日本腦炎病毒非結構蛋白NS4A與NS4B阻撓第一型干擾素訊息傳遞及其與細胞因子之交互作用; Inhibition of INF-α/β Signaling by the NS4A and NS4B of Japanese Encephalitis Virus and its interactions with cellular factor
96
0
0
全文
(2) 中文摘要 日本腦炎病毒是屬於黃病毒科之中的一員,含單股正性的 RNA 病 毒,基因體可轉譯生成三個結構蛋白(核蛋白,前膜蛋白,套膜蛋白) 以及七個非結構蛋白 (NS1, NS2a, NS2b,NS3,NS4a,NS4b,and NS5)。 日本腦炎病毒已經被發現可以抑制干擾素反應的訊息傳遞路徑,且在 先前的研究已證實登革熱病毒的 NS2A、NS4A 與 NS4B,以及西尼羅河 病毒的 NS4B 蛋白可以抑制第一型干擾素的訊息傳遞。因此,本篇論 文目的在於研究日本腦炎病毒 NS4A 與 NS4B 蛋白拮抗第一型干擾素反 應的可能性及其機轉。首先藉 interferon-stimulating response element (ISRE)啟動子結合冷光報導基因之細胞內訊息傳遞偵測系 統證實細胞先感染日本腦炎病毒再加干擾素處理,ISRE 啟動冷光報 導基之活性明顯受到抑制;但先加干擾素處理後再感染病毒時,冷光 報導基之活性無明顯抑制。然後建立穩定表現病毒蛋白 NS4A 與 NS4B 的 BHK-21 與 TE671 細胞,經由干擾素-α/β處理後,利用以及利用 西方墨點法偵測 Stat2 蛋白質的磷酸化,觀察 NS4A 與 NS4B 對第一型 干擾素的訊息傳遞是否受影響。結果發現病毒蛋白 NS4A 在 BHK-21 細 胞表現分別可以抑制 100%的α-干擾素所誘發 ISRE 冷光報導基之活 性與 60%β-干擾素所誘發 ISRE 冷光報導基之活性。病毒蛋白 S4B 在 BHK-21 細胞表現可以抑制 15%干擾素-α/β所誘發 ISRE 冷光報導基 i.
(3) 之活性。同樣地,NS4A 與 NS4B 在 TE671 細胞表現可抑制 35~45%干擾 素-α/β所誘發 ISRE 冷光報導基之活性。接下來,運用噬菌體表現 人腦 cDNA 基因庫的技術,找尋到 NS4A 交互作用的神經細胞因子 Dead Box42 。利用共同免疫沉澱-西方墨點法,驗證 NS4A 重組蛋白與 Dead Box42 之 C 端重組蛋白間專一性結合。經共軛焦顯微鏡的觀察,證明 NS4A 重組蛋白與 Dead Box42 之 C 端重組蛋白在 BHK-21 與 TE671 細 胞內表現有同位化的現象。進一步在過度表現 Dead BOX42 的 BHK-21 細胞中可以抵抗日本腦炎病毒感染時抑制干擾素所誘發 ISRE 冷光報 導基之活性,使原本冷光值明顯上升將近七倍。本研究結果將可應用 在生藥技術方面,NS4A 與 Dead Box42 可做為抗病毒藥物發展之標的。. ii.
(4) Abstract Japanese encephalitis virus (JEV) is a member of Flavivirus. JEV contains a single-stranded positive-sense RNA genome.. JEV genome. encodes for 3 structural proteins (C, prM, and E) and 7 nonstructural proteins (NS1, NS2a, NS2b, NS3, NS4a, NS4b, and NS5). JEV has been demonstrated the inhibitory effect on the IFN-α/β response by blocking Jak-Stat singling pathway. In addition, dengue virus type 2 NS2A, NS4A, and NS4B, and West Nile virus NS4B were associated with the inhibition of IFN-α/β signaling. In this study, we intend to investigate the effect of JEV NS4A and NS4B proteins on IFN-α/β signaling, and to identify the interaction of NS4A and NS4B proteins with neuron cellular factors. Our results indicated that JEV NS4A and NS4B inhibited IFN-α/β signaling using interferon stimulating response element (ISRE)-luciferase cis-acting reporter gene, and reduced stat2 protein phosphorylation with western blotting. Moreover, expression of NS4A in BHK-21 cells inhibited about 100% and 60% luciferase activity of ISRE-luciferase cis-acting reporter by INF-α and INF-β, respectively. Expression of NS4B in BHK-21 cells inhibited 15% luciferase activity of ISRE-luciferase cis-acting reporter by INF-α/β. Expression of NS4A or NS4B in TE671 cells also inhibit around 35~45% luciferase activity of ISRE-luciferase cis-acting reporter by INF-α/β. Dead box42 protein that interacted with the NS4A protein has been identified from a phage display human brain cDNA library.. Co-immunoprecipitation showed. the binding interaction of recombinant NS4A protein with C-terminus of Dead box42 protein.. In addition, confocal imaging indicated the iii.
(5) co-localization of NS4A with DDX42 in BHK-21 and TE671 cells. Overexpression of DDX42 in BHK-21 cell significantly blocked the inhibitory affect of INF-β by IFN-α/β signaling.. This study could be. helpful for understanding the JEV pathogenesis and developing antiviral drugs.. iv.
(6) 致謝 曾經,一個對未來懵懂的大學畢業生在人生抉擇的岔路上,選擇 了前往研究所的道路,兩年的碩士班生活,有像甘露般甜美的回憶, 也有像咖啡般苦澀的往事。曾經不只一次問過自己,辛苦嗎?開心 嗎?值得嗎?無論如何,終究還是走到了一個里程碑,無論是開心的 走過抑或辛苦的承受,兩年中的每一分每一秒,編織成生命中最璀璨 的ㄧ頁。對所有曾經幫助過的人,傑文心中只有滿滿的感激;然而, 由衷感謝的人太多,請恕劣者無法一一說明。 感謝兩年來高銘欽老師的提攜與照顧,不只實驗與課業方面令我 獲益良多,也在高老師身上學習到做為學者應有的氣度;感謝林振文 老師這兩年在實驗上細心的教導與督促,也讓我明白科學研究應該有 的嚴謹態度。還有楊翠青老師、蕭乃文老師、張清堯老師對於論文的 寶貴建議。兩年裡一起奮鬥的炳峯、鈞萍、善恩,相信彼此間的革命 友情會讓我們有相同的喜悅,你們是傑文這兩年來最重要的朋友;感 謝大學的好友景堂、小鳴、亮傑,在彼此討論課業與實驗讓我受益良 多;還有學妹素連、佳芳、詩雯,我們的精神和經驗就得靠你們傳承 了!大學部的學弟妹政怡、欣怡、靜宜、光庭、永瑞、尹萍、韋華、 紹晟、佳文還有是學妹也是嘉義的鄰居育存,謝謝你們在實驗室裡幫 助我大大小小的事情,有了你們實驗室也充滿了喜悅與歡樂。兩年最. v.
(7) 後的這段時間是最難熬的,由衷感激怡蘭、紀盈、小元、怡靜的支持 鼓勵,給了我堅持到底的勇氣,讓我能完成了這小小的成就。奈何紙 短情長,所不盡者尚有萬千,我會永遠感激且銘記每一位曾經幫助 過、鼓勵過傑文的人。 最後感謝從小到大扶持我的父母親還有家人們這兩年來的關懷 與無私奉獻。謹以此將這研究成果獻給以上各位,與傑文分享所有的 成長與喜悅。. vi.
(8) 目. 錄. 中文摘要. i. 英文摘要. iii. 致謝. vi. 目 ㄧ、. 錄. vii. 緒論. 1. 1.1. 日本腦炎簡介. 1. 1.2. 干擾素. 5. 1.3. 研究動機. 7. 1.4. 實驗策略. 9. 二、. 材料與方法. 13. 2.1. 實驗材料. 13. 2.1.1 2.1.2 2.1.3 2.1.4 2.1.5 2.1.6 2.1.7 2.1.8. 病毒株 細胞株 大腸桿菌菌株 質體 抗體 引子 噬菌體呈現人腦 cDNA 基因庫 實驗試劑及緩衝液. 13 13 13 14 15 16 17 17. 2.2. 實驗方法. 18. 2.2.1 2.2.2 2.2.3 2.2.4. 病毒增幅放大 病毒蝕班試驗 病毒 RNA 的抽取 反轉錄酵素-聚合酵素連鎖反應. 18 18 19 19. vii.
(9) 2.2.5 2.2.6 2.2.7 2.2.8 2.2.9 2.2.10 2.2.11 2.2.12 2.2.13 2.2.14 2.2.15 2.2.16 2.2.17 2.2.18 2.2.19 2.2.20 2.2.21 2.2.22 2.2.23 2.2.24 2.2.25 2.2.26. 聚合酵素連鎖反應 質體純化 限制酵素的水解 質體的構築 適應性細胞的製備 大腸桿菌之轉型 誘發大腸桿菌上乳糖啟動子(lac promoter)表現蛋白 IMAC 純化重組病毒蛋白質 (FPLC) 蛋白質濃度測定 蛋白質之電泳分析(SDS-PAGE) 西方墨點法 (Western blotting) DNA 轉染(DNA transfection) 冷光測試 ISRE 啟動子的活性(measure ISRE activity by luciferase assay) 噬菌體的生長與儲存 測定噬菌體的效價 噬菌體篩選 噬菌體酵素連結免疫吸附分析 噬菌體 DNA 製備 噬菌體胜肽酵素連結免疫吸附分析 共同免疫沉澱法 共同轉染免疫螢光染色法(confocal immunofluorescence assays) 免疫染色. 19 20 21 21 21 22 22 23 23 23 24 25 26 26 26 27 28 29 30 31 31 32. 三、 結果. 33. 3.1. 日本腦炎抵抗干擾素反應即訊息傳遞的機制. 33. 3.1.1 3.1.2 3.1.3 3.1.4. 日本腦炎病毒感染後,可以抑制第一型干擾素反應 日本腦炎病毒 NS4a、NS4b DNA 重組的質體構築 細胞內表現日本腦炎病毒 NS4a、NS4b 蛋白 JEV-NS4A 與 JEV-NS4B 降低 ISRE 啟動子的活性 JEV-NS4A 與 JEV-NS4B 藉由降低 STAT2 磷酸化來阻撓干擾 素的訊息傳遞. 33 33 34 35. 3.1.5. 36. 鑑定與日本腦炎病毒非結構蛋白 NS4A 作用的細胞 36. 3.2 因子. viii.
(10) 3.2.1 3.2.2 3.2.4 3.2.5 3.2.6. 日本腦炎病毒 NS4A 蛋白的質體構築 日本腦炎病毒 NS4A 蛋白質大量表現及純化 以日本腦炎病毒 NS4A 重組蛋白進行噬菌體篩選 Dead box42 蛋白的質體構築與純化 人腦細胞蛋白 DDX42 protein 與病毒蛋白 NS4A 共同免疫沉 澱. 36 37 37 38 39. 3.2.7. 共同轉染質體觀察細胞內蛋白質交互作用的關係. 40. 3.2.8. 過度表現 DDX42 的 BHK-21 細胞,抵抗日本腦炎病毒抑制第 一型干擾素反應. 40. 四、 討論 4.1 4.2. 42. 日本腦炎的抗干擾素作用 NS4A 蛋白與細胞內蛋白的交互作用. 42 44. 五、 參考文獻. 46. 六、 圖表. 52. ㄧ、附圖 二、附表. 52 77. ix.
(11) 一、緒論 文獻探討 1.1 日本腦炎簡介 日本腦炎是以節肢動物(Arthropod-borne)為媒介,主要是三 斑家蚊(Culex tritaeniorhynchus)與環斑家蚊(Culex annulex) 所傳染的病毒性疾病,因為日本腦炎是在西元 1871 年最早在日本報 告出來,在十九世紀初,日本爆發數次急性腦膜炎的大流行。西元 1935 年,病毒首先由病人腦部的病理組織被分離出來(Mitamura et al. 1936 ,Monath 1988),並且命名為日本腦炎 B 型病毒( Japanese B encephalitis virus),現今則稱為日本腦炎病毒(Japanese encephalitis virus,簡稱 JEV),是屬於黃病毒科(Flavivirudae) 的病毒成員之ㄧ。 人類感染日本腦炎病毒後會有持續發燒、嘔吐和呼吸衰竭等急性 腦炎臨床症狀(2001. Flaviviridae),有些則會有腦炎、出血性疾 病、肝炎等嚴重的症狀,嚴重則會死亡。發病後許多痊癒者會有永久 性神經損壞。日本腦炎病毒的毒性由老鼠動物實驗中病毒的神經侵襲 力(neuroinvasiveness)及神經毒力(neurovirulence)所組成。神 經侵襲力是指病毒在宿主週邊組織之繁殖複製引起病毒血症之後,侵 襲宿主中樞神經系統的能力。神經毒力則是病毒引起宿主中樞神經系. 1.
(12) 統細胞病變與引發腦炎的能力(McMinn et al.,1997) , 日本腦炎病毒利用宿主細胞外表接受器媒介的內吞噬作用進入 細胞內,而在細胞核附近,大量的進行轉譯(translation) 與蛋白質 修飾 (protein processing) 以及 RNA 的複製 (RNA replication) 。日本腦炎病毒為正股的 RNA 病毒,其基因組成大小 約為 11 個 kb 的 RNA,其中包含有 5'及 3'兩端的非轉譯區 (Untranslation Region ,UTR)還有中間開放讀碼區(open reading frames),而中間 ORF 的區域經過轉譯形成約將近 2500 個胺基酸的聚 胜肽鏈(polypeptide),而這個聚胜肽鏈經過細胞的胜肽水解酶及病 毒本身 NS2B-NS3 蛋白酶切割後,分成三個結構蛋白分別為核心蛋白 (C;capsid protein)、前膜蛋白 (prM;membrane protein)以及套 膜蛋白 (E;envelope protein),及七個非結構蛋白 NS1、NS2A、NS2B、 NS3、NS4A、NS4B 及 NS5。 ORF UTR 5‘m7g. C prM. ns4a. ns2a. E. ns2b. 3 structural proteins. NS. NS. NS. 7 non-structural. 2. UTR. ns4b. -3‘OH.
(13) 核蛋白、膜蛋白以及套膜蛋白等結構蛋白主要負責組合成病毒 顆粒(virion)。核心蛋白(C)含 126 胺基酸分子量大約 13.6kDa,組 成了病毒的外殼而包覆著病毒的核酸。而膜蛋白(M)(~8kDa)與套膜蛋 白(E)皆為醣蛋白,兩者以 heterodimer 的形式呈現在病毒的外殼 上,以穩定套膜蛋白的摺套膜蛋白(E)(~50KDa),是日本腦炎病毒 表面最主要的抗原蛋白,約為 500 個胺基酸大小。黃病毒的感染起始 於與宿主細胞的受器(receptor)結合後,藉由胞飲作用 (endocytosis)進入細胞內。在功能上,套膜蛋白與病毒進入細胞 時和細胞上病毒接受體的結合有關。 日本腦炎的非結構蛋白NS4A分子量約14kDa,其主要功能目前並 無太多的相關文獻探討。不過有文獻指出,四個原本功能不明的厭水 性非結構性蛋白質(NS2A、2B、4A和4B),在JEV感染生物體時,其 生活史中可能扮演膜通透性改變,而造成細胞病變的角色(J. Virol., Aug 1999)。在同樣屬於黃病毒科的C型肝炎病毒(HCV)中,HCV的病毒 蛋白NS4A在蛋白質多胜肽鏈切割時,扮演NS3蛋白酶的輔助因子,與 NS3形成穩定複合物,增加NS3裂解非結構蛋白質的能力(Failla et. al.,2002)。而其另外的功能也有作為NS5 RNA聚合酶之輔助因子 (Koch et al.,1999)。 非結構蛋白 NS4B 為含有多數疏水性胺基酸的膜蛋白質,如同. 3.
(14) NS4A,被發現於病毒感染時,使細胞膜通透性改變,造成細胞病變 (CPE)。而在 HCV 中 NS4B 被發現其功能可能是作為 NS5 RNA 聚合酶之 輔助因子(Koch et al.,1999),且被發現病毒感染時可誘導含有病毒 蛋白質的囊狀小泡的形成(Egger et al.,2002)。. 4.
(15) 1.2 干擾素 干擾素(interferon,INF)是ㄧ群蛋白質的家族,依照性質不同, 大小、製造的細胞、穩定性、還有功能細分後,大概可以分成兩型三 種干擾素。INF-α 與 INF-β 的性質十分相近,像是作用方式、結構 上的相似性,所以歸類為第一型干擾素,INF-α 是由白血球產生, 又稱 leukocyte INF,有抗病毒作用;INF-β 是由纖維原母細胞 (fibroblast)產生,又稱 fibroblast INF,亦有抗病毒作用。INF-γ 由 B lymphocyte、單核球(monocyte)及巨噬細胞(macrophage)經過 抗原刺激以後產生,又稱 immune INF,有抗病毒作用、活化巨噬細 胞、誘發 MHC 表現,所以被歸類為第二型干擾素。(最新醫用微生物 學 1998) RNA病毒複製的時候,所產生的中間產物(replicative intermediates),雙股RNA(ds-RNA),會誘發第一型干擾素(INF-α/β) 的生成,第一型干擾素為病毒感染時,生物體產生最直接的抗病毒產 物(Antiviral actions of interferons. 2001)。而第一型干擾素是 如何抗病毒呢?當細胞膜表面的第一型干擾素接受器(INFAR1與 INFAR2)形成二聚體,在跟INF-α/β蛋白 結合的時候,會使接受器 上的兩個激酶(kinase),Jak1與Tyk2磷酸化,而這兩個激酶會使訊息 傳遞路徑下游的兩個蛋白質Stat1與Stat2蛋白磷酸化,而磷酸化的. 5.
(16) stat1與stat2會形成異形二聚體,再與細胞中的IRF-9結合,形成一 個轉錄因子(transcriptional factor)再轉位進入細胞核,與一段 ISG(Interferon-α/βstimulated gene)基因的 ISRE(Interferon-α/βstimulated response element)啟動子結 合,使ISG這段基因製造出一些蛋白質來抗病毒,而這些蛋白包括三 百多種不同的蛋白質,其中包含一些酵素、熱休克蛋白、訊息傳遞蛋 白、轉錄因子以及一些細胞凋亡的蛋白等等,例如其中一種蛋白質, PKR(protein kinase R)為雙股RNA誘發干擾素產生所形成最重要的抗 病毒產物(J. Virol Feb. 2005)。 但是病毒為了生存、複製及繁衍下一代,必須演化出一些策略 與方式來對抗干擾素的免疫反應(J. Virol. 2001),像是抑制干擾素 的生成及訊息傳遞,或是阻礙干擾素誘發的抗病毒蛋白等等。例如 說,某些痘病毒會製造出一些與干擾素接受器相似的物質,藉由誘騙 干擾素結合而抑制;黃病毒的E1A蛋白阻礙干擾素的訊息傳遞;而一 些副黏液病毒科的病毒,例如腮腺炎病毒會使細胞內訊息傳遞路徑中 的STAT1蛋白降解,流行性感冒病毒會使STAT2的蛋白量下降及STAT 磷酸化的減少。而黃病毒科的病毒是如何來抵抗干擾素的作用呢?在 先前的研究已經有指出,黃病毒科中的登革熱病毒(Dengue virus,DEN-2)的非結構蛋白NS2A、NS4A、NS4B可以抵抗第一型干擾素. 6.
(17) 的作用(Proc. Natl. Acad. Sci. USA. 2003),西尼羅河(West Nile. virus,WNV)病毒的非結構蛋白NS2A與NS4B (J. Virol., July 2005), 黃熱病毒(Yellow fever virus,YFV)的NS4B都可以抑制干擾素(J. Virol., July 2005),而壁虱性腦炎病毒(Tick-borne encephalitis virus,TBEV)的非結構蛋白NS5也可以當作干擾素的抑制劑( J. Virol., Oct. 2005)。而日本腦炎病毒已經被發現能藉由抑制干擾素 接受器上的Tyk2激酶的磷酸化來阻擾訊息傳遞 (J. Virol., Sept. 2004)。. 1.3 研究動機 由前面描述得知,日本腦炎病毒感染細胞時,能夠明顯抵抗第 一型干擾素的抗病毒作用,且可以抑制細胞內干擾素的訊息傳遞;而 在登革熱病毒也被發現有相同的作用,且登革熱病毒的非結構蛋白 NS2A、NS4A、NS4B 已經被發現可以抵抗干擾素的作用,且同屬於黃 病毒科的 YFV、WNV 的 NS4B 以及 TBEV 的 NS5 皆可做為第一型干擾素 的抑制劑。但是,對於日本腦炎是藉由何種病毒蛋白抑制干擾素反應 卻無研究文獻探討。因此,本實驗我們選擇了黃病毒科中的日本腦炎 病毒作為研究的模型,利用 ISRE-Lucuferase 的報導基因來檢測是否 日本腦炎病毒的感染,能夠降低 ISRE 啟動子的活性。以及在細胞中,. 7.
(18) 能夠穩定表現日本腦炎的非結構蛋白 NS4A 與 NS4B 對干擾素訊息傳遞 的影響。再利用噬菌體表現技術,觀察蛋白質之間的交互作用,研究 細胞內是何種蛋白質類的細胞因子與日本腦炎病毒的 NS4A 蛋白質有 交互作用的現象,進而推論是否與日本腦炎抑制干擾素反應有所相 關。. 8.
(19) 1.4 實驗策略 本研究的實驗策略,分成幾個方向。 第ㄧ部分: 先將日本腦炎病毒蛋白基因進行質體選殖,將帶有日本腦炎蛋白 (NS4a NS4b)序列的質體在大腸桿菌中做大量的表現質體 DNA,抽取 細菌的質體以後,轉染進入 BHK-21 及 TE671 細胞,再藉由 ISRE-luc 的報導基因來偵測日本腦炎病毒顆粒本身以及日本腦炎病毒的 NS4A、NS4B 蛋白抑制干擾素反應的程度,再收取細胞的蛋白,利用 觀察蛋白磷酸化的程度,來判別干擾素訊息傳遞路徑是否被阻斷;再 將可以大量表現日本腦炎蛋白(NS4a NS4b)序列的質體在大腸桿菌中 大量表現蛋白質,並將蛋白質純化回收。. 第二部份: 是利用 T7 噬菌體質體在套膜蛋白位置 ( capsid protein )10B 之後,接上實驗所要篩選的人類腦細胞 cDNA 的基因庫能表現出人類 腦細胞各種蛋白因子。讓噬菌體表現人類腦細胞的胜肽庫針對日本腦 炎 NS4A 蛋白分別進行噬菌體篩選試驗,篩選出相關的人類腦細胞蛋 白因子之後,將噬菌體所表現蛋白的 DNA 序列選殖出來,再做大量的 蛋白質表現及純化回收並進行探討其功能性或是做進一步的運用。主. 9.
(20) 要流程是: T7 人類腦細胞的 cDNA library 表現出人類腦細胞分子,來對 日本腦炎病毒蛋白進行噬菌體篩選(bio-panning)之後,將挑選到 的噬菌體再與日本腦炎病毒蛋白進行 ELISA,再將 ELISA 反應 OD. 450nm. 吸光值較高的噬菌體,吸光值越高表示噬菌體與 NS4A 蛋白親合力越 高。接下來抽出噬菌體的 DNA 進行序列分析。於噬菌體中得到的人類 腦細胞 cDNA 在 NCBI 網站(http://www.ncbi.nlm.nih.gov/BLAST/) 進行核苷酸序列比對確實為人類基因序列。所以將在噬菌體中 cDNA 核苷酸從插入位置之後開始進行轉譯成蛋白質的位置(EcoR I : AAT/Asn),將核苷酸序列轉換成氨基酸序列,之後在 NCBI 網站 (http://www.ncbi.nlm.nih.gov/BLAST/)進行氨基酸比對。希望能 從噬菌體人類腦細胞因子所呈現 cDNA 基因庫中篩選出與日本腦炎病 毒蛋白有關的蛋白配體(protein ligand)。. 第三部份: 主要在於利用數種方法證明,所尋得的蛋白質與病毒蛋白是否 確實有交互作用的關係。在噬菌體蛋白質的功能性分析中,進行噬菌 體胜肽酵素連結免疫吸附分析(phage peptide ELISA)之後,將噬 菌體表面呈現蛋白的 DNA 序列接到 pET32a 的載體上,使蛋白大量表. 10.
(21) 現。之後進行蛋白質表現與純化,利用兩個蛋白質的共同免疫沉澱 (Co-immunoprecipitation)做進一步確定蛋白質-蛋白質之間的交 互作用關係。接下來再利用免疫螢光染色法(confocal immunofluorescence assays )使兩個蛋白表現紅色及綠色螢光,觀 察蛋白質是否座落在細胞內同一位置,進而推論兩者是否有互相作 用。 干擾素的生成與雙股 RNA(ds-RNA)有密切的相關,而日本腦炎病 毒其 NS4A 蛋白利用噬菌體表現所尋得的 DEAD BOX(DDX)蛋白,其為 ATP 依賴的 RNA 解旋酶(ATP-dependent RNA helicases)之其中一員, 有類似於 RNA 解旋酶的功能,可以打開雙股 RNA 螺旋。藉由此推測, 日本腦炎病毒抵抗干擾素反應,其機轉也許與細胞內 Dead box 蛋白 有關。而本論文研究藉由免疫沉澱法以及利用共軛焦顯微鏡觀察的結 果,發現日本腦炎的 NS4A 蛋白質與 Dead box42 蛋白彼此之間有交互 作用的現象。. 第四部份: 在確認日本腦炎病毒 NS4A 蛋白與細胞蛋白 DDX42 蛋白有交互作 用的現象以後,觀察細胞如果有過度表現 DDX42 蛋白,在日本腦炎病 毒感染時,是否影響第一型干擾素反應的表現。當細胞過度表現 DDX42. 11.
(22) 在病毒感染後,可以發現,ISRE 啟動子的活性會因為病毒感染而升 高。. 12.
(23) 二、材料與方法 2.1 實驗材料 2.1.1 病毒株 台灣本土株 日本腦炎 (T1P1) 2.1.2 細胞株 ( cell line ) Vero cell : 非洲綠猴腎臟細胞(African green monkey kidney cell)。培 養於 90%的 DMEM ( Dulbecco Modified Eagle Medium) , 包含有 10% 的 FBS(fetal bovine serum)。 BHK-21:幼小倉鼠腎臟纖維母細胞( baby hamster kidney cell )。培養 於 90%的 DMEM ( Dulbecco Modified Eagle Medium) , 包含有 10%的 FBS(fetal bovine serum)。 TE671:人類腦胚胎瘤細標(Human Caucasian medulloblastoma) 。培 養於 90%的 MEM,包含有 10%的 FBS(fetal bovine serum)。. 2.1.3 大腸桿菌菌株( Escherichia coli ) Top10 : 基因型為 F-,mcrA,∆(mrr-hsdRMS-mcrBC),φ80lacZ ,∆M1, ∆lacX74, recA1,araD139,galU,galK ,∆(ara-leu)7697,rpsL (StrR), endA1, nupG,用於質體轉形(transformation) 、質體製備。. BL21 (DE3):基因型為 F-,ompT ,hsdSB ( rB- mB- ),gal, dcm ( DE3 ) (camR), 用於蛋白質表現(protein induction) 。 13.
(24) 2.1.4 質體 ( Vector ) pET24a (Novagen):含有 5.3kb 個鹼基配對,可在大腸桿菌 BL21 品 系中,經 IPTG 誘導表現蛋白質,N’端帶有 T7-taq,C’端帶有 His-tag。 pET32a (Novagen):含有 5.9kb 個鹼基配對,可在大腸桿菌 BL21 品 系中,經 IPTG 誘導表現蛋白質,N’端帶有 Trx-tag、S-tag、His-tag, C’端帶有 His-tag。 pcDNA3.1/His B(Invitrogen): 含有5.5kb個鹼基配對,可在哺乳動物細 胞(mammalian cells)中,經 human cytomegalovirus immediate-early (CMV)promoter表現蛋白質,N’端帶Anti-Xpress epitope tag、His-tag。 pEGFP-N1(BD Biosciences Clontech): 含有4.7個鹼基配對,可在哺 乳動物細胞(mammalian cells)中,經 human cytomegalovirus immediate-early (CMV) promoter表現Enhanced green fluorescent protein (EGFP)蛋白質,在顯微鏡下產生螢光。. pDsRed1-1(BD Biosciences Clontech):含有 4.11kb,經表現 DsRed-Monomer Fluorescent Protein,在螢光顯微鏡下產生紅色螢光。. 14.
(25) 2.1.5 抗體(Antibody) Mouse Anti-His tag antibody (Novagen TM) Mouse Anti-Xpress tag antibody(Invitrogen) Anti-Thio antibody(Invitogen) Anti-Phospho-Tyk(Tyr1054/1055)(Cell Signaling) Anti-Phospho-Jak1(Tyr1022/1023)(Cell Signaling) Anti-Phospho-Jak2(Tyr1007/1008)(Cell Signaling) Anti-Stat(Cell Signaling) Anti-T7 tag(Novagen TM) Anti-Rabbit IgG AP-linked(Cell Signaling) Anti-mouse IgG(goat),AP labeled antibody(PerKlin-ElmerTM) Anti-mouse IgG(goat),FITC(PerKlin-ElmerTM). 15.
(26) 2.1.6 引子(primer) NS4a JEV-NS4a-F ATCCGAATTCTCAGCCGTTAGCTTC (EcoR I) JEV-NS4a-R GGTGCTCGAGCCTCTGTTTTTCCGG (Xho I) NS4b JEV-NS4b-F ATCCGAATTCAACGAGTACGGGATG (EcoR I) JEV-NS4b-R GGTGCTCGAGCCTTTTCAAGGAGGG (Xho I) DEAD BOX DDX23-F ATCCGAATTCCAACAGGGCCATAAC(EcoR I) DDX23-R CGACAAGCTTAACCAGTTGCAGCAT(Hind III) DDX23/pDsRed-forward ATCCGAATTCTGATGCAACAGGGCCATAAC(EcoR I) DDX23/pDsRed-reverse CGGTGGATCCAACCAGTTGCAGCAT(BamH I). 16.
(27) 2.1.7 噬菌體呈現人腦 cDNA 基因庫 本實驗室使用選購自 Novagen 的 T7 Select Phage Display System,噬菌體的載體為 T7Select 10-3b,宿主細菌為 BLT5403。 T7Select 10-3b 載體在噬菌體基因表現 capsid 蛋白質的區域,gene 10 的位置做了修改,原本 gene 10 由 10A (344aa)和 10B(397aa)所組合而 成,T7Select 10-3b 載體則將 gene 10A 去除並在 10B 後以 EcoRI/HindIII 的酵素切位接上人類腦細胞的 cDNA,其片段長度從 300bp 到 3Kbp。. 2.1.8 實驗試劑及緩衝液 試劑與緩衝液如表 2. 17.
(28) 2.2、實驗方法 2.2.1 病毒增幅放大 日本腦炎病毒是利用貼附性的幼小倉鼠腎細胞 ( BHK-21 cell ) 當作病毒的宿主細胞。BHK-21 細胞在 37℃ 恆溫培養箱中以 75T 培 養瓶 (75ml T flask )培養,等到細胞長滿單層時,移掉舊的培養液, 加入新的培養液(2% FBS) 10ml。以 M.O.I=10 的病毒顆粒對細胞數 的比例感染細胞。在感染病毒後四十八至七十二小時之間收集培養 液。以 3,000 rpm 離心 10 分鐘,取上清液,以每管 1ml 分裝,並儲 存於-80℃冰箱。. 2.2.2 病毒蝕班試驗 (plaque assay) BHK-21 細胞先在 6-well plate 上培養單層細胞約 6-7 成滿,吸 去舊的培養液。病毒經連續稀釋後,每個 well 加入 100μl,每個濃度 作兩個 well 重複。在 37℃,5%CO2 培養箱培養,每隔十五分鐘輕拍 輕搖 6-well plate 讓病毒液能均於分散。 一小時後加覆蓋液。將 6-well plate 至於培養箱培養三天。之後將上清液吸除,每個 well 加入 2-3ml 的染色液,並輕微搖晃。染色後第二天,到掉染色液,用清水清洗剩 餘的染色液。倒放乾燥後觀察病毒蝕班,並計算病毒蝕班數目。判定 病毒效價以 PFU/ml (plaque forming unit/ml)表示。 18.
(29) 2.2.3 病毒 RNA 的抽取 (purification of viral RNA) 以 QlAmp Viral RNA Mini Kit 抽取。將 0.14ml 病毒液加入 0.56ml AVL 震盪混合均勻,室溫靜置 10 分鐘後加入,加入純酒精 0.56ml 並混合均勻靜置 15 分鐘,在移入 spin collumn 中,離心 8000 轉 3 分鐘,加入 AW1 並離心 8000 轉 3 分鐘,再加入 AW1 離心 8000 轉 3 分鐘。換新的微量離心管,加入 60μl AVE 靜置一分鐘,離心 8000 轉 1 分鐘,取適量在 UV260/280 測吸光值。分析 RNA 的純度與定量。 之後將 RNA 儲存在-80℃冰箱。. 2.2.4 反轉錄酵素-聚合酵素連鎖反應 ( Reverse Transcription-PCR) 在 0.5ml 微量離心管中,取病毒 RNA 11μl 加入 dNTP (10 mM)1 μl 和針對病毒蛋白序列所設計的反股引子(reverse primer,10μM),65℃加熱五分鐘後快速的置於冰上,加入 DTT 2 μl 和 5 X buffer 4μl,在 42℃, 二分鐘後加入 superscript II Reverse Transcriptase (Invitrogen) 1 μl,於 42℃作用一小時進行第一股 DNA (cDNA)的合成,然後 70℃,十五分鐘終止反應。. 2.2.5 聚合酵素連鎖反應 (polymerase chain reaction,PCR) 在 0.5ml 微量離心管中,取病毒反轉錄的 cDNA 5 μl 做為模 19.
(30) 版( template),依序加入 ddH2O 79μl,10X Optipol buffer (GeneTeks)10 μl ,dNTP (10mM) 1 μl,Primer F 與 Primer R (10mM)各 2 μl ,最後 加 Optipol Taq polymerase 1 μl,總體積為 100 μl。混合均勻後加入約 20 μl 的礦物油在置於聚合酵素連鎖反應器(thermohybrid)的反應槽 中。先以 94℃作用五分鐘,再以每個週期,94℃作用 1 分鐘的 DNA 變性反應;52℃作用一分鐘三十秒的鏈合反應;72℃作用一分鐘的 DNA 合成反應。進行 35 個反應週期,之後以 72℃作用十五分鐘將 DNA 片段補齊。最後設定 4℃保存。將 PCR DNA 產物跑 2%瓊酯凝 膠(2% agarose gel)進行電泳分析。確定 PCR 產物片段大小正確, 以 Spin PCR Clean-Up system 試劑套組(Viogene)將 DNA 純化。. 2.2.6 質體純化 ( plasmid purification ) 含有質體的單一菌落,以 2ml LB 在 37°C 培養箱搖隔夜後取 出,以最大速離心 12,000rpm 五分鐘後,倒掉上清液,用 Gene-SpinTM Miniprep Purification Kit 試劑組 ( Protech technology )來純化質體。 以 solution I 溶液 200μl resuspend pellet,加入 solution II 溶液 200μl 上下倒置數次後,再加入 solution III 溶液 200μl 上下輕搖數次,最大 速離心 10 分鐘。將 spin column 插入一收集管(collection tube)中, 離心後的上清液移至 spin collumn 中,離心一分鐘,移除濾液後,加. 20.
(31) 入 700μl Washing solution,離心一分鐘。再將收集管以 1.5ml 微量離 心管置換,加入 20μl ddH2O 回收質體。. 2.2.7 限制酵素的水解 PCR 得到的產物,以 Spin PCR Clean-Up system 試劑套組 ( Viogene )將 DNA 純化後,以 80 μl 的 ddH2O 將 DNA 溶出,加 入 10X reaction buffer( Fermentas ) 10μl 及切位的限制酵素依照最佳比 例配製,使總體積為 100 μl,以 37℃作用隔夜。相對應的質體載體亦 以同樣的方式進行限制酵素的水解,來構築實驗的表現載體。 將作用隔夜的 DNA 產物跑 1%的 agarose gel,染 EtBr 十分 鐘,在 UV 燈下將正確位置的 DNA 切除下來,Gel Extraction System 試劑套組( Viogene )將 DNA 純化。. 2.2.8 質體的構築 : DNA 接合( DNA ligation ) 質體載體 1μl,加上欲接合的 Target DNA 7μl,10X ligation buffer 1μl,ligase 1μl,以 22℃反應 3-4 小時或 16℃作用隔夜。. 2.2.9 適應性細胞(competent cell)的製備 培養隔夜的 E. coli cell ( Top10 、BL21 ) 2ml,倒入 200ml. 21.
(32) LB 中,以 37℃ 培養箱培養二至三小時達到 log phase,取出置於冰 上靜置十分鐘後,再以 3000rpm、4℃、離心 15 分鐘,取出倒掉上清 液,以 0.1M CaCl2 resuspend pellet。於冰上靜置培養三十分鐘後,同 樣再以 3000rpm,4℃,離心十五分鐘,取出後小心地倒掉上清液, 再加入 0.1M CaCl2 1.6ml,glycerol 0.4ml( 20% )將 pellet resuspend, 然後靜置於冰上二小時後分裝至 1.5ml 離心管中,每管約含 200μl 左 右。. 2.2.10 大腸桿菌之轉型 (transformation) 將構築好的質體加入適應性細胞中,混合後至於冰上三十分 鐘。在經 42℃反應 90 秒後,迅速至於冰上三分鐘,之後加入 800μl 不含抗生素的 LB 培養液。在 37℃ 培養箱培養一小時後,離心去掉 上清液,均勻塗在含抗生素的 LB 培養基上。. 2.2.11 誘發大腸桿菌上乳糖啟動子(lac promoter)表現蛋白 將含有融合蛋白的實驗菌株培養在 10ml LB 培養液,隔天加 到 500ml 的 LB broth ,培養箱中 37℃培養二到三小時,吸光值 OD600nm 約為 0.4 到 1 之間,加入 IPTG 0.8M 500 μl,最終濃度為 0.8mM,再經 16℃培養四到五小時,將菌液收集並且離心,之後加. 22.
(33) 入 20ml 的 10mM imidazole。再置於超音波擊碎機(sonicator)將細 胞打碎。. 2.2.12 IMAC 純化重組病毒蛋白質 (FPLC) 利用 FPLC 純化蛋白,在 column 中裝填 10ml 的 Chelating SepharoseTM(Amersham Biocsciences),將以打碎的細菌溶液離心,上 清液流過鎳離子親和性層析凝膠,先用沖洗緩衝液(100mM imidazole)沖洗,再用析出緩衝液(400mM imidazole)將蛋白質析 出。. 2.2.13 蛋白質濃度測定 純化後的蛋白質濃度用 Bio-Rad protein assay 染劑(取 10ml, 以二次水稀釋到 50ml)進行測試。用牛血清蛋白(bovine serum albumin;BSA)1mg/1ml 為樣品,分配成濃度為 0.2、0.4、0.6、0.8、 1.0 mg/1ml 各 20μl,各別加入 1ml 染劑中,算出線型回歸公式,做出 標準曲線。再取蛋白質樣本 20μl 加入 1ml 染劑,測 OD595nm 的吸光值, 並分析蛋白質濃度。. 2.2.14 蛋白質之電泳分析(SDS-PAGE). 23.
(34) 實驗所使用的電泳裝置為 Bio-Rad 蛋白質電泳槽,待鑄膠裝 置組合後,先加入分離膠體溶液(separating gel),再加入酒精將上 層膠壓平,待凝固後倒掉酒精並擦拭乾後加入集膠溶液(stacking gel),插上齒膜,膠體凝固後置於電泳槽中,注入電泳緩衝液(running buffer)。取蛋白質樣本溶液 20μl 加入等量的 sample loading dye 20 μl,混勻後在 100 ℃加熱五分鐘後,迅速置於冰上再注入樣本槽 10 μl,並注入蛋白質標準溶液以為對照,先以 80V 進行電泳,待藍色染 劑跑到分離膠體時,將電壓調整為 110V 繼續電泳,直到藍色染劑跑 到膠體底部。關閉電源將膠體取出,用 Commassie brilliant blue 染色 再用脫色液(Destain solution)脫色到蛋白質色帶清楚。. 2.2.15 西方墨點法 (Western blotting) 實驗所使用的電泳裝置為 Bio-Rad 半乾式的電泳轉漬槽, SDS-PAGE 後,切除集膠體部份,膠片浸在轉漬緩衝液(Transfer buffer)中。取轉印紙(硝化纖維紙 nitrocellulose paper)切成膠片大 小浸在轉漬緩衝液,先鋪上三張濕潤的 3M 濾紙,後鋪上濕潤的轉 印紙,疊上已濕潤的膠片,再鋪上三張濕潤的 3M 濾紙。(注意! 過程需趕走氣泡),轉印紙那面位向正極,膠片那面位向負極,以. 24.
(35) 200 mA 轉印 100 分鐘。之後,將轉印紙放入 1x TBST 含 5% 脫脂 牛奶,室溫搖盪一小時,以填塞沒有蛋白質轉印上去的空間。倒掉 進行 Blocking 的牛奶,加入第一次抗體(1:1000 稀釋),置於室 溫搖擺一至二小時或放置 4℃隔夜。倒去第一次抗體或回收重覆使 用,以 1 x TBST 清洗七分鐘三次。再加入第二次抗體(1:1000 稀 釋),置於室溫搖擺一至二小時。倒去二次抗體,轉印紙以 1 x TBST 清洗七分鐘三次。加入 NBT/TCIP 顯色劑,反應至呈色(約二十分 鐘以內)。加入二次水以終止反應,晾乾後保存。. 2.2.16 DNA 轉染(DNA transfection) 細胞的轉染是利用 Arrest In(AI) Transfection Kit 試劑。 轉染實驗前日將細胞分盤到 6 well plate 中培養至隔夜,細胞約六分 滿進行轉染。將 3μg/well 的 DNA 以 0.5m/well 不含胎牛血清的培養 液稀釋。將加約 15μl 的 AI Transfection Kit Reagents 以 0.5ml/well 不 含胎牛血清的培養液稀釋,將以上兩者混合均勻靜置於室溫十五分 鐘,使得 AI/DNA complexes 形成。將要轉染的細胞移去舊的培養液, 將 AI/DNA complexes 加入,於 37.0°C, 5% CO2 下培養 4 到 5 小時, 在更換含胎牛血清的培養液繼續培養 48 至 72 小時。 25.
(36) 2.2.17 冷光測試 ISRE 啟動子的活性(measure ISRE activity by luciferase assay) 將細胞單層培養於 6-well plate 約 6-7 成滿,再共同轉染 pISRE-luc 及 pRunilla-luc 兩種報導基因質體。隔天以 trypsin 將細胞 重新懸浮,平均分於 24- well plate。待細胞穩定貼附於 well 中,分別 加入干擾素(INF-α/β),四小時後以 PBS 清洗三次,加入 Lysis buffer 100 μl (Sigma)反應 20 分鐘,吸取 lysis 後的細胞上清液 20 μl 加入 96-well,再加入 luciferase substrate 100 μl (sigma) ,偵測冷光值。 之後加入 stop solution100 μl (sigma)再次偵測冷光值。. 2.2.18 噬菌體的生長與儲存(Growth and Storage of Bacteriophage T7 ) 先將BLT5403的大腸桿菌劃在含Ampcillin抗生素的培養基 中,隔夜,挑單一個菌落搖菌,37℃ 培養箱培養到OD600nm值到 0.6~1 後加入噬菌體,持續搖晃培養到有細菌溶解發生,就可以離心收集上 清液,分管儲存。每管加入1/10體積的80%glycerol在-80℃ 冰箱保存。. 2.2.19 測定噬菌體的效價(phage titer determination) 將噬菌體做序列稀釋,取適當濃度噬菌體 100 μl 加到 250 μl. 26.
(37) BLT5403 菌液混合均勻,再加上約 55℃的 Top agar 3ml 搖晃一下, 未凝固前迅速倒在 plate 中。37℃培養二到三小時後噬菌斑產生後計 數噬菌斑(plaque)的數目。(圖 2.1). 2.2.20 噬菌體篩選(bio-panning) 將欲實驗的日本腦炎病毒蛋白溶於 coating buffer,且蛋白質濃 度調整 5μg/ml,分注各 100μl 於 30 孔的 96 well ELISA 測試盤中,三 種濃度各兩盤,於 4℃靜置隔夜,未進行 bio-panning 的 plate 先置於 4℃ 備用,倒去孔內液體。分別在每個孔加入 100μl 的 5% 脫脂牛奶 在室溫中搖晃一個小時進行 blocking。倒掉 blocking 的牛奶用 1 X TBST 200μl,清洗七分鐘各三次。在只有牛奶的 well 加入噬菌體在 室溫中搖晃三十分鐘。比例分三區(80μl phage. /. 40μl phage+ 40μl. TBST / 20μl phage+ 60μl TBST)之後再將噬菌體加到相對的 well 在 室溫中搖晃三十分鐘。再用 1X TBST 200μl,清洗七分鐘各五次。加 入濃度 100μg/ml 的相同蛋白質溶液進行 elution 在室溫中搖晃三十分 鐘。將沖洗下來的噬菌體加到 10ml BLT5403 E.coli / 50ml 離心管。 搖到有看到細菌溶解,就可以離心收上清液。其篩選流程如下頁圖示。. 27.
(38) 2.2.21 噬菌體酵素連結免疫吸附分析(phage enzyme-linked immunosorbent assay) 經過六次噬菌體篩選後,將最後一次噬菌體篩選的噬菌體進 行效價測定(phage plaque assay) ,從噬菌體溶菌班中挑選 50 管的噬 菌體,各以 5mlM9TB 培養液, 37℃培養隔夜後,離心 8000rpm 15 分鐘收集上清液。將噬菌體 coated 在 96 well 的 ELISA plate,4℃靜 置隔夜或 37 ℃ 1 小時 (10μl/well 的 phage 加 90μl/well 的 coating buffer)。到掉上清液,加 5%脫脂牛奶,每一個 well 200μl,室溫中搖 晃一個小時。再以 1 X TBST 200μl,清洗七分鐘各三次。在每個孔 28.
(39) 中加入蛋白質樣本濃度約 20μg/ml,100μl,室溫中搖晃一個小時。之 後以 1X TBST 100μl/well, 室溫中清洗七分鐘各三次。加第一次抗 體,anti-his Ab ( 1μl/ml TBST , 100μl / well ) 室溫中搖晃一個小時。 之後以 1X TBST 100μl/well,室溫中清洗七分鐘各三次。加第二次抗 體 anti-mouse Ab – peroxidase ( 1μl/ml TBST , 100μl / well ) 室溫中搖 晃一個小時。之後以 1X TBST 100μl/well,室溫中清洗七分鐘各三次. 接著加入 TMB solution(50μl / well)約四十分鐘。到顏色有改變, 加入 stop solution(50μl / well )立刻測 OD 450nm 吸光值不可超過三 十分鐘。. 2.2.22 噬菌體 DNA 製備( Rapid purification of phage sequencing templates) 經過噬菌體酵素連結免疫吸附分析,挑出十六個OD 450nm 吸光值 反應較高的噬菌體依上述方法進行增殖。第一次離心後取含噬菌體上 清液1ml到新的1.5ml離心管,加入200 μl (PEG2000/ 2.5M NaCl)混合 後,室溫中搖晃一小時。離心,10K rpm十分鐘,小心的完全去掉上清 液,將沈澱物溶於200 μl 二次水中,再加入200 μl phenol/ chloroform。 離心12,000 rpm 十五分鐘。取上清液 150μl 加入300μl ethanol。在室 溫中靜置十分鐘。離心,10,000 rpm 十 分鐘,小心的完全去掉上清液. 29.
(40) 用70% ethanol清洗沈澱物置於室溫中晾乾,再加入50 μl 二次水。之 後將噬菌體DNA進行PCR或是進行定序反應。. 2.2.23 噬菌體蛋白質-酵素連結免疫吸附分析(phage protein- enzyme linked immunosorbent assay) 將噬菌體篩選出的所表現的胜肽序列接到 pET32a 載體上進行 蛋白質表現並純化。一組先將相對應的日本腦炎病毒蛋白 coated 在 96 well 的 ELISA plate 的一半,另一半加入 BSA (濃度 20μg/ml, 100μl/well 的),4℃靜置隔夜或 37 ℃ 1 小時。隔天,到掉上清液,加 5%脫脂牛奶,每一個 well 200μl,室溫中搖晃一個小時。再以 1 X TBST 200μl,清洗七分鐘各三次。之後,兩組各加入序列稀釋的噬菌 體胜肽蛋白( 100μg/ml、10μg/ml、1μg/ml、10-1μg/ml、10-2μg/ml、 10-3μg/ml , 100μl / well ) 室溫中搖晃一個小時。之後以 1X TBST 100μl/well,室溫中清洗七分鐘各三次。加第抗體 anti-S protein Ab –HRP ( 1μl/ml TBST , 100μl / well ) 室溫中搖晃一個小時。之後以 1X TBST 100μl/well,室溫中清洗七分鐘各三次. 接著加入 TMB solution(50μl / well)約四十分鐘。到顏色有改變,加入 stop solution (50μl / well )立刻測 OD 450nm 吸光值不可超過三十分鐘。 2.2.24 共同免疫沉澱法(Co-immunoprecipitation). 30.
(41) 在每組試管加入蛋白質,取其中一個蛋白質的專一性抗體 3 μg,加 TSET buffer 使總體積至 300 μl,在 4℃混合作用 4 小時,接 著加入 50 μl (5 mg)的 protein A-Sepharose beads 懸浮液。接著於 4℃ 下旋轉作用 1 小時,在 4℃10,000 rpm 離心 20 秒鐘,去上清液。以 TSET buffer 洗一次,NET buffer 洗三次,每次沖洗均以 10,000 rpm 離心 20 秒,並去上清液。加入 2X sample loading buffer 50 μl,混勻 後在 100 ℃加熱 5 分鐘後迅速置於冰上,離心 10,000 rpm,20 秒鐘, 取上清液進行 SDS-PAGE 電泳。. 2.2.25 共同轉染免疫螢光染色法(confocal immunofluorescence assays) 將細胞平均培養於有蓋玻片放置的 6-well plate 槽內,培養至隔 天,使細胞約六成滿。再把可以產生綠色或紅色螢光的質體,利用 AI Transfection Kit 共同轉染進入細胞內,其轉染技術與前述相同。 36-48 小時後,以 PBS 小心清洗,再以試鏡紙吸乾 PBS。再將蓋玻片 慢慢傾斜貼附於載玻片上,蓋玻片的四周圍以指甲油固定。指甲油乾 燥後再以共軛焦顯微鏡(Leisa TCS SP2)觀察之。. 2.2.26 免疫染色(immunostain). 31.
(42) 將細胞平均培養於有蓋玻片放置的 6-well plate 槽內,隔天吸去 培養液,用 PBS 洗滌兩次去除殘留的培養液,以福馬林固定(fixing), o. 乾燥後再加入 acetone。加入 anti-JEV 抗體 37 C 反應一小時,再以 PBS 洗滌三次去除抗原抗體非專一性結合,在加入 mouse-conjugated FITC(1:200),PBS 清洗後利用倒立式螢光顯微鏡觀察。. 32.
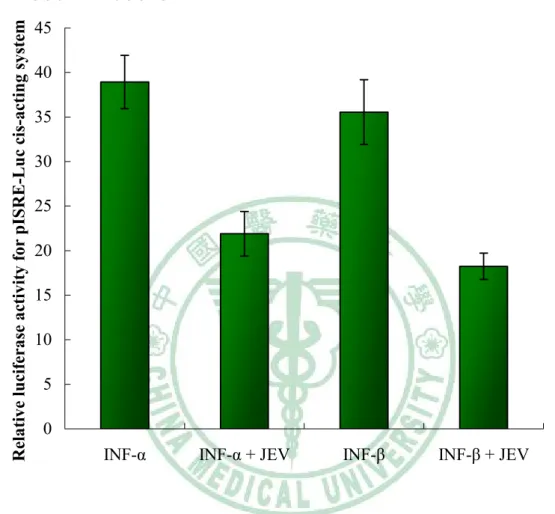
(43) 三、 結果 3.1 日本腦炎抵抗干擾素反應即訊息傳遞的機制 3.1.1 日本腦炎病毒感染後,可以抑制第一型干擾素反應 將 BHK-21 細胞單層平鋪培養在 6-well 盤中,再共同轉染 pISRE-luc 及 pRunilla-luc(9:1) ,以 Runilla 作為 internal control,,隔天分盤於 24-well 盤。將細胞分為先感染病毒後治療 干擾素,以及先治療干擾素後感染病毒兩組。先感染病毒:感染病毒 一小時後,再治療第一型干擾素四小時,再測冷光值。結果顯示,無 論是以α-干擾素或是β干擾素來治療 BHK-21 細胞,在先感染日本腦 炎病毒後治療干擾素的實驗中,可以明顯發現細胞受到日本腦炎病毒 的感染,可以抵抗第一型干擾素的治療。其 ISRE 活性所表現的冷光 值被抑制(Fig. 1B);而在先治療第一型干擾素而後再感染病毒的實 驗中,可以發現第ㄧ型干擾素受到日本腦炎病毒感染,卻沒有十分明 顯的抑制現象(Fig. 1A)。. 3.1.2 日本腦炎病毒 NS4a、NS4b DNA 重組的質體構築 第一步先將日本腦炎 T1P1 病毒株進行病毒增殖放大,再用病毒 蝕斑試驗測試病毒濃度效價(Fig.2),從病毒液中抽取出日本腦炎病. 33.
(44) 毒的 RNA,進行反轉錄進而得到病毒的 cDNA。利用反轉錄所得的 cDNA 進行聚合酶連鎖反應(PCR),用 NS4a-forward、NS4a-Reverse 及 NS4b-forward、NS4b-Reverse 的引子(primer)進行 PCR。將 PCR2 的產物進行 DNA 純化後,利用 EcoR I 和 Xho I 的限制酵素作用切位 處理,再接到同樣切位的 pcDNA3.1-HisB 的載體上,進行質體選殖。 選殖到的質體轉型到大腸桿菌(Top 10)中,大量增殖細菌表現 DNA。 抽取大量細菌的質體,進行質體 DNA 的純化。. 3.1.3 細胞內表現日本腦炎病毒 NS4a、NS4b 蛋白 將質體 DNA pcDNA3.1,pcDNA3.1-NS4a,pcDNA3.1-NS4b 分別轉 染近 BHK-21 細胞與 TE671 細胞。載體上有 neomycin resistance 基 因,因此可以利用 G418 加入細胞培養液中篩選細胞。經過約 20 天的 篩選,將細胞培養於含玻片的 6-well 盤,進行細胞免疫染色,以抗 日本腦炎的老鼠血清作為第一級抗體,所得細胞免疫染色法的結果 為,有轉染 pcDNA3.1-NS4a,pcDNA3.1-NS4b 的 BHK-21 與 TE671 細 胞經過免疫染色法後,於螢光顯微鏡下觀察可見到強烈的綠色螢光 (Fig. 3)。接下來再將細胞收取下來,進行免疫沉澱的實驗來濃縮細 胞中質體表現的蛋白。利用 cell lysis buffer(含蛋白酶抑制劑)溶 O. 解細胞,加入 anti-Xpress(1:100)於 4 C 隔夜,而以 protein A 作用. 34.
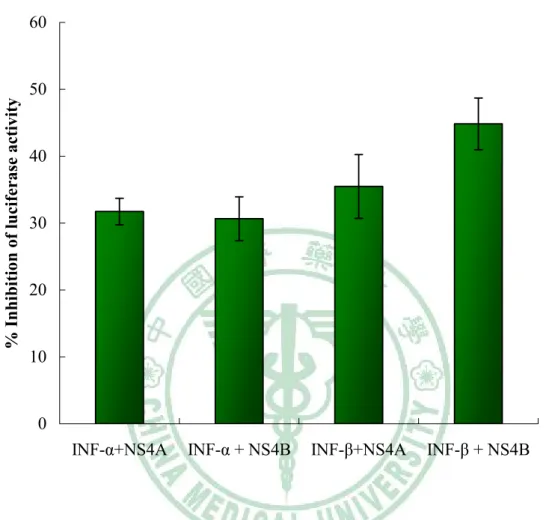
(45) 後的沉澱物進行西方墨點法檢測蛋白,可發現 BHK-21 細胞(Fig. 4A) 及 TE671 細胞(Fig. 4B)中日本腦炎病毒非結構蛋白 NS4A 及 NS4B 蛋 白的表現。. 3.1.4 JEV-NS4A 與 JEV-NS4B 降低 ISRE 啟動子的活性 將穩定表現日本腦炎病毒非結構蛋白 NS4A 及 NS4B 的 BHK-21 及 TE671 細胞,培養於 6-well 盤中。將 pISRE-luc 及 pRunilla-luc 以 9:1 的比例轉染進入細胞中,而 Runilla 作為 internal control。 可以發現在 BHK-21 細胞中,穩定表現 NS4A 蛋白的細胞,加入 3000 U/ml 的α-干擾素後其 ISRE 啟動子的活性抑制的比例高達約 105%, 而加入 3000 U/mlβ-干擾素後其 ISRE 抑制比例約 60%;但穩定表現 NS4B 蛋白的細胞,再加入α/β-干擾素後,其 ISRE 活性抑制程度約 10~20%(Fig. 5A)。在 TE671 細胞中,穩定表現 NS4A 蛋白其抑制 ISRE 活性的程度約 30~40%;而 NS4B 蛋白抑制α-干擾素程度約 30%,β干擾素受到的抑制程度大約 45%(Fig. 5B)。 利用 VSV 病毒可誘發細胞產生第一型干擾素的特性,將表現日本 腦炎病毒非結構蛋白 NS4A 及 NS4B 的 BHK-21 細胞以 pISRE-luc 及 pRunilla-luc 以 9:1 的比例轉染進入細胞中,而後再感染 VSV 病毒. 35.
(46) (MOI=5)36 小時。可以發現在 BHK-21 細胞中,穩定表現 NS4A 及 NS4B 蛋白的細胞可以有效抑制 VSV 病毒感染時,所誘發的干擾素(Fig. 5C)。 3.1.5 JEV-NS4A 與 JEV-NS4B 藉由降低 STAT2 磷酸化來阻撓干擾素 的訊息傳遞 將穩定表現日本腦炎病毒非結構蛋白 NS4A 及 NS4B 的 TE671 細 胞培養於 75T-flask,培養至 7 分滿時再加入 3000 U/ml 的干擾素α/β ,反應半小時後刮取細胞,離心後將沉殿的細胞加入 lysis buffer,再以西方墨點法檢測 stat2 磷酸化的情形。結果發現,TE671 細胞內穩定表現 NS4A 及 NS4B 蛋白,Stat2 磷酸化的程度明顯降低許 多(Fig. 6)。因此,日本腦炎病毒的 NS4A 及 NS4B 蛋白可以藉由降低 Stat2 的磷酸化,來阻饒第一型干擾素的訊息傳遞。. 3.2 鑑定與日本腦炎病毒非結構蛋白 NS4A 作用的細胞因子 3.2.1 日本腦炎病毒 NS4A 蛋白的質體構築 將 NS4a PCR 產物純化後,利用 EcoR I 與 Xho I 的酵素作用切 位處理,再接到同樣切位的 pET24a 載體上,利用質體轉型進入大腸 桿菌(TOP 10)中進行質體選殖(fig 7),抽取質體 DNA 後,經由限制 36.
(47) 0. 酵素切割 DNA 反應 37 C 3 小時,利用洋菜膠電泳,觀察切割之後的 DNA 片段(Fig. 8)。選殖到的質體轉型到能表現蛋白質的大腸桿菌 (BL21 DE3)中,進行大量蛋白表現。. 3.2.2 日本腦炎病毒 NS4A 蛋白質大量表現及純化 能表現日本腦炎病毒 NS4A 蛋白的質體經由轉型到能表現蛋白 質的大腸桿菌(BL21 DE3)中,進行大量蛋白的過度表現,再經由 FPLC(IMAC,His-binding)將蛋白質純化回收。日本腦炎非結構蛋白 NS4A 為 92 個胺基酸所構成的蛋白質,經由西方墨點法(Western Blotting)確定得到約為 14 KDa 的 NS4A 蛋白質(Fig 9B)。亦可從 SDS-PAGE 利用考馬仕藍染劑染色,觀察 NS4A 蛋白質純化後的程度 (Fig 9A)。. 3.2.3 以日本腦炎病毒 NS4A 重組蛋白進行噬菌體篩選 用 His-bind 的方法經由 FPLC 純化後所得的 NS4A 蛋白,coated 在 96 –well 上進行噬菌體的親和性篩選(affinity selection), 與 NS4A 蛋白具有高度結合力的噬菌體沖洗出來加到 BLT5403 細菌中 增殖放大再進行下一次的篩選(bio-panning)。經過至少六次(NS4A 蛋白濃度:20μg/ml)的噬菌體篩選,每次篩選過後都必須用噬菌體 37.
(48) 蝕菌斑試驗(phage plaque assay)確定噬菌體的濃度是否足夠。而在 最後一次的噬菌體噬菌斑實驗中,選擇四十七個噬菌斑(Fig. 10)進 行噬菌體增值放大。隔天,離心取上清液的噬菌體做噬菌體酵素連結 免疫吸附分析(ELISA) (Fig. 11),從 ELISA 試驗結果中挑選出 OD 450nm 吸光值較高十個噬菌體,將其所帶有人腦基因的片段經由 PCR 後,由 洋菜膠電泳觀察 PCR 片段大小(Fig. 12)。再將 PCR 產物定序,從噬 菌體開始表現人腦因子的位置轉譯成氨基酸序列,再將轉譯出的胜肽 序列在 NCBI 網站上進行比對,比對的結果可以分為兩群,結果得知 其中ㄧ群樣品 11. 23.37.39.43 為 DDX42(DEAD BOX)protein(表 1.1) 有高達 100%相似性,佔所挑選出的噬菌體中的十分之五,其序列主 要是在於 DDX42 蛋白的 C'端的 Domain;另ㄧ群樣品 42.44.43 定序 為 SYNAPTOSOMAL-ASSOCIATED protein(SNAP25)有 91%相似性,佔所 挑選出的噬菌體中的十分之三;樣品 16.19 為未知的蛋白,帶有不同 人腦因子 cDNA 所表現的蛋白質序列。. 3.2.4 Dead box42 蛋白的質體構築與純化 設計好帶有限制酶酵素切位的引子後,將噬菌體所帶有 DDX 基 因的片段經由 PCR 後(第 23 號樣品),利用 BamH I 和 Hind III 酵素 切割,再接合到同樣切位的 pET32a 載體上,,轉型進入大腸桿菌 BL21. 38.
(49) 大量表現蛋白,其載體轉譯後所表現的蛋白會帶有約 20kDa 的 Tag fusion protein。而載體上帶有的 His tag 經由轉譯後,也可以利用 FPLC(IMAC,His-binding)將表現的蛋白質純化回收,利用 SDS-PAGE 考馬仕藍染色以及 western blotting 觀察 DDX42 蛋白純化的結果 (fig. 13A.B)。. 3.2.6 人腦細胞蛋白 DDX42 protein 與病毒蛋白 NS4A 共同免疫沉澱 為了證明經由噬菌體表現所得的人腦細胞帶蛋白 DDX42,其是 否與日本腦炎病毒的非結構蛋白 NS4A 有所交互作用,因此利用共同 免疫沉澱法證明之。實驗分為實驗組與對照組,實驗組主要是觀察 DDX 是否與病毒蛋白 NS4A 有交互作用的情形;而對照組主要是排除 載體 pET32a 上帶有 Thioredoxin 的 fusion protein 會與病毒蛋白 NS4A 結合。在實驗中利用抗體(NS4A 蛋白上的 T7 tag)作用經過共同 免疫沉澱,再用西方墨點法觀察。結果顯示,從西方墨點法可以觀察 到在實驗組中的 pellete 裡含有 NS4A 蛋白和 DDX42 蛋白,則表示兩 蛋白質有交互作用的現象(fig.14 ,lan3 lan4 );在對照組中可以明 顯觀察到,病毒蛋白與載體上的 fusion protein 與病毒蛋白 NS4A 分 別分布在 pellete 與上清液中,表示兩者彼此之間無交互作用. 39.
(50) (fig.14 ,lan1 lan2 )。. 3.2.7 共同轉染質體觀察細胞內蛋白質交互作用的關係 將日本腦炎病毒蛋白 NS4A 接合到可以產生綠色螢光蛋白的載 體 pcDNA3.1-EGFP,經過質體選殖(Fig.15)轉型進入大腸桿菌(TOP 10) 中;在將細胞蛋白 DDX42 接合到可以產生紅色螢光的 pDsRed 載體上, 經過質體選殖(Fig. 16)轉型進入大腸桿菌(Top 10)中。抽取兩種細 菌的質體後,以相同濃度共同轉染進入 BHK-21(Fig. 17A)和 TE671(Fig. 17B)細胞,利用共軛焦顯微鏡(confocol)觀察之。觀察 結果發現,在不同的兩種細胞中,紅色螢光的 DDX 蛋白與綠色螢光的 病毒蛋白 NS4A 會坐落在相同的位置,融合後產生明顯的黃色結果 (Fig. 17)。. 3.2.8 過度表現 DDX42 的 BHK-21 細胞,抵抗日本腦炎病毒抑制第一 型干擾素反應 將 BHK-21 細胞單層平鋪培養在 6-well 盤中,再共同轉染 pDsRed-DDX,pISRE-luc 及 pRunilla-luc(5:5:1) ,以 Runilla 作 為 internal control,,隔天分盤於 24-well 盤。將細胞分為先感. 40.
(51) 染病毒後治療干擾素,以及先治療干擾素後感染病毒兩組。先感染病 毒:感染病毒一小時後,再治療第一型干擾素四小時,再測冷光值。 結果顯示,以β干擾素來治療有 DDX42 過度表現的 BHK-21 細胞,在 先感染日本腦炎病毒後治療干擾素的實驗中,可以明顯發現細胞受到 日本腦炎病毒的感染,其第一型干擾素反應沒有被抑制,反而因為病 毒感染而促進干擾素生成,其 ISRE 活性所表現的冷光值升高 (Fig.18A );而在先治療第一型干擾素而後再感染病毒的實驗中,並 無發現沒有十分明顯的升高的現象(Fig. 18B)。. 41.
(52) 四、討論 噬菌體呈現技術是利用噬菌體基因所呈現的蛋白質來跟目標蛋 白進行蛋白質之間的交互作用。隨著研究生命科學技術的層面進步與 噬菌體表現技術配合,對於分子生物的基礎研究、生物科技和臨床醫 學已經有廣泛的運用。噬菌體呈現技術可以用來進行尋找配位體、抗 原決定位(epitope) 、單株抗體的 Fab 或是找出蛋白質功能性區域, 再生物藥學方面進一步可以開發藥物,或是製造疫苗、抗體等。 在本研究所要探討的是日本腦炎病毒蛋白,其在感染宿主細胞 時,其非結構蛋白 NS4A 與 NS4B 是否抑制第一型干擾素的反應。而其 NS4A 在感染宿主細胞時,是與哪個宿主細胞內的何種蛋白結合交互 作用來進行對宿主細胞下一個影響反應。. 4.1 日本腦炎的抗干擾素作用 在先前的文獻報導(J.Viology 2004 Sep)指出,日本腦炎病毒可 以阻礙第一型干擾素的訊息傳遞,進而抑制干擾素反應,且主要抑制 現象,在於日本腦炎病毒可以阻撓細胞膜上的第一型干擾素接受器的 Tyk2磷酸激酶進行磷酸化的現象,進而使下游的stat無法磷酸化。而 再另外的文獻已經有發現,同屬黃病毒科內的西尼羅河病毒、黃熱病. 42.
(53) 毒、登革熱病毒的NS4B可以抑制訊息傳遞路徑的stat1磷酸化 (JOURNAL OF VIROLOGY, July 2005)。因此,本篇實驗第一部分主要 發現,日本腦炎病毒確實能夠抵抗第一型干擾素的反應。且在病毒先 感染細胞而後再進行干擾素的治療時,抑制的現象較為明顯。所以, 當已經確定日本腦炎病毒確實能夠抵抗第一型干擾素後,再構築能表 現日本腦炎非結構蛋白NS4A與NS4B的質體DNA,並穩定轉染進入細胞 (BHK-21與TE671),使BHK-21及TE671細胞能夠穩定表現NS4A、NS4B, 再轉染ISRE啟動子的報導基因,進而藉由冷光值的高低來判別NS4A、 NS4B是否可以當作日本腦炎病毒對干擾素的抑制劑,我們發現在 BHK-21細胞中,病毒蛋白NS4A蛋白能夠非常明顯的抑制干擾素的反 應,而NS4B則較少;然而在TE671細胞中,病毒蛋白NS4A與NS4B其抑 制ISRE活性相差不多。 因此,當證明了 NS4A 可以抑制干擾素作用後,接下來研究的部 分主要是在於訊息傳遞的部份。主要在證明,穩定表現日本腦炎病毒 非結構蛋白 NS4A、NS4B 是否能夠藉由抑制干擾素接受器的磷酸激酶 磷酸化,來阻撓訊息傳遞,進而使 NS4A 及 NS4B,抗干擾素更加確定, 然而本篇實驗發現,病毒蛋白 NS4A 與 NS4B 可以降低細胞內 Stat2 的 磷酸化現象,藉以影響干擾素的訊息傳遞。所以,接下來的就是在病 毒感染細胞時,NS4A 與 NS4B 到底是與細胞內的何種蛋白質有交互作. 43.
(54) 用的現象。. 4.2 NS4A 蛋白與細胞內蛋白的交互作用 經過六次不同日本腦炎病毒蛋白質濃度的噬菌體篩選,噬菌體已 有相當的篩選,但是還是進一步以 ELISA 實驗,所測的 OD. 450nm. 吸光. 值高低來作為挑選噬菌體的依據。再將噬菌體 DNA 純化進行 PCR 後, 進行 PCR 產物的基因定序。將定序的結果從噬菌體開始表現人腦 cDNA 的位置開始進行轉譯,所得到氨基酸序列進行比對。所得大概為三種 蛋白,其一 100%序列相似者為所謂的 DEAD BOX43 蛋白,其主要功能 為類似 ATP 依賴的 RNA 的解旋酶;其二 100%相似者為 SNAP 蛋白,其 主要功能與蛋白質的運輸有關;其三為未知的蛋白(novel protein)。 本篇論文證明利用噬菌體表現找出的兩個蛋白質,是否與日本腦 炎病毒的 NS4A 有所交互作用。因此,將所挑選到的噬菌體表現在表 面蛋白基因序列接在 pET32a 載體上進行噬菌體蛋白質表現,再進行 純化。接著純化後的 NS4A 與純化後的 DDX 蛋白利用共同免疫沉澱-西 方墨點法來證明兩者之間確實有所交互結合作用的關係。進而構築噬 菌體表現在表面蛋白基因序列於 pDsRed 這個可以表現紅色螢光的載 體上;而 NS4a 則接在能表現綠色螢光蛋白(GFP)的載體 pcDNA3.1-EGFP 上。而這兩者以等量的核酸濃度,共同轉染進入細胞 內,於共軛焦顯微鏡下觀察是否兩者會座落於細胞同一個地方. 44.
(55) (co-locolization)。而本論文也證明出,在 TE671 與 BHK-21 細胞中, 病毒蛋白 NS4A 與細胞蛋白 DDX43 在細胞內有 co-locolization 的現 象。 而未來研究計畫的部份:除了證明細胞內 DDX42 蛋白如果有過度 表現,若在病毒感染時,是否會影響病毒抵抗第一型干擾素反應,而 在 C 型肝炎病毒裡面已經被發現,C 型肝炎的 NS4A 蛋白可作為 NS3 蛋白的輔因子,而 NS3 蛋白裡有一段 Domain 亦為 DDX box,其中相 互關係也有待證明;還有日本腦炎病毒非結構蛋白 NS4B 與細胞因子 之交互作用的關係,藉由分子生物學及蛋白質體學的方法,證明病毒 蛋白 NS4A、NS4B 與細胞因子交互作用關係後,再探討這數種蛋白質 之間的關係。. 45.
(56) 五、參考文獻 1. Amberg, S. M., Nestorowicz, A., McCourt, D. W., and Rice, C. M. (1994). NS2B-3 proteinase-mediated processing in the yellow fever virus structural region: in vitro and in vivo studies. J Virol 68(6), 3794-802. 2. Andre, S., Seed, B., Eberle, J., Schraut, W., Bultmann, A., and Haas, J. (1998). Increased immune response elicited by DNA vaccination with a synthetic gp120 sequence with optimized codon usage. J Virol 72(2), 1497-503. 3. Chang, T. H., Liao, C. L., and Lin, Y. L. (2006). Flavivirus induces interferon-beta gene expression through a pathway involving RIG-I-dependent IRF-3 and PI3K-dependent NF-kappaB activation. Microbes Infect 8(1), 157-71. 4. Chang, Y. S., Liao, C. L., Tsao, C. H., Chen, M. C., Liu, C. I., Chen, L. K., and Lin, Y. L. (1999). Membrane permeabilization by small hydrophobic nonstructural proteins of Japanese encephalitis virus. J Virol 73(8), 6257-64. 5. Chen, Y., Maguire, T., and Marks, R. M. (1996). Demonstration of binding of dengue virus envelope protein to target cells. J Virol 70(12), 8765-72. 6. Chu, P. W., and Westaway, E. G. (1992). Molecular and ultrastructural analysis of heavy membrane fractions associated with the replication of Kunjin virus RNA. Arch Virol 125(1-4), 177-91. 7. Deblandre, G. A., Marinx, O. P., Evans, S. S., Majjaj, S., Leo, O., Caput, D., Huez, G. A., and Wathelet, M. G. (1995). Expression cloning of an interferon-inducible 17-kDa membrane protein implicated in the control of cell growth. J Biol Chem 270(40), 23860-6. 8. Der, S. D., Zhou, A., Williams, B. R., and Silverman, R. H. (1998). Identification of genes differentially regulated by interferon alpha, beta, or gamma using oligonucleotide arrays. Proc Natl Acad Sci U S A 95(26), 15623-8. 9. Diamond, M. S., and Harris, E. (2001). Interferon inhibits dengue virus infection by preventing translation of viral RNA through a PKR-independent mechanism. Virology 289(2), 297-311. 10. Diamond, M. S., Roberts, T. G., Edgil, D., Lu, B., Ernst, J., and 46.
(57) 11.. 12.. 13.. 14.. 15.. 16.. 17.. 18.. 19.. 20.. 21.. Harris, E. (2000). Modulation of Dengue virus infection in human cells by alpha, beta, and gamma interferons. J Virol 74(11), 4957-66. Diaz, M. O., Ziemin, S., Le Beau, M. M., Pitha, P., Smith, S. D., Chilcote, R. R., and Rowley, J. D. (1988). Homozygous deletion of the alpha- and beta 1-interferon genes in human leukemia and derived cell lines. Proc Natl Acad Sci U S A 85(14), 5259-63. Durbin, J. E., Hackenmiller, R., Simon, M. C., and Levy, D. E. (1996). Targeted disruption of the mouse Stat1 gene results in compromised innate immunity to viral disease. Cell 84(3), 443-50. Falgout, B., and Markoff, L. (1995). Evidence that flavivirus NS1-NS2A cleavage is mediated by a membrane-bound host protease in the endoplasmic reticulum. J Virol 69(11), 7232-43. Fitzgerald, K. A., McWhirter, S. M., Faia, K. L., Rowe, D. C., Latz, E., Golenbock, D. T., Coyle, A. J., Liao, S. M., and Maniatis, T. (2003). IKKepsilon and TBK1 are essential components of the IRF3 signaling pathway. Nat Immunol 4(5), 491-6. Foy, E., Li, K., Wang, C., Sumpter, R., Jr., Ikeda, M., Lemon, S. M., and Gale, M., Jr. (2003). Regulation of interferon regulatory factor-3 by the hepatitis C virus serine protease. Science 300(5622), 1145-8. Gale, M., Jr., Blakely, C. M., Kwieciszewski, B., Tan, S. L., Dossett, M., Tang, N. M., Korth, M. J., Polyak, S. J., Gretch, D. R., and Katze, M. G. (1998). Control of PKR protein kinase by hepatitis C virus nonstructural 5A protein: molecular mechanisms of kinase regulation. Mol Cell Biol 18(9), 5208-18. Garcia-Sastre, A., Durbin, R. K., Zheng, H., Palese, P., Gertner, R., Levy, D. E., and Durbin, J. E. (1998). The role of interferon in influenza virus tissue tropism. J Virol 72(11), 8550-8. Crance, J. M., Scaramozzino, N., Jouan, A., and Garin, D. (2003). Interferon, ribavirin, 6-azauridine and glycyrrhizin: antiviral compounds active against pathogenic flaviviruses. Antiviral Res 58(1), 73-9. Goodbourn, S., Didcock, L., and Randall, R. E. (2000). Interferons: cell signalling, immune modulation, antiviral response and virus countermeasures. J Gen Virol 81(Pt 10), 2341-64 Guo, J. T., Hayashi, J., and Seeger, C. (2005). West Nile virus inhibits the signal transduction pathway of alpha interferon. J Virol 79(3), 1343-50. Harcourt, B. H., Sanchez, A., and Offermann, M. K. (1999). Ebola 47.
(58) 22.. 23.. 24.. 25.. 26.. 27.. 28.. 29.. 30.. virus selectively inhibits responses to interferons, but not to interleukin-1beta, in endothelial cells. J Virol 73(4), 3491-6 Heinz, F. X., Stiasny, K., Puschner-Auer, G., Holzmann, H., Allison, S. L., Mandl, C. W., and Kunz, C. (1994). Structural changes and functional control of the tick-borne encephalitis virus glycoprotein E by the heterodimeric association with protein prM. Virology 198(1), 109-17. Jones, M., Davidson, A., Hibbert, L., Gruenwald, P., Schlaak, J., Ball, S., Foster, G. R., and Jacobs, M. (2005). Dengue virus inhibits alpha interferon signaling by reducing STAT2 expression. J Virol 79(9), 5414-20. Khromykh, A. A., and Westaway, E. G. (1997). Subgenomic replicons of the flavivirus Kunjin: construction and applications. J Virol 71(2), 1497-505. Kinney, R. M., Butrapet, S., Chang, G. J., Tsuchiya, K. R., Roehrig, J. T., Bhamarapravati, N., and Gubler, D. J. (1997). Construction of infectious cDNA clones for dengue 2 virus: strain 16681 and its attenuated vaccine derivative, strain PDK-53. Virology 230(2), 300-8. Koch, J. O., and Bartenschlager, R. (1999). Modulation of hepatitis C virus NS5A hyperphosphorylation by nonstructural proteins NS3, NS4A, and NS4B. J Virol 73(9), 7138-46. Lee, C. J., Liao, C. L., and Lin, Y. L. (2005). Flavivirus activates phosphatidylinositol 3-kinase signaling to block caspase-dependent apoptotic cell death at the early stage of virus infection. J Virol 79(13), 8388-99. Liu, W. J., Wang, X. J., Clark, D. C., Lobigs, M., Hall, R. A., and Khromykh, A. A. (2006). A single amino acid substitution in the West Nile virus nonstructural protein NS2A disables its ability to inhibit alpha/beta interferon induction and attenuates virus virulence in mice. J Virol 80(5), 2396-404. Liu, W. J., Wang, X. J., Mokhonov, V. V., Shi, P. Y., Randall, R., and Khromykh, A. A. (2005). Inhibition of interferon signaling by the New York 99 strain and Kunjin subtype of West Nile virus involves blockage of STAT1 and STAT2 activation by nonstructural proteins. J Virol 79(3), 1934-42. Lundin, M., Monne, M., Widell, A., Von Heijne, G., and Persson, M. A. (2003). Topology of the membrane-associated hepatitis C virus 48.
(59) 31.. 32.. 33.. 34.. 35.. 36.. 37.. 38.. protein NS4B. J Virol 77(9), 5428-38. Mahalingam, S., Meanger, J., Foster, P. S., and Lidbury, B. A. (2002). The viral manipulation of the host cellular and immune environments to enhance propagation and survival: a focus on RNA viruses. J Leukoc Biol 72(3), 429-39. Mashimo, T., Lucas, M., Simon-Chazottes, D., Frenkiel, M. P., Montagutelli, X., Ceccaldi, P. E., Deubel, V., Guenet, J. L., and Despres, P. (2002). A nonsense mutation in the gene encoding 2'-5'-oligoadenylate synthetase/L1 isoform is associated with West Nile virus susceptibility in laboratory mice. Proc Natl Acad Sci U S A 99(17), 11311-6. Matikainen, S., Siren, J., Tissari, J., Veckman, V., Pirhonen, J., Severa, M., Sun, Q., Lin, R., Meri, S., Uze, G., Hiscott, J., and Julkunen, I. (2006). Tumor necrosis factor alpha enhances influenza A virus-induced expression of antiviral cytokines by activating RIG-I gene expression. J Virol 80(7), 3515-22. Menozzi, F. D., Reddy, V. M., Cayet, D., Raze, D., Debrie, A. S., Dehouck, M. P., Cecchelli, R., and Locht, C. (2006). Mycobacterium tuberculosis heparin-binding haemagglutinin adhesin (HBHA) triggers receptor-mediated transcytosis without altering the integrity of tight junctions. Microbes Infect 8(1), 1-9. Meraz, M. A., White, J. M., Sheehan, K. C., Bach, E. A., Rodig, S. J., Dighe, A. S., Kaplan, D. H., Riley, J. K., Greenlund, A. C., Campbell, D., Carver-Moore, K., DuBois, R. N., Clark, R., Aguet, M., and Schreiber, R. D. (1996). Targeted disruption of the Stat1 gene in mice reveals unexpected physiologic specificity in the JAK-STAT signaling pathway. Cell 84(3), 431-42. Munoz-Jordan, J. L., Laurent-Rolle, M., Ashour, J., Martinez-Sobrido, L., Ashok, M., Lipkin, W. I., and Garcia-Sastre, A. (2005). Inhibition of alpha/beta interferon signaling by the NS4B protein of flaviviruses. J Virol 79(13), 8004-13. Munoz-Jordan, J. L., Sanchez-Burgos, G. G., Laurent-Rolle, M., and Garcia-Sastre, A. (2003). Inhibition of interferon signaling by dengue virus. Proc Natl Acad Sci U S A 100(24), 14333-8. Ng, L. F., and Liu, D. X. (2002). Membrane association and dimerization of a cysteine-rich, 16-kilodalton polypeptide released from the C-terminal region of the coronavirus infectious bronchitis virus 1a polyprotein. J Virol 76(12), 6257-67. 49.
(60) 39. Niyomrattanakit, P., Winoyanuwattikun, P., Chanprapaph, S., Angsuthanasombat, C., Panyim, S., and Katzenmeier, G. (2004). Identification of residues in the dengue virus type 2 NS2B cofactor that are critical for NS3 protease activation. J Virol 78(24), 13708-16. 40. Olle-Goig, J. E., Rodes, A., and Barrera, J. M. (1993). Prevalence of HIV-1 infection in a rural medical clinic in Haiti. J Trop Med Hyg 96(4), 222-4. 41. Perelygin, A. A., Scherbik, S. V., Zhulin, I. B., Stockman, B. M., Li, Y., and Brinton, M. A. (2002). Positional cloning of the murine flavivirus resistance gene. Proc Natl Acad Sci U S A 99(14), 932242. Percy, N., Barclay, W. S., Garcia-Sastre, A., and Palese, P. (1994). Expression of a foreign protein by influenza A virus. J Virol 68(7), 4486-92. 43. Qu, L., McMullan, L. K., and Rice, C. M. (2001). Isolation and characterization of noncytopathic pestivirus mutants reveals a role for nonstructural protein NS4B in viral cytopathogenicity. J Virol 75(22), 10651-62. 44. Sharma, S., tenOever, B. R., Grandvaux, N., Zhou, G. P., Lin, R., and Hiscott, J. (2003). Triggering the interferon antiviral response through an IKK-related pathway. Science 300(5622), 1148-51. 45. Shresta, S., Kyle, J. L., Snider, H. M., Basavapatna, M., Beatty, P. R., and Harris, E. (2004). Interferon-dependent immunity is essential for resistance to primary dengue virus infection in mice, whereas T- and B-cell-dependent immunity are less critical. J Virol 78(6), 2701-10. 46. Singh, I., and Helenius, A. (1992). Role of ribosomes in Semliki Forest virus nucleocapsid uncoating. J Virol 66(12), 7049-58. 47. Singh, I. R., Suomalainen, M., Varadarajan, S., Garoff, H., and Helenius, A. (1997). Multiple mechanisms for the inhibition of entry and uncoating of superinfecting Semliki Forest virus. Virology 231(1), 59-71. 48. Stadler, K., Allison, S. L., Schalich, J., and Heinz, F. X. (1997). Proteolytic activation of tick-borne encephalitis virus by furin. J Virol 71(11), 8475-81. 49. Taylor, D. R., Shi, S. T., Romano, P. R., Barber, G. N., and Lai, M. M. (1999). Inhibition of the interferon-inducible protein kinase PKR by HCV E2 protein. Science 285(5424), 107-10. 50. Wathelet, M. G., Lin, C. H., Parekh, B. S., Ronco, L. V., Howley, P. 50.
(61) M., and Maniatis, T. (1998). Virus infection induces the assembly of coordinately activated transcription factors on the IFN-beta enhancer in vivo. Mol Cell 1(4), 507-18. 51. Weaver, B. K., Kumar, K. P., and Reich, N. C. (1998). Interferon regulatory factor 3 and CREB-binding protein/p300 are subunits of double-stranded RNA-activated transcription factor DRAF1. Mol Cell Biol 18(3), 1359-68. 52. Yoneyama, M., Suhara, W., Fukuhara, Y., Fukuda, M., Nishida, E., and Fujita, T. (1998). Direct triggering of the type I interferon system by virus infection: activation of a transcription factor complex containing IRF-3 and CBP/p300. Embo J 17(4), 1087-95.. 51.
(62) 六、圖表 ㄧ、附圖 Fig. 1A. Relative luciferase activity for pISRE-Luc cis-acting system. A. Pre-infection 40 35 30 25 20 15 10 5 0 INF-α. INF-α + JEV. INF-β. INF-β + JEV. 圖 1A. 細胞先治療 α/β-干擾素,在日本腦炎病毒感染時, ISRE 活性的影響 將 pISRE-luc 報導基因、pRunilla-luc 共同轉染進入 BHK-21 細胞,先 治療干擾素兩小時後再感染病毒兩小時,在偵測冷光強度。可以發現, 日本腦炎病毒的感染,使 α/β-干擾素誘發的 ISRE 啟動子活性降低程 度不太明顯。 52.
(63) Fig. 1B. Relative luciferase activity for pISRE-Luc cis-acting system. B. Post-infection 45 40 35 30 25 20 15 10 5 0 INF-α. INF-α + JEV. INF-β. INF-β + JEV. 圖 1B 細胞先感染日本腦炎病毒時,而後治療 α/β-干擾素, 對 ISRE 活性的影響 先將 pISRE-luc 報導基因、 pRunilla-luc 共同轉染進入 BHK-21 細胞,分成先感染日本腦炎病毒而後治療干擾素四小時。可以發現,細 胞先受到日本腦炎病毒的感染,再治療干擾素時,可以使α/β-干擾素 誘發的 ISRE 啟動子活性明顯降低。. 53.
(64) Fig. 2. 病毒濃度稀釋倍數:. 10-4. 10-5. 10-6. 圖 2. 利用病毒蝕班試驗測試病毒增幅放大後的效價 將增幅放大後的日本腦炎病毒(T1P1 strain) 序列稀釋後,再加到 6-well 單層的 BHK-21 細胞加上覆蓋液,三天後觀察及計算病毒蝕斑 (plaque)的數目並計算效價。. 54.
(65) Fig. 3 pCDNA3.1. A.. pcDNA3.1-NS4A. pcDNA3.1-NS4B. B.. C.. E.. F.. BHK-21 cells. D.. TE671 cells. 圖 3.TE671 與 BHK-21 細胞免疫螢光染色 將穩定表現 NS4A 及 NS4B 的 BHK-21 及 TE671 細胞,經過福馬林固定後, 利用抗日本腦炎病毒蛋白的抗體作為一級抗體,Anti-mouse(FITC)做為 二級抗體。經由螢光顯微鏡觀察,發現兩種細胞能表現 NS4A 及 NS4B 蛋 白,其螢光強度甚強。. 55.
(66) Fig. 4A. 55kDa 40 kDa 35 kDa NS4B. 25 kDa 15kDa. NS4A. 10 kDa. 圖 4A 免疫沉澱-西方墨點法檢測 BHK-21 細胞 JEV-NS4A, JEV-NS4B 蛋白表現 BHK-21 含有 pcDNA3.1-empty,pcDNA3.1-NS4a,pcDNA3.1-NS4b 的細胞 收取溶解後,加入 anti-Xpress 抗體(1:100)作為免疫沉澱的抗體,於 4 度 C 搖晃至隔天,而後利用西方墨點法檢測蛋白的存在。Lan 1 為 Marker,Lan 2 為單獨載體,Lan 3 為穩定表現 NS4A 的 TE671 細胞,Lan 4 則為穩定表現 NS4B 的 TE671 細胞。. 56.
(67) Fig. 4B. 55kDa 40 kDa 35 kDa NS4B. 25 kDa. NS4A 15kDa 10 kDa. 圖 4B 免疫沉澱-西方墨點法檢測 TE671 細胞 JEV-NS4A, JEV-NS4B 蛋白表現 同 Fig 5A。TE671 細胞內含有 pcDNA3.1-empty,pcDNA3.1-NS4a, pcDNA3.1-NS4b 的細胞收取溶解後,加入 anti-Xpress 抗體(1:100)作 為免疫沉澱的抗體,於 4 度 C 搖晃至隔天,而後利用西方墨點法檢測蛋 白的存在。Lan 1 為 Marker,Lan 2 為單獨載體,Lan 3 為穩定表現 NS4A 的 BHK-21 細胞,Lan 4 則為穩定表現 NS4B 的 BHK-21 細胞。. 57.
(68) Fig. 5 A BHK-21 cells. % Inhibition of luciferase activity. 120. 100. 80. 60. 40. 20. 0 INF-α+NS4A INF-α + NS4B INF-β+NS4A. INF-β + NS4B. 圖 5A 在治療干擾素下,偵測穩定表現日本腦炎非結構蛋白 NS4A、NS4B 的 BHK-21 細胞所影響 ISRE 的效能 將 pISRE-Luc 報導基因及 pRunilla-luc 分別共同轉染進入三種細胞, BHK21-NS4a ,BHK21-NS4b,隔天加入干擾素並在 4 小時後偵測冷光 I. 在加入 INF-α 之下,可發現穩定表現 NS4A 的 BHK-21 細胞,其 ISRE promoter activity 抑制程度明顯,而 NS4B 則較低 II. 在加入 INF-β 之下,亦可發現 NS4A 可明顯抑制 ISRE 的活性,而 NS4B 較不明顯。. 58.
數據



+7





相關文件
查我國皮蛋已可輸往美國、日本,熟煮 蛋(包括鹹熟蛋)亦可輸往美國、加拿 大、日本、新加坡等國。我國依據世界 動物衛生組織(OIE)規範,禽流感病 毒及新城病病毒經攝氏 60
十七、 因應新型冠狀病毒肺炎(COVID-19)疫情,防疫期間請隨時至衛生福利部疾病 管制署全球資訊網(https://www.cdc.gov.tw)之新型冠狀病毒肺炎(COVID-
SF15140A 楊勝舜 利用 JKB-122 評估對於經干擾素(長效 型或短效型)或干擾素和 Ribavirin 組合 治療沒有反應的 C 型肝炎病毒陽性患 者之肝臟功能 (丙胺酸轉胺酶
拾柒、為因應新型冠狀病毒肺炎(COVID-19)疫情,請所有參與人員配合本局訂定因應新型冠狀病
澳洲爆發傳染力高的 Delta 變異病毒疫情,為了阻止病毒擴散,雪梨(Sydney)自 6 月 26 日起開始封城,墨爾本(Melbourne)、南澳洲也相繼封城,導致澳洲自 7 月
和富蘭克林·史達(Franklin Stahl)於 1958 年進行研究而得以證實。 1952 年赫希與蔡斯利用噬菌 體與細菌進行研究,證明 DNA 在噬菌體可作為遺傳物質(Hershey
碘液測試 澱粉 碘液保持褐色 碘液轉為深藍色 尿蛋白試紙 蛋白質 黃色
一、對抗生素Meropenem Trihydrate 具抗藥性(R 菌)和不具抗藥性(S 菌)的 Acinetobacter baumannii 菌在蛋白質電泳實驗中,比較蛋白質表現的差異。(文中所用代號: 「R
