Lycopene inhibits growth of human colon cancer cells via suppression
of Akt signaling pathway
Feng-Yao Tang*, Chung-Jin Shih and Ya-Chi Sun
Biomedical Science Laboratory, Department of Nutrition, China Medical University, 40402 Taichung, Taiwan
*
Corresponding author: Dr. Feng-Yao Tang
Biomedical Science Laboratory Department of Nutrition
China Medical University 91 Hsueh-Shih Road, Taichung, 40402
Taiwan, Republic of China Telephone: (886-4) 22060643 Facsimile: (886-4) 22062891
Running Title: Anti-proliferation mechanism of lycopene
Financial support: This material is based upon work supported by the National Science Council grant, under agreement NSC 95-2320-B-039-053-MY2 .
Abstract
The aberrant regulation of the phosphoinositide 3-kinase/Akt survival signaling pathway in cancer has prompted significant interest in suppression of this pathway to treat cancer. Previous studies identified an important role for PI3K/Akt in colon cancer progression. Lycopene, a major component in tomato, exhibited potential anti-carcinogenic activity. Consumption of tomato has been associated with reduced risk of several types of human cancer such as lung and stomach cancer. However, the inhibitory mechanisms of lycopene on the proliferation of human colon cancer have not been studied well yet. Thus we investigated the inhibitory effects of lycopene on Akt signaling pathway in human colon cancer HT-29 cells.
Lycopene inhibited cell proliferation in human colon cancer HT-29 cells with IC50 value of 10 M. Lycopene treatment suppressed Akt activation and non-phosphorylated -catenin protein level in human colon cancer cells. Immunocytochemical result indicated that lycopene increased phosphorylated form of -catenin proteins and could lead to ubiquitin mediated degradation of this protein. These effects were also associated with reduced promoter activity and protein expression of cyclin D1. Furthermore, lycopene significantly increased nuclear CDK inhibitor p27kip abundance and inhibited phosphorylation of the retinoblastoma tumor suppressor protein in human colon cancer cells.
In conclusion, lycopene inhibited cell proliferation of human colon cancer cells via suppression of Akt signaling pathways and downstream targeted molecules.
Introduction
Colorectal cancer is one of the leading causes of cancer death in Western countries, including North America. In the United States alone, nearly thousands of deaths are attributed to this cancer annually [Landis et al., 1998]. Animal study showed that consumption of tomato has been associated with reduced malignant lesion in a rodent model [Sengupta et al., 2003;Guttenplan et al., 2001]. Epidemiologic studies reported statically significant inverse associations between tomato consumption and colon cancer risk for men and women [Franceschi et al., 1994;Franceschi et al., 1997;Erhardt et al., 2003;Murtaugh et al., 2004;Terry et al., 2002]. Lycopene, a major component in tomato, exhibited potential anti-carcinogenesis activity in many types of cancer [Amir et al., 1999;Karas et al., 2000;Nahum et al., 2001]. However, lack of the results across numerous studies and unclear action of lycopene may not be able to demonstrate the benefits of lycopene supplementation against human colon cancer cells.
During the carcinogenesis processes, the Akt/protein kinase B (PKB) molecule plays an important role leading to the tumorogenesis of cancer [Morgensztern and McLeod, 2005]. Akt represents a subfamily of the serine/theronine kinase. Under the extracellular stimuli, Akt is activated in a phosphatidylinositol 3-kinase (PI-3K) dependent manner [Morgensztern and McLeod, 2005]. Suppression of Akt signaling pathway could block the development of several types of tumors including colon cancer [Arico et al., 2002;Arico et al., 2001]. One possible mechanism could be mediated by down regulation of cyclin D1 expression since downstream signaling pathways of Akt could affect nuclear translocation of -catenin and uncovered essential roles in the transcriptional control of cyclin D1 which impact on cell growth and cell cycle progression. Increased phosphorylated N-terminal serine (Ser) of -catenin could promote -catenin degradation and suppress cellular -catenin protein level through
proteosomal pathway [Gwak et al., 2006].
Several proteins under the control of Akt are known to control the cell cycle and play crucial roles in cancer development. One notable example is the cyclin-dependent kinase inhibitor (CKI), p27kip, which is an inhibitor of the cell cycle, and thus a candidate tumor suppressor. Increasing level of p27kipcould inhibit activity of CDK4/6 /cyclin D complex and reduce phosphorylation of retinoblastoma protein (Rb). Reduced phosphorylation or hypo-phosphorylation of Rb leads to inactivation of E2F transcription factor and suppression of S-phase cyclin A. Thus, accumulation of nuclear p27kipcould up-regulate the activity of Rb tumor suppressor and blockade proliferation of cancer cells. A decrease in p27kiplevels, due to p27kipprotein degradation occurs in roughly half of carcinoma and correlates with aggressive, high-grade tumors and poor prognosis. However, certain carcinomas of the breast, thyroid, esophagus or colon contain normal levels of p27kip but the protein has shifted location in these cancers. Studies indicated that p27kipmainly resides in the cytoplasm rather than in the nucleus of those malignant cancer cells. It suggested that the activated kinase Akt/PKB disables the nuclear translocalization of p27kipin those malignant cancer cells.
Thus, in the current study, we determined the inhibitory effect of lycopene on Akt signaling pathway and cell cycle progression in human colon cancer HT-29 cells. Materials and methods
Reagents and Antibodies
Lycopene was purchased from Extrasynthese, Genay, France. Anti-phosphorylation Akt polyclonal antibody and anti-p27kipantibody were purchased from R&D Systems, Inc. (Minneapolis, MN). Anti-phosphorylation -catenin monoclonal antibody was purchased from Cell signaling Technology Inc. (Danvers, MA). Anti- phosphorylation Rb polyclonal antibody, anti--catenin and anti- cyclin D1 polyclonal antibody were purchased from Santa Cruz Biotech Inc. (Santa Cruz, CA). Quantum dot 565
secondary antibody was purchased from Invitrogen Inc. (Carlsbad, CA). Anti--actin antibody and wortamannin were purchased from Sigma (St Louis, MO). Human colon cancer cells HT-29 was purchased from American Type Culture Collection (Walkersville, MD). Tetrahydrofuran (THF), containing 0.025% butylated hydroxytoluene as an antioxidant, was purchased from SIGMA-Aldrich (USA). McCoy’smedium and phosphatebuffered saline(PBS)werepurchased from GIBCO. Lycopene was dissolved in THF at a concentration of 10 mM stock solution and stored at -20oC. Immediately before the experiment, the stock solution was added to the cell culture medium, as described previously.
Cell culture
Briefly, HT-29 colon cancer cells were cultured in a 37oC humidified incubator with 5% CO2 and grown to confluency using fetal bovine serum (FBS) supplemented McCoy’smedia.Cells used in different experiments have the similar passage number. McCoy’s medium were supplemented with 10% heat-inactivated fetal bovine serum, 2 mM L-glutamine and 1.5 g/L sodium bicarbonate.
Supplementation with lycopene
HT-29 colon cancer cells were incubated with different concentrations (0, 2, 5, and 10 M) of lycopene for variable time points. For efficient uptake of lycopene by HT-29 colon cancer cells, lycopene was incorporated into FBS for 30 min and mixed with medium. In control groups, cells were incubated with equivalent volume of solvent THF as a carrier vehicle.
Assessment of cell proliferation
MTT assay was conducted to detect the cell proliferation. HT-29 colon cancer cells were seeded in 24- well plates, each well containing 1x 105cells. After 24 h, the culture
medium was replaced by media in which lycopene concentrations were 0, 2, 5 and 10 µM respectively. There were triplicate for each concentration. From 1 to 3 days, one of the plates was taken out and fresh 3-[4,5-dimethhylthiaoly]-2,5-diphenyltetrazolium bromide (MTT, final concentration 0.5 mg/mL in PBS) was added to each well. After 2 hr incubation, the culture media were discarded, 200 µL of acidic isopropanol was added to each well and vibrated to dissolve the depositor. The optical density was measured at 570 nm with a microplate reader.
Western Blotting Analysis
HT-29 human colon cancer cells were cultured in 10% FBS culture media in the presence or absence of lycopene for various length of time. Cells were lysed in a buffer containing: 1X PBS, 1% Ipegal CA-630 (Sigma, St. Louis, MO), 0.5% sodium deoxycholate, 0.1% SDS with 100 M of phenylmethylsulfonyl fluoride, aprotinin and specific phosphatase inhibitors, sodium orthovanadate. Cell lysates were cleared by centrifugation. Cellular proteins were fractioned on 10% SDS-PAGE, transferred to nitrocellulose membrane, and blotted with anti-phosphorylation Akt polyclonal antibody,according to themanufacturer’sinstructions.Theblotswerestripped and reprobed with-actin antibody as loading control. Protein levels of -catenin, cyclin D1, p27kip, and level of phosphorylated Rb were measured by using the same procedure described above.
Quantum Dot Based Immunofluorescence and Imaging Techniques
Human colon cancer cells,cultured in McCoy’smedium with 10% FBS in atissue culture dish, were lifted off by trypsinization, pelleted by centrifugation, and resuspended in the same medium. Cells were cultured on glass 8-well Tek Chamber and treated with various concentration of lycopene (0, 2, and 10M) for 24 hr. At the end of experiment, cells were fixed in 4% paraformaldehyde in 20 mM of HEPES and 150 mM of NaCl for 20 min, permeabilized in 0.01% Triton X-100 in PBS for 10 min,
blocked with 1% bovine serum albumin (BSA)/PBS for 1h, and then incubated at room temperature for 1 h with anti-phosphorylation -catenin primary antibody at 1:50 in blocking solution. At the end of incubation, cells were washed with PBS and incubated with Quantum dot (Q-dot) 565 secondary antibody for another 1 hr in 1.5% BSA/PBS. Images were acquired on an Olympus BX-51 microscope using the Olympus DP-71 digital camera and imaging system.
Transient transfection and luciferase reporter assays
Cyclin D1 promoter construct –1745 CDLUC and pAL3 basic construct were transiently transfected into HT-29 cells using Lipofectamine (purchased from Invitrogen Inc.). Luciferase reporter assays (Promega) were performed in triplicate asperthemanufacturer’sinstructions.Luciferase fluorescenceactivity was measured in a luminometer (DRL ready TD 20/20).
Statistical analysis
The biostatistic methodology was used to determine whether the difference in the cell viability among experimental sets of colon cancer cells. In brief, statistical analyses of the differences in viability among triplicate sets of experimental conditions were performed using SPSS. Confirmation of difference in cell viability as being statistically significant requires rejection of the null hypothesis of no difference between mean viability indices obtained from replicate sets at the P=0.05 level with the Student t test.
Results
Inhibitory effects of lycopene on the proliferation of colon cancer cells
dose-dependent manner (0, 2, 5 and 10 M). At a concentration of 10 M, lycopene effectively suppressed the proliferation of colon cancer cells up to 47, 57 and 71% for treatment of 24, 48, and 72 hr, respectively (Fig. 1). It suggested that lycopene could effectively suppress the proliferation and survival of human colon cancer HT-29 cells.
Lycopene inhibited consecutive activation of Akt in human colon cancer cells
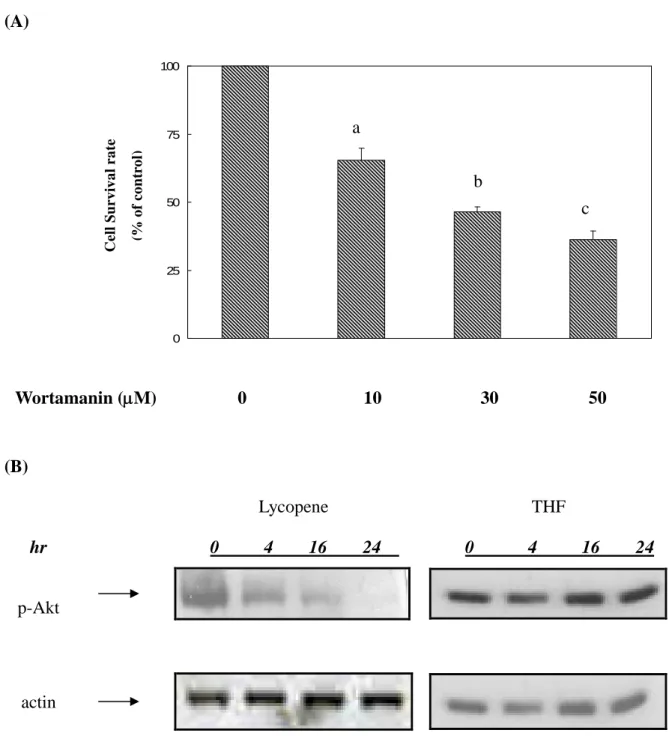
To investigate whether activation of Akt plays an important role in determining survival of human colon cancer cells, we used PI-3K specific inhibitor, wortamannin, to investigate the molecular mechanism of action. In the presence of wortamannin, cell survival rate decreased in a dose-dependent manner (0, 10, 30, 50M) in human colon cancer cells (Fig 2A). It indicated that blockade of Akt activation may suppress the survival of human colon cancer cells.
Thus we examined the inhibitory effect of lycopene on the activation of PI3K/Akt via using western blotting analysis. We found that Akt molecule is consecutively activated in human colon cancer HT-29 cancer cells (Fig 2B). However, lycopene treatment still significantly suppressed the activation of Akt compared to untreated colon cancer HT-29 cells in carrier vector (THF only). Thus, it suggested that lycopene could interrupt PI3K/Akt signaling pathway and inhibit cell survival in human colon cancer HT-29 cells.
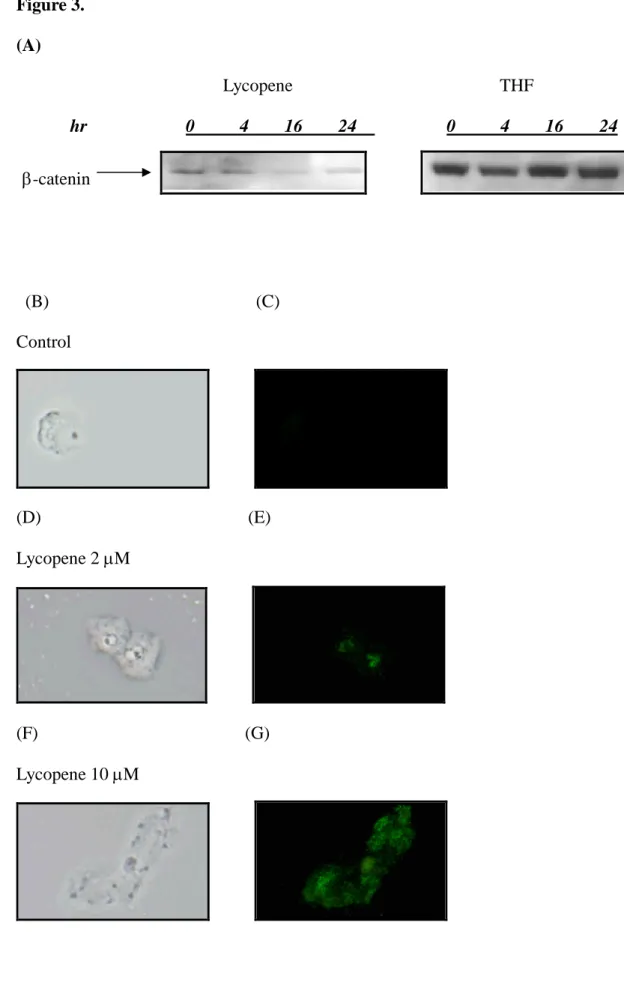
Lycopene modulated the cytoplasmic levels of-catenin
To investigate whether lycopene affected the Akt/-catenin pathways, we determined the level of -catenin molecule via measuring the protein level and cytoplasmic localization. As shown in Fig 3A, lycopene inhibited the cytoplasmic levels of -catenin in human colon cancer HT-29 cells. We also further analyzed the phosphorylated -catenin via quantum dot immunocytostaining. As shown in Fig 3 (B-G), lycopene dose-dependently (0, 2, 10 M) enhanced the accumulation of
phosphorylated -catenin. Since serine phosphorylation of -catenin leads to its ubiquitin degradation, it is plausible that lycopene reduced cytoplasmic level of -catenin via increased phosphorylation level of -catenin.
Lycopene suppressed cyclin D1 promoter activity and protein expression in human colon cancer cells
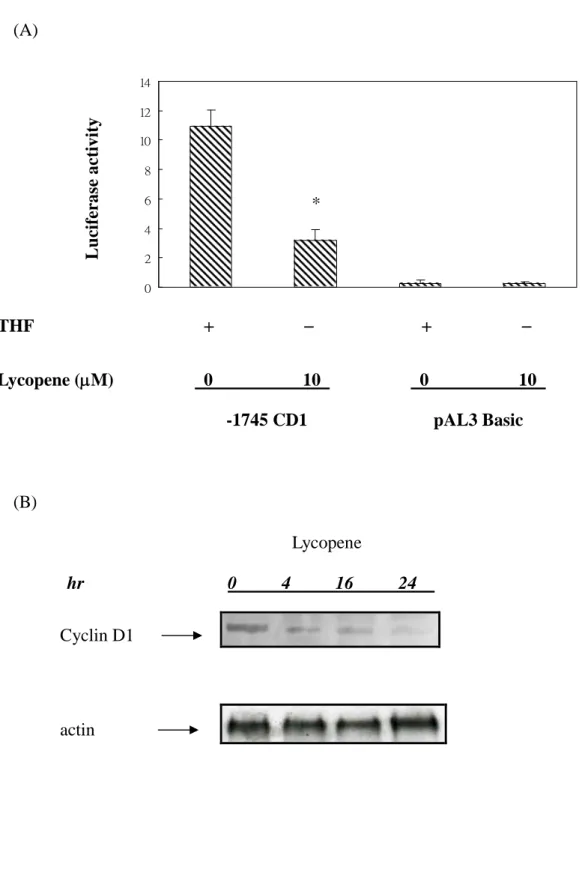
To determine whether lycopene suppressed the gene expression of cyclin D1, we used cyclin D1-Luciferase construct (-1745 CD1) and basal construct (pAL3 basic) to test the possibilities. In Fig 4A, it showed that lycopene inhibited the promoter activity of cyclin D1 gene. No promoter activities were detected in the group of basal construct. Thus, it’s plausible that lycopene could inhibit the promoter activity of cyclin D1. To determine whether lycopene could further block the downstream of Akt signaling pathways, we examine the expression of cyclin D1 proteins. As shown in Fig 4B, expression of cyclin D1 protein is suppressed under the treatment of lycopene in a time-dependent manner. These results suggested that lycopene might suppress Akt signaling pathways and expression of downstream cyclin D1 protein.
Lycopene induced accumulation of nuclear p27Kipprotein and activity of Rb in colon cancer cells
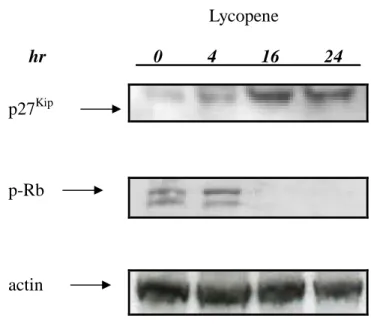
Since PI-3K/Akt has been found to be an important mediator of cell proliferation via p27kip translocation, we tested the effect of lycopene on nuclear localization of p27kip in colon cancer cells. As shown in Fig 5, lycopene significantly induced the nuclear localization of p27kip compared to unstimulated HT-29 cells. Furthermore, lycopene indirectly blocked the phosphorylation of Rb and enhanced its activity. These findings suggest that lycopene could inhibit cell growth via reduced phosphorylation level of Akt and Rb proteins.
Discussion
In this study, we showed that lycopene inhibited the proliferation of colorectal cancer cells in a dose-dependent manner (Fig. 1). At 24 hour time point, at a concentration of 5 M, lycopene suppressed cell proliferation by almost 25 % and at 10 M, almost inhibited cell growth up to 47%. At 72 hour time point, at a concentration of 5 M, lycopene suppressed cell proliferation by almost 30 % and at 10M, almost abolished cell growth up to 71 %. Normal range of human serum lycopene level is around 2-3 M. However, 10 M of human serum lycopene level could be reached by dietary supplementation. These results demonstrated that supplementation of lycopene effectively block the growth and survival of human colorectal cancer cells. Phosphatidylinositol 3-kinase (PI-3K) and Akt have come onto to the scene and are reaching central status as an important regulator of mammalian cell proliferation and survival during tumor development. Several components of the PI3K –Akt pathways are dysregulated in a wide spectrum of human cancers [Kaufmann and Gores, 2000;Vivanco and Sawyers, 2002]. The inhibitory regulatory subunit of PI-3 kinase is mutated in some colon cancer cells[Bates et al., 2001]. Recent studies reported that mutation of regulatory subunit p85- and constitutively activated Akt molecules support the proliferation of colorectal cancer cells [Philp et al., 2001]. Constitutive activation of PI-3K/Akt induces not only tumor angiogenesis but also the progression of tumor development [Sun et al., 2001b;Sun et al., 2001a;Yokoyama et al., 2000]. Increasing evidence implicates inhibition of PI-3K pathways would inhibit the growth of tumor. Thus therapeutic strategies that target the PI3K/Akt pathway are now in development. In the present study, we speculated that lycopene whether could block the active site of Akt and prevent activation of its downstream signaling pathways required for tumor growth.
We found that Akt is consecutively activated in human cancer cells (Fig 2.). However, lycopene treatment effectively suppressed Akt activity. Lycopene significantly suppressed the survival of colon cancer cells via inhibition of Akt kinase activity (Fig 2.). It suggested that lycopene could be an effective compound with anti-cancer activity.
In colorectal carcinoma cells with intact APC genes, -catenin was shown to contribute to the activation of the wingless/ wnt pathway [He et al., 1998;Morin et al., 1997]. Accumulation of -catenin in the nucleus and the uncontrolled activation of target gene expression is believed to contribute to tumor progression. Previous studies suggested cyclin D1 gene as a target for the -catenin/LEF-1 complex. Increased elevated -catenin levels in colon cancer cells result in the activation of cyclin D1 gene promoter by the heterodimeric complex formed between -catenin and LEF-1, which, in turn, results in the elevation of cyclin D1 gene expression, protein level, and cell cycle progression [Shtutman et al., 1999]. These data indicate that -catenin molecules may mediate the activation of the wnt signaling pathway in up to 80% of human colorectal carcinomas. It suggested, therefore, that activation of this pathway is the main initial event in colorectal tumorigenesis [Bodmer, 1997;Peifer and Polakis, 2000;Peifer, 1996;Peifer, 1997]. Recently, cyclin D1 was identified as a target of the -catenin pathway in HT29 cells [He et al., 1998]. However, the effects of lycopene on genetic targets of this signaling pathway and their contribution to the neoplastic process have not yet been characterized. The purpose of the present work is to investigate whether lycopene inhibits cell cycle through suppression of the target genes in the development of human colorectal carcinomas. Our results showed that lycopene inhibited protein level of-catenin in human colorectal cancer HT-29 cells. It suggested that lycopene might block the Akt signaling pathways and protein level of downstream -catenin molecule (Fig 3.). Recent study indicated that increased
phosphorylated level of-catenin is correlated with downregulation of Wnt/-catenin pathway due to -catenin proteosomal degradation [Gwak et al., 2006]. To further investigate the effects of lycopene on the regulation of -catenin, we used immunocytostaining assay conjugated with quantum dot techniques to identify the cytoplasmic localization of phosphorylated -catenin protein. . As shown in Fig 3 B-G, lycopene treatment dose dependently enhanced the cytoplasmic accumulation of phosphorylated -catenin. We further investigated the effects of lycopene on the promoter activity of cyclin D1. Results showed that lycopene significantly inhibited promoter activity and protein expression of cyclin D1 (Fig 4.). These results suggested that lycopene could block cell cycle progression via modulation of cyclin D1.
A decrease in p27Kiplevels, due to p27Kipprotein degradation occurs in roughly half of carcinoma and correlates with aggressive, high-grade tumors and poor prognosis. Certain carcinomas of the breast, thyroid, esophagus or colon contain normal levels of p27Kipbut, strangely, the protein has shifted location in these cancers. Normally p27Kip resides in the nucleus, but in these tumors it can be found in the cytoplasm. During the translocation of p27Kip, the activated kinase Akt/PKB disables the nuclear localization capacity of p27 Kip. Thus barred from the nucleus, p27Kip can no longer rein in cell division, providing the tumor cell with a growth advantage and contributing to genome instability. Similar to the loss of p27 Kip, this accumulation of cytoplasmic, Akt-phosphorylated p27 Kip correlated with tumor aggressiveness. Collectively, the current evidence indicates that Akt activation is an important component in the cell proliferation. Recent studies have demonstrated that the signals of tyrosine phosphorylation in Akt complex markedly enhanced the phosphorylation of p27Kip and prevented its nucleus translocation [Shin et al., 2002;Viglietto et al., 2002].
Thus, it is conceivable that the changes in tyrosine phosphorylation of Akt are associated with the changes in the cell cycle. To further examine other possible mechanism of actions, we also investigated the effects of lycopene on cell cycle modulators. While p27 Kip through dephosphorylation signaling would induce a pleiotropic response allowing cell cycle arrest, our data indicate that, lycopene induced this process leading to the inhibition of cell growth. It suggested that abundance of nuclear p27Kipand activity of Rb were increased under the treatment of lycopene.
These results indicated that the blockade of Akt activation by lycopene treatment is the major modulator for their effect on cell proliferation. In the present study, we demonstrated for the first time that lycopene inhibited the tyrosine phosphorylation of Akt and-catenin protein level in human colon cancer HT-29 cells. We also demonstrated that lycopene enhanced phosphorylation level of -catenin protein and possibly lead to proteosomal degradation. To examine the exact mechanism of this inhibition, we speculate this inhibitory effect to be evolved with lycopene, in a possible model of signal transduction via the cyclin changes. As previously suggested, cyclin D1 is reported to be important for progression of cell cycle, therefore changed levels of cyclins are likely a consequence of a cascading signal through Akt signaling pathways. In this study, we demonstrated that lycopene may specifically inhibit PI-3K/Akt tyrosine phosphorylation cascade during tumor progression and suppress cyclin D1 promoter activity and protein level. Furthermore, lycopene increased nuclear p27kip protein but suppressed the phosphorylation of Rb protein. Increased protein level of p27kip and activity of Rb leads to the blocking of cell cycle progression.
antitumorgenic effect, is in part through inhibition of Akt phosphorylation as needed for cell cycle arrest during tumor growth. These findings provide a novel mechanistic insight into the inhibitory effects of lycopene on the growth of human colorectal cancer. Although we only test the effects of lycopene on human colon cancer HT-29 cell lines, our in vitro results are also consistent with the concept that consumption of tomato, in which lycopene is the major polyphenol, is associated with reduced risk of cancer development as evident from population studies and animal experiments showing suppression of tumor progression and cancer development.
Acknowledgements
Financial support: This material is based upon work supported by the National Science Council grant, under agreement NSC 95-2320-B-039-053-MY2. Any
opinions, findings, conclusions, or recommendations expressed in this publication are those of the author(s) and do not necessarily reflect the view of the National Science Council and China Medical University.
Figure Legends
Figure 1.Inhibitory effects of lycopene on the proliferation of colon cancer cells
Human colon cancercells,cultured in McCoy’smedium with 10% FBS in atissue culture dish, were lifted off by trypsinization, pelleted by centrifugation, and resuspended in the same medium.
Approximately 30,000 cells were seeded on each well of 24-well plate. The human colon cancercellswerethen cultured in lycopene atin McCoy’smedium with or without different concentration (0, 2, 5, 10 M) of lycopene for 24, 48, and 72 h until measurement of cell proliferation. The incubation was stopped at different time points and measured with MTT assays for cell proliferation. The analysis of cell proliferation was described in Materials and Methods. Data analysis from three separate experiments is shown as the MeanS.E.M (standard error of mean). Similar results were observed from three independent experiments. Different letters represent statistically significant difference, p<0.05.
Figure 2.
Lycopene inhibited consecutive activation of Akt in human colon cancer cells
Human colon cancercells,cultured in McCoy’smedium with 10% FBS in atissue culture dish, were lifted off by trypsinization, pelleted by centrifugation, and resuspended in the same medium.
(A) After washing out the media, human colon cancer cells were incubated in McCoy’smedium with 10% FBS in atissueculturedishwith various concentration (0, 10, 30, 50 M) of wortamanin for 24 hr. Different letters represent statistically significant difference, p<0.05.
(B) Total cell lysates were blotted with anti- phosphorylated -Akt antibody as described in Materials and Methods. The levels of detection in cell lysates represent the amount of phosphorylated- Akt in human colon cancer cells. The blots were stripped and reprobed with anti-actin antibody as loading control. The immunoreactive bands are noted with arrow.
Figure 3.
Lycopene modulated the cytoplasmic levels of-catenin
(A) Human colon cancer cells supplemented with various concentration (0, 10 M) of lycopene were cultured in McCoy’smedium with 10% FBS in a10 cm tissue culture dish. Total cell lysates were blotted with anti--catenin antibody as described in Materials and Methods. The levels of detection in cell lysates represent the amount of -catenin in human colon cancer cells. The blots were stripped and reprobed with anti-actin antibody as loading control. The immunoreactive bands are noted with arrow.
(B)-(G) Human colon cancer cells were incubated in McCoy’smedium with 10% FBS in an 8-well Tek Chamber with various concentration (0, 2, 10 M) of lycopene for 24 hrs. At the end of incubation, cells were blotted with anti-phosphorylation -catenin primary antibody and quantum dot 565 secondary antibody as described in Materials and Methods. The levels of detection in cell represent the amount and localization of phosphorylation-catenin in human colon cancer cells. Images (1000x) were acquired using Olympus DX-51 fluorescent stereomicroscope and DP-71 imaging system. (B),(D), (F) represent images taken in light fields. (C), (E), (G) represent images taken in dark fields.
Figure 4.
Lycopene suppressed cyclin D1 promoter activity and protein expression in human colon cancer cells.
(A) Ninety percent confluent human colon cancer HT-29 cells were treated with lycopene for different time points. The cyclin D1 luciferase reporter construct (1 g) and pAL3 basic construct were transiently transfected into HT-29 cells, respectively. Triplicate experiments were performed at 24 h after treatment with or without lycopene (10M). The experiment was performed for three times; results were highly repeatable. *p<0.05 compared with control.
(B) Human colon cancercells,cultured in McCoy’smedium with 10% FBS in atissue culture dish, were lifted off by trypsinization, pelleted by centrifugation, and resuspended in the same medium. After washing out the media, human colon cancer cells were incubated in McCoy’smedium with 10% FBS in atissueculturedishwith 10 M of lycopene for different time points (0, 4, 16, 24 hrs). Total nuclear cell lysates were blotted with anti-cyclin D1 antibody as described in Materials and Methods. The levels of detection in nuclear cell lysates represent the amount of cyclin D1 in human colon cancer cells. The blots were stripped and reprobed with anti-actin polyclonal antibody as loading control. Each experiment was repeated three times with very similar results in repeat trials. The immunoreactive bands are noted with arrow.
Figure 5.
Lycopene induced accumulation of nuclear p27Kip protein and activity of Rb in
colon cancer cells
Confluent human colon cancer cells were incubated with 10 M of lycopene at 37oC for different time points (0, 4, 16, 24 hrs). After washing out the media, cytoplasma cell lysates and nuclear extracts of human colon cancer cells were prepared as described in the Materials and Methods section. (A) Total nuclear cell lysates were blotted with anti-p27kip antibody as described in Materials and Methods. The levels of detection in nuclear cell lysate represent the amount of p27kip in human colon cancer cells. The levels of detection in nuclear extracts represent the amount of p27Kip in human colon cancer cells. The immunoreactive bands are noted with arrow. (B) Total nuclear cell lysates were blotted with anti- phosphorylated-Rb antibody as described in Materials and Methods. The levels of detection in nuclear cell lysate represent the amount of tyrosine phosphorylated-Rb in human colon cancer cells. The immunoreactive bands are noted with arrow.
Figure 1. 0 25 50 75 100 0 uM 2 uM 5 uM 10 uM 24 48 72 (Hr) c f h e g C e ll S u r v iv a l r a te (% o f c o n tr o l) a b i d
Figure 2. (A) 0 25 50 75 100 (B) Lycopene THF hr 0 4 16 24 0 4 16 24 p-Akt actin Wortamanin (M) 0 10 30 50 C e ll S u r v iv a l r a te (% o f c o n tr o l) a b c
Figure 3. (A) Lycopene THF hr 0 4 16 24 0 4 16 24 (B) (C) Control (D) (E) Lycopene 2M (F) (G) Lycopene 10M -catenin
Figure 4. (A) 0 2 4 6 8 10 12 14 (B) Lycopene hr 0 4 16 24 THF Lycopene (M) 0 10 0 10 L u ci fe ra se a c ti v it y -1745 CD1 pAL3 Basic * Cyclin D1 actin
Figure 5. Lycopene hr 0 4 16 24 p-Rb p27Kip actin
References
Amir H, Karas M, Giat J, Danilenko M, Levy R, Yermiahu T, Levy J, Sharoni Y (1999) Lycopene and 1,25-dihydroxyvitamin D3 cooperate in the inhibition of cell cycle progression and induction of differentiation in HL-60 leukemic cells. Nutr Cancer 33, 105-112.
Arico S, Pattingre S, Bauvy C, Gane P, Barbat A, Codogno P, Ogier-Denis E (2002) Celecoxib induces apoptosis by inhibiting 3-phosphoinositide-dependent protein kinase-1 activity in the human colon cancer HT-29 cell line. J Biol Chem 277, 27613-27621.
Arico S, Petiot A, Bauvy C, Dubbelhuis PF, Meijer AJ, Codogno P, Ogier-Denis E (2001) The tumor suppressor PTEN positively regulates macroautophagy by inhibiting the phosphatidylinositol 3-kinase/protein kinase B pathway. J Biol Chem 276, 35243-35246.
Bates RC, Edwards NS, Burns GF, Fisher DE (2001) A CD44 survival pathway triggers chemoresistance via lyn kinase and phosphoinositide 3-kinase/Akt in colon carcinoma cells. Cancer Res 61, 5275-5283.
Bodmer W (1997) The somatic evolution of cancer. The Harveian Oration of 1996. J
R Coll Physicians Lond 31, 82-89.
Erhardt JG, Meisner C, Bode JC, Bode C (2003) Lycopene, beta-carotene, and colorectal adenomas. Am J Clin Nutr 78, 1219-1224.
Franceschi S, Bidoli E, La Vecchia C, Talamini R, D'Avanzo B, Negri E (1994) Tomatoes and risk of digestive-tract cancers. Int J Cancer 59, 181-184.
Franceschi S, Favero A, La Vecchia C, Negri E, Conti E, Montella M, Giacosa A, Nanni O, Decarli A (1997) Food groups and risk of colorectal cancer in Italy. Int J
Cancer 72, 56-61.
Guttenplan JB, Chen M, Kosinska W, Thompson S, Zhao Z, Cohen LA (2001) Effects of a lycopene-rich diet on spontaneous and benzo[a]pyrene-induced mutagenesis in prostate, colon and lungs of the lacZ mouse. Cancer Lett 164, 1-6.
Gwak J, Cho M, Gong SJ, Won J, Kim DE, Kim EY, Lee SS, Kim M, Kim TK, Shin JG, Oh S (2006) Protein-kinase-C-mediated beta-catenin phosphorylation negatively regulates the Wnt/beta-catenin pathway. J Cell Sci 119, 4702-4709.
He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT, Morin PJ, Vogelstein B, Kinzler KW (1998) Identification of c-MYC as a target of the APC pathway. Science 281, 1509-1512.
Karas M, Amir H, Fishman D, Danilenko M, Segal S, Nahum A, Koifmann A, Giat Y, Levy J, Sharoni Y (2000) Lycopene interferes with cell cycle progression and
insulin-like growth factor I signaling in mammary cancer cells. Nutr Cancer 36, 101-111.
Kaufmann SH, Gores GJ (2000) Apoptosis in cancer: cause and cure. Bioessays 22, 1007-1017.
Landis SH, Murray T, Bolden S, Wingo PA (1998) Cancer statistics, 1998. CA Cancer
J Clin 48, 6-29.
Morgensztern D, McLeod HL (2005) PI3K/Akt/mTOR pathway as a target for cancer therapy. Anticancer Drugs 16, 797-803.
Morin PJ, Sparks AB, Korinek V, Barker N, Clevers H, Vogelstein B, Kinzler KW (1997) Activation of beta-catenin-Tcf signaling in colon cancer by mutations in beta-catenin or APC. Science 275, 1787-1790.
Murtaugh MA, Ma KN, Benson J, Curtin K, Caan B, Slattery ML (2004) Antioxidants, carotenoids, and risk of rectal cancer. Am J Epidemiol 159, 32-41.
Nahum A, Hirsch K, Danilenko M, Watts CK, Prall OW, Levy J, Sharoni Y (2001) Lycopene inhibition of cell cycle progression in breast and endometrial cancer cells is associated with reduction in cyclin D levels and retention of p27(Kip1) in the cyclin E-cdk2 complexes. Oncogene 20, 3428-3436.
Peifer M (1996) Regulating cell proliferation: as easy as APC. Science 272, 974-975. Peifer M (1997) Beta-catenin as oncogene: the smoking gun. Science 275, 1752-1753. Peifer M, Polakis P (2000) Wnt signaling in oncogenesis and embryogenesis--a look outside the nucleus. Science 287, 1606-1609.
Philp AJ, Campbell IG, Leet C, Vincan E, Rockman SP, Whitehead RH, Thomas RJ, Phillips WA (2001) The phosphatidylinositol 3'-kinase p85alpha gene is an oncogene in human ovarian and colon tumors. Cancer Res 61, 7426-7429.
azoxymethane-induced colon carcinogenesis in rats. Eur J Cancer Prev 12, 195-200. Shin I, Yakes FM, Rojo F, Shin NY, Bakin AV, Baselga J, Arteaga CL (2002) PKB/Akt mediates cell-cycle progression by phosphorylation of p27(Kip1) at threonine 157 and modulation of its cellular localization. Nat Med 8, 1145-1152.
Shtutman M, Zhurinsky J, Simcha I, Albanese C, D'Amico M, Pestell R, Ben Ze'ev A (1999) The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl
Acad Sci U S A 96, 5522-5527.
Sun M, Paciga JE, Feldman RI, Yuan Z, Coppola D, Lu YY, Shelley SA, Nicosia SV, Cheng JQ (2001a) Phosphatidylinositol-3-OH Kinase (PI3K)/AKT2, activated in breast cancer, regulates and is induced by estrogen receptor alpha (ERalpha) via interaction between ERalpha and PI3K. Cancer Res 61, 5985-5991.
Sun M, Wang G, Paciga JE, Feldman RI, Yuan ZQ, Ma XL, Shelley SA, Jove R, Tsichlis PN, Nicosia SV, Cheng JQ (2001b) AKT1/PKBalpha kinase is frequently elevated in human cancers and its constitutive activation is required for oncogenic transformation in NIH3T3 cells. Am J Pathol 159, 431-437.
Terry P, Jain M, Miller AB, Howe GR, Rohan TE (2002) Dietary carotenoid intake and colorectal cancer risk. Nutr Cancer 42, 167-172.
Viglietto G, Motti ML, Bruni P, Melillo RM, D'Alessio A, Califano D, Vinci F, Chiappetta G, Tsichlis P, Bellacosa A, Fusco A, Santoro M (2002) Cytoplasmic relocalization and inhibition of the cyclin-dependent kinase inhibitor p27(Kip1) by PKB/Akt-mediated phosphorylation in breast cancer. Nat Med 8, 1136-1144. Vivanco I, Sawyers CL (2002) The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev Cancer 2, 489-501.
Yokoyama Y, Wan X, Shinohara A, Takahashi S, Takahashi Y, Niwa K, Tamaya T (2000) Expression of PTEN and PTEN pseudogene in endometrial carcinoma. Int J