國 立 台 東 大 學 生 命 科 學 系 研 究 所 Department of Life Science
National Taitung University
碩士論文 Master Thesis
於大腸桿菌 EPI300 中建立噬菌體 Smp14 的突變平台
Establishment of mutation plateform of Stenotrophomonas maltophilia bacteriophage Smp14 in E. coli EPI300
指導教授:陳芝融 博士 (Dr. Chiy-Rong Chen)
研究生:李俊毅 (Jyun-Yi Li)
中華民國 105 年 7 月
July 2016
謝 誌
漫長的四個寒暑,終於完成了碩士的論文,回想撰寫的過程,受 到許多師長、同學及家人的鼓勵與協助,內心感謝萬分。
感謝指導教授陳芝融博士的指導,在求學研究的過程,總是鉅細 靡遺與條理分明地分析、歸納並指出實驗上問題癥結所在。以及國立 中興大學教授曾義雄博士不遠千里前來擔任口試委員,並給予論文上 的建議與修正;鼓勵學生努力向上學習。再者,感謝段文宏教授指導 學生口試,並於實驗儀器上給予學生不少協助,使人獲益良多。老師 們的諄諄教誨都令學生感念於心。
感謝同學鈞鼎、政嘉、宇平與昭如,實驗室學弟妹財修、顥翰、
玨豪、宗憲和月芳,在實驗階段提供許多的幫助和意見,在此階段有 你們的相伴帶給我許多歡樂的時光。
感謝女友麗子,於求學階段可以給予我最大包容和鼓勵,陪我度 過這幾年的艱辛和歡樂的時光。
最後感謝我的父母,給予我求學上的支助和開導,讓我可以無顧 慮的完成學業。
謹向研究過程中不吝指導與協助的貴人致上最衷心的感謝。
俊毅 謹誌 中華民國一○五年七月
i
摘要
Stenotrophomonas maltophilia 是需氧氣的革蘭氏陰性桿菌,廣泛分佈於環境 中,亦是人類伺機性病原菌,可造成肺炎病患 50% 以上的致死率。S. maltophilia 是多重抗藥性菌株,對大多數的抗生素都有抗藥性,如 β-lactams 類抗生素。噬 菌體 Smp14 為 S. maltophilia 的溶裂型噬菌體,屬 T4-type 噬菌體,基因體大 小為 159,910 鹼基對,具 230 ORFs,其中有 120 ORFs 功能性未知。為了近一 步探討噬菌體 Smp14 ORFs 的功能性,欲藉由跳躍子 EZ-Tn5 隨機突變的方 式,於 E. coli EPI300 中建立噬菌體突變株基因庫,結果卻造成 E. coli EPI300 於培養過程中溶裂死亡。上述結果推測與噬菌體 Smp14 溶菌相關基因 ( 例如 lysozyme 和 holin ) 於 E. coli 系統被表現有關,因此決定於 E. coli EPI300 平台中建立噬菌體 Smp14 holin 和 lysozyme 的 antisense RNA 表現系統,以抑 制噬菌體 Smp14 的溶菌相關基因的轉譯作用。首先分別針對噬菌體 Smp14 溶 菌基因 holin 和 lysozyme 的啟動子區和反股區,以及 pOK12 上的終止子區,
分別設計具適當限制酶切位之引子對,再以 PCR 方式擴增目標 DNA 片段後,
將其分別轉接至載體 pTA 載體後,再轉接至 pSKII 與 pOK12 載體上,最終構 築 pSKII-asholin-aslysozyme 的 antisense RNA 表現系統,並將此質體轉殖入 E.
coli EPI300 中。後續實驗藉由跳躍子 EZ-Tn5 系統進行噬菌體 Smp14 DNA 的 突變作用後,利用電穿孔法將突變噬菌體 DNA 樣品轉殖入 E. coli EPI300 (pSKII-asholin-aslysozyme) 中,利用抗生素 Km 進行篩選,目前初步獲得 118 顆疑似含有噬菌體 Smp14 突變 DNA 的菌落。再以 colony PCR 對這些菌落進 行噬菌體 smp14 gp23 的片段偵測,於上述菌落中有 4 顆菌落於 DNA 電泳膠 上,顯示出具有相對應於 smp14 gp23 PCR 產物之條帶訊號。
ii
Abstract
Stenotrophomonas maltophilia is an aerobic gram-negative bacterium, found in various environments. It is an opportunistic pathogen that can cause pneumonia with more than 50% fatality rate. Multidrug-resistant S. maltophilia are capable of resisting many antibiotics, such as β-lactams. The lytic phge Smp14 of S. maltophiliais belongs to T4-type phage, with a genome consisting of 159,910 bp which contains 120 hypoththetical protein genes among 230 putative protein-encoding ORFs. To study the function of the unknow Smp14 genes, we used transposon EZ-Tn5 <oriV/kan-2> to constract a phage mutant library in E.coli EPI300. Cell lysis was found to occure after electroporation-mediated transformation of mutant DNA. It is possible that holin and lysozyme genes of Smp14 were expressed in E. coli EPI300 to cause cell lysis. Then, we constructed the antisense RNA system in E. coli EPI300 that was expected to inhibit the translation of the holin and lysozyme genes of Smp14. First, we ampified the target DNA fragment, promoter region and antisense region of phage Smp14 holin and lysozyme as well as terminator region of pOK12, by PCR. For expression of antisense RNA of phage Smp14 holin and lysozyme in E. coli EPI300, plasmids were constructed using pTA, pOK12 and pSKII as vectors. One plasmid (named pSKII-asholin-aslysozyme) was obtained and transformed into E. coli EPI300 to inhibit the translation of electroporated phage Smp14 :: EZ-Tn5 <oriV/Kan-2> holin and lysozyme genes. Preliminary results showed that among 118 colonies which might contain mutated Smp14 DNA, contained Smp14 gp23 four in PCR analyes.
iii
目錄
摘要 ... i
Abstract ... ii
表目錄 ... vi
圖目錄 ... vii
前言 ... 1
壹、嗜麥芽窄食單胞菌 (Stenotrophomonas maltophilia, Sm) ... 1
甲、微生物特性 ... 1
乙、人體伺機性 (opportunistic ) 病原菌... 3
丙、抗生素殺菌機制介紹 ... 5
丁、細菌抗藥機制分類 ... 5
戊、S. maltophilia 抗藥機制分類 ... 6
一、合成抗生素降解酶 ... 6
二、外排幫浦 (Efflux pumps)... 7
三、抗生素修飾酶 ... 10
貳、噬菌體療法 (phage therapy) ... 11
甲、噬菌體療法於動物實驗 ... 11
乙、噬菌體療法於人體實驗 ... 12
參、噬菌體 ... 13
甲、噬菌體構造與分類 ... 13
乙、S. maltophilia 的噬菌體 ... 16
丙、具廣泛宿主範圍的 S. maltophilia 的噬菌體 ... 17
丁、可感染 Xanthomonas 屬的噬菌體 ... 17
肆、可感染 S. maltophilia 噬菌體 smp14 ... 18
伍、常見的 DNA 突變方法介紹 ... 19
iv
甲、紫外線光(UV)突變法 ... 19
乙、藥劑突變法 ... 22
丙、跳躍子 (transposon) 突變法 ... 22
陸、反義 (antisense) RNA 干擾系統... 27
研究動機 ... 27
研究目的 ... 28
材料方法 ... 29
壹、研究設備及器材 ... 29
一、菌種、噬菌體與質體 ... 29
二、一般藥品 ... 29
三、酵素 ... 29
四、引子 (primer) ... 29
五、培養基 ... 29
六、抗生素使用濃度 ... 29
七、試劑與緩衝液 ... 29
貳、研究方法 ... 31
一、菌種之培養及保存 ... 31
二、噬菌體培養 ... 31
三、溶菌斑測試 ... 32
四、DNA 備製 ... 32
五、聚合酶連鎖反應 (polymerase chain reaction, PCR) ... 34
六、洋菜膠電泳分析 (Agarose gel electrophoresis) ... 34
七、限制酶切割作用 ... 35
八、DNA 之回收 ... 35
九、DNA 黏接反應 (DNA ligation) ... 35
十、啟動子活性分析 ... 36
十一、轉形作用 ... 36
v
結果與結論 ... 39
一、lysozyme 和 holin 啟動子序列分析的活性測試 ... 39
二、溶菌基因 holin 的 antisense RNA 之系統構築 ... 41
三、溶菌基因 lysozyme 的 antisense RNA 之系統構築 ... 43
四、antisense RNA 系統轉殖入 E. coli EPI300 ... 45
五、噬菌體 Smp14 突變株 DNA 篩選... 46
討論 ... 47
引用文獻 ... 50
vi
表目錄
表 一、S. maltophilia 的生長特性... 2
表 二、12 種抗生素對 68 株 S. maltophilia 的藥物敏感性測試... 4
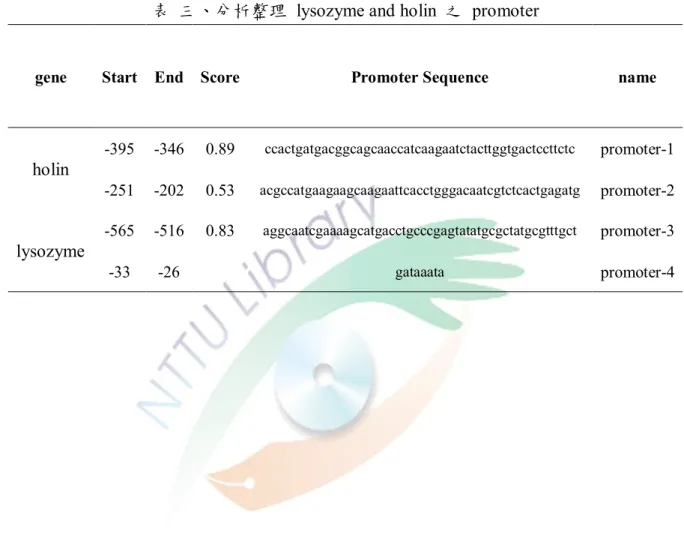
表 三、分析整理 lysozyme and holin 之 promoter ... 55
表 四、PCR 實驗中所使用之引子 ... 85
vii
圖目錄
圖 一、細菌的耐藥機制。 ... 5
圖 二、噬菌體的繁殖過程可以分成兩類,溶裂型噬菌體與潛溶型噬菌體 (lytic and lysogenic phage)。 ... 15
圖 三、噬菌體外型。 ... 15
圖 四、UV 所引起的 CC 嘧啶二聚體突變影響。 ... 21
圖 五、鳥嘌呤 (G) 受到烷化形成 O6-methylguanine,在 DNA 複製過程,最終 會造成 GC→AT 置換。 ... 24
圖 六、亞硝酸會將腺嘌呤 (A) 去胺化成次黃嘌呤,鍵結的鹼基將從胸腺嘧啶 (T) 換成胞嘧啶 (C)。 ... 25
圖 七、5-溴尿嘧啶誘變機制。這種分子有兩種互變異構亞型,酮形式(BUk)與 鹼基 A 鍵結,而烯醇式(BUe)與鹼基 G 鍵結,因而在 DNA 複製期間, 會造成 AT 鍵結變成 GC 鍵結。 ... 25
圖 八、2-氨基嘌呤會與胸腺嘧啶 (T) 或胞嘧啶 (C) 形成鍵結。 ... 26
圖 九、EZ-Tn5 系統造成 DNA 突變的複製過程。 ... 26
圖 十、holin promoter 之程式分析及設計的 promoter 區域。 ... 56
圖 十一、lysozyme promoter 之程式分析及設計的 promoter 區域。 ... 57
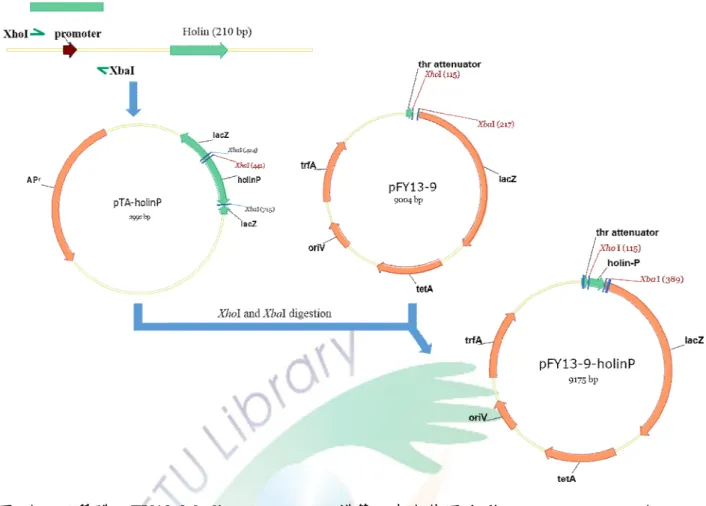
圖 十二、質體 pFY13-9-lysozyme-promoter 之構築。 ... 58
圖 十三、質體 pFY13-9-holin-promoter 之構築。 ... 59
圖 十四、 E. coli DH5α (pFY13-9-n) 轉殖株於 LB 中之啟動子活性分析測試。 ... 60
圖 十五、holin promoter 之程式分析及設計的 promoter 區域。 ... 61
圖 十六、lysozyme promoter 之程式分析及設計的 promoter 區域。 ... 62
圖 十七、antisense holin 之分析結果及設計的 antisense RNA 區域。... 63
圖 十八、antisense lysozyme 之分析結果及設計的 antisense RNA 區域。 ... 64
圖 十九、terminator 之程式分析結果。 ... 65
圖 二十、針對 holin 的 antisense RNA 系統中 terminator 區域的引子設計。 ... 66
圖 二十一、針對 lysozyme 的 antisense RNA 系統中 terminator 區域的引子設 計。 ... 67
圖 二十二、藉由 PCR 增幅出構築 antisense holin 和 antisense lysozyme 系統 之六種 PCR 產物。 ... 68
圖 二十三、質體 pTA-holin-promoter、pTA-as-holin 和 pTA-holin-terminator 之 構築。 ... 69 圖 二十四、質體 pTA-lys-promoter、pTA-as-lysozyme 和 pTA-lys-terminator 之
viii
構築。 ... 70
圖 二十五、以 PCR 法確認 pTA-holin-promoter、pTA-antisense-holin 和 pTA-holin-terminator 的構築。 ... 71
圖 二十六、藉由 PCR 增幅出包含 promoter、antisense RNA 和 terminator 之 五種 PCR 產物。 ... 72
圖 二十七、質體 pSKII-antisense-holin 構築。... 74
圖 二十八、以 PCR 法確認 pSKII-holin-promoter 的構築。 ... 75
圖 二十九、以 PCR 法確認 pSKII-holin-promoter-terminator 的構築。 ... 76
圖 三十、以 PCR 法確認 pSKII-as-holin 的構築。 ... 77
圖 三十一、質體 pOK12-lysozyme-terminator 之構築。 ... 78
圖 三十二、質體 pOK12-lys-promoter-terminator 之構築。 ... 79
圖 三十三、質體 pOK12-antisense-lysozyme 之構築。 ... 80
圖 三十四、以 PCR 法確認 pOK12-lys-terminator 的構築。 ... 81
圖 三十五、以切割酶確認 pOK12-lys-promoter-terminator 的構築。... 82
圖 三十六、以 PCR 法確認 pOK12-as-lysozyme 的構築。 ... 83
圖 三十七、噬菌體 Smp14 突變株 DNA 篩選。 ... 84
ix
縮寫字對照表
DTT dithiothreitol
EDTA ethylenediaminetetraacetic acid
EtBr ethidium bromide
g gram
kb kilobase
mg milligram
ml milliliter
mM millimolar
OD600 optical density at 600 nm
SDS sodium dodecyl sulfate
μg microgram
μl microliter
1
前言
壹、嗜麥芽窄食單胞菌 (Stenotrophomonas maltophilia, Sm)
甲、微生物特性
嗜麥芽窄食單胞菌 (Stenotrophomonas maltophilia, S. maltophilia) 最早 於 1943 年從胸腔液中分離出來命名為 Bacterium booker ,於 1961 年被 歸類為 Pseudomonas 屬,然後在 1983 年用 rRNA 的同源性分析以及基因 體 GC 的含量 (P. maltophilia 63 - 67.5%; Xanthomonas 63 - 70% 和 S.
maltophilia 66.1 - 66.9%),更改為 Xanthomonas 屬,最終於 1993 年定義為 Stenotrophomonas 屬 (Looney et al., 2009)。S. maltophilia 是一隻需要氧氣、
不會產孢子、非發酵型之格蘭氏陰性桿菌,菌體全長約為 0.5 至 1.5 μm,是 一隻代謝型 γ - 變形菌,廣泛的存在環境中,包括潮濕的土壤、植物根周、
植物和動物本體中都可分離出來。Margesin 和 Schinner 的報告指出,曾經在 10℃ 的高山環境下發現此菌株 (1991)。S. maltophilia 具有一至多根鞭毛,
可藉由鞭毛運動而移動,典型的菌落色澤為黃色或薰衣草綠,菌落平滑,表 面具光澤,邊緣完整 (Denton et al., 1998)。Joanna 參考前人的實驗數據整理 出 S. maltophilia 生長特性 (如表一)。從表中得知 S. maltophilia 以麥芽醣 為菌體主長的主要營養碳源,生長過程中另需要甲硫氨酸 (Methionine) 或 胱氨酸 (cystine),但也不是所有的菌株都需要 (Ikemoto et al., 1980;Marraro and Mitchell, 1974 )。S. maltophilia 生長溫度範圍從 4℃ 至 40℃,而最適 宜的生長溫度約為 35℃ (Joanna, 2012)。S. maltophilia 可以耐受極寬的 pH 範圍 (pH 4-10),但是鹽濃度就比大多數常見的實驗室細菌更加敏感 (<4.5%
NaCl);培養於極豐富的培養基中,S. maltophilia 分裂的周期大約是 30 至 40 分鐘,隔夜培養菌數列可達到 109 CFU/ml 以上 (Osama et al., 2014)。
2
表 一、S. maltophilia 的生長特性
生長特性 反應a
直桿或是彎曲桿狀,0.5 至 1.5 μm 氧化酶 (Oxidase)
過氧化氫酶 (Catalase)
蛋氨酸 (Methionine) 是生長必需 最適生長溫度 35°C
無生長 4°C 或 41°C 冷藏溫度的存活率 運動性 (Motility)
硝酸鹽還原性,但硝酸鹽不被用作氮源 吲哚 (Indole)
賴氨酸脫羧酶 (Lysine decarboxylase) 鳥氨酸脫羧酶 (Ornithine decarboxylase) 甲基紅 (Methyl red)
伏普反應 (Voges-Proskauer reaction) 硫化氫 (Hydrogen sulfide)
檸檬酸 (Citrate)
苯胺脫氨酶 (Phenylamine deaminase) β-半乳糖苷 (β-Galactosidase/ONPGb) 碳水化合物利用 (Carbohydrate utilization) 從麥芽糖產酸 (Acid production from maltose) 從葡萄糖酸生產酸 (Acid production from glucose) 生長碳源
核糖 (Adonitol) 阿拉伯糖 (Arabinose
β-羥基丁 (β-Hydroxybutyrate) 纖維二糖 (Cellobiose)
衛矛醇 (Dulcitol) 葡萄糖 (Glucose) 果糖 (Fructose) 半乳糖 (Galactose) 乳糖 (Lactose) 麥芽糖 (Maltose) 甘露醇 (Mannitol) 甘露糖 (Mannose) 鼠李糖 (Rhamnose) 水楊甙 (Salicin) 山梨醇 (Sorbitol) 海藻糖 (Trehalose)
七葉靈水解 (Esculin hydrolysis) 液化明膠 (Gelatin liquefaction) Tween 80 水解 (Tween 80 hydrolysis) DNA 酶生產 (DNase production) 澱粉水解 (Starch hydrolysis) 尿素水解 (Urea hydrolysis)
+/- + + + + + - + - - - - v - v + - - - - v - + v v + + - v - - - +/- + + + + - -
a +,>85% 菌株有效;v,16 – 84% 菌株有效;-,≦15% 菌株 有效;bONPG;o-nitrophenyl-β-D-galactopyranoside
(Joanna S. Brooke, 2012)。
3
乙、人體伺機性 (opportunistic ) 病原菌
S. maltophilia 是一人體伺機性病原菌,並可以形成生物膜附著在生物或 非生物 (例如醫療儀器) 的表面上,導致近年來醫院院內感染 S. maltophilia 的比例持續升高。S. maltophilia 在兒童和成人感染都曾出現過案例 (Joanna S. Brooke, 2012),Schable 等人亦報導過在重度病房醫護人員的手上分離出 S.
maltophilia 菌株,透過直接接觸傳染 (1991)。此外, S. maltophilia 菌株可 以藉由儀器傳播病菌感染免疫力不足和身體虛弱的病人,進而讓病人加重病 情,甚至發展成威脅生命的感染 (Giovanni et al., 2015)。
纖維囊腫(Cystic Fibrosis, CF)或黏液黏稠病 (mucoviscidosis) 是一種致 命的染色體隱性遺傳疾病,近年各地的 CF 中心發現 S. maltophilia 所導致 的患者數量持續上升 ( Cystic Fibrosis Foundation Patient Registry;CFFPR,
2012)。對於囊腫性纖維化患者,常見的是 S. maltophilia 引起的呼吸道感 染,並且讓肺部發炎加重與慢性阻塞性肺疾病急性的惡化 (Nseir et al.
2006)。於動物實驗中 S. maltophilia 感染小鼠會引起小鼠呼吸道上皮組織之 白介素-8 (Interleukin-8,IL-8) 的表現量提高和促使巨噬細胞表現腫瘤壞死 因子 (Tumor necrosis factor-α,TNF-α),促使小鼠肺部產生發炎 (Giovanni et al., 2010)。此外,S. maltophilia 也會導致人類血液感染而引發菌血症,以及 引發泌尿道感染、腹內感染、心內膜炎、腦膜炎和眼部感染等症狀 (George et al., 2012)。S. maltophilia 雖然不是高度有毒病原體,但其引發菌血症所造 成的患者死亡率高達 70% (王,2011)。如表二所示,George 等人於 2012 年 使用市面常見的 12 種抗生素對 68 株 S. maltophilia 病原菌進行抗菌測 試,發現多數抗生素都有 60% 以上的敏感性,這顯示 S. maltophilia 可被 多數抗生素抵制其生長,但在臨床治療上卻沒有如此高的效果 (George et al., 2012)。目前治療 S. maltophilia 感染的患者,大多數都使用混合型抗生素來 醫治,例如氟喹諾酮類藥物 (fluoroquinolones) 結合多黏菌素 (colistin) 或是
4
加 環 素 (tigecycline) , 來 替 代 之 前 的 甲 氧 芐 啶 / 磺 胺 甲 噁 唑 (trimethoprim/sulfamethoxazole) 的治療方案 (George et al., 2012)。
台灣院內感染監視資訊系統(TNIS系統) 是一個台灣監測研究單位,
此研究是在監測從病患中分離出的病原菌株比例。西元 2006 – 2015 年期 間,醫學中心加護病房不分年度前 4 名醫療照護相關感染菌種依序為 Candida species、Acinetobacter baumannii、Pseudomonas aeruginosa 及 S.
maltophilia。2015 年第 2 季醫療照護相關感染菌種前 5 名依序為 Candida species、Escherichia coli、Enterococcus faecium、Staphylococcus aureus 及 S.
maltophilia。
在 2006 – 2015 年期間,於醫學中心加護病房的肺炎患者分離出 S.
maltophilia 之排名大約都在 4 – 5 名之間;血流感染 (bloodstream infection) 患者 於 8 – 10 名之間。
表 二、12 種抗生素對 68 株 S. maltophilia 的藥物敏感性測試
抗菌藥物 S (%) I (%)
多粘菌素 Colistin 乙基西梭黴素 Netilmicin
甲氧芐啶/磺胺甲基異噁唑 Trimethoprim/sulfamethoxazole 氯黴素 Chloramphenicol
丁胺卡那黴素 Amikacin 環丙沙星 Ciprofloxacin 慶大霉素 Gentamicin 妥布黴素 Tobramycin 四環素 Tetracycline 頭孢他啶 Ceftazidime
替卡西林/克拉維酸 Ticarcillin/clavulanic acid
62 (91.2) 58 (85.3) 58 (85.3) 57 (83.8) 56 (82.4) 56 (82.4) 56 (82.4) 48 (70.6) 47 (69.1) 18 (26.5) 18 (26.5)
0 (0.0) 4 (5.9) 1 (1.5) 7 (10.3) 3 (4.4) 5 (7.4) 3 (4.4) 1 (1.5) 8 (11.8) 6 (8.8) 10 (14.7) S:敏感 (susceptible),I:中等敏感(intermediately susceptible) (George et al., 2012)。
5
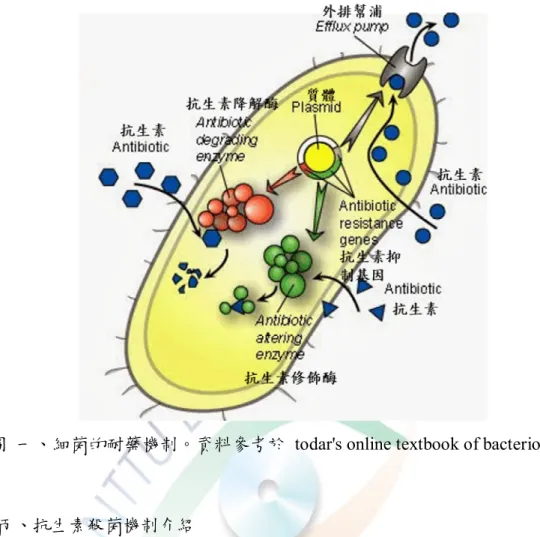
圖 一、細菌的耐藥機制。資料參考於 todar's online textbook of bacteriology。
丙、抗生素殺菌機制介紹
抗生素影響細菌的方式可分為下列四種,(1) 抑制細菌細胞壁合成,導 致細菌在低滲透壓環境下膨脹破裂死亡,主要以 β-lactam 類抗生素為主;
(2) 抑制細菌蛋白質合成,核糖體 (ribosome) 是細菌合成蛋白質的地方,是 由 30S 與 50S 兩個次單元蛋白組合而成,若 30S 或 50S 的核糖體次單 元受干擾,可影響細菌合成蛋白質的能力,如 tetracycline 和 erythromycin;
(3) 抑 制 細 菌 核 酸 合 成 , 有 些 抗 生 素 會 干 擾 DNA 隻 複 製 和 轉 錄 , 如 rifampin;(4) 影響細菌生長繁殖,抗生素透過抑制新陳代謝作用,讓細菌無 法生長,如 Penicillin。
丁、細菌抗藥機制分類
相對於上,細菌亦會產生抗藥性來抵抗抗生素作用,其抗藥性機制可分
6
成下述三類,(1) 合成抗生素降解酶,引起抗生素失活;(2) 表現藥物外排 幫浦,減少菌體類抗生素含量,將抗生素外排,或通過對菌體細胞壁增厚作 用,抑制抗生素進入菌體;(3) 產生抗生素修飾酶,與抗生素結合後改變其 構型,使得抗生素無法發揮藥效,例如鏈黴素修飾後,即不具與核糖體結合 的能力,因而不影響菌體蛋白質的合成 (圖一)。
戊、S. maltophilia 抗藥機制分類
一、合成抗生素降解酶
近年來抗生素使用普及,使得一些多重抗藥性的 S. maltophilia 菌 株於醫院中被篩選出來,這些菌株對市面上大部分的抗生素都有抗性的 效果,所以在治療上十分棘手。因 S. maltophilia 染色體 DNA 上具有 數個可抗抗生素的基因,如 β-lactamases 可水解 β-lactam 類的抗生素 (Lin et al., 2008;Hu et al., 2008;Spencer et al., 2001)。目前 β-lactamase 分子的分類根據是以酶的核苷酸和氨基酸序列分析為主,目前共分為四 類 (class A - D)。A、C 和 D 類的機制以絲氨酸 (Serine) 為主,B 類 或稱金屬類則需要鋅離子的輔助 (Ambler, 1980)。S. maltophilia 具有兩 個誘導型 β-lactamase,分別是 L1 和 L2。L1 是屬 B 類的鋅 2+ 依賴 性-β-lactamases ,而 L2 則是屬 A 類克拉維酸敏感的頭孢菌素分子 (class A clavulanic acid-sensitive cephalosporinase)。L1 和 L2 β-lactamase 是由 ampR (抗藥基因) 同時調節,轉錄調節位在 L2 的上游區域 (Okazaki and Avison, 2008)。ampR-L2 模組同源於 ampR-ampC 系統,
抗藥基因 (ampR) 調控 ampC 產生 β-lactamase ,此系統廣泛分佈於 腸桿菌科的菌株和綠膿桿菌 (Lodge et al., 1990)。但是與綠膿桿菌不同 的是, S. maltophilia 由 AmpN-AmpG 操縱子(啟動子 (ampN) 和內膜 通透酶 (AmpG) 所組成),調控 β-lactamase 表現 (Huang et al., 2010)。
7
二、外排幫浦 (Efflux pumps)
微生物利用外排幫浦排出抗生素的類型分為五種,分別為抗性結瘤 細胞分裂類(resistance-nodulation-cell-division, RND)、主要協調超族類 (major facilitator superfamily, MFS)、小多重抗藥類 (small multidrug resistance, SMR)、ATP 結合盒家族 (ATP binding cassette, ABC) 和耐藥 /有毒化合物排出類 (multidrug and toxic compound extrusion, MATE)。 S. maltophilia 目前的外排幫浦有四種類,分別為 RND 類、ABC 類、
MFS 類與三方鐮孢菌酸外排幫浦。
(一) RND 類
第一類 RND 外排幫浦在革蘭氏陰性菌是由三種蛋白質所構 成,其中內膜蛋白 (inner membrane protein) 為鍵結於基質,外膜 蛋白(porin,孔蛋白),和膜融合蛋白(membrane fusion protein, MFP),在週質空間中連結外膜蛋白和內膜蛋白。
S. maltophilia 是一種重要的人類伺機性病原體,具可以抵抗大 多數的抗生素之特色。S. maltophilia K279a 基因體序列分析顯示 基因體編碼多達六種已知的 RND 外排系統,分別是 SmeABC、
SmeDEF、SmeIJK、SmeOP,SmeVWX 和 SmeYZ 外排系統(Alonso and Martinez, 2000; Li et al., 2002; Crossman et al., 2008)。
SmeABC 主要與 β-lactam 類、氨基糖苷類 (aminoglycosides) 和喹諾酮類 (quinolones) 的抗生素抗性有關。SmeC (外膜蛋白,孔 蛋白) 功能獨立於 SmeAB (SmeA,膜融合蛋白;SmeB,內膜蛋 白),當 SmeC 基因缺失時會影響某些抗生素的敏感性,例如硝噻 (Nitrocefin) (Li et al., 2002)。
8
SmeDEF 是 一 個 複 合 型 外 排 幫 浦 , 主 要 是 由 內 膜 蛋 白 (SmeE)、外膜蛋白 (SmeF) 和周質膜融合蛋白 (SmeD) 所組成,
其參與氯黴素 (chloramphenicol)、四環素 (tetracyclines)、大環內酯 類 (macrolides) 和喹諾酮類 (quinolones) 的抗藥性,以及獲得抵抗 非抗生素化合物的能力(如二氯苯氧氯酚;triclosan (Alonso and Martinez, 2000;Sanchez et al., 2005; Hernandez et al., 2011)。目前研 究指出 S. maltophilia 感染植物時,植物根部會產生類黃酮來抵禦 細菌的感染,而菌體的 SmeDEF 外排幫浦可以排出植物根部產生 的類黃酮,使得 S. maltophilia 得以感染植物,若 SmeE 基因缺失 將會導致細菌感染植物的能力消失 (García-León et al., 2014a)。
SmeVWX 外排幫浦於 S. maltophilia KJ 主要含有 5 個基因 操縱子編碼蛋白,這些基因分別為 smeU1、smeV、smeW、smeU2 與 smeX。若 SmeVWX 過度表現可促成對氯黴素、喹諾酮和四環素 的抗性,但對氨基糖苷類的敏感性卻增加 (Chen et al., 2011)。
García-León 等人的研究中也證實,在 S. maltophilia 分離株中增 量表現 smeXVW 基因可以提高 S. maltophilia 抵抗喹諾酮類的抗 生素。將 smeV 和 smeW 兩個基因失活則會徹底導致 SmeVWX 外 排幫浦的功能性喪失,而單獨失活 smeX 則會降低 SmeVWX 外排 幫浦的功能 (Chen et al., 2011)。
外排幫浦系統 SmeYZ 和 SmeIJK 的相對基因為 smeY、
smeZ、smeJ 和 smeK 是在 S. maltophilia K M5 中被發現。SmeZ 有 助 於 提 高 細 菌 抵 抗 氨 基 糖 苷 類 抗 生 素 的 MIC (minimum inhibitory concentration) 值;SmeJ 和 SmeK 共同作用則可以提高細 菌抵抗四環素、米諾環素 (minocycline) 和環丙沙星 (ciprofloxacin) 的 MIC 值,並且可以增加抵抗左氧氟沙星 (levofloxacin) (Gould et
9
al., 2013)。外排幫浦除了可以排出藥物之外,將外排幫浦系統 SmeIJK 的基因失活時,細菌的細胞膜完整性會受損,影響多黏菌 素的抗性 (Huang et al., 2014)。在最新的研究指出 SmeYZ 可以對 細菌生理有所幫助,也有毒力相關的功能,包含了細菌的移動,鞭 毛形成,氧化壓力的敏感性,生物膜的形成和蛋白酶的分泌 (Lin et al., 2015)。
SmeO、SmeP 與 TolCSm (S. maltophilia 的 TolC 蛋白) 有關 聯性,TolCSm 是 RND 外排幫浦的配件之一,TolCSm 基因位於 smeOP 操縱子的上游。SmeOP 提供菌體對於氨基糖苷類、萘啶酸 (nalidixic acid)、多西環素 (doxycycline)、大環內酯類和某些不是 抗 生 素 化 合 物 , 例 如 羰 基 氰 化 物 3- 氯 苯 ( carbonyl cyanide 3-chlorophenylhydrazone, CCCP),結晶紫 (crystal violet),十二烷 基硫酸鈉(sodium dodecyl sulfate, SDS),和 tetrachlorosalicylanilide
(TCS) 具低敏感性 (Lin et al., 2014)。
此 外 TolCSm 協 同 SmeDEF 和 SmeYZ 外 排 幫 浦 與 trimethoprim / sulfamethoxazole 耐 藥 性 有 所 關 聯 (Huang et al., 2013b;Lin et al., 2015;Sánchez and Martínez, 2015)。
(二) ABC 類
ABC-類外排幫浦可調節疏水性藥物之傳遞與調節細胞膜內外 的脂質排出藥物,是革蘭氏陽性菌主要耐藥機制,但於革蘭氏陰性 生物體 ABC-型外排幫浦也慢慢被發現,例如,E. coli 中發現的 MsbA 和 MacAB (脂質翻轉酶)、Vibrio cholera 的 VcaM,Neisseria gonorrhoeae 的 MacAB 和 Serratia marcescens 的 SmdAB (Lin et al., 2014)。 S. maltophilia 菌株中兩個 ABC-類外排幫浦被發
10
現,分別是 SmrA 和 MacABCsm;SmrA 是 S. maltophilia 菌株中 第一個被發現的 ABC-類外排幫浦成員,主要排出的抗生素為喹諾 酮類和四環素 (Al-Hamad et al., 2009)。細菌內含有 MacABCsm 外 排幫浦會賦予對氨基糖苷類、大環內酯類和多黏菌素的耐藥性,也 會參與細胞膜的生成作用 (Lin et al., 2014)。
(三) MFS 類
EmrRsm 和 EmrCABsm 隸 屬 於 MFS Family 一 員 。 S.
maltophilia 基因的 emrCABsm 與 E. coli 的 emrAB 有較高的同 源性(Lomovskaya and Lewis,1992)。此幫浦是由四個基因操縱子 調控,EmrRsm 調節 EmrCABsm 參與疏水性化合物 (包括抗生素 萘啶酸和紅黴素) 的排出 (Huang et al., 2013)。
(四) 三方鐮孢菌酸外排幫浦
在 S. maltophilia 中發現一種新型的三方鐮孢菌酸 (Fusaric acid) 的外排幫浦即為 FuaABC,是三方外排幫浦的一個新亞家 族;該 fuaABC 操縱子被證明透過鐮孢菌酸誘導,增量表現 fuaABC 時會促進對鐮孢菌酸的抗性 (Hu et al., 2012)。
三、抗生素修飾酶
在 S. maltophilia 中耐氨基糖苷類機制主要是以氨基糖苷類修飾酶 和外排幫浦為主。氨基糖苷類修飾酶包含了 AAC(6′)-Iz (氨基糖苷類乙 酰) (Li et al., 2003)、APH(3′)-IIc (氨基糖苷磷酸) (Okazaki and Avison, 2007) 和 AAC(6′)-Iak,這是最新於尼泊爾發現的一株多重抗藥菌株 (Tada et al., 2014)。
11
貳、噬菌體療法 (phage therapy)
近年來因為抗生素的濫用,導致環境中細菌出現多重抗藥性或抗藥菌株,因 此研究學者們開始尋找其他的療法來殺死細菌,而其中以噬菌體療法 (phage therapy)備受注目,此療法為利用一種具有專一性的噬菌體來殺死病源菌抵制細 菌生長的治療方法 (Levin & Bull, 1996; Lederberg, 1996; Barrow & Soothill, 1997;
Alisky et al., 1998; Kutter, 1998)。噬菌體療法的實踐跟噬菌體被發現息息相關,
在西元 1915 年至 1917 年就有發現噬菌體的案例出現。但是隨著抗生素的發 現、大量研發和應用,大多數的噬菌體研究就漸漸停止運作,只有少數國家 (東 歐和蘇聯),噬菌體研究和抗生素研發是同時並存 (carlton, 1999)。因為多重抗藥 菌株抵抗抗生素因素,所以噬菌體殺死細菌十分熱門。
噬菌體療法具有比抗生素治療更佳的特性,例如: (1) 噬菌體具有極高的宿 主專一性特徵,只會針對特定的細菌進行抑制細菌生長或殺菌,比較不會破壞人 體內部正常細菌族群平衡性,但是抗生素就沒此特性;(2) 如果成功感染,溶裂 型噬菌體將會裂解細菌,造成菌體死亡。相較之下,某些抗生素只能抑制細菌的 生長而無法殺死細菌,如四環黴素(tetracycline);(3) 對抗生素產生抗藥性的 細菌,對於同類型抗生素依舊會有抗藥性,但是抗噬菌體之細菌,對於相似之噬 菌體可能依舊是敏感的;(4) 發現一隻新的噬菌體所花費的時間可能只要數天至 一個多月的分離與純化,但是研發一個新的抗生素可能需花費幾年的時間才可能 完成 (Loc-Carrillo & Abedon, 2011)。以上特性使得噬菌體療法更具有研發潛力。
甲、噬菌體療法於動物實驗
至目前為止,已有許多有關噬菌體療法的動物試驗研究報告發表,舉例 如下: (1) 由於致病菌導致水產養殖魚類幼魚死亡,尤其是高死亡率的 Vibrio anguillarum (V. anguillarum),目前尚未無有效的預防方法。如於水產 養殖中添加噬菌體將是一種適合對抗 V. anguillarum 的方法,Yolanda 等人
12
直接將噬菌體添加入養殖水中發現對於提高幼魚存活率是有益的 (2014)。(2) 於 Pseudomonas aeruginosa 所感染的鯰魚中,添加噬菌體處理 8-10 天,
顯示噬菌體療法可有效治癒 P. aeruginosa 所造成的魚潰爛病,比起未接受 噬菌體處理的感染魚存活率提高了 7 倍 (Krishna et al., 2013)。(3)另外,由 P.
aeruginosa 所引發狗的中耳炎,在治療過程中,施用 P. aeruginosa 溶裂型 噬菌體混合物,於犬耳內,並無明顯的毒性產生 (Catherine et al., 2010)。(4) 耐甲氧西林金黃色葡萄球菌 ( Methicillin-resistant Staphylococcus aureus, MRSA)是動物骨髓炎的最常見的原因。由 MRSA 所感染的 20 隻骨髓炎 兔子,在持續兩週的噬菌體治療實驗組中,兔子可有效治癒,並且兔子身上 的局部水腫、紅斑和硬塊也消退,並且提高了兔子的食慾和活動量,顯示噬 菌體療法具有抵抗多重抗藥細菌 (Kishor et al., 2016)。
乙、噬菌體療法於人體實驗
噬菌體無所不在,在任何地方都可能存在噬菌體。文獻指出從人類唾液 中分離出的噬菌體對糞腸球菌有抑制生長的影響,因此可以考慮用噬菌體療 法作為一種控制口腔細菌的方法 (Bachrach et al., 2003)。噬菌體療法在人體 試驗方面也有一些研究報告,大部分的人體試驗病例都是細菌引發慢性疾病 的案例,並且細菌對所有現有的抗生素都呈現抗藥性,並且於其他治療方法 都失敗了以後,才開始使用噬菌體療法來治療病人。在國外有三名工人因受 傷而感染 S. aureus,住院治療後,對患者使用各種藥物,包括抗生素和外 用藥膏治療,雖傷口能癒合,但體內的 S. aureus 的感染卻不能完全被消 除,所以最終處以施打抗生素與噬菌體混合劑七天後,才有效降低體內金黃 色葡萄球菌的含量 (Jikia et al., 2005)。另外,人體在皮膚受損時,P.
aeruginosa 是常見引起傷口感染的菌株之一,此菌株也會引起其他疾病,如 肺炎、菌血症、腦膜炎和尿路感染;於人體皮膚試驗中,發現噬菌體 PA709 可有效地失活多重耐藥菌株 P. aeruginosa 709 (Vieira et al., 2012)。而在開發
13
中的國家,孩童因 Escherichia coli (E. coli) 引發的腹瀉的案例十分普及,多 數的抗生素也無法治療此腹瀉,並且沒有高效率的 E. coli 疫苗 (Ahmed et al., 2013)。在孟加拉,給急性細菌性腹瀉住院兒童口服 T4-like 噬菌體、商 業俄羅斯大腸桿菌噬菌體產品或安慰劑,4 天後。結果顯示口服噬菌體藥物 沒有不良現象發生,但也未能改善改善腹瀉,推測口服噬菌體量不足以抑制 E. coli (Shafiqul et al., 2016)。
參、噬菌體
甲、噬菌體構造與分類
噬菌體(bacteriophage 或 phage)簡單的定義就是可以生長在細菌細胞 的病毒,經常都伴隨着細菌,通常在一些充滿細菌群落的地方,如泥土和動 物的腸臟裡,都可以找到噬菌體的蹤影。目前世上蘊含最豐富噬菌體的地方 就是海水,在海平面平均每毫升的海水含有 109 個病毒粒子(virions),且海 水中高達七成 的細菌可受到這些噬菌體的感染 (Wommack and Colwell, 2000)。
不同的噬菌體有不同的外形構造,基本上噬菌體的構造不外是核酸分 子 , 外加 一個 外殼蛋白 (coat 或 capsid) 保護,使核酸 不被核酸 酵素 (nuclease) 或 有 害 物 質 傷 害 ; 噬 菌 體 的 核 酸 分 子 可 以 是 單 股 或 雙 股
(single-stranded or double-stranded)、直線或環型(linear or circular)DNA,
或 RNA。
噬菌體依照其繁殖過程可以分成兩類,溶裂型噬菌體與潛溶型噬菌體 (lytic and lysogenic phage)。如圖二所示,溶裂型噬菌體感染細菌後,將噬菌 體遺傳物質送至細胞內,並破壞宿主的遺傳物質後,控制被感染的細菌製造
14
噬菌體所需的蛋白質和核酸;當細胞內噬菌體顆粒組裝完成後,噬菌體釋放 溶裂素 (lysozyme),細菌就被溶解(lysis)而破裂死亡,釋放出眾多的噬菌 體子代來感染其他細菌,代表性噬菌體如 T2、T4 和 T7。另一種潛溶型噬 菌體是比較溫和性 (temperate) 的噬菌體,它可以進行溶裂型或潛溶型的 生活史,在潛溶時期不會產生噬菌體子代,而是會將自己的 DNA 插入到宿 主的 chromosome DNA 上,進入休眠狀態,細菌的 chromosome DNA 複製 也會隨之複製;而當受到一些誘導因子的影響 (例如 UV 光照射) 或處於不 利於生長因子中時 (例如環境乾燥或是寒冷),噬菌體將會停止潛溶時期轉 而進入溶裂時期,這類型的代表性噬菌體包含了噬菌體 λ、P1 和 P2(Maloy et al., 1994)。
目前發現的噬菌 體 , 大多數 都歸屬於 有尾 噬菌體 目 Myoviridae、
Siphoviridae 和 Podoviridae 三 科 中 , 少 部 分 屬 於 Inoviridae 和 Microviridae 科。Myoviridae 科的噬菌體外觀型態具有頭部構造以外,有一 個可以伸縮的尾部,例如噬菌體 T4;Siphoviridae 科的噬菌體有頭部構造 以外,另具無法伸縮的長尾,最具代表的噬菌體為 λ;Podoviridae 科的噬 菌體外觀型態為具有頭部構造和十分短的尾部結構,以噬菌體 T7 最為大家 所 熟 知 (chen et al., 2007) 。 Inoviridae 科的 噬 菌體 外觀 型態 為 直現 型
(filamentous),如 M13;而 Microviridae 科噬菌體外觀型態則有一二十面 體的頭部(icosahedral hesd),但無尾巴(tailless),代表性噬菌體為 ψ-X174 (圖三)。
15
圖 二、噬菌體的繁殖過程可以分成兩類,溶裂型噬菌體與潛溶型噬菌體 (lytic and lysogenic phage)。
圖 三、噬菌體 T4-like virus、噬菌體 HK97 和噬菌體 P22 分別是 Myoviridae, Siphoviridae, and Podoviridae 科 (Mart et al., 2011)、噬菌體 M13 為 Inoviridae 科 (Karl Johnson, 2011) 和噬菌體 ψ-X174 為 Microviridae 科 (Sanger et al. , 1978)。
16
乙、S. maltophilia 的噬菌體
目前已被報導的 S. maltophilia 噬菌體共有 14 株,相關訊息如下。最 早被報導的溫和型噬菌體 ψSMA5 在電子顯微鏡下觀察其外型,顯示它類 似於 Myoviridae 噬菌體,其基因體為雙股 DNA,大小約為 250 Kb;在於 台灣醫院 中所收集的 87 株 S. maltophilia,只有 61 株可以被噬菌體 ψSMA5 所影響 (Chang et al., 2005)。另有報導指出 S. maltophilia 噬菌體 ψSMA9 具有約有 6.9 kb 大小的基因組,內包含 7 個基因,其中最大的基 因編碼相似於霍亂弧菌的絲狀噬菌體 ψLf 的毒素基因 Zot (Hagemann et al., 2006 )。 溶 裂 型 噬 菌 體 Smp14 於 電 子 顯 微 鏡 觀 察 其 外 型 , 類 似 Myoviridae 噬菌體的成員,其基因體大小約 150 kb,於 87 株臨床分離菌 株中可裂解 37 株菌 (chen et al., 2007)。T7-like 噬菌體是一類溶裂型噬菌 體,相較於其他類型的噬菌體,其基因體功能大多數都已解序分析並有較小 的基因組;S. maltophilia 噬菌體 IME15 是屬於 T7-like 的噬菌體,其基因 體大小為 38,513 bp,GC 比為 53.7%,IME15 基因體具有 45 個 ORFs,
其中 41 ORFs 與已知的蛋白具有同源性 (Huang et al., 2012);另一 S.
maltophilia 噬菌體 IME13 完整的基因組長度為 162,327 bp,其 GC 比為 41.2%,具 182 個 ORF 和 15 個 tRNA,其中 95 個 ORFs 為已知的基 因 (Huahao et al., 2012)。S. maltophilia 噬菌體 φSHP1 於電子顯微鏡觀察,
得知其屬於 Inoviridae,全長 2.1 μm,基因體長度為 6,867 bp 的環型單股 DNA,具有十個 ORFs ,其中六個與已知蛋白有同源性 (Liu et al., 2012)。
兩隻絲狀噬菌體 phiSMA6 和 phiSMA7 是從 S. maltophilia Khak84 被分 離出來,其基因體分別為 7,648 bp 和 7,069 bp,都有 11個 ORFs;這兩隻 噬菌體對於從環境 (例如湖水和凍土) 分離出的 17 株 S. maltophilia 細菌 的抑菌效果不佳 (抑菌率分別是 23% 和 11%) (Petrova et al., 2014)。
17
溫和型噬菌體 S1 基因體大小為 40,287 bp ,具有 48 個 open reading frames (ORFs) 與長度為 12 bp 的 5 端突出結合性末端。噬菌體 S1 與噬 菌體 ψSMA5 的高感染率 (70%) 相較下,噬菌體 S1 因其 DNA 容易被切 割酶切斷而失去功能,所以能夠感染 S. maltophilia 的能力只有 4/26 株(15
%)(Pilar et al., 2008)。另噬菌體 S3 和 S4 基因體分別為 33 kb 和 200 kb,其基因體對常見的切割酶 (BamHI 、EcoRI、HindIII 和 XbaI 等切割 酶) 有抗性,相較噬菌體 S1 有較高的宿主範圍。作者認為這個特性可能有 助於噬菌體具有較廣泛的宿主範圍 (Pilar et al., 2008)。溫和性噬菌體 Smp131 是從 S. maltophilia T13 中被分離出來,可以感染其他三株 S.
maltophilia (ATCC 13637 、 BCRC 11901 和 T16) ; 研 究 並 指 出 噬 菌 體 Smp131 其基因體為 33,525 bp ,具有 47 個 ORFs。;噬菌體 Smp131 根 據基因體序列分析其基因組結構、操縱子的排列和在尾部基因具有一個不穩 定 的 翻 譯 碼 序 列 T7G , 等 特 性 推 斷 此 噬 菌 體 與 P2-like 噬 菌 體 和 S.
maltophilia 與 Xanthomonas 屬中的 prophage 相似 (Lee et al., 2014)。
丙、具廣泛宿主範圍的 S. maltophilia 的噬菌體
噬菌體 DLP1 和 DLP2 是從 S. maltophilia D1585 分離出。兩隻噬菌 體的基因組長度分別為 42,887 bp 和 42,593 bp,GC 含量都是 53.7%。噬 菌體通常只能感染相關的細菌種類,而且很少感染橫跨分類科的細菌,但這 兩 隻 噬 菌 體 DLP1 和 DLP2 表 現 出 異 乎 尋 常 的 廣 泛 宿 主 範 圍 (S.
maltophilia 和 P. aeruginosa);P. aeruginosa 是株人類伺機性病原菌可感染 人體肺部及泌尿道。(Danielle et al., 2015)。
丁、可感染 Xanthomonas 屬的噬菌體
早期 S. maltophilia 被歸類於 Xanthomonas 屬,因此 S. maltophilia 噬 菌體可能會與 Xanthomonas 屬噬菌體相近。絲狀噬菌體 XacF1 可以感染 Xanthomonas axonopodis pv. citri (Xac) 菌株,噬菌體 XacF1 以電顯觀察其
18
外型屬於 noviridae 科,顆粒長度約 600 nm,基因體長度為 7,325 bp,具 有 13 個 ORFs。噬菌體 XacF1 可感染菌的能力很高,研究中測試了 11 株 Xanthomonas axonopodis pv. citri (Xac) 可以感染 7 株菌株。細菌宿主受到 噬菌體 XacF1 感染會引起生理變化,包括胞外多醣的生產減少、細菌活力 降低、生長速度較慢和毒力顯著下降等變化。其中在毒力的降低部分顯示噬 菌 體 XacF1 具 作 為 柑 桔 潰 瘍 病 的 生 物 控 制 劑 潛 力 。 另 一 株 可 感 染 Xanthomonas campestris pv. campestris (Xcc) 的裂解型噬菌體 phiL7,其形態 上屬於 Siphoviridae 科家族,包含線性雙股 DNA 基因組(44,080 bp,56%
GC 比),有一個 3'-突出 COS 位(5'-TTACCGGAC-3')和 59 個可能基因 (Lee et al., 2009)。
肆、可感染 S. maltophilia 噬菌體 smp14
前人在篩選噬菌體時,於高雄醫院廢水分 離噬菌體,之後 培養 S.
maltophilia T14 中,所以命名為 Smp14,屬溶裂型噬菌體。噬菌體 Smp14 在 87 株台灣院內 S. maltophilia 分離株中抑菌比例為 56% (49/87),殺菌 比例為 32%。利用穿透式電子顯微鏡,觀察噬菌體外觀型態,顯示噬菌體 具有平面六角形頭部,可以伸縮尾部構造,屬於 Myoviridae。噬菌體 Smp14 基因體長度為 159,910 bp,GC 比為 54%,推測 Smp14 基因體具有 20 tRNA 基因,230 ORFs,其中 70 個 ORFs 與已知功能之蛋白質具同源性,
40 個 ORFs 與 hypothetical proteins 具同源性,其他 120 ORFs 為 Smp14 基因體上 unique hypothetical genes (陳,2007) 。前人分析噬菌體 Smp14 基 因體序列得知歸類於 T4-like 噬菌體,噬菌體 Smp14 基因體除包含了 T4-like 的噬菌體基因,也包含了一些少數其他類型噬菌體的同源基因,例 如 ORF80 為 弧 菌 噬 菌 體 VP4 lysozyme 同 源 基 因 。 VP4 於 NCBI database 中是 unclassified T7-like 噬菌體。顯示噬菌體 Smp14 的 lysozyme
19
基因親緣性較接近於 T7-type 的 lysozyme,而非 T4-type 的 lysozyme。
噬菌體的溶菌相關蛋白主要有三個:lysozyme、holin 和 hydrolase。前 兩種蛋白通常會相互配合以提高溶菌作用,lysozyme 無法單獨穿越細胞 膜,所以通常藉由 holin 蛋白的協助通過細胞膜到達細胞壁,接觸到細胞壁 後進行分解作用 (Wang et al., 2000)。因 holin 蛋白具有穿透模的功效,不同 類的 holin 蛋白其結構上的 transmembrane doain (TMD) 的數量不同,holin 共可分成 3 類:Class I holin 具有 3 個 TMDs;Class II holin 具有 2 個 TMDs 和 Class III holin 具有 1 個 TMDs (Ry Young, 2002)。T7-type holin 屬於 Class II holin 具有 2 個 TMDs,大小約為 64 至 71 aa ,前人經由 程式分析噬菌體 Smp14 ORF194 具有 66 aa,推測為噬菌體 Smp14 的 holin 基因 (陳,2007)。
伍、常見的 DNA 突變方法介紹 甲、紫外線光(UV)突變法
UV (非電離輻射) 對 DNA 上的核苷酸鹼基胞嘧啶 (C) 和胸腺嘧啶 (T) 最容易利用輻射改變它們的屬性。UV 光可誘導相鄰的嘧啶鹼基在 DNA 鏈上形成共價結合的嘧啶二聚體 (圖四),此嘧啶二聚體於 DNA 複製和基 因表現時會阻礙複製與轉錄作用,直到其被移除。UV 也可能導致 DNA 產 生氧化損傷現象,尤其是波長較長的 UVA (圖四),首先 UV 光使兩個相鄰 的胞嘧啶殘基形成二聚體,胞嘧啶二聚體可能導致腺嘌呤於 DNA 複製期間 被加入至新股合成,隨後的 DNA 複製會產生鹼基序列 CC 變成 TT 造成 突變。雖然胞嘧啶二聚體可能會被修正,但新合成 DNA 的突變不會被 DNA 修復系統檢測出。Kozmin 等人觀察 UVA 輻射對於酵母菌 Saccharomyces cerevisiae 的存活和誘變是否會有影響,結果顯示紫外線輻射的致命作用會 對酵母細胞的 DNA 修復途徑產生不敏感作用,如鹼基切除修復 (base
20
excision repair, BER)、核苷酸切除修復 (nucleotide excision repair, NER)、同 源重組 (homologous recombination, HR)、複製後的修復或是 translesion DNA synthesis (TLS) 的失活作用。此外,UV 亦可以強烈誘變 DNA,形成氧化 性 DNA 損傷,對於人類的皮膚的惡性轉化可以做為參考依據 (Kozmin et al., 2005)。
21
圖 四、UV 所引起的 CC 嘧啶二聚體突變影響。UV 可能造成的突變機制,於 DNA 複製時 CC 二聚體序列突變成 TT 序列 (Molecular Biology Web Book)。
22
乙、藥劑突變法
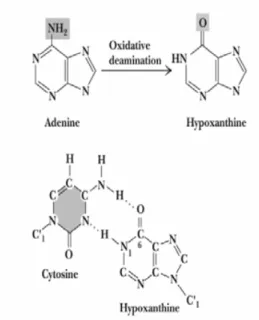
可導致 DNA 突變的化學劑稱為誘變劑。大多數的誘變劑也是致癌物 質 。 於 DNA 突 變 實 驗 上 , 烷 化 劑 (dimethyl sulfate 與 methyl trifluorosulfonate) 可將烷基 (甲基或乙基) 加到嘌呤或嘧啶的 N 或 O 上使 鹼基烷基化,其中以鳥嘌呤 (G) 和腺嘌呤 (A) 最容易受攻擊,形成烷基化 的嘌呤鹼基,導致複製時鹼基配位錯誤。Yarosh 於 1985 年發表 DNA 鹼 基 烷化形成 O6-methylguanine 會 造 成 DNA 產 生 突 變 和 致 癌 作 用 ( 圖 五)。原本鹼基 A 與 T 配對,亞硝酸 (nitrous acid) 與含胺基的鹼基反應,
使得 A 被氧化,進行去胺反應變成為亞黃嘌呤 (hypoxanthine);而於 DNA 複製期間,亞黃嘌呤與鹼基 C 鍵結,而不會與 T 鍵結,造成鹼基 A 突變 成 G (圖六)。
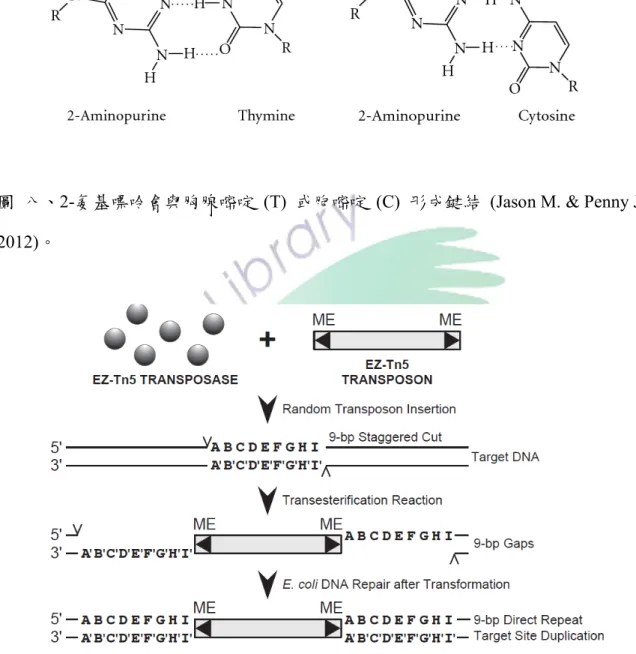
鹼基類似物 (Base analogs) 是化學物質可以代替正常的鹼基的核酸,當 添加鹼基類似物時,在 DNA 複製過程中將因改變鹼基配對的結果而導致置 換突變。如 5-溴尿嘧啶 (5-bromouracil;5-BU) 為胸腺嘧啶 (T) 的相似物,
正常情況下鹼基 T 與 A 配對,也會與 G 形成氫鍵;在 DNA 複製期間,
添加 5-溴尿嘧啶後,DNA 複製,但 AT 鍵結突變成 CG 鍵結 (圖七)。如 圖八顯示,2-氨基嘌呤 (2-aminopurine) 通常與鹼基 T 配對,但也可與鹼基 C 形成單一的氫鍵 (Griffiths et al., 2005)。
丙、跳躍子 (transposon) 突變法
DNA 在換位 (transposition) 的過程中會產生各種基因重排,如插入、
缺失 、 倒位 和染 色體融 合等 情形 。 DNA 的換位 過程 ,一 個跳躍 蛋白
(transposase protein, TNP)會介導基因組的遺傳信息的移動 (移動 DNA 區段或跳躍子)。跳躍子已廣泛於生活中被發現 (Craig et al., 2002)。這些跳 躍子可以攜帶重要因子 (例如抗藥基因) 來幫助細菌適應新環境 (Davies,
23
1994)。逆轉錄病毒家族蛋白包括 Tnps、逆轉錄病毒整合酶和 RAG-1,跳躍 蛋白 TNP 是其中一員。逆轉錄病毒家族蛋白質催化核心 (catalytic cores) 的結構是具保守性的,蛋白質功能的化學機制亦是十分相似 (Davies et al., 1999; Rice and Baker, 2001)。跳躍子 Tn5 系統是一個研究 DNA 換位的模 組系統,Tn5 系統是一個相對簡單的系統,只需要 Tn5 跳躍蛋白 TNP、Tn5 DNA、目標 DNA、Mg2+ 和水,就可以進行目標 DNA 的突變,此系統已 有很多的遺傳、生化和結構的數據報導 (Richard et al., 2008)。而 EZ-Tn5 系 統比 Tn5 系統有更高的轉錄活性,系統造成 DNA 突變的方式如圖九所 示,TNP 會在目標 DNA 隨機作用造成 DNA 切割,跳躍子將會插入切割 位,經由 DNA 修復後產生突變 DNA。
於 2010 年,Martin 等人為了探討肺炎克雷伯菌的耐熱機制,利用 EZ-Tn5 <oriV/KAN-2> 系統將肺炎克雷伯菌的基因體片段隨機突變後,轉 殖入大腸桿菌中,獲得克雷伯菌的基因片段突變株。再將這些突變株進行耐 熱實驗,經耐熱篩選後得知克雷伯菌的 clpK 基因會影響大腸桿菌的耐熱 性,所以作者認為 clpK 基因是肺炎克雷伯菌耐熱性基因 (Martin et al., 2010)。此外,為了探討人類腸道微生物的膳食纖維分解酶的功效,Lena 等 人將人類糞便中細菌染色體片段接至質體並轉殖入大腸桿菌中,再利用 EZ-Tn5 <oriV/KAN-2> 系統隨機突變,產生含糞便細菌突變基因的大腸桿 菌,並建立基因庫後,分析這些基因功能性 (Lena et al., 2010)。另有生物學 家 Phillip 等人想了解胃腸細菌如何存活於有袋動物體內,於是將袋鼠胃腸 內容物的細菌分離後,抽取其染色體 DNA,並將染色體片段轉接至 pCC1Fos 中,再利用 EZ-Tn5 <oriV/KAN-2> 系統得到大量的突變 DNA,
之後利用電穿孔法將這些含突變 DNA 的 pCC1Fos 轉殖至大腸桿菌 EPI300,進行細胞膜形成實驗,得知含溶菌酶 (Muramidase) 會影響細胞聚 集和生物膜的形成 (Phillip et al., 2010)。由於抗生素的濫用或誤用加速抗生
24
素耐藥性細菌的生成。因此,Tong Zhang 等人想探討汙泥中抗藥細菌比例,
於是提取汙泥細菌的總 DNA 後,利用 EZ-Tn5 <oriV/KAN-2> 搭配大腸桿 菌 EPI300 系統得到大量的含突變 DNA 的大腸桿菌,之後再將這些突變株 DNA 與耐藥性數據庫分析比對,得知汙水處理廠的汙泥具有大量的抗藥菌 株,進而成為生成抗藥菌株的溫床 (Zhang et al., 2011)。探討節桿菌分離株 JBH1 對於碳和氮的利用,利用 EZ-Tn5 <oriV/KAN-2> 系統獲得 JBH1 分 離株的突變 DNA,分析得知 pfvC 基因所形成的蛋白會影響細菌對於碳和 氮的利用 (Johana et al., 2012)。
圖 五、鳥嘌呤 (G) 受到烷化形成 O6-methylguanine,在 DNA 複製過程,最終會造 成 GC→AT 置換。
25
圖 六、亞硝酸會將腺嘌呤 (A) 去胺化成次黃嘌呤,鍵結的鹼基將從胸腺嘧啶 (T) 換 成胞嘧啶 (C)。
圖 七、5-溴尿嘧啶誘變機制。這種分子有兩種互變異構亞型,酮形式(BUk)與鹼基 A 鍵結,而烯醇式(BUe)與鹼基 G 鍵結,因而在 DNA 複製期間,會造成 AT 鍵結 變成 GC 鍵結 (Molecular Biology Web Book)。
26
圖 八、2-氨基嘌呤會與胸腺嘧啶 (T) 或胞嘧啶 (C) 形成鍵結 (Jason M. & Penny J., 2012)。
圖 九、EZ-Tn5 系統造成 DNA 突變的複製過程。突變需要目標 DNA、跳躍酶與跳 躍子,跳躍酶會隨機將目標 DNA 切割,跳躍子插入切割位,待 DNA 修復後生成突 變 DNA (EPICENTRE EZ-Tn5™ <oriV/KAN-2> Insertion Kit)。
27
陸、反義 (antisense) RNA 干擾系統
在 細 菌 中 基 因 表 達 是 由 許 多 不 同 的 機 制 進 行 調 節 。 其 中 , 轉 錄 作 用 (transcrption) 的調控主要是藉由蛋白質或 RNA 所控制。在許多真核系統中,反 義 RNA 和 RNA 干擾 (RNA interference, RNAi) 介導的基因沉默是常見的,而 在細菌的 RNAi 機制報導卻十分少 (Nobutaka et al., 2006)。反義 RNA 的作用為 標靶 RNA 與互補以阻止轉譯作用 (translation),並誘導 mRNA 降解,進而抑 制 transcripsome 的引子鍵結或誘導標靶 RNA 的轉錄提前終止。表現反義 RNA
(asRNAs) 似乎是適合讓細菌條件性基因沉默 (Yin D and Ji Y., 2002;Engdahl et al., 1997)。
研究動機
因 S. maltophilia 噬菌體 Smp14 有許多功能性未知的 ORFs,為探討其功 能性,預計以突變互補實驗確認其 ORF 功能性。以往突變反應需要在菌體內進 行,但最新技術 EZ-Tn5 系統可於體外利用跳躍子將噬菌體 Smp14 DNA 進行 突變。本實驗室曾將是菌體 Smp14 DNA 以 EZ-Tn5 系統突變後,運用電穿孔 方式轉殖入 E. coli EPI300 中,卻造成 E. coli EPI300 溶裂死亡,推測噬菌體 Smp14 溶菌相關基因於 E. coli 系統中表現之故,因此欲藉由表現 antisense RNA 表現方式來抑制噬菌體溶菌基因 holin 與 lysozyme mRNA 的轉譯作用,
以利未來應用於噬菌體突變株庫的建立,進而可探討噬菌體未知 ORF 的功能 性。
28
研究目的
為建構噬菌體 Smp 14 holin 和 lysozyme 的 antisense RNA 系統平台,首 先利用電腦程式 (vector NTI) 分析 antisense RNA 系統的三個元素 (啟動子,反 股 DNA 和終止子);再利用 PCR 技術將目標 DNA 擴增,將目標 DNA 逐步 轉接至適當載體上後,於 E. coli 平台上建立 antisense RNA 系統,應用於抑制 噬菌體 Smp14 lysozyme 和 holin 溶裂基因的表現,之後利用電穿孔法將已突變 的噬菌體 smp14::EZ-Tn5 DNA 轉殖入此平台中,藉由菌內 antisense RNA 系統 表現可抑制噬菌體 Smp14 溶菌基因的 antisense RNA,進而讓 E. coli EPI300 不會溶裂死亡。進而於此平台建立噬菌體 Smp14 突變株基因庫,之後可應用於 未來噬菌體 smp14 ORFs 基因功能性分析研究。
29
材料方法
壹、研究設備及器材 一、菌種、噬菌體與質體
本實驗所使用的菌種為 E. coli DH5α,E. coli EPI300,Stenotrophomonas maltophilia (S. maltophilia ) ,S. maltophilia 之噬菌體 Smp14。
二、一般藥品
本實驗所用的藥品大多數購自 AMRESCO。DNA 回收和質體抽取所需 的 Kit 購自波士特生物科技公司和 Favorgem Biotech CORP.。抗生素購自 AMRESCO 。TA vector 和勝任細胞購自波士特生物科技公司和益生生技公 司。
三、酵素
限制酶購自 Yeastern Biotech CO., Lid 。Taq DNA polymerase 購自波士 特生物科技公司。T4 DNA Ligase 購自 New England BioLabs。
四、引子 (primer)
RCR 和 RT-PCR 實驗中所使用的引子列於表四。
五、培養基
LB medium: 每公升含有 10 g tryptone,5 g yeast extract and 5 g NaCl 六、抗生素使用濃度
1. Ampicillim: 25 μg/ml 和 50 μg/ml 2. Kanamycin: 50 μg/ml
3. Tetracycline: 15 μg/ml 七、試劑與緩衝液
1. 抽取質體 DNA 試劑
(1) Solution I: 25 mM Tris-HCl (pH 8.0), 10 mM EDTA and 50 mM
30
glucose。
(2) Solution II: 0.2 N NaOH and 1% SDS。
(3) Solution III: 60 ml 5 M Potassium acetate, 11.5 ml Glacial acetic acid and 28.5 ml H2O
2. 噬菌體保存劑
(1) SM buffer: 5.8 g NaCl, 2 g MgSO4•7H2O, 50 ml Tris-HCl (pH 7.5) and add ddH2O to 1000 ml。
3. DNA 電泳試劑
(1) 0.5 X TAE buffer: 20 mM Tris-acetate and 1 mM EDTA (pH 8.0)。
(2) 6 X loading dye: 0.25% Bromophenol blue and 30% Glycerol。
(3) Staining buffer: 0.5 μg/ml ethidium bromide (EtBr)。
4. β-galactosidase 活性測試
(1) 1 X assay buffer: 8.04 g Na2HPO4•7H2O, 2.76 g NaH2PO4•H2O, 0.37 g KCl, 0.12 g MgSO4•7H2O and β-mercaptoethanol
(2) O-Nitrophenyl-β-D-galactopyranoside: 19.5 ml 0.2 M KH2PO4, 30.5 ml 0.2 M K2HPO4, 0.4 g o-nitrophenyl-β-d-galactopyranoside add ddH2O to 100 ml
(3) Stop buffer: 10.6 g Na2CO3 add ddH2O to 100 ml 5. RNA 純化試劑
(1) Stop solution: 95% ethanol, 5% Phenol
(2) TEN buffer: 500 mM NaCl, 100 mM Tris-HCl (pH 8.0), 20 mM EDTA (pH 8.0) Adiust to pH 9.0
(3) 4 M LiCl DEPC H2O: 1 ml DEPC and 1000 ml dd H2O
31
貳、研究方法
一、菌種之培養及保存
依照陳,2007 論文方法修改,將欲分析的菌種先進行單株分離後,挑 取個單一菌落接種於 6 ml 的 LB 培養液中培養。E. coli 置於 37℃ 培 養。假如培養之菌株含有抗生素基因時,需於培養液中(10 g trypton, 5 g yeast extract and 5 g NaCl/L),並加入適量的抗生素分別為: Ampicillim 25 μg/ml;
Ampicillim 50 μg/ml;Kanamycin 50 μg/ml。如需長期保存菌種則取隔夜培養 之菌液,加入最終濃度為 15 % 之 glycerol,混和均勻後保存於 -80℃ 冰 箱。
二、噬菌體培養
1. 小量噬菌體培養
依照陳,2007 論文之方法修改,從噬菌體培養平盤上挑選一 顆單一溶菌斑進行培養,將此溶菌斑加入至新鮮的菌液培養基 (OD600 > 0.7),放置於 37℃ 培養箱震盪培養,培養到菌液澄清 (約 5 - 7 小時),再以 12,000 g 離心 10 分鐘,取其上清液經由 filter (0.45 μm) 過濾,其液體保存於 4℃。
2. 大量噬菌體培養
依照陳,2007 論文之方法修改,取 50 ml 隔夜培養菌液,加 入至 450 ml 新鮮 LB 培養液中,於 37℃ 培養至 OD600 為 0.3 左右,加入 MOI 比例為 1:100 的噬菌體,培養至菌液澄清。以 12,000 g 離心 30 分鐘,取其上清液經由 filter (0.45 μm) 過濾,
其液體保存於 4℃。
先將噬菌體原液加入 DNase I and RNase A 於 37℃ 下反應 兩小時,再添加 1/6 體積之 20% PEG 8000 和 2.5 M NaCl 後混 和均勻,置於冰上一小時,進行噬菌體顆粒凝集。之後將此溶液以
32
16,000 g 離心六十分鐘之後,將其 pellet 回溶於 SM buffer 中。
三、溶菌斑測試
1. 點測試驗 (spot test)
依照陳,2007 論文之方法,取 0.1 ml 隔夜培養之 Sm14 (OD600
約 0.4),加入至 5 ml soft ager (50-55℃)中,混和均勻後倒入 LB ager 上,作為 indicator host。待 ager 凝固後,在 ager 表層滴上 1 μl 之噬菌體液 (titer = 1 × 1011 PFU/ml),再放於 37℃ 培養箱培 養,並觀察有無溶菌斑 (plaque) 形成。
2. Phage titer assay
初 步 做 法 與 spot test 一 致 , 先 準 備 Sm14 之 菌 液 作 為 indicator host,不同的地方在於噬菌體加入的時機不同,噬菌體加 入時機如同 Sm14 之菌液加入 soft ager 中,此外 Smp 14 於分析 前也需適當的序列稀釋,每管 soft ager 加入的體積亦為相同 100 μl,於 37℃ 培養 16-18 小時後觀察,其表面將因 Smp14 會照成 的溶菌斑之生成,數其溶菌斑數量並回推原始噬菌體液中其 titer 濃度。
四、DNA 備製
1. 小量質體 DNA 製備
此方法是依照 Gene-SpinTM –V Miniprep purification Kit 之方 法。將 6 ml 隔夜培養的菌液以 8,000 g 離心 10 分鐘,去除上清 液。以 200 μl Solution I 重新懸浮菌液,加入 200 μl Solution II 溫 和混勻,再加入 200 μl Solution III 溫和混合均勻。以 12,400 g 離 心 10 分鐘,將 spin column 放入 Collection Tube,再吸取離心後 的上清液移置 spin column 中,以 6,000 g 離心 2 分鐘重複兩
33
次,離心後去除濾液,加入 700 μl Wash buffer,以 12,400 g 離心 2 分鐘,離心後倒掉濾液,在離心 3 分鐘,去除殘留於管壁的酒 精。將 spin column 移至新的微量離心管,靜置 5 分鐘,讓殘餘 的酒精揮發。加入 35-50μl 無菌去離子水,靜置 10 分鐘,以 12,400 g 離心 2 分鐘即可得到純化後的 DNA,保存於 -20℃ 冰 箱。
2. 大型質體 DNA 備製
方法依照 NucleoBond PC100 kit 之流程。從隔夜培養的菌液 轉接至 100 ml 新鮮 LB 培養基中,培養液加入適當的抗生素。於 37℃ 培養箱培養 30 分鐘,加入適當濃度的 inhibition solution 在 培養 3-4 小時 (OD600 0.5-0.6)。以 6,000 g 離心 10 分鐘去除上清 液,加入 8 ml solution S1 將菌落重新懸浮;加入 8 ml solution S2 溫和上下混合;加入 8 ml 冰的 solution S3,輕輕搖勻,冰上反應 5 分鐘。以 12,000 g 離心 10 分鐘,離心期間,用 5 ml 的 solution N2 潤洗 column。將離 心後的 上清液晶 過 濾紙過濾至 column 中,之後加入 6 ml solution N3 將多餘的離子試劑去除,最終加入 4 ml solution N5 將 DNA 回溶至液體中,加入 3.5 ml isopropanol 與液體混合並於 -20℃ 反應,之後於 4℃ 中,以 12,000 g 離心 30 分鐘沉澱 DNA,再用 70% 冰酒精潤洗後,將 DNA 晾乾後加 入無菌去離子水回溶,並保存於 -20℃ 備用。
3. 噬菌體 DNA 備製
依照陳,2007 論文之方法,先將純化過的噬菌體原液加入 DNase I and RNase A 於 37℃ 下反應兩小時,噬菌體加入 1/10 體
34
積的 chloroform 後劇烈搖晃,離心 15000 g 30 秒,將大多的菌體 雜質去除,只留上清液。之後加入 SDS、EDTA 和 proteinase K (最 終濃度分別為 0.1%、0.5 mM 和 50 μg/ml) 放於 65℃ 下反應 30 分鐘。再以等體積的 phenol/chloroform (100 μl /100 μl)分別萃取,
至界面無雜質為止,之後吸取上清液添加 1/10 體積的 3M sodium acetate 及 2 倍體積的 95% 冰酒精,於 -80℃ 中靜置反應 4 小 時,之後於 4℃ 中,以 12,000 g 離心 30 分鐘沉澱 DNA,再用 70% 冰酒精潤洗後,將 DNA 晾乾後加入無菌去離子水回溶,並 保存於 -20℃ 備用。
五、聚合酶連鎖反應 (polymerase chain reaction, PCR)
取 5-1000 ng DNA 最為模板,加入 0.1 - 1 mM 的引子、0.25 mM dNTP、終濃度 1 倍的 PCR 緩衝液和 0.4 倍 5 U/μl Taq 酵素,最後補無 菌去離子水至總體積。
利用儀器進行 PCR 反應。溫度設定 95℃ 進行一個循環,再以 95℃ 1 分鐘、引子黏合 45 秒 (溫度為 primer 之 Tm 值低 2℃) 和 72℃ 1 分鐘 進行 25 個循環。再以 72℃ 5 分鐘一個循環,最後維持在 4℃ 。取適量 PCR 產物進行電泳分析。
六、洋菜膠電泳分析 (Agarose gel electrophoresis)
將洋菜粉加熱溶於 0.5 倍的 TAE 電泳緩衝液中,其洋菜膠體濃度依需 要分析 DNA 片段大小而定,其濃度範圍約 0.7% - 2.5% (w/v)。洋菜粉完全 溶解後,待溫度降至 50 - 60℃ 時,倒入模板中再插上齒梳 (comb)。等待膠 體完全凝固,移除齒梳將其移至電泳槽中,加入 0.5X TAE 緩衝液。將 DNA 樣品與 6X loading dye 混合後加入膠體孔洞中,於同塊膠體加入 DNA
35
marker 作為對照組。以 110 V 進行電泳,於適當的時間將膠體浸漬於 ethidium bromide (EtBr) 之水溶液,再以去離子水浸泡退染。放置於 UV Box 觀察並且照相,藉由 DNA marker 比較,推斷其 DNA 大小及濃度。
七、限制酶切割作用
利用限制酶判斷特定序列的特性,選擇切割 DNA 樣品所需的限制酶酵 素和適當的限制酶緩衝液。將 DNA 樣品、限制酶酵素和十分之一體積的緩 衝液混合均勻,加入無菌的去離子水補至總體積。於 37℃ 進行反應 12 - 14 小時。
八、DNA 之回收
實驗流程依照 Gene-SpinTM – V2 1-4-3 DNA Extraction Kit 之步驟。將需 回收的 DNA 片段於 UV box 上,以刀片切割下來放入微量離心管中,加 入適當的 FAGP buffer 於 55℃ 反應 10 分鐘至膠體溶解,期間每 2 分鐘 溫和混合液體,放於室溫冷卻 2 分鐘。將 FAGP column 放入 Collection Tube,再將含有 DNA 溶液移置 FAGP column,以 6,000 g 離心 2 分鐘重 複 2 次。去除濾液,加入 700 μl Wash buffer,以 12,400 g 離心 2 分鐘,
離心後倒掉濾液,在離心 3 分鐘,去除殘留於管壁的酒精。將 FAGP column 移至新的微量離心管,靜置 5 分鐘,讓殘餘的酒精揮發。加入 35 - 50 μl 無 菌去離子水,靜置 10 分鐘,以 12,400 g 離心 2 分鐘即可得到純化後的 DNA,保存於 -20℃ 冰箱。
九、DNA 黏接反應 (DNA ligation)
將選定的質體 (vecter) 經由限制酵素切割回收後與回收 DNA 片段 (insert) 以適當的比例混合均勻。加入 1/10 總體積的酵素緩衝液和 0.3 倍 400 U/μl T4 DNA ligase,再加入無菌去離子水補至總體積 (20 μl),置於 16
℃ 反應 14 - 16 小時。