Braak Stages III and IV)
幾乎所有的邊緣系統都出現 NFT 之分布,甚至是副邊緣系統,同時還有視丘及黑 質。新皮質中會出現 NFT 的區域只有扇形腦迴、下顳葉與中線側 (medial) 顳葉,但 為數稀少,這情形好像是從附近的 PHG 擴散開來一般。此期的前額葉、後頂葉及枕 葉,都還沒有看到 NFT。這階段的老人日常生活還可自理,但神經心理學測驗已開始 出現異常,尤其是記憶。病人可能出現些微認知功能退化或人格改變,但不致於被診 斷為失智,相當於 MCI 或 AD 前驅。
Low neocortical stage (相當於 Braak-Braak Stage V)
前期僅少數 NFT 的新皮質扇形腦迴、下及中線側顳葉腦迴,如今已出現為數眾多 的 NFT,並且開始向中及顳迴、前額葉與後頂葉聯合區擴展,至於初級運動及感覺皮 質,仍然只有極少數的 NFT。病人的表現是早期的失智,尤其是記憶力為主的障礙,
判斷力變差,認路障礙越來越明顯,無法完成一長串的動作或計劃,聽理解的能力也 開始出現問題,其他的認知功能逐漸敗壞。病人一再重覆話語、忘記他人名字,而且 常錯置個人物品。近期記憶差,但久遠的記憶、或高度情緒相關的近程記憶,還算良 好。病人可以靠索引或選擇項選出正確的答案,進一步施以神經心理學測驗,可發現 其他如複雜的注意力、命名、理解、視空間能力,多少也有影響。病人可能會起伏不 定,但如果有病識感病人,會陷入憂鬱狀態。病人與先前的職業、社交與娛樂逐漸脫 節,病人顯得懶散、沒鬥志,病人也開始出現對飲食與性愛的需求減弱。一開始,病
人尚可獨立生活,可以打掃房子、付款、開車、打牌、參與所有社交活動,甚至是較 複雜的專業活動,後者常常是因為有其他專業助理的協助保護下,但是病人已顯得沒 效率、敷衍、果決不斷,而且經常需人從旁協助。接著,病人開始出現語言、理解、
空間定向感、執行功能的障礙,日常生活已明顯受影響,失憶日趨嚴重,到無法再認 的地步,連幾分鐘之前發生的事情,一樣也記不起來。注意力的障礙,壞到無法持續 一段思路、無法執行逐步完成的動作或目標。失語症愈發明顯,影響溝通,多半是流 利型的失語症。病人判斷力大降,沒有病識感,而變得無所謂、不在乎。病人已無法 煮食、持家、付款或開車。後來,病人開始出現醒睡節律混亂及睡眠障礙,也常出現 日落 (sunset) 現象,個人衛生變差,精神症狀逐漸明顯,如妄想 (尤其是不貞及被偷 忘想)、幻覺、激動、收集東西、打鬥等行為,病人變得非常依賴配偶或重要的照顧者。
High neocortical stage (相當於 Braak-Braak Stage VI)
幾乎所有的聯合區以及臥核、olfactory tubercles 都可看到高密度的 NFT。值此階 段,病人所有的認知功能都出現問題,如語言、視覺空間技巧、推理及社交禮儀。病 人開始出現大小便失禁,認不出家人,連走動或吃東西都有困難,幾乎所有的認知障 礙、精神症狀與舉止行為障礙都會出現。病人可能開始有錐體外徵候,如肌躍動、僵 硬、面無表情、步態不穩等,因為黑質及紋狀體也有 NFT 的大量出現。值得的注意的 是初級運動及感覺皮質,仍然只有少數的 NFT,即使是重度的 AD 病人,其基本的運 動及感覺仍然保有一定的功能。
Braak 與 Braak (1996) 之病理分期也可對應到與認路有關的解剖結構,即先影響 海馬,進而頂葉,最後是枕葉之舌狀腦迴,見 Table 2.4。
不論從病理變化、腦血流研究、或者影像學,都顯示 AD 的病灶開始於顳葉的 EC 及 SC,逐漸向邊緣系統及新皮質延伸拓展。從 AD 病人的症狀學看來,病人認知功能 與生活能力的逐漸喪失,恰好與學習發展的順序相反,即晚來早去,這個理論稱為逆 生 (retrogenesis) (Reisberg, Kenowsky, Franssen, Stefanie & Souren, 1999)。Retrogenesis 說明 AD 病人的日常功能、使用能力及飲食能力的變遷與發展順序呈反相關。除此之 外,AD 病人的退化恰好與發展順序相反的情形,尚可見於語言與認知能力、大腦葡
萄糖代謝、腦波變化與神經反射,以及前述的神經解剖病理 (Braak & Braak, 1996)。
與人類生活息息相關的認路行為,在 AD 進展時,是否也遵循 retrogensis 理論呢?
此乃本研究中心議題之一,見 Table 2.4。
輕度認知障礙是一個介於正常者與阿氏症的中間地帶
AD 是一個大腦連續退化的過程,在病人尚未發展成 AD 之前,會有一段處於「輕 度認知障礙」(mild cognitive impairment;MCI) 的階段 (Petersen, 1999)。MCI 的病人 中有很高的比例後來演變成失智,因而被認為與正常老化所帶來之記憶力退步
(age-associated memory impairment; AAMI) (Crook, Bartus, Gerris, Whitehouse, Cohen &
Gershon1986) 不同。由於與 AD 關連性極大的病理變化 AP 及 NFT,可見於正常老人 (Braak & Braak, 1996; Price & Morris, 1999),代表這種病理過程在真正的 AD 症狀表現 出來之前,已經開始進行,MCI 的病人是有發生迷路的可能。
研究目的與假設
本研究目的有三,a) 解決臨床診斷的疑惑、b) 驗證行為神經科學的假說及 c) 發 現早期診斷失智症的工具。
首先,臨床診療中經常可發現失智病人的走失,甚至導致嚴重的意外,顯示研究 有其重要性。其次,學習的過程如結網築塔,後來的連結需靠既存的網狀結構 (Mesulam, 1998; Mesulam, 2000),若將這樣的概念應用於認路,應可驗證後述之假說;最後,由於 目前治療 AD 的藥物成效不錯,除了能改善病人日常生活能力、行為障礙及認知外,也具有 延緩病程惡化的效果,本研究利用有效的工具以期早期發現病人,進而接受治療。
本研究的假說是,就認路行為能力而言,從正常人的狀態、經由 AD 前驅狀態 (MCI)、進入早期 AD,病人首先會出現「他中心」的認路異常,接著是「自我中心」
之認路失能,最後才是失去學習地標的能力。此假說建立在發展心理學與 retrogenesis 的理論架構上,且能配合 Braak-Braak 病理分期和腦傷的研究,見 Table 2.4。此假說可 稱為「認路功能退化階段假說」(retrogenetic hypothesis of wayfinding function)。
Table 2.4. 「認路功能退化階段假說」簡圖。
認路步驟 地標學習 自我中心認路 他中心認路
發展心理學
神經解剖 舌狀腦迴 後頂葉 海馬
病理變化 阿氏症
同時,本假說預測不同階段的病人在認路行為的表現不同,MCI 病人在「地標學 習」上應接近於正常者,在「他中心」的認路學習則應似 AD 病人,而「自我中心認 路」應是三組分開,見 Table 2.5。
Table 2.5. 預測三組受測者的結果。
地標學習 自我中心認路 他中心認路
AD X XX XXX
MCI O X XX
正常者 O O O
Note: O: normal; X: abnormal.
第三章 方法論
研究對象
受測者分為正常者、MCI 及輕度 AD 病人。病人來自成大醫院神經部行為神經學 特別門診,此門診創始於 1993 年 5 月,每月均有來診病人分類統計,過去一季規則來 診者,計有 AD 病人 96 人,主訴記憶力障礙 (含 MCI) 25 人,血管性失智症 46 人,
巴金森氏症病人 39 人。正常受測者,則選擇病人之配偶,或年齡、教育、及生活環境 相似之配對。正常者,MCI 及輕度 AD 病人,每組各 30 人,共計 90 人。
AD 病人的診斷根據 NINCDS-ADRDA (National Institute of Neurological and Communicative Disorders and Stroke-Alzheimer's Disease and Related Disorders
Association, Mckhann, Drachman, Folstein, Katzman, Price & Stadlan, 1984) 準則 (附錄 1) 及 DSM-IV 準則 (APA, 1994) (附錄 2);MCI 則依據 Petersen (1999) 之診斷依據 (附 錄 CogMap 24)。AD 嚴重度以臨床失智評量 (Clinical Dementia Rating Scale, CDR) (Hughes, Berg, Danziger, Coben & Martin, 1982) 分級 (附錄 CogMap 15);CDR 評估項 目包括記憶 (memory)、定向感 (orientation)、判斷及解決問題 (judgment and problem solving ability)、社區活動能力 (social skill)、家居嗜好 (habit and hobbies) 及自我照料 (self-care),其判定依據原文之準則;MCI 病人的 CDR 等於 0.5,輕度 AD 病人的 CDR 等於 1.0。臨床診斷由同一位神經科專科醫師完成。
神經心理學測驗包含 Consortium to Establish a Registry for Alzheimer's Disease (CERAD, Morris, Heyman, Mohs et al, 1989) 神經心理學測驗 [中文版由劉景寬醫師等 人翻譯] (附錄 CogMap 3-14)、認知能力篩檢 (Cognitive Ability Screening Instrument, CASI, Teng, Hasegawa, Homma et al, 1994) (附錄 CogMap 19-23),其中 CERAD 神經心 理學測驗包含項目有語言流暢 (Verbal fluency)、波士頓命名測驗 (Bonston Naming Test)、物件命名 (Object Naming)、語言記憶之登錄 (Registration of verbal memory)、
語言記憶之回憶 (Recall of verbal memory)、語言記憶之再認 (Recogniton of verbal memory)、視空間能力 (Visualspatial ability)、視空間記憶之再認 (Recognition for visualspatial memory)、及連線測驗 (Trail Making test)。
另外,為了解病人的認路行為與認路現狀,也設計了一個問卷,對象是受測者家 屬 (附錄 CogMap 25)。所有實驗室檢查及造影術,均於成大醫學中心完成。所有神經 心理學評估、問卷及實驗,由同一位施測者完成。
為了避免與主題無關之干擾因素,設立排除條件如下:1) 病人有明顯語言問題,
無法理解指導語者;2) 有心肺功能障礙或骨骼關節系統問題,無法連續行走 1000 公 尺者;3) 基本感覺器官嚴重障礙者,例如眼、耳等機能;4) 無家屬陪同前來者;5) 病 人有明顯憂鬱症狀或其他精神症狀,有影響認知功能之可能者。
病人及家屬經詳細說明後,簽署同意書;完成者,可領取酬金新台幣 500 元整 (NSC 計劃補助)。
研究工具及施行程序
實驗一 地標再認 Landmark recognition
路景或地標之自由回憶 (free recall) 在認路上用處不大,除非是要描繪或敘述到訪 之地;地標或路景的再認反而較為重要,視線真正觸及地標才會激起看過與否的感覺,
因此,本研究利用「地標再認」檢驗,較符合實際認路的行為。然而,地標的存在、
被選取、乃至於在腦中形成表徵,與個人的經驗極為相關,這造成研究施行上之困難,
利用陌生 (novel) 的建築物重新學習與再認,可提供較為客觀的訊息。
本實驗以具有地標作用的建築物照片測驗病人之再認能力,見 Figure 3.1。此等照 片避免非專一性的地點,如十字路口、尋常小學等;避免人潮而賦予的特徵,如大賣 場、市場;避免個人特殊經驗有關者;以及避免熟悉的地點,如臺南車站、政府機關,
警察局等。部份來源為文建會「百年台灣攝影史料整理」。


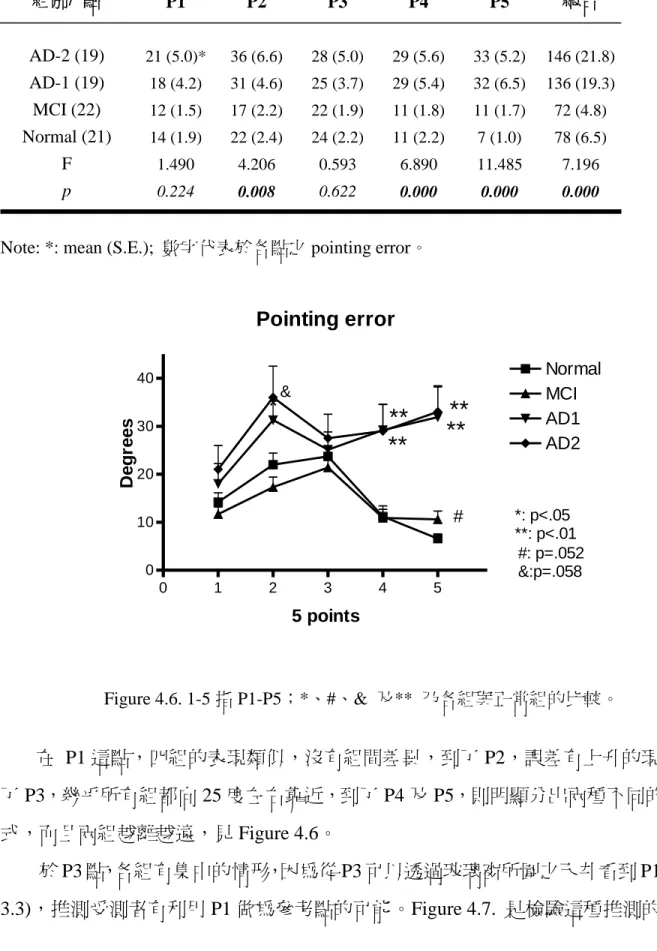
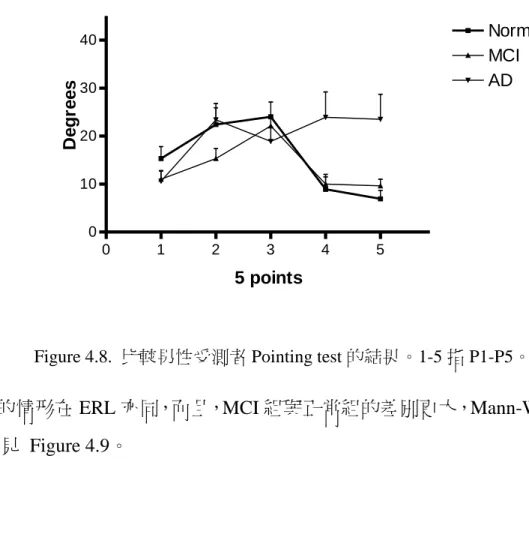
![Figure 4.5. Pointing test 的原始結果,其中 1-5 指要求指出出發點方位的五個點,粗線 表示正確角度。 四組於各點之組間比較如下,AD2 在 P2 [ t (23) = 1.99, p = .058] (邊際效應)、P4 [ t (24) = 2.944, p = .007] 及 P5 [ t (19) = 4.925, p = .000] 三點,均劣於正常組;AD1 於 P4 [ t (24) = 3.07, p = .005]、P5 [ t (19) = 3.815,](https://thumb-ap.123doks.com/thumbv2/9libinfo/9328534.536617/47.891.240.644.360.640/FigurePointing原始結果其中指要求指出出發點方位五個點粗表示正確三點均.webp)