建立驗證調節脂肪酸代謝之細胞模式與改善動物代謝症候群模式
133
0
0
全文
(2) 目錄. 縮寫對照表……………………………………..…………….……………………….Ⅰ Ⅰ 縮寫對照表 Ⅲ 組別縮寫對照 縮寫對照 表…………………..……………..……….…………………………….Ⅲ 組別 Ⅳ 摘要……………………………………..……………………..……….……………….Ⅳ. Abstract……………………………………..……………….……..………………….Ⅵ Ⅵ 第一章 前言……………………………………..…………………………………….1 第二章 文獻回顧………………………………….………………………………….2 一、代謝症候群……………………………………….…………………………………2 代謝症候群 (一)代謝症候群定義…………………………………………………………………..2 代謝症候群定義 (二)判定標準………………..………………................................................................2 判定標準 (三)代謝症候群發生原因……………………………………………………………..6 代謝症候群發生原因 (四)代謝症候群治療方式………………………………………….………………….7 代謝症候群治療方式 二、乙醯輔酶羧 A 化酶……………………………………………….…………………8 化酶 (一)何謂 Acetyl-CoenzymeA carboxylase……………….…………………………..8 (二)ACC 調控………………………………………………….…………………….10 調控 1. 異位調節…………………………………………………..….…………………10 異位調節 2. 共價修飾………………………………………………………………..……….11 共價修飾 (三)抑制 ACC 可改善代謝症候群之相關研究………………….…………………12 可改善代謝症候群之相關研究 三、高糖飲食誘發代謝症候群 高糖飲食誘發代謝症候群…………………………………………….…………..13 飲食誘發代謝症候群 (一)高糖( 高糖(蔗糖) 蔗糖)飲食引發代謝症候群之動物模式…………………..………….13 飲食引發代謝症候群之動物模式 (二)高糖誘發肝脂堆積及胰島素阻抗之細胞模式……………...…………………14 高糖誘發肝脂堆積及胰島素阻抗之細胞模式 (三)肝臟中脂質生合成相關轉錄因子……………………….……………………..15 肝臟中脂質生合成相關轉錄因子 1. Liver X receptors……………………………………………………………….15.
(3) 2. Carbohydrate responsive element binding protein………….………………..16 3. Sterol regulatory element binding protein-1c…………………………………17 (四)蔗糖誘發脂質新生( )之生化與分子機制………………19 蔗糖誘發脂質新生(de novo lipogenesis) 之生化與分子機制 (五)Microsomal triglyceride transfer protein 與三酸甘油酯分泌……….……….21 與三酸甘油酯分泌 四、紅雞屎藤……………………………………………………………………………23 紅雞屎藤. 第三章 材料與方法…………………………………………………………………25 實驗一……………………………………………………………………………………27 實驗一 實驗二……………………………………………………………………………………28 實驗二 (一)細胞培養…………………………………………………………………………30 細胞培養 (二)ACC 活性分析…………………………………………………………………..30 活性分析 (三)FAS 活性分析……………………………………………………………………34 活性分析 (四)培養基中 ApoB 含量測定………………………………………………………39 含量測定 (五)細胞內 MTTP mRNA 表現量分析…………………………………………….39 表現量分析 (六)動物飼養…………………………………………………………………………39 動物飼養 (七)動物犧牲與樣品收集……………………………………………………………39 動物犧牲與樣品收集 (八)三酸甘油酯含量測定……………………………………………………………40 三酸甘油酯含量測定 (九)膽固醇含量測定…………………………………………………………………40 膽固醇含量測定 (十)肝臟、 肝臟、肌肉脂質均質液製備……………………………………………………41 肌肉脂質均質液製備 (十一) 十一)肝臟、 肝臟、肌肉三酸甘油酯含量測定……………………………………………41 肌肉三酸甘油酯含量測定 (十二) 十二)肝臟、 肝臟、肌肉膽固醇含量測定…………………………………………………41 肌肉膽固醇含量測定 (十三) 十三)全血葡萄糖含量測定…………………………………………………………42 全血葡萄糖含量測定 (十四) 十四)口服葡萄糖耐受測試 Oral Glucose Tolerance Test (OGTT) ……………..42 (十五) 十五)胰島素耐受測試 Insulin Tolerance Test (ITT) …………………………….43 (十六) 十六)禁食血清胰島素含量測定……………………………………………………43 禁食血清胰島素含量測定 (十七) 十七)禁食血清瘦素含量測定……………………………………………………….44 禁食血清瘦素含量測定.
(4) (十八) 十八)尿酸測定………………………………………………………………………46 尿酸測定 (十九) blot)分析肝臟 分析肝臟、 十九)西方點墨法(Western 西方點墨法 分析肝臟、肌肉和脂肪組織 PTP1B 蛋白質表現…………………………………………………………..46 蛋白質表現 (二十) 二十)脂肪組織切片…………………………………………………………………47 脂肪組織切片 (二十一) 二十一)脂肪組織中 TNF-α 量測定………………………………………………..48 量測定 (二十二) 二十二)大鼠血壓測定………………………………………………………………49 大鼠血壓測定 (二十三) 二十三)統計分析……………………………………………………………………49 統計分析. 第四章 結果…………………………………………………………………………..50 第五章 討論…………………………………………………………………………..84 第六章 結論…………………………………………………………………………..97 第七章 參考文獻…………………………………………………………………….99.
(5) 表 目 錄 表 2–1 台灣代謝症候群 台灣代謝症候群之判定標準 代謝症候群之判定標準…………………………………………………...4 之判定標準 表 2–2 台灣代謝症候群及組成因子盛行率…………………………………………...5 台灣代謝症候群及組成因子盛行率 表 2-3. 95 年國人代謝症候群相關疾病醫療費用……………………………………..6 年國人代謝症候群相關疾病醫療費用. 表 4-1. Wistar 大鼠餵食紅雞屎藤酒精萃四週之體重及白色脂肪量……………….59 大鼠餵食紅雞屎藤酒精萃四週之體重及白色脂肪量. 表 4-2. Wistar 大鼠餵食紅雞屎藤酒精萃四週之血液生化質……………………….60 大鼠餵食紅雞屎藤酒精萃四週之血液生化質. 表 5-1. 大鼠給予紅雞屎藤 4 週後之結果統整…………………………….………….93 週後之結果統整.
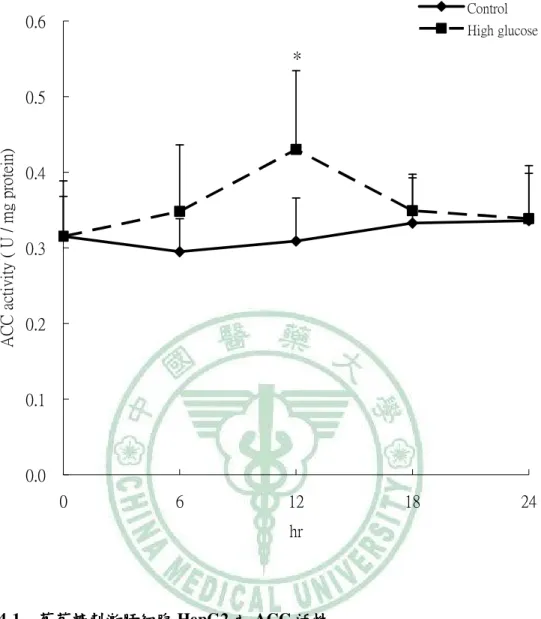
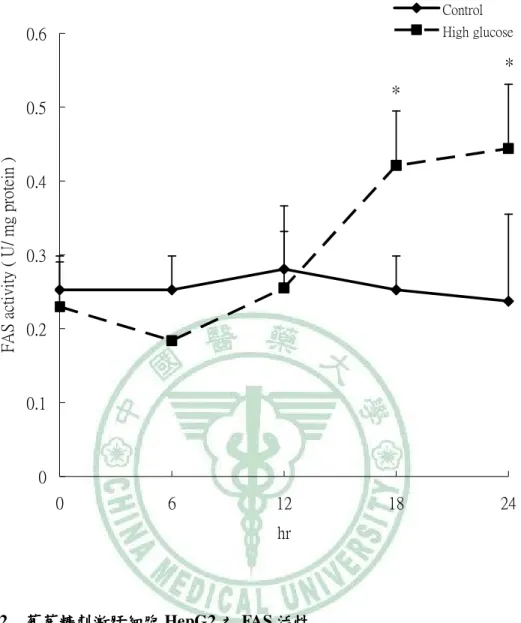
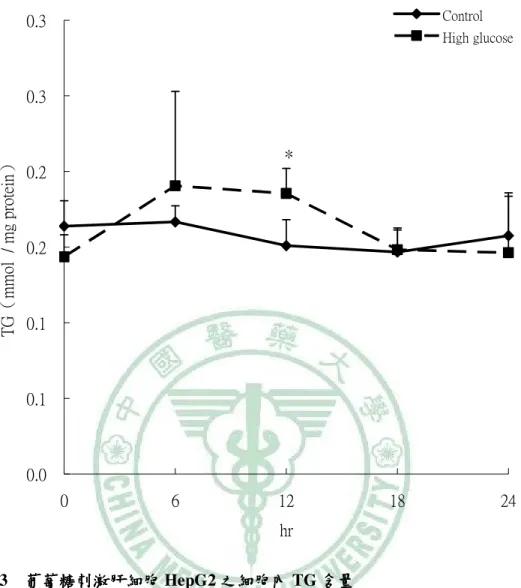
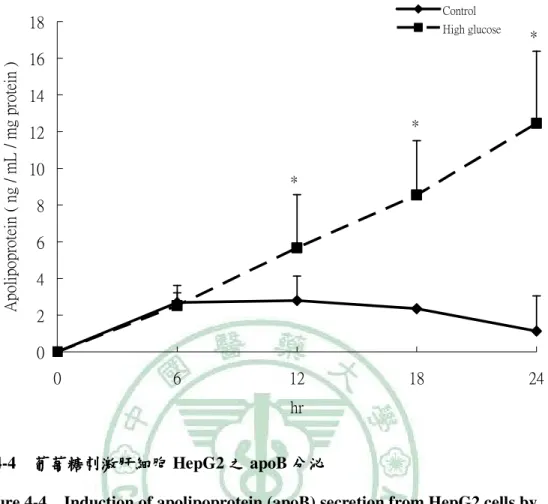
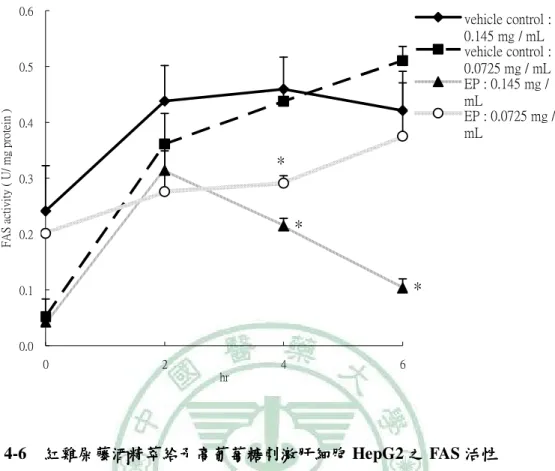
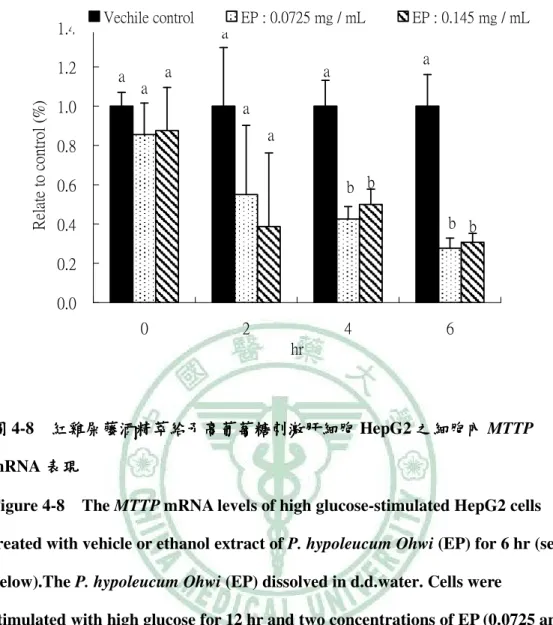
(6) 圖 目 錄 圖2-1. 參與ACC催化反應的兩個步驟 催化反應的兩個步驟…………………………………………………9 參與 催化反應的兩個步驟. 圖2-2 TOFA 結構…………………………………………………………………..…13 結構 圖2-3 T0901317之結構圖 之結構圖……………………………………………………………..16 之結構圖 圖2-4 GW3965之結構圖 之結構圖……………………………………………………………...16 之結構圖 圖3-1. 本論文實驗研究大綱 本論文實驗研究大綱…………………………………………………………..26 大綱. 圖3-2. 細胞給予红雞屎藤酒精萃之實驗流程………………………………………..27 細胞給予红雞屎藤酒精萃之實驗流程. 圖3-3. 大鼠實驗流程…………………………………………………………………..29 大鼠實驗流程. 圖 4-1 葡萄糖刺激肝細胞 HepG2 之 ACC 活性…………………………………….61 活性 圖 4-2. 葡萄糖刺激肝細胞 HepG2 之 FAS 活性……………………………………..62 活性. 圖 4-3. 葡萄糖刺激肝細胞 HepG2 之細胞內 TG 含量……………………………….63 含量. 圖 4-4 葡萄糖刺激肝細胞 HepG2 之 apoB 分泌…………………………………….64 分泌 圖 4-5. 紅雞屎藤酒精萃給予高葡萄糖刺激肝細胞 HepG2 之 ACC 活性………….65 活性. 圖 4-6. 紅雞屎藤酒精萃給予高葡萄糖刺激肝細胞 HepG2 之 FAS 活性…………..66 活性. 圖 4-7 紅雞屎藤酒精萃給予高葡萄糖刺激肝細胞 HepG2 之細胞內 TG 含量……67 含量 圖 4-8. 紅雞屎藤酒精萃給予高葡萄糖刺激肝細胞 HepG2 之細胞內 MTTP mRNA 表現……………………………………………………………………………..68 表現. 圖 4-9. Wistar 大鼠餵食紅雞屎藤酒精萃四週之收縮壓和舒張壓 大鼠餵食紅雞屎藤酒精萃四週之收縮壓和舒張壓………………….69 四週之收縮壓和舒張壓. 圖 4-10. Wistar 大鼠餵食紅雞屎藤酒精萃四週之葡萄糖耐量試驗( )和葡萄糖曲 大鼠餵食紅雞屎藤酒精萃四週之葡萄糖耐量試驗(A) 線下面積( )…………………………………………………………………70 線下面積(B). 圖 4-11 Wistar 大鼠餵食紅雞屎藤酒精萃四週之胰島素耐量試驗( )和葡萄糖曲 大鼠餵食紅雞屎藤酒精萃四週之胰島素耐量試驗(A) 線下面積( )…………………………………………………………………71 線下面積(B) 圖 4-12. Wistar 大鼠餵食紅雞屎藤酒精萃四週之肝臟及肌肉三酸甘油酯含量……72 大鼠餵食紅雞屎藤酒精萃四週之肝臟及肌肉三酸甘油酯含量. 圖 4-13. Wistar 大鼠餵食紅雞屎藤酒精萃四週之肝臟中膽固醇含量………………73 大鼠餵食紅雞屎藤酒精萃四週之肝臟中膽固醇含量.
(7) 圖 4-14. Wistar 大鼠餵食紅雞屎藤酒精萃四週之腹膜後脂肪組織切片與脂肪細胞平 均直徑…………………………………………………………………………..74 均直徑. 圖 4-15 Wistar 大鼠餵食紅雞屎藤酒精萃四週之禁食血清 Leptin…………………75 圖 4-16 Wistar 大鼠餵食紅雞屎藤酒精萃四週之腹睪脂組織中 TNF-α 含量……..76 含量 圖 4-17. Wistar 大鼠餵食紅雞屎藤酒精萃四週之肝臟 ACC 活性………………….77 活性. 圖 4-18 Wistar 大鼠餵食紅雞屎藤酒精萃四週之肝臟 FAS 活性…………………..78 活性 圖 4-19. 以 Western blot 分析大鼠餵食蔗糖水 5 個月之肝臟 PTP1B 蛋白質表現量.79 蛋白質表現量. 圖 4-20. 以 Western blot 分析大鼠餵食蔗糖水 5 個月之肌肉 PTP1B 蛋白質表現量.80 蛋白質表現量. 圖 4-21. 以 Western blot 分析大鼠餵食蔗糖水 5 個月之脂肪組織 PTP1B 蛋白質表 現量……………………………………………………………………………..81 現量. 圖 4-22. Wistar 大鼠餵食紅雞屎藤酒精萃 2 小時後之肝臟 ACC 活性……………82 活性. 圖 4-23. Wistar 大鼠餵食紅雞屎藤酒精萃 大鼠餵食紅雞屎藤酒精萃 2 小時後之肝臟 FAS 活性…………….83 活性. 圖 5-1. 糖水模式誘發代謝症候群之可能分子機制………………………………….88 糖水模式誘發代謝症候群之可能分子機制.
(8) 縮寫對照表 縮. 寫. 全. ACC. 名. Acetyl - CoA carboxylase. AMPK. AMP - activated protein kinase. ABC. ATP-binding cassette transporters. ApoB48. Apolipoprtein B48. ApoB100. Apolipoprtein B100. bHLH. basic helix-loop-helix. BC. Biotin carboxylase. BP. Blood pressure. CT. Carboxyltransferase. CYP7a1. Cholesterol 7-α-hydroxylase. ChoRE. Carbohydrate response element. ChREBP. Carbohydrate response element binding protein. CPT - 1. Carnitine palmitoyltransferase – 1. Evolv-6. Long chain fatty acid elongase. EMSA. Eletrophoretic mobility shift assay. FAS. Fatty acid synthase. HDL HPRT. High density lipoprotein Hypoxanthine phosphorylribosyltransferase. IDF. International Diabetes Federation. IL-6. Interleukin -6. IL-1β. Interleukin-1β. IR. Insulin receptor. IRS. Insulin receptor substrate. IRE. Insulin response element. LXRs. Liver X receptors. I.
(9) LXREs. LXR response elements. LDL. Low density lipoprotein. MTTP NCEP - ATP - III. Microsomal triglyceride transfer protein National Cholesterol Education Program –Adult Treatment Panel – III. PKA. Protein kinase A. PTP1B. Protein tyrosine phosphatase 1B. PDI. Protein disulfide isomerase. PP2A. Protein phosphatase 2 A. RXR. Retinoid X receptor. SCD1 SREBP-1c S. Stearoyl - CoA desaturase 1 Sterol regulatory element binding protein-1c. SRE. Sterol responsive element. SCAP. SREBP cleavage-activating protein. TOFA. 5-(Tetradecyloxy)-2-furancarboxylic acid. UCP2. Uncoupling protein 2. UCP3. Uncoupling protein 3. VLDL. Very low density lipoprotein. II.
(10) 組別縮寫對照表 組別縮寫對照表 縮 SW. 寫. 全. 名. Chow diet+ +30% % sucrose containing water. SWL. Chow diet+ +30% % sucrose containing water+94 mg EP / ㎏ 大鼠體重. SWM. Chow diet+ +30% % sucrose containing water+188 mg EP / ㎏ 大鼠體重. SWH. Chow diet+ +30% % sucrose containing water+470 mg EP / ㎏ 大鼠體重. III.
(11) 建立驗證調節脂肪酸代謝之細胞模式與改善動物代謝症候群模式 中國醫藥大學營養學系碩士班 林毓舜. 摘要 代謝症候群(Metabolic 代謝症候群 syndrome)其盛行率在全球日益增加 其盛行率在全球日益增加, 其中發生原因之ㄧ 其盛行率在全球日益增加,其中發生原因之ㄧ 為過多脂質堆積於組織中。 為過多脂質堆積於組織中。理論上促進脂肪酸氧化或抑制脂肪酸合成應可有效降低 理論上促進脂肪酸氧化或抑制脂肪酸合成應可有效降低 代謝症候群危險。 代謝症候群危險。已知 acetyl-CoA carboxylase (ACC) 是脂肪酸合成關鍵酵素, 是脂肪酸合成關鍵酵素,許 多研究已證實利用基因轉殖抑制 ACC 表現或使用 ACC 抑制劑 抑制劑之藥物均可有效改善 代謝症候群。 代謝症候群。 本實驗室之前自鼠肝成功分離 ACC 酵素, 酵素,利用此 利用此平台篩選具有 ACC 活性抑制 活性抑制 功效之中草藥 酒精萃有最強的抑制 功效之中草藥, 之中草藥,其中發現紅雞屎藤(P. 其中發現紅雞屎藤 hypoleucum Ohwi)酒精 酒精萃有最強的抑制 ACC 活性功效(IC ,因此本論文目標為建立細胞與動物模式驗證紅雞屎 活性功效 50=0.029 mg / ml), 藤酒精萃功效。 藤酒精萃功效。因紅雞屎藤酒精萃已知含有 因紅雞屎藤酒精萃已知含有 ACC 活性抑制物 活性抑制物, 抑制物,故在細胞與動物模 故在細胞與動物模 式上如能 )反應, 式上如能誘發出 如能誘發出 ACC 活性並促進脂質新生( 活性並促進脂質新生(de novo lipogenesis) 反應,再投予紅 投予紅 雞屎藤酒精萃應可觀察到最佳功效 雞屎藤酒精萃應可觀察到最佳功效。 應可觀察到最佳功效。 在細胞實驗方面, 在細胞實驗方面,利用 HepG2 人類肝癌細胞株, 人類肝癌細胞株,給予高糖培養液刺激細胞中 ACC 活性, 活性,發現在給予高糖 12 小時後可顯著增加 ACC 活性和增加細胞內三酸甘 油酯含量, 油酯含量,增加細胞內脂質生合成並增加細胞 apoB 分泌, 分泌,在此條件下預先以高糖 培養 6 小時後再給予紅雞屎藤酒精萃不同濃度( ) 小時後再給予紅雞屎藤酒精萃不同濃度(0.145 mg / mL 和 0.0725 mg / mL) 4 小時, 小時,可有效抑制 ACC 和 FAS 活性, 活性,細胞內三酸甘油酯含量與極低密度膽固醇 細胞內三酸甘油酯含量與極低密度膽固醇 分泌有關蛋白 Microsomal triglyceride transfer protein(MTTP) )之表現亦顯著降低 之表現亦顯著降低。 動物實驗中, 動物實驗中,以四週齡公 Wistar 大鼠分為對照及誘發組, 大鼠分為對照及誘發組,均餵以 chow diet 但 飲水分別為 plain water 及 30%蔗糖水 蔗糖水, 蔗糖水,實驗給予 20 週糖水後即誘發肥胖、 週糖水後即誘發肥胖、高胰島 素血症、 素血症、高血壓及高三酸甘油酯血症、 高血壓及高三酸甘油酯血症、葡萄糖不耐和胰島素阻抗等症狀, 葡萄糖不耐和胰島素阻抗等症狀,顯示糖水. IV.
(12) 誘發大鼠體內脂質新生反應。 誘發大鼠體內脂質新生反應。誘發組再分別不給予或給予紅雞屎藤酒精萃三種不同 劑量( )管餵 4 週,結果發現給予低劑量紅雞屎藤酒精萃可有效 劑量(94, 188, 469 mg / kg) 抑制肝臟中 ACC 活性, 活性,藉由抑制 ACC 活性減少脂肪酸生成, 活性減少脂肪酸生成,進一步改善脂肪組織 肥大、 肥大、高胰島素血症、 高胰島素血症、高三酸甘油酯血症、 高三酸甘油酯血症、葡萄糖不耐和胰島素阻抗等代謝症候群 之功效。 之功效。 本實驗成功建立用以評估 ACC 活性篩選平台之細胞和動物模式, 活性篩選平台之細胞和動物模式,並驗證紅雞 屎藤確實具有抗代謝症候群生理功效, 屎藤確實具有抗代謝症候群生理功效,值得開發。 值得開發。. 關鍵詞: 、脂質新生、 關鍵詞:代謝症候群、 代謝症候群、acetyl-CoA carboxylase、 脂質新生、红雞屎藤酒精萃. V.
(13) Establishing the in vitro and in vivo models for testing the fatty acid metabolic regulatory effect and anti-metabolic syndrome function Yu-Shun Lin. Abstract Metabolic syndrome is a global healthy problem with a soaring morbidity. Abnormal fatty acid metabolism and tissue fat burden may lie at the core of this disease. Modulation of fatty acid metabolism, either by stimulating fatty acid oxidation or inhibiting fatty acid synthesis, is therefore been suggested as a potential treatment for obesity and metabolic disorders. Acetyl–coenzyme A carboxylase (ACC) plays a crucial role in fatty acid metabolism, and its inhibition had been shown to be an effective approach for treating metabolic syndrome. In our previous study, partially purified ACC from rat liver was used to screen herbs commonly used in Taiwanese folk medicine for ACC inhibitory effects. An ethanol extract of Polygonum hypoleucum Ohwi (EP), the Taiwan tuber fleece flower, was found to have the highest inhibitory activity (half-maximal inhibitory concentration = 30 µg/mL). Therefore, this study was aimed at establishing the in vitro and in vivo models for testing compounds with fatty acid metabolic regulatory effect and anti-metabolic syndrome function. For this purpose, the cellular and animal models with induced de novo lipogenesis was needed for testing the physiological benefits of EP with ACC inhibitory activity. Induction of ACC activity in HepG2 cells was successfully achieved by incubating the cells with high glucose (30 mM) for 12 hours. At this time point, the TG content and apoB secretion of the high glucose–stimulated cells were also significantly increased compared with the low-glucose control. To test if EP had an inhibitory effect on lipogenesis, cells were stimulated with high glucose for 12 hours;. VI.
(14) and 2 doses (0.0725 and 0.145 mg/mL, i.e. 2.5- and 5-fold higher respectively than the IC50) of EP were added at 6 hours of high-glucose stimulation. As a result, the EP treated cells had significantly lower ACC and FAS activities, TG content and MTTP mRNA levels than their corresponding vehicle controls at 4 hr of EP treatment. In animal model, Wistar rats given a chow diet were divided into a control group (C) and an inducing group given, respectively, plain water or 30% sucrose water (SW) to drink ad libitum. After 20 weeks, rats with syndromes like obesity, hyperinsulinemia, hypertension, hypertriglyceridemia, glucose intolerance and insulin resistance were successfully induced by sucrose water drinking. Thereafter, rats in the inducing group were subdivided into 4 groups, without (SW) or with supplementation of EP at 94 ( SWL ), 188 ( SWM ) and 469 (SWH) mg/ kg body weight dosages for 4 wk. Results showed the SWL group had a significant reduction in ACC activity in liver, accompanied with the most striking improving effects on alleviating the symptoms associated with metabolic syndrome. In conclusion, we had successfully established the in vitro and in vivo models for testing compounds with fatty acid metabolic regulatory effect and anti-metabolic syndrome function in this study. By using high glucose-stimulated HepG2 cells and sucrose drinking water-treated animals, P. hypoleucum Ohwi, a Chinese herb, is shown to be effective in alleviating the metabolic syndrome, a function which is worthy to be further explored.. VII.
(15) 第一章 前言. 第一章 前言. 代謝症候群( )意指肥胖、 代謝症候群(Metabolic syndrome) 意指肥胖、血脂紊亂、 血脂紊亂、高血糖和高血壓等 多種特徵同時存在一個人身上, 多種特徵同時存在一個人身上,已成為世界性重要的公共衛生問題, 已成為世界性重要的公共衛生問題,且是第 2 型糖 尿病和心血管疾病之危險因子。 尿病和心血管疾病之危險因子。在台灣罹患代謝症候群人數有逐年增加趨勢, 在台灣罹患代謝症候群人數有逐年增加趨勢,已造 就國家醫療成本的龐大支出。 就國家醫療成本的龐大支出。若能有效開發出預防或抗代謝症候群之藥品、 若能有效開發出預防或抗代謝症候群之藥品、保健食 品,將有助於降低罹患代謝症候群人數。 將有助於降低罹患代謝症候群人數。. Acetyl-CoA carboxylase( (ACC) )為體內 de novo lipogenesis 之重要關鍵酵素, 之重要關鍵酵素, 調控脂肪酸生合成 調控脂肪酸生合成。其產物 malonyl - CoA 為肝臟中合成三酸甘油酯的原料外, 為肝臟中合成三酸甘油酯的原料外,亦 能調控脂肪酸氧化酵素 Carnitin palmitoyltransferase - 1 (CPT - 1), ,抑制其活性降 低脂肪酸氧化。 低脂肪酸氧化 。 若是過多脂肪酸合成造成脂肪酸溢流到其他非脂肪組織, 若是過多脂肪酸合成造成脂肪酸溢流到其他非脂肪組織 , 造成 lipotoxicity。 。因此, 因此,若能有效抑制 ACC 活性, 活性,即可抑制脂肪酸生合成和/或促進脂 即可抑制脂肪酸生合成和 或促進脂 肪酸氧化, 肪酸氧化,也許能降低組織中三酸甘油酯堆積導致肥胖和代謝症候群的發生。 也許能降低組織中三酸甘油酯堆積導致肥胖和代謝症候群的發生。. 本實驗室先前研究發現, 本實驗室先前研究發現,红雞屎藤中草藥具有抑制 ACC 活性之功效, 活性之功效,因此將 進一步應用於細胞和動物實驗上 進一步應用於細胞和動物實驗上, 實驗上,驗證红雞屎藤之抗代謝症候群之功效, 驗證红雞屎藤之抗代謝症候群之功效,首要之務 即是建立適合細胞和動物實驗, 即是建立適合細胞和動物實驗,以方便、 以方便、科學化評估红雞屎藤之有效性。 科學化評估红雞屎藤之有效性。. 1.
(16) 第二章 文獻回顧. 第二章 文獻回顧. 一、. 代謝症候群( ) 代謝症候群(Metabolic syndrome). (一) 代謝症候群定義. 1923 年 , 代謝症候群最早被敘述為包括有高血壓( ) 、 高血糖 代謝症候群最早被敘述為包括有高血壓( Hypertension) (Hyperglycaemia) )和高尿酸血症( )的症候群。 和高尿酸血症(Hyperuricaemia) 的症候群。在 1947 年的研究 指出腹部肥胖(Abdominal obesity)和脂質分佈與糖尿病及其他疾病的相關性 和脂質分佈與糖尿病及其他疾病的相關性; 指出腹部肥胖 和脂質分佈與糖尿病及其他疾病的相關性;接著 在 1965 年再次敘述到此症候群是含有高血壓 年再次敘述到此症候群是含有高血壓、 此症候群是含有高血壓、高血糖以及肥胖的症候群。 高血糖以及肥胖的症候群。Reaven 於 1988 年將腹部肥胖、 )、高三酸甘油酯血症 年將腹部肥胖、高胰島素血症( 高胰島素血症( Hyperinsulinemia)、 )、高三酸甘油酯血症 (Hypertriglyceridemia)、 )、高血壓以及 ,這一群對糖尿病 )、高血壓以及 Low - HDL - cholesterol, 和心血管疾病的危險因數命名為 Syndrome X, ,並指出代謝症候群主因是與胰島素 抗性有關(1)。 抗性有關 。. (二) 判定標準. 根據2001美國國家膽固醇教育計畫 美國國家膽固醇教育計畫 National Cholesterol Education Program 根據 Adult Treatment Panel - III( (NCEP – ATP - III) )主要針對心血管疾病的危險所建立 的判斷標準, (Ι) 判斷標準,建立造成代謝症候群五種危險因子: 建立造成代謝症候群五種危險因子: ( ) 腹部肥胖, 腹部肥胖,男性腰圍大於 102 cm和女性腰圍大於 和女性腰圍大於 88 cm; ;(Ⅱ ; (Ⅲ Ⅲ)高 高 (Ⅱ) 血中三酸甘油酯濃度大於 150 mg / dl; 密度脂蛋白 - 膽固醇( ) 濃度, 和女性小於 膽固醇(HDL – Cholesterol) 濃度,男性小於 40 mg / dl和女性小於 50 mg / dl; ;(Ⅳ ;(Ⅴ mg / dl。 。 (Ⅳ)血壓超過 135 / 85 mmHg; (Ⅴ)禁食血糖濃度大於110 禁食血糖濃度大於 NCEP判定須符合五項因子的其中三項以上即認為有代謝症候群且增加罹患心血管 判定須符合五項因子的其中三項以上即認為有代謝症候群且增加罹患心血管. 2.
(17) 第二章 文獻回顧. 疾病的風險(2)。 疾病的風險 。 在美國成年人罹患代謝症候群比率為24%。 在美國成年人罹患代謝症候群比率為 %。且已知與動脈硬化及 %。且已知與動脈硬化及 T2DM患者死亡率增加有正相關 患者死亡率增加有正相關(3, 。 患者死亡率增加有正相關 4)。. 在 2004 年,International Diabetes Federation( (IDF) )建立出適用於任何國家 的判斷標準。 的判斷標準。所建立的判斷標準包括有評估腹部脂肪堆積、 所建立的判斷標準包括有評估腹部脂肪堆積、胰島素抗性、 胰島素抗性、atherogenic dyslipidaemia、 、血壓增加、 。目前 IDF 血壓增加、proinflammatory state 和 pothrombotic state。 在 2005 年又建立了最新的判斷標準,特別強調中央型肥胖是判定代謝症候群的必要 條件, 條件,再加上高三酸甘油酯、 再加上高三酸甘油酯、低的高密度脂蛋白、 低的高密度脂蛋白、高血壓、 高血壓、高空腹血糖, 高空腹血糖,四項中兩 項。而中央型肥胖的判定則是依各民族自訂之標準(5)。 而中央型肥胖的判定則是依各民族自訂之標準 。. 依我國行政院衛生署國民健康局在 2006 年對代謝症候群的判定標準建議採 年對代謝症候群的判定標準建議採 94 年美國 NECP - ATP - Ⅲ的診斷標準, 的診斷標準,並將空腹血糖向下修正, 並將空腹血糖向下修正,於 96 年 1 月 18 日公 告,即以下 5 項危險因子中, 項危險因子中,若包含 3 項或以上者可判定為代謝症候群。 項或以上者可判定為代謝症候群。. 3.
(18) 第二章 文獻回顧. 表 2 – 1 台灣代謝症候群之判定標準 台灣代謝症候群之判定標準 危 險 因 子. 異 常 值 腰圍( ): 腰圍(waist). 腹部肥胖(Central obesity) 腹部肥胖. 男性 ≧90 cm 女性 ≧80 cm SBP ≧130 mmHg /. 血壓(BP)上升 上升 血壓 DBP ≧85 mmHg 男性 <40 mg/dl 高密度酯蛋白膽固醇(HDL-C)過低 過低 高密度酯蛋白膽固醇 女性 <50 mg/dl 空腹血糖值(Fasting glucose)上升 上升 空腹血糖值. FG ≧100 mg/dl. 三酸甘油酯(Triglyceride)上升 上升 三酸甘油酯. TG ≧150 mg/dl. 備註: 、「空腹血糖值上升 備註:上項危險因子中「 上項危險因子中「血壓上升」 血壓上升」 「空腹血糖值上升」 空腹血糖值上升」之判定, 之判定,包括依醫師 處方使用降血壓或降血糖等藥品(中 處方使用降血壓或降血糖等藥品 中、草藥除外), 草藥除外 ,血壓或血糖之檢驗值正常者。 血壓或血糖之檢驗值正常者。. 在盛行率方面, 在盛行率方面,國民健康局 91 年三高( 年三高(高血壓、 高血壓、高血糖、 高血糖、高血脂) 高血脂)調查研究顯 示,國人代謝症候群盛行率: %(男 %,女 %) ,且隨年 國人代謝症候群盛行率:15 歲以上為 14.99%( %(男 16.9%, %,女 13.8%) ,且隨年 齡上升而有增加的趨勢, 齡上升而有增加的趨勢,見表 2 - 1 所示。 所示。又據衛生署統計顯示, 又據衛生署統計顯示,代謝症候群所衍生 之腦血管疾病、 之腦血管疾病、心臟病、 心臟病、糖尿病、 糖尿病、高血壓等慢性疾病, 高血壓等慢性疾病,皆年居台灣十大死因榜中, 皆年居台灣十大死因榜中, 此現象也造就了國家對於醫療保健費用龐大的支出, 此現象也造就了國家對於醫療保健費用龐大的支出,見表 2 - 2 所示。 所示。因此如何預防 代謝症候群已成為我國及世界之新興重要公共衛生議題。 代謝症候群已成為我國及世界之新興重要公共衛生議題。. 4.
(19) 第二章 文獻回顧. 表 2 – 2 台灣代謝症候群及組成因子盛行率 91*年三高盛行率調查 男. 女. 合計. 16.9%. 13.8%. 14.99%. 高血壓(15 歲) 高血壓. 24.9%. 18.2%. 21.38%. 高三酸甘油酯. 20.3%. 11.3%. 15.6%. 高低密度膽固醇. 6.5%. 6.3%. 6.4%. 高血糖(19 歲↑) 高血糖. 8.8%. 7.2%. 8.0%. 高血糖(45 高血糖 歲↑). 15.5%. 14.0%. 高血糖(65 歲↑) 高血糖. 18.8 %. 22.8%. 糖尿病前期. 3.2%. 3.2%. 3.2%. 高膽固醇. 10.8%. 10.9%. 10.9%. 代謝症候群* 代謝症候群. 組成因子. 資料來源: 資料來源:行政院衛生署國民健康局. 5.
(20) 第二章 文獻回顧. 表 2 - 3 95年國人代謝症候群相關疾病醫療費用 年國人代謝症候群相關疾病醫療費用 就診人數. 住診費用. (萬 萬). (千點數 千點數) 千點數. 糖尿病. 120.2. 14,003,298. 高血壓. 258.0. 15,769,193. 心臟病. 133.6. 17,811,943. 腦中風. 52.8. 12,628,941. 小計. 564.6. 60,213,375. 癌症. 39.6. 30,983,734. 項目. 代謝症候群/癌症 代謝症候群 癌症. 1.94. 資料來源: 資料來源:衛生署95年全民健康保險醫療統計年報 衛生署 年全民健康保險醫療統計年報. (三) 代謝症候群發生原因. 代謝症候群發生原因, 代謝症候群發生原因,主要由於現代人飲食和生活型態改變有關 主要由於現代人飲食和生活型態改變有關。 變有關。隨著飲食西 化,飲食偏向過度精緻化的糖類食品、 飲食偏向過度精緻化的糖類食品、含糖飲料、 含糖飲料、高油飲食等, 高油飲食等,並缺乏運動進而產 生肥胖。 生肥胖。脂肪組織為調節身體熱量平衡之重要組織, 脂肪組織為調節身體熱量平衡之重要組織,今若攝取過多的熱量, 今若攝取過多的熱量,過多的 熱量會以脂肪形式儲存在白色脂肪組織中。 熱量會以脂肪形式儲存在白色脂肪組織中。長期熱量攝取過多, 長期熱量攝取過多,產生過多脂肪造成 脂肪細胞肥大並且脂肪組織無法負荷過多脂肪, 脂肪細胞肥大並且脂肪組織無法負荷過多脂肪,轉而將多餘的脂肪酸堆積流向其他 非脂肪組織, ,造成身體 非脂肪組織,如:肝臟、 肝臟、肌肉和胰臟。 肌肉和胰臟。而這樣的過程稱之為 lipotoxicity, 各組織對胰島素產生抗性、 各組織對胰島素產生抗性、減少胰島素分泌、 減少胰島素分泌、第 2 型糖尿病、 型糖尿病、心血管疾病接踵而來. 6.
(21) 第二章 文獻回顧. (6)。 。另種理論認為肥大的脂肪細胞也會釋出不利胰島素傳訊之脂肪細胞激素如: 另種理論認為肥大的脂肪細胞也會釋出不利胰島素傳訊之脂肪細胞激素如: resistin、 、plasminogen activator inhibitor I、 、Interleukin-6( (IL-6) ) 、Tumor necrosis factor α (TNF-α)及 及 free fatty acids 等,加重胰島素阻抗(7)。 加重胰島素阻抗 。. (四) 代謝症候群治療方式. 由 NCEP 建議預防代謝症候群的目標鎖定在飲食與運動, 建議預防代謝症候群的目標鎖定在飲食與運動,有研究顯示飲食控制 有研究顯示飲食控制 和增加活動量可降低體重、 和增加活動量可降低體重、促進全身氧化代謝和降低胰島素阻抗(2)。 促進全身氧化代謝和降低胰島素阻抗 。近年來有學者 提出調節脂肪酸代謝對改善肥胖和代謝症候群是具有潛力的對策(8)。 提出調節脂肪酸代謝對改善肥胖和代謝症候群是具有潛力的對策 。在肥胖者中發 現脂肪組織增加的原因可能來自於體內增加 de novo lipogenesis 和降低 fatty acid oxidation 。 因 〝 lipotoxicity 〞 導 致 體 內 發 生 Non - alcoholic steatohepatitis 、 dyslipidemia 和 insulin resistance。 。所以與脂質代謝相關的重要調控酵素成為治療代 謝症候群的研究焦點之ㄧ 謝症候群的研究焦點之ㄧ。例如抑制脂質生合成關鍵酵素 acetyl - CoA carboxylase (ACC)可能有助於抗肥胖與代謝症候群 可能有助於抗肥胖與代謝症候群。 可能有助於抗肥胖與代謝症候群。. 在藥物治療方面已證實亦有針對不同的代謝紊亂給予治療藥物, 在藥物治療方面已證實亦有針對不同的代謝紊亂給予治療藥物, 例如 Statins 治 療 高 膽 固 醇 血 症 、 Fibrates 和 Niacin 治 療 Hypertriglyceridemia 與 hypoalphalipoproteinemia、 、Angiotensin - converting enzyme inhibitors 治療高血 壓、Aspirin 治療 Hypercoagulability、 、Biguanides 和 Glitazones 治療高血糖(9)。 治療高血糖 。但 少有藥物能同時解決以上多種問題。 少有藥物能同時解決以上多種問題。. 7.
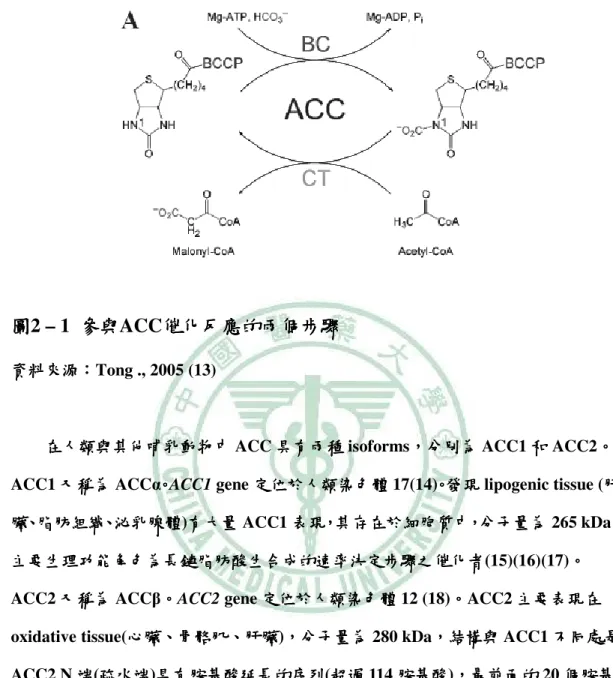
(22) 第二章 文獻回顧. 二、. 乙醯輔酶羧 A 化酶( ) 化酶(Acetyl-CoenzymeA carboxylase). (一) 何謂 Acetyl-CoenzymeA carboxylase( (ACC) ). 在 1958 年由 Salih 等人發現 ACC 是屬於 Biotin - dependent enzyme, ,其結構與 催化受質能力有關(10, 11)。 。ACC(EC 6.4.1.2)對脂質貯存和所有能量代謝有強力的影 對脂質貯存和所有能量代謝有強力的影 催化受質能力有關 響,ACC 為催化細胞質中脂質生合成的關鍵步驟 (Rate limiting reaction)之 之羧化反 應,將受質 acetyl - CoA 反應生成產物 malonyl - CoA,而 malonyl - CoA 是 Fatty acid synthase (FAS) 必需受質也是脂肪酸鏈延長的原料, 必需受質也是脂肪酸鏈延長的原料,特別在脂肪、 特別在脂肪、肝臟和泌乳腺; 肝臟和泌乳腺; 同時亦為粒線體中脂肪酸氧化作用的抑制者。 同時亦為粒線體中脂肪酸氧化作用的抑制者。. ACC 所催化的反應為 ACC 由 holo - carboxylase synthetase 將 apo - ACC 結構 lysine residue 上 ε - amino group 與 biotin 共價鍵結, 共價鍵結,又 biotin 稱為羧 稱為羧基的 carrier。 。 ACC 有兩個催化步驟: Ⅰ)需要 需要 ATP 參與提供能量於 biotin ureido ring 上的 N1 分 有兩個催化步驟:(Ⅰ 子,藉由 biotin carboxylase (BC)催化下提供 催化下提供 bicarbonate 進行羧 進行羧基的轉移, 基的轉移,形成 carboxy - biotinyl - ACC。(Ⅱ Ⅱ)將 將羧基自 biotin 轉移至 acetyl - CoA,形成產物 malonyl - CoA(12), ,此步驟則不需能量參與反應。 此步驟則不需能量參與反應。因此 ACC 催化反應整個過程需要兩個酵 素的參與, 、carboxyltransferase(CT), ,如圖 2 - 1 所示(13)。 。 素的參與,biotin carboxylase(BC)、 所示. 8.
(23) 第二章 文獻回顧. 圖2 – 1 參與ACC催化反應的兩個步驟 催化反應的兩個步驟 參與 資料來源: 資料來源:Tong ., 2005 (13). 在人類與其他哺乳動物中 ACC 具有兩種 isoforms, ,分別為 ACC1 和 ACC2。 。 ACC1 又稱為 ACCα。ACC1 gene 定位於人類染色體 17(14)。發現 lipogenic tissue (肝 肝 臟、脂肪組織、泌乳腺體)有大量 泌乳腺體 有大量 ACC1 表現,其存在於細胞質中,分子量為 265 kDa, , 主要生理功能角色為長鏈脂肪酸生合成的速率決定步驟之催化者(15)(16)(17)。 。 主要生理功能角色為長鏈脂肪酸生合成的速率決定步驟之催化者 ACC2 又稱為 ACCβ。 。ACC2 gene 定位於人類染色體 12 (18)。 。ACC2 主要表現在 oxidative tissue(心臟 心臟、 ,結構與 ACC1 不同處是 心臟、骨骼肌、 骨骼肌、肝臟), 肝臟 ,分子量為 280 kDa, ACC2 N 端(疏水端 疏水端)具有胺基酸延長的序列 疏水端 具有胺基酸延長的序列(超過 具有胺基酸延長的序列 超過 114 胺基酸), 胺基酸 ,最前面的 20 個胺基 酸可通透粒線體膜, 酸可通透粒線體膜,而產物 malonyl - CoA 為粒線體膜上調控脂肪酸氧化之酵素 carnitine palmitoyltransferase - Ι(CPT - 1)的異位抑制劑 的異位抑制劑(allosteric inhibitor), ,進而 的異位抑制劑 抑制脂肪酸氧化作用(33)。 。相較於 ACC1, ,ACC2 具有與 ATP 和 HCO3-較低的 Km, 抑制脂肪酸氧化作用 與 acetyl - CoA 則有較高的 Km(19)(20)。 。. 9.
(24) 第二章 文獻回顧. (二) ACC 調控. 在不同的生理狀況下 ACC 的半衰期為 1 至 3 天,其濃度並不會迅速的顯著改 變(21)。 。但仍受到營養狀況與荷爾蒙調控 ACC 基因表現。 基因表現。短時間 ACC 活性的調控 是經由異位調節和共價修飾( 是經由異位調節和共價修飾(磷酸化與去磷酸化) 磷酸化與去磷酸化)所調控。 所調控。. 1.. 異位調節. ACCs 活性會受到正回饋 feedforward 和負回饋 feedback 調控作用。 調控作用。Citrate 對 哺乳動物 ACCs 為正回饋的異位活化調控者, 為正回饋的異位活化調控者,且 Citrate 是 ACC 受質 acetyl - CoA 之前驅物, ,與 oxaloacetate 共同反應形成 citrate, ,其 之前驅物,在粒線體形成的 acetyl - CoA, 催化酵素為 tricarboxylic acid cycle 中的 citrate synthase。 。過多的 citrate 會通過粒 線體, 。因此 citrate 線體,出現在細胞質中, 出現在細胞質中,藉由 ATP - citrate lyase 轉生成 acetyl - CoA。 也是 acetyl - CoA 從粒線體轉運至細胞質中的決定者。 從粒線體轉運至細胞質中的決定者。細胞內的 citrate 生理濃度範 圍約在 0.1 ~ 1 mM, ,活化 ACC 的 Ka 約為 2 mM。 。而 citrate 調控 ACC 是透過改變 ACC 的原始結構, 。不只 的原始結構,促進 ACC 聚合作用表現為活化態, 聚合作用表現為活化態,增加 ACC 的活性(22)。 的活性 是 citrat 會促進 ACC 聚合作用, 、malonate、 、sulfate 和 聚合作用,像其他離子包括 isocitrate、 phosphate 也會促進 ACC 聚合增加其活性。 聚合增加其活性。. Long - chain acyl - CoA esters 為 ACC 活性之負回饋抑制者, 活性之負回饋抑制者,會促進 ACC 聚合 物的分離恢復成原始構型。 物的分離恢復成原始構型。16 碳或以上的飽和或不飽和 fatty acyl - CoA 之 Ki 範圍 在 1 ~ 150 nM(23)。 。此抑制作用可經由缺乏 stearoyl - CoA desaturase 1(SCD1)的 的 mice 試驗來解釋, 試驗來解釋,SCD1 為產生不飽和脂肪酸的酵素(24)。 為產生不飽和脂肪酸的酵素 。SCD1 的缺乏會產生比正 常情形下更大量的飽和 long - chain acyl - CoAs 而導致 ACCs 抑制作用和提高脂肪. 10.
(25) 第二章 文獻回顧. 酸氧化(25)。 。在 SCD - 1 缺乏小鼠 AMP - activated protein kinase (AMPK)的刺激也 的刺激也 酸氧化 扮演了代謝上的重要角色(25)。 。 扮演了代謝上的重要角色. 2.. 共價修飾. ACCs 活性也受到共價修飾的調控, 活性也受到共價修飾的調控,特別是 critical Ser residues 的磷酸化作用 (16)。 。AMPK 是在生理環境下可磷酸化 ACCs 的關鍵酵素, 的關鍵酵素,AMPK 是與各種壓力訊 息傳導和運動的反應之主要與調節代謝有關的酵素(26)。 。活化 AMPK 會調控相當多 息傳導和運動的反應之主要與調節代謝有關的酵素 的催化酵素活性和細胞內磷酸化反應, 的催化酵素活性和細胞內磷酸化反應,最後都是導致 ATP - consuming pathways 被 抑制和活化 ATP - producing pathways。 。於是 AMPK 磷酸化 ACCs 藉此降低酵素的 Vmax 和 citrate 敏感性, 敏感性,達到顯著抑制酵素活性。 達到顯著抑制酵素活性。而 AMPK 磷酸化位置包括 ACC1 的 Ser79、Ser1200、Ser1215 和 ACC2 的 Ser218。ACC1 Ser79 相當於 ACC2 Ser218 皆位 於 BC domain 之前,這兩個位置若發生磷酸化作用則能足夠抑制 ACCs 活性,ACC2 則缺乏與 ACC1 Ser1200 和 Ser1215 相同的磷酸化位置。 相同的磷酸化位置。. 除了 AMPK 外,Protein kinase A( (PKA) )也會磷酸化 ACC1 Ser79、Ser1200 和 ACC2, ,降低 ACC 的活性, 17)。 。 的活性,但此機制還尚未被證實(16, 但此機制還尚未被證實. 11.
(26) 第二章 文獻回顧. (三) 抑制 ACC 可改善代謝症候群之相關研究. Acetyl - CoA carboxylase(ACCs)在人類和大部分的有機生物體為脂肪酸代謝的 在人類和大部分的有機生物體為脂肪酸代謝的 重要角色。 重要角色。在抑制 ACC 的藥物發展作為治療各種疾病的目標, 的藥物發展作為治療各種疾病的目標,包括糖尿病、 包括糖尿病、肥胖、 肥胖、 癌症和細菌感染已相當令人重視。 癌症和細菌感染已相當令人重視。抑制 ACC 也抑制脂質生合成(肝臟 也抑制脂質生合成 肝臟、 肝臟、脂肪組織) 脂肪組織 同時也刺激氧化組織(肝臟 同時也刺激氧化組織 肝臟、 肝臟、心臟、 心臟、骨骼肌)脂肪酸氧化 骨骼肌 脂肪酸氧化。 脂肪酸氧化。提供了一個引人注意的改 善代謝症候群之相關模式。 善代謝症候群之相關模式。以下為抑制 ACC 運用在改善心血管疾病有關的危險因 子 (例如肥胖 例如肥胖、 例如肥胖、糖尿病、 糖尿病、胰島素阻抗、 胰島素阻抗、代謝症候群)之相關文獻報告 代謝症候群 之相關文獻報告. 利用 ACC2 cDNA probe 分離出 ACC2 genomic clone, ,以胚胎幹細胞建構一段 ACC2 突 變 的 對 偶 基 因 , 也 就 是 在 ACC2 序 列 上 的 biotin binding site 以 hypoxanthine phosphorylribosyltransferase (HPRT)取代 取代, 取代,經同質互換後, 經同質互換後,目標剔除 ACC2 基因表現。 基因表現。此方法可降低由高脂或高糖飲食誘導肥胖和 Type 2 DM 的小鼠肝 臟脂質含量和胰島素阻抗之現象(27)(28), ,提高肝臟和骨骼肌脂肪酸氧化、 臟脂質含量和胰島素阻抗之現象 提高肝臟和骨骼肌脂肪酸氧化、心臟和脂 肪組織 uncoupling protein(UCP2)、 、骨骼肌 uncoupling protein 3 (UCP3)濃度 濃度, 濃度,增加 能量消耗(28)、 、減少體重、 能量消耗 減少體重、血糖、 血糖、組織肝醣含量。 組織肝醣含量。 有學者也嘗試將動物體內 ACC1 knockout, ,但卻發現剔除 ACC1 後的小鼠無法 正常生長, 。 正常生長,胎死於母體腹中(29)。 胎死於母體腹中. TOFA 為類似長鏈脂肪酸的藥物, 為類似長鏈脂肪酸的藥物,當 TOFA 在肝臟微粒體中轉生成為 ToFyl CoA 則能抑制 ACC 活性(30), ,在肝細胞(31)或動物模式 或動物模式(87-89)中發現可降低脂肪酸 中發現可降低脂肪酸 活性 在肝細胞 或動物模式 合成與三酸甘油酯分泌、 、 降低體重(33)。 。 合成與三酸甘油酯分泌 、 降低血膽固醇與三酸甘油酯(31)(32)、 降低血膽固醇與三酸甘油酯 降低體重. 12.
(27) 第二章 文獻回顧. 圖 2 -2. 三、. TOFA 結構. 高糖飲食誘發代謝症候群. (一) 高糖( 高糖(蔗糖) 蔗糖)飲食引發代謝症候群之動物模式. 在建立代謝症候群動物模式上, 在建立代謝症候群動物模式上,給予齧齒類動物高油或高糖(包括蔗糖或果糖 給予齧齒類動物高油或高糖 包括蔗糖或果糖) 包括蔗糖或果糖 飲食是常用方法, 飲食是常用方法,相較於高脂飲食穩定誘發動物肥胖及胰島素阻抗, 相較於高脂飲食穩定誘發動物肥胖及胰島素阻抗,文獻顯示高糖 飲食誘發效果不ㄧ 飲食誘發效果不ㄧ。有研究指出若不提高飼料油脂比例, 有研究指出若不提高飼料油脂比例,僅以蔗糖置換同等量澱粉 給予 C57BL/6J 小鼠, 35)。 。Sumiyoshi 等人給予 小鼠,無法有效誘發代謝症候群現象(34, 無法有效誘發代謝症候群現象 等人給予 C57BL/6J 小鼠 50% %蔗糖添加於飼料中( ,無法誘發小鼠肥胖與體脂增 蔗糖添加於飼料中(為期 55 週) ,無法誘發小鼠肥胖與體脂增 加(34)。 。El Hafidi et al (2001) 利用 30% %蔗糖水成功誘發 Wistar rats 代謝症候群 (36), ,之後此模式之應用可見於數篇文獻(37)。 。他們發現持續給予 Wistar 大鼠蔗糖 之後此模式之應用可見於數篇文獻 水 39 週後, 週後,會出現肥胖、 會出現肥胖、高血糖、 高血糖、高胰島素血症與高三酸甘油酯血症, 高胰島素血症與高三酸甘油酯血症,並且出現葡 萄糖不耐與胰島素阻抗現象之代謝症候群相關症狀(37); ;給予糖水動物雖然固體飼 萄糖不耐與胰島素阻抗現象之代謝症候群相關症狀 料攝取減少, 料攝取減少,總熱量攝取卻大幅增加。 總熱量攝取卻大幅增加。此結果顯示高糖來自固體食物或液體飲料會 有不同效果, 有不同效果,事實證明蔗糖給予的形式( 事實證明蔗糖給予的形式(固體或液體) 固體或液體)會影響誘發效果, 會影響誘發效果,當熱量以 液體形式供應引起的代償反應(抑制食慾 液體形式供應引起的代償反應 抑制食慾)效果較差 抑制食慾 效果較差(38), 效果較差 ,因此蔗糖取代飼料中澱粉通 常無法有效提高動物熱量攝取, 常無法有效提高動物熱量攝取,但以糖水供應時則會造成熱量攝取失控。 但以糖水供應時則會造成熱量攝取失控。糖水模式 類似於人類過度攝取含糖飲料, 類似於人類過度攝取含糖飲料,造就營養素缺乏但熱量過剩之營養狀態。 造就營養素缺乏但熱量過剩之營養狀態。. 13.
(28) 第二章 文獻回顧. (二) 高糖誘發肝脂堆積及胰島素阻抗之細胞模式. 在誘發肝脂堆積及胰島素阻抗之肝細胞模式方面, 在誘發肝脂堆積及胰島素阻抗之肝細胞模式方面,Zang 等人於 2004 年發表 (39), ,利用人類肝癌細胞 HepG2, ,給予 24 小時高糖濃度培養液( 小時高糖濃度培養液(葡萄糖濃度為 30 mM) )後,再給予胰島素一起培養, 再給予胰島素一起培養,發現細胞給予高糖濃度培養液, 發現細胞給予高糖濃度培養液,會減弱胰島素 所誘發之胰島素訊息傳遞路徑 所誘發之胰島素訊息傳遞路徑, 發之胰島素訊息傳遞路徑,其中又以胰島素訊息傳遞路徑中 Akt 磷酸化減少有 很大關係, 很大關係,已知 Akt 為胰島素訊息傳遞中胰島素接受器之下游蛋白, 為胰島素訊息傳遞中胰島素接受器之下游蛋白,負責細胞對於 葡萄糖擷取之調控(39)。 。給予高濃度葡萄糖刺激下不僅阻礙胰島素訊息傳遞, 葡萄糖擷取之調控 給予高濃度葡萄糖刺激下不僅阻礙胰島素訊息傳遞,同時 發現, 發現,細胞給予高濃度葡萄糖刺激下, 細胞給予高濃度葡萄糖刺激下,細胞內 AMPK 磷酸化減少, 磷酸化減少,造成細胞中 ACC 磷酸化減少, 磷酸化減少,細胞傾向脂質生合成反應, 細胞傾向脂質生合成反應,使細胞中三酸甘油酯大量產生。 使細胞中三酸甘油酯大量產生。. 根據以上細胞和動物實驗可知, 根據以上細胞和動物實驗可知,持續給予高糖下, 持續給予高糖下,會使肝細胞產生胰島素阻抗, 會使肝細胞產生胰島素阻抗, 減弱胰島素功能 減弱胰島素功能, 素功能,造成細胞對於葡萄糖擷取困難, 造成細胞對於葡萄糖擷取困難,便形成高血糖, 便形成高血糖,而過高的血糖一 再刺激胰臟分泌大量胰島素, 再刺激胰臟分泌大量胰島素,造成了高胰島素血症進一步加重胰島素阻抗現象。 造成了高胰島素血症進一步加重胰島素阻抗現象。同 時,給予高糖會增加肝細胞大量合成脂肪酸, 給予高糖會增加肝細胞大量合成脂肪酸,過多脂肪酸形成便引發高三酸甘油酯 血症。 血症。而脂肪酸儲存在脂肪細胞內, 而脂肪酸儲存在脂肪細胞內,造成肥胖、 造成肥胖、體脂肪增加和脂肪細胞肥大。 體脂肪增加和脂肪細胞肥大。過多 脂肪酸若是溢流到其他組織, 脂肪酸若是溢流到其他組織,如:肝臟、 肝臟、肌肉、 肌肉、胰臟等非脂肪組織中, 胰臟等非脂肪組織中,造成 lipotoxicity 現象, 現象,進一步加重胰島素阻抗。 進一步加重胰島素阻抗。此外, 此外,脂肪細胞肥大也會分泌不利健康的脂肪細胞 激素, 激素,更不利於胰島素訊息之傳遞。 更不利於胰島素訊息之傳遞。種種現象的累積, 種種現象的累積,便形成現代文明病-代謝 便形成現代文明病 代謝症候 代謝症候 群。. 14.
(29) 第二章 文獻回顧. (三) 肝臟中脂質生合成相關轉錄因子. 1.. Liver X receptors( (LXRs) ). 在 1994 年於大鼠肝臟中發現 Liver X receptors( (LXRs) )為 ligand-activated 轉 錄因子, 。LXRs 包含 LXRα(NR1H3) )和 LXRβ(NR1H2) ) 錄因子,屬於核受器的一種(40)(41)。 屬於核受器的一種 兩種異構物, % 兩種異構物,此兩異構物被辨識在它們的配位子結合區和 DNA 結合區將近有 結合區將近有 78% 胺基酸序列相同(42)。 。LXRα 主要表現在肝臟、 胺基酸序列相同 主要表現在肝臟、脂肪組織、 脂肪組織、脾臟、 脾臟、小腸、 小腸、腎臟和肺 臟中, 。 臟中,相反的 LXRβ 卻在身體各部組織都會表現(40)(41)。 卻在身體各部組織都會表現. LXRs 最初因它在自然界中的配位子並沒有被發現, 最初因它在自然界中的配位子並沒有被發現,故被視為一種 orphan nuclear receptor, ,直到近年才被發現(42)。 。當細胞中有 LXRs 配位子出現時, 直到近年才被發現 配位子出現時,便會 活化 LXRs, ,此時會與 retinoid X receptor( (RXR) )結合形成 heterodimers, ,進入細 胞核中結合至含有 LXR response elements( (LXREs) )之 promoter regions(43)。 。 LXREs 是由六個核甘酸( )重複兩次, 是由六個核甘酸(AGGTCA) 重複兩次,其中間夾雜四個隨機的核甘酸所 形成之特定序列(43)。 。 形成之特定序列. 研究發現, 之內生性配位子大多為一些氧化固醇類, 研究發現,LXR之內生性配位子大多為一些氧化固醇類 之內生性配位子大多為一些氧化固醇類,其中又以下列氧化固 醇類活化LXR最為強烈 最為強烈。 和 20(S)-hydroxycholesterol屬於 屬於 醇類活化 最為強烈 。 22(R)-hydroxycholesterol和 固醇類賀爾蒙之中間代謝物, 產生於大腦, 固醇類賀爾蒙之中間代謝物, 24(S)-hydroxycholesterol產生於大腦 產生於大腦,是血清中氧化 固醇最主要的形式。 大量存在於肝臟中(44-47)。 。大部分的 固醇最主要的形式。24(S),25-epoxycholesterol大量存在於肝臟中 大量存在於肝臟中 氧 化 固 醇 對 LXRs 之 親 和 力 皆 差 不 多 , 但 5,6-24(S),25-dieopxycholesterolc 和 6a-hydroxy bile acids這兩種氧化固醇只專一性接合至 這兩種氧化固醇只專一性接合至LXRα(48)。 。除了天然的配位子 這兩種氧化固醇只專一性接合至 外,也開發合成出人工LXRs配位子 配位子T0901317和 和GW3965這兩種藥物 這兩種藥物(49, 50)。 。 也開發合成出人工 配位子 這兩種藥物. 15.
(30) 第二章 文獻回顧. 圖2-3 T0901317之結構圖 之結構圖. 圖2-4 GW3965之結構圖 之結構圖. 近年來研究發現, 在膽固醇、 近年來研究發現,LRX在膽固醇 在膽固醇、脂質和糖類代謝上扮演重要的角色。 脂質和糖類代謝上扮演重要的角色。在膽固 醇代謝上, 為調控體內膽固醇之恆定, 基因後, 醇代謝上,LXR為調控體內膽固醇之恆定 為調控體內膽固醇之恆定,在剔除小鼠LXR基因後 在剔除小鼠 基因後,餵予高膽固醇 飲食發現, 缺乏小鼠會造成肝藏中膽固醇堆積、 飲食發現,與正常小鼠相比, 與正常小鼠相比,LXR缺乏小鼠會造成肝藏中膽固醇堆積 缺乏小鼠會造成肝藏中膽固醇堆積、肝腫大和高 膽固醇血症(51)。 。研究發現LXR調控膽酸合成之速率限制酵素 調控膽酸合成之速率限制酵素cholesterol 膽固醇血症 研究發現 調控膽酸合成之速率限制酵素 7-α-hydroxylase( (CYP7a1) ),此酵素為膽固醇代謝成膽酸之重要酵素 剔 ,此酵素為膽固醇代謝成膽酸之重要酵素, 此酵素為膽固醇代謝成膽酸之重要酵素,若將LXR剔 若將 除後, 合成, 除後,會造成無法增加CYP7a1合成 會造成無法增加 合成,因而使膽固醇代謝為膽酸減少, 因而使膽固醇代謝為膽酸減少,阻礙膽固醇 的排出(52)。 。此外, 也會促進膽酸排出, cassette 的排出 此外,LXR也會促進膽酸排出 也會促進膽酸排出,藉由調控ATP-binding 藉由調控 transporters( (ABC) )酵素, 54)。 。由此可知, 酵素,此酵素功用為加速膽酸被運往膽囊(53, 此酵素功用為加速膽酸被運往膽囊 由此可知, LXR對於膽固醇之代謝扮演重要角色 對於膽固醇之代謝扮演重要角色。 對於膽固醇之代謝扮演重要角色。. 2.. Carbohydrate responsive element binding protein( (ChREBP) ). 在2001年 年Uyeda等人 等人(55)利用質譜儀發現 利用質譜儀發現Carbohydrate responsive element 等人 利用質譜儀發現 binding protein( (ChREBP) )的存在, 構造為basic helix-loop-helix( (bHLH) ) 的存在,ChREBP構造為 構造為 為一種核受器, kDa。 。ChREBP主要表現在與脂質生合成相關組織 主要表現在與脂質生合成相關組織 為一種核受器,分子量大約為100 分子量大約為 中,如:肝臟、 肝臟、小腸、 小腸、腎臟和白色、 腎臟和白色、棕色脂肪組織。 棕色脂肪組織。. 16.
(31) 第二章 文獻回顧. ChREBP為主要調控葡萄糖代謝之轉綠因子 為主要調控葡萄糖代謝之轉綠因子。 基因小鼠, 為主要調控葡萄糖代謝之轉綠因子。在剔除ChREBP基因小鼠 在剔除 基因小鼠,給予 正常飲食下, 正常飲食下,剔除基因小鼠出現肝臟中肝糖負荷過多、 剔除基因小鼠出現肝臟中肝糖負荷過多、脂肪組織過小和減少血清中 游離脂肪酸含量, 游離脂肪酸含量,除了醣類和脂質代謝出現異常, 除了醣類和脂質代謝出現異常,並有胰島素和葡萄糖不耐的現象 (56)。 。抑制ob/bo小鼠中 小鼠中ChREBP表現 表現, 抑制 小鼠中 表現,則有助於減少肝脂堆積和增加肝臟、 則有助於減少肝脂堆積和增加肝臟、肌肉和 脂肪組織對於胰島素敏感性, 脂肪組織對於胰島素敏感性,有助於改善葡萄糖不耐、 有助於改善葡萄糖不耐、高血酯症、 高血酯症、高血糖和高胰島 素血症, 。由相關研究顯示, 對於發展肥胖和 素血症,有助於改善代謝症候群(57)。 有助於改善代謝症候群 由相關研究顯示,ChREBP對於發展肥胖和 第2型糖尿病過程中 型糖尿病過程中, 型糖尿病過程中,扮演舉足輕重的角色。 扮演舉足輕重的角色。. ChREBP活化會被葡萄糖所刺激 活化會被葡萄糖所刺激, 活化會被葡萄糖所刺激,而被昇糖激素所抑制。 而被昇糖激素所抑制。葡萄糖代謝物會促進 ChREBP轉位至細胞核中 轉位至細胞核中, 等人研究發 轉位至細胞核中,而昇糖激素反而會抑制此情況發生。 而昇糖激素反而會抑制此情況發生。Uyeda等人研究發 現(55), ,ChREBP上有三個磷酸化位點 上有三個磷酸化位點, 196、 、Ser 568和 和Thr 666。 。當葡萄 上有三個磷酸化位點,分別是Ser 分別是 糖代謝經由五碳糖代謝路徑, 5-phosphate, ,xylulose 糖代謝經由五碳糖代謝路徑,由葡萄糖形成xylulose 由葡萄糖形成 5-phosphate會進一步活化 會進一步活化protein phosphatase 2 A( (PP2A) )使PP2A將 將Ser 196上磷 上磷 會進一步活化 酸化給去除, 將剩餘磷酸根給去除 酸化給去除,進入細胞核中。 進入細胞核中。在進入細胞核後再被PP2A將剩餘磷酸 在進入細胞核後再被 將剩餘磷酸根給去除, 根給去除,露出 DNA結合區域 結合區域, response element( (ChoRE) )之 結合區域,進一步結合到含有carbohydrate 進一步結合到含有 promoter位置 位置。 序列皆主要為調控葡萄糖糖解 位置。含有ChoRE序列皆主要為調控葡萄糖 含有 序列皆主要為調控葡萄糖糖解和脂質生合成代謝之相關 糖解和脂質生合成代謝之相關 酵素, 、ACC、 、FAS和 和stearoyl-CoA desaturase, ,促進脂 酵素,如:L-type pyruvate kinase、 質生合成。 質生合成。. 3.. Sterol regulatory element binding protein-1c( (SREBP-1c) ). SREBPs 為協助控制細胞中 de novo lipogenesis 狀態之重要轉錄因子, 狀態之重要轉錄因子,負責調控 細胞中和細胞膜上脂質的多寡(58-60)。 細胞中和細胞膜上脂質的多寡 。SREBPs 其結構為 basic helix-loop-helix (bHLH) )為一種核受器。 、SREBP-1c 和 SREBP-2 為一種核受器。SREBPs 家族中包含 SREBP-1a、. 17.
(32) 第二章 文獻回顧. 三種異構物, 三種異構物,分別由 SREBF-1 和 SREBF-2 兩個基因所轉譯出來。 兩個基因所轉譯出來。而 SREBPs 的表 現會因組織的不同而表現在特定組織中 ,SREBP-1c 會表現於飽食後的肝臟 現會因組織的不同而表現在特定組織中(61-63), 因組織的不同而表現在特定組織中 中;SREBP-1a 和 SREBP-2 並沒有特定表現的組織, 並沒有特定表現的組織,在全身組織中皆會表現。 在全身組織中皆會表現。而 SREBP-1a 和 SREBP-1c 蛋白, 蛋白,負責調控單元不飽和和多元不飽和脂肪酸合成相關 酵素之基因表現(61-63), ,其中 SREBP-1c 會促進脂質生合成相關酵素 ACC、 、FAS、 、 酵素之基因表現 long chain fatty acid elongase( (Evolv-6) )和 stearoyl-CoA desaturase 1 之基因表現。 之基因表現。. SREBPs與代謝症候群的發展有著重要角色 與代謝症候群的發展有著重要角色。 transgenic/LDL 與代謝症候群的發展有著重要角色。利用SREBP-1a 利用 receptor KO小鼠和 小鼠和ob/ob LDL receptor KO小 小鼠顯示, 小鼠和 鼠顯示,會造成高血脂症別是高三酸甘 油酯血症, 脂蛋白清除能力(64, 65)。 。SREBP-1c調控肝臟中脂質生合 調控肝臟中脂質生合 油酯血症,並會阻礙apoB脂蛋白清除能力 並會阻礙 脂蛋白清除能力 成,與三酸甘油酯合成和極 與三酸甘油酯合成和極低密度脂蛋白的產生有著很大的關係, 低密度脂蛋白的產生有著很大的關係,在利用SREBP-1c 在利用 過度表現之小鼠, 脂蛋白含量增加和HDL脂蛋白含量減少 脂蛋白含量減少(66)。 。 過度表現之小鼠,出現血清中VLDL脂蛋白含量增加和 出現血清中 脂蛋白含量增加和 脂蛋白含量減少. SREBP由內質網中被 由內質網中被合成後 由內質網中被合成後, 合成後,便停留在內質網膜上等待活化訊號出現。 便停留在內質網膜上等待活化訊號出現。而 SREBPs的活化過程包含了調節膜上蛋白質的裂解 的活化過程包含了調節膜上蛋白質的裂解、 的活化過程包含了調節膜上蛋白質的裂解、轉位至細胞核和保持SREBPs 轉位至細胞核和保持 的穩定性。 合成完畢後, 活性的次單位-SREBP 的穩定性。SREBP合成完畢後 合成完畢後,其調節SREBP活性的次單位 其調節 活性的次單位 cleavage-activating protein (SCAP)結合在 結合在SREBP後面 後面, 結合在 後面,用以感應細胞內膽固醇多 寡,此時SREBPs為不活化態 為不活化態。 此時 為不活化態。當有活化訊息產生時, 當有活化訊息產生時,如:細胞中膽固醇減少或有 胰島素存在下, 會藉由Sar 1( (一個小的GTP結合蛋白 結合蛋白) 胰島素存在下,SCAP-SREBP complex會藉由 會藉由 一個小的 結合蛋白)作用 招喚其他vesicular transport coat proteins-Sec23和 和Sec24, ,幫助SCAP-SREBP 招喚其他 幫助 complex轉位至高基氏體上 轉位至高基氏體上, proteases和 和Site-2 轉位至高基氏體上,隨後會在高基氏體膜上藉由Site-1 隨後會在高基氏體膜上藉由 proteases作用下 作用下, ,進入細胞核中, 作用下,便形成活化態的nSREBP, 便形成活化態的 進入細胞核中,結合至含有sterol 結合至含有 responsive element( (SRE) )的目標基因上, 的目標基因上,促進其轉錄活性(67)。 促進其轉錄活性 。. 18.
(33) 第二章 文獻回顧. (四) 蔗糖誘發脂質新生( )之生化與分子機制 蔗糖誘發脂質新生(de novo lipogenesis). 蔗糖組成是由一分子葡萄糖與一分子果糖所形成。 蔗糖組成是由一分子葡萄糖與一分子果糖所形成。當攝取糖水( 當攝取糖水(蔗糖) 蔗糖)時,蔗 糖會在小腸內分解形成葡萄糖與果糖, 糖會在小腸內分解形成葡萄糖與果糖,再分別吸收進入血液中, 再分別吸收進入血液中,經由肝門靜脈流入 肝臟, 肝臟,肝臟會調節血液中葡萄糖與果糖濃度, 肝臟會調節血液中葡萄糖與果糖濃度,將葡萄糖與果糖進一步合成脂質, 將葡萄糖與果糖進一步合成脂質,並 將合成的脂質包裹於極 )從肝臟分泌出, 將合成的脂質包裹於極低密度膽固醇( 低密度膽固醇(VLDL) 從肝臟分泌出,藉以提供身體其他 組織利用, 組織利用,過多的脂質將貯存在脂肪組織中, 過多的脂質將貯存在脂肪組織中,以供給日後能量所需。 以供給日後能量所需。. 葡萄糖在肝臟中藉由醣解反應形成acetyl-CoA, ,進一步藉由ACC、 、FAS合成脂 合成脂 葡萄糖在肝臟中藉由醣解反應形成 進一步藉由 肪酸, )。另一 肪酸,此一連串由醣轉變成脂質步驟被稱為脂質新生( 此一連串由醣轉變成脂質步驟被稱為脂質新生(de novo lipogenesis) 。另一 方面, 方面,葡萄糖會促進胰臟分泌胰島素, 葡萄糖會促進胰臟分泌胰島素,加速脂質新生。 加速脂質新生。而果糖代謝對於葡萄糖和脂 質代謝有很重要的影響, )磷酸化 質代謝有很重要的影響,果糖進入肝細胞後, 果糖進入肝細胞後,經由果糖激酶( 經由果糖激酶(fructokinase) 形成fructose-1-phosphate後 後,藉由aldolase B作用下裂解成 作用下裂解成glyceraldehyde和 和 形成 藉由 作用下裂解成 dihydroxyacetone phosphate直接進入醣解反應 直接進入醣解反應。 直接進入醣解反應。相反的肝臟中葡萄糖代謝受到肝臟 中肝糖、 和ATP濃度所影響 濃度所影響, 減慢醣解反應 中肝糖、citrate和 濃度所影響,進而抑制了phosphofructokinase減慢 進而抑制了 減慢醣解反應。 醣解反應。 但果糖代謝不受到phosphofructokinase所影響 所影響, 但果糖代謝不受到 所影響,因此果糖代謝跳過醣解反應之限制 步驟, 。 步驟,因此果糖相較於葡萄糖代謝較快, 因此果糖相較於葡萄糖代謝較快,更有利於脂質合成(68)。 更有利於脂質合成. 就分子機制層面,Mitro 等人於 2007 年(69)研究發現 研究發現,葡萄糖和其代謝物 glucose 6-phosphate 可直接接合到 LXRs 之 ligand binding domain 上,促進 LXRs 轉位活 性,由細胞質中轉位到細胞核中結合到 LXREs 上,由此可知葡萄糖和其代謝物 glucose 6-phosphate 為 LXRs 之促進子。 之促進子。文獻指出, 文獻指出,ChREBP 和 SREBP-1c 兩個葡 萄糖代謝和脂質代謝調控之重要轉錄因子, 萄糖代謝和脂質代謝調控之重要轉錄因子,其 promoter regions 中含有 LXREs 序 列(70, 71)。 。餵予小鼠 LXR 之促進劑-T0901317 其肝臟中 SREBP-1 基因表現大量增 之促進劑. 19.
數據
+7
Outline
相關文件
國立高雄師範大學數學教育研究所碩士論文。全國博碩士論文資訊網 全國博碩士論文資訊網 全國博碩士論文資訊網,
低脂奶類
Borrowing this concept, we think that relevance, skillful means, and the elective affinity among lay practitioners might be able to explain and prove that modernity exists
Dialogic inquiry, collaborative inquiry, community of inquiry.
因此在表 5-4 評估次項目中,統計結果顯示政治穩定度、房產政 策、官僚政治以及景氣是接受度最高的,可以顯示政局安定以及當局
在軟體的使用方面,使用 Simulink 來進行。Simulink 是一種分析與模擬動態
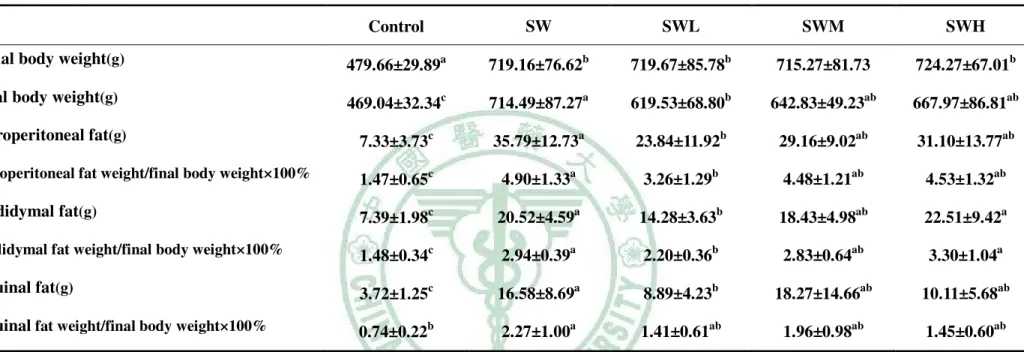
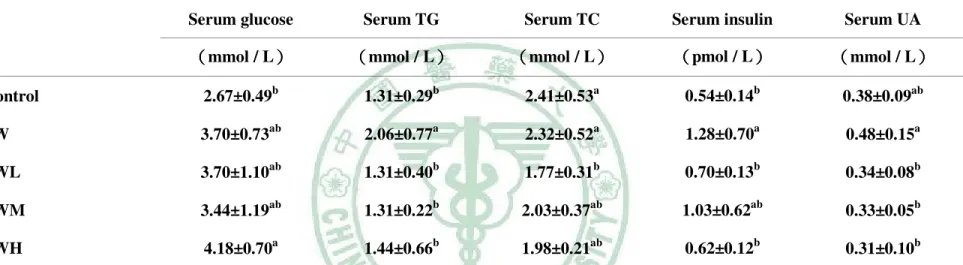
表 2.1 停車場經營管理模型之之實證應用相關文獻整理 學者 內容 研究方法 結論
辣椒 有利於促進胃黏膜的再生,維持胃腸 細胞功能,防治胃潰瘍。常食可降低 血脂、血糖減少血栓形成,且有效地
