國立臺灣大學生物資源暨農學院 植物病理與微生物學系
碩士論文
Department of Plant Pathology and Microbiology College of Bioresources and Agriculture
National Taiwan University Master Thesis
天然素材 Tryptanthrin 對隱球菌的抗菌效果及細胞週期之影響 Tryptanthrin, a natural product, exhibits novel antifungal activity
and cell cycle arrest against Cryptococcus species
張雅琳 Ya-Lin Chang
指導教授:陳穎練 博士 Advisor: Ying-Lien Chen, Ph.D.
中華民國 106 年 10 月
October 2017
誌謝
似乎經歷了一個比其他人更長的碩士生涯,長到回到校園後每個 人都疑惑你怎麼還在這。回想起來還是完成不少事,研討會、學術發 表、論文口試、海外實習,因為有許多人的幫助才得以圓滿達成這些。
當初念碩士班最自己最大的期許就是培養獨立思考與解決問題的能力,
目標太抽象,至今不確定自己在經過碩士班訓練後是否有符合預期,
畢竟研究從來不是獨立可以完成的;而問題也鮮少真的被完全解決,
學術的道路太漫長,往往只留下更多需要被探究的未知。學到更多的 是如何與人溝通,在每次討論後,能夠更務實的找出有價值的資訊,
從中找到最適切的研究方向。
謝謝指導教授陳穎練老師,在這三年多的日子提供這麼多的協助,
給我許多機會磨練,成就了一些,也搞砸了不少。謝謝口試委員,曾 祥洸醫師、楊玉良老師、薛雁冰老師,對於我的研究成果給予的指導 與肯定。謝謝 novel 305,你們給予的心靈上的支持與生活中的調劑永 遠是讓人繼續前行的動力。謝謝家人,雖然總是難以與你們分享研究 上的苦難與喜悅,一無所知仍無條件支持是讓我覺得自己一點也沒資 格放棄的理由。
在臺大的日子終要告一段落,希望走過的日子不會白費,下一站,
還要更努力。
中文摘要
本研究致力於尋找對人類病原真菌念珠菌及隱球菌具抗真菌效果之活性物質,
希望能找出具抑制能力或和現有藥物有協同作用的化合物。透過快速大量篩 選小分子化合物,包括天然素材與美國食品藥品管理局核准之藥品,找出有 抗真菌能力的化合物。從篩選結果中發現,來自蜜蜂腸胃道共生之放線菌所
分泌的次級代謝物色胺酮 tryptanthrin 對隱球菌屬有抑制能力,對新型隱球菌
的最小抑制濃度為 2 g/mL,對格特隱球菌為 4 g/mL。進一步實驗結果發 現,tryptanthrin 與鈣調蛋白免疫抑制劑(FK506)有協同作用。Tryptanthrin 的 抗隱球菌能力為劑量依賴性的,並且為抑菌型藥物而非殺菌型。我們也發現 tryptanthrin 在 37 ºC 時抑菌能力比 30 ºC 或 25 ºC 更佳。同時,tryptanthrin 對 臨床抗藥性隱球菌菌株 T1 和 89-610 也具抑菌作用。為了找到 tryptanthrin 可 能參與的藥物標靶或途徑,我們用藥物敏感性的測試篩選突變株庫。相較於 野生株,49 個突變株對藥物更敏感或有抗性,其中有一部分突變株參與在細 胞週期的調控。螢光激發流式細胞分選和基因表現定量結果顯示隱球菌的細
胞週期調控可能和 tryptanthrin 呈現的抑制作用相關。綜合以上研究,天然素
材tryptanthrin 具有抗真菌能力並和 FK506 有協同作用,有潛力作為隱球菌症 的治療策略,尤其是抗藥性菌株。
關鍵字: 色胺酮、隱球菌、天然素材、病原真菌、抗真菌藥劑
ABSTRACT
This study aims to identify bioactive compounds that exert novel antifungal activity alone or exhibit synergistic effect with an existing antifungal agent against human fungal pathogens Cryptococcus and Candida. We screened several compound libraries including natural products, agricultural fungicides and FDA-approved drugs in order to identify compounds that exert antifungal activity. Among selected compounds, tryptanthrin secreted from Nocardiopsis alba, an actinobacterium existed in the intestine of honeybee, was chosen for further characterization because of its potent inhibition activity against Cryptococcus species with the minimal inhibition concentration (MIC) of 2 g/mL for C. neoformans and 4 g/mL for C.
gattii. We further found that tryptanthrin showed synergistic effect with FK506, an
immunosuppressant, based on broth dilution and checkerboard assays. Tryptanthrin inhibits the growth of Cryptococcus cells in a dose-dependent manner and shows fungistatic activity instead of fungicidal. We also found that tryptanthrin is more effective at 37 ºC compared with to 30 ºC or 25 ºC. Meanwhile, tryptanthrin demonstrated antifungal activity against two clinical azole-resistant C. neoformans isolates, T1 and 89-610. In order to identify potential targets or pathways that tryptanthrin involves in, we screened the Cryptococcus deletion mutant library by drug susceptibility test. Forty-nine deletion mutants were found to be more susceptible or resistant than that of the wild-type, and some of these mutants were involved in in cell cycle regulation. Fluorescence-activated cell sorting and gene
expression quantification results support that cell cycle regulation in C. neoformans may be linked to tryptanthrin. In summary, the natural product tryptanthrin shows novel antifungal activity alone or in combination with FK506, leading possible therapeutic strategies for cryptococcosis caused by Cryptococcus species, especially the azole-resistant isolates.
Keywords: Tryptanthrin, Cryptococcus, Natural product, Fungal pathogen, Antifungal agent
Table of Contents
口試委員審定書...i
誌謝...ii
中文摘要...iii
Abstract...iv
1. Introduction 1.1 Antifungal drug research and development...1
1.2 Cryptococcus neoformans...1
1.3 Currently used antifungal agents for Cryptococcus...2
1.4 Natural product tryptanthrin...4
2. Materials and methods 2.1 Strains, media and chemicals...6
2.2 Drug library screening...6
2.3 Disc diffusion assay...7
2.4 Determination of minimum inhibitory concentration (MIC) and minimum fungicidal concentration (MFC)...7
2.5 Fractional inhibitory concentration assay for antifungal activity...8
2.6 Growth kinetics assay...8
2.7 Serial dilution growth assay...8
2.8 Mutant library screening...9
2.9 Fluorescence-activated cell sorting assay...9
2.10 Quantitative real-time RT-PCR...10
2.11 Murine model and antifungal treatment...11
3. Results and discussion
3.1 Screening results of compound libraries…...11
3.2 Tryptanthrin exhibits novel antifungal activity against Cryptococcus
species...12
3.3 Growth of C. neoformans is sensitive to tryptanthrin at 37
ºCbut not at 25
ºC...13
3.4 Tryptanthrin exhibits synergistic activity with calcineurin inhibitors FK506 against Cryptococcus species...14
3.5 Fitness test of Cryptococcus deletion mutant library...15
3.6 Tryptanthrin exhibits G
1/S cell cycle arrest in C. neoformans...16
3.7 Cell cycle associated genes were up-regulated after treating with tryptanthrin...17
3.8 Limited anti-cryptococcal activity of tryptanthrin in murine infection model...18
4. Conclusion...19
5. Future work...19
6. Acknowledgement...21
7. References...22
8. Appendix...43
List of Figures
Figure 1. Screening results of the compounds libraries against Cryptococcus neoformans and Candida albicans...33 Figure 2. The structure and antifungal activity of tryptanthrin...34 Figure 3. Tryptanthrin exhibits dose-dependent inhibition against C.
neoformans...35 Figure 4. Growth of C. neoformans is sensitive to tryptanthrin at 37 ºC but
not at 30 or 25 ºC...36 Figure 5. Tryptanthrin can inhibit drug-resistant C. neoformans, and clinical
or environmental C. gattii isolates at 37 ºC...37 Figure 6. Tryptanthrin exhibits synergistic effect with FK506 against
Cryptococcus species...38 Figure 7. Tryptanthrin exhibits dose-dependent manner on resting of G
1/S
phase in C. neoformans (H99)...39 Figure 8. Cell cycle associated genes were up-regulated after treating with
tryptanthrin...40 Figure 9. Limited anti-cryptococcal activity of tryptanthrin in murine
infection model...41 Figure S1. Rapamycin and fluconazole cause cell cycle arrest in G
1/S phase
but exhibit different gene expression pattern...42
List of Tables
Table 1. Fungal strains used in this study...27 Table 2. Minimum inhibitory concentration of tryptanthrin against human
fungal pathogen...28 Table 3. Fractional inhibitory concentration index of tryptanthrin with
FK506, fluconazole (FLC) or amphotericin B (AmB)...29 Table 4. Deletion mutants of C. neoformans with altered susceptibility to
tryptanthrin...30
Table 5. Oligonucleotide primers used in this study...32
1. Introduction
1.1 Antifungal drug research and development
Approximately 1.2 billion individuals worldwide suffer from fungal infections, and the occurrence of these infections has significantly increased in recent years due to a rise in the number of immunocompromised patients, such as patients with AIDS or those with cancer, organ transplant, or autoimmune disease who require immunosuppressive therapy 1. Unlike superficial infections that cause local, benign, or self-limiting diseases, invasive fungal infections (IFIs) are deep-seated and include bloodstream and systemic infections as well as infection of specific organs. IFIs are frequently caused by yeast pathogens such as Candida and Cryptococcus, filamentous fungi such as Aspergillus, Fusarium, or Mucor, or, less frequently, dimorphic fungi, including Coccidioides, Blastomyces, or Histoplasma.
Currently, only three main classes of antifungals are approved for treatment of patients with IFIs: polyenes, triazoles, and echinocandins. These agents target ergosterol, lanosterol 14- α-demethylase, and β-1,3 glucan synthase, respectively 2. Because our current antifungal therapies have problems like great efficacy but with significant toxicities, emergence of drug resistance and low potency against specific fungal pathogen, new drugs are needed to extend the limited antifungal arsenal 3.
1.2 Cryptococcus neoformans
The pathogenic yeast Cryptococcus species belong to Basidiomycota and are divergent from other human fungal pathogens Candida and Aspergillus, which belong to ascomycetous fungi. Two major Cryptococcus species including C. neoformans and C.
gattii attribute to cryptococcosis in human. The virulence factors of the pathogen include the capability of survival in host temperature (37 ºC), the production of capsule and
melanin which are specific structures for escaping from immune system and restricted environment 4. Current studies also reported quorum sensing between fungal populations and unique changing in cell morphology as important virulence factors in host 5. Cryptococcus spp. are ubiquitous in the environment and usually abundant in avian excreta and various trees with waxier cuticles 6. Therefore, the epidemiology of cryptococcosis demonstrated the worldwide distribution of clinical cases. It is estimated that one million cases of cryptococcosis were diagnosed each year and over half of the patients were died from cryptococcal meningoencephalitis 7, 8. Meanwhile, high mortality in different countries is noticed. Two main reasons contributed to the high mortality possibly include drug shortage in low-resources countries and strong association with human immunodeficiency virus infection 9. Others people at risk are the immunocompromised patients who take immunosuppressive medications 9. Hence, the adequate treatment and new antifungal for prophylaxis and management of cryptococcosis is important.
1.3 Currently used antifungal agents for Cryptococcus
C. neoformans is an opportunistic pathogen that colonizes in different parts of human body and is life-threatening if no appropriate therapeutic treatments performed. The pathogen infects individuals by inhalation into lungs and thus cause pulmonary cryptococcosis.
Afterward, C. neoformans can spread quickly to other organs and disseminate to central nerve system and lead to overwhelming cryptococcal meningoencephalitis. Cryptococcosis is a challenging disease due to limited utilizable antifungal drugs. Currently, the practical guideline for management of cryptococcosis only articulate two regimens: fluconazole monotherapy and amphotericin B with flucytosine as combination therapy 10. Besides,
amphotericin B can cause severe nephrotoxicity, thus newer formulation liposomal amphotericin B is used when it comes to renal impairment patients. Fluconazole is a fungistatic agents widely used as an initial therapy. It is a safer and cheaper antifungal drugs compared with others. However, the cases of fluconazole resistance in patients with cryptococcal meningitis have more frequently been reported in Africa 11. Due to the risk of terrible side effects and emergence of drug resistant isolates 12, more novel antifungal agents are urgent to be discovered and applied to the clinic.
Multiple approaches have been launched to discover lead compounds for antifungal investigation. We could follow the target-based or phenotype-based methods to find out the potential drug targets or the candidate compounds. Nowadays, several antifungal agents are in pre-clinical stages. T-2307, an arylamidines derivatives, has been reported its broad- spectrum antifungal activity 13, 14. Structure of arylamidine is similar to pentamidine which is utilized in anti-parasitic and anti-pneumocystis. Though the modes of action are still unclear, the in vitro and in vivo results both demonstrated that T-2307 was more effective than the conventional echinocandins or polyenes against human fungal pathogens 14. Besides, a series of 1,2-benzisothiazol-3(2H)-one derivatives (BZT) were also reported to exhibit antifungal activity. The mode of action of BZT was proposed by interfering fungal mitochondrial respiration system 13, 15. In addition, a group of phosphoinositide dependent kinase inhibitors KP-372-1, OSU-03012 and UCN-01 have been discovered as potent antifungal compounds against Candida and Cryptococcus species 16. BHBM and D0 have been demonstrated as new class of antifungal agents with the ability to target the fungal sphingolipids, GlcCer 17. It has been demonstrated that these compounds exhibit antifungal activity against wide ranges of human fungal pathogens and are highly effective in in vivo
test.
Repurposing of off-patent drugs is another strategy for antifungal discovery. Zhai et al.
screened the Johns Hopkins Clinical Compound Library and found an early stage antibiotics, polymyxin B, for Gram-negative bacterial infection and later for, human fungal infections 18, 19. Furthermore, polymyxin B is synergistic with fluconazole and exhibits fungicidal effect at lower concentration. Other repurposing cases such as tamoxifen and toremifene, acted as estrogen receptor antagonists in breast cancer therapy, have been found to exhibit anti-cryptococcal activity 20. Miltefosine which is a medication for treating leishmaniasis was reported its in vitro activity against Cryptococcus species 21. The miltefosine analogs also exhibits broad-spectrum activity to human fungal pathogen 22. While the in vivo test of miltefosine and its analogs revealed unsuccessful efficacy. In spite of the fact that more and more antifungal agents have been discovered and identified, none of them represents as a new class of antifungal agent approved in pharmacy market yet.
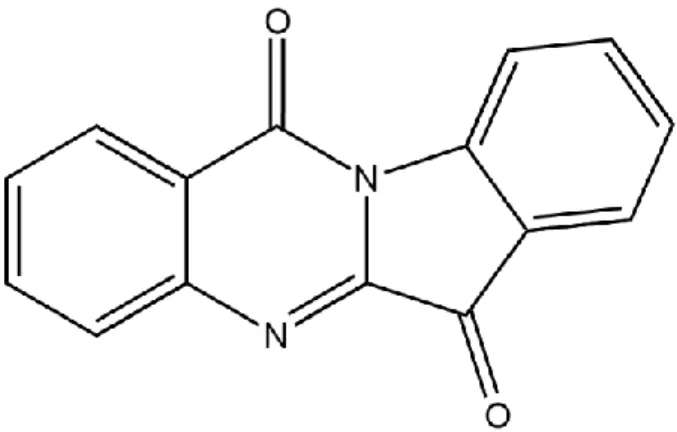
1.4 Natural product tryptanthrin
Natural alkaloid tryptanthrin is a yellow needle-like crystal which could be purified from several natural sources or be synthesized through organic procedure. Tryptanthrin sublimated from a plant Indigo is first identified in 1879 by Sommargua 23, 24. Afterward, it is denominated tryptanthrin due to the high production of this compound by culturing Candida lipolytica in tryptophan-contained media 25, 26. The empirical formula of tryptanthrin is C15H8N2O2 (M.W. = 248.24) and its chemical structure is planar.
Tryptanthrin could be isolated from many organisms, from microorganism Schizophyllum commune, Leucopaxillus cerealis to higher plants Isatis indigotica, Polygonum tinctorium
and Wrightia tinctoria. In this study, we identify tryptanthrin from the secondary metabolites of Nocardiopsis alba, an actinobacterium existed symbiotically in the intestine of honeybee. Tryptanthrin has been mentioned to its various medical activities, including anti-microbial, anti-trypanosomal, anti-inflammatory and anti-cancer properties.25, 27, 28
Tryptanthrin possesses anti-bacterial activity against both Gram positive and negative bacteria including Bacillus subtilis, Mycobacterium tuberculosis, Porphyromonas gingivalis and methicillin-resistant Staphylococcus aureus. It could inhibit the growth of different pathogenic protozoa such as Trypanosoma brucei, Leishmania donovani and Plasmodium falciparum. Otherwise, tryptanthrin inhibits cellular expression of inflammation-related enzyme COX-2 and 5-LOX. Tryptanthrin exhibits various anti- cancer activity via decrease the secretion of growth factor, inhibition of multidrug resistance gene, cytotoxicity to specific cancer cells. All of the above demonstrated that tryptanthrin is a multifunction bioactive compound potential for pharmacological use. But little of studies reported that tryptanthrin also exhibits anti-fungal activity 29, 30. Few studies described that tryptanthrin exhibits antifungal activity to topical infection fungus such as Trichophyton mentagrophytes and Malassezia furfur 31, 32. As to systemic infection fungus, none of the report has mentioned tryptanthrin exhibits antifungal activity.
Studies about the mechanism of tryptanthrin bioactivity are still finite and incomplete.
Bandekar et al. indicated that tryptanthrin inhibits the growth of Escherichia coli by DNA intercalation 33. On the other hand, few studies reported that tryptanthrin exhibits anti- cancer activity by arresting cell cycle at G0/G1 phase and inhibiting the proliferation on myeloid leukemia and neuroblastoma cells 34-36. Conversely, tryptanthrin exhibits anti- angiogenesis by arresting cell cycle at G2/M phase 37. However, the definite drug targets or
associated pathways that tryptanthrin interacts with remain elusive 38.
2. Materials and methods
2.1 Strains, media and chemicals
Fungal strains used in this study were listed in Table 1. YPD (1% yeast extract, 2% peptone and 2% glucose) and RPMI-1640 medium (Sigma) buffered to PH 7.0 with 0.2% glucose and 0.165 M MOPS were used in this study. Compound libraries (Selleckchem Chemicals) of natural products, FDA-approved drugs were used for antifungal activity screening.
Tryptanthrin used in this study was obtained from the laboratory of Dr. Yu-Liang Yang in Academia Sinica. FK506 (Astellas Pharma Inc.), fluconazole (FLC, Bedford Laboratories), amphotericin B (AmB, Sigma), rapamycin (Selleckchem Chemicals) were used for drug susceptibility test and cell cycle analysis.
2.2 Drug library screening
C. albicans and C. neoformans cells were grown overnight at 30 ºC then washed twice with sterile water. The concentration of cells suspension was adjusted to 0.0005 OD/mL in RPMI-1640 medium. 98 L of the strain culture was added to each well in a 96 well plate.
2 L of drug library compounds were added into the indicated wells to 10 M in final volume. The plates were incubated at 35 ºC for 24 or 48 hr. The results were read visually.
The potential candidates with obvious antifungal activity were chosen for further drug susceptible test.
2.3 Disc diffusion assay
Yeast cells were grown overnight at 30 ºC, and 0.1 OD 600 (in 100L) was spread on the surface of YPD medium. Paper discs (6 mm) were placed on the surface of the medium, and 12.5L of tryptanthrin (25 g) and 5L of DMSO (as a control) were added to each disc. The plates were incubated at 37 ºC for 48 hr and photographed.
2.4 Determination of minimum inhibitory concentration (MIC) and minimum fungicidal concentration (MFC)
We determined MIC by the Clinical and Laboratory Standards Institute (CLSI) protocol M27-A3. Testing was done in RPMI-1640, buffered to pH 7.0 with 0.165 M MOPS. Yeast strains were grown in YPD medium overnight, incubated at 30 ºC with shaking, and washed twice with ddH2O. The OD600 was measured and each strain was diluted to 1 OD600
/mL. This inoculum was diluted to 0.0005 OD 600 /mL in RPMI-1640. 98 L of the strain culture was added to each well in a 96 well plate format. 2 L of serially diluted drugs were added to the wells, yielding a final volume of 100 L per well. The concentration of tryptanthrin which ranged from 0.25–64 g/mL was added across the plate with the highest to the lowest from the left to the right well. The plates were incubated for 48 hr at 35 ºC, and OD 600 value of each plate was read by spectrophotometer. The in vitro drug studies were performed at least twice. The minimum inhibitory concentration (MIC100) of drug was defined as the clear well with the lowest concentration of each drug. The minimum fungicidal concentration were determined by transferring 3 L of solution from indicated well to drug-free YPD plates. The viability of fungus cells was checked after the plates were incubated for 48 hr at 30 ºC.
2.5 Fractional inhibitory concentration assay for antifungal activity
Fractional inhibitory concentration (FIC) was assessed via checkerboard titration assays.
The inoculum were prepared as MIC method. Tryptanthrin was added across the plate with the highest concentration in the left well and the lowest concentration in the right well. The complement antifungal agents FK506, fluconazole or amphotericin B with concentration ranged from 0.25-16 g/mL was added from top to bottom, with the highest concentration in the top row and the lowest concentration in the bottom row. This manipulation demonstrated 70 different combined drug concentration to be tested on one plate. The plates were incubated for 48 hr at 35 ºC and the MIC value was based on spectrophotometric determination at OD600 comparing to the control well. The FIC was calculated by: (MIC combined drug A/MIC alone drug A) + (MIC combined drug B/MIC alone drug B). A FIC index of < 0.5 indicates synergy, > 4.0 indicates antagonism, and an index between 0.5 and 4 indicates no interaction.
2.6 Growth kinetics assay
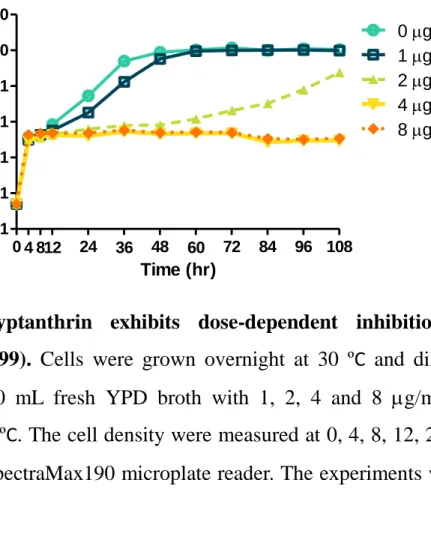
Cells were grown overnight at 30 ºC and washed twice with sterile water and then diluted to 0.0005 OD/mL with fresh YPD broth. 10 mL of mixtures were incubated at 37 ºC (200 rpm) with 1, 2, 4 and 8 g/mL tryptanthrin. The cell density were measured after 0, 4, 8, 12, 24 and every 12 hr later using SpectraMax190 microplate reader (Molecular Devices).
The experiments were performed in triplicate and the results were plotted using Prism 5.03.
2.7 Serial dilution growth assay
Cells were grown overnight at 30 ºC and washed twice with sterile water. The optical density of cells was measured and then the fungus cells were diluted to 1 OD600/mL in sterile water. 5-fold serial dilution of each strain was accomplished in 96-well microplate.
3 L of serial diluted suspension was spotted onto the YPD soli agar. The plates were incubated at 25, 30 and 37 ºC for 72 hr and photographed.
2.8 Mutant library screening
The C. neoformans mutant library was used in the fitness test. This deletion mutant library contains 1201 genes knockout strains and cover ~30% genes in C. neoformans 39. Mutants were recovered from frozen stock to liquid YPD medium and incubated at 30 ºC for 48 hr.
Cells were washed twice with sterile water. Mutants in each well were diluted to 0.05 OD/mL and transferred by pin replicator to one well YPD-agar plate without or with tryptanthrin (1, 2 or 4 g/mL). Plates were incubated at 35 ºC for 48 hr. Mutants that displayed growth defect in plate with 1 g/mL tryptanthrin were defined as hypersensitive (HS) and mutants which could be able to grow on YPD plate containing 4 g/mL tryptanthrin were considered as resistant (R).
2.9 Fluorescence-activated cell sorting assay
Yeast cells treated with 0, 1, 2, 4, 8 g/mL tryptanthrin for 12 hr were harvested and washed twice with PBS. Cells were fixed in 70% ethanol overnight at -20 ºC. Cells were centrifuged at 850 g for 5 min, washed twice with PBS and later re-suspended in 1 mL Tris/NaCl/MgCl2 buffer (200 mM Tris PH7.5, 211 mM NaCl, 78 mM MgCl2 ).RNase was added to final concentration of 100 g/mL and incubated at 37 ºC for 2 hr. Propidium
iodide was added to final concentration of 5 g/mL. The samples were analyzed by flow cytometry (Beckman Coulter FC500) and percentages of each phase were calculated.
2.10 Quantitative real-time RT-PCR
Strains were grown overnight at 30 ºC and washed twice with sterile water. The yeast cells were diluted to 0.02 OD/mL in 50 mL fresh YPD containing indicated compounds. Cells were harvested after 12 h incubation at 37 ºC (200 rpm), washed twice and lyophilized overnight. The total RNAs were extracted using TRIZOL (Thermo Fisher Scientific) and chloroform (Fisher Scientific). The RNA was precipitated in pre-cooled isopropanol, washed by 75% ethanol and finally eluted in RNase-free dH2O. Total RNA were evaluated the quality and quantity by RNA electrophoresis and Nanodrop spectrophotometer. The qualified RNA was treated with TURBO DNA-free Kit (Invitrogen) to remove the remaining genomic DNA. The DNase-treated RNA was reverse-transcribed to cDNA with the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Real-time PCR reactions of 20 l included 6 l cDNA (6 ng), 10 µl of 2x qPCR master mix (Fast SYBR Green Master Mix; Applied Biosystems), 2 l of 2.5 M forward primer and 2 l of 2.5
M reverse primer. Primers were designed using Primer 3 (Applied Biosystems) and listed in Table 5. Quantitative PCR conditions were shown below: 95°C /10 min for denaturation;
95°C /3 sec, 60°C /30 sec (40 cycles); 95°C /15 sec, 60°C /60 sec, 95°C /15 sec (melting curve). The StepOnePlus System and StepOne v2.2 (Applied Biosystems) were used to determine the cycle threshold (Ct) and the relative expression levels were calculated based on 2 -∆∆Ct. The bar graphs of ACT1 normalized relative quantity compared with drug- untreated samples were created with Prism 5.03. The P-values between samples were
determined by using unpaired t test.
2.11 Murine model and antifungal treatment
Four-week male outbred ICR mice (BioLasco Co. Ltd, Taiwan) were utilized in this study.
Mice were housed five mice per cage and maintained with sufficient food and water until weighing approximately 27 g. To establish disseminated cryptococcosis, mice were inoculated intravenously via the lateral tail vein with C. neoformans (H99). C. neoformans were grown in 10 mL YPD medium overnight at 30 ºC. The cells were washed twice with sterile water, counted with hemocytometer and adjusted the concentration to obtain an infection inocula concentration of 5 x 106 cells/mL. 200 L were inoculated in mice by lateral tail vein injection. The inocula were appropriate diluted and spread on YPD medium for confirmation of viability. Antifungal treatment with tryptanthrin was initiated at 4, 24, 48 and 96 hr post inoculation at doses 2, 8, 32 mg/kg of body weight by oral gavage or intraperitoneal injection. Tryptanthrin were dissolved in DMSO and prepared to indicated concentration with phosphate buffered saline (DMSO: PBS=1:9, DMSO should not above 10% of the final volume).
3. Results and discussion
3.1 Screening results of compound libraries
The high-throughput screening of nearly 1060 natural products and FDA-approved drugs provided several compounds with novel antifungal activity against C. neoformans (H99) and C. albicans (SC5314) (Fig. 1). Screening ~140 natural products, 12 compounds with antifungal activity against C. neoformans (H99) and C. albicans (SC5314) were identified
and four of them including sclareol, azomycin, parthenolide and tryptanthrin have not been reported. These four compounds could inhibit the growth of Cryptococcus neoformans but not Candida albicans. Meanwhile, 61 FDA-approved drugs demonstrated anti-Candida or anti-cryptococcal activity and five of them such as carmofur, ponatinib, eltrombopag, penfluridol and otilonium bromide were unpublished.
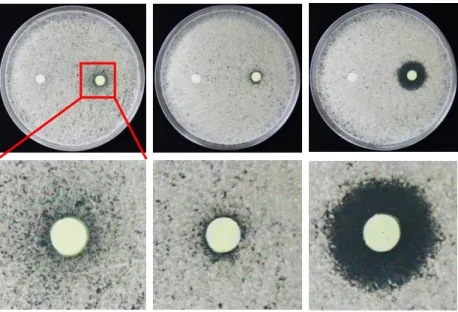
3.2 Tryptanthrin exhibits novel antifungal activity against Cryptococcus species Several compounds including tryptanthrin were selected for further investigation based on the results of the high-throughput screenings. We validated the antifungal activity of tryptanthrin against C. neoformans. The disc diffusion assay indicated that tryptanthrin has antifungal activity against C. neoformans and Saccharomyces cerevisiae, but not Candida albicans (Fig. 2). The obvious inhibition zone produced by tryptanthrin could be seen in C.
neoformans. However, tryptanthrin showed a slight inhibition against S. cerevisiae.
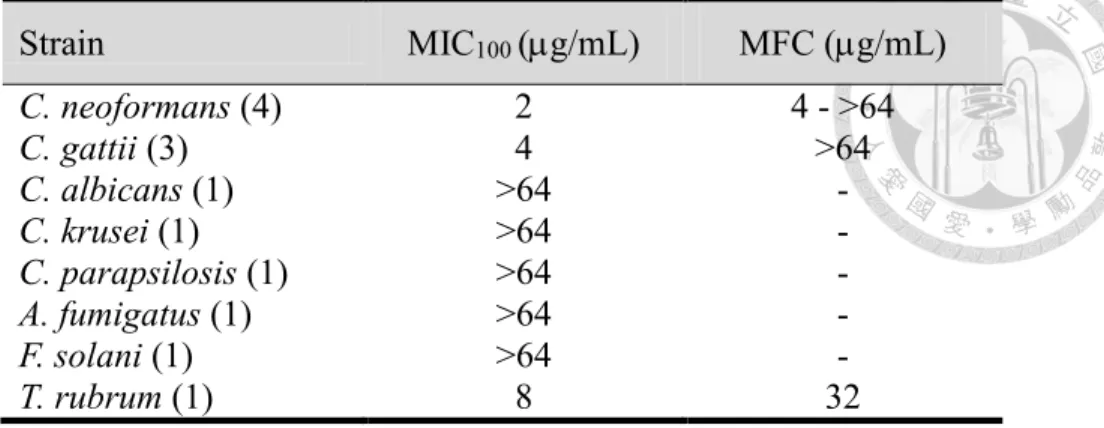
Therefore, we focused on the anti-cryptococcal activity of tryptanthrin. In C. neoformans, we first tested serotype A strain, which is the most widespread strain causing human fungal disease 40. Serotype D, a lesser virulent strain and causing fewer cases of cryptococcosis than serotype A, was also tested 41, 42. The antifungal susceptibility test via broth ilution assays showed that tryptanthrin exhibited equivalent growth inhibition for both A and D serotypes (Table 1), suggesting no serotype susceptibility differences in response to tryptanthrin and it is consistent to other antifungal drugs 43. In Cryptococcus gattii, we have tested three strains including two isolates (R265 and R272) from Vancouver Island outbreak and one isolate (WM276) from the environment (Table 1). The MIC values for C.
neoformans and C. gattii is 2 g/mL and 4 g/mL, respectively. The MFC values of C.
neoformans and C. gattii strains were all above 64 g/mL, indicating tryptanthrin may be a fungistatic agent. The growth kinetic assay of C. neoformans in different concentration demonstrated that tryptanthrin exhibits antifungal activity at 2 g/mL and in a dose- dependent manner (Fig. 3). Besides, C. neoformans continued to grow at 2 g/mL after incubating for 60 hr but still inhibited at 4 g/mL. These result may resulted from different nutrient conditions. Nevertheless, the growth curve indicated that tryptanthrin exhibits potent growth inhibition activity against C. neoformans but cannot kill the fungal cells.
Emergence of drug resistant isolates is a serious problem in clinical therapy and leads to treatment failure. Thus finding a potential antifungal agent that could conquer clinically drug-resistant isolates is important. Surprisingly, the MIC value for tryptanthrin against C.
neoformans T1 and 89-610, azole-resistant isolates, is 2 g/mL (Table 3), which is the same as the azole sensitive C. neoformans (H99), leaving a hint that tryptanthrin may target C.
neoformans via a pathway distinct from ergosterol biosynthesis pathway. Nevertheless, it needs more clinical isolates from different sources for antifungal susceptibility assay to validate the potency of tryptanthrin. Our results suggested that tryptanthrin revealed a narrow-spectrum antifungal activity due to ineffective inhibition activity against other human fungal pathogen such as Aspergillus fumigatus, Fusarium solani, Candida krusei, and Candida parapsilosis. Interestingly, tryptanthrin exhibits antifungal activity against Trichophyton rubrum, a filamentous fungus causing onychomycosis, as well as the previous study in 1979 against Trichophyton mentagrophytes (Table 3) 32.
3.3 Growth of C. neoformans is sensitive to tryptanthrin at 37 ºC but not at 25 ºC C. neoformans is capable to grow at the human body temperature of 37 ºC which is an
important virulence factors. To determine the impact of tryptanthrin on thermotolerance of C. neoformans, we tested the susceptibility of drug at different temperatures by disc diffusion and spotting assay. The results demonstrated that C. neoformans is more sensitive to tryptanthrin at higher temperature (Fig. 4). The agar plate incubated at 37 ºC exhibits a clear inhibition zone and the MIC value is 1 g/mL which reveals great antifungal activity.
Interestingly, the disc diffusion assay on solid agar demonstrated slightly inhibition of growth at 30 ºC and 25 ºC but tryptanthrin could still exhibit antifungal activity in liquid medium (Fig. 4). These results may be caused by the differences between liquid and solid- based test 44. The spotting assay demonstrated the similar results as shown above (Fig. 5).
In solid YPD agar plate containing tryptanthrin (2 g/mL), C. neoformans could hardly grow at 37 ºC but appeared a normal growth at 25 ºC. Otherwise, C. gattii demonstrated increased sensitivity to tryptanthrin at higher temperature as well. Multiple genes involve with stress responses were up-regulated at high temperature for altering the tough conditions 45. FK506 is an example of antifungal agent that targets calcineurin which requires for fungal growth at 37 ºC. It demonstrated toxicity to C. neoformans at 37 ºC but not at 24 ºC46. In addition to calcineurin, pathways such as Ras, HOG and MAPK are also play the role in temperature-sensing signaling 47. Therefore, we hypothesized that tryptanthrin may involve in specific pathways essential at high temperature.
3.4 Tryptanthrin exhibits synergistic activity with the calcineurin inhibitor FK506 against Cryptococcus species
Combination therapy is a strategy that aim to enhance the clinical outcome by blocking the drug resistance, declining dosage of drugs and improving drug efficacy 48. Therefore, we
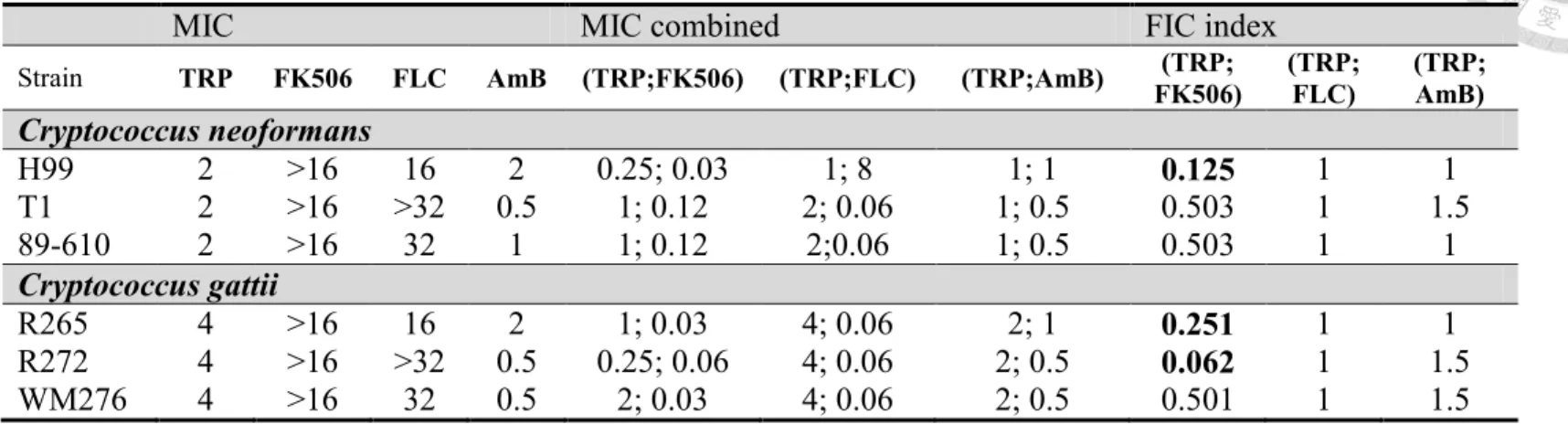
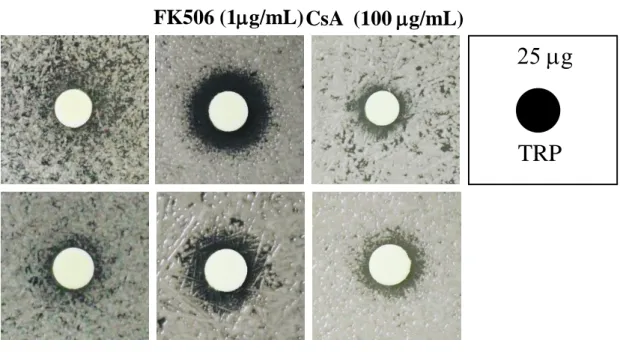
tested the interactive effects between tryptanthrin and current antifungal agents for cryptococcosis treatment. Our results suggested that there is no interaction between tryptanthrin and fluconazole or amphotericin B against the tested Cryptococcus species (Table 3). Surprisingly, the calcineurin inhibitor FK506, which can inhibit the growth of C.
neoformans at 37 ºC, exhibits synergistic activity with tryptanthrin against C. neoformans H99 and C. gattii R265 and R272 (FIC<0.5; Table 3) 49. FK506 has been reported of the synergistic effect with bafilomycin A1 or fluconazole against C. neoformans and with posaconazole against C. albicans 50, 51. We also tested another calcineurin inhibitor, cyclosporine A, by disc diffusion assay, but cyclosporine A has no synergy with tryptanthrin (Fig. 6A) 52. As the evidence of synergistic effect between tryptanthrin and FK506, we further investigated that whether this combination may transform fungistatic activity of tryptanthrin into fungicidal effect. However, the medium transferring from the checkerboard onto drug-free YPD solid agar has grown into colonies and this result indicated that the combination treatment cannot kill cryptococcal cells (Fig. 6B).
3.5 Fitness test of Cryptococcus deletion mutant library
The modes of action for each lead compound should be heavily focused on during the drug development and targets at some elsewhere in fungus cell would be a pioneering innovation for antifungal agents 53. We utilized the deletion collection mutants of C. neoformans to identify their susceptibility to tryptanthrin and aim to find the direct or indirect clues for drug target identification. In these fitness assay conditions, we defined mutants unable to grow on the YPD plate with 1 g/mL tryptanthrin as hypersensitive and able to grow on YPD plate with 4 g/mL as resistant strains. Our results demonstrated 49 mutants revealed
differential drug susceptibility; 27 of them were resistant and 22 were hypersensitive.
Microplate antifungal susceptibility assays were used to verify the fitness results from agar plates. The final results were classified by the function of the deleted mutants. A category of genes were identified and the may related directly or indirectly to the pathway interfering by tryptanthrin (Table 4). We noticed that a group of cell cycle-related genes such as RAD53, SSN801, MLN2, BUB1 (CNAG_03184), ACP2 (CNAG_03900), and KIP2 (CNAG_06335).
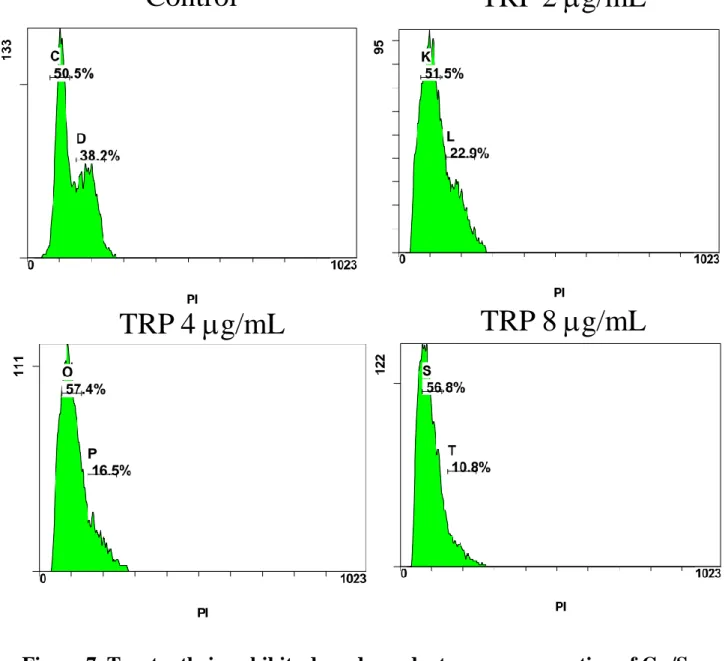
3.6 Tryptanthrin exhibits G1/S cell cycle arrest in C. neoformans
Based on the fitness test above, we suggest that the variation of susceptibility in mutants compared with wild-type may reflect cell cycle dysfunction in tryptanthrin-treated cells, thus the following experiments related cell cycle analysis were conducted. The DNA contents of each single cell were measured by fluorescence-activated cell sorting (FACS) assay and the distribution of cell cycle were plotted. Our results showed that tryptanthrin exhibits a dose dependent manner in arresting the G1/S phase of C. neoformans (H99). The percentage of G2 was declined when the concentration of tryptanthrin increased (i.e. 8
g/mL) in culture medium (Fig. 7). Meanwhile, rapamycin which induces G1 arrest in yeast were used as a positive control, and it demonstrated a pattern similar to tryptanthrin (Fig.7 and Fig. S1A) 54. Surprisingly, fluconazole also exhibits G1 arrest as well as tryptanthrin (Fig. S1A). These results suggested that arresting in cell cycle of C. neoformans may occur when treating with fungistatic agents. By treating with fungistatic agents, cells were impeded to enter the G2 phase and thus the budding process were inhibited 55.
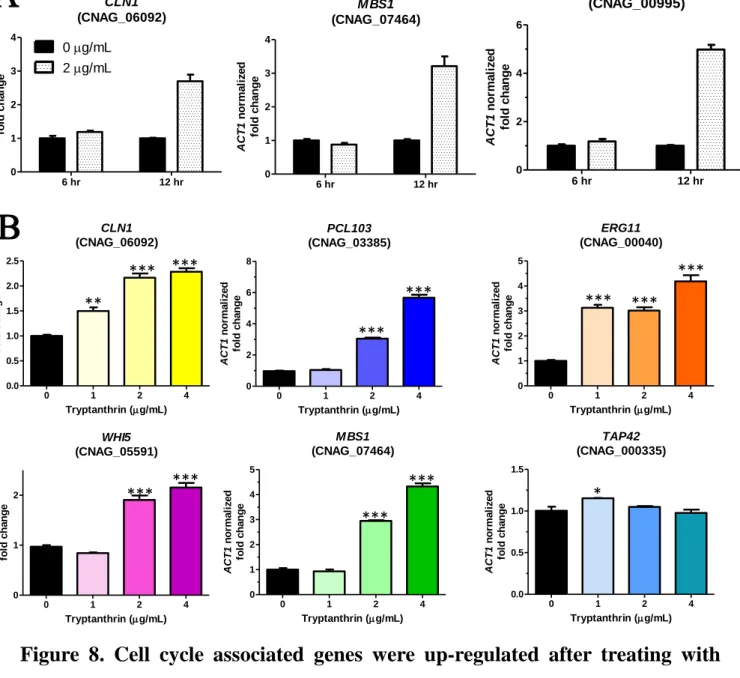
3.7 Cell cycle associated genes were up-regulated after treating with tryptanthrin Due to the fact that tryptanthrin blocks cell cycle of C. neoformans in G1/S phase, we further examine whether the regulation of cell cycle was interfered by tryptanthrin. We analyzed the mRNA expression of genes associate with the regulation of cell cycle, especially focused on the G1 and G1/S transition genes. Tryptanthrin exerted a dose dependent effect on the mRNA levels of CLN1, PCL103, WHI5, MBS1, CNAG_00995 and ERG11 (Fig. 8B). Cln1 and Mbs1 have been reported to play important roles on cell cycle progression in C. neoformans 56. Mbs1 is a transcription factor homologous to the yeast Mbp1 and Swi4 57. Proteins Mbp1 and Swi4 in S. cerevisiae form the MBF and SBF complex with Swi6 and involve in the G1/S transition by activating the G1 cyclins, Cln1 and Cln2 58. Our data suggested that tryptanthrin blocks the cell cycle in G1 phase and therefore causes the up-regulated the expression of G1/S transition genes. CNAG_00995 encoded a hypothetical protein that predicted to function in meiotic recombination was also up-regulated 54, 59. Since fluconazole and rapamycin revealed the cell cycle arrest effect in G1/S phase, as well as tryptanthrin, we wondered the mRNA expression of ERG11 and TAP42 which associated with ergosterol biosynthesis and TOR pathway, target of fluconazole and rapamycin, respectively. Interestingly, ERG11 was up-regulated after tryptanthrin treatment and is similar to the results of fluconazole 59. Conversely, expression of TAP42, encodes an essential protein implicated with TOR pathway, was not affected by tryptanthrin 60. Our data suggested that tryptanthrin and fluconazole may regulate cell cycle in a similar way than rapamycin even though these three fungistatic agents share the same pattern in cell cycle analysis.
3.8 Limited anti-cryptococcal activity of tryptanthrin in murine infection model To evaluate the application potential of tryptanthrin, we tested the efficacy of tryptanthrin against disseminated cryptococcosis. The intravenous injection of C. neoformans in murine model could simulate the situation of cryptococcosis. Tryptanthrin was treated 4 hr post- inoculation and once-daily dosing for three days. Due to the uncertainty of the best regimen for the delivery of tryptanthrin, we delivered tryptanthrin via oral gavage or intraperitoneal injection (IP). As the results of two delivery strategies, tryptanthrin did not improve the survival rate at dose of 2, 8 or 32 mg/kg as compared to PBS control group (Fig. 9). In the oral gavage administration group, every tryptanthrin-treated group died earlier than PBS group, indicating the potential toxicity of tryptanthrin to mice during cryptococcal infection.
Furthermore, we found that mortality rate of tryptanthrin at 32 mg/kg dose at the fastest pace and the P value is 0.0313 (Log-rank test) as compared with the PBS group. In the IP administration group, all of the drug treatment group demonstrated no anti-cryptococcal activity and had equivalent survival rate. Our results indicated that high dose of tryptanthrin administration via oral gavage may cause negative effect in murine infection model.
The failure of tryptanthrin therapeutic test in murine model may be due to three possible disadvantages of tryptanthrin as an antifungal agent in animal bodies. First, tryptanthrin was slightly precipitated and crystallized in PBS solvent, which may cause a difficulty in diffusion of tryptanthrin to the bloodstream. Second, C. neoformans could transmigrate into brain of murine model in 24 hr post-inoculation and it is the major cause of fatality61. Jähne et al. indicated that tryptanthrin has high blood-brain barrier penetration activity based on the in vitro human and animal models 62. Nevertheless, an in vivo pharmacokinetic study of tryptanthrin in rats reported that tryptanthrin remained in liver the most and the
longest time, but did not detect any in brain 63. Thus, the inefficacious therapeutic results may be attributed to the difficulty of tryptanthrin crossing blood-brain barrier. Third, on the other hand, the toxicity and side effect of tryptanthrin could also be concerned.
4.
Conclusion
This study demonstrated that tryptanthrin exhibits anti-cryptococcal activity against C.
neoformans and C. gattii which has not been reported in the past. It also exhibits antifungal activity against clinical azole-resistant isolates. We found the synergistic effect between tryptanthrin and FK506. Though the direct tryptanthrin-involved pathways has not been identified, we proved that the cell cycle regulation is interfered by tryptanthrin and cause cell cycle arrest in G1/S phase. The phenomenon of cell cycle arrest and the change in mRNA expression of cell-cycle related genes may attribute to other upstream pathways or transcription factors. Besides, we also tested the potential therapeutic efficacy in murine model. In further experiments, we could modify its chemical structure or invent a better formulation to make tryptanthrin an utilizable natural product.
5.
Future work
Our data suggested that tryptanthrin exhibits anti-cryptococcal activity in vitro, but only few isolates were tested. If more clinical isolates could be obtained and tested, the antifungal activity against Cryptococcus would be more solid. We found that tryptanthrin is synergistic with FK506 in vitro. Meanwhile, FK506 is an immunosuppressant which was reported its drawback outweighed antifungal activity in vivo. The combination therapy of tryptanthrin and FK506 may improve the efficacy. As to the toxicity of tryptanthrin in mice,
we may conduct toxicology assay in mammalian cell line. In addition, the derivatives of tryptanthrin could also be tested for antifungal activity. Derivatives with great solubility and better potency is our expectation. We have verified the effect of tryptanthrin on cell cycle related genes in C. neoformans, the further confirmation related to the changes in protein level is needed. Hopefully a clarified pathway of the growth inhibition by tryptanthrin can be established.
6.
Acknowledgement
We thank Dr. Yu-Liang Yang for technical support for the synthesis of the natural product tryptanthrin and Dr. Hsiang-Kuang Tseng for generously sharing the C. neoformans deletion mutant library. We especially thank Ya-Lin’s committee members Dr. Yu-Liang Yang, Dr. Hsiang-Kuang Tseng and Dr. Yen-Ping Hsueh for sage advice. YL Chen receives research grants MOST 102-230-B-002-041-MY2 and 104-2320-B-002-063-MY3 from the Ministry of Science & Technology, and 104AS-10.7.3-BQ-B1 (5) from the Bureau of Animal and Plant Health Inspection and Quarantine in Taiwan. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest or conflict with the subject matter or materials discussed in the manuscript.
7. References
1. Denning DW, Bromley MJ. Infectious Disease. How to bolster the antifungal pipeline. Science (New York, NY) 2015; 347:1414-6.
2. Roemer T, Krysan DJ. Antifungal drug development: challenges, unmet clinical needs, and new approaches. Cold Spring Harbor perspectives in medicine 2014;
4:a019703.
3. Sanglard D. Emerging threats in antifungal-resistant fungal pathogens. Front Med 2016; 3:1-10.
4. Loftus BJ, Fung E, Roncaglia P, Rowley D, Amedeo P, Bruno D, et al. The genome of the basidiomycetous yeast and human pathogen Cryptococcus neoformans. Science (New York, NY) 2005; 307:1321-4.
5. May RC, Stone NR, Wiesner DL, Bicanic T, Nielsen K. Cryptococcus: from environmental saprophyte to global pathogen. Nature reviews Microbiology 2016;
14:106-17.
6. Hull CM, Heitman J. Genetics of Cryptococcus neoformans. Annual review of genetics 2002; 36:557-615.
7. Desalermos A, Kourkoumpetis TK, Mylonakis E. Update on the epidemiology and management of cryptococcal meningitis. Expert opinion on pharmacotherapy 2012;
13:783-9.
8. Park BJ, Wannemuehler KA, Marston BJ, Govender N, Pappas PG, Chiller TM.
Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS (London, England) 2009; 23:525-30.
9. Sloan DJ, Parris V. Cryptococcal meningitis: epidemiology and therapeutic options.
Clinical Epidemiology 2014; 6:169-82.
10. Perfect JR, Dismukes WE, Dromer F, Goldman DL, Graybill JR, Hamill RJ, et al.
Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the Infectious Diseases Society of America. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America 2010; 50:291-322.
11. Bicanic T, Harrison T, Niepieklo A, Dyakopu N, Meintjes G. Symptomatic relapse of HIV-associated cryptococcal meningitis after initial fluconazole monotherapy: the role of fluconazole resistance and immune reconstitution. Clinical infectious
diseases : an official publication of the Infectious Diseases Society of America 2006;
43:1069-73.
12. Smith KD, Achan B, Hullsiek KH, McDonald TR, Okagaki LH, Alhadab AA, et al.
Increased antifungal drug resistance in clinical isolates of Cryptococcus neoformans in Uganda. Antimicrobial Agents and Chemotherapy 2015; 59:7197-204.
13. Krysan DJ. Toward improved anti-cryptococcal drugs: novel molecules and repurposed drugs. Fungal genetics and biology : FG & B 2014; 78:93-8.
14. Mitsuyama J, Nomura N, Hashimoto K, Yamada E, Nishikawa H, Kaeriyama M, et al. In vitro and in vivo antifungal activities of T-2307, a novel arylamidine.
Antimicrobial Agents and Chemotherapy 2008; 52:1318-24.
15. Dou D, Alex D, Du B, Tiew K-C, Aravapalli S, Mandadapu SR, et al. Antifungal activity of a series of 1,2-benzisothiazol-3(2H)-one derivatives. Bioorganic &
Medicinal Chemistry 2011; 19:5782-7.
16. Baxter BK, DiDone L, Oga D, Schor S, Krysan DJ. Identification, in vitro activity
and mode of action of phosphoinositide-dependent-1 kinase inhibitors as antifungal molecules. ACS chemical biology 2011; 6:502-10.
17. Mor V, Rella A, Farnoud AM, Singh A, Munshi M, Bryan A, et al. Identification of a new class of antifungals targeting the synthesis of fungal sphingolipids. mBio 2015;
6: e00647-15.
18. Zhai B, Lin X. Evaluation of the anticryptococcal activity of the antibiotic polymyxin B in vitro and in vivo. International journal of antimicrobial agents 2013; 41:250-4.
19. Zhai B, Zhou H, Yang L, Zhang J, Jung K, Giam C-Z, et al. Polymyxin B, in combination with fluconazole, exerts a potent fungicidal effect. Journal of antimicrobial chemotherapy 2010; 65:931-8.
20. Butts A, Koselny K, Chabrier-Rosello Y, Semighini CP, Brown JC, Wang X, et al.
Estrogen receptor antagonists are anti-cryptococcal agents that directly bind EF hand proteins and synergize with fluconazole in vivo. MBio 2014; 5:e00765-13.
21. Wiederhold NP, Najvar LK, Bocanegra R, Kirkpatrick WR, Sorrell TC, Patterson TF.
Limited activity of miltefosine in murine models of cryptococcal meningoencephalitis and disseminated cryptococcosis. Antimicrob Agents Chemother 2013; 57:745-50.
22. Ravu RR, Chen YL, Jacob MR, Pan X, Agarwal AK, Khan SI, et al. Synthesis and antifungal activities of miltefosine analogs. Bioorganic & medicinal chemistry letters 2013; 23:4828-31.
23. von Sommaruga E. Über die Moleculargröfse des Indigos. European Journal of Organic Chemistry 1879; 195:302-13.
24. Pathania AS, Kumar S, Guru SK, Bhushan S, Sharma PR, Aithagani SK, et al. The synthetic tryptanthrin analogue suppresses STAT3 signaling and induces caspase dependent apoptosis via ERK up regulation in human leukemia HL-60 cells. PloS one 2014; 9:e110411.
25. Chiang YR, Li A, Leu YL, Fang JY, Lin YK. An in vitro study of the antimicrobial effects of indigo naturalis prepared from Strobilanthes formosanus Moore. Molecules (Basel, Switzerland) 2013; 18:14381-96.
26. Jahng Y. Progress in the studies on tryptanthrin, an alkaloid of history. Archives of pharmacal research 2013; 36:517-35.
27. Ishihara T, Kohno K, Ushio S, Iwaki K, Ikeda M, Kurimoto M. Tryptanthrin inhibits nitric oxide and prostaglandin E(2) synthesis by murine macrophages. European journal of pharmacology 2000; 407:197-204.
28. Recio MC, Cerda-Nicolas M, Potterat O, Hamburger M, Rios JL. Anti-inflammatory and antiallergic activity in vivo of lipophilic Isatis tinctoria extracts and tryptanthrin.
Planta medica 2006; 72:539-46.
29. Honda G, Tabata M. Isolation of antifungal principle tryptanthrin, from Strobilanthes cusia O. Kuntze. Planta medica 1979; 36:85-90.
30. Seifert K, Unger W. Insecticidal and fungicidal compounds from Isatis tinctoria.
Zeitschrift fur Naturforschung C, Journal of biosciences 1994; 49:44-8.
31. Kawakami J, Matsushima N, Ogawa Y, Kakinami H, Nakane A, Kitahara H, et al.
Antibacterial and antifungal activities of tryptanthrin derivatives. Transactions of the Materials Research Society of Japan 2011; 36:603-6.
32. Honda G, Tabata M. Isolation of antifungal principle tryptanthrin, from Strobilanthes cusia O. Kuntze. Planta medica 1979; 36:85-6.
33. Bandekar PP, Roopnarine KA, Parekh VJ, Mitchell TR, Novak MJ, Sinden RR.
Antimicrobial activity of tryptanthrins in Escherichia coli. Journal of medicinal chemistry 2010; 53:3558-65.
34. Chan HL, Yip HY, Mak NK, Leung KN. Modulatory effects and action mechanisms of tryptanthrin on murine myeloid leukemia cells. Cellular & molecular immunology 2009; 6:335-42.
35. Miao S, Shi X, Zhang H, Wang S, Sun J, Hua W, et al. Proliferation-attenuating and apoptosis-inducing effects of tryptanthrin on human chronic myeloid leukemia K562 cell line in vitro. International Journal of Molecular Sciences 2011; 12:3831-45.
36. Liao X, Leung KN. Tryptanthrin induces growth inhibition and neuronal differentiation in the human neuroblastoma LA-N-1 cells. Chemico-biological interactions 2013; 203:512-21.
37. Chang H-N, Huang S-T, Yeh Y-C, Wang H-S, Wang T-H, Wu Y-H, et al. Indigo naturalis and its component tryptanthrin exert anti-angiogenic effect by arresting cell cycle and inhibiting Akt and FAK signaling in human vascular endothelial cells.
Journal of Ethnopharmacology 2015; 174:474-81.
38. Ohkusu M, Hata K, Takeo K. Bud emergence is gradually delayed from S to G2 with progression of growth phase in Cryptococcus neoformans. FEMS microbiology letters 2001; 194:251-5.
39. Liu OW, Chun CD, Chow ED, Chen C, Madhani HD, Noble SM. Systematic genetic analysis of virulence in the human fungal pathogen Cryptococcus neoformans. Cell 2008; 135:174-88.
40. Shapiro RS, Robbins N, Cowen LE. Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiology and molecular biology reviews : MMBR 2011; 75:213-67.
41. Lin X, Nielsen K, Patel S, Heitman J. Impact of mating type, serotype, and ploidy on the virulence of Cryptococcus neoformans. Infection and Immunity 2008; 76:2923- 38.
42. Mitchell AP. Updated view of Cryptococcus neoformans mating type and virulence.
Infection and Immunity 2003; 71:4829-30.
43. Thompson GR, 3rd, Wiederhold NP, Fothergill AW, Vallor AC, Wickes BL, Patterson TF. Antifungal susceptibilities among different serotypes of Cryptococcus gattii and Cryptococcus neoformans. Antimicrob Agents Chemother 2009; 53:309-11.
44. Matar MJ, Ostrosky-Zeichner L, Paetznick VL, Rodriguez JR, Chen E, Rex JH.
Correlation between E-test, disk diffusion, and microdilution methods for antifungal susceptibility testing of fluconazole and voriconazole. Antimicrob Agents Chemother 2003; 47:1647-51.
45. Kraus PR, Boily MJ, Giles SS, Stajich JE, Allen A, Cox GM, et al. Identification of Cryptococcus neoformans temperature-regulated genes with a genomic-DNA microarray. Eukaryotic cell 2004; 3:1249-60.
46. Odom A, Del Poeta M, Perfect J, Heitman J. The immunosuppressant FK506 and its nonimmunosuppressive analog L-685,818 are toxic to Cryptococcus neoformans by inhibition of a common target protein. Antimicrobial Agents and Chemotherapy 1997;
41:156-61.
47. Yang DH, Jung KW. Rewiring of signaling networks modulating thermotolerance in the human pathogen Cryptococcus neoformans. Genetics 2017; 205:201-19.
48. Shekhar-Guturja T, Gunaherath GM, Wijeratne EM, Lambert JP, Averette AF, Lee SC,
et al. Dual action antifungal small molecule modulates multidrug efflux and TOR signaling. Nat Chem Biol 2016; 12:867-75.
49. Odom A, Muir S, Lim E, Toffaletti DL, Perfect J, Heitman J. Calcineurin is required for virulence of Cryptococcus neoformans. The EMBO Journal 1997; 16:2576-89.
50. Chen YL, Lehman VN, Averette AF, Perfect JR, Heitman J. Posaconazole exhibits in vitro and in vivo synergistic antifungal activity with caspofungin or FK506 against Candida albicans. PloS one 2013; 8:e57672.
51. Del Poeta M, Cruz MC, Cardenas ME, Perfect JR, Heitman J. Synergistic antifungal activities of bafilomycin A(1), fluconazole, and the pneumocandin MK-
0991/caspofungin acetate (L-743,873) with calcineurin inhibitors FK506 and L- 685,818 against Cryptococcus neoformans. Antimicrob Agents Chemother 2000;
44:739-46.
52. Liu J, Farmer JD, Lane WS, Friedman J, Weissman I, Schreiber SL. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991;
66:807-15.
53. Odds FC, Brown AJ, Gow NA. Antifungal agents: mechanisms of action. Trends in microbiology 2003; 11:272-9.
54. García-Rodas R, Cordero RJB, Trevijano-Contador N, Janbon G, Moyrand F,
Casadevall A, et al. Capsule growth in Cryptococcus neoformans is coordinated with cell cycle progression. mBio 2014; 5:e00945-14.
55. Hawser S, Islam K. Comparisons of the effects of fungicidal and fungistatic antifungal agents on the morphogenetic transformation of Candida albicans. The Journal of antimicrobial chemotherapy 1999; 43:411-3.
56. Kelliher CM, Leman AR, Sierra CS, Haase SB. Investigating conservation of the cell- cycle-regulated transcriptional program in the fungal pathogen, Cryptococcus
neoformans. PLoS Genetics 2016; 12:e1006453.
57. Song M-H, Lee J-W, Kim MS, Yoon J-K, White TC, Floyd A, et al. A flucytosine- responsive Mbp1/Swi4-like protein, Mbs1, plays pleiotropic roles in antifungal drug resistance, stress response, and virulence of Cryptococcus neoformans. Eukaryotic cell 2012; 11:53-67.
58. Haase SB, Wittenberg C. Topology and control of the cell-cycle-regulated transcriptional circuitry. Genetics 2014; 196:65-90.
59. Florio AR, Ferrari S, De Carolis E, Torelli R, Fadda G, Sanguinetti M, et al. Genome- wide expression profiling of the response to short-term exposure to fluconazole in Cryptococcus neoformans serotype A. BMC microbiology 2011; 11:97.
60. Cardenas ME, Cutler NS, Lorenz MC, Di Como CJ, Heitman J. The TOR signaling cascade regulates gene expression in response to nutrients. Genes & development 1999; 13:3271-9.
61. Liu T-B, Kim J-C, Wang Y, Toffaletti DL, Eugenin E, Perfect JR, et al. Brain inositol is a novel stimulator for promoting Cryptococcus penetration of the blood-brain barrier. PLOS Pathogens 2013; 9:e1003247.
62. Jähne EA, Eigenmann DE, Sampath C, Butterweck V, Culot M, Cecchelli R, et al.
Pharmacokinetics and in vitro blood-brain barrier screening of the plant-erived alkaloid tryptanthrin. Planta medica 2016; 82:1021-9.
63. Zhang N, Hua Y, Wang C, Sun Y, Wang Z, Liu Z, et al. Distribution study of tryptanthrin in rat tissues by HPLC and its relationship with meridian tropism of
Indigo naturalis in traditional Chinese medicine. Biomedical chromatography : BMC 2014; 28:1701-6.
64. Perfect JR, Ketabchi N, Cox GM, Ingram CW, Beiser CL. Karyotyping of Cryptococcus neoformans as an epidemiological tool. Journal of clinical microbiology 1993; 31:3305-9.
65. Wickes BL, Mayorga ME, Edman U, Edman JC. Dimorphism and haploid fruiting in Cryptococcus neoformans: association with the alpha-mating type. Proceedings of the National Academy of Sciences 1996; 93:7327-31.
66. Cruz MC, Del Poeta M, Wang P, Wenger R, Zenke G, Quesniaux VFJ, et al.
Immunosuppressive and Nonimmunosuppressive Cyclosporine Analogs Are Toxic to the Opportunistic Fungal Pathogen Cryptococcus neoformans via Cyclophilin- Dependent Inhibition of Calcineurin. Antimicrobial Agents and Chemotherapy 2000;
44:143-9.
67. Kidd SE, Hagen F, Tscharke RL, Huynh M, Bartlett KH, Fyfe M, et al. A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proceedings of the National Academy of Sciences of the United States of America 2004; 101:17258-63.
68. Kidd SE, Guo H, Bartlett KH, Xu J, Kronstad JW. Comparative gene genealogies indicate that two clonal lineages of Cryptococcus gattii in British Columbia resemble strains from other geographical areas. Eukaryotic cell 2005; 4:1629-38.
69. Gillum AM, Tsay EY, Kirsch DR. Isolation of the Candida albicans gene for
orotidine-5′-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E. coli pyrF mutations. Molecular and General Genetics MGG 1984; 198:179-82.
70. Pfaller MA, Bale M, Buschelman B, Lancaster M, Espinel-Ingroff A, Rex JH, et al.
Selection of candidate quality control isolates and tentative quality control ranges for in vitro susceptibility testing of yeast isolates by National Committee for Clinical Laboratory Standards proposed standard methods. Journal of clinical microbiology 1994; 32:1650-3.
71. Winston F, Dollard C, Ricupero-Hovasse SL. Construction of a set of convenient Saccharomyces cerevisiae strains that are isogenic to S288C. Yeast (Chichester, England) 1995; 11:53-5.
72. Nierman WC, Pain A, Anderson MJ, Wortman JR, Kim HS, Arroyo J, et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus.
Nature 2005; 438:1151-6.
73. Ghannoum MA, Arthington-Skaggs B, Chaturvedi V, Espinel-Ingroff A, Pfaller MA, Rennie R, et al. Interlaboratory study of quality control isolates for a broth
microdilution method (modified CLSI M38-A) for testing susceptibilities of Dermatophytes to antifungals. Journal of clinical microbiology 2006; 44:4353-6.
Table.1 Fungal strains used in this study.
Strain Description Background Reference
Cryptococcus neoformans
H99 Wild-type MATa, serotype A Clinical isolate 64
JEC21 Wild-type MATa, serotype D Clinical isolate 65
T1 fluconazole resistant Clinical isolate 66
89-610 fluconazole resistant Clinical isolate 66
Cryptococcus gattii
R265 Wild-type VG IIa MATa, serotype B Clinical isolate 67 R272 Wild-type VG IIb MATa, serotype B Clinical isolate 67
WM276 Wild-type VG I Environmental isolate 68
Candida albicans
SC5314 Wild-type Clinical isolate 69
Candida krusei
ATCC 6258 Wild-type Clinical isolate 70
Candida parapsilosis
ATCC 22019 Wild-type Clinical isolate 70
Saccharomyces cerevisiae
BY4741 MATa his3Δ1 leu2Δ0 met15Δ0
ura3Δ0 Laboratory strain 71
Aspergillus fumigatus
AF293 Wild-type Clinical isolate 72
Trichophyton rubrum
MYA-4438 Wild-type Clinical isolate 73
Fusarium solani
Fungus III-6 Wild-type Clinical isolate
Table 2. Minimum inhibitory concentration of tryptanthrin against human fungal pathogen.
Strain MIC100 (g/mL MFC (g/mL
C. neoformans (4) 2 4 - >64
C. gattii (3) 4 >64
C. albicans (1) >64 -
C. krusei (1) >64 -
C. parapsilosis (1) >64 -
A. fumigatus (1) >64 -
F. solani (1) >64 -
T. rubrum (1) 8 32
Table 3. Fractional inhibitory concentration of tryptanthrin with FK506, fluconazole (FLC) or amphotericin B (AmB).
MIC MIC combined FIC index
Strain TRP FK506 FLC AmB (TRP;FK506) (TRP;FLC) (TRP;AmB) (TRP;
FK506)
(TRP;
FLC)
(TRP;
AmB)
Cryptococcus neoformans
H99 2 >16 16 2 0.25; 0.03 1; 8 1; 1 0.125 1 1
T1 2 >16 >32 0.5 1; 0.12 2; 0.06 1; 0.5 0.503 1 1.5
89-610 2 >16 32 1 1; 0.12 2;0.06 1; 0.5 0.503 1 1
Cryptococcus gattii
R265 4 >16 16 2 1; 0.03 4; 0.06 2; 1 0.251 1 1
R272 4 >16 >32 0.5 0.25; 0.06 4; 0.06 2; 0.5 0.062 1 1.5
WM276 4 >16 32 0.5 2; 0.03 4; 0.06 2; 0.5 0.501 1 1.5
Table 4. Deletion mutants of C. neoformans with altered susceptibility to tryptanthrin.
deletion mutant CNAG_*****
C.n.
name homolog fitness description of function Enzymes
03981 PFA4(S.c.) HS palmitoyltransferase PFA4
01232 PMC1 HS calcium-translocating P-type ATPase, PMCA-type
03525 NTH2 R alpha,alpha-trehalase
04735 MEP1 R metalloproteinase
Metabolism
01981 HMT2(S.p.) HS sulfide:quinone oxidoreductase
02833 R NADH dehydrogenase (ubiquinone) subcomplex 7
05540 URE1 R urease
07180 R NAD-dependent histone deacetylase
Transporter
04704 JEN1(S.c.) R MFS transporter, SHS family, lactate transporter 03664 NIC1 R high-affinity nickel-transporter
05330 MPH2(S.c.) R
MFS transporter, SP family, glucoside: H+
symporter
05812 KHA1(S.c.) R potassium: hydrogen antiporter Cell cycle
03184 BUB1(S.p.) R BUB protein kinase
04162 PKA2 R AGC/PKA protein kinase
05216 RAD53 R non-specific serine/threonine protein kinase 03900 ACP2(S.p.) R F-actin-capping protein beta subunit
06335 KIP2(S.c.) R centromeric protein E MAPK signaling pathway
04090 ATF1 R activating transcription factor 03938 CPR2 R pheromone a factor receptor 02531 CPK1 R mitogen-activated protein kinase Transcription
04545 LSG1(S.p.) R CTD kinase subunit gamma
00440 SSN801 R cycin C /Cyclin subunit of Mediator subcomplex
01217 MPE1(S.c.) R protein MPE1
05221 HTZ1(S.c.) R histone H2A.Z
00073 FAR8(S.p.) R nuclear mRNA splicing protein 02215 HAP3 R transcriptional activator
01551 GAT201 R transcription factor
03409 SKN7 R osomolarity two-component system, response regulator
Nucleus
06224 MLN2 HS nuclear movement protein nudC
01747 PMS1 R DNA mismatch repair protein PMS1
05301 CRN1 R deoxyuridine 5'-triphosphate nucleotidohydrolase 02090 GPA3 R guanine nucleotide-binding protein subunit alpha 00445 NHP6B01 R non-histone chromosomal protein
Ubiquitin system
03151 CSN4 HS COP9 signalosome complex subunit 4
05765 UBC6 R ubiquitin--protein ligase
04208 R Ataxin-3
Table 5. Oligonucleotide primers used in this study.
Primer Use Sequence (5’ →3’)
JC1052 C.n. ACT1 CCACACTGTCCCCATTTACGA
JC1053 C.n. ACT1 CAGCAAGATCGATACGGAGGAT
JC1054 C.n. CLN1 CATTCCCATAGCATTGATTTCCT
JC1055 C.n. CLN1 TTCGGACATCCCATACCATGAT
JC1363 C.n. CNAG_00995 CGCCACTTCTGCTCCTTACCTT
JC1364 C.n. CNAG_00995 TAGTCTCTCGGATGGCAGCAAG
JC1367 C.n. MBS1 CGCCCGAATGTATGTGGTATGT
JC1368 C.n. MBS1 TTGTCGAAACCGGCTACTTTCA
JC1411 C.n. ERG11 AACTTGCCCCTTCCCAGTTACA
JC1412 C.n. ERG11 CTTGCAGCTCTGGAGGTTTTCA
JC1415 C.n. PCL103 TGTTTCCGGTTAGCGAGATCAA
JC1416 C.n. PCL103 CAAGAACGGTTGGAAGTTGTCG
JC1417 C.n. WHI5 TGCATCAACTGCAACAACGACT
JC1418 C.n. WHI5 TTGAAAGGGAGTCGAAGGAAGC
JC1413 C.n. TAP42 TGTCTGCTTCAACTCCCGTCTC
JC1414 C.n. TAP42 TCTTCCGCCATGTCATCTTCAT