國立臺灣大學生命科學院生化科技學系 博士論文
Department or Graduate Institute of Biochemical Science & Technology College of Life Science
National Taiwan University Doctoral Dissertation
基因改造抗胡瓜嵌紋病毒番茄安全性與 非預期效應評估之研究
Study on the Safety Assessment and Unintended Effects of Genetically Modified Cucumber Mosaic Virus Resistant
Tomato
林志輝 Chih-Hui Lin
指導教授:潘子明 博士
謝 誌
在許多人的支持與協助之下,這本論文終於完成了。其中尤其感謝我的指導教授潘子明 博士在學業實驗上的全力支持。感謝老師給予我的歷練和對於實驗工作的全力支持,也感謝 老師對於我這個喜歡異想天開,沒事弄東弄西的怪學生總是採取鼓勵包容的態度,放手讓我 可以去探索新的領域。潘老師在我的博士班生涯不知道該如何走下去時,不吝伸出援手指引 方向。如果沒有老師,今天就不會有這本論文的誕生,對於老師的指導與照顧萬分感謝。
本論文在資格考、預口試及口試期間承蒙蘇遠志教授、黃鵬林教授、徐源泰教授、方繼 教授、陳良築教授、鄭石通教授、陳明汝教授、廖啟程博士、朱文深博士與柳建安博士的審 查,感謝各位教授對於本論文的悉心指導。
感謝一路走來在實驗工作上熱心幫助我的許多人,首先感謝好友宗偉在實驗室生活的幫 助與支持,沒有你我大概博士班唸不完了。感謝已經退休的亞洲蔬菜中心孫宏吉先生在採樣 工作上的協助,同時讓我學習了許多種植番茄的知識。感謝吳素心小姐在行政工作方面的支 持與教導,感謝蔡宗佑學長在學業與研究生生涯的照顧與鼓勵,也感謝潤榮學長在實驗以及 行政工作上的支援。在這裡也感謝實驗室的伙伴們,和我一起渡過了那段難忘的日子的承先、
俊帆、貞幼、瑋盈、大頭、小猴和侯董。而俊霖、瑞蘭、建德、立涵、俊傑、小惠、汶佩、
俊賢、小路、楷謙、小黑、珮祺、超人、大牛、小花、佳穎、阿包、伊絢還有姿杏,你們的 歡笑讓實驗室生活變得有趣。建利感謝你願意承接這個重擔,讓我在快畢業的時候可以有比 較多的時間整理論文。
最後必須感謝我的父母,在一個其實不是很清楚我每天做了哪些事的情況下,很諒解的 支持我完成漫長的學業,感謝妹妹與姿杏在精神上的完全支持,家人們是我每天努力的原動 力,謹將此論文獻給你們。
林志輝 謹誌於國立臺灣大學 中華民國九十九年八月
目 錄
中文摘要………XI 英文摘要………..……….XII 縮寫表………..………..………….IX 第一章 緒論
1-1 基因改造生物(genetically modified organisms, GMO) 的定義………...1
1-2 基因改造作物現況………..1
1-3 基改作物安全性評估………..3
1-4 基改作物的管理與檢測……….5
1-5 基改作物的非預期效應………8
1-6 基改抗胡瓜嵌紋病毒番茄………11
1-7 研究架構與研究目的………12
第二章 基改抗胡瓜嵌紋病毒番茄過敏性評估 2-1 基改抗胡瓜嵌紋病毒番茄安全性評估研究計畫………15
2-2 基改抗胡瓜嵌紋病毒番茄基本資料………15
(一) 「基因改造植物」之描述………..………15
(二) 宿主植物及其食物用途………..….………15
1. 俗名、學名及分類學資訊………..………15
2. 育種栽培及發展之歷史,特別是對有礙人體健康之性狀說明………16
3. 與宿主植物安全性相關之基因型及表現型資訊……….…16
a. 毒性………..…16
b. 過敏誘發性………..…16
c. 宿主類緣種與對宿主植物之遺傳背景具顯著貢獻之植物與安全性相關之 基因型及表現型………..16
4. 安全食用歷史……….….18
a. 傳統育種、運輸及貯存的方法………18
b. 安全食用所需之特殊加工方法………..…18
c. 在一般膳食中所扮演的角色………..…18
(三) 基因提供生物(可包括類緣種) 之描述………..……….…18
c. 中間宿主………20
2. 提供欲轉殖入宿主植物之 DNA 的相關資訊………..……20
a. 所有遺傳物質之特徵……….…20
b. 大小及辨識特徵………23
c. 於最終重組體中的位置及方向及功能………..…23
(五) 基因改造之特徵……….…23
1. 嵌入植物基因體之轉殖 DNA 的相關資訊……….…23
2. 提供基因改造植物體內任何新表現物質之資料………23
3. 其他必要資料………..…23
2-3 材料與方法………....23
(一) 基改抗胡瓜嵌紋病毒番茄過敏誘發性評估………23
1 . 探 討 基 因 來 源 … … … . . … … … . . … … 2 3 2. 胺基酸序列過敏原資料庫搜尋比對………..……24
3. 基改番茄中轉殖基因表現量分析………..……24
a. 胡瓜嵌紋病毒鞘蛋白基因序列選殖與確認……….……26
b. 基改番茄中轉殖基因於不同生長時期的表現量……….……26
(1) 番茄樣品……….26
(2) Total RNA 製備與純化………...……….26
(3) RNA 定量………28
(4) 反轉錄即時定量 PCR 分析 (RT-qPCR) ……….28
(a) cDNA 合成………..28
( b ) 即 時 定 量 P C R 引 子 設 計 … … … 2 8 (c) 即時定量 PCR 反應條件……….30
(d) 基因表現量的計算方式………30
4. 重組胡瓜嵌紋病毒鞘蛋白生產………32
a. 重組大腸桿菌培養與誘導生產胡瓜嵌紋病毒鞘蛋白….………32
b. rCMVCP 的純化與確認………...………33
(1) rCMVCP 的純化…….………...…33
(2) SDS-PAGE、Western、N 端定序以及 LC/MS/MS 分析………33
5 . 模 擬 胃 液 消 化 試 驗 … … … . . 3 5 2-4 結果………..………..36
(一) 蛋白質來源、基因表現量與蛋白質表現量分析………..36
1. 蛋白質來源與過敏誘發性相關資料………..36
2. 新表現蛋白質的含量及表現位置………..…..36
(二) 序列搜尋比對………..………..36
(三) rCMV cp 生產純化………..………..36
3-1 前言……….………..………..47
3-2 材料與方法……….………..……….…..51
(一) 以 GenomeWalkerTM 試劑套組選殖轉殖基因鄰近序列….……….…..51
1. Genomic DNA 之製備與純化….……….51
2. 基改抗胡瓜嵌紋病毒番茄轉殖基因定序….……….52
3. 選殖轉殖基因鄰近序列.……….52
(二) 以 Inverse PCR 選殖轉殖基因鄰近序列.………..56
(三) 以 TAIL-PCR 選殖轉殖基因 3’ 與 5’ 端序列.……….58
(四) 選殖片段的確認.………..58
(五) 即時定量 PCR 檢測系統的品系專一性引子設計及確認.……….………….65
(六) 即時定量 PCR 檢測系統的建立.………..…….65
1. 探 針 設 計 與 內 源 性 基 因 的 選 擇. … … … . 6 5 2. 參考質體的構築與標準曲線的建立.……….65
3. 即時定量 PCR 反應條件.………..69
4. 系檢測統建立與確認.……….69
(七) 基改番茄 R8 番茄果實的模擬胃液消化試驗.………73
3-3 結果.………...73
(一) 轉殖基因鄰近序列選殖與確認.………...73
(二) 即時定量 PCR 檢測系統建立.……….74
(三) 基改番茄 R8 番茄果實的模擬胃液消化試驗.……….82
3-4 討論.………...88
3-5 結論………...90
第四章 基改抗胡瓜嵌紋病毒番茄對於土壤微生物菌相影響評估 4-1 前言.……….91
(一) 基改作物的基因水平轉移現象.………...91
(二) 土壤微生物菌相與植物.………...92
(三) 變性梯度膠體電泳 (denaturing gradient gel electrophoresis, DGGE) .………. 94
(四) 評估策略.……….95
4-2 材料與方法.………..……….98
(一) 基改番茄試驗田與土壤樣本處理.……….98
(二) 土壤基本性質與微生物計數分析.……….101
(三) PCR-DGGE 分析條件.……….103
1. 土壤 DNA 抽取.……….……….103
2. PCR 條件.……….………….……….103
第五章 以蛋白質二維電泳、cDNA 微陣列及反轉錄-即時定量聚合酶鏈反應進行基改抗胡瓜 嵌紋病毒番茄非預期效應的評估
5-1 前言.………125
5-2 材料與方法.……….…125
(一) 植物樣品.……….125
(二) Total RNA 抽取.……….125
(三) RNA 樣本處理與 cDNA 微陣列晶片製備方法.……….130
(四) RNA 品質確認及濃度測定.………..………….130
(五) cDNA 微陣列結果分析.……….……….131
(六) RT-qPCR 確認基因表現.………..…….131
1. RT-qPCR 分析條件.……….….132
(七) 蛋白質樣品抽取.………..…….132
1. rehydration buffer 配製.……….……….132
2. 蛋白質樣品抽取.……….132
(八) 等電集焦電泳.………..……….134
(九) 第二維電泳分析.……….……….134
1. Blue Silver Stain 染色劑配製.………..134
2. 第二維蛋白質電泳與影像分析.……….135
3. 蛋白質質譜分析鑑定 .………..…….135
4. mRNA 表現量確認.……….136
(十) 轉殖基因嵌入位置鄰近基因的表現量分析.……….138
(十一) 花芽分化相關基因的表現量分析.……….138
5-3 結果.………..……….138
(一) RNA 與 cDNA 微陣列品質確認.………...……….138
(二) cDNA 微陣列分析結果.……….144
(三) 以 RT-qPCR 確認 cDNA 微陣列結果.……….144
(四) 蛋白質二維電泳分析結果.……….151
(五) 轉基因嵌入鄰近基因表現.……….…….160
(六) 花芽分化基因的表現量分析.……….165
5-4 討論.……….170
5-5 結論………178
總結………..………179
參考文獻………181
附錄………..………198
附錄一 基因改造食品之安全性評估方法修正草案.……….198
附錄二 「混合型基因改造食品」安全性評估原則.……….……….223
圖 目 錄
圖 1-1、1996 至 2008 年間全球基改作物種植面積(單位:百萬公頃) ………..2
圖 1-2、基改作物 PCR 檢測引子的四種設計方式………..7
圖 1-3、本論文研究架構……….14
圖 2-1、質體 pBI121 (GenBank AF48578) ………..21
圖 2-2、基改番茄 R8 的轉基因構築……….22
圖 2-3、基改番茄 R8 的胡瓜嵌紋病毒鞘蛋白基因與胺基酸序列………..25
圖 2-4、番茄果實成熟度的六個判定標準……..………27
圖2-5、即時定量 PCR 放大曲線與 Ct 值的計算……..………...31
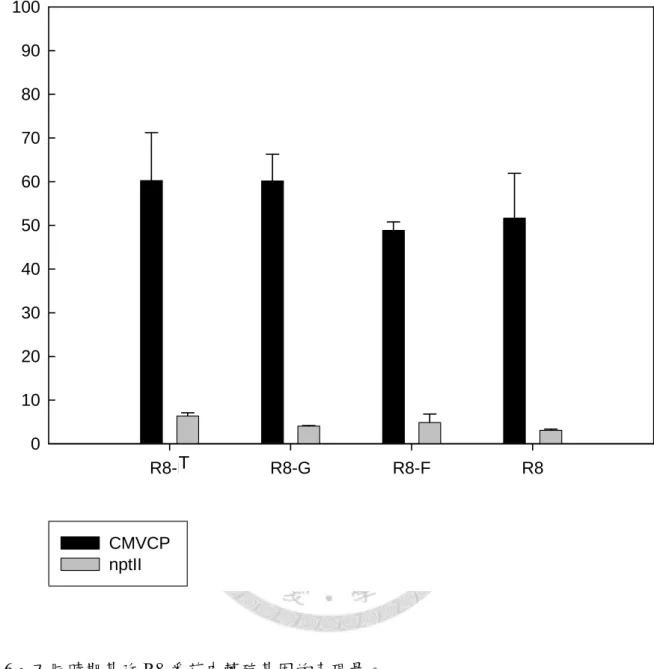
圖 2-6、不同時期基改 R8 番茄中轉基因的表現量……..……….37
圖2-7、始花時期基改 R8 番茄中 cmvcp 表現量的 REST 軟體統計分析結果………..………38
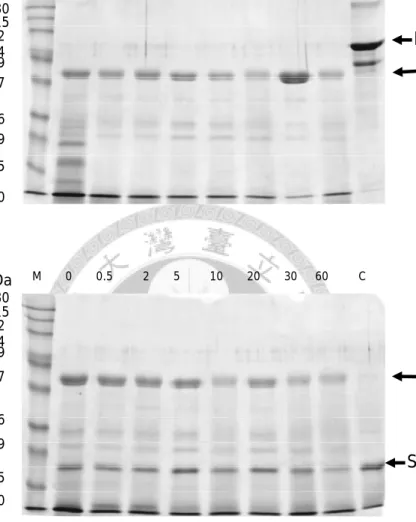
圖2-8、重組胡瓜嵌紋病毒鞘蛋白純化………42
圖 2-9、重組胡瓜嵌紋病毒鞘蛋白 LC/MS/MS 與 N 端定序分析……….45
圖 2-10、模擬胃液正負控制組試驗結果………43
圖 2-11、重組胡瓜嵌紋病毒鞘蛋白模擬胃液消化試驗結果………44
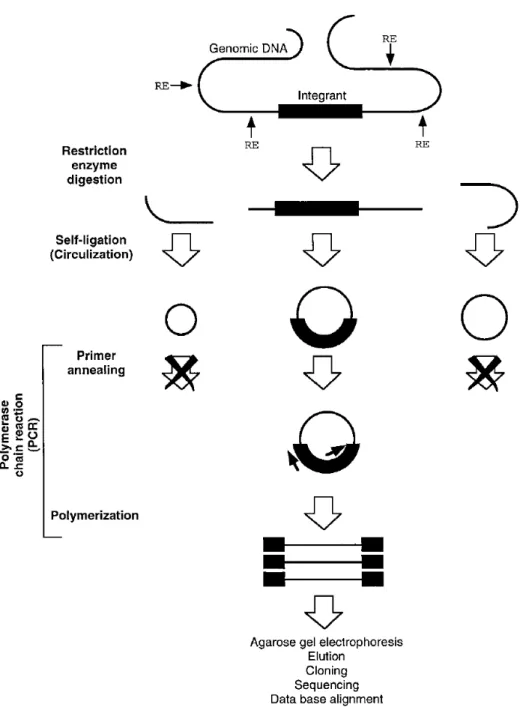
圖 3-1、GenomeWalkerTM 原理簡圖(http://www.clonetech.com) ...48
圖 3-2、Inverse PCR 原理簡圖………49
圖 3-3、TAIL-PCR 原理簡圖………50
圖3-4、抗胡瓜嵌紋病毒番茄轉基因片段圖譜………...………….54
圖3-5、GenomeWalkerTM 試劑套組 PCR 產物電泳分析………..……….57
圖3-6、Inverse PCR 產物電泳分析………59
圖3-7、TAIL-PCR 產物電泳分析………..……….62
圖3-8、三種不同的選殖方法所產生的最終 PCR 產物電泳分析………..63
圖3-9、Inverse PCR 產物 DNA 序列(#111) ……….66
圖3-10、基改抗胡瓜嵌紋病毒 R8 番茄的轉基因以及 flanking sequence………….………67
圖 3-11. 品系專一性引子測試……….70
圖3-12、TA cloning 質體 pyT&A 圖譜……….………71
圖3-13、SYBR Green 與 TaqMan/MGB 即時定量 PCR 原理簡圖……….……75
圖3-14、品系專一性 TaqMan 即時定量聚合酶鏈反應定量系統放大曲線和標準曲線…….…76
圖3-15、番茄內源性基因 TaqMan 即時定量聚合酶鏈反應定量系統放大曲線和標準曲線.…77 圖3-16、品系專一性 SYBR Green 即時定量 PCR 定量系統定量曲線………83
圖3-17、品系專一性 TaqMan 即時定量 PCR 定量系統定量曲線………84
圖3-18、番茄果實 DNA 於模擬消化液中的分解………..85
圖4-9、畦部土壤樣品的 DGGE profile PCA 分析結果………...……….114
圖4-10、溝部土壤樣品的 DGGE profile PCA 分析結果………..………115
圖4-11、土壤樣品 DGGE profile cluster 分析結果……….117
圖4-12、土壤樣品 DGGE profile cluster 分析結果……….118
圖4-13、部分土壤樣品間 DGGE profile 的相似性………..119
圖4-14、以合併畦部及溝部土壤 16S rRNA gene 及 ammonium-oxidizing bacteria 的 DGG E profile 進行 PCA 以及叢集分析結果………...………..120
圖5-1、本研究所採取非預期效應評估的策略……….129
圖5-2、本研究的評估非預期效應的實驗設計方向……….132
圖5-3、AK322797.1 基因與轉基因序列的相對位置………..139
圖5-4、阿拉伯芥的花芽分化基因調控………..…………...……….141
圖 5-5、與花芽分化相關的基因突變株番茄莖的生長與構成………142
圖5-6、部分 RNA 樣品的 quality check 結果……….145
圖5-7、Affymetrix 番茄 cDNA 微陣列原始影像……….146
圖5-8、四至六片真葉時期的番茄 cDNA 微陣列分析結果………148
圖5-9、四至六片真葉時期的番茄 cDNA 微陣列的叢集分析結果………..149
圖 5-10、四至六片真葉時期的特定番茄基因表現量分析比較………..153
圖 5-11、果實採收時期的特定番茄基因表現量分析比較………..154
圖5-12、四至六片真葉時期的番茄二維電泳圖譜………156
圖5-13、開花、綠色果實與果實收成時期的番茄二維電泳圖譜……….157
圖5-14、四至六片真葉時期的番茄二維電泳圖譜中具有顯著差異的蛋白質……..………….158
圖5-15、始花時期的番茄二維電泳圖譜中具有顯著差異的蛋白質………159
圖5-16、綠果時期的番茄二維電泳圖譜中具有顯著差異的蛋白質………161
圖5-17、果實採收時期的番茄二維電泳圖譜中具有顯著差異的蛋白質………162
圖5-18、AK322 於不同生長時期的番茄中表現量的比較………...164
圖5-19、4-6 片真葉時期基改與非基改番茄開花分化相關基因表現量比較……….166
圖5-20、始花時期基改與非基改番茄開花分化相關基因表現量比較………167
圖5-21、綠果時期基改與非基改番茄開花分化相關基因表現量比較………168
圖5-22、果實採收時期基改與非基改番茄開花分化相關基因表現量比較………169
圖5-23、參與光合作用的基因與代謝路徑…..……….174
圖5-24、參與固碳作用的基因與代謝路徑……..……….175
表 目 錄
表2-1、已知的番茄過敏原………17
表2-2、轉基因與 housekeeping 基因表現測定用引子………29
表2-3、胡瓜嵌紋病毒鞘蛋白的過敏原資料庫搜尋比對結果………39
表3-1、基改抗胡瓜嵌紋病毒番茄轉基因放大選殖引子...53
表3-2、Flanking sequence 選殖所用的探針及引子...………55
表3-3、第一輪 TAIL-PCR 條件………60
表3-4、第二及第三輪 TAIL-PCR 條件。………..………61
表3-5、以三種不同選殖方法所得部分 PCR 產物序列進行 Blastn 比對分析結果………..64
表3-6、即時定量聚合酶鏈反應所用的探針及引子……….……….………..68
表3-7、品系專一性 SYBR Green I 即時定量聚合酶鏈反應放大參考質體標準曲線的重復性 及再現性……….78
表3-8、內源性基因 SYBR Green 即時定量聚合酶鏈反應放大參考質體標準曲線的重復性 及再現性………...79
表3-9、品系專一性 SYBR Green 即時定量聚合酶鏈反應定量系統的重復性及再現性…….80
表3-10、品系專一性 TaqMan 即時定量聚合酶鏈反應定量系統的重復性及再現性………….87
表 4-1、基改植物基因水平轉移至細菌的部分實驗室與田間研究...93
表4-2、於 DGGE 研究中所使用的引子………104
表4-3、土壤特性分析結果………..109
表4-4、土壤樣品微生物菌相的 Shannon-Wiener Diversity Index………..109
表4-5、基改作物對於土壤微生物菌相影響研究的相關文獻………..123-124 表5-1、本研究中所使用的番茄品種………..128
表5-2、用來確認番茄 cDNA 微陣列結果的基因定量用引子………..133
表5-3、用來確認蛋白二維電泳結果的基因定量用引子……….137
表5-4、番茄花芽分化相關基因定量用引子……….143
表5-5、微陣列控制組探針 3 端與 5 端比值………..147
表5-6、果實收成時期的番茄 cDNA 微陣列基因表現分析結果……….150
表5-7、進行 RT-qPCR 表現量測定的番茄基因………152
表5-8、以 RT-qPCR 確認番茄基因表現 cDNA 微陣列分析結果………...155
表5-9、蛋白質二維電泳分析中具有差異的蛋白質點………..163
表5-10、基改與非基改番茄在不同生長時期花芽分化相關基因表現量的差異………175
縮寫表
ADFS (Allergen Database for Food Safety):食品安全過敏原資料庫 APEC (Asia-Pacific Economic Cooperation):亞太經濟合作組織
ARDRA (Amplified ribosomal DNA restriction analysis):核糖體去氧核糖核酸擴增限制酶分析 AVRDC (Asian Vegetable Research and development centre):亞洲蔬菜研究發展中心
BA (6-Benzylaminopurine):苯基胺基嘌呤
BLAST (Basic local alignment search tool):基礎本域比對搜尋工具 BSA (Bovine serum albumin):牛血清白蛋白
CaMV 35S (Cauliflower Mosaic Virus 35S promoter):花椰菜嵌紋病毒 35S 啟動子 CAPS (N-cyclohexyl-3-aminopropanesulfonic acid):3-(環己胺)-1-丙磺酸
CBD (Convention on Biological Diversity):生物多樣性公約 CBR (Coomassie Blue R-250):考馬斯亮藍 R-250
cDNA (Complementary DNA):互補 DNA
CLPP (Community level physiological profiles):族群生理圖譜 CMV (Cucumber Mosaic Virus):胡瓜嵌紋病毒
CP-MR (Coat protein-mediated resistance):鞘蛋白衍生抗性 Ct (cycle threshold):增幅循環閥值
CTAB (Cetyltrimethylammonium bromide):十六烷基三甲基銨溴化物 DEPC (Diethyl pyrocarbonate):二乙基焦碳酸
DGGE (Denaturing gradient gel electrophoresis):變性梯度膠體電泳 DNA (Deoxynucleic acid):去氧核糖核酸
DRT (degenerated random tagged):退化隨機標示 DTT (Dithiothreitol):二硫代蘇糖醇
EDTA (Ethylenediaminetetraacetic acid):乙烯二胺四乙酸 EEA (European Environment Agency):歐洲環境局
ELISA (Enzyme-linked immunosorbent assay):酵素結合免疫吸附分析 EtBr (Ethidium bromide):溴化乙錠
FAME (Fatty acid methyl ester analysis):脂肪酸甲酯分析
FAO (Food and Agriculture Organization):聯合國糧食暨農業組織 FISH (Fluorescence in situ hybridization):原位螢光雜交
FRET (fluorescence resonance energy transfer):螢光共振能量轉換 FT-IR (Fourier transform infrared spectroscopy):傅立葉轉換紅外線光譜 GMO (Genetically modified organisms):基因改造生物
IAA (Indole-3-acetic acid):3-引朵醋酸
LC/MS/MS (Liquid chromatography-tandem mass spectrometry):液相層析串聯質譜
PEG (Polyethylene glycol):聚乙二醇
PBS (Phosphate buffered saline):磷酸緩衝生理食鹽水 PLFA (Phospholipid fatty acid analysis):磷脂質分析
PTGS (Post-transcriptional gene silencing):轉錄後基因靜默 PVDF (Polyvinylidene fluoride):聚偏二氟乙烯樹脂
QCM (Quartz crystal microbalance):石英晶體微天平
RFLP (Restriction fragment length polymorphism):限制酵素切割長度多形性 RISA (Ribosomal intergenic spacer analysis):核醣體轉錄間隔區分析
RNA (Ribonucleic acid):核糖核酸
RSDr (Relative standard deviation of repeatability):重覆性相對標準差 RSDR (Relative standard deviation of reproducibility):再現性相對標準差
RT-qPCR (Reverse transcription eal-time quantitative PCR):反轉錄即時定量 PCR SAM (Significant analysis of microarray):微陣列顯著性分析
SDAP (Structural Database of Allergenic Proteins):過敏原蛋白結構資料庫 SDS (Sodium dodecyl sulfate):鈉十二烷基的硫酸鹽
SDS-PAGE (Sodium dodecyl sulfate- polyacrylamide gel electrophoresis):鈉十二烷基的硫酸鹽丙 烯醯胺膠體電泳
SGF (Simulated gastric fluid):模擬胃液
SSCP (Single strand conformation polymorphism):單股構形多形性 STI (Soy trypsin inhibitor):大豆胰蛋白酶抑制因子
TAIL-PCR (The thermal asymmetric interlaced PCR):熱不對稱性交錯 PCR
ICGEB (The International Center for Genetic Engineering and Biotechnology):國際遺傳工程與生 物技術研究中心
T-RFLP (Terminal restriction fragment length polymorphism):端粒限制片段長度多型性分析 ToMV (Tomato MosaicVirus):番茄嵌紋病毒
TSWV (Tomato Spotted Wilt Virus):番茄斑點萎凋病毒 TYLCV (Tomato Yellow Leaf Curl Virus):番茄捲葉病毒
UNEP (United Nations Environment Programme):聯合國世界環保總署
UNIDO (United Nations Industrial Development Organization):聯合國工業發展組織
UPGMA (Unweighted pair-group method using arithmetic averages):平均非加權成對叢集演算法 WHO (World Health Organization):世界衛生組織
WTO (World Trade Organization):世界貿易組織
中文摘要
隨著基改作物的栽種生產日趨普及,以歐盟為首之諸多國家,對於基改作物的安全性與 管制也越來越重視。本研究以亞洲蔬菜發展中心所研發出的國產基改抗胡瓜嵌紋病毒番茄R8 為材料,進行安全性評估以及非預期效應的研究。本研究於第二章首先遵循衛生署所公布「基 因改造食品」之安全評估方法進行過敏性評估。綜合本研究以及其他評估研究計畫的結果,
雖然在農藝性狀與果實總糖含量上有顯著變化,但是沒有實際的安全性風險。第三章進行品 系專一性即時定量聚合酶鏈反應檢測系統的建立,成功地建立具有足夠專一性、定量線性與 靈敏度的檢測方法。此外藉由模擬胃液的分解,證實基改番茄 R8 的果實 DNA 並未有殘留 於消化道的疑慮。第四章進行種植基改番茄 R8 對於土壤微生物菌相的影響評估,以瞭解種 植基改番茄 R8 是否會對於土壤微生物生態帶來衝擊。結果顯示相較於試驗田中的其他環境 因子,種植基改番茄 R8 對於土壤微生物不會顯著的衝擊土壤微生物生態。本研究的最後一 部份為整合 cDNA 微陣列分析與蛋白質二維電泳以及反轉錄-即時定量聚合酶鏈反應技術進 行基改番茄 R8 的非預期效應評估。結果顯示基改番茄 R8 在轉殖過程中的確產生了非預期 的改變,對於植株的生理發育以及果實的糖度可能產生了影響,但是並沒有發現具有安全性 疑慮的非預期改變。在涵蓋性足以將所有的代謝變化進行描述的研究方法出現之前,整合多 個分子層次的非預期效應研究方法有其必要性。綜合本研究結果,國產基改抗胡瓜嵌紋病毒 番茄R8 雖然基改番茄 R8 在農藝性狀、基因表現與蛋白質表現上與非基改番茄 L4783 有部分 具有顯著差異,但是這些改變並未發現明顯的安全性的疑慮。過敏性與毒理學的評估的結果 也顯示基改番茄 R8 可以安全食用。近年來高通量分析技術的快速進步,以及生物分子資料 庫的快速成長,使得有關植物生理代謝機制知識的快速進展。越來越深入的分子層次研究結 果顯示出現行基於「實質等同」的法則所提供的資訊有可能不足以完全評估基改作物的安全 性。雖然目前非預期效應的評估仍因為成本偏高及知識和技術的限制無法普遍實行,評估的 結果也往往無法有清楚而完整的解釋,但對於基改作物的安全性評估來說,仍然具有相當的
Abstract
Following the widespread of genetically modified (GM) crop in the world, many countries such as EU have great concern about GM crop regulation and safety. A transgenic CMV-resistant tomato line R8 which developed by Asian Vegetable Research and Development Center was used to study the safety assessment and unintended effect of GM crop in this research. We start with the conventional safety assessment of GM crop in the chapter 2, and the GM tomato R8 was concluded as safe following the guideline of safety assessment. In the chapter 3 of this research, a real-time PCR detection system of GM tomato R8 was established with feasible specificity, linearity and sensitivity. The digestive fate of tomato fruit DNA was also revealed by simulated gastric fluid (SGF) digestion and real-time polymerase chain reaction analysis. Rapid degradation of tomato fruit DNA in the SGF showed that there is a minimal risk of gene flow. The chapter 4 of this study is the evaluation of the effect of GM tomato on soil microbial communities using denaturing gradient gel electrophoresis. The result of soil microbial analysis showed that the effect of GM tomato R8 was minor than the environmental factors such as soil position. Thus we conclude that GM tomato R8 has no detrimental impact on soil microbial communities. The last part of this study is the evaluation of the unintended effect of GM tomato R8 with proteomic and transcriptomic analysis.
The results of this study have revealed the unintended changes of gene and protein expression in GM tomato R8, which may caused the significant changes of floral development and sugar content of GM tomato R8. In summary, GM tomato R8 did have several unintended changes in the gene and protein expression which may relate to the significant changes of floral development and sugar content. However, these unintended changes do not relate to any significant safety risk. The results of allergenicity and toxicity assessment were also showed no significant safety concern. Due to the complex and uncertain process of plant gene transformation, unintended changes GM crop should always exist. As the rapid development of high-throughput analysis, the unintended effects of GM crops have caused the contradiction of “substantial equivalence” concept used in the GM crop
第一章 緒論
1-1 基因改造生物 (genetically modified organisms,GMO) 的定義
「基因改造生物」係指生物體基因的改變,為使用基因工程或分子生物技術,將遺傳 物質轉移 (或轉殖) 入活細胞或生物體,而非由於天然之交配或天然的重組所產生的; 不 包括傳統育種、細胞及原生質體融合、雜交、誘變、體外受精、體細胞變異及染色體倍增 等技術 (「基因改造食品之安全性評估方法修正草案」,2010) 。
1-2 基因改造作物現況
由於植物基因轉殖技術的逐漸成熟,使得生物產業與大規模農業產生了連結。北美地 區自1990 年代開始快速並大規模的推出基因改造 (或稱轉殖基因、或基因轉殖) 作物,並 迅速的向全球市場擴展。自 1996 年基因改造作物正式開始商業栽培算起,十年間這些作 物已成功席捲全球農業生產。2009 年全球基因改造作物的種植面積已突破 1.34 億公頃 (圖 1-1) ,年成長率 8 % (ISAAA, 2010) 。這種史上少見的接受與推廣速度反映出農業生物 技術應用已是世界趨勢,但也引起了全球消費者對基改作物與其衍生食品的安全顧慮。美 國目前仍然是基改作物最主要的開發及推廣國家,其基改作物栽培面積6 千 4 百萬公頃占 全球總量的48%。2009 年有 15 個種植面積在 50 萬公頃以上的大規模種植國。按基改作物 種植面積排列,分別是美國、阿根廷、巴西、印度、加拿大、中國、巴拉圭、南非、烏拉 圭、玻利維亞、澳洲、布吉納法索、西班牙與墨西哥。
在全球基改作物市場中,耐除草劑基改作物一直占有主導地位,其次是具有抗蟲性狀 的基改作物。目前全球基因改造作物市場主要集中於黃豆、玉米、棉花及油菜等四項作物,
在2009 年基改品種種植比例分別佔該作物全球生產面積之 50%、37.2%、10.5%及 3% 。 種植基改作物農民分佈於25 個國家共 1400 萬人,其中九成以上為資源相對貧乏的開發中 國家農民。根據 Cropnosis 的估算 2009 年基改作物的全球市場總值為 105 億美元,相當 於2009 年全球作物保護市場總值 (522 億美元) 的 20%,以及全球商業種子市場 (340 億
圖1-1、1996 至 2009 年間全球基改作物種植面積 (單位:百萬公頃) (ISAAA, 2010) 。 Fig. 1-1. Global area of biotech corps, 1996 to 2009 (million hectares) .
10 個品系、大豆9 個品系等 (ISAAA, 2010) 。
目前世界各國對於以基改作物進行雜交所產生的混合型基改品系 (stacked traits) 的 評估及管理態度有很大的不同,例如美國及加拿大認為只要親本的基改品系通過審查評 估,即可認定雜交子代亦通過審查,但歐盟及阿根廷則認為混合型基改品系必須重新進行 審查評估。無論如何,混合型基改品系已漸漸的成為目前基改作物市場的主流。以最大的 基改作物生產國-美國為例,種植混合型基改品系作物的面積已達到全國基改作物生產總面 積的41%。
根據研究報告顯示1996 至 2009 年間,基改作物對於全球總體經濟效益為 92 億美元 (開發中國家 47 億美元,工業國家 45 億美元) 。1996 至 2009 年間,因為種植基改作物所 使用殺蟲劑減少的累計量為356 百萬噸的活性成分,相當於降低了 16.1% 因為使用殺蟲 劑對環境作物產生的破壞,也間接降低了不少基改作物對於環境衝擊的爭議。此外基改作 物的大規模種植因為減少殺蟲劑的生產以及噴灑殺蟲劑所需要的機械動力,也直接與間接 的減少二氧化碳排放,估計2008 年因為種植基改作物而減低的二氧化碳排放量約有 1440 萬噸。由於國際原油價格高漲及溫室效應議題受到重視,各國近年紛紛開始生物燃料 (biofuel) 的研發與生產。現行生物燃料主要是以生質柴油 (biodiesel) 及乙醇為主,其中生 質柴油多以油菜、大豆及棕櫚為原料,乙醇則是大多利用甘蔗、玉米或其他澱粉穀類為原 料。由於原料作物中的油菜、玉米及大豆目前均已大規模的應用基改品種進行生產,因此 在不影響地區糧食供應的前提下,生質燃料應該是未來具相當潛力的應用市場。鑒於基改 作物市場快速成長、減低糧食問題以及生質能源需求等議題受到重視,基改作物在可預見 的未來將會持續的快速發展,尤其是開發中國家以及糧食供應不足的國家。
1-3 基改作物安全性評估
隨著基改作物的栽種生產日趨普及,以歐盟為首之諸多國家,對於基改作物的安全 性與管理也越來越重視。現今全球貿易活動日漸頻繁,國際間基因改造作物的商業流通已
則。其中,以 FAO/WHO 的安全性評估決策樹 (FAO/WHO, 2001) 及 Codex 的基因改造 食品風險分析原則 (Codex, 2003) ,是全球主要遵循的依據。聯合國下生物多樣性公約組 織 (Convention on Biological Diversity, CBD) 於 2000 年 1 月 29 日通過全球矚目的卡塔黑 納生物安全議定書 (Cartagena Protocol on Biosafety) ,並於 2001 年 5 月起開放各國簽署,
至2003 年 6 月 13 日第五十個國家帛琉簽署後,已具備成為國際公法所需條件,直至 2010 年3 月已有 157 個國家簽約加入且立法實施 (www.biodiv.org) 。依據議定書規定,締約國 應採取必要且適當的法律、行政及其它措施,以防止或減少因基改生物發展、處理 (handling) 、運輸、使用及釋出 (release) ,而對於生物多樣性及人類健康產生不利影響及 風險,也就是說這個名為「卡塔黑納生物安全議定書」的國際條約,授予簽署的國家相當 權力,在考量可能造成環境及人類健康危害時,對於特定的基因改造生物,不論是動物、
植物或微生物,簽約國得禁止其輸入。議定書賦予締約國的權利,造成基改作物出口國與 其他國家在管理方面的嚴重歧見,同時也形成卡塔黑納生物安全議定書與世界貿易組織 (WTO) 相關規範之間的衝突。我國雖非「生物安全議定書」之締約國,但其內容亦涉及 WTO 有關基因改造產品等農業談判議題,故對我國亦產生規範作用,因此國人不得不重 視此議定書內之規範。我國於 2001 年衛生署公告基因改造之黃豆及玉米應辦理查驗登 記,並於2010 年修正公布「基因改造食品之安全性評估方法修正草案」(附錄一),以供 欲製造或輸入「基因改造食品」之業者及衛生主管機關確認該產品及製程之安全性。此評 估方法適用於具有安全食用歷史之植物經基因改造技術改造所得之基因改造植物食品的 安全與營養相關議題,並未涉及基因改造之動物飼料與用該飼料餵養之動物,亦未涉及基 因改造植物之環境風險議題。此外目前世界各國對於以基改作物進行雜交所產生的混合型 基改品系 (stacked traits) 的評估及管理態度有很大的不同。我國於 2008 年公布「混合型 基因改造食品」安全性評估原則,其適用對象為各殖入之性狀基因均源自已經審核通過之 基因改造食品,並以傳統育種方法育成之混合型基因改造植物所衍生之混合型基因改造食
1-4 基改作物的管理與檢測
自1996 年基因改造作物正式開始商業栽培,迄今已將近 15 年。基改作物由於政策、
成本及產量上的優勢,透過全球的大規模作物交易,已廣泛的進入世界各地的市場 (ISAAA, 2010) 。為了降低基改作物的安全性疑慮,世界各國不斷的持續進行基改作物安全性研究,
並且在科學研究成果的基礎上,制訂了安全性評估方法。即使如此,社會大眾對於基改作 物的疑慮和爭議仍然持續不斷,甚至情緒化的反應有越演越烈的趨勢 (Poppy, 2000) 。為 了保護消費大眾、食品製造商、農民及種子公司的利益,世界各國當局紛紛立法對於基因 改造食品進行規範和管理。目前已經有超過 50 個國家訂定食品中基改成分需要標示的上 限,也就是說當食品中的基改成份超過一定的限度時,就必須標示該食品含有基改成分。
目前各國對於食品中基改成份標示的規定不盡相同,美國為自願性標示,我國與日本為 5
% (DOH Food No. 0900011746, 2001; Notification 1775, 2000) ,韓國為 3% (Notification 200-31,2000) ,澳洲與紐西蘭為 1% (Standard 1.5.2, 2001) ,歐盟與俄羅斯為 0.9% (EC No. 1829/2003 and 1830/2003,2003; Russian Federation Biotechnology GMO Labeling Requirement, 2006) ,中國大陸為 0% (Order 10, 2002) 。由於各國准許上市的基改作物品 系和有關基改食品標示的法規並不相同,為了避免紛爭,可靠而準確的基改作物檢測與定 量方法是基改作物管理上一個不可缺少的環節。歐盟已於 2004 年開始明確要求申請許可 上 市 的 基 改 作 物 必 須 提 供 品 系 專 一 性 (event-specific) 檢 測 方 法 (EC No. 641/2004, 2004) 。
基改作物的檢測與定量方法依據所檢測的目標可以大致分成三類,第一類是針對 DNA 進行檢驗,以聚合酶鏈反應 (polymerase chain reaction,PCR) 和 DNA 微陣列 (DNA microarray) 技術最常見。第二類是針對特定的蛋白質進行檢驗,通常是以抗體進行偵測,
市售產品常見技術包含ELISA (enzyme-linked immunosorbent assay) 及蛋白質試紙 (protein test strip) 。第三類為偵測基改作物中特定的代謝物或是其代謝物指紋圖譜,常用的技術包 含FT-IR (fourier transform infrared spectroscopy) 及質譜分析 (mass spectrometry) 。目前以
由於各國國情的不同,先進但高成本的技術有實際推廣應用的困難,所以低成本且可靠的 PCR 技術目前仍然是主流方法。雖然 DNA 微陣列技術具有高通量的特性,可以滿足未來 對於基改作物篩檢的需求,但是目前DNA 微陣列仍有成本偏高和設備不普及等缺點,尚 無大量應用的產品出現。其他新穎的偵測基改作物 DNA 技術包含 QCM (quartz crystal microbalance) 生物感測器、乾式檢驗試劑條 (dry reagent dipstick-type sensor) 和表面電漿 共 振 (surface plasmon resonance sensor) 式感測器則仍在開發階段 (Michelini et al., 2008) 。PCR 偵測方法是以具有專一性的引子對於基改作物 DNA 進行放大,引子的設計 可以分成4 個等級 (圖 1-3) 。第一級為針對一些常用轉殖基因元件所設計的引子,由於目 前的基改作物內所帶有的轉殖基因元件有很高的重複性,這類引子不具有品系的辨認能 力。相對的,第一級引子因為可以檢測出多種基改作物而適合作為篩檢用途 (Lu et al., 2010) 。第二級引子為針對基改作物所帶有的異源基因進行檢測,此類引子可用於基改作 物的分類,但是因為偵測對象不廣,也幾乎不具有品系辨認能力,因此這類引子多半搭配 第一級引子作為篩檢用途。第三級引子為針對基改作物轉殖基因元件之間的接合處 (junction) 進行設計,具有 construct-specific 的辨認能力,可以分辨大多數的品系,但是由 於辨認專一性不如品系專一性引子,也無法像第一和第二級引子一樣作為篩檢用途,所以 目前已經很少使用。第四級就是所謂的品系專一性引子,為針對基改作物轉殖基因與宿主 染色體之間的接合處 (junction) 進行設計,具有最高的專一性,是目前公認的基改作物標 準檢測方法 (Demeke and Jenkins, 2010) 。
雖然目前基改作物的檢測技術已漸趨成熟,但由於基改作物的品系和種類急速增 加,使得基改作物檢測的未來挑戰日漸龐大複雜。目前上市的基改作物品系已達到 155 種,而且持續快速的增加中。基改作物品系的急速增加使得篩檢基改作物的時間、金錢與 人力的負擔不斷的上升,高通量檢測技術無可避免的將成為未來發展的重點。目前尚無方 法可以準確的分辨樣品中混合型基改作物來源是來自單一混合型基因改作物品系,或是來
圖1-2、基改作物 PCR 檢測引子的四種設計方式 (Holst-Jensen et al., 2003) 。 Fig. 1-2.Four-type of primer design of PCR assay for GM crop detection.
Terminator Stop signal
Gene coding for new feature (the modified gene) Host DNA
Left flank
1 1
3
2
3
4 4
Category 1: screening Category 2: gene specific Category 3: construct specific Category 4: event specific
Promoter Start signal
Host DNA Right flank
1-5 基改作物的非預期效應
隨著基改作物在消費市場的普及,世界上許多國家均立法規範基改作物的安全性評 估流程,以科學評估來維護大眾的權益。即使如此,人們對於基改作物的安全性疑慮並 沒有因此消減,反而越趨重視。對於社會大眾來說,存在於若干目前尚無完整科學定論 或是缺乏系統性評估方法的議題仍然是主要的疑慮。這些議題受到日積月累的誤解濫 用,已成為未來基改作物安全性評估的最大挑戰,其中最為常見的就是「非預期效應」
(Rischer and Oksman-Caldentey, 2006) 。「非預期效應」 (unintended effects) 為概指基因改 造過程中所產生非預期的影響,而使得基改作物有非預期的改變,常被認為帶有負面的 意涵。非預期效應的存在也是目前基改作物安全性評估所遵循的「實質等同」概念受到 爭論的主要原因。Rischer and Oksman-Caldentey 於 2006 年提出對於基改作物有關此方面 的完整科學性描述應考慮:
(一) 預期效應 (intended effects)
1. 由於轉殖基因的影響而達成預期的改造目標 2. 造成表現型、組成分或其他農藝性狀的改進 3. 可由特定的定量方法對於目標代謝物進行分析
(二) 非預期效應
1. 在統計學上具有顯著差異的非預期變化
2. 評估比較時應將生物個體間存在的差異納入考量,也就是必須包含適當數量的對 照樣品數
3. 所發生的變化對於農藝表現或許會造成衝擊,但與威脅人類健康或環境生態並不 具有必然之關連
(1) 在統計學上具有顯著差異的非預期變化,但無法由目前對於科學知識的瞭 解加以進一步闡釋的
(2) 微 小 但 可 能 重 要 的 改 變 可 藉 由 整 體 的 代 謝 分 析 描 寫 方 法 (metabolic profiling methods) 提供相關資訊
由上述原則可得知,有關非預期效應涉及的層面相當廣泛,包含直接與作物本身相 關 的 基 因 體 (genome) 、 轉 錄 體 (transcriptome) 、 蛋 白 質 體 (proteome) 和 代 謝 體 (metabolome) 評估,或是土壤微生物菌相改變等環境衝擊評估。目前有關基改作物非預 期效應的研究比較集中在與人類有切身關係的方面,也就是直接與食用安全有關的作物 本身。此類研究主要有兩個重點:其一為針對轉殖基因的表現與功能進行特定目標的分 析,以評估轉殖基因的影響是否如預期。另一方面則利用代謝分析描寫方法,對於基改 作物及各種傳統育種品系進行比較,以進行非預期變化的評估 (Cellini et al., 2004;
Filipecki and Malepszy, 2006) 。有多種「體學」 (omics) 方法已經被用於非預期效應研究,
包括功能性基因體學 (functional genomics) 、轉錄體學 (transcriptomics) 、蛋白質體學 (proteomics) 以及代謝體學 (metabolomics) 。然而目前尚無任何一種代謝分析描寫方法具 有足夠的涵蓋性 (comprehensiveness) 可以將所有的代謝變化進行描述 (Cellini et al., 2004) 。並且由於成本等實際上的限制,也無法對同一個目標用所有的方法都加以研究。
以蛋白質二維電泳質譜分析為主要工具的蛋白質體學研究方法目前已經相當成熟,
並且已有多篇應用在非預期效應研究的文獻發表 (Lehesranta et al., 2005; Ruebelt et al., 2006a; Ruebelt et al., 2006b; Ruebelt et al., 2006c) 。與其他代謝分析描寫方法相比較,應用 蛋白質體學方法進行研究具有相對較為簡易、成本相對較低、容易標準化等優點。隨著 全基因定序技術以及 cDNA 微陣列的逐漸普及,以轉錄體學方法進行非預期效應研究的 潛力也開始受到注意 (Filipecki and Malepszy, 2006) 。除了 cDNA 微陣列進行基改作物的 非預期效應評估之外 (Cheng et al., 2008) ,亦有文獻結合 cDNA 微陣列與反轉錄-即時定
分析 (Cellini et al., 2004) 。有鑑於單一體學研究方法僅能涵蓋有限的分子層次,目前基 改作物非預期效應的研究的趨勢為整合多個體學方法進行整體性評估,可以更精確的描 述基改作物中的非預期效應 (Barros et al., 2010) 。
雖然Codex 已於 2003 年將非預期效應納入基改食品的風險評估考量中 (Haslberger, 2003) ,但目前對於非預期效應的評估仍因科學技術的限制,而無公訂的研究方法與策略 (Rowland, 2002; Cellini et al., 2004; Rischer and Oksman-Caldentey, 2006) 。我國於 2010 年 公布的「基因改造食品之安全性評估」中附件五 『非預期效應』之評估方法如下:
安全性評估得提供非預期效應相關之試驗數據與資料,包括基因改造所嵌入之DNA 序列、基因改造植物後續之傳統育種過程、源於DNA 序列隨機嵌入植物基因體中而導致 植物現有基因之分裂或靜默、靜默基因之活化,或是改變現有基因之表現等現象,以減 少基因改造植物食品對人體產生非預期危害之可能性。基因改造植物食品得以比較評估 之原則分析其可能之非預期效應。
(一) 觀察下列項目,並提出試驗數據資料說明
1. 基因改造植物之表現型及農藝特性,以比較判斷其他性狀表現之變異情形 2. 基因改造導致新代謝物之產生或原有代謝產物式樣之改變情形,例如原有酵素
之大量表現,可能會引發間接之生化效應,或是改變代謝途徑之調控或代謝物 之含量
(二) 具非預期效應之基因改造植物食品應評估非預期效應之生物相關性及其對食品安全 之潛在影響
本研究針對基改抗胡瓜嵌紋病毒番茄進行基因改造所嵌入之DNA 序列選殖、cDNA
1-6 基改抗胡瓜嵌紋病毒番茄
胡瓜嵌紋病毒 (Cucumber Mosaic Virus, CMV) 為 Bromoviridae 科 Cucumovirus 屬的 RNA 植物病毒,自 1916 年在美國發現此種病毒以來 (Doolittle, 1916) ,胡瓜嵌紋病毒就 是世界上的重要植物病原。胡瓜嵌紋病毒的寄主範圍廣泛 (目前已知約有 191 種植物) , 又可經由機械及蚜蟲快速傳播,防治極為困難,對於農業經濟的衝擊相當大,也使得研 究人員不斷的利用轉殖基因技術研發抗胡瓜嵌紋病毒品系 (Morroni et al., 2008) 。1986 年 Powell-Abel 等 人 首 先 發 表 在 植 物 體 內 表 現 病 毒 鞘 蛋 白 基 因 可 以 產 生 coat protein-mediated resistance (CP-MR) 作用而使得轉殖基因植物具有病毒抗性。接著首例藉 由表現胡瓜嵌紋病毒鞘蛋白基因產生抗胡瓜嵌紋病毒性狀的轉殖基因植物在隔年發表 (Bryan et al., 1987) 。此外 Carr et al. (1994) 指出,帶有 truncated CMV RNA-2 replicase 基 因的煙草轉殖植株,可成功地表現出對胡瓜嵌紋病毒的抗性,且其 T1、T2後代亦具有此 抗病性。此類技術不斷的應用在煙草、番茄和甜瓜等多種作物之抗病育種,而且抗病基 因可遺傳至後代,仍表現出很好的抗病性 (Xue et al. 1994,Provvidenti and Gonsalves, 1995) 。
番茄 (Solanum lycopersicon) 是世界上重要的蔬菜,2008 年世界番茄總產量約為 130 萬噸,約佔世界蔬菜生產總量的14.2% (FAOSTAT,http://faostat.fao.org/) 。2008 年臺灣 地 區 生 產 量 為 11 萬公噸,在蔬菜生產量中排行第四名 (行政院農委會農糧署,
http://www.afa.gov.tw/index.asp) 。番茄一直是重要的基改目標作物,世界上第一個商業化 的基改作物即為番茄 (CGN-89564-2, FLAVR SAVR) 。世界上有 6 個品系的基改番茄曾 經商業化,目前有 5 個品系仍然在商業化生產。由於消費習慣與品種的不同,目前以商 業化生產的基改番茄主要是以延緩成熟/軟化的性狀為主(5 個品系為延緩成熟/軟化,另 一個為品系性狀抗蟲),並沒有抗病毒性狀。由於臺灣地區的番茄消費習慣是以小果「聖 女」以及大果「黑柿」類型品種為主,採收運送時果實硬度較高,而且臺灣番茄產地與 市場的運送距離與時間較短,並沒有因為運送時果實軟化而造成嚴重損失的問題。臺灣
由此可知植物病毒是臺灣番茄生產的主要威脅之一。
由於抗胡瓜嵌紋病毒天然的番茄種源不易取得,目前亞蔬中心所收集的六千多種番 茄種源中,未發現對胡瓜嵌紋病毒抗性強且穩定的種源。因此為有效防治胡瓜嵌紋病毒,
亞蔬中心利用基因轉移技術,將胡瓜嵌紋病毒鞘蛋白基因轉移到番茄栽培品種,育成抗 病基改植株,並篩選園藝性狀優良的品系,以降低防治成本及提高番茄的產量與品質。
在過去的研究中已經有許多藉由在植物中表現胡瓜嵌紋病毒基因而產生抗胡瓜嵌紋病毒 轉殖植物的例子 (Morroni et al., 2008) 。亞蔬中心柳建安博士所建立的基改抗胡瓜嵌紋病 毒番茄,是藉由在番茄內表現胡瓜嵌紋病毒鞘蛋白基因來抑制病毒的繁殖,具有良好之 抗胡瓜嵌紋病毒能力。基改抗胡瓜嵌紋病毒番茄使用的品種 (受體植物) 是臺灣農民廣泛 使用的大果鮮食番茄品種彌陀黑柿,甜度高,是一豐產品種。但近年來因為胡瓜嵌紋病 毒與番茄捲曲病毒的危害,使得農民已漸少使用此一品種。如果能推廣此一抗胡瓜嵌紋 病 毒 的 基 改 番 茄 , 可 減 少 番 茄 生 產 因 病 毒 與 真 菌 感 染 所 造 成 之 損 失 , 使 產 量 提 昇 10~15%,更可減少農藥之使用量,使農民之收益增加 7.5~10%, 每年經濟效益預估可增 加3-4 千萬台幣 (柳,2001;柳等,2005) ,具有相當的發展潛力。
1-7 研究架構與研究目的
由於植物轉殖的過程具有不確定性,加上目前對於複雜的植物整體生理代謝以及環 境生態平衡還無法完全明瞭,這些科學知識上的不確定使得許多人仍然對於基改作物的 安全性存疑。目前基改作物已經成為全球大規模農業生產的一部分,基改作物安全與否 也已經成為有關於經濟生產、國際貿易、法律管理、食品安全、科學研究、環境保護甚 至是政治考量的複雜議題。現行的基改作物安全性評估方法是基於已知的科學證據,使 用最為可靠有效率的的研究方法進行評估。在經過不斷的研究驗證,以及近十年的實際 執行結果顯示,現行的基改作物安全性評估方法具有相當的可靠度。即使如此,因為現
2003 年將納入基改食品的風險評估考量中 (Haslberger, 2003) ,成為現今的基改作物安 全性評估研究的重要發展趨勢。但目前對於非預期效應的評估仍因科學技術的限制,而 無 公 訂 的 研 究 方 法 與 策 略 (Rowland, 2002; Cellini et al., 2004; Rischer and Oksman-Caldentey, 2006) 。因此有關基改作物非預期效應評估的研究目前仍不斷的進行 中,希望藉由研究人員的不斷努力,非預期效應的評估可以為基改作物安全性提供更多 的可靠評估資訊。
我國尚未准許種植基改作物,目前已准許上市的基改作物均為外國研發生產,其安 全性評估與相關的研究都是在研發國進行,也因此雖然我國有公訂的基改作物安全性評 估方法,相關的安全性評估執行經驗與成果卻相當少。本研究以國產基改抗胡瓜嵌紋病 毒番茄為研究材料,希望藉由此一本土開發基改作物的安全性評估研究,獲取有關安全 性評估的實際經驗,並且在現行安全性評估方法之外,進行品系專一性檢測方法的建立、
土壤微生物菌相分析以及非預期效應的研究。希望透過這些研究,建立非預期效應的研 究模式,並探討基改作物安全性評估研究架構,以提供未來我國基改作物安全性評估工 作及發展的參考。
本論文研究架構如圖1-3 所示。首先遵循衛生署所公布「基因改造食品」之安全評估 方法進行過敏性評估 (第二章) 。由於嚴謹可靠的定量檢測系統為基改作物商業化的重要 環節,為進一步提升此一基改番茄的商業化潛力,本研究接著建立的品系專一性即時定 量 PCR 檢測系統,並進行了番茄果實 DNA 於模擬胃液中的分解研究 (第三章) 。本論 文第四章進行基改抗胡瓜嵌紋病毒番茄對於環境的非預期效應研究,以變性梯度膠體電 泳技術 (denaturing gradient gel electrophoresis,DGGE) 評估基改抗胡瓜嵌紋病毒番茄對 於土壤微生物菌相的影響。本研究的最後一部份結合第二和第三章所得到的轉殖基因嵌 入與表現分析結果,以cDNA 微陣列、蛋白質二維電泳和 RT-qPCR 方法針對基改抗胡瓜 嵌紋病毒番茄本身進行非預期效應的研究。藉由整合基因層次-轉錄體學層次以及蛋白質 體學層次的研究結果,整體性的評估基改抗胡瓜嵌紋病毒番茄非預期效應 (第五章) 。
mRNA表現分析 (RT-qPCR and cDNA
microarray)
品系專一性定量 PCR 檢測方法建立
過敏性評估 轉殖基因嵌入位置
確認與分析
基改抗胡瓜嵌紋病毒番茄
蛋白質二維電 泳分析 轉殖基因鄰近序列選殖
過敏原序列比 對分析
模擬胃液 分解試驗
14
第二章 基改抗胡瓜嵌紋病毒番茄過敏性評估
2-1 前言
亞蔬中心柳建安博士所建立的基改抗胡瓜嵌紋病毒番茄在T0-T6均表現出穩定而良 好的抗胡瓜嵌紋病毒能力。2005 年的田間栽培試驗中,T6 含轉殖基因番茄花粉以人工 授粉方式,分別與龍葵、酸漿番茄、茄子與枸杞等近親植物雜交。結果並未發現轉殖基 因轉移至近緣野生物種 (柳等,2005) 。另外基改番茄在果實採收後,經淹水處理二週 後,並無殘株再生與遺留種子萌芽之疑慮。種子發芽力測定資料顯示,不論是發芽皿法 或是埋土發芽測定法,基改番茄種子與對照組 (非基改種子) 並無明顯差異,其發芽力 均介於90-95% (柳等,2005) 。由於此一基改抗胡瓜嵌紋病毒番茄具有相當發展潛力,
自2006 年起衛生署補助進行「國產基因轉殖抗胡瓜嵌紋病毒番茄安全評估模式之建立」
群體型研究計畫 (農業生物技術國家型科技計畫-基因改造食品安全評估環境建置) 。 該計畫共分成6 個子計畫以進行基改抗胡瓜嵌紋病毒番茄的安全性評估,分別為子計畫 一:基因轉殖抗胡瓜嵌紋病毒番茄之食品安全性評估; 子計畫二:園藝生理變化對基改 抗胡瓜嵌紋病毒番茄外源基因差異表現之影響與安全評估; 子計畫三:胡瓜嵌紋病毒基 改番茄之毒理研究; 子計畫四:製備番茄抗胡瓜嵌紋病毒之重組嵌蛋白與建構其免疫分 析平台; 子計畫五:抗胡瓜嵌紋病毒基因轉殖番茄之食品成分分析; 子計畫六:抗胡瓜 嵌紋病毒基因轉殖番茄之栽培與園藝特性評估,本論文第二章為子計畫一的研究成果。
2-2 基改抗胡瓜嵌紋病毒番茄基本資料 (一) 「基因改造植物」之描述
基因改抗胡瓜嵌紋病毒番茄,利用表現胡瓜嵌紋病毒鞘蛋白基因,可抑制病毒 的繁殖,具有良好之抗胡瓜嵌紋病毒能力。
2. 育種栽培及發展之歷史,特別是對有礙人體健康之性狀說明
番茄原產於南美洲的秘魯,人類在史前時代墨西哥中部的阿茲提克 (Aztec) 文化已經有栽培食用的紀錄。隨著歐洲人探索美洲大陸以及西班牙殖民南美洲,在 1544 年歐洲出現了最早有關番茄的文獻紀錄,當時主要作為觀賞及醫藥用途。西 元1600 年之後,在歐洲番茄逐漸開始食用,在 18 世紀時已經發展出眾多的品種,
並且逐漸的傳播到全世界。隨著持續不斷的育種改良,原生番茄品種中含有的有毒 生物鹼- tomatine 已經降低許多,現代番茄品種中 tomatine 含量相當低,甚至無法 測出 (Barceloux, 2008) 。人類食用番茄的歷史至少已經 400 年,目前全球各地都 有栽培和食用,並無明顯的安全疑慮。
3. 與宿主植物安全性相關之基因型及表現型資訊,包括 a. 毒性
原生番茄品種中含有的有毒生物鹼- tomatine,現代番茄品種中 tomatine 含量 相當低,甚至無法測出 (Barceloux, 2009) 。
b. 過敏誘發性
目前已知番茄的過敏原有6 種 (表 2-1) ,分成 3 個 group,包含 profilin (Lyc e 1) 、β-fructofuranosidase (Lyc e 2) 和 nonspecific lipid transfer protein (Lyc e 3) ,番茄並非高過敏誘發性的食物。
c. 宿主類緣種與對宿主植物之遺傳背景具顯著貢獻之植物與安全性相關之基因型 及表現型
茄科植物包含許多重要作物,包括菸草 (Nicotiana tabacum) 、茄子 (Solanum melongena) 、甜椒/辣椒 (Capsicum sp.) 和馬鈴薯 (Solanum tuberosum) 。除了菸
表2-1、已知的番茄過敏原 (FARRP allergen database 10.0,2010) Table 2-1 Known tomato allergens
IUIS Allergen* Group Length (a.a.) GI number Lyc e 1 Lycopersicon Lyc e 1 131 17224229 Lyc e 1 Lycopersicon Lyc e 1 131 16555787 Lyc e 2.0101 Lycopersicon Lyc e 2 553 18542113 Lyc e 2.0102 Lycopersicon Lyc e 2 636 18542115 Unassigned Lycopersicon Lyc e 3 114 71360928 Unassigned Lycopersicon Lyc e 3 114 71360930 IUIS: International Union of Immunological Societies.
* Allergen nomenclature: Allergens are designated according to the accepted taxonomic name of their source as follows: the first three letters of the genus, space, the first letter of the species, space, and an Arabic number.
4. 安全食用歷史,可包括
a. 傳統育種、運輸及貯存的方法
現代品種的番茄為自花授粉優勢植物,進行育種時必須採取人工授粉的方 式。目前人工育種產生的品種相當多,目前亞蔬中心所收集的番茄品種約有六千 多種。番茄運送貯存方式與一般蔬菜水果類似,紅熟的番茄質地較軟。運送貯存 時要避免重壓。此外番茄是典型的後熟型園藝作物,採收之後會不斷的繼續成 熟,需要低溫保存才能存放比較久的時間。
b. 安全食用所需之特殊加工方法
一般市面上的番茄可直接生食、熟食或是加工後安全食用,並不需要特殊的 加工方法。
c. 在一般膳食中所扮演的角色
番茄營養價值高,容易取得,而且廣為世人接受。番茄為重要的蔬菜,在許 多國家的日常飲食中都是不可或缺的食材。同時番茄也常被直接生食,當成水果 來食用。近來番茄因為富含具有多種生理活性的茄紅素 (lycopene) 而受到保健 食品相關研究的矚目。
(三) 基因提供生物 (可包括類緣種) 之描述 1. 俗名、學名及分類學資訊
胡 瓜 嵌 紋 病 毒 (Cucumber Mosaic Virus, CMV) 為 Bromoviridae 科 Cucumovirus 屬的 RNA 植物病毒。
3. 作為食品用途之資訊,以及除食品用途外之可能攝取途徑
胡瓜嵌紋病毒並無食品用途,僅有可能曾經隨著被感染的植物一起被人類食 用。食品用途外之可能的攝取途徑可能不存在。
(四) 基因改造之描述 1. 轉殖之描述
a. 轉殖方法
基改抗胡瓜嵌紋病毒番茄原受體番茄品種 (非基改品種) 為臺灣農民廣泛 栽 培 大 果 鮮 食 品 種 彌 陀 黑 柿 ( 亞 蔬 中 心 編 號 L4783) , 使 用 的 農 桿 菌 (Agrobacterium tumefaciens) 品系為 LBA4404。轉殖步驟簡述如下:番茄種子以 含有 0.1 % Tween-20 之 25 %家用漂白水 (Clorox,有效成份為 5.25 %次氯酸 鈉) ,表面消毒 10~15 分鐘後,播於 1/2× Murashige and Skoog (MS) 固體培養基 (Murashige and Skoog, 1962) 表面,置於 25 oC 黑暗中,發芽後移至光照 16 小時 黑暗8 小時 (16L/8D) 光週下生長。當番茄小苗之真葉 (true leaf) 尚未長出,或 真葉尚小時,取子葉中段成為培植體。將培植體以葉面朝上置於含 2 mg/L BA (6-benzylaminopurine) 、0.5 mg/L IAA (indole-3-acetic acid) 之 MS 培養基,置於 25 oC、16L/8D 光週下 2 天,進行前培養。將農桿菌以含有 200 μM acetosyringone 之MS 液體培養基懸浮,使其濃度約為 6 × 108 cell/mL (OD660 = 0.2) 。培植體以 農桿菌液感染 5 分鐘後,移至含 2 mg/L BA、0.5 mg/L IAA 及 200 μM acetosyringone 之 MS 固體培養基進行 2 天之共培養,共培養之環境條件與前培 養相同 (25oC,16L/8D) 。共培養 2 天後,將培植體移至無菌培養皿,以 carbenicillin (500 mg/L) 及 cefotaxime (100 mg/L) 溶液泡洗 10 分鐘以除去農桿菌。將經由 carbenicillin 及 cefotaxime 溶液泡洗後之培植體移至含 2 mg/L BA、0.5 mg/L IAA、
50 mg/L kanamycin、50 mg/L cefotaxime 及 500 mg/L carbenicillin 之 MS 固體培養
elongation) 。將抽長至約 3~4 cm 之幼芽切下,移至 0.15 mg/L NAA、25 mg/L kanamycin、25 mg/L cefotaxime 及 25 mg/L carbenicillin 之 MS 固體培養基中長根。
將長根後之小植株 (plantlet) 移至栽培介質中生長 (柳,2001) 。
b. 載體各結構片段之資訊
(1) 來源,如植物、微生物、病毒、合成
轉 殖 載 體 (transformation vector) 是 由 中 興 大 學 葉 錫 東 教 授 提 供 的 pBICBI2 質體,pBICBI2 是將合成的 binary vector pBI121 (圖 2-1) 上面的 gus 基因由胡瓜嵌紋病毒臺灣品系的鞘蛋白基因 (cmvcp) 取代構築而成。
(2) 辨識特徵
CaMV 35S 啟動子 (Cauliflower Mosaic Virus 35S promoter) 及 nos 終結子 (Agrobacterium Ti plasmid nopaline synthase gene terminator) 序列,以 nptII (neomycin phosphotransferase II) 基因為選殖標識,並帶有胡瓜嵌紋病毒臺灣品 系的鞘蛋白基因 (cmvcp) 。
(3) 於植物中預期之功能
嵌入RB 至 LB 間的 DNA 片段 (圖 2-1) 進入宿主染色體 DNA,提供胡瓜 嵌紋病毒抗病性。
c. 中間宿主
以農桿菌為中間宿主。
2. 提供欲轉殖入宿主植物之 DNA 的相關資訊
圖2-1、質體 pBI121 圖譜 (GenBank AF48578) 。 Fig. 2-1 Plasmid map of pBI121.
pBI121 embl AF485783 AF485783
14758 bp
npt II
gus A
traF hom ologue s
te tA hom ologue s
te tA hom ologue s trfA homologue s
npt III hom ologue s Tn IS 1 hom ologue s
npt III hom ologue s k ilA hom ologue s
G US F
CaM V 35 S promote r NOS prom ote r ColE 1 ori
ori V
NOS term ina tor
NOS term ina tor
T-RB
T-LB
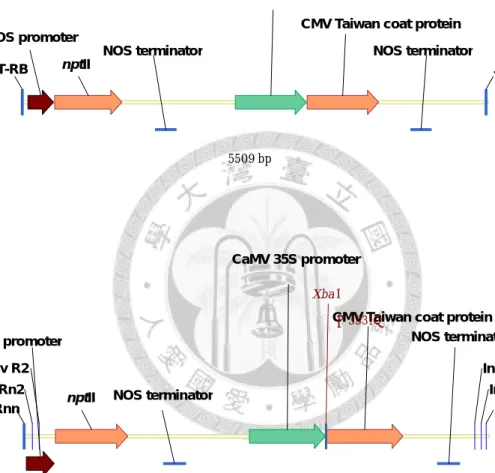
圖2-2、基改番茄 R8 的轉殖基因構築。
Fig. 2-2 Transgenic construct of GM tomato R8.
RB and LB: right and left T-DNA border. Pnos: nopaline synthase promoter. NptII: neomycin phosphoesterase II. Tnos: nopaline synthase terminator. P35S: Cauliflower Mosaic Virus 35S promoter. Cmvcp: Cucumber Mosaic Virus coat protein gene
b. 大小及辨識特徵
轉殖入宿主植物的部分,DNA 大小為 5509 bp,辨識特徵胡瓜嵌紋病毒臺 灣品系的鞘蛋白基因 (cmvcp) 與 nos 終結子序列接合處。
c. 於最終重組體中的位置及方向及功能將本論文第三及第五章詳述。
(五) 基因改造之特徵
下列資料將於本論文第三及第五章詳述:
1. 嵌入植物基因體之轉殖 DNA 的相關資訊 2. 提供基因改造植物體內任何新表現物質之資料 3. 其他必要資料
2-3 材料與方法
本研究所進行的基改抗胡瓜嵌紋病毒番茄過敏誘發性評估依據我國「基因改造食品」
之安全評估方法 (六) 安全性評估之 2. 新表現物質之毒性與過敏誘發性的初步評估之 b.
新表現蛋白質之過敏誘發性初步評估進行 (附錄一)
(一) 基改抗胡瓜嵌紋病毒番茄過敏誘發性評估
基改作物之過敏誘發反應可分為三類:第一類為在轉殖過程中改變了內生蛋白 質; 第二類為經由基因改造將過敏原轉殖進作物中; 第三類為經由基因改造改變了作 物的代謝路徑,產生新的可能是過敏原的蛋白質。因為引發過敏之機制極其複雜,造 成過敏誘發性必須採取評估的方式; 評估方法分為三項:探討基因來源、序列比對與 模擬胃液消化試驗。
1. 探討基因來源
該基因轉殖產物是ㄧ個未知是否具有過敏誘發性之蛋白質,或是基因來自從未接觸 的基因提供者,則此方法將難以預測改造作物是否因此具有過敏誘發性。
2. 胺基酸序列過敏原資料庫搜尋比對
抗原與抗體間之結合反應是藉抗原表面的抗原簇 (epitope) 與抗體結合,因此 若是未知蛋白質與已知過敏原蛋白具有類似或相同之胺基酸序列,則此未知蛋白質 即可能具有過敏原性 (Astwood and Fuchs, 1996; Metcalfe et al., 1996) 。利用生物資 訊學可將待測蛋白質及已知過敏原蛋白進行胺基酸序列比對,即可評估其過敏原性;
若兩者間具有高度相似性,則其可能具有具有過敏誘發性。
胡瓜嵌紋病毒鞘蛋白胺基酸序列 (GenBank AAY21660) 如圖 2-3 所示,共有 218 個胺基酸。所使用的過敏原資料庫包含 AllergenOnline version 10.0 database (1417 peer reviewed sequences; http://www.allergenonline.com/) 、 Structural Database of Allergenic Proteins (SDAP; 891 allergen sequences; http://fermi.utmb.edu/SDAP/) 和 Allergen Database for Food Safety (ADFS; 1285 registered allergens;
http://allergen.nihs.go.jp/ADFS/index.jsp) 。依據資料庫所提供搜尋工具的不同,在 這3 個資料庫進行 80 mer amino acid FASTA alignment 搜尋,在 ADFS 以及 SDAP 資料庫進行滑動連續胺基酸片段搜尋,並在AllergenOnline 以及 SDAP 資料庫進行 full FASTA overall search。在 80 mer amino acid FASTA alignment 搜尋中,如果超過 35% 的胺基酸序列相同為陽性 (具有顯著的過敏原相似度,Codex, 2003) 。在滑動 連續胺基酸片段搜尋中,如果有連續 8 個線性胺基酸序列相同則結果為陽性 (Aalberse, 2000) 。 在 full FASTA overall search 中 具 有 顯 著 低 期 望 值 (AllergenOnline 資料庫為小於 1×10-7,SDAP 資料庫為小於 0.01) 的結果為陽性 (Shin et al., 1998; Reese et al., 1999; Chatchatee et al., 2001) 。
圖2-3、基改番茄 R8 的胡瓜嵌紋病毒鞘蛋白基因與胺基酸序列。
Fig. 2-3. DNA and amino acid sequence of CMV coat proetin gene of transgenic tomato R8.
25
a. 胡瓜嵌紋病毒鞘蛋白基因序列選殖與確認
以 試 劑 套 組 (GeneMark plant genomic DNA purification kit , Hopegen Biotechnology Development Enterprises,Taiwan) 進行基改番茄葉片 DNA 抽取,
再 以 引 子 對 35S1 (5’-GCTCCTACAAATGCCATCA-3’) - NOS3 (5’-GCTCCTACAAATGCCATCA-3’) 進行 PCR 放大胡瓜嵌紋病毒鞘蛋白基因片 段。PCR 反應條件如下:先以 95oC 反應 1 min,再以 95oC 30 sec- 55oC 30 sec-72oC 1 min 為一循環,進行 35 次增幅循環,最後於 72oC 反應 10 min。PCR 放 大產物經由定序後取得基改番茄中胡瓜嵌紋病毒鞘蛋白之核酸序列。
b. 基改番茄中轉殖基因於不同生長時期的表現量
本研究測定基改番茄中轉殖基因於四個不同生長時期的表現量,包含 4-6 片真葉時期、始花時期、綠色果實時期與「黑柿」品種番茄果實採收時期 (turning,俗稱「一點紅」,圖 2-4) 。
(1) 番茄樣品
本研究所使用的種子由亞蔬中心提供,於恆溫植物培養室中種植傳統 番茄品系L4783 和基改番茄品系 R8,培養溫度 25oC,光照設定為早上 4 點 至晚上8 點,每日 16 小時光照與 8 小時黑暗(16L/8D)。
(2) Total RNA 製備與純化
Total RNA 以 ConcertTM Plant RNA Reagent (Invitrogen,Carlsbad,USA) 進行抽取。將番茄植株樣品以液態氮急速冷凍後研磨成細粉,在未解凍的情 況下取約0.5 g 樣品粉末加入 4 mL ConcertTM Plant RNA Reagent 中,混勻後
圖2-4、番茄果實成熟度的六個判定標準 (Goud, 1983) 。 Fig. 2-4. Tomato color chart showing the six official classifications.
GREEN
The tomato surface is completely green. The shade of green may vary from light to dark.
BREAKERS
There is a definite break of color from green to bruised fruit.
Tannish - TURNING
Tannish - yellow, pink or red color shows on over 10% but not more than 30% of the tomato surface.
PINK
Pink or red color shows on over 30% but not more than 60% of the tomato surface.
LIGHT RED Pinkish -
not more than 90% of the tomato surface.
RED
Red color shows on over 90% of the tomato surface.
yellow, pink or red on 10% or less of the tomato surface.
red or red color shows on over 60% but red color covers