以葉綠體基因組之非密碼區及核核糖體 DNA 之序列探討台灣地區
鬼針草(
Bidens pilosa L.)之遺傳多樣性
蔡麗琴
1,2廖培鈞
1謝幸媚
2李俊億
3王震哲
1* 1 國立臺灣師範大學生命科學系 2中央警察大學鑑識科學系 3 國立臺灣大學醫學院法醫學科 (收稿日期:2007.4.12,接受日期:2007.5.9) 摘 要 根據台灣植物誌之記載鬼針草(Bidens pilosa L.)於台灣地區共有三個變種,分別為大花咸豐草 (B. pilosa var. radiata)、小白花鬼針(B. pilosa var. minor)及白花鬼針(B. pilosa var. pilosa)。本研究採集此三個變種共 11 個族群 161 株個體,加上 7 株當控制組的鬼針舅(Bidens biternata)共 168 個樣本,分別進行 ITS1、 5.8S 及 ITS2 等三個細胞核核糖體 DNA(nrDNA)的基因位與 trnL intron 及trnL-trnF IGS 等兩個葉綠體(cpDNA)的基因位之序列分析,將 nrDNA 之三個基因位及 cpDNA
之兩個基因位之序列分別合併再進行排比後其序列長度分別為 637 及 799 bp,本研究即以此排比後 之 nrDNA 及 cpDNA 序列進行此三個變種間、變種內、族群間及族群內等不同階層之遺傳多樣性 及基因交流之探討。根據核苷酸變異位點(S)、序列型(H)、基因型歧異度(gene diversity,Hd)、
核苷酸歧異度(nucleotide diversity,π)、遺傳歧異度(genetic diversity,θ)等相關指數顯示 nrDNA 其遺傳多樣性均遠高於 cpDNA。至於族群內及變種內之遺傳分化程度(FST)為大花咸豐草<小白 花鬼針<白花鬼針,而基因交流值(Nm)則反之(大花咸豐草>小白花鬼針>白花鬼針)。AMOVA
分析之結果顯示Bidens pilosa 遺傳變異的主要貢獻為族群內個體間的變異,於 nrDNA 及 cpDNA
均佔 50 %以上,其次則為變種間及變種內族群間的遺傳變異所致。大花咸豐草與其他兩變種間的 基因交流程度遠低於白花鬼針與小白花鬼針之間的基因交流值,極可能較早歸化的白花鬼針與小白 花鬼針間已出現明顯的雜交現象,至於較晚歸化的大花咸豐草與此二變種間的雜交現象則可能尚不 明顯。
關鍵詞:遺傳變異、鬼針草、nrDNA、ITS1、ITS2、5.8S、cpDNA、trnL intron、trnL-trnF IGS
緒 言
鬼針草(Bidens pilosa L.)為菊科植物鬼針屬 之一種,於世界各地常應用於傳統醫學或民俗療 法 ( Chiang et al., 2003; Andrade-Neto et al.,
2004),相關研究多集中於植株萃取物有效成分
之分離及鑑定(Chang et al., 2004)、在現代醫學 之疾病治療(Sundararajan et al., 2006; Yang et al., 2006)等方面的應用。根據台灣植物誌之記載 B.
pilosa 於台灣地區共有三個變種,分別為大花咸
豐草(B. pilosa var. radiata)、小白花鬼針(B.
pilosa var. minor)及白花鬼針(B. pilosa var. pilosa)(Peng et al., 1998)。2003 年鄧書麟等人將
頭花為輻射狀且舌狀花花冠成深裂狀者處理為
另一新的變種即裂瓣鬼針(Bidens pilosa var.
trivalvis)(Deng et al., 2003)。本研究則仍以台灣 植物誌第二版記載之各變種為對象。這些變種在 台灣地區均廣泛分布於中低海拔,其中大花咸豐 草原生於美洲,目前則廣泛分布於南美、北美、 北非及南亞,而台灣地區則於近二十年來才出現 此侵略性的新歸化種,其棲地幾乎與較早歸化的 小白花鬼針及白花鬼針重疊,甚至於其族群及個 體的數量已凌駕於此兩變種之上。由於三個變種 之形態類似且呈共域分布,故此三個變種間、族 群 間 及 族 群 內 之 遺 傳 變 異 與 基 因 交 流 ( gene flow)之現象均值得深入探討。 鄧書麟等人依據簡單序列重複分析法(inter- simple sequence repeat, 簡稱 ISSR)之研究,認為
B. pilosa 各變種間可能存在雜交及基因交流之現 象(Deng et al., 2005),但其研究並未比較兩兩變 種間、各族群間及族群內之遺傳分化及基因交流 之程度。此外,以ISSR 所得之 DNA 指紋圖譜於 植物之品系鑑定及親緣關係之分析上可能為一 有用的工具,然而此種 DNA 指紋圖譜之數據並 不適合資料庫之建立或實驗室間結果之比對,加 上植物樣品若屬於較為陳舊之檢體,其基因組內 之 DNA 將遭降解,且可能會有細菌或真菌之污 染,使得ISSR 圖譜的再現性面臨挑戰。 本研究以特定基因位之 DNA 序列來探討大 花咸豐草、小白花鬼針及白花鬼針此三個變種間 於不同階層之遺傳變異與遺傳分化。由於非基因 區等不表現的序列(non-coding DNA regions)具 有較快的演化速率,因此常應用於低階類群之鑑 定、分類研究、演化探討及親緣分析,其中位於
葉 綠 體 基 因 組 DNA(chloroplast DNA, 簡稱
cpDNA)之 trnL intron (UAA)、 trnL-trnF (GAA) IGS (intergenic spacer)及位於細胞核之核糖體 DNA(nuclear ribosomal DNA,簡稱 nrDNA)的 ITS1(internal transcribed spacer)、5.8S 及 ITS2 (internal transcribed spacer)等基因位均常應用 於植物之科內到種內之鑑別或遺傳變異之相關 研究 (Taberlet et al., 1991; Fangan et al., 1994; Baldwin and Markos, 1998; Terry et al., 2000; Holt
et al., 2004; Tsai et al., 2006a., 2006b ),因此本研
究以此五個基因位之 DNA 序列特徵來探討 B. pilosa 之三個變種於不同階層的遺傳多樣性及基 因交流情形。
材料與方法
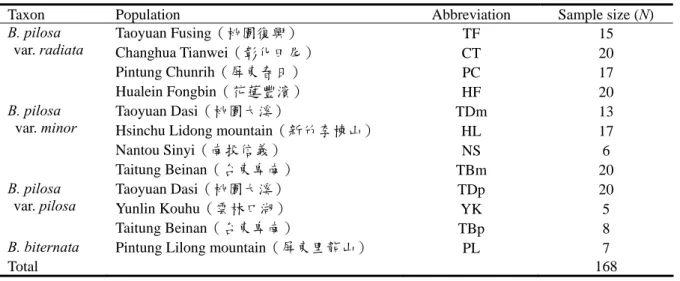
材料 本研究採集台灣地區鬼針草(Bidens pilosa L.)的三個變種各三或四個族群共 161 株個體及 本屬之另一種鬼針舅(Bidens biternata)7 株個體 當控制組,共計168 株樣本,各採集地點及樣品 數量如表一所示。各變種之分類鑑定乃依據台灣 植物誌第二版之形態特徵(Peng et al., 1998)。 方法 DNA 之萃取與定量 摘取嫩葉一片先以蒸餾水將表面附著物洗 除,接著置入含有液態氮之研缽中研磨成粉狀, 取約10 mg 粉末置入 1.5 mL 的離心管中,以萃取 植物 DNA 專用之商業套組進行 DNA 之萃取(Plant Genomic DNA Miniprep System, Viogene, Taipei, Taiwan),萃取之基因組 DNA 則以 0.7 % 瓊酯膠電泳(agarose gel electrophoresis)及分光
光度計進行DNA 之定量,最後於–20 oC 冰箱中
儲存。
PCR 複製
本研究trnL intron、trnL-trnF IGS、ITS1、5.8S
及ITS2 等五個基因位進行 PCR 複製所需之引子
直接採用前人研究(White et al., 1990; Taberlet et
al., 1991; Fangan et al., 1994)之通用引子,各基
因位進行 PCR 複製所需的條件均相同,分述如
下:反應溶液之總體積為50 μL,內含各引子 0.3
μM、pH 8.3 的緩衝液(10 mM Tris-HCl, 2.5 mM MgCl2, 50 mM KCl, 0.1 % (w/v) gelatin)、200 μM
dNTP 、 2.5 unit 的 VioTaq DNA polymerase (Viogene) 和 10 ng 模板 DNA。 使用之熱循環 儀為 2400 Perkin-Elmer thermal cycler (Applied
Biosystems),其熱循環條件設定為 94 oC 1 分
鐘,50 oC 1 分鐘 及 72 oC 2 分鐘進行 35 個循
環,接著72 oC 7 分鐘,最後維持在 4 oC。
DNA 序列分析
PCR 產物於 2 % 瓊酯膠進行電泳確認並 以 PCR-MTM Clean Up System (Viogene)進行純
化,接著使用 ABI PRISMTM BigDyeTM Terminator
Cycle Sequencing Ready Reaction Kit 並分別以正 向(forward)及反向(reverse)引子進行 PCR 產物之循環定序(cycle sequencing)PCR 複製,
此循環定序 PCR 產物以 POP-7TM performance
optimized polymer (Applied Biosystems)進行分 析並以ABI 3130 DNA Analyzer 進行 DNA 序列之 偵 測 。 各 基 因 位 之 序 列 則 以 國 家 衛 生 研 究 院 (http://bioinfo.nhri.org.tw ) 之 GCG computer package 之 PileUp 程式進行序列間之排比。 統計分析 ( 一 ) 遺 傳 多 樣 性 分 析 ( Analysis of genetic diversity) 排序後的 DNA 序列利用 Arlequin v. 3.0.1 (Excoffier et al., 2005)程式計算各種遺傳多樣性 指數,包含基因型歧異度(gene diversity,Hd)、 核苷酸歧異度(nucleotide diversity,π)、遺傳歧異 度(genetic diversity,θ),本研究中的 θ 由同質 型期望值(expected homozygosity)計算而來 (Zouros, 1979)。本研究分別以族群、變種及
表一、本研究之採樣分布。
Table 1. Sample distribution in this study.
Taxon Population Abbreviation Sample size (N)
Taoyuan Fusing(桃園復興) TF 15 Changhua Tianwei(彰化田尾) CT 20 Pintung Chunrih(屏東春日) PC 17 B. pilosa var. radiata Hualein Fongbin(花蓮豐濱) HF 20 Taoyuan Dasi(桃園大溪) TDm 13
Hsinchu Lidong mountain(新竹李棟山) HL 17
Nantou Sinyi(南投信義) NS 6 B. pilosa var. minor Taitung Beinan(台東卑南) TBm 20 Taoyuan Dasi(桃園大溪) TDp 20 Yunlin Kouhu(雲林口湖) YK 5 B. pilosa var. pilosa Taitung Beinan(台東卑南) TBp 8
B. biternata Pintung Lilong mountain(屏東里龍山) PL 7
Total 168 Bidens pilosa 複合群為單位計算各種遺傳多樣性 指數,以了解各族群、變種及複合群內的多樣性 程度。 ( 二 ) 遺 傳 結 構 分 析 (Analyses of genetic structure) 本研究將Bidens pilosa 區分為變種及族群兩 個階層,以分子變方分析(analysis of molecular variance, AMOVA)判斷各階層的遺傳變異程度, 判斷變種間、變種內族群間以及族群內的變異程 度,並佐以分類群之間的遺傳分化程度(Φ 值)的 統計,判斷族群結構的顯著性。AMOVA 的分析 亦利用 Arlequin v. 3.0.1 (Excoffier et al., 2005) 程 式分析,進行 1023 次變更取樣(permutation)以計 算統計之支持度。
( 三 ) 族 群 分 化 及 基 因 交 流 (Population differentiation and gene flow)
利用 Arlequin v. 3.0.1 (Excoffier et al., 2005) 程式進行序列分析以比較各族群間、各變種間的 遺傳分化指數FST (genetic distance,pairwise FST)
(Reynolds et al., 1983; Slatkin, 1995),並利用 FST
與基因交流值(Nm)的倒數關係評估各族群間、各 變種間基因交流的程度。核 DNA 的基因交流值 的計算為 Nm=(1-FST)/4FST;由 cpDNA 估計的基 因交流值為Nm=(1-FST)/2FST (Wright, 1951)。此 外,亦利用鄰近歸群法(neighbor-joining method) 以 FST描繪族群及變種的分支樹(cladogram)。分
支樹的構圖則利用 MEGA 3.1 (Kumar et al., 2004) 程式進行。
結 果
序列分析(Sequence analysis) 所有樣本之trnL intron、trnL-trnF IGS、ITS1、 5.8S 及 ITS2 等五個基因位之序列經序列排比 (alignment)後其長度分別為 438、362、256、 164 及 217bp。將 trnL intron 及 trnL-trnF IGS 等 兩個 cpDNA 的基因位合併後之序列排比長度為 800bp;而 ITS1、 5.8S 及 ITS2 等三個 nrDNA 的 基因位合併後之序列排比長度則為 637bp。經由 序列排比之結果發現 nrDNA 之變異位點數目(S) 遠高於 cpDNA,Bidens pilosa 之三個變種 161 個 樣本中具有 26 個變異位點(佔總排比長度之 4.08 %),分屬 73 種序列型(H)(表二),7 個鬼針舅 樣本間則具有 4 個變異位點分屬 3 種序列型;而 cpDNA 於 Bidens pilosa 則僅具 8 個變異位點(佔總排比長度之 1.00 %),分屬 5 種序列型(表三),
至於鬼針舅則只出現單一型別,由分析結果發現 鬼針草類群 nrDNA 鹼基變異率約為 cpDNA 之 4 倍。不論是 nrDNA 或 cpDNA 序列,在 Bidens
pilosa 三個變種之所有個體中均未發現與鬼針舅 相同的序列型。 Bidens pilosa 於不同階層之遺傳多樣性及基因交 流 Bidens pilosa 三個變種其 11 個族群內於 nrDNA 及 cpDNA 序列之遺傳多樣性各有不同程 度的差異,由表二及表三之結果顯示 nrDNA 序 列於基因型歧異度(Hd)、核苷酸歧異度(π)及 遺傳歧異度(θ)等三項指數之值均遠高於 cpDNA
表二、Bidens pilosa 三個變種內及其族群內於 nrDNA 之遺傳多樣性。
Table 2. The genetic diversity of 11 populations in three varieties of Bidens pilosa in nrDNA.
Population N H S Hd π θ FST Nm
TF 30 18 26 0.956 0.0177 20.23 0.131 1.658 CT 40 15 18 0.905 0.0114 8.22 0.143 1.498 PC 34 14 21 0.895 0.0093 7.25 0.147 1.451 HF 40 16 23 0.926 0.0126 11.00 0.141 1.523
B. pilosa var. radiata 144 33 26 0.938 0.0139 13.58 0.141 1.523 TDm 26 11 20 0.914 0.0087 9.23 0.178 1.154
HL 34 17 20 0.931 0.0143 11.90 0.162 1.293 NS 12 4 3 0.727 0.0021 2.05 0.195 1.032 TBm 40 25 20 0.980 0.0139 45.91 0.163 1.284
B. pilosa var. minor 112 38 20 0.963 0.0131 24.31 0.170 1.221 TDp 40 19 20 0.949 0.0143 16.89 0.219 0.892
YK 10 4 9 0.822 0.0067 3.71 0.248 0.758 TBp 16 6 18 0.817 0.0104 3.56 0.234 0.818
B. pilosa var. pilosa 66 27 20 0.962 0.0141 23.45 0.227 0.851 Total 322 73 26 0.993 0.0193 132.70 0.380 0.407
N is the sample size (the sample size of diploid nrDNA is doubled), H is the number of haplotypes, S is the number of
polymorphic sites, Hd is the gene diversity, π is the nucleotide diversity, and θ is the genetic diversity, FST is genetic distance, Nm is gene flow.
表三、Bidens pilosa 三個變種內及其族群內於 cpDNA 之遺傳多樣性。
Table 3. The genetic diversity of 11 populations in three varieties of Bidens pilosa in cpDNA.
Population N H S Hd π θ FST Nm
TF 15 3 7 0.648 0.0026 1.39 0.030 16.167 CT 20 2 1 0.505 0.0006 0.76 0.120 3.667
PC 17 1 0 0 0 0 0.150 2.833
HF 20 3 7 0.416 0.0013 0.53 0.090 5.056
B. pilosa var. radiata 72 3 7 0.440 0.0012 0.59 0.100 4.500 TDm 13 2 7 0.385 0.0035 0.47 0.350 0.929
HL 17 2 6 0.515 0.0040 0.79 0.337 0.984
NS 6 1 0 0 0 0 0.429 0.666
TBm 20 1 0 0 0 0 0.429 0.666
B. pilosa var. minor 56 3 7 0.383 0.0029 0.46 0.383 0.805 TDp 20 4 7 0.732 0.0044 2.10 0.418 0.696
YK 5 1 0 0 0 0 0.516 0.469
TBp 8 1 0 0 0 0 0.516 0.469
B. pilosa var. pilosa 33 4 7 0.583 0.0041 1.05 0.457 0.594 Total 161 5 8 0.939 0.0029 13.84 0.272 1.335
N is the sample size, H is the number of haplotypes, S is the number of polymorphic sites, Hd is the gene diversity, π is
the nucleotide diversity, and θ is the genetic diversity, FST is genetic distance, Nm is gene flow.
序列分析之結果。至於各族群內之遺傳分化指數 (FST)於 nrDNA 與 cpDNA 之分析結果幾乎均具 有相同的趨勢,即大花咸豐草最低,其次為小白 花鬼針,最高為白花鬼針;而由 FST值估算此二 基因位之基因交流值(Nm)則發現大花咸豐草各 族群內之基因交流值最高,其次為小白花鬼針, 最低為白花鬼針。根據 nrDNA 序列所得各族群 內之Nm 值,於大花咸豐草及小白花鬼針均大於 1,而白花鬼針則均小於 1;至於 cpDNA 序列分 析之結果除了大花咸豐草各族群內之Nm 值大於 1 以外,其他兩個變種各族群內之 Nm 值則均小 於 1。 各變種內於 nrDNA 及 cpDNA 序列之遺傳多 樣 性 也 發 現 各 有 不 同 程 度 的 差 異 ( 表 二 及 表 三),而代表遺傳多樣性的三項指數(基因型歧 異 度 、 核 苷 酸 歧 異 度 及 遺 傳 歧 異 度 ) 也 發 現
nrDNA 序列分析之結果均遠高於 cpDNA 者。至 於各變種內之遺傳分化指數(FST)於 nrDNA 與 cpDNA 之分析結果亦均具有相同的趨勢,即大花 咸豐草最<小白花鬼針<白花鬼針;而 Nm 值則相 反為大花咸豐草最>小白花鬼針>白花鬼針,其中 nrDNA 的 Nm 值於大花咸豐草為 1.523,小白花 鬼針為 1.221,白花鬼針為 0.851,至於 cpDNA 則分別為 4.500、0.805 及 0.594。 將此三個變種共 11 個族群間之 FST 值以 neighbor-joining 方法進行歸群分析所建立之分支 樹如圖一及圖二所示,nrDNA 序列之分析結果顯 示大花咸豐草之四個族群均歸屬在同一分支,但 小白花鬼針及白花鬼針此二變種之族群並無法 有效歸群;cpDNA 分析之結果亦與 nrDNA 歸群 之結果相類似,但大花咸豐草於桃園復興鄉之族 群 歸 入 小 白 花 及 白 花 鬼 針 的 分 支 中 。 由 表 四 nrDNA 序列之分析結果顯示大花咸豐草之四個 族群間有較顯著的基因交流;而 cpDNA 分析之 結果雖然發現此變種的族群間亦有明顯的基因 交流,但桃園復興鄉的族群卻發現與其他兩個變 種之多個族群間也有明顯的基因交流情形。 至於各變種間的遺傳分化及基因交流程度 則如表五及表六所示,大花咸豐草與小白花鬼針 兩變種間其 FST 值為 0.450 (nrDNA) 及 0.317 (cpDNA),Nm 值分別為 0.306 及 1.077;大花咸 豐 草 與 白 花 鬼 針 間 之 FST 值 分 別 為 0.416 及 0.418,Nm 值分別為 0.351 及 0.696;小白花鬼針 與白花鬼針間之 FST值分別為 0.035 及 0.028,Nm 值分別為 6.893 及 17.357。至於鬼針舅與此三個 變種間均具有較高的 FST值,其 Nm 值則遠低於 此三個變種間的值。而由表五 Bidens pilosa 三個 變種及鬼針舅間的 FST值進行歸群分析所建立之 分支樹(圖三)則顯示,不論以 nrDNA 或 cpDNA 序列歸群之結果均具有相同的趨勢,即小白花鬼 針及白花鬼針間具有最小的遺傳分化,接著為大 花咸豐草,遺傳分化最大者為鬼針舅。
圖一、Bidens pilosa 11 個族群間於 nrDNA 序列之遺傳 分化指數所建構之 neighbor-joining 樹狀圖。
Figure 1. The neighbor-joining tree of genetic distance
(FST) based on nrDNA of the populations of Bidens
pilosa.
圖二、Bidens pilosa 11 個族群間於 cpDNA 序列之遺傳 分化指數所建構之 neighbor-joining 樹狀圖。
Figure 2. The neighbor-joining tree of genetic distance
(FST) based on cpDNA of the populations of Bidens
pilosa.
圖三、以鄰近歸群法建構 B. pilosa 三個變種及 B. biternata 的親緣關係。左圖為以 nrDNA 之遺傳分化指數建構 之親緣關係圖,右圖為以 cpDNA 之遺傳分化指數建構之親緣關係圖。
Figure 3. The neighbor-joining tree of genetic distance (FST) based on nrDNA (left) and cpDNA (right) of three varieties of B. pilosa and B. biternata.
表四、Bidens pilosa 三個變種其族群間之基因交流。
Table 4. Pairwise gene flow (Nm) calculated among populations of B. pilosa by Wright’s (1951) equation. B. pilosa var. radiata B. pilosa var. minor B. pilosa var. pilosa
TF CT PC HF TDm HL NS TBm TDp YK TBp TF 9.92 1.42 18.73 7.19 1.00 4.93 1.56 0.91 6.85 3.44 CT 1.66 0.95 ∞ 1.04 0.36 0.63 0.36 0.37 0.68 0.56 PC 0.60 1.15 5.60 0.45 0.26 0.00 0.00 0.28 0.00 0.00 HF 1.47 5.18 5.43 1.17 0.45 0.80 0.45 0.45 0.87 0.71 TDm 0.36 0.15 0.12 0.18 2.34 6.75 1.70 2.23 10.86 4.22 HL 0.72 0.24 0.20 0.29 1.59 0.65 0.35 ∞ 0.70 0.57 NS 0.27 0.12 0.08 0.14 1.87 0.64 ∞ 0.65 ∞ ∞ TBm 0.75 0.25 0.22 0.32 0.89 16.42 0.43 0.37 ∞ ∞ TDp 0.78 0.24 0.21 0.31 1.04 ∞ 0.53 83.08 0.70 0.58 YK 0.41 0.16 0.11 0.19 1.03 0.83 0.50 0.64 0.78 ∞ TBp 0.61 0.20 0.17 0.27 0.28 1.28 0.14 3.66 1.87 0.23 The upper values are calculated by cpDNA; the bottom values are calculated by nrDNA.
表五、Bidens pilosa 之三個變種及 B. biternata 於 nrDNA 及 cpDNA 之遺傳分化。
Table 5. Genetic distance (FST) based on nrDNA and cpDNA in the three varieties of Bidens pilosa and B. biternata.
B. pilosa var. radiata B. pilosa var. minor B. pilosa var. pilosa B. biternata
B. pilosa var. radiata 0.317 0.418 0.634
B. pilosa var. minor 0.450 0.028 0.361
B. pilosa var. pilosa 0.416 0.035 0.378
B. biternata 0.887 0.890 0.889 The upper values are calculated by cpDNA; the bottom values are calculated by nrDNA.
表六、Bidens pilosa 之三個變種及 B. biternata 於 nrDNA 及 cpDNA 之基因交流。
Table 6. Pairwise gene flow (Nm) calculated among three varieties of Bidens pilosa and B. biternata. B. pilosa var. radiata B. pilosa var. minor B. pilosa var. pilosa B. biternata
B. pilosa var. radiata 1.077 0.696 0.289
B. pilosa var. minor 0.306 17.357 0.885
B. pilosa var. pilosa 0.351 6.893 0.823
B. biternata 0.032 0.031 0.031 The upper values are calculated by cpDNA; the bottom values are calculated by nrDNA.
分子變方分析(Analysis of molecular variance, AMOVA)
以 AMOVA 分析 Bidens pilosa 三個變種之 11 個族群之遺傳變異如表七及表八所示,nrDNA 之 分析結果顯示三個變種間的變異成分佔總變異 的 34.75 %,變種內族群間的變異佔 10.82 %, 族群內個體間的變異則佔 54.43 %,而其遺傳分 化程度(Φ 值)則分別為 0.3475、0.1658 及 0.4557。至於 cpDNA 分析之結果也顯示族群內個 體間所佔的變異最大為 53.52 %,其次為變種內 族群間的變異佔 27.41 %,最小為三個變種間的 差異佔 19.06 %,其 Φ 值則分別為 0.4648、0.3387 及 0.1906。這些數值除了 cpDNA 序列其變種間 的變異之 P 值為 0.1017 之外,其餘之 P 值均小 於 0.0001。
討 論
由 序 列 分 析 之 結 果 發 現 nrDNA 序 列 較 cpDNA 具有較多的變異位點及序列型,同時代表 遺傳多樣性之參數如Hd、π 及 θ 等指數也表現出 較高的數值,本研究由 161 個 Bidens pilosa 樣本 中,於 nrDNA 之序列分析即出現 73 種序列型, 表示個體間存在極大的差異。經由 AMOVA 分析表七、Bidens pilosa 三個變種之族群間於 nrDNA 序列之分子變方分析。
Table 7. Analyses of molecular variance (AMOVA) among populations of three varieties in Bidens pilosa using the
sequences of nrDNA.
Source of variation d.f. % total variance Φ P Among varieties 2 34.75 0.3475 <0.0001
Among populations within varieties 8 10.82 0.1658 <0.0001 Within populations 311 54.43 0.4557 <0.0001 Variance explained by differences among varieties and its significance using probabilities derived from 1023 permutations are indicated.
表八、Bidens pilosa 三個變種之族群間於 cpDNA 序列之分子變方分析。
Table 8. Analyses of molecular variance (AMOVA) among populations of three varieties in Bidens pilosa using the
sequences of cpDNA.
Source of variation d.f. % total variance Φ P Among varieties 2 19.06 0.1906 0.1017
Among populations within varieties 8 27.41 0.3387 <0.0001 Within populations 150 53.52 0.4648 <0.0001 Variance explained by differences among varieties and its significance using probabilities derived from 1023 permutations are indicated.
之結果(表七及表八)亦顯示 Bidens pilosa 遺傳變 異的主要貢獻為族群內個體間的變異,於 nrDNA 及 cpDNA 均佔 50 %以上,其次則為變種間及變 種內族群間的遺傳變異所致。此結果亦與鄧書麟 等人之結果相符(Deng et al., 2005)。但 AMOVA 分 析 之 結 果 ( 表 七 及 表 八 ) 仍 出 現 nrDNA 及 cpDNA 部份結果不甚一致的現象,如變種內族群 間的變異及變種間的變異所佔之變異百分比之 趨勢略有不同,且 cpDNA 序列其變種間的變異 之P 值超過 0.05,此差異現象可能是由於 nrDNA 與 cpDNA 經歷不同的演化歷史所致,且 cpDNA 為母系遺傳,不受花粉(父系)影響,亦不如 nrDNA 可能受到重組事件之影響。再者,二者之演化速 率 不 同 , 一 般 而 言 nrDNA 的 鹼 基 變 異 率 較 cpDNA 為高,例如菊科植物菜薊族(Cardueae) 其 nrDNA 之鹼基變異率約為 cpDNA 之 8 倍 (Wang et al., 2007)、黑藥花科(Melanthiaceae)之
Paris 屬約為 4 倍(Ji et al., 2006),,而本研究則發
現鬼針草類群 nrDNA 鹼基變異率約為 cpDNA 之 4 倍。可能也因此於進行序列分析時影響其解析 力,而由於本研究對象為同種之不同變種,在兩 支系演化時間相距太短之情形下,所產生之新的 突變其出現僵化(fixed)的時間也將不同,因而導 致 譜 系 撿 選 (lineage sorting) 速 率 的 差 異 (Nei, 1987; Pamilo and Nei, 1988; Lyons-Weiler and Milinkovitch, 1997)。因此在以 cpDNA 分析結果 進行族群間之歸群分析時,出現大花咸豐草於桃 園復興鄉之族群歸入小白花鬼針及白花鬼針的 分支中(圖二),可能即為譜系撿選速率的差異影 響到親緣樹的建構。 比較各變種內及變種內各族群內之遺傳分 化指數及基因交流值(表二及表三),則發現不論 是 nrDNA 或 cpDNA 序列分析之 FST值均為大花 咸豐草<小白花鬼針<白花鬼針,而Nm 值的大 小幾乎均為大花咸豐草>小白花鬼針>白花鬼 針,此趨勢恰與鄧書麟等人(Deng et al., 2005)以 ISSR 分析之指紋圖譜的結果中,認為大花咸豐草 變種內的遺傳分化程度高於其他兩個變種之結 果相反,但由於該篇論文所使用之分析技術與本 研究所使用之特定基因位之序列比較不同,且並 未進行如表二及表三各族群內之 FST值的完整呈 現,由本研究於 nrDNA 及 cpDNA 所分析之結果 發現大花咸豐草之遺傳分化指數 (FST) 不論於 各族群內或變種內均呈現一致之趨勢,即 FST值 均明顯低於白花鬼針草及小白花鬼針者,故本研 究之結果應屬可信。而根據楊國楨(2001)野外觀 察發現大花咸豐草由於其多年生的競爭優勢,在 很多地區已經取代了較早移居台灣的小白花鬼 針與白花鬼針之現象,族群的快速擴散使得其變 種內及變種內各族群內擁有較高的基因交流應 屬合理。 表六之結果顯示各變種之間的基因交流情 形,其中白花鬼針與小白花鬼針的基因交流值分 別約為與外群(鬼針舅)的基因交流值的 220 倍
(nrDNA)與 20 倍(cpDNA)左右;大花咸豐草與小 白花鬼針、大花咸豐草與白花鬼針的基因交流值 分別約為大花咸豐草與鬼針舅的 10 倍(nrDNA) 與 2-4 倍 (cpDNA)。此結果顯示白花鬼針與小白 花鬼針兩變種之間雜交的機會高於與大花咸豐 草雜交的機會,而此三變種之間雜交的機會更高 於同屬不同種(鬼針舅)間的雜交機會。表五之結 果顯示三變種之間的遺傳分化程度均不高(特別 是白花鬼針與小白花鬼針兩變種之間),其原因 除了此三變種在遺傳上仍未完全分化,致使擁有 較多相同的基因型外,物種之間的分化不完全以 及共域分佈亦導致較高的雜交機會,進而消除變 種間在遺傳組成上的阻隔藩籬。而大花咸豐草與 白花鬼針及小白花鬼針間的基因交流程度遠低 於後二者之間的基因交流值,可能與此三者歸化 的時間有關,白花鬼針與小白花鬼針歸化時間可 能均已超過百年,長期的共域分佈導致頻繁的雜 交,而大花咸豐草之歸化時間僅約二十餘年,其 與白花鬼針和小白花鬼針之間的雜交情形自未 若後二者之間的頻繁。根據 Bidens pilosa 三個變 種及鬼針舅間 FST 值所建立之分支樹(圖三)顯 示,小白花鬼針及白花鬼針間具有較近的遺傳距 離,接著為大花咸豐草,最遠者為鬼針舅,此歸 群之結果與與上述之推論相符,亦與鄧書麟等人 (Deng et al., 2005)之研究結果一致。將此三個變 種 共 11 個 族 群 間 之 遺 傳 分 化 值 以 neighbor-joining 方 法 進 行 歸 群 分 析 ( 圖 一 及 圖 二),發現小白花鬼針及白花鬼針之族群並無法依 類群有效歸群,此結果亦支持此二變種間的遺傳 分化並不明顯之論證,而大花咸豐草與其他兩變 種間的基因交流程度亦遠低於此二變種間的基 因交流值(表六),因此,極可能較早歸化的白 花 鬼 針 與 小 白 花 鬼 針 間 已 出 現 明 顯 的 雜 交 現 象,至於較晚歸化的大花咸豐草與此二變種間的 雜交現象則可能尚不明顯。
誌 謝
本研究承行政院國家科學委員會計畫(NSC 92-2320-B-015-003)經費補助。參考文獻
楊國楨 2001. 外來植物大車拼(二)大花咸豐草 以及它的弟兄們。 http://tnl.org.tw/article/column/exotic/exotic2 /exotic2.htmAndrade-Neto VF, Brandao MG, Oliveira FQ, Casali VW, Njaine B, Zalis MG, Oliveira LA and Krettli AU. 2004. Antimalarial activity of
Bidens pilosa L. (Asteraceae) ethanol extracts
from wild plants collected in various localities or plants cultivated in humus soil. Phytother. Res. 18: 634-639.
Baldwin BG and Markos S. 1998. Phylogenetic utility of the external transcribed spacer (ETS) of 18S-26S rDNA: congruence of ETS and ITS Trees of Calycadenia (Compositae). Mol. Phylogenet. Evol. 10: 449-463.
Chang SL, Chang CL, Chiang YM, Hsieh RH, Tzeng CR, Wu TK, Sytwu HK, Shyur LF and Yang WC. 2004. Polyacetylenic compounds and butanol fraction from Bidens pilosa can modulate the differentiation of helper T cells and prevent autoimmune diabetes in non-obese diabetic mice. Pl. Med. 70: 1045-1051.
Chiang LC, Chang JS , Chen CC , Gg LT and Lin CC. 2003. Anti-Herpes simplex virus activity of Bidens pilosa and Houttuynia cordata. Am. J. Chin. Med. 31: 355-362.
Deng SL, Lu FY and Ho KY. 2003. The genus
Bidens (Compositae) in Taiwan. Quart. J.
Chin. For. 33: 331-348.
Deng SL, Lu FY, Ho KY and Kuo YW. 2005. Genetic variation of the Bidens pilosa complex (Compositae) in Taiwan. Quart. J. Chin. For. 38: 397-408.
Excoffier L, Laval G and Schneider S. 2005. Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evol. Bioinform. Online 1: 47-50.
Fangan BM, Stedje B, Stabbetorp OE, Jnsen ES and Jakobsen KS. 1994. A general approach for PCR-amplification and sequencing of chloroplast DNA from crude vascular plant and algal tissue. Biotechniques 16: 484-494. Holt SD, Horova L and Bures P. 2004. Indel
patterns of the plastid DNA trnL- trnF region within the genus Poa (Poaceae). J. Pl. Res. 117: 393-407.
Ji Y, Fritsch PW, Li H, Xiao T and Zhou Z. 2006. Phylogeny and classification of Paris (Melanthiaceae) inferred from DNA sequence data. Ann. Bot. 98: 245-256.
Kumar S, Tamura K and Nei M. 2004. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief Bioinform 5: 150-163. Lyons-Weiler J and Milinkovitch MC. 1997. A
phylogenetic approach to the problem of differential lineage sorting. Mol. Biol. Evol. 14: 968-975.
Nei, M. 1987. Molecular evolutionary genetics. Columbia University Press, New York.
Pamilo P and M. Nei. 1988. Relationships between gene trees and species trees. Mol. Biol. Evol. 5: 568-583.
Peng CI, Chung KF and Li HL 1998. Bidens. In Huang TC et al. (eds.), Flora of Taiwan, 2nd edition. Editorial Committee of the Flora of Taiwan, Vol. 4, pp. 868-870.
Reynolds J, Weir BS and Cockerham CC. 1983. Estimation for the coancestry coefficient: basis for a short-term genetic distance. Genetics 105: 767-779.
Slatkin M. 1995. A measure of population subdivision based on microsatellite allele frequencies. Genetics 139: 457-462.
Sundararajan P, Dey A, Smith A, Doss AG, Rajappan M and Natarajan S. 2006. Studies of anticancer and antipyretic activity of
Bidens pilosa whole plant. Afr. Health Sci. 6:
27-30.
Taberlet P, Gielly L, Pautou G and Bouvet J. 1991. Universal primers for amplification of three non-coding regions of chloroplast DNA. Pl. Mol. Biol. 17: 1105-1109.
Terry RG, Nowak RS and Tausch RJ. 2000. Genetic variation in chloroplast and nuclear ribosomal DNA in Utah Juniper (Juniperus
osteosperma, Curpressaceae): evidence for
interspecific gene flow. Am. J. Bot. 87: 250-258.
Tsai LC, Hsieh HM, Huang LH, Wang JC, Linacre A and Lee JCI. 2006. Cannabis seed identification by chloroplast and nuclear DNA. Forensic Sci. Int. 158 (2-3): 250-251. Tsai LC, Yu YC, Hsieh HM, Wang JC, A. Linacre
A and Lee JCI. 2006. Species identification using sequences of the trnL intron and the trnL-trnF IGS of chloroplast genome among popular plants in Taiwan. Forensic Sci. Int. 164: 193-200.
White TJ, Bruns T, Lee S and Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In Innis MA et al. (eds.), PCR Protocols: a guide to methods and applications. Academic Press, Inc. California USA., pp. 315-322.
Wang YJ, Liu JQ and Miehe G. 2007. Phylogenetic origins of the Himalayan endemic Dolomiaea,
Diplazoptilon and Xanthopappus (Asteraceae:
Cardueae) based on three DNA regions. Ann. Bot. 99: 311-322.
Wright S. 1951. The genetical structure of populations. Ann. Eugenics. 15: 323-354. Yang HL, Chen SC, Chang NW, Chang JM, Lee
ML, Tsai PC, Fu HH, Kao WW, Chiang HC, Wang HH and Hseu YC. 2006. Protection from oxidative damage using Bidens pilosa extracts in normal human erythrocytes. Food Chem. Toxicol. 44: 1513-1521.
Zouros E, 1979 Mutation rates, population sizes and amounts of electrophoretic variation of enzyme loci in natural populations. Genetics 92: 623-646.
The Genetic Diversity of Bidens pilosa L. in Taiwan Analyzed by
Chloroplast Noncoding and Nuclear rDNA Sequences
Li-Chin Tsai
1,2, Pei-Chun Liao
1, Hsing-Mei Hsieh
2, James Chun-I Lee
3, Jenn-Che Wang
1*1
Department of Life Science, National Taiwan Normal University Taipei, Taiwan
2
Department of Forensic Science, Central Police University Taoyuan, Taiwan
3
Department of Forensic Medicine, National Taiwan University Taipei, Taiwan
(Received: 12 April 2007, accepted: 9 May 2007)
ABSTRACT
According to the Flora of Taiwan, there are three varieties of Bidens pilosa L. They are var. radiata, var. minor and var. pilosa. To discover the genetic diversity and gene flow in different hierarchy (inter-varieties, intra-varieties, inter-populations and intra-populations) in these varieties, this study investigated 161 samples from these varieties comprising 11 different populations and 7 samples of
Bidens biternata for out-group control. The internal transcribed spacer (ITS1 and ITS2) and the 5.8S of
nuclear ribosomal DNA (nrDNA), trnL intron and trnL-trnF IGS loci of the chloroplast genome were amplified and sequenced in all samples. For further analysis, the sequences of ITS1, ITS2 and 5.8S were combined and aligned as nrDNA sequence which covered 637 bp. And the sequences of trnL intron and
trnL-trnF IGS were combined as cpDNA with 799 bp alignment. It showed that the genetic diversity of
nrDNA sequence was higher than that of cpDNA from the data of the number of polymorphic sites (S), number of haplotypes (H), gene diversity (Hd), nucleotide diversity (π) and genetic diversity (θ). The genetic distances (FST) of intra-populations and intra-varieties was with the following order, var. radiata < var. minor < var. pilosa, and the gene flow (Nm) has shown var. radiata > var. minor > var. pilosa. From the results of the analysis of molecular variance (AMOVA), there are more than 50 % variance contributed by the variance of “within populations” in the loci of nrDNA and cpDNA. The rest were contributed by the variance of “among varieties” and “among populations within varieties”. The gene flow between var. radiata and var. minor or var. pilosa showed lower than those between var. minor and var. pilosa, it indicated the significant hybridization phenomenon between var. minor and var. pilosa which naturalized earlier than var. radiata.
Key words:genetic diversity, Bidens pilosa L., nrDNA, ITS1, ITS2, 5.8S, cpDNA, trnL intron, trnL-trnF