國立臺灣大學生物資源暨農學院動物科學技術學系 碩士論文
Department of Animal Science and Technology College of Bioresources and Agriculture
National Taiwan University Master Thesis
豬隻運輸過程中緊迫源與唾液腎上腺 皮質醇之關聯性
The Correlation between Stressors and Salivary Cortisol during Transportation in Swine
陳正軒
Jeng-Shiuan Chen
指導教授:鍾德憲、邱智賢 博士
Advisor: De-Shien Jong, Chih-Hsien Chiu, Ph.D.
中華民國 108 年 7 月
July 2019
致謝
人生漫漫,充斥著各種機緣與巧合,就像數年前我曾信誓旦旦覺得自己不會念 研究所,現在卻正在提筆為碩士班生活寫下總結。是的,這兩年多來的經歷像是場 驚奇的旅程,我在其中經歷了許多,有過情緒,有過懷疑,曾為了研究有所進展而 興奮、為了研究遭遇困難與挫折而失落,曾質問自己是否有能力面對接踵而至的挑 戰,也曾想逃避和放棄。若不是一路上有許多人點燈,我想我可能沒法走到今天。
為此,我十分感激,難以忘懷。
在碩士班七百多個日夜裡,我接受過許多人的幫助。感謝鍾德憲老師、邱智賢 老師與吳兩新老師的諄諄教誨,提供研究的大方向讓我去思考,並且修正思路的邏 輯性,同時要求留意細節,讓我在科學上有所累積,解決問題與表達陳述的能力亦 得以培養。感謝中興大學林怡君老師,提供了本研究的雛型,並在執行的過程中多 次給予建議與協助,且為我在動物福利科學上指點迷津。感謝賴俊甫先生對我的歡 迎和照顧,讓我在屏東可以順利進行研究,感謝賴英傑先生與林原聰先生載我見識 豬隻運輸的現況,不嫌麻煩地分享運輸中的見聞。感謝屏東科技大學楊國泰老師借 用研究相關器材,讓我在屏東採集到檢體後能做初步的處理。感謝生理研究室的夥 伴,沐恩學長、孟傑學長、黃謙學長、恩加學長、秀如、思芃、宜軒、張珉、于翔 與助理們,無論在實驗或生活上都給予了我許多幫助。能接受這麼多人的幫助,我 認為自己非常地幸運,也由衷地感謝大家。
最後感謝我的父母從小到大對我的扶養與教導。兩年的時間我完成了碩士學 業,而父母卻更加年邁,白駒過隙,前些日子意外地發現雙親的衰老更甚以往,是 他們的青春成就了我,希望我這一點時光淬鍊的結果能讓他們驕傲。
中文摘要
隨著家畜飼養方式集約化與規模化,活體運輸在現代肉豬生產過程中時屬必 要,運輸期間動物是否感到緊迫、痛苦逐漸引起社會大眾的關心。因此,動物運輸 在近年來已成為動物福利探討的重要議題之一。本試驗旨在以高屏地區商用肉豬 為對象,調查運輸過程中豬隻唾液腎上腺皮質醇含量與緊迫源的關聯。每趟運輸隨 機採集 1 至 3 隻豬隻,於牧場繫留欄位內、牧場車體上和肉品市場車體上之唾液 樣本,以酵素聯結免疫吸附分析法(ELISA)檢測腎上腺皮質醇(Cortisol)含量,
記錄運輸過程中各項緊迫源變化,再以簡單迴歸、多元複迴歸與非線性迴分別歸探 討兩者間之關連,並在調查前針對唾液採集裝置對皮質醇的回收率、唾液腎上腺皮 質醇的日夜節律以及豬隻的緊迫試驗進行基礎測驗,確保調查之準確性。
基礎測驗結果顯示:本試驗所使用的唾液採集裝置 Cortisol-Salivette®有良好 的皮質醇回收率;唾液皮質醇的日夜節律高峰在09:00、11:00 以及 15:00;緊迫試 驗中,豬隻唾液皮質醇可以即時地反映豬隻緊迫程度。運輸調查結果發現:電擊棒 使用頻率、豬隻承載密度和豬隻驅趕前後的皮質醇變化無顯著關聯(P=0.9184, 0.5294)。豬隻承載密度和豬隻運輸前後皮質醇變化亦無顯著關聯(P=0.2456)。驅 趕與運輸時間和豬隻在驅趕及運輸前後的皮質醇變化無顯著關聯(P=0.6760, 0.5520),但當運輸時間界於 35 至 70 分鐘時,運輸時間與皮質醇變化有顯著關聯
(R2=0.5699, P=0.0187)。而豬隻驅趕前後的皮質醇變化與豬隻運輸前後的皮質醇 變化有顯著的關聯(R2=0.3947, P=0.0161)。總時間與豬隻在整趟運輸前後的皮質 醇變化則存在幾乎顯著的非線性關聯(P=0.0871)。
本研究以調查台灣運輸現況為主軸,調查結果顯示,在現行豬隻運輸過程與操 作中,電棒使用頻率與豬隻承載密度等被視為影響動物福利的緊迫源,與豬隻皮質 醇變化沒有顯著關聯性。而運輸時間(介於35 至 70 分鐘)與皮質醇變化的顯著關
聯顯示豬隻在此時間範圍內,運輸時間越長,皮質醇含量越低,說明豬隻有適應運 輸緊迫的能力;豬隻驅趕前後皮質醇變化與運輸前後皮質醇變化之間的顯著關聯,
則顯示部分豬隻對於驅趕緊迫並不敏感。
總上所述,雖無法明確得知影響豬隻驅趕和運輸的主要緊迫源為何,但可確定 豬隻驅趕前後與運輸前後的腎上腺皮質醇含量變化,兩者之間有關連。肉豬場應於 豬隻在進入車體前給予良好照護,以提升豬隻運輸時之動物福利。
關鍵語:豬隻動物福利、動物運輸、緊迫、腎上腺皮質醇
Abstract
With the scale-intensive production of animal husbandry, whether animals feel stressed during transportation has gradually aroused the concern of the public. Thus, animal transportation has become an important issue in animal welfare discussion in recent years. The study aimed to investigate the transportation of commercial pigs in Southern Taiwan. The saliva samples were taken from several pigs randomly before loading, after loading and arrival at the meat market in each transportation. The cortisol level was tested via ELISA. The relationships between cortisol level and various stressors were analyzed via simple linear regression, multiple regression and nonlinear regression.
Basal experiments were done before investigation, including recovery rate of saliva collection device, circadian rhythm of salivary cortisol in swine and response of salivary cortisol to stress in swine, in order to ensure the accuracy of investigation.
The results of basal experiments showed the saliva collection device in this study has acceptable recovery rate; peaks of circadian rhythm of salivary cortisol are at 09:00, 11:00 and 15:00; salivary cortisol in swine can instantly reflect the degree of stress. The results of investigation of transportation showed no significant correlation between cortisol change after loading and stressors, for example, the use of electric sticks (P=0.9184), loading density (P=0.5294) and loading time (P=0.6760). Same results were
found between loading density and cortisol change after transport (P=0.2456). Similarly, no significant correlation was found between cortisol change after transport and transport time (P=0.5520). However, there was a significant correlation between time and cortisol change after transport when the duration is between 35 to 70 minutes (R2=0.5699, P=0.0187). Also, a significant correlation was found between cortisol change after loading and after transport (R2=0.3947, P=0.0161). There is an almost significant nonlinear correlation between total time and cortisol changes (P=0.0871).
This study focused on the current status of swine transportation in Taiwan.
Interestingly, during the process and the handling of transportation, the use of electric sticks and loading density, the stressors that assumed to compromise animal welfare, have no significant correlation with cortisol changes in pigs. Whether the pigs are able to adapt in the situation of transportation requires further studies. Correlation between cortisol change after loading and after transport showed part of pigs are not sensitive to loading stress.
In conclusion, it’s sure that cortisol change after transport is related to cortisol change after loading although the main stressors that exactly affect loading and transport are unclear. Good animal care should be provided when loading pigs into vehicle, in order to improve animal welfare during transportation.
Key Words: Animal welfare, Animal transportation, Stressors, Cortisol
目錄
口試委員會審定書 ... i
致謝 ... ii
中文摘要 ... iii
英文摘要 ... v
目錄 ... vii
圖目錄 ... ix
表目錄 ... x
第一章 前言 ... 1
第二章 文獻探討 ... 2
2.1 認識動物福利 ... 2
2.2 動物之緊迫生理 ... 5
2.3 運輸過程中緊迫源與其對動物福利之影響 ... 25
第三章 材料與方法 ... 30
3.1 試驗動物與場地 ... 30
3.2 檢體採集方法 ... 31
3.3 試驗設計 ... 31
3.4 腎上腺皮質醇酵素免疫分析法 ... 34
3.5 統計模式 ... 36
第四章 結果 ... 37
4.1 基礎試驗結果 ... 37
4.2 運輸緊迫源與腎上腺皮質醇之關聯 ... 44
第五章 討論 ... 63
5.1 研究限制 ... 63
5.2 唾液採集裝置 Cortisol-Salivette®之回收率 ... 67
5.3 豬隻唾液皮質醇之日夜節律 ... 67
5.4 豬隻緊迫試驗 ... 69
5.5 運輸緊迫源與腎上腺皮質醇之關聯 ... 70
第六章 結論 ... 73
第七章 參考文獻 ... 74
第八章 附錄 ... 84
圖目錄
圖1. 動物對緊迫之生理反應之模型框架 ... 8
圖2. 類固醇內泌素之生成路徑 ... 12
圖3. 腎上腺皮質醇代謝分解示意圖 ... 14
圖4. 腎上腺皮質醇代謝分解路徑圖 ... 14
圖5. 動物應對緊迫源之假想進程 ... 18
圖6. 唾液採集裝置 ... 32
圖7. 運輸各階段 ... 32
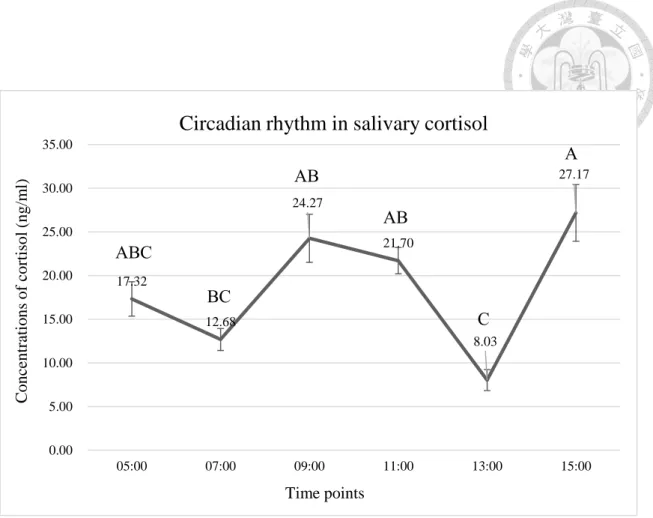
圖8. 豬隻唾液皮質醇之日夜節律 ... 41
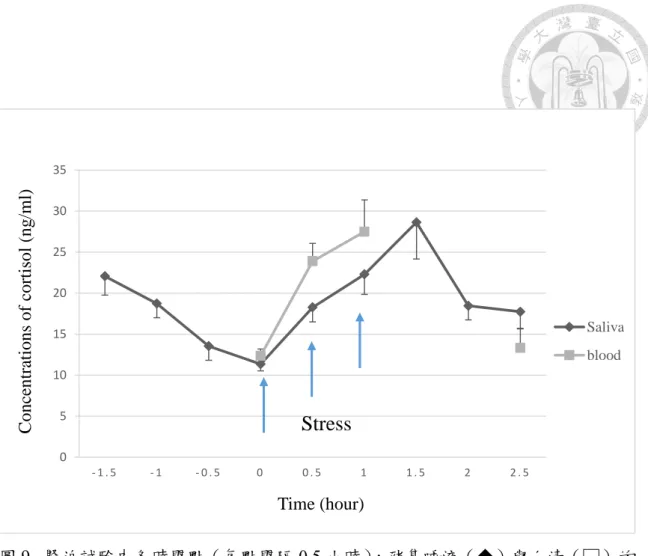
圖9. 緊迫試驗中各時間點的豬隻與血清皮質醇濃度 ... 42
圖10. 表現相近的豬隻在緊迫試驗中各時間點的平均唾液皮質醇濃度 ... .43
圖11. 運輸中三個採集點之平均唾液皮質醇濃度 ... 53
圖12. 驅趕時間與驅趕緊迫之簡單直線迴歸圖 ... 54
圖13. 豬隻承載密度與驅趕緊迫之簡單直線迴歸圖 ... 55
圖14. 電棒使用頻率與驅趕緊迫之簡單直線迴歸圖 ... 56
圖15. 運輸時間與運輸緊迫之簡單直線迴歸圖 ... 57
圖16. 運輸時間介於 35 至 70 分鐘時,與運輸緊迫之簡單直線迴歸圖 ... 58
圖17. 豬隻承載密度與運輸緊迫之簡單直線迴歸圖 ... 59
圖18. 驅趕緊迫與運輸緊迫之簡單直線迴歸圖 ... 60
圖19. 驅趕緊迫較低組與較高組在運輸中三個採集點之平均唾液皮質醇濃度 ... 61
圖20. 總運輸時間與總緊迫之間簡單直線迴歸圖 ... 62
圖21. 總運輸時間與以比值表現的總緊迫之散佈圖 ... 62
表目錄
表1. 豬隻唾液皮質醇濃度之日夜節律 ... 39
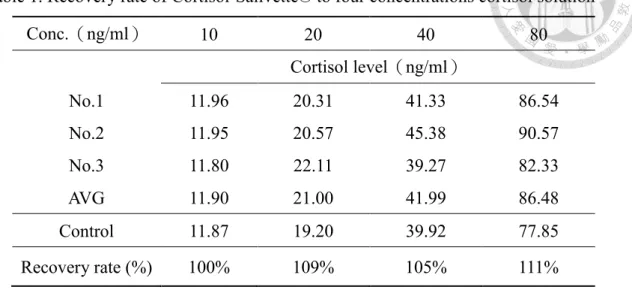
表2. Cortisol-Salivette®對四個皮質醇濃度溶液的回收率 ... 39
表3. 豬隻唾液與血清皮質醇濃度在緊迫測試中之表現 ... 40
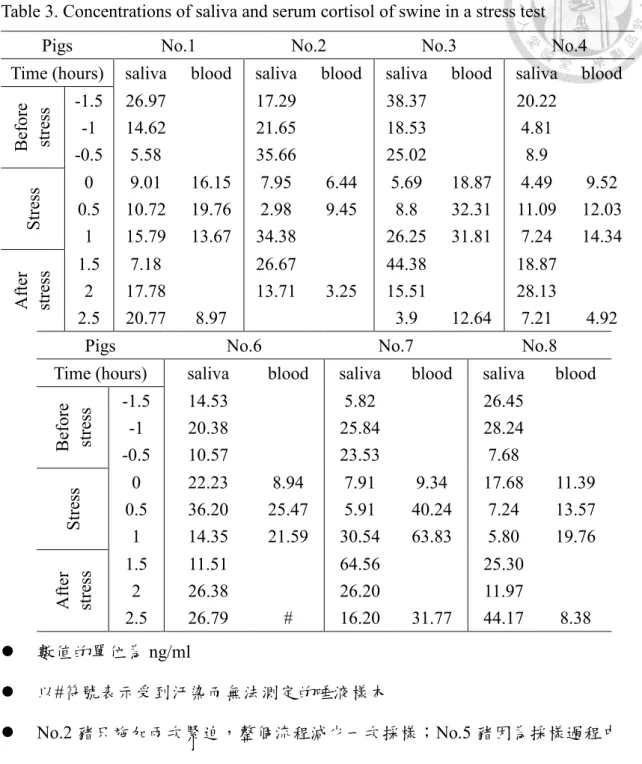
表4. 豬隻在運輸過程中三個採集點的唾液皮質醇濃度 ... 51
表5. 豬隻運輸時每趟旅程之條件 ... 52
第一章 前言
動物運輸是家畜禽在屠宰前必經的程序,且隨著家畜飼養方式的集約化與規 模化,經濟動物的活體運輸在畜產品生產中實屬必要。在運輸期間動物是否受到緊 迫與痛苦,逐漸引起社會大眾的關心。因此,近年來動物運輸已成為動物福利所探 討的重要議題之一。
一般而言,動物運輸被認為是農場動物最主要且無法避免的緊迫源,對動物健 康、福利、表現及產品品質皆具有負面影響(Borell and Schaffer, 2005)。運輸緊 迫源包括對環境改變之恐懼、運輸間禁食造成的飢餓與脫水、車輛微氣候不適造成 的冷熱緊迫、車輛乘載過高密度之動物所造成的擠壓與疼痛以及車輛設計和路況 不良時造成的震動等(Roldan-Santiago et al., 2013),並在運輸過程中對動物造成 傷害,如皮膚與肌肉損傷、肌肉出血、腎上腺皮質醇過度分泌、暈眩嘔吐、昏厥、
無法行走甚至死亡,且動物屠體品質亦會下降,如肌肉肝醣過度消耗所導致的暗乾 肉(DFD)與水漾肉(PSE)等。運輸中緊迫導致了畜產業的重大經濟損失(Ritter et al., 2009),更迫使農場動物在其中承受巨大的痛苦,動物福利受到損害。而這
些緊迫源主要與運輸距離、時間、季節以及人員管理操作等多項因素有關,以非侵 入式的緊迫指標—唾液腎上腺皮質醇,釐清兩者之間的關聯,有助於減輕農場動物 在運輸過程中所遭遇的緊迫,進一步提升運輸過程中的動物福利。
台灣豬隻產業發達,產值佔畜牧總產值約四成六,豬隻在養頭數約540 萬,運 輸遍佈全國,故本試驗以豬隻運輸為主軸,試驗對象為商用肉豬種,試驗地區為高 屏地區,取豬隻唾液中腎上腺皮質醇含量作為緊迫指標,探討各項運輸中的緊迫源 如時間、豬隻密度、電棒使用等與腎上腺皮質醇變化的關聯,期許未來能藉此改善 台灣豬隻運輸之過程。
第二章 文獻探討
2.1 認識動物福利
現今的動物福利(animal welfare)係指動物在人為飼養環境下,應該擁有而非 被給予的基本權利與自由,以避免痛苦之情形及追求正向愉快之經驗,其定義應將 動物基本需求、健康與緊迫等層面,經過科學性的評估與測量後納入考量,對象應 涵蓋伴侶動物、經濟動物、展演動物、實驗動物與任何被人類使用之動物。動物福 利亦包括人類對動物利用系統(animal-usage systems)中的永續性、道德與食物品 質是否符合社會大眾之要求(Broom, 2010)。
而動物福利並非一成不變,隨著社會變遷,動物的飼養與利用不停變化,人類 對待動物的方式與倫理道德亦不斷地更動,動物福利的基本概念便隨著年代的進 展而不停蛻變。因此,認識動物福利的緣由與演變,方能對動物福利有較為通盤的 了解。
2.1.1 動物福利起源與演變
無論東西方文化,自亙古人類馴養動物以來,皆具有禁止虐待動物的倫理與傳 統,避免動物在人類飼養下遭受不必要的痛苦、傷害。直到20 世紀中葉,主流思 想或哲學對於對待動物之倫理已有足夠的共識及關心,然而此時社會正逢劇烈變 遷,使得當時動物之倫理與傳統面臨挑戰與檢討(李,2015)。
動物在農業與畜牧業上的利用,為人類使用動物之主要目的,在20 世紀前葉 以使用經濟動物為主的年代,動物福利與倫理便建立在傳統的畜牧業之上。彼時,
動物提供食物、勞力、纖維、原料,而人類給予庇護,維持動物生存之條件、保護 其不受惡劣天候及掠食者之影響,傳統畜牧業像是動物與人類之間的契約,提供一
個雙贏局面,讓雙方都可以獲益(Rollin, 1995)。在這樣的共生關係下,人類自然 而然會去關心動物的健康、禁止虐待動物、避免動物生病,確保動物能有最佳經濟 表現。此時,對動物的關懷出自於人類需要動物維持健康狀態的人本位思考,而非 動物在人為飼養環境下應該擁有的自由與福利。然而在當時的社會環境下,如此的 動物倫理,已足夠滿足動物之需求,良好的畜牧者會照顧好動物而謀取利益;反之,
當業者不善待動物,會因為動物之表現低落,導致經濟利益損失而被淘汰。
直到20 世紀中葉,拜工業發展所賜,畜牧業迎接了交通運輸與畜產品保存的 技術突破,使畜產品可以銷售至更遠的區域,劇烈地改變當時的產業結構。屠宰與 加工產業可以從廣大腹地取得動物及畜產品,因此邁向集約,畜牧產業需要大量地 生產動物,而促成規模化與企業化,伴隨而來的是生產者之間高度競爭與生產每頭 動物之利益降低。此時飼養動物的模式大幅改變,機械化的設備與科技應用,協助 人類以低人力資本飼養大量的動物,動物不僅獲得較少的照護,還被飼養在與牠們 本質不同的環境中,以方便管理。同時,生產每頭動物所獲得的利益降低勢必影響 提供給動物的福利,當動物在飼養過程中不安適時,經濟的考量會限制生產者能負 擔的照顧成本,使動物福利受損。
這樣的情形在1964 年被 Ruth Harrison 登載於動物機器(Animal Machines)一 書中,此書反映在當時集約飼養制度下,動物遭遇痛苦的狀況,引發社會大眾的震 撼與關注。英國議會因此委託Roger Brambell 成立委員會進行調查,隔年 1965 年 的調查報告(Brambell report)確認了遭受痛苦的動物已然是當時普遍之事實,並 指出動物在人為飼養環境下應該擁有可以站立、躺下、轉身、自我清理以及伸展四 肢的自由。以提升經濟動物之福利為目標。英國於1967 年成立了「農場動物福利 諮詢委員會」(Farm Animal Welfare Advisory Committee),並於 1979 年轉型為「農 場動物福利委員會」(Farm Animal Welfare Council, FAWC),同年提出的五大自由
(Five Freedoms)擴充了 Brambell report 的內容,更明確指出動物在人為飼養下理 應受的待遇,成為舉世皆準的普遍性動物福利目標。
1980 年代開始,隨著社會穩定、文明發展,社會大眾對動物福利的關心大幅 增加,相關的研究亦與日俱增,使動物福利逐漸發展成一門科學,維護及推廣動物 福利的組織相繼成立(Broom, 2010)。動物福利出現明確定義,個體之福利在於其 試圖應對環境的狀態(Broom, 1986),應對環境意味著個體對身體與心理的穩定有 能力控制。至今,動物福利關心的層面亦有了擴張,相較以往著重避免動物遭遇負 面經驗,現今動物福利更強調促使動物擁有正面的經歷或情緒(Mellor, 2009),正 向動物福利評估亦有所發展(Yeates and Main, 2008)。此外,動物福利的優劣也被 消費者作為評估生產系統或程序的標準,成為畜產品價值的一部份,生產過程中擁 有優良動物福利的畜牧產品較能被社會大眾接受,而具備永續性。綜上所述,動物 福利的蓬勃發展,影響到社會的無數層面,人類與動物之間關係與倫理的更迭,正 全面性地改變人們的生活。
2.1.2 淺談五大自由
FAWC 在 1979 年提出的五大自由(Five Freedom)具有歷史性的意義,其內 容涵蓋了維持正常生物功能、自然行為表現與避免負面心理感受等層面,引導社會 大眾對於動物福利的關懷應該有全盤性的考量,因此被普遍地認為是現今動物福 利之基礎。
五大自由分別為:免於飢渴的自由(Freedom from hunger and thirst)、免於不 適的自由(Freedom from discomfort)、免於疼痛受傷與疾病的自由(Freedom from pain, injury or disease)、能表現自然行為的自由(Freedom to express normal behavior)
以及免於恐懼與痛苦的自由(Freedom from fear and distress)。飢渴的狀態會直接影 響動物的存活與否,而身體不適、疼痛與疾病,無法表現自然行為及遭遇痛苦、恐 懼之經驗則降低動物的生活品質,兩者皆非人們樂見。設立五大自由讓動物福利的 關注與評估得以聚焦,避免特定的威脅發生在動物上。然而現今,五大自由僅作為 評估動物福利的初步指南,若需要科學證據佐證,則得針對物種考量動物的諸多需
求,方能更進一步探討動物福利。
2.1.3 動物福利之檢驗
福利(welfare)的概念起源於來自哲學,它並非客觀可測量的事情,因此評估 時難跳脫高度的主觀詮釋,為避免口說無憑,對動物福利的評估應從其定義出發。
根據Broom 於 1986 年所提,動物福利的定義為個體動物試圖適應環境的狀態,其 優劣取決於動物應付環境的狀況,當動物很艱難地應付或無法應付環境時,動物福 利較差。承此定義,欲評估動物福利,需了解動物原先的正常身體功能,以及在環 境的挑戰下動物應對環境之策略與生理狀態之變化,而這些知識屬於緊迫生理學 的範疇。
緊迫生理學(stress physiology)是現今評估動物福利的基礎,主要探討不同緊 迫源刺激下,動物如何反應與適應來維持身體持恆,以及其後果對生物體之影響。
面臨緊迫時,動物擁有許多應對的方式,例如行為、內分泌與免疫等反應,測量其 中各項參數以及原先生理功能參數的變化,能為評估動物福利提供可量化的科學 證據,同時考量遺傳、環境和時間的因素,方能對動物福利做出最客觀的判斷。
2.2 動物之緊迫生理 2.2.1 緊迫之認識
緊迫的意涵經歷了數次演變,其最早的概念出現在 1929 年,由 Cannon 氏所 提出的「戰鬥或逃跑(Fight-or-flight response)」理論。該理論認為動物在認知到一 個可能有害之事件或威脅時,體內的交感神經系統(sympathetic nervous system)
會 被 刺 激 , 促 使 腎 上 腺 髓 質 分 泌 腎 上 腺 素 (adrenaline ) 及 正 腎 上 腺 素
(norepinephrine),引起生理變化,如血糖升高、心跳與呼吸加速、血壓上升等,
最終迅速將動物預備能成戰鬥或逃跑的狀態,使其能有效排解緊急狀況(Cannon, 1929)。1932 年,Cannon 氏提出持恆(homeostasis)的觀念,並假定當持恆受到緊
迫的擾亂時,生物體會設法透過自主神經系統的作用回復到先前的平穩狀態
(Cannon, 1932)。
Cannon 氏的研究與理論被 Selye 氏延續並擴充,1936 年 Selye 氏提出「一般 適應徵候群(general adaptation syndrome, GAS)」,認為有危害之事件或緊迫源會刺 激下視丘-腦垂腺-腎上腺皮質軸(hypothalamo- pituitary- adrenocortical axis, H-P-A 軸),使腎上腺髓、皮質分泌下游內泌素影響生理,並將緊迫引起的反應分為三階 段,戰鬥或逃跑反應為警告階段(stage of alarm reaction),動物設法脫離緊迫之環 境,同時活化 H-P-A 軸與自主神經以備不時之需。其次為動物對緊迫之抵抗階段
(stage of resistance),此時腎上腺皮質開始活動,分泌醣類皮質素(cortisol and corticosterone)促使後續生理反應,試圖對抗緊迫並回復持恆狀態。最後,當緊迫 超出動物所能負荷,生理功能開始失常、疲勞並產生病態反應,即為竭盡階段(stage of exhaustion)。在GAS 理論中,Selye 氏認為醣類皮質素是主要的傳訊因子,並率 先探討了緊迫與醣類皮質素的關聯,開啟了此領域一新扉頁,爾後更多科學家從醣 類皮質素的角度投入研究,擴展對緊迫之認識。
綜合Cannon 氏與 Selye 氏的理論,緊迫與 H-P-A 軸、醣類皮質素以及生物體 持恆機制有密切關聯。然而動物受到緊迫後的生理反應十分複雜,以內分泌反應為 例,除了H-P-A 軸,泌乳素(prolactin)、生長素、升糖素(glucagon)、胰島素、
血管加壓素(vasopressin)、褪黑激素(melatonin)、性內泌素(sex hormone)、甲 狀腺素以及某些神經傳導物質等皆會受到影響(Ranabir and Reetu, 2019),故僅以 H-P-A 軸、醣類皮質素與持恆之間的關聯來探討緊迫仍不夠全面。此外,動物對緊 迫的反應亦受到心理學層面影響,動物對一個事件的認知,會決定此事件是否成為 緊迫以及嚴重程度,這取決於動物的先前經驗(農場動物與工作人員相處之經驗將 決定動物是否將人的出現視為緊迫)與遺傳特徵如脾氣暴躁或溫馴等(Grandin and Shively, 2015)。
隨著研究成果累積,對緊迫的了解越發透徹,緊迫的定義與日俱增。其中與動
物福利最相關的說法係由Moberg 所提出,他定義緊迫為個體經過認知的威脅(緊 迫源,影響持恆)所引起的生物反應,當緊迫反應超出動物原先持恆機制的負荷,
需要額外改變生理機能來適應緊迫,動物便會經歷痛苦(distress),動物福利受到 損害(Moberg, 2000)。因此,如何確定緊迫造成動物之痛苦以及如何偵測緊迫之程 度,成為以緊迫生理角度探討動物福利時之重大挑戰。
2.2.2 動物之緊迫模式
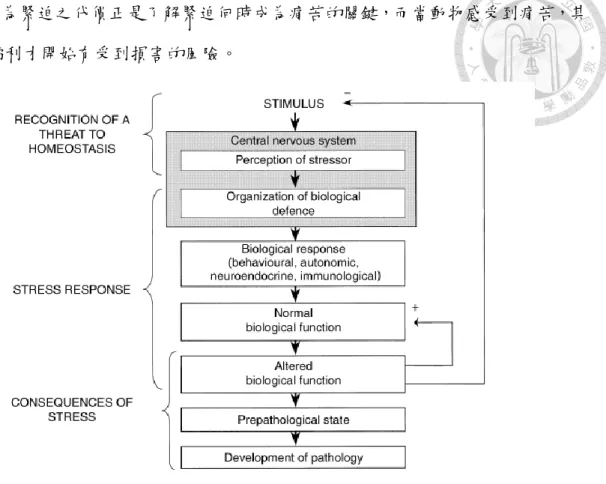
為解決上述的挑戰,Moberg 發展出一個框架模型,將動物緊迫相關知識整合 至統一且可經測試的觀念,協助科學家專注在緊迫與動物福利之間的關聯。此模型 如圖1 所示,將動物應對緊迫的過程分為三個階段:緊迫源認知、緊迫反應、緊迫 反應之後果,各階段再包含數個部分。首先,緊迫起源於中樞神經系統認知到一個 可能威脅生物體持恆的刺激,其中涉及到個體先前之經驗、基因、年齡以及當下心 理狀態。一旦中樞神經系統認知到某一刺激並視之為威脅,此刺激便成為緊迫,引 發後續之緊迫反應,包含行為、自主神經、神經內分泌以及免疫反應,單獨或共同 作用來協助生物體逃離、抵抗或適應緊迫源。最後,當緊迫反應打破生物體原先的 持恆,意味著生物體必須額外改變自身生理功能來應付緊迫源,並且面對緊迫反應 之後果,例如痛苦、預病理狀態甚至發展成疾病。
Moberg 認為不論動物使用何種手段來應對緊迫源,影響到動物福利的都是動 物應付緊迫源之後的結果,如果動物能以原先的持恆狀態應對緊迫源,其生理功能 維持正常,動物福利上不會受到影響;相反地,當緊迫源無法以原先的持恆狀態應 對,生物體會改變自身生理功能(緊迫的代價,Biological cost of stress)以應付緊 迫源,此時動物逐漸感到痛苦,若緊迫過於嚴峻或期間過長,動物極有可能發展成 預病理狀態甚至疾病,則動物福利儼然受到損害。
對動物福利而言,生物功能是否因為應對緊迫而改變比牽涉在緊迫反應中的 機制來得重要。Moberg 相信應該以緊迫源導致的生理功能改變作為動物福利的衡
量,因為緊迫之代價正是了解緊迫何時成為痛苦的關鍵,而當動物感受到痛苦,其 動物福利才開始有受到損害的風險。
圖1. 動物對緊迫之生理反應之模型框架
Fig 1. Model framework of stress response of animal (Moberg, 2000)
2.2.3 動物對緊迫之生理反應
緊迫源引發諸多生理反應,包含行為、自主神經、神經內分泌以及免疫反應,
協助生物體逃離、抵抗或適應緊迫源。行為反應是許多緊迫源最快速直接引起的,
動物可以藉由「戰鬥或逃跑」或「暫止」(freeze)(Bracha et al., 2004)將自己免於 處在緊迫源的環境下,若動物無法逃離緊迫源,則可能發展出刻板行為來協助適應。
自主神經系統反應,為動物體應對緊迫源的第二道防線,透過自主神經快速地引起 心血管、胃腸系統與腎上腺髓質的反應,促使動物體心律、血壓及腸胃道活動力改 變,即時地協助生物體。相較之下,神經內分泌系統擁有更長時間及更廣泛的作用 效果,例如 H-P-A 軸系統,可透過下視丘、腦垂腺與腎上腺之間的訊息傳遞,來
釋放醣類皮質素影響生物體能量代謝、生殖與免疫等系統。至於免疫反應,除了受 到醣類皮質素的抑制外,亦會因應特定緊迫源而產生變化,如紅白血球數的改變、
細胞激素(Cytokines)的分泌等。儘管這些緊迫反應十分重要,但以反應之嚴峻程 度來探討動物福利,則以神經內分泌的H-P-A 軸反應最為適合,因為 H-P-A 軸的 普遍性,多數緊迫源都可引起反應,且反應耗時較長,動物在反應過程中有可能經 歷痛苦(Distress),較具有討論動物福利的意義。
(1) 下視丘-腦垂腺-腎上腺皮質軸
H-P-A 軸由下視丘、腦垂腺前葉與腎上腺皮質所組成,藉由彼此間的內泌素傳 訊,共同調控醣類皮質素的分泌,並透過醣類皮質素的生理功能,參與代謝之持恆,
決定能量流向(energy flux)(Dallman et al., 1993)。一般認為,緊迫反應時 H-P-A 軸的致活,可協助生物體將能量用在應對緊迫源上。
i. H-P-A 軸之調節
生物體認知到威脅後,下視丘之腦室旁核(paraventricular nucleus, PVN)其中 的小細胞神經元軸突會自正中隆突(median eminenxce)分泌激腎上腺皮質素釋放 素(corticotrophin releasing hormone, CRH)與精胺酸加壓素(arginine vasopressin, AVP)進入門脈毛細管床,並作用在腦垂腺前葉的激腎上腺皮質素細胞(cortictrophs)
上,使其分泌激腎上腺皮質素(adrenocorticotropic hormone, ACTH)至血液中,最 終ACTH 刺激腎上腺皮質束狀帶合成及分泌醣類皮質素(Mormède et al., 2007)。
進入循環系統的醣類皮質素將運輸至各器官組織內發揮生理功能,作為緊迫反應 以應對緊迫源,同時參與對CRH 與 ACTH 的負回饋調控(negative feedback),作 用在下視丘與腦垂腺前葉,抑制 CRH、ACTH 的分泌,避免過度刺激 H-P-A 軸
(Manteuffel, 2002)。
H-P-A 軸線受到數種內泌素的調控,在下視丘,CRH 為主要且較強效的內泌素
(River and Plotsky, 1986),負責在急性緊迫反應中刺激 ACTH 分泌;而 AVP 可維 持 ACTH 的基礎分泌以及在長期緊迫下輔助刺激分泌 ACTH(Scott and Dinan, 1998)。腦垂腺亦受其他內泌素刺激而產生 ACTH,例如腎上腺素(Giguere and Labrie, 1983)以及催產素(oxytocin)(Link et al., 1993)。此外,在認知威脅時,
H-P-A 亦受到複雜的上游調控,腦室旁核可從下視丘其它核區、腦幹、下穹窿體
(subfornical organ)與邊緣系統(limbic system)接受生理或心理的訊號引發下游 腦垂腺與腎上腺的反應(Mormède et al., 2007),賦予 H-P-A 軸的高度敏感性與相 對非特異性,能針對多數緊迫源引起反應。
H-P-A 軸線調控的複雜度,說明此系統牽涉的生理反應眾多,在持恆作用上扮 演關鍵角色,其功能完整性之維持十分重要。在諸多能調控 H-P-A 軸功能的因素 中,晝夜節律(circadian rhythm)與緊迫源是重要內源性及外源性因素。
ii. H-P-A 軸與日夜節律之關聯
生物有許多生理現象具有節律性,會隨著時間呈週期變化。若節律之週期接近 二十四小時,即稱為日夜節律(diurnal rhythm),具有此特性的生理反應除了 H-P- A 軸,還有新陳代謝、體溫、採食飲水、睡眠、活動力與其他內泌素分泌等(Ingram and Dauncey, 1985)。節律性被生物體內的生理時鐘(endogenous oscillators)調控
(Aschoff, 1963;1966)。生理時鐘能感應呈週期性變化的環境因子(zeitgeber),並 依此調整生理反應之節律,協調身體各項功能共同或獨立運作以適應環境週期。
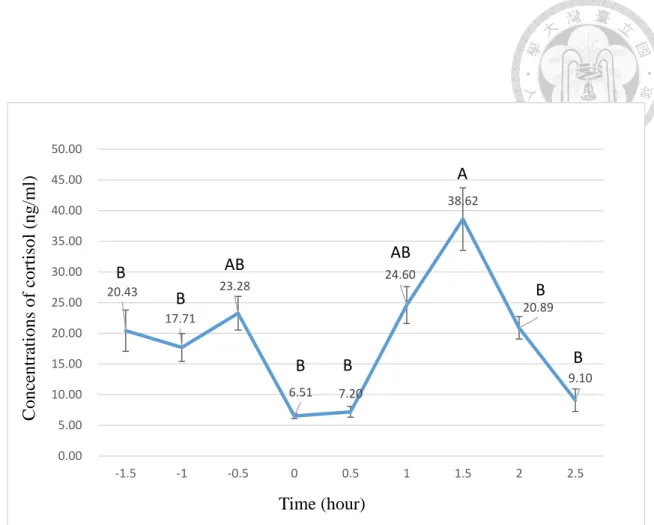
一般而言,光亮/黑暗循環是影響 H-P-A 軸日夜節律的重要環境因子,光的訊號 可透過傳入下視丘前葉的視交叉上核(suprachiasmatic nuclei, SCN)進行調節。日 行性動物如人、猴、牛、馬以及綿羊,其 ACTH 與醣類皮質素在清晨或睡醒前會 達到高峰;相反地,夜行性動物如齧齒類,則在傍晚達到高峰。本次試驗對象豬隻 屬日行性動物,故醣類皮質素的分泌高峰在上午(Koopmans et al., 2005)。以限飼
圈養豬隻為例,血漿中皮質類固醇濃度在下午為15 ng/ml,而在上午可達 45 ng/ml 的高峰值(Barnett et al., 1981);然而,有研究指出 H-P-A 軸反應的高峰出現在下 午,且受到豬隻採食行為的影響(Hillmann et al., 2008),顯示出 H-P-A 軸的日夜 節律性除了受光照外,亦受其他因素調控,例如年齡、性別與緊迫等(Ruis et al., 1997; Boumans et al., 2017)。
H-P-A 軸線之日夜節律性對於緊迫反應的檢測十分重要,將動物一天各時間醣 類皮質素的基礎值納入考量,方能做出有效的緊迫測量,準確地解讀動物的動物福 利狀態。
iii. 醣類皮質素之特性
醣類皮質素種類會因動物而異,在綿羊、豬、貂、狐狸、犬、貓與魚中主要為 皮質醇(Cortisol);在鳥類與齧齒類中主要為皮質酮(Corticosterone);在反芻動物,
兩者分泌量相近(白等, 1996)。以本次試驗對象豬隻所分泌的皮質醇為例,說明其 合成、運輸、代謝相關機制以及生理功能。
A. 皮質醇之合成與化學特性
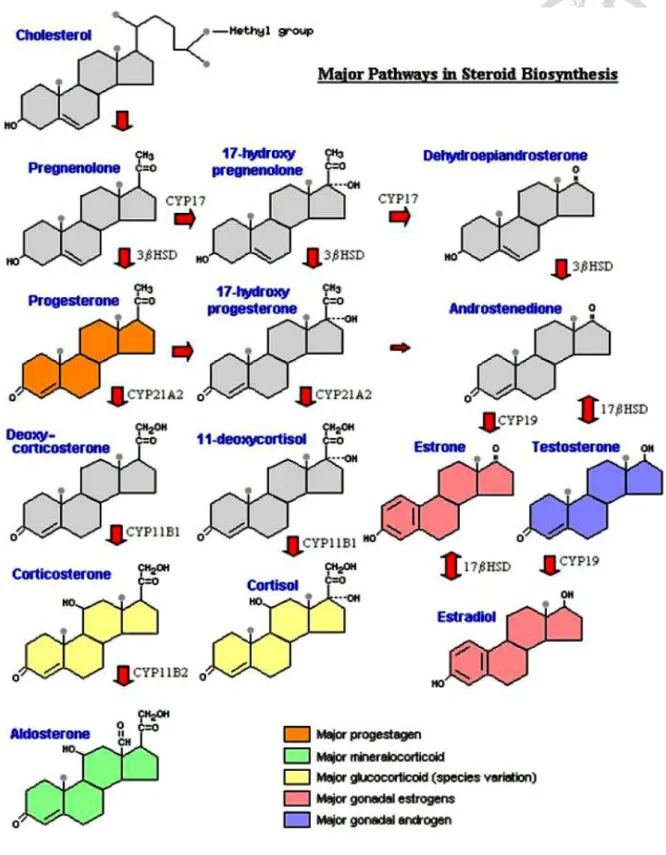
膽固醇(cholesterol)為皮質醇的前驅物,可透過腸胃吸收或體內醋酸鹽(acetate)
合成。膽固醇在腎上腺皮質的束狀帶細胞內的平滑內質網與粒線體中逐步合成為 皮質醇如圖 2 所示,主要參與的酵素為膽固醇側鏈裂解酶(Cholesterol side-chain cleavage enzyme)、17α-羥化酶(17α-hydroxylase)、3-β 羥化類固醇脫氫酶(3-β Hydroxysteroid dehydrogenases)、21α-羥化酶(21α-hydroxylase)與 11β-羥化酶(11β- hydroxylase)。皮質醇由四個碳環組成,為脂溶性分子,能自由擴散血腦屏障與細 胞膜,發揮生理作用。
圖2. 類固醇內泌素之生成路徑。
Fig 2. Pathway for the biosynthesis of steroid hormones.
(Bowen, 2001)
B. 皮質醇之運輸與代謝
由於皮質醇為脂溶性非極性小分子,在血液中需要與大分子之蛋白質結合,避 免被肝臟、腎臟或血中酵素分解,故在循環系統中將近 90%的皮質醇與運送蛋白 結合成結合態,未與蛋白質結合的皮質醇即為游離態(free forms),具有生理活性,
能被吸收進細胞內引發反應。運送蛋白為對皮質醇具有親和力的蛋白分子,包含白 蛋白(albumin)與皮質類固醇轉運蛋白(corticosteroid-binding globulin, CBG)。兩 者對於皮質醇的親和力不一,白蛋白親和力較低但血中含量高,能與大量皮質醇結 合;相反地,皮質類固醇轉運蛋白親和力較高,總結合量卻較低(白等, 1996)。功能 上,皮質類固醇轉運蛋白負責皮質醇基礎量的結合,而白蛋白在皮質醇分泌量增加 時負責與增多的皮質醇結合,兩者濃度的高低,可共同調控皮質醇的生物可利用性
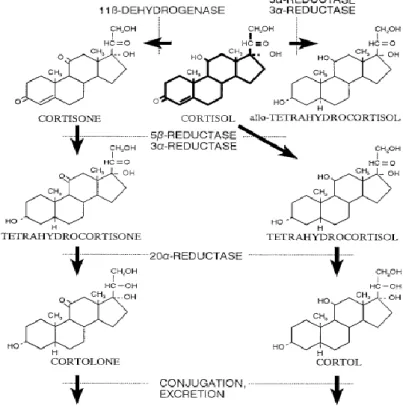
(Gayrard et al., 1996)。進入循環系統後,皮質醇作用於標的器官並且隨即被代謝 排出體外,其半衰期因物種而異,一般不超過兩個小時。皮質醇代謝分解之示意圖 如圖3 所示,主要發生在肝臟,部分在腎臟與其他器官進行。除了使皮質醇失去生 理活性,分解異化作用目的在增加類固醇分子的親水性,將葡萄醣醛酸(glucuronic acid)或硫酸鹽連接至皮質醇,使之成為親水的化合物,隨後,親水皮質醇代謝物 可藉肝臟膽汁的分泌進入消化道中,由糞便(25%)排出,亦可由腎臟從血液過濾 至尿液,由尿液(75%)排出體外(Honour, 1984)。而皮質醇在肝臟與腎臟進行代 謝時參與的酶和反應路徑如圖 4 所示。此外,皮質醇以及相關代謝產物也存在乳 汁、唾液與毛髮中。
C. 皮質醇之作用機制與生理功能
皮質醇透過簡單擴散進入標靶細胞後,於細胞質內與特定受體結合成複合體,
此複合體再經過其他條件的活化,如溫度、時間、離子等,進入細胞核影響轉錄,
活化或抑制基因組上的某些片段,隨後新蛋白質形成,發揮生理機能,最後皮質醇
圖3. 腎上腺皮質醇代謝分解示意圖
Fig 3. Schematic diagram of the metabolism of adrenal cortisol (Möstl and Palme, 2002)
圖4. 腎上腺皮質醇代謝分解路徑圖
Fig 4. Pathway for the metabolism of adrenal cortisol (White et al., 1997)
代謝後喪失功能。由於大多數器官組織都具備皮質醇受體,能接受其訊號,故皮 質醇的生理功能十分廣泛,影響諸多生理功能。其主要功能分項說明如下:
a. 對能量代謝之影響
皮質醇能引起周圍組織(peripheral tissues)中蛋白質與脂質的異化作用
(catabolism)分解出碳骨架,並抑制周圍組織的蛋白質同化作用、降低細胞對 胺基酸的吸收;在肝臟,皮質醇會促使肝細胞碳骨架合成為葡萄糖,增加葡萄 糖新生成作用(gluconeogenesis),同時增進肝醣與血漿蛋白質的合成(McMahon et al., 1988)。另外,皮質醇能抑制細胞對葡萄糖的利用,進而增加血糖與胰島 素分泌,且皮質醇亦能作用在下視丘造成食慾、採食增加。藉由中央與周圍機 制的合作,皮質醇增加生物體內的能量可利用性(Tempel et al., 1994)。皮質醇 對能量代謝調控,在應對緊迫源時,可使動物有充足的血糖水平以供使用(通 常藉由行為反應)。
過量皮質醇作用會促使肌肉組織過度分解,導致肌肉損失及衰弱,若皮 質醇引發的能量流動沒有被生物體以行為方式立即應對緊迫,能量會重新以 脂肪形式儲存,淨效應為生物體將蛋白質轉成脂肪(Devenport et al., 1989)。
脂肪組織的重新分布與累積,即為醣類皮質素過度分泌導致的病徵—月亮 臉、水牛肩,此時動物福利即有受到損害的疑慮。
b. 對水分及電解質代謝之影響
皮質醇與鹽類皮質素功能上有部分重疊效果,原因為細胞核內的鹽類皮質 素受體能接受皮質醇的訊號引發反應(Pascual-Le et al., 2005)。雖然皮質醇對 水分與電解質之影響遠小於鹽類皮質素,但仍有回收鈉離子、排出鉀離子之功 能,甚至過量的皮質醇也會引起水腫、高血鈉、低血鉀、多尿與頻渴等症狀。
c. 對抗發炎與免疫系統之影響
發炎與免疫反應可視為動物體應對外來感染的緊迫反應,皮質醇對於 兩者的抑制,作用在於防止過度的緊迫反應威脅到生物體之持恆(Munck et al., 1984)。對於發炎反應,皮質醇可抑制初級發炎反應如纖維蛋白沉積、微血管擴 張和白血球移動與吞噬,以及後期發炎反應如纖維細胞增生與膠原細胞沉積。
對於免疫反應,皮質醇能減緩磷脂酶A2(phospholipase A2)的釋出,抑制由磷 脂酶 A2引起的下游反應—釋出花生四烯酸(arachidonic acid)。花生四烯酸為 前列腺素(Prostaglandin)、白三烯(Leukotriene)以及免疫相關內分泌物的前 驅物,藉由抑制花生四烯酸的分泌,皮質醇可壓抑免疫反應。
過度的皮質醇分泌會導致免疫反應被過度壓抑,此時生物體對於外來病 原失去可以抵禦的機制,容易患病而影響動物福利。
d. 對內分泌系統之影響
過多的皮質醇可透過負回饋抑制下視丘與腦垂腺分泌 CRH 與 ACTH,促 使皮質醇分泌回到基礎量。若長期過量的皮質醇分泌,會對下視丘、腦垂腺造 成嚴重的抑制,導致腎上腺皮質萎縮、腎上腺功能降低。其他內泌素系統亦會 受到皮質醇影響,例如性腺系統。在皮質醇過量下,下視丘的促性腺激素釋放 激素(gonadotropin-Releasing Hormone, GnRH)被抑制,其下游腦垂腺促濾泡 激素與黃體素的分泌以及性腺功能皆被影響,生殖生理功能受到干擾。在嚴峻 的緊迫下,動物的內分泌系統紊亂,失去原先的生理功能,動物福利受到損害。
(2) 持恆與變穩態
持恆為 Cannon 所提出的概念,是生物體內在的穩定能力,意即生物體有能力 藉由生理系統的回饋圈(feedback circle),將各項生理反應或跡象穩定在固定範圍
(適合生物體存活的範圍)內,諸如體溫、血糖、水分與電解質等等,其過程無需
付出任何代價。當緊迫源威脅到生物體原先持恆,擾亂生理反應或跡象,生物體會 藉由許多機制作用(即緊迫反應),嘗試應對緊迫源、重新恢復持恆。如果緊迫源 僅輕微干擾,則生物體無需花費太多代價便能恢復;反之,當生物體遭遇到強烈或 長期緊迫,超過生物體所能負荷,其持恆之恢復則需要付出相當的代價,甚至會改 變原先持恆所固定的生理範圍。
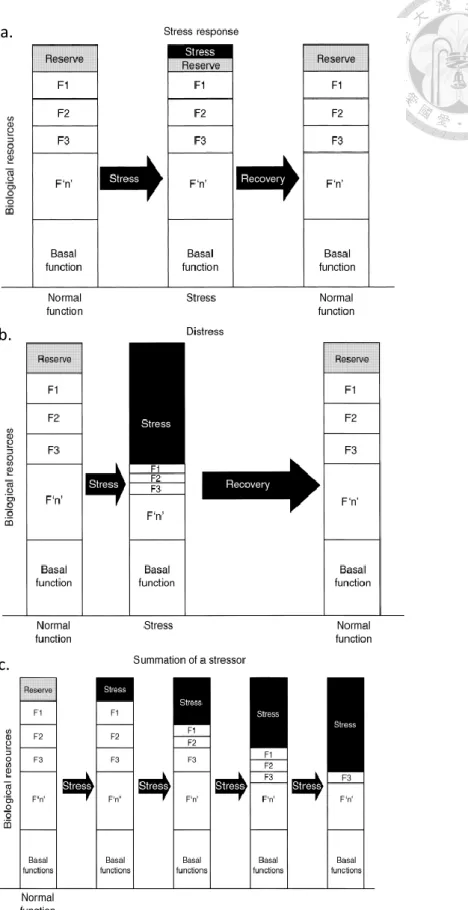
為了解釋持恆範圍的變化,McEwen(1998)提出了變穩態(allostasis)的觀 念。Allostasis 是指生物體經由改變持恆範圍來達到並維持穩定的能力。當生物體 遭遇到無法逃離或克服的緊迫源,僅能設法適應,過程中生物體整合各項資訊,調 整生理系統回饋圈,改變原先持恆,將生物體準備成能適應緊迫源的狀態,並且維 持在那個狀態直到緊迫源消失。藉由 Allostasis,生物體得以適應緊迫源並在正常 生理範圍外獲得一新持恆,穩定下來與緊迫源共存。當然,改變正常生理範圍的適 應機制必有其代價,長期的Allostasis 反應所要求生物體支付的代價稱為變穩態負 擔(Allostasis loads),通常伴隨著慢性疾病的發生。
用Allostasis 的觀念詮釋動物福利狀態,可以較準確地區分動物之正常生理狀 態與適應緊迫源時之變穩態,藉此判斷動物何時感受到痛苦(Distress),當Allostasis loads 出現在動物時,無疑是動物福利的損害。故近年來,Allostasis 已成為評估動 物福利的重要觀念(Korte et al., 2007)。圖5 可以說明動物如何應對緊迫源(Moberg, 2000),功能正常運作下,生物體除了基礎功能外還具備若干生理功能(F1,F2,F3 至F’n’),並有餘裕(reserve)能維持正套生理系統的持恆,若動物無須改變原先持 恆狀態便可應對緊迫源如圖5.a 所示,即動物應對緊迫源時仍有餘裕維持恆定,生 理功能與動物福利皆不受負面影響;反之,當出現強烈或長期的緊迫源如圖 5.b,c 所示,僅憑動物原先餘裕不足以應對,得調整生理系統的持恆範圍方可應付,其他 生理功能受到壓抑,此時動物開始經歷痛苦,動物福利開始受損。
若以H-P-A 軸說明 Allostasis 時可能發生的生理現象,當外在威脅引起激烈的 H-P-A 軸反應,釋放過量皮質醇進入循環系統,在腦部原本能改善認知記憶的皮質
圖5. 動物應對緊迫源之假想進程
Fig 5. Hypothetical process of how animal respond to stressors
(Moberg, 2000)
a.
b.
c.
醇長期分泌下會導致神經源萎縮、記憶衰退及恐懼感增加(Roozendaal, 2000)。對 於免疫系統,適量的皮質醇可以避免免疫反應過度打破持恆,而過量皮質醇會壓抑 免疫系統導致疾病發生(Dhabhar and McEwen, 1999)。能量代謝方面,適量皮質醇 能增加能量流以供給動物利用,然而過量的醣類皮質素分泌,導致能量流無法全部 被動物使用,多餘的能量將以脂肪儲存,導致肥胖甚至胰島素阻抗或心血管的疾病
(Brindley and Rolland, 1989)。綜上所述,H-P-A 軸影響生理甚鉅,若反應超出持 恆的範圍進入Allostasis 就有可能造成動物的不適與痛苦,進而對動物福利造成負 面的影響。
2.2.4 緊迫源與緊迫指標
以動物福利觀點定義,能挑戰動物五大自由的內外在因子,便為緊迫源,在家 畜禽的飼養過程中,緊迫源通常為不安適的環境因素或人為操作。緊迫源可從質與 量去區分,質的部分包括物理性(冷熱、機械性傷害、電擊、噪音)、化學性(麻 醉、毒素、藥物)、營養性(食物或水的缺乏)與心理性(隔離、併欄、保定、動 物社會位階更動)的緊迫源(Martínez-Miró et al., 2016),甚至有結合多個緊迫源 的威脅如運輸(Roldan-Santiago et al., 2013)與分娩(Fell and Shutt, 1986)等。量 的部分則依照緊迫源的強度(分貝、安培或將緊迫源量化的單位)與持續時間(急 性、慢性與亞急性)作區別。緊迫源的量化能協助科學家了解緊迫源的性質,然而 卻無法反映動物的真實感受與生理狀態。動物體內緊迫反應的程度,是否打破持恆 造成痛苦,才是動物福利學關心的事物,因此需要緊迫指標(Stress indicator),來 反映動物在應對緊迫源之過程中的真實體驗與生理狀態。
緊迫指標是可量化及可重複測量的生理反應、跡象或表現,用以反映緊迫源所 引起反應之程度,並藉此評估動物之生理與動物福利狀態。各種不同的緊迫源,能 引起各式各樣的緊迫反應,故作為緊迫程度的指標很多。分類上,緊迫指標有普遍 性與特異性的區別,普遍性之緊迫指標表示多數緊迫源能引起某一生理跡象的變
化,反映出生物整體的狀態,例腎上腺皮質醇的分泌量、體溫、心律與呼吸速率等。
而特異性的緊迫指標只受特定緊迫源刺激,僅能顯示某項生物功能或跡象的狀態 以及程度,例如血中游離脂肪酸、酮體、葡萄糖與尿素等濃度可以反映動物體飢餓 程度,滲透壓、總蛋白、白蛋白與血球容積(Packed-cell volume)等數值可以反映 脫水狀態,而乳酸與肌酸激酶(creatine kinase)的濃度可以凸顯動物活動的劇烈與 肌肉受損之程度,血管加壓素(vasopressin)的多寡則可以顯示動物運輸過程中暈 眩的程度(Knowles and Warriss, 2000)。
緊迫指標的選用需配合緊迫源之性質與其可能引發的緊迫反應,本次試驗為 調查現行運輸下豬隻的動物福利,運輸過程即為豬隻所遭遇的緊迫源,常見適用於 分析運輸的緊迫指標如下:
(1) 死亡率與屠體品質
死亡率是非常直接的評估標準,若動物無法應對所遭遇的緊迫,其生理機制無 法維持持恆,最糟糕的結果就是無法存活。在動物運輸的動物福利研究中,動物的 死亡率可以對運輸過程的緊迫程度,提供非常直觀的證據。一趟運輸過程中,死亡 率越高表示動物所面對的環境越加嚴峻,其動物福利越有可能受到挑戰(Knowles and Warriss, 2000)。然而,死亡率作為緊迫指標有其缺點。以動物運輸研究為例,
運輸中較貧弱的動物才會有死亡的可能,整體的死亡率相當低,大約在 1%左右,
意味著需要大量足夠的動物數量才能準確地估計預探討的變因之影響。
屠體品質作為緊迫指標,以動物屠宰後的損傷狀態來評估動物生前的情況。以 動物運輸研究為例,常見的屠體損傷可分為兩類:物理機械性損傷與肌肉代謝異常。
前者如皮膚損傷、肌肉瘀傷、肌肉撕裂出血以及骨骼斷裂,來自車體上或屠宰廠內 的擠壓或打鬥、駕乘等行為。後者為水漾肉(PSE)與暗乾肉(DFD),為運輸過程 中肌肉過度使用,肌肉中的肝醣大量消耗,屠後肌肉中酸鹼值失衡所導致。兩者的
成因都跟動物遭遇的緊迫有關,且過程中動物會感受到疼痛與不適。因此,屠體的 狀態確實地可以反應動物生前的動物福利狀態。
(2) 動物行為
動物行為科學可為動物的感受提供證據,各種動物都具備一定的行為反應準則,
趨向喜歡的環境、逃避討厭的環境,因此,動物對各種不同刺激的反應與選擇,可 以作為評估動物福利最直觀的指標(李,2015)。此外,動物行為的改變是重要的 適應機制,亦是動物經歷痛苦的最早徵象(Ayo et al., 2002)。
運輸過程中豬隻的行為變化,以站姿、臥姿與坐姿的轉換以及侵略性行為(騎 乘、頂咬)為主。藉由站姿、臥姿與坐姿的比例,可以推論動物是否疲勞、暈眩或 緊張。例如短期運輸(低於3 小時)的豬隻以站姿為主(Bradshaw et al., 1996),
原因為豬隻尚未習慣運輸環境,但若運輸環境較佳,豬隻有可能適應環境,並以躺 姿為主(Lambooij and Engel, 1991);然而動物表現躺姿亦有負面意涵之可能,
Knowles et al.(1999)發現長期運輸(高於 24 小時)的牛隻多以躺姿為主,且血 漿中皮質醇比站姿的牛隻高,表示牛隻不勝負荷長期的運輸,而無力站立。當豬隻 進行社會位階重新排位(social regrouping)或競爭車體上資源(充足可休息的空間)
時,容易發生侵略性行為,導致為疼痛、皮膚損傷、屠體瘀傷、肌肉過度運動與肝 醣耗盡以及皮質醇水平上升等,對動物福利造成傷害。
(3) 腎上腺皮質醇
根據 Cannon 與 Style 氏的理論,H-P-A 軸活動為非特異性反應能接受多數緊迫 源之刺激而引發,因此在測量緊迫時,用直接或間接的方法評估腎上腺之分泌功能,
為常見的緊迫測量方式(Stott, 1981)。縱使現已證實並非所有緊迫源皆會刺激 H- P-A 軸反應,但評估腎上腺分泌的能力,也就是腎上腺皮質醇的含量多寡,仍舊為 評估緊迫時的主流方法。除了腎上腺皮質醇外,亦可藉由腎上腺大小、血液與腦垂
腺中ACTH 濃度與下視丘中 CRH 濃度來作為判斷 H-P-A 軸活動的依據(Vernikos- Danellis and Heybach, 1980)。對於長期的緊迫反應,可進行反應性(responsiveness)
的評估,如施加外源性 ACTH 或 CRH 刺激 H-P-A 軸線,檢查各器官分泌的能力
(Gaillard and AI-Damluji, 1987)。
使用腎上腺皮質醇做為緊迫指標有其限制,除了日夜節律的變化需要考量,皮 質醇的分泌為脈衝性(pulsatile),以約九十分鐘為周期。此性質出現在某些物種上,
例如人(Follenius et al., 1987)、牛(Thun et al., 1981)與羊(Fulkerson et al., 1979)。 另外,腎上腺功能的高變異性已在實驗室動物(Popova et al., 1981;Sarrieau et al., 1998)與經濟動物(Garcia-Belenguer et al., 1996;Desautes et al., 1997)上被充分 描述,此個體差異之存在乃受到基因背景調控。在豬隻,皮質類固醇運轉蛋白(CBG)
的基因多態性會影響循環的皮質醇水平(Desautes et al., 2002)。腎上腺分泌功能亦 具有極限,隨著緊迫源的強度上升或持續,皮質醇分泌量會達高原期而無法反映緊 迫源之程度,有研究指出豬隻在僅暴露於新環境的刺激下,其皮質醇分泌量已達高 峰(Choi et al., 2012)。最後,緊迫源的持續時間與檢驗腎上腺功能的時間點為重 要解讀 H-P-A 軸活動力的因素。H-P-A 軸運作需要各器官的整合,故皮質醇在循 環系統中的增加為一緩慢過程,面對緊迫源時需要數分鐘的時間血中皮質醇方會 上升,緊迫源消失後約一小時皮質醇含量才會回到基底水平(Mormède et al., 2007)。 若緊迫源維持或重複一段時間,則有可能加速動物適應,而導致腎上腺對緊迫源不 產生反應。研究指出,豬隻在經歷連續一個月的不定期電擊刺激後,對於刺激仍有 行為反應,但其腎上腺皮質醇濃度卻與未處理組相同(Jensen et al., 1996)。
綜上所述,腎上腺皮質醇做為非特異性緊迫反應之緊迫指標,可以反映動物生 理狀態、判讀動物福利。然而,將腎上腺皮質醇分泌方式與規律、個體間之高度差 異性、分泌極限量與緊迫源之持續時間納入考量,方能作出精確之判斷。
2.2.5 腎上腺皮質醇之評估
欲瞭解動物腎上腺分泌功能,需檢測動物檢體中的內泌素含量,而常見的腎上 腺皮質醇評估方法,從競爭性蛋白質分析法(Competitive protein binding analysis, CPB)、放射免疫分析法(Radioimmunoassay, RIA)逐漸發展到酵素免疫分析法
(Enzyme immunoassay, EIA ), 或 稱 酵 素 連 結 免 疫 吸 附 法 ( Enzyme-linked immunosorbent assay, ELISA)。ELISA 的原理為將酵素標幟在抗原上,經由免疫反 應,抗原與抗體結合後,加入基質(substrate)產生酵素呈色反應,並利用呈色之 吸光值對檢體中抗原濃度加以定量。酵素連結免疫吸附法有許多優點,如成本較便 宜、步驟與操作簡單、無放射性之廢料與汙染、容易進行判別或定量(Blake and Gould, 1984)。近年來免疫與生化技術進步,改善ELISA 的靈敏度,已逐漸取代其 他測量方法,並廣泛應用於生物醫學、環境科學等產業(Monroe, 1984)。ELISA 有 許多種類,本實驗室使用競爭型酵素免疫分析法(competitive EIA)。原理為利用酵 素標幟抗原(抗體與過氧化酶之聯結體),與檢體內標的物競爭抗體之結合位,並 與基質產生呈色反應,透過標準溶液內不同濃度之抗原所建立的標準曲線,經由吸 光值換算出動物檢體內未知標的物的濃度。
從動物取得檢體的方式又可分為侵入式(invasive)與非侵入式(non-invasive)。 前者會使用器具侵入動物身體,如以針具取得血液、腦脊隨液、部分器官組織等,
過程中人為操作、疼痛甚至麻醉都是額外緊迫源,對動物福利造成傷害,同時影響 皮質醇之分泌,使評估的數值有高估之疑慮;而以非侵入式獲取檢體較不會造成動 物不適,能避免皮質醇分泌的變動,減少採集過程中動物福利的損害。常見的非侵 入式檢體為唾液、尿液、糞樣、乳樣與毛髮等,甚至動物行為之變化亦可提供充足 資訊以解讀其動物福利。本節將說明以各檢體為來源,評估腎上腺皮質醇含量與動 物生理狀態的優缺點。
(1) 血漿
在動物福利研究中,血漿是分析腎上腺皮質醇時最常被使用的檢體,可以檢測 動物總皮質醇濃度。血漿作為檢體,其優點為可以即時地測出最直觀且準確的皮質 醇濃度,同時亦可檢測激腎上腺皮質素(ACTH)、兒茶酚胺(catecholamines)、血 糖和白血球數量與型態等與H-P-A 軸功能致活相關的指標(Mormède et al., 2007)。 而缺點為血漿是侵入性的檢體,取得的過程會造成額外的緊迫,不但造成動物福利 的損害,而影響了皮質醇的評估與判讀。
(2) 糞便與尿液
尿液是代謝醣類皮質醇的主要路徑之一,血液中皮質醇經肝臟、腎臟代謝之產 物,經由腎臟過濾而形成尿液排出體外,可作為非侵入式的檢體,採集過程中不會 額外造成動物緊迫。另外,尿液作為排泄產物,會累積數小時才會排出,因此尿液 皮質醇與其代謝產物之濃度可以提供一段時間內 H-P-A 軸活動力的綜合評估,亦 可校正血漿皮質醇濃度之波動。同理,血漿中皮質醇經由肝臟代謝,同膽汁一起進 入消化道,最後以糞便排出體外,使得糞便中皮質醇之檢測亦成為非侵入式之單位 時間內的腎上腺功能綜合評估方式。
(3) 唾液
使用唾液作為檢體提供了非侵入且無壓力的採集方式,因此近年來廣受歡迎。
唾液主要由三對唾腺(salivary gland)產生,形成於唾腺內特化的分泌性末梢—腺 泡或分泌小管。腺泡藉由鈉離子幫浦耗能運輸將血液中鈉離子移往分泌小管內引 起滲透壓失衡,促使液體從血液移至管腔成為唾液(Vining and McGinley, 1987)。
由於腺泡細胞的緊密連結(tight junctions),血液中許多有機物無法自由過濾至唾 液中,例如尿素、葡萄糖、脂質、大分子蛋白等,這些物質在唾液中的濃度遠低於 血液。而游離態的固醇類內泌素具有脂溶性質,能快速地以被動擴散方式進入腺泡
細胞(Riad-Fahmy et al., 1982),這使得唾液中的皮質醇以游離態為主,約為血液 中皮質醇濃度的5-10%左右,因此需要較靈敏的分析系統。游離態皮質醇被動擴散 的速率非常快,足以維持血漿與唾液中的皮質醇濃度平衡,因此唾液中皮質醇濃度 不受到唾液流速的影響(Vining and McGinley, 1984)。當緊迫源引發H-P-A 軸反應 促使血中皮質醇上升時,其快速的被動擴散使唾液能及時地反映動物生理狀態
(Bushong et al., 2000)。綜上所述,以唾液中腎上腺皮質醇做為緊迫指標監測動物 之生理狀況,有其實用性與潛力,得在不侵擾動物下,提供立即性之資訊以供解讀 其生理狀態與動物福利狀態。
然而以唾液做為檢體檢測皮質醇濃度有其限制,唾液中以游離態存在的皮質 醇濃度僅有血液中皮質醇濃度的5-10%。除了需要靈敏的分析系統之外,唾液中較 低的皮質醇含量,會導致當唾液中部分蛋白質、微生物、食物殘渣、齦溝液(gingival crevicular fluid)或者外界汙染時,唾液皮質醇濃度會受到嚴重干擾,造成評估誤 差。在採集人類的唾液之前,通常會注意被採集者的口腔清潔、是否留有食物殘渣 或是否有出血之情形;在動物的唾液採集,則避免在動物進食後採集,且採集前數 週便開始讓動物習慣被唾液採集之過程,避免造成額外刺激。然而,在本研究中,
以調查台灣豬隻運輸現況為主,這些事前須注意之細節不易周全。
2.3 運輸過程中緊迫源與其對動物福利之影響
動物運輸是現今畜牧業重要的組成部分(Speer, Slack, & Troyer, 2001),且已 被證實會對動物福利造成影響(Grandin, 2000),動物在運輸過程中會遭遇受傷、
疲勞、疾病甚至死亡之威脅,其屠體品質亦有下降之可能(Schwartzkopf-Genswein et al., 2012)。運輸過程中的各項條件如運輸時間(duration)、豬隻承載密度(Loading density)、工作人員操作手法(handling)與車輛微環境(trailer microclimate)等,
將影響運輸緊迫之劇烈程度以及動物於運輸時的動物福利,本節即為說明各項條 件之變動與運輸過程中動物福利之關聯。
2.4.1 運輸時間
動物運輸既已被認定為緊迫源,其時間長短便是量化此緊迫源之首要條件,運 輸距離同理。一般認為運輸時間或距離越長,對動物造成的緊迫越大,如Voslarova et al.(2017)將運輸距離分為五個段落(< 50 Km, 50-100 Km, 101-200 Km, 201-300 Km, >300 Km),發現豬隻死亡率隨運輸距離增長而上升,在運輸時間上也有相似 的研究結果(Warriss, 1993)。然而,亦有研究顯示出相反的結果,Haley et al.(2008)
以普查方式研究豬隻運輸條件與死亡率的關聯,發現當運輸距離增加50 Km 時,
豬隻死亡率會下降。在血液生化值上,運輸3 小時豬隻血中的乳酸(lactate)與皮 質醇含量比運輸 15 分鐘來得低,而且屠體品質較無水樣肉的可能(Perez et al., 2002)。
這樣相反的結果顯示出時間或距離對豬隻的影響可能為非線性的關係,
Sutherland et al.(2009)發現豬隻死亡率在運輸時間 30 分鐘至 4 小時間上升,但 在運輸時間超過4 小時後有下降的情形。動物行為上亦發現,比起長期運輸,動物 經歷短期運輸後(短於30 分鐘),有較難以驅趕的情況,且較可能有水樣肉之發生
(Grandin, 1994),顯示出動物尚未適應運輸環境,仍處於較躁動的狀態。運輸環 境之優劣也需要被考量,如果整趟旅程平順、流暢,長時間的運輸可提供豬隻適應 與休息之可能;反之,當旅程粗糙且充滿干擾,即使是短期的運輸也會對豬隻造成 很大的傷害。
動物運輸前的操作亦會影響動物運輸的品質。運輸前的餵飼會使豬隻容易在 運輸過程中感到不適、暈眩,通常需要禁食(Bradshaw et al., 1996),而在熱季時 運輸前對車體上豬隻進行灑水,也能有效降低其熱緊迫,改善運輸環境。由此可見,
在探討動物運輸時間或距離時,許多運輸條件的異同,會造成試驗結果的分歧,應 該將諸多的條件均納入考量。
2.4.2 豬隻承載密度
豬隻承載密度對動物福利與經濟收益有重要之影響,密度過低會造成豬隻在 移動車體上容易失去平衡而摔倒,密度過高則會造成豬隻彼此擠壓,兩者都會讓豬 隻發生皮膚損傷、肌肉瘀傷、受傷(Eldridge and Winfield, 1988),嚴重者在抵達目 的地時無法行走(non-ambulatory, non-injured, NANI),甚至死亡(death on arrival, DOA),這無疑是對動物福利與經濟收益的傷害。另外,承載密度也決定了一趟車 次可容納多頭數豬隻,對於牧場而言是重要的成本考量。
關於合適的豬隻承載密度,Ritter et al.(2006)在北美地區的研究發現當承載 密度從0.5 m2/100 kg(200 Kg / m2)提高至0.4 m2/100 kg(250 Kg / m2)時,豬隻 死亡率會從0.08%上升至 0.27%,並認為密度是 0.48 m2/100 kg(208.3 Kg / m2)時,
最能減少運輸時的損失。在加拿大亦有同樣的研究結果,單位面積上的豬隻頭數越 多,死亡率越高(Whiting and Brandt, 2002)。而近年來的研究更指明承載密度與運 輸時間之間具有交互作用,當承載密度較低且短程運輸時,豬隻容易發生水樣肉
(Guàrdia et al., 2004),有研究者甚至建議在短程運輸時使用高密度的承載;歐盟
(EU)對密度的規範(235.3 Kg / m2, CEC, 2005)亦被建議應適用在超過三小時的 運輸。承載密度對動物的影響,除了自身變化外,亦需考慮與時間、溫度甚至是操 作強度的交互關係,台灣目前法規對豬隻承載密度的規範為每平方公尺不得超過 315 公斤,實際上對動物的影響仍不清楚。
2.4.3 工作人員操作手法
運輸過程中,工作人員操作主要集中在驅趕上下車的階段 (loading and unloading),且會帶給動物相當大的緊迫。有許多變因影響驅趕過程是否緊迫,工 作人員接觸動物所引起的恐懼(Hemsworth et al., 1997)、過程中器具的使用、畜舍 的設計與動線規劃。由此可見,動物在牧場端的經歷存有很大的差異,根據Dewey
et al.(2004)的調查,各趟運輸過程中豬隻死亡率的變異有 55%的部分可由牧場端
的差異解釋。
驅趕耗時是一個重要的變因,當工作人員嘗試加快速度,儘管動物維持同樣的 移動方向,仍有錯置腳步、滑倒、摔傷與瘀傷的可能(Atkinson, 2000)。當牧場動 線規劃不佳(過陡的上升坡度、走道充滿障礙物、走道寬度過窄或長度過長、走道 具有大角度彎道等)及工作人員不了解豬隻移動習性(習慣以小群體移動、會緩慢 探索未知環境、會反抗推擠的力量而非順從、驚嚇時容易集聚而造成驅趕困難等), 此時若迫使豬隻快速移動會造成很嚴峻的緊迫。
驅趕器具也很重要,擋豬板是主要使用的器具,來驅趕大部分的豬隻,輔以 電擊棒來對付較頑強的豬隻。社會大眾普遍地認為電棒的使用會對動物福利造成 損害,然而,在較高豬隻承載密度下,使用電棒能快速迫使豬隻移動到車體上,
進而減少豬隻屠體瘀傷的情形(Guise and Penny, 1989a)。的確電擊棒會造成豬隻 疼痛,不該使用在豬隻敏感的部位上,如眼、鼻、耳與陰部,台灣目前法令規範 不得不當電擊便是此意。
綜上所述,操作人員的訓練、豬場硬體設施的優劣會決定驅趕的難度,若有 良好的教育訓練及牧場規劃,並且適當地使用驅趕工具,可在驅趕過程中盡量地 減少豬隻感受的緊迫。
2.4.4 車輛微環境
車輛微環境(trailer microclimate)通常是指在運輸過程中車體內部的溫度與濕 度,與該運輸發生的地理位置、季節與車輛設計有關。以溫度為例,當車上環境溫 度沒有維持在豬隻臨界溫度(critical temperature)的上下限內,也就是 26 °C 至 31
°C 時,豬隻就會經歷冷熱緊迫,其動物福利會受到損害,嚴重者更會導致死亡。
Voslarova et al.(2017)發現在捷克地區的豬隻,在冬季時有最高的死亡率,夏季次 之,春秋季死亡率最低,顯示該地區冷緊迫的影響大於熱緊迫。而Haley et al.(2008)
則發現加拿大地區的運輸過程中,溫度每上升 1°C,死亡率則增加為原先的 1.26 倍,顯示該地區的運輸較容易面對熱緊迫的挑戰,Grandin (1994)的研究也有相 似的結果,當外在溫度超過35°C 時,死亡率會從 0.15%上升至 0.30%。而 Sutherland et al.(2009)則使用 NANI 與 DOA 作為緊迫指標,發現車體溫度達 25°C 以上時 死亡率最高,而 0°C 以下最低,但豬隻無法行走的數量則是隨溫度下降而上升,
0°C 以下時最高,這顯示高低溫對豬隻的影響不盡相同,但皆對動物福利造成損害。
在炎熱的地區,Randall(1993)建議運輸車輛內的溫度不應該超過 30°C,而 台灣主要為亞熱帶及熱帶地區,豬隻在運輸過程中時常要面對嚴峻的熱緊迫,卻缺 乏對溫度的相關法令規範。在台灣熱緊迫對豬隻的影響,仍待更詳盡的研究。
第三章 材料與方法
3.1 試驗動物與場地
3.1.1 運輸緊迫源與腎上腺皮質醇之關聯
運輸試驗的豬隻來自屏東地區的台糖豬場—大響一畜殖場、大響二畜殖場、四 林繁殖場與四林畜殖場,以及部份的私人豬場。豬隻為商用肉豬,品系主要為LYD
(藍瑞斯母豬x 約克夏公豬 x 杜洛克公豬)與 YLD(藍瑞斯公豬 x 約克夏母豬 x 杜洛克公豬),體重約介於110 至 120 公斤間,主要運輸至台南市善化肉品市場、
高雄市岡山、旗山與鳳山肉品市場以及屏東縣肉品市場、台糖生鮮肉品工廠與信功 肉品廠。豬隻在運輸前一天移至豬場繫留欄位等待並且禁食,隔日司機抵達後進行 驅趕,驅趕時司機利用豬擋板、電擊棒等工具迫使豬隻移動至車體上。運輸貨車有 兩層,可利用液壓系統升降,裝載完畢後經過過磅便開始運輸。運輸過程中,司機 會視情況使用車體上的灑水系統或遮蔭裝置,為車上的豬隻降溫。抵達至目的地後,
司機會用高壓水柱清洗車上的豬隻,等待豬隻穩定後驅趕豬隻下車進入目的地繫 留欄位中,等待活體拍賣或屠宰。運輸時程因目的地而異,若運輸至肉品市場,通 常為上午八點到十二點,若運輸至台糖生鮮肉品市場及信功肉品廠,運輸時程則可 能延遲至下午兩點。當貨車承載兩層時,上下層豬隻有可能要送往不同目的地,司 機得依照當天出豬的豬場以及目的地優化出最佳路線。
3.1.2 日夜節律試驗與緊迫試驗
此二試驗之豬隻來自高雄地區的私人肉豬場,品系為LYD 豬種,試驗時約四 個月齡,共8 隻豬隻參與試驗。該豬場為水廉式豬舍,溫度恆介於 24-25℃,例行 操作為早上八點至九點、下午四點至五點的餵飼與定期性的投藥。
3.2 檢體採集方法 3.2.1 唾液採集
本研究以唾液中腎上腺皮質醇作為緊迫指標,因此唾液的採集十分重要。本研 究使用的唾液採集裝置為Cortisol-Salivette®(Sarstedt),由口蓋、棉條、內管與採 集管組成,如圖6 所示。事先在棉條上綁棉線,並將棉條投入豬隻口腔中,等待豬 隻咀嚼約30 秒至 1 分鐘唾液浸潤棉條後,扯動棉線收回棉條。之後,將棉條放入 內管與採集管、蓋上口蓋,以1000 g 離心 2 分鐘,便可將唾液離心至採集管內,
並以-20℃保存唾液樣本,等待測定皮質醇含量。
3.2.2 血液採集
採集血液前以鼻圈套索進行保定,確保豬隻不會隨意移動,並以 18 G 針頭
(TERUMO)與 10 ml 的針筒(TERUMO)進行採血。採血的方式為豬隻前腔靜 脈採血,將豬頭部仰起後,可見豬胸部兩側第一對肋骨與胸骨結合處前側(胸骨柄 兩側旁約 2 公分左右)各有一個前腔靜脈凹陷窩,取針具由下而上以垂直角度向 凹陷窩進針,進針後拉緊活塞使針筒內呈現負壓,扎破前腔靜脈壁後即會有血液流 入針筒。若無血液流出,可上下左右微調針頭,直到有血液流入針管。收集之血液 以室溫暫置,等待凝集後以1,000 x g 離心後取上清液血清,再將血清置於-20℃保 存,等待測定皮質醇含量。
3.3 試驗設計
3.3.1 運輸緊迫源與腎上腺皮質醇之關聯
本研究欲探討豬隻運輸過程中唾液中腎上腺皮質醇變化與緊迫源之間的關聯,
因此需要在運輸過程中進行豬隻唾液採樣。唾液採集的時間點分別為,豬隻位於豬 場繫留欄位時(第一點)、經過驅趕移動至車體時(第二點)、經過運輸抵達目的地 時(第三點)如圖7 所示。採樣後,唾液採集管先後以 4℃、-20℃暫存,等待離
圖6. 唾液採集裝置,由上而下為口蓋、棉條、內管與採集管。
Fig 6. Saliva collection device are made of cover, swab, inner tube and collecting tube.
圖7.運輸各階段。圖 7-1 為驅趕前豬隻於牧場繫留欄位內、圖 7-2 為驅趕過程、圖 7-3 為驅趕後豬隻於車體上、圖 7-4 為豬隻運輸後於車體上。
Fig 7. Each stage of transportation. Fig 7-1 show pigs in the picked-up facility before loading. Fig 7-2 show the processing of loading. Fig 7-3 show pigs on the vehicle after loading. Fig 7-4 show pigs on the vehicle after transport.