Background and Purpose
Diabetes mellitus is defined by the presence of chronic hyperglycemia and disturbances of carbohydrate, fat, and protein metabolism related to absolute or relative deficiencies in insulin secretion and/or insulin action (Bennett, 1994). It is associated with an enormous amount of morbidity and mortality resulting from diabetic microangiopathy, macroangiopathy, and neuropathic complications (Ruderman and Haudenschild, 1984). Particularly, type 1 diabetes mellitus (insulin-dependent diabetes mellitus) is characterized by the deficits in insulin secretion that leads to many metabolic consequences, such as severe hyperglycemia and its related symptoms (Bennett, 1994). Patients with type 1 diabetes mellitus have a three to six fold increased risk of cardiovascular death compared with the
non-diabetic subjects (Krolewski et al., 1987; Head and Fuller, 1990). The mechanisms underlying the development of microvascular and macrovascular angiopathy in type 1 diabetes mellitus are complex and not fully understood.
Endothelial dysfunction and decreased availability of endothelium-derived nitric oxide have been suggested to be the early events in diabetic vascular disorders (Hink et al., 2001; Hsueh and Quinones, 2003; Poston and Taylor, 1995; Schalkwijk and Stehouwer, 2005). Prolonged hyperglycemia could possibly lead to permanent endothelial dysfunction and thereby development of vascular complications in the diabetic status (Hink et al., 2001; Schalkwijk and Stehouwer, 2005). Many
investigators have indicated that endothelium-dependent vasodilating responses to acetylcholine are impaired in type 1 diabetic animals and in patients with type 1 diabetes mellitus (Johnstone et al., 1993; Kamata et al., 1989; Oyama et al., 1986;
Poston and Taylor, 1995; Targher et al., 2005). In addition, endothelium-derived nitric oxide plays a principle factor in the pathogenesis of diabetic vascular complications (Calver et al., 1992; Chan et al., 2000; Poston and Taylor, 1995). Nitric oxide, a potent vasodilator, is released by endothelium and plays a pivotal role in maintaining
vascular homeostasis (Palmer et al., 1987; Stamler et al., 1994). It not only relaxes vascular smooth muscles, but also inhibits LDL oxidation, platelet aggregation, monocyte adhesion, and smooth muscle proliferation/migration (Boger et al, 1996;
Kubes et al., 1991; Radomski et al., 1990). Decreased release or activity of nitric oxide could contribute to abnormalities of vasomotor function, as well as to
atherogenesis (Boger et al, 1996; Vanhoutte, 1997). Evidences have demonstrated that impaired endothelium-dependent vasodilation in patients with type 1 diabetes mellitus is probably as a result of the decreased bioavailability of endothelial nitric oxide (Chan et al., 2000; Cosentino and Lüscher, 1998; Poston and Taylor, 1995). Especially, the hyperglycemia-induced endothelial dysfunction may result from the decreased
production of nitric oxide, inactivation of nitric oxide by oxygen-derived free radical, and increased production of endothelium-derived contracting factors, which oppose the protective activity of nitric oxide (Chan et al., 2000; Poston and Taylor, 1995).
Endothelial damage and nitric oxide abnormalities may mainly involve in the
development of cardiovascular diseases in the type 1 diabetic status. Furthermore, one previous study has indicated that an increase in endothelium-dependent vasodilation early, which is followed by a reverse phase in which vasodilation is normal, and subsequently followed by a final phase of impaired vasodilation during the time course of type 1 diabetes mellitus (Pieper, 1999). It implies that the disease duration might be a critical factor in the development of diabetes-related cardiovascular disorders. However, limited information exists in evaluating the fluctuations of cardiovascular changes during the time course of type 1 diabetes mellitus.
It has been known that insulin, as well as insulin-like growth factor-1 (IGF-1), plays important roles in the modulation of glucose metabolism and the development of diabetic cardiovascular diseases (Goke and Fehmann, 1996; Ren et al., 1999;
Sowers, 1997; Spallarossa et al., 1996). Many reports have indicated that
hyperglycemia and insulin resistance deteriorate the alteration of vascular function leading to cardiovascular disease (Chan et al., 2000; Hsueh and Quinones, 2003;
Poston and Taylor, 1995). In type 1 diabetic patients, chronic hyperglycemia causes insulin resistance by inducing a defect in glucose extraction and insulin-stimulated blood flow (Baron et al., 1991; Yki-Järvinen and Utriainen, 1998). Otherwise, many cardiovascular risk factors, such as insulin resistance and diabetes mellitus, have been associated with the deficiency of serum IGF-1 level, which may subsequently result in endothelial dysfunction and cardiovascular disease (Conti et al., 2004). Clinically, both insulin and IGF-1 have potential effects on the treatment of diabetes mellitus and cardiovascular disorders (Bondy et al., 1994; Bretzel, 2004; DeWitt and Hirsch, 2003;
Dunger and Acerini, 1997; Duron, 1995; Kolaczynski and Caro, 1994). They can facilitate glucose metabolism, reduce hyperglycemia, lower insulin levels, increase insulin sensitivity, and improve lipid profiles (Bretzel, 2004; DeWitt and Hirsch, 2003;
Duron, 1995; Kolaczynski and Caro, 1994). Furthermore, IGF-1 has been believed to promote cardiac growth, improve cardiac contractility, cardiac output, stroke volume and ejection fraction (Bondy et al., 1994; Dunger and Acerini, 1997; Kajstura et al, 2001). Insulin and IGF-1 are structurally related hormones and share many signaling components (Hsueh and Law, 1999; Ren et al., 1999; Sowers, 1997). Many studies have indicated that insulin, as well as IGF-1, has vasodilatory effects in vivo that depend on the production of endothelium-derived nitric oxide (Isenovic et al, 2001;
Sowers, 1997; Steinberg et al., 1994). Insulin stimulates the production of nitric oxide
serine/threonine kinase Akt pathways, following by increasing endothelial nitric oxide synthase (eNOS) activity (Kuboki et al., 2000; Zeng et al., 2000). Animal studies have indicated that insulin treatment can ameliorate endothelial dysfunction by inducing overexpression of eNOS and production of nitric oxide in diabetic rats (Kobayashi and Kamata, 2001; Kobayashi et al., 2006). IGF-1 is a homologue of insulin and has insulin-like cellular responses. In vitro studies have shown that IGF-1 induces eNOS activity in endothelial cells through PI3-K/Akt-related pathways as well (Isenovic et al, 2001; Michell et al, 1999; Walsh et al., 1996). Therefore, IGF-1 also mediates the release of nitric oxide and subsequently vascular relaxation (Isenovic et al, 2001;
Michell et al, 1999; Sowers, 1997). Taken together, both insulin and IGF-1 can regulate vascular function through the stimulation of nitric oxide production. In our laboratory, we found that in the obese rat model, the decreased insulin-mediated vasorelaxation appeared to be counteracted by the increased IGF-1-mediated
vasorelaxation. Also, the PI3K-NOS-dependent pathway was mainly involved in the altered vasorelaxant responses. (Yang et al., 2007). However, in type 1 diabetic status, whether endothelial function, vasodilating responses, and signaling pathways
mediated by insulin and IGF-1 are deranged has not been fully investigated.
It is well known that exercise intervention or increased physical activity has beneficial effects for patients with diabetes mellitus and cardiovascular disease (Chandrashekhar and Anand, 1991; Chipkin et al., 2001; Fuchsjäger-Mayrl et al., 2002; Green et al., 2004; Hambrecht et al., 2000; Ornish et al., 1998; Shephard and Balady, 1999). Many studies have reported that life style changes, such as regular physical activity, may influence endothelial function and in turn reduce the occurrence of cardiovascular disorders (Chipkin et al., 2001; Green et al., 2004; Laughlin and McAllister, 1992). Exercise training induces many adaptive responses, including the enhanced insulin action on the glucose transport system of skeletal muscles, the reduced hormonal stimulation of hepatic glucose production, the improved blood flow to skeletal muscles, and the normalization of abnormal blood lipid profiles (Chipkin et al., 2001; Henriksen, 2002; Green et al., 2004). Furthermore, previous studies have indicated that chronic exercise significantly improves endothelial function and endothelium-dependent vasorelaxation to acetylcholine in different vascular beds in type 1 diabetic animals and patients with type 1 diabetes mellitus (Chakraphan et al., 2005; Fuchsjäger-Mayrl et al., 2002; Heidarianpour et al., 2007). There is substantial evidence that exercise training corrects the endothelial function by enhancing
acetylcholine (ACh)-induced endothelium-dependent vasorelaxation in vessels of normal, atherosclerotic, and diabetic animals (Chakraphan et al., 2005; Chen and Li, 1993; Delp et al., 1993; Heidarianpour et al., 2007; Yang et al., 2003). In addition, an increase in the agonist-stimulated NO release and the induction of NOS gene
expression have been shown to contribute to the beneficial exercise effects (Delp et al., 1993; Heidarianpour et al., 2007; Higashi, 1999; Sessa et al., 1994; Yang et al., 2002).
Recently, our laboratory has found that in normal animals, exercise training enhances both insulin-induced and IGF-1-induced vasodilating responses in the aortas (Yang et al., 2006). The increases of PI3-K and NO release are involved in the training effects on these vasoactive responses (Yang et al., 2006). However, little information exists about effects of exercise intervention on insulin-mediated and IGF-1-mediated vascular function and their underlying mechanisms in the disease status, such as type 1 diabetes mellitus.
The overall objective of this proposed project is to characterize effects of exercise intervention and insulin treatment on insulin-mediated and IGF-1-mediated vascular function and their underlying mechanisms in different time periods of type 1 diabetic rat model. This project will be carried out in three years. In the second year, the effects of exercise training on insulin-mediated and IGF-1-mediated vascular function and their underlying mechanisms will be characterized in type 1 diabetic rat model. This project would provide important information for elucidating the action of insulin and IGF-1 on the vascular function in type 1 diabetes mellitus with the
influence from exercise intervention. Better understanding of the link between type 1 diabetes mellitus and cardiovascular function could help clinicians to build a
comprehensive strategy aiming at reducing diabetes-related medical problems. This study might provide useful findings and basic mechanisms for lifestyle modification (i.e. exercise intervention) in patients with type 1 diabetes mellitus clinically.
Methods
Experimental animals
This study was approved by Institutional Animal Care and Use Committee of National Cheng Kung University. All procedures followed the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (NIH). Male Wistar rats (8-week old), purchased from Laboratory Animal Center of National Cheng Kung University, were randomly assigned to non-diabetic control group (Con), diabetic (DM), and diabetes with exercise training (DM+Ex) groups. Food and water were given ad libitum. All animals were housed in a temperature-controlled room (22-25℃) with a 12 h dark-light cycle at Laboratory Animal Center of National Cheng Kung University. No statistical difference in body weight and blood glucose level was found in the beginning of the experiment.
Diabetes induction
To induce type 1 diabetes mellitus, rats were fasted for three days before diabetic induction. On the day of induction, rats were generally anaesthetized by an
intraperitoneal imjection of sodium pentobarbital in the concentration of 45 mg/kg of body weight. After anaesthetization, femoral artery pulse was palpated to identify the position of femoral vein (femoral artery and vein are parallels nearby). When the femoral vein was exposed, streptozotocin (65 mg/kg of body weight) was injected immediately into femoral vein. The incision then sewed up and disinfected. To confirm the induction of type 1 diabetes, fasting blood glucose level (fasting
overnight) was determined one week after STZ injection by glucose test of tail vein blood using glucometer (Abbott Laboratories, Inc., USA). STZ-induced rats failed to exhibit an elevated blood glucose level greater than 200 mg/dL were excluded from the study.
Measurement of blood glucose level
Blood glucose level and body weight were monitored every week. All animals were fasted for 12 hours, and then blood from tail vein was collected as the samples to determine fasting blood glucose level by the glucometer.
Exercise training protocol
The 8-week exercise training protocol was implemented according to the study of Chen et al. Before the beginning of exercise training, rats from the DM+Ex group had run on a horizontal treadmill (Model T510E, Diagnostic and Research Instruments Co., Taiwan) at the speed of 12 m/min for the familiarization. After one week of familiarization, all rats had run on the treadmill 60 minutes per session, 5 sessions per week, for 8 weeks totally. During the training period, the running speed was gradually increased 3 m/min every two weeks, from 15 m/min to 24 m/min finally. In contrast, rats from sedentary groups were placed on the treadmill without running for 15 minutes each session. To avoid acute effect of exercise, all animals were sacrificed 48 hours after exercise training. Body weight and fasting blood glucose level were measured before sacrificing.
Assay of citrate synthase activity
Citrate synthase is one of the main oxidative enzymes in mitochondria. Previous
studies have proven that citrate synthase activity increased significantly after chronic exercise. Therefore an increase in citrate synthase activity is commonly used as a surrogate index for the effectiveness of exercise training. The assay of citrate synthase activity in this study was measured by using the method described by Srere. Soleus muscles (weight ~0.1g) isolated from three groups were immediately stored at -80℃
until analyzed. When measuring, the defrosted muscle samples were homogenized by homogenizer (Model 398, Biospec Products Inc, Mexico) with quintuple volumes of Tris buffer (0.1 mol/L) containing Triton X-100 (0.1%). After homogenizing, the samples were centrifuged by centrifuge at the speed of 13000 rpm for 15 minutes, and then the supernatants were collected. The 50 µl of supernatant was added into cuvette which had already contained 100 µl of DTNB (1 mM), 30 µl of acetyl-CoA (10 mM), and 770 µl of H2O. The absorption at 412 nm of wave length was measured by spectrophotometer (Beckman Instruments, Inc., DU640B, CA, USA) for 10 minutes to measure the possible acetyl-CoA deacylase activity. With the addition of 50 µl of oxaloacetate (10 mM), the measurement of enzymatic activity was begun also by spectrophotometeric assay for 5 minutes. Every sample was tested twice and averaged.
The result was expressed as micromoles of substrate utilized per minute per gram of wet tissue.
Preparation of vessels
At the end of experiments, all rats were anesthetized with diethyl ether and killed.
Thoracic aortas were then carefully excised from open-chest animals and placed immediately in oxygenated Krebs-Ringer solution at room temperature. Each aorta was cleaned of loosely adhering fat and connective tissues, then cut into 3-mm long of aortic strips for the measurement of vasorelaxation. One of these aortic strips from each sample was denuded from endothelium layer carefully by a stick. By removing the endothelium layer, vasorelaxation mediated by insulin and IGF-1 via
endothelium-dependent pathway could be confirmed. To ensure the endothelium layer was actually removed, ACh (10-6 M)-mediated vasorelaxation on the denuded aortic strip was tested prior to the formal vasorelaxation measurements. Since
ACh-mediated vasorelaxation is endothelium-dependent, the aortic strip was considered to be truly denuded when the result of ACh (10-6 M)-mediated vasorelaxation is less than 20% of precontraction. All processes had to be done carefully to ensure the vessels were not damaged during aortic strip preparation.
Measurement of vasorelaxation
Each aortic strip was mounted by a pair of stainless steel hooks on the
force-displacement transducer (Models FT3E, Grass, West Warwick, RI, USA) and submerged in organ chamber containing Krebs-Ringer solution bubbling with 95%
O2-5% CO2 at the constant temperature of 37℃. The mounted aortic strips were gradually stretched to the optimal passive tension, at which the contractile force evoked by phenylephrine was maximal. Previous studies have shown that the optimal passive tension of rat thoracic aorta is 2.0 g. After the optimal passive tension was achieved, all mounted aortic strips were equilibrated under the tension for 30 minutes.
Before the formal relaxation measurement, ACh (10-6 M)-mediated vasorelaxation was tested to ensure the vascular activity. For the formal relaxation measurements, the aortic strips were precontracted with the equieffective concentration of phenylephrine (10-7 M). When precontraction reached a plateau level, insulin (3x10-8 - 3x10-6 M), IGF-1 (10-9 - 10-7 M), ACh (3x10-9 - 3x10-7 M), and SNP (3x10-11 - 3x10-9 M) were added respectively in a cumulative manner. The output of the force-displacement transducer was registered by a polygraph recorder (TA11, GOULD, USA).
The roles of PI3-K and eNOS in the vasorelaxation
In order to characterize possible roles of PI3-K and eNOS responsible for insulin- and IGF-1-induced vasorelaxation, two selective inhibitors, wortmannin (3×
10-7 M) and L-NAME (NG-nitro-L-arginine methyl ester, 10-5 M), were pre-incubated with the vessel ring for 15 min, respectively. After pretreatment of the inhibitor, insulin- and IGF-1-induced vasorelaxation was determined as the percentage of the phenylephrine-evoked tension. After each experiment, the organ chamber was immediately washed with Krebs-Ringer solution at least three times. Before starting the next experiment, the vessel ring was incubated for more than 45 min for
equilibration. All results of these relaxation responses were expressed as percentages of phenylephrine (10-7 M)-evoked precontractile force (Kobayashi and Kamata, 2002;
Yang et al., 2003).
Statistical Analysis
Data were expressed as mean ± SEM; “n” indicated the sample sizes in each group. Response curves in time-dependent and concentration-dependent fashion among three groups were compared by ANOVA with repeated measures design and LSD post hoc test further. The difference of vasorelaxation responses between pre- and post-treatment of the inhibitors were determined by paired t test. P < 0.05 was considered as a significant level.
Results
Body weight and blood glucose level
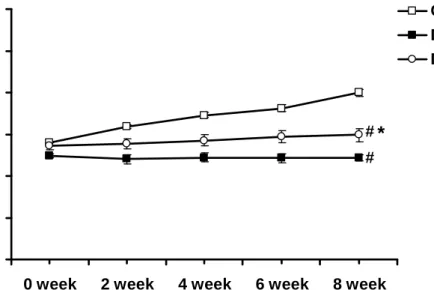
As indicated in Figure 1, body weight in the Con group was gradually increased with age. As compared with the Con group, body weight in the DM group was significantly lowered (P<0.05). Furthermore, the 8-week exercise training
significantly improved the lowered body weight in the DM+Ex group (P<0.05). Blood glucose level in the Con group was constantly in the normal range. After STZ
induction, blood glucose level in the DM group was significantly elevated as compared with the Con group (P<0.05). Eight-week exercise training significantly decreased the elevated blood glucose levels in the DM+Ex group (P<0.05), as shown in Figure 2.
Citrate synthase activity
Exercise training for 8 weeks significantly increased the citrate synthase activity of soleus muscle in the DM+Ex group as compared with the Con and DM groups
(P<0.05), as shown in Table 1. These results indicated that our exercise program was effective.
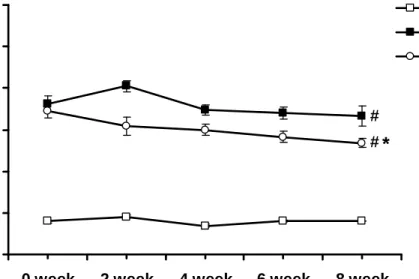
Concentration-dependent responses to insulin
When the phenylephrine (10-7 mol/L)-induced contraction had reached a plateau level, insulin (3x10-8 - 3x10-6 M) was added cumulatively. In aortic rings from the Con, DM, and DM+Ex rats, insulin (3x10-8 - 3x10-6 M) caused a
concentration-dependent vasorelaxation (Fig. 3A). In the DM group, insulin-mediated vasorelaxation was significantly weaker than that in the Con group (P<0.05). In the DM+Ex group, the exercise training significantly restored insulin-mediated
vasorelaxation (P<0.05) to the normal range (Fig. 3A). After the endothelium was removed, there was no significant difference among three groups (Fig. 3B). These results indicated that the effects of exercise intervention on the alternations of insulin-mediated vasorelaxation among three groups were regulated via the endothelium-dependent pathway.
Concentration-dependent responses to IGF-1
Similarly, IGF-1 (10-9 - 10-7 M) was added cumulatively when the aortic ring was pre-contracted with phenylephrine (10-7 mol/L). The administration of IGF-1 (10-9 - 10-7 M) induced a concentration-dependent vasorelaxation in the Con, DM, and DM+Ex rats (Fig. 4A). IGF-1-mediated vasorelaxation was also impaired in the DM group as compared with that in the Con group (P<0.05); the vasorelaxation in the DM+Ex group could be significantly improved after 8-week exercise training (P<0.05), even greater than that in the Con group (Fig. 4A). After the endothelium was removed, no significantly difference was found among three groups (Fig. 4B), which indicated the effects of exercise intervention on the alternations of
IGF-1-medated vasorelaxation among three groups were regulated though the endothelium layer as well.
Roles of PI3-K and eNOS in the vasorelaxant responses mediated by insulin and IGF-1
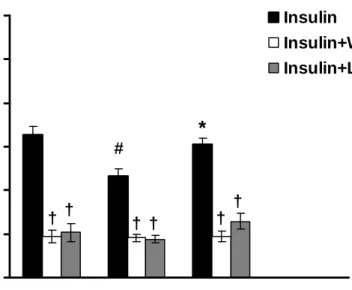
In the insulin-mediated vasorelaxation, wortmannin and L-NAME (the inhibitors of PI3-K and eNOS respectively) significantly inhibited the vascular responses among three groups (Fig. 5). Without the pre-treatments of wortmannin or L-NAME, the decreased insulin-mediated vasorelaxation was found in the DM group as compared with that in the Con group (P<0.05), whereas 8-week exercise training restored the decreased vasorelxation in the DM+Ex group (P<0.05). After the administration of wortmannin or L-NAME, there was no significant difference among three groups in the insulin-mediated vasolaxation.
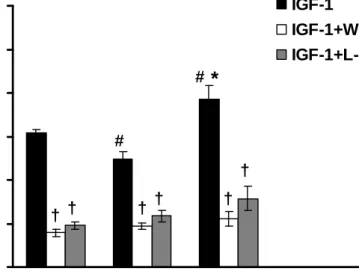
Similarly, wortmannin and L-NAME significantly inhibited IGF-1-mediated vasorelxation among three groups (Fig. 6). Without the pre-treatments of wortmannin and L-NAME, IGF-1-mediated vasorelaxation in the DM group was found to be impaired significantly as compared to that in the Con group (P<0.05), whereas the exercise training could reverse this impairment in the DM+Ex group (P<0.05). In contrast, after the administrations of wortmannin and L-NAME, there was no significant difference among three groups in IGF-1-mediated vasorelaxation. These results revealed that the effects of exercise training on insulin- and IGF-1-mediated vascular functions in the DM+Ex group were mainly due to the alternation of PI3-K and eNOS activities.
ACh-induced vasorelaxation
The administration of ACh (3x10-9 - 3x10-7 M) induced a concentration-dependent vasorelaxation in the Con, DM, and DM+Ex rats (Fig. 7). ACh-mediated
vasorelaxation was weaker in the DM group than that in the Con group (P<0.05), whereas exercise training restored the vasorelaxation (P<0.05) to normal range in the DM+Ex group. These results of our study were consistent with previous studies.
SNP-induced vasorelaxation
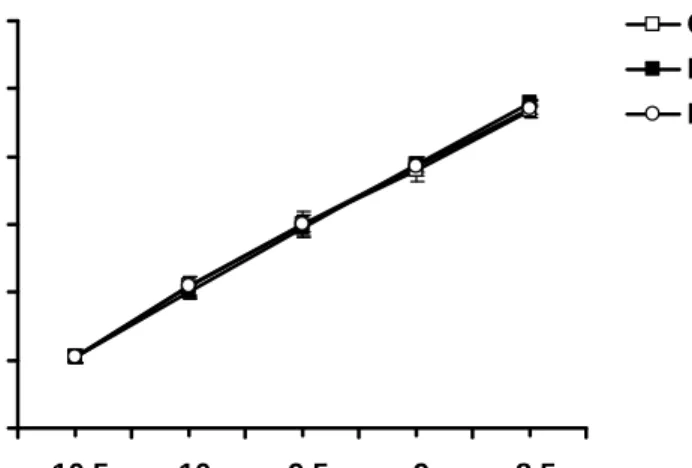
SNP primarily releases NO and causes vasorelaxation intracellularly within vascular smooth muscle cells without the involvement of endothelium. To examine the endothelium-independent vasorelaxation, the SNP-mediated vascular responses were examined and our results showed that there was no significant difference among three groups (Fig. 8). These results further suggested that exercise training exerted their positive effects on the vascular function in type 1 diabetes without affecting vascular smooth muscle layer.
Discussion
Our findings indicated that, 1) insulin- and IGF-1-mediated vasorelaxation was significantly impaired in the DM group compared with that in the Con group (P<0.05);
2) 8-week exercise training significantly ameliorated these impaired vascular responses to insulin and IGF-1 in the DM+Ex group (P<0.05); 3) the alternations of vascular responses to insulin and IGF-1 were mainly due to the altered activities of PI3-K and eNOS; 4) consistent with previous studies, ACh-mediated vasorelaxation was impaired in the DM group, whereas exercise training reversed the impaired vascular response in the DM+Ex group; 5) no significant difference was found in SNP-mediated vasorelaxation among three groups; 6) in the denuded vessels, the insulin- and IGF-1-mediated vasorelaxation was comparable among three groups. We suggested that insulin- and IGF-1-mediated vasorelaxant responses were significantly impaired in type 1 diabetes; whereas exercise intervention effectively reversed these vascular impairments. The benefits of exercise intervention were mainly due to the increased activities of PI3-K and eNOS in the NO-dependent pathway.
This project would provide important information for elucidating the action of insulin and IGF-1 on the vascular function in type 1 diabetes mellitus with the influence from exercise intervention. Better understanding of the link between type 1 diabetes mellitus and cardiovascular function could help clinicians to build a
comprehensive strategy aiming at reducing diabetes-related medical problems. This study might provide useful findings and basic mechanisms for lifestyle modification (i.e. exercise intervention) in patients with type 1 diabetes mellitus clinically.
References
Baron, A. D., Laakso, M., Brechtel, G., & Edelman, S. V. (1991). Mechanism of insulin resistance in insulin-dependent diabetes mellitus: a major role for reduced skeletal muscle blood flow. J Clin Endocrinol Metab, 73(3), 637-643.
Bennett, P. H. (1994). Definition, diagnosis, and classification of diabetes mellitus and impaired glucose tolerance. In C. R. Kahn & G. Weir (Eds.), Diabetes Mellitus.
(pp.193-200). USA.
Boger, R. H., Bode-Boger, S. M., & Frolich, J. C. (1996). The L-arginine-nitric oxide pathway: role in atherosclerosis and therapeutic implications. Atherosclerosis, 127, 1-11.
Bondy, C. A., Underwood, L. E., Clemmons, D.R., Guler, H.P., Bach, M. A., &
Skarulis, M. (1994). Clinical uses of insulin-like growth factor I. Ann Intern med, 120, 593- 601.
Calver, A., Collier, J., & Vallance, P. (1992). Inhibition and stimulation of nitric oxide synthesis in the human forearm arterial bed of patients with insulin-dependent diabetes. J Clin Invest, 90(6), 2548-2554.
Chakraphan, D., Sridulyakul, P., Thipakorn, B., Bunnag, S., Huxley, V. H., &
Patumraj, S. (2005). Attenuation of endothelial dysfunction by exercise training in STZ-induced diabetic rats. Clin Hemorheol Microcirc, 32(3), 217-226.
Chandrashekhar, Y., & Anand, I. S. (1991). Exercise as a coronary protective factor.
Am Heart J, 122, 1723-1739.
Chan, N. N., Vallance, P., & Colhoun, H. M. (2000). Nitric oxide and vascular responses in Type I diabetes. Diabetologia, 43(2), 137-147.
Chen, H. I., & Li, H. T. (1993). Physical conditioning can modulate
endothelium-dependent vasorelaxation in rabbits. Arterioscler Thromb, 13, 852-856.
Chipkin, S. R., Klugh, S. A., & Chasan-Taber, L. (2001). Exercise and diabetes.
Cardiol Clin, 19(3), 489-505
Conti, E., Carrozza, C., Capoluongo, E., Volpe, M., Crea, F., Zuppi, C., et al. (2004).
Insulin-like growth factor-1 as a vascular protective factor. Circulation, 110(15), 2260-2265.
Cosentino, F., & Lüscher, T. F. (1998). Endothelial dysfunction in diabetes mellitus. J Cardiovasc Pharmacol, 32(3), S54-61.
Delp, M. D., McAllister, R. M., & Laughlin, M. H. (1993). Exercise training alters endothelium-dependent vasoreactivity of rat abdominal aorta. J Appl Physiol, 75, 1354-1363.
Dunger, D. B., & Acerini, C. L. (1997). Does recombinant human insulin-like growth factor-1 have a role in the treatment of diabetes? Diabet Med, 14, 723-731.
Fuchsjäger-Mayrl, G., Pleiner, J., Wiesinger, G. F., Sieder, A. E., Quittan, M., Nuhr, M.
J., et al. (2002). Exercise training improves vascular endothelial function in patients with type 1 diabetes. Diabetes Care, 25(10), 1795-1801.
Goke, B., & Fehmann, H. C. (1996). Insulin and insulin-like growth factor-I: their role as risk factors in the development of diabetic cardiovascular disease Diabetes Res Clin Pract, 30, 93-106.
Green, D. J., Maiorana, A., O'Driscoll, G., & Taylor, R. (2004). Effect of exercise training on endothelium-derived nitric oxide function in humans. J Physiol, 561, 1-25.
Hambrecht, R., Wolf, A., Gielen, S., Linke, A., Hofer, J., Erbs, S., et al. (2000). Effect of exercise on coronary endothelial function in patients with coronary artery disease. N Engl J Med, 342, 454-460.
Head, J., & Fuller, J. H. (1990). International variations in mortality among diabetic patients: the WHO Multinational Study of Vascular Disease in Diabetics.
Diabetologia, 33(8), 477-481.
Heidarianpour, A., Hajizadeh, S., Khoshbaten, A., Niaki, A. G., Bigdili, M. R., &
Pourkhalili, K. (2007). Effects of chronic exercise on endothelial dysfunction and insulin signaling of cutaneous microvascular in streptozotocin-induced diabetic rats. Eur J Cardiovasc Prev Rehabil, 14(6), 746-752.
Henriksen, E. J. (2002). Invited review: Effects of acute exercise and exercise training on insulin resistance. J Appl Physiol, 93, 788-796.
Higashi, Y., Sasaki, S., Kurisu, S., Yoshimizu, A., Sasaki, N., Matsuura, H., et al.
(1999). Regular aerobic exercise augments endothelium-dependent vascular relaxation in normotensive as well as hypertensive subjects: role of
endothelium-derived nitric oxide. Circulation, 100, 1194-1202.
Hink, U., Li, H., Mollnau, H., Oelze, M., Matheis, E., Hartmann, M., et al. (2001).
Mechanisms underlying endothelial dysfunction in diabetes mellitus. Circ Res, 88(2), E14-22.
Hsueh, W. A., & Law, R. E. (1999). Insulin signaling in the arterial wall. Am J Cardiol, 84, 21J-24J.
Hsueh, W. A., & Quinones, M. J. (2003). Role of endothelial dysfunction in insulin resistance. Am J Cardiol, 92(4A):10J-17J.
Isenovic, E., Muniyappa, R., Milivojevic, N., Rao, Y., & Sowers, J. R. (2001). Role of PI3-kinase in isoproterenol and IGF-1 induced ecNOS activity. Biochem Biophys Res Commun, 285, 954-958.
A. (1993). Impaired endothelium-dependent vasodilation in patients with insulin-dependent diabetes mellitus. Circulation, 88(6), 2510-2516.
Kajstura, J., Fiordaliso, F., Andreoli, A. M., Li, B., Chimenti, S., Medow, M. S., et al.
(2001). IGF-1 overexpression inhibits the development of diabetic
cardiomyopathy and angiotensin II-Mediated oxidation stress. Diabetes, 50, 1414-1424.
Kamata, K., Miyata, N., & Kasuya, Y. (1989). Impairment of endothelium-dependent relaxation and changes in levels of cyclic GMP in aorta from
streptozotocin-induced diabetic rats. Br J Pharmacol, 97(2), 614-618.
Kobayashi, T., & Kamata, K. (2001). Effect of chronic insulin treatment on NO production and endothelium-dependent relaxation in aortae from established STZ-induced diabetic rats. Atherosclerosis, 155(2), 313-320.
Kobayashi, T., Oishi, K., Hayashi, Y., Matsumoto, T., & Kamata, K. (2006). Changes in aortic endothelial gene expressions and relaxation responses following chronic short-term insulin treatment in diabetic rats. Atherosclerosis, 185(1), 47-57.
Kolaczynski, J. W., & Caro, J. F. (1994). Insulin-like growth factor-1 therapy in diabetes: physiologic basis, clinical benefits, and risks. Ann Intern Med, 120, 47-55.
Krolewski, A. S., Kosinski, E. J., Warram, J. H., Leland, O. S., Busick, E. J., Asmal, A.
C., et al. (1987). Magnitude and determinants of coronary artery disease in juvenile-onset, insulin-dependent diabetes mellitus. Am J Cardiol, 59(8), 750-755.
Kubes, P., Suzuki, M., & Granger, D. N. (1991). Nitric oxide: an endogenous modulator of leukocyte adhesion. Proc Natl Acad Sci USA, 88, 4651-4655.
Kuboki, K., Jiang, Z. Y., Takahara, N., Ha, S. W., Igarashi, M., Yamauchi, T., et al.
(2000). Regulation of endothelial constitutive nitric oxide synthase gene
expression in endothelial cells and in vivo: a specific vascular action of insulin.
Circulation, 101, 676-681.
Laughlin, M. H., & McAllister, R. M. (1992). Exercise training-induced coronary vascular adaptation. J Appl Physiol, 73, 2009-2025.
Michell, B. J., Griffiths, J. E., Mitchelhill, K. I., Rodriguez-Crespo, I., Tiganis, T., Bozinovski, S., et al. (1999). The Akt kinase signals directly to endothelial nitric oxide synthase. Curr Biol, 9, 845-848.
Ornish, D., Scherwitz, L. W., Billings, J. H., Brown, S. E., Gould, K. L., Merritt, T. A., et al. (1998). Intensive lifestyle changes for reversal of coronary heart disease.
JAMA, 280, 2001-2007.
Oyama, Y., Kawasaki, H., Hattori, Y., & Kanno, M. (1986). Attenuation of
endothelium-dependent relaxation in aorta from diabetic rats. Eur J Pharmacol, 132(1), 75-78.
Palmer, R. M., Ferrige, A. G., & Moncada, S. (1987). Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature, 327, 524-526.
Pieper, G. M. (1999). Enhanced, unaltered and impaired nitric oxide-mediated endothelium-dependent relaxation in experimental diabetes mellitus:
importance of disease duration. Diabetologia, 42(2), 204-213.
Poston, L., & Taylor, P. D. (1995). Endothelium-mediated vascular function in insulin-dependent diabetes mellitus. Clin Sci, 88(3), 245-255.
Radomski, M. W., Palmer, R. M., & Moncada, S. (1990). An L-arginine/nitric oxide pathway present in human platelets regulates aggregation. Proc Natl Acad Sci USA, 87, 5193-5197.
Ren, J., Samson, W. K., & Sowers, J. R. (1999). Insulin-like growth factor I as a cardiac hormone: physiological and pathophysiological implications in heart disease. J Mol Cell Cardiol, 31, 2049-2061.
Ruderman, N. B. & Haudenschild, C. (1984). Diabetes as an atherogenic factor. Prog Cardiovasc Dis, 26, 373-412.
Schalkwijk, C. G., & Stehouwer, C. D. (2005). Vascular complications in diabetes mellitus: the role of endothelial dysfunction. Clin Sci, 109(2), 143-159.
Sessa, W. C., Pritchard, K., Seyedi, N., Wang, J., & Hintze, T. H. (1994). Chronic exercise in dogs increases coronary vascular nitric oxide production and endothelial cell nitric oxide synthase gene expression. Circ Res, 74, 349-353.
Shephard, R. J., & Balady, G. J. (1999). Exercise as cardiovascular therapy.
Circulation, 99, 963-972.
Sowers, J. R. (1997). Insulin and insulin-like growth factor in normal and pathological cardiovascular physiology. Hypertension, 29, 691-699.
Spallarossa, P., Brunelli, C., Minuto, F., Caruso, D., Battistini, M., Caponnetto, S., et al. (1996). Insulin-like growth factor-I and angiographically documented coronary artery disease. Am J Cardiol, 77, 200-202.
Srere, P. A. (1969). Citrate synthase. Methods Enzymol, 13, 3-5.
Stamler, J. S., Loh, E., Roddy, M. A., Currie, K. E., & Creager, M. A. (1994). Nitric oxide regulates basal systemic and pulmonary vascular resistance in healthy humans. Circulation, 89, 2035-2040.
Steinberg, H. O., Brechtel, G., Johnson, A., Fineberg, N., & Baron, A. D. (1994).
Insulin-mediated skeletal muscle vasodilation is nitric oxide dependent. A novel action of insulin to increase nitric oxide release. J Clin Invest, 94,
Targher, G., Bertolini, L., Zoppini, G., Zenari, L., & Falezza, G. (2005). Increased plasma markers of inflammation and endothelial dysfunction and their association with microvascular complications in Type 1 diabetic patients without clinically manifest macroangiopathy. Diabet Med, 22(8), 999-1004.
Vanhoutte, P. M. (1997). Endothelial dysfunction and atherosclerosis. Eur Heart J, 18, 19-29.
Walsh, M. F., Barazi, M., Pete, G., Muniyappa, R., Dunbar, J. C., & Sowers, J. R.
(1996). Insulin-like growth factor I diminishes in vivo and in vitro vascular contractility: role of vascular nitric oxide. Endocrinology, 137, 1798-1803.
Yang, A. L., Chao, J. I., & Lee, S. D. (2007). Altered insulin-mediated and insulin-like growth factor-1-mediated vasorelaxation in aortas of obese Zucker rats. Int J Obes, 31, 72-77.
Yang, A. L., Jen, C. J., & Chen, H. I. (2003). Effects of high cholesterol diet and parallel exercise training on the vascular function of rabbit aortas: a time course study. J Appl Physiol, 95, 1194-1200.
Yang, A. L., Su, C. T., Lin, K. L., Chao, J. I., & Lee, S. D. (2006). Exercise training improves insulin-induced and insulin-like growth factor-1-induced
vasorelaxation in rat aortas. Life Sci, 79, 2017-2021.
Yang, A. L., Tsai, S. J., Jiang, M. J., Jen, C. J., & Chen, H. I. (2002). Chronic exercise increases both inducible and endothelial nitric oxide synthase gene expression in endothelial cells of rat aorta. J Biomed Sci, 9, 149-155.
Yki-Järvinen, H., & Utriainen, T. (1998). Insulin-induced vasodilatation: physiology or pharmacology? Diabetologia, 41(4), 369-379.
Zeng, G., Nystrom, F. H., Ravichandran, L. V., Cong, L. N., Kirby, M., Mostowski, H., et al. (2000). Roles for insulin receptor, PI3-kinase, and Akt in
insulin-signaling pathways related to production of nitric oxide in human vascular endothelial cells. Circulation, 101, 1539-1545.
Self-Evaluation
This research was consistent with my previously proposed projects for NSC. The results indicated that, 1) insulin- and IGF-1-mediated vasorelaxation was significantly impaired in the DM group compared with that in the Con group (P<0.05); 2) 8-week exercise training significantly ameliorated these impaired vascular responses to insulin and IGF-1 in the DM+Ex group (P<0.05); 3) the alternations of vascular responses to insulin and IGF-1 were mainly due to the altered activities of PI3-K and eNOS; 4) consistent with previous studies, ACh-mediated vasorelaxation was
impaired in the DM group, whereas exercise training reversed the impaired vascular response in the DM+Ex group; 5) no significant difference was found in
SNP-mediated vasorelaxation among three groups; 6) in the denuded vessels, the insulin- and IGF-1-mediated vasorelaxation was comparable among three groups. We suggested that insulin- and IGF-1-mediated vasorelaxant responses were significantly impaired in type 1 diabetes; whereas exercise intervention effectively reversed these vascular impairments. The benefits of exercise intervention were mainly due to the increased activities of PI3-K and eNOS in the NO-dependent pathway.
This project would provide important information for elucidating the action of insulin and IGF-1 on the vascular function in type 1 diabetes mellitus with the influence from exercise intervention. Better understanding of the link between type 1 diabetes mellitus and cardiovascular function could help clinicians to build a
comprehensive strategy aiming at reducing diabetes-related medical problems. This study might provide useful findings and basic mechanisms for lifestyle modification (i.e. exercise intervention) in patients with type 1 diabetes mellitus clinically.
The plan in my NSC proposal was executed, and parts of our results have been presented in the international scientific conference and further prepared for the publication for sharing our findings in the relative field.
Table 1. Comparison of citrate synthase activity among three groups
Group Citrate Synthase Activity
(µmol/min/g wet wt)
Con 1.91±0.01
DM 1.88±0.07
DM+Ex 2.15±0.08
Data are presented as mean±SEM. Abbreviations: Con- non-diabetic control; DM- diabetic; DM+Ex- diabetes with exercise training.
# P<0.05 vs. Con; * P<0.05 vs. DM. (n=5~6)
* #
0 100 200 300 400 500 600
0 week 2 week 4 week 6 week 8 week
Body weight (g)
Con DM DM+Ex
#
#
*
Figure 1. Comparison of time-dependent curves of body weight among the Con, DM, and DM+Ex groups. The abscissa represents the disease duration. 0 week indicates one week after STZ induction. Abbreviations: Con- non-diabetic control; DM- diabetic; DM+Ex- diabetes with exercise training.
# P<0.05 vs. Con; * P<0.05 vs. DM. (n=10)
0 100 200 300 400 500 600
0 week 2 week 4 week 6 week 8 week
Blood glucose level (mg/dL) Con
DM DM+Ex
#
#
*
Figure 2. Comparison of time-dependent curves of blood glucose levels among the Con, DM, and DM+Ex groups. The abscissa represents the disease duration. 0 week indicates one week after STZ induction. Abbreviations are the same as those in Figure 1. # P<0.05 vs. Con; * P<0.05 vs. DM. (n=10)
(A) Endothelium-intact vasorelaxation
0 10 20 30 40 50 60
-7.5 -7 -6.5 -6 -5.5
[Insulin] (Log M) Vasorelaxation (% precontraction)
Con DM DM+Ex
#
*
(B) Endothelium-denuded vasorelaxation
0 10 20 30 40 50 60
-7.5 -7 -6.5 -6 -5.5
[Insulin] (Log M) Vasorelaxation (% precontraction)
Con DM DM+Ex
Figure 3. (A) Insulin-induced vasorelaxation with intact endothelium and (B) Insulin-induced vasorelaxation with denuded endothelium in aortic rings obtained from the Con, DM, and DM+Ex groups. The ordinate shows the vasorelaxation of aortic strips as a percentage of the precontraction induced by phenylephrine (10-7 M).
Abbreviations are the same as those in Figure 1.
# P<0.05 vs. Con; * P<0.05 vs. DM. (n=10)
(A) Endothelium-intact vasorelaxation
0 10 20 30 40 50 60
-9 -8.5 -8 -7.5 -7
[IGF-1] (Log M) Vasorelaxation (% precontraction)
Con DM DM+Ex
#
#*
(B) Endothelium-denuded vasorelaxation
0 10 20 30 40 50 60
-9 -8.5 -8 -7.5 -7
[IGF-1] (Log M) Vasorelaxation (% precontraction)
Con DM DM+Ex
Figure 4. (A) IGF-1-induced vasorelaxation with intact endothelium and (B) IGF-1-induced vasorelaxation with denuded endothelium in aortic rings obtained from the Con, DM, and DM+Ex groups. The ordinate shows the vasorelaxation of aortic strips as a percentage of the precontraction induced by phenylephrine (10-7 M). Abbreviations are the same as those in Figure 1.
# P<0.05 vs. Con; * P<0.05 vs. DM. (n=10)
0 10 20 30 40 50 60
Con DM DM+Ex
Vasorelaxation (% precontraction) Insulin
Insulin+Wortmannin Insulin+L-NAME
†
† †
† †
†
#
*
Figure 5. Effects of wortmannin (3x10-7 M) and L-NAME (10-5 M) on the insulin (3x10-7 M)-mediated vasorelaxation of aortic strips obtained from the Con, DM, and DM+Ex groups.
The ordinate shows the vasorelaxation of aortic strips as a percentage of the precontraction induced by phenylephrine (10-7 M). Abbreviations are the same as those in Figure 1.
# P<0.05 vs. Con; * P<0.05 vs. DM; † P<0.05 without the inhibitor vs. with the inhibitor (wortmannin or L-NAME) (n=8)
0 10 20 30 40 50 60
Con DM DM+Ex
Vasorelaxation (% precontraction) IGF-1
IGF-1+Wortmannin IGF-1+L-NAME
†
†
† †
† †
# *
#
Figure 6. Effects of wortmannin (3x10-7 M) and L-NAME (10-5 M) on the IGF-1 (10-8 M) -mediated vasorelaxation of aortic strips obtained from the Con, DM, and DM+Ex groups.
The ordinate shows the vasorelaxation of aortic strips as a percentage of the precontraction induced by phenylephrine (10-7 M). Abbreviations are the same as those in Figure 1.
# P<0.05 vs. Con; * P<0.05 vs. DM; † P<0.05 without the inhibitor vs. with the inhibitor (wortmannin or L-NAME) (n=8)
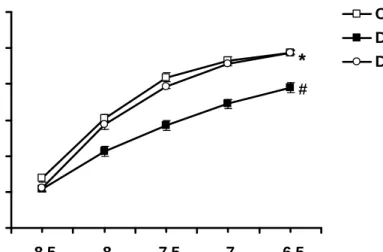
0 20 40 60 80 100 120
-8.5 -8 -7.5 -7 -6.5
[ACh] (Log M) Vasorelaxation (% precontraction)
Con DM DM+Ex
#
*
Figure 7. Concentration-dependent curves of the ACh-mediated vasorelaxation of aortic strips obtained from the Con, DM, and DM+Ex groups. The ordinate shows the vasorelaxation of aortic strips as a percentage of the precontraction induced by phenylephrine (10-7 M).
Abbreviations are the same as those in Figure 1.
# P<0.05 vs. Con; * P<0.05 vs. DM. (n=10)
0 20 40 60 80 100 120
-10.5 -10 -9.5 -9 -8.5 [SNP] (Log M)
Vasorelaxation (% precontraction)
Con DM DM+Ex
Figure 8. Concentration-dependent curves of the SNP-mediated vasorelaxation of aortic strips obtained from the Con, DM, and DM+Ex groups. The ordinate shows the vasorelaxation of aortic strips as a percentage of the precontraction induced by phenylephrine (10-7 M).
Abbreviations are the same as those in Figure 1. (n=10)