國立交通大學
生物醫學研究所
碩士論文
MTGFP 融合蛋白摺疊過程中分子內螢光
能量轉移研究
Intramolecular Fluorescence Energy
Transfer Study of MTGFP fusion protein
during protein folding process
研究生:姜芳馨
指導教授:張家靖
MTGFP 融合蛋白摺疊過程中分子內螢光能量轉移研究
Intramolecular Fluorescence Energy Transfer Study of MTGFP fusion proteinduring protein folding process
研究生:姜芳馨 Student:Fang-Hsing Chiang
指導教授:張家靖 Advisor:Chia-Ching Chang
國 立 交 通 大 學
生物醫學所
碩 士 論 文
A Thesis
Submitted to Institute of Biomedical Science
College of Biological Science and Technology
National Chiao Tung University
In partial Fulfillment of Requirement for the Degree of Master inScience
July 2009
Hsinchu,Taiwan,Republic of China
中華民國九十八年七月
I
MTGFP 融合蛋白摺疊過程中分子內螢光能量轉移研究
學生:姜芳馨 指導教授:張家靖教授
國立交通大學生物醫學研究所 碩士班
摘要
為能即時觀測蛋白質摺疊的過程中做蛋白質摺疊狀態的變化,在這個研究中,我們設計 MTGFP (金屬硫蛋白融合綠色螢光蛋白) 蛋白質分子內的螢光共振能量系統(FRET),可 用來偵測金屬結合在金屬硫蛋白結合上。利用過臨界點摺疊路徑進行蛋白質的摺疊,發 現MT 蛋白質在摺疊過程中同時有 FRET 效應的產生。這是個新的技術利用螢光能量共 振轉移做蛋白質摺疊的即時偵測。 金屬硫蛋白具有 61 個胺基酸內有 20 個硫基,並具 有2 個金屬鍵結環。利用波長 395 nm 激發綠色螢光蛋白(GFP),可在 508 nm 放出螢光, 提供螢光環做為FRET 的系統。利用準靜態熱平衡透析法(TED)摺疊 MTGFP 蛋白質, 利用動態光散射儀(DLS)、圓二色光譜儀(CD)去證明蛋白質結構摺疊回自然態.利用感應 耦合電漿原子發射光譜法(ICP-AES)偵測到金屬離子已經摺疊到 MTGFP 蛋白質的內 部,證明MTGFP 具有結合金屬的能力。然而 MTGFP 蛋白質在摺疊過程中,508nm 螢 光波長的強度逐步降低。發現MT 可以做為一個粹熄子(quencher)會將 GFP 的螢光衰減 掉。換句話說,MT 及 GFP 蛋白之間彼此有螢光共振能量轉移(FRET)現象也有粹熄 (quenching)的現象。這是一個新的方式可以用來研究蛋白質摺疊的過程。II
Intramolecular Fluorescence Resonance Energy Transfer Study of
MTGFP fusion protein during protein folding process
Advisee:Fang-Hsing Chiang Adviser:Dr.Chia-Ching Chang
Institute of Biomedical Science
National Chiao Tung University
Abstract
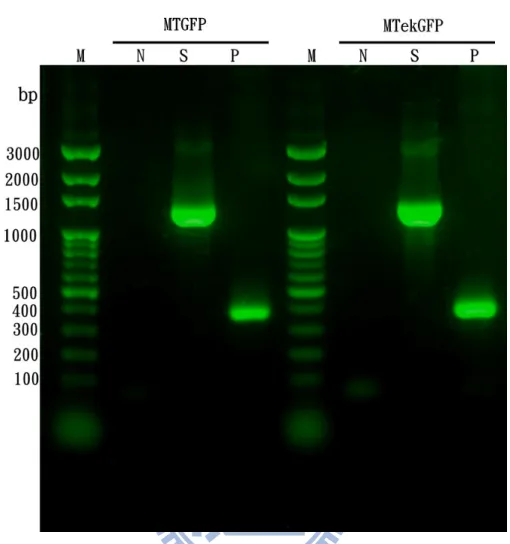
Protein folding status indicator is highly desired for revealing the folding process in real time. Therefore, ultra sensitive detection system in molecular scale is required. In this study, we designed an intramolecular fluorescence resonance energy transfer (FRET) system to monitor the folding status of metal binding protein, metallothionein (MT) by liking with green fluorescence protein (GFP). Both FRET and quench effect took place when the protein folding followed the over-critical reaction path. This new perspectives may provide significant strategy for investigation of protein folding process in real time.Metallothionein (MT) is a 61 amino-acids protein which contains 20 cysteine residues and forms two metal binding clusters (α-, β-domains). Green fluorescent protein (GFP), which emits green fluorescent light (λ max = 508 nm) by exciting with ultraviolet light (395 nm), can be used as a fluorophore in fluorescence resonance energy transfer (FRET) system. A fusion protein of MTGFP was cloned and refolded to native form by stepwise thermal equilibrium dialysis (TED). Both dynamic light scattering (DLS) and circular dichroism (CD) spectra indicate that the fusion protein has been refolded to its native state. The inductively couple plasma-mass (ICP-mass) spectrum also indicates that the refolded-MTGFP has restored its metal binding capability However, the fluorescence intensity of MTGFP at 508 nm declines during the refolding process. Therefore the MT seems acting as a quencher of GFP when it folds to native state. Namely, there is intramolecular fluorescence energy transfer taking place between MT and GFP. This new perspectives may provide significant strategy for investigation of protein folding process.
III
誌謝
謝謝上帝讓我在三年前的颱風天來到張家靖老師這兒參加面試,感謝張老師給我機會 使我能從您的助理變成學生,還有要謝謝老師三年來的教導,常給您製造意外的大小驚 喜,而您總是很忍耐的給我重新學習的機會,尤其是當您在教我數學時,常常會大大的 深呼吸,然後再繼續慢慢的教我時,真的很感謝老師…謝謝您!^_^= 感謝阿木學長(鄭博)謝謝您常常鼓勵我,每次都會跟我說:『很好、很好、這次比上 次更好了。』,每當實驗結果不如預期時,您都叫我不要害怕,阿木學長您就像爸爸一 樣給我安定丸,尤其是口試那天站在台上緊張的傻了,可是您還是一直豎著大拇指,唸 著不錯不錯的唇語,謝謝您! 曾博謝謝您常幫我想辦法解決實驗的問題,也謝謝您跟我 分享工作、家庭、休閒等等的看法,最常聽到您對我說:『不用功』,我會把您的教導記 在心裡,遇到問題時會安靜思考解決的方法。在您一年的指導下獲益良多,謝謝您! 謝 謝師母在實驗上的協助;謝謝親愛的美儀姊姊在實驗及精神上的鼓勵;謝謝銘松學長幫 我測量ICP-AES,謝謝葉桑每天幫我們送信及整理環境,讓我們能專心的做實驗;謝謝 阿亮爸爸這三年辛苦ㄌ,把你的脾氣越磨越好了;愛欺騙的人簡哥,謝謝你讓我提早瞭解 社會真實面;突然當爸的建彰學長謝謝你在我念碩班製造的歡樂;無緣的辰哥謝謝你教我 做實驗、看 paper、還有難過時安慰我;謝謝佩欣、昌祐、輔宣、姿亘對我的教導;謝 謝可愛的小薇、阿良、靜怡、真宜、光凱學長、惠芳學姊、偉賢、昌翰、子歡、明勳、 久安、秀敏,因為有你們大家才能讓整棟生科實驗二館充滿熱鬧的氣氛;終於輪到我的 好朋友們-瑤貞我親愛的同居人,每天跟你一起作息真是幸福;靖淳無理頭搞的笑,讓 實驗室每天都有充滿生氣;子正謝謝你的日出小吃,常幫我們加菜謝謝拉!感謝爸媽、 阿姨們、教會的朋友們-惇方、家薇、佩熹及大學和五專的同學們,謝謝大家的鼓勵及 常常為我禱告,給我很大的力量支持我前進;最後要感謝宏穆-我的寶貝小哥哥,交大 三年的日子,不管我開心難過,你都在我身邊,幾乎每天不管多晚幾乎都護送我平安到 宿舍,又常帶我去吃好料補身體,假日也常陪我在實驗室或教會渡過,有你在我身邊,真 幸福。 在張老師這的三年,讓我學到做研究應有的態度、解決問題的能力、合作團隊的重要, 這三年的點點滴滴-酸、甜、苦、辣,要好好謝謝各位陪我一起成長,深深的謝謝大家-親愛的上帝、老師、曾博、鄭博、家人、情人、及實驗室所有的人,謝謝你們!IV
目錄
摘要... I ABSTRACT ... II 誌謝... III 目錄...IV 表目錄... VII 圖目錄... VIII 第一章 緒論... 1 1-1 研究動機...1 1-2 螢光原理介紹...11-3 螢光共振能量轉移(FLUORESCENCE RESONANCE ENERGY TRANSFER,FRET) ...2
1-4 淬熄效應 (QUENCHING)...5
1-5 生物體發光...5
1-6 綠色螢光蛋白(GREEN FLUORESCENCE PROTEIN,GFP)介紹...6
1-7 金屬硫蛋白(METALLOTHIONEIN,MT)介紹...10
1-7.1 蛋白質結構與摺疊的重要性...12
1-7.2 蛋白質的變性 (denaturation) 與沈澱...12
1-7.3 蛋白質的復性 (renaturation) 與摺疊 (fold) ...13
第二章 實驗原理:... 16
2-1DNA 聚合酶連鎖反應 (POLYMERASE CHAIN REACTION,PCR) ...16
2-2DNA 定序 ...16 2-3 誘導劑 IPTG...17 2-4 內涵體 (INCLUSION BODY)...17 2-5 蛋白質變性與沈澱...18 2-6 各種常見的化學變性劑...18 2-7 蛋白質摺疊 (PROTEIN REFOLDING)...20 2-8 圓二色光譜儀 (CIRCLULAR DICHRORISM,CD)...22
V
2-9 動靜態光散射儀 (DYNAMIC LIGHT SCATTERING,DLS) ...23
2-10 螢光儀(FLUORESCE) ...23
2-10.1 螢光共振能量轉移(Fluorescence Resonance Energy Transfer, FRET)...23
2-11 淬熄效應 (QUENCHING)...25
2-12 感應耦合電漿原子發射光譜法(INDUCTIVELY COUPLED PLASMA ATOMIC EMISSION SPECTROMETRY,ICP-AES) ...26
2-13 腸激酶 (ENTEROKINASE,EK) 酵素切割...28 第三章 實驗方法... 29 3-1 實驗材料的製備...29 3-1.1 重組蛋白質 pET200-MTGFP 及 pET200-MTekGFP 基因鑑定...29 3-2 蛋白質的小量表現與蛋白質的大量表現...30 3-3 蛋白質純化...30 3-3.1 破菌...30 3-3.2 Gel filtration...31 3-4 西方墨點法(WESTERN)...31 3-5 蛋白質摺疊(PROTEIN FOLDING) ...32 3-6 圓二色光譜儀 (CIRCLULAR DICHRORISM,CD)...34
3-7 動態光散射儀 (DYNAMIC LIGHT SCATTERING,DLS) ...34
3-8 感應耦合電漿原子發射光譜法(INDUCTIVELY COUPLED PLASMA ATOMIC EMISSION SPECTROMETRY,ICP-AES) ...34
3-9 螢光光譜儀(FLUORESCENCE SPECTROSCOPY) ...35 3-10ENTEROKINASE 酵素切割...36 第四章 實驗結果... 37 4-1 基因鑑定...37 4-1.1 重組蛋白質 pET200-MTGFP 及 pET200-MTekGFP 基因鑑定結果...37 4-2MTGFP 重組蛋白質的表現...44 4-3GEL FILTRATION 純化蛋白質...45 4-4 西方墨點法確認MTGFP 蛋白質...48 4-5 圓二色光譜儀 (CIRCLULAR DICHRORISM,CD)測量二級結構變化...49
4-6 動靜態光散射儀(DYNAMIC LIGHT SCATTERING,DLS)分析粒子大小...52
VI 4-8 利用摺疊中間物探討蛋白質之穩定度及螢光螢光蛋白的表現...54 4-8.1 利用螢光儀分析 MTGFP 蛋白質其疏水性核心的形成 ...54 4-8.2 利用螢光儀分析 MTGFP 蛋白質觀察綠色螢光蛋白的表現 ...55 4-8.3 3D 全光譜尋找 MT 與 GFP 的螢光基團 ...58 4-8.4 MTGFP 蛋白質在摺疊過程中 FRET 的效應 ...60 4-8.5 螢光圖譜分析...62 第五章 討論... 64 第六章 參考文獻... 66
VII 表目錄 表1 突變 GFP 螢光蛋白種類...10 表2 蛋白質變性的方法 ...13 表3 減少蛋白質分子間錯誤鍵結的添加物 ...14 表4 直接透析與階段性熱平衡透析法比較 ...15 表5 階段性熱平衡透析法的變性藥劑和蛋白質摺疊緩衝液的化學成分 ...33 表6 MTGFP 蛋白質各摺疊中間體的二機結構分佈表...50 表7 ICP – AES 分析摺疊回自然態的 MTGFP 蛋白質金屬含量比例 ...53
VIII 圖目錄 圖1 螢光能階示意圖 ...2 圖2 螢光能量共振轉移 FRET 效應能階圖 ...3 圖3 螢光能量共振轉移 FRET 與距離示意圖 ...4 圖4 CFP 螢光蛋白及 YFP 螢光蛋白的 FRET 螢光光譜...5 圖5 Aequorin 的發光機制示意圖 ...6 圖6 GFP 的發光基團結構...8 圖7 GFP 發光機團與其機制...9 圖8 金屬硫蛋白(Metallothionein, MT)結構示意圖...11 圖9 能量改變示意圖 ...22 圖10 FRET 效應與距離的關係 ...24 圖 11 FRET 強度與距離的示意圖 ...25 圖 12 ICP-AES 架構原理圖 ...26 圖 13 感應耦合電漿激發源之構造示意圖 ...27 圖14 MTekGFP 重組蛋白質內 enterokinase 的切點介紹...36
圖15 pET200-MTGFP 及 pET200-MTekGFP 其 PCR 產物做 DNA 電泳分析 ..38
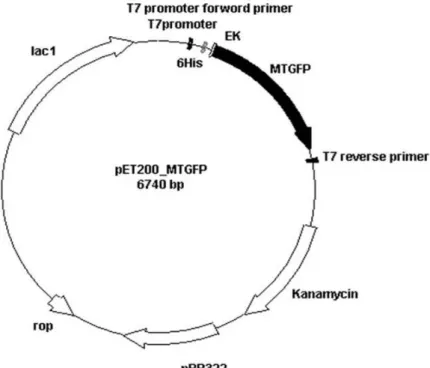
圖16 pET200-MTGFP 質體建構圖...39
圖17 pET200-MTekGFP 質體建構圖...39
圖18 MTGFP 之 SDS-PAGE 分析...44
圖 19 利用 Gel filtration 方式分離 MTekGFP 的蛋白質 ...45
圖 20 Gel filtration (Superdex 200 xk16/40) 分離 GFP 跑膠確認圖 ...46
圖 21 Gel filtration (Superdex 200 xk16/40) 分離 MTekGFP 所收集的樣品 ...46
圖 22 利用 Gel filtration 來第二次純化 MTekGFP 蛋白 ...47
圖23 MTGFP Western 的結果 ...48 圖24 MTGFP 蛋白質各摺疊中間體的 CD 光譜 ...50 圖25 MTGFP 蛋白質各摺疊中間體的二機結構柱狀分佈圖...50 圖26 MTGFP M5 與 GFP 與 MT 與 MTGFP 理想值的二級結構比較...51 圖27 動態光散射儀分析 MTGFP 蛋白質在各摺疊過程中分子粒徑大小分佈 52 圖28 MTGFP 摺疊過程中(激發光 280 nm)各中間體的螢光光譜 ...54 圖29 MTGFP 在摺疊過程中螢光光譜的位移分析(I320/I365)折線圖 ...55 圖30 比較相同濃度下 GFP 與 MTGFP 其(激發光 395 nm)螢光表現...56 圖31 MTGFP 摺疊過程中(激發光 395 nm)各中間體的螢光光譜 ...56
IX 圖32 MTGFP 摺疊過程中各中間體的螢光強度分佈圖(激發光 395 nm) ...57 圖33 MTGFP 摺疊過程中其摺疊中間體的螢光強度衰減圖 (激發光 395 nm) 57 圖34 Commerial MT 3D 螢光立體光譜 ...58 圖35 GFP 的 3D 螢光立體光譜...59 圖36 MTGFP 摺疊過程中(激發光 350 nm)各中間體螢光光譜分佈圖 ...60 圖37 用 350 nm 激發觀察 MTGFP、GFP、MT 是否有 FRET 效應 ...61 圖38 MTGFP 螢光扣掉 GFP 螢光後,分析 MT 在 440 nm 左右的螢光變化 ..62
圖39 Native page 觀察 Enterokinas 切割 MTekGFP 的結果...63
1
第一章 緒論
1-1 研究動機 蛋白質、去氧核醣核酸、生物膜皆屬於生物巨分子,這些巨分子具有自我組織 (Self-organiziation) 形成穩定結構的能力,當生物巨分子處在開放系統的環境中,其 系統內部的分子會與外在環境產生交互作用,而改變構形,使分子趨於能量最低態 的特殊構形[1,2]。 金屬硫蛋白 (Metallothionein, MT) 具有α-,β- 兩個金屬鍵結團簇 (metal cluster/domain)[3-7]。2006 年研究指出,螯合金屬後的 MT ,其 α-,β- metal cluster 形狀類似兩個環狀結構,當β-metal cluster 形成(Mn2CdS3)3-時,可形成磁矩;而α-metal cluster 雖沒有磁性卻扮演穩定β-metal cluster 的角色[8]。然而這環狀結構上的 金屬原子具有自由電子,且這些自由電子會在金屬原子上自由跳動,類似於電子共 振的現象[8],所以我們推測 metal cluster 可能會吸收能量放出螢光或影響其他發光 機團的發光效應。
生物冷光(Bioluminescence)是在大自然中生物所發出的光之總稱,而本實驗中主 要是探討綠色螢光蛋白GFP (Green Florescence protein),而此蛋白質是 1962 年從 水母細胞中被發現,且使用395 及 475 nm 的光激發後,會在 508 及 503 nm 呈現螢 光[9,10]。
螢光蛋白在生物技術上有許多貢獻,例如用螢光蛋白做為指標,分析蛋白質摺 疊狀況的指標[11,12],以及藉由螢光共振能量轉移 (Fluorescence Resonance Energy Transfer, FRET) 的效應快速檢測蛋白質在摺疊過程中能量的變化[13-15]。
然而在掃瞄MT 及 GFP 蛋白質螢光光譜時,發現其 MT 的放射光譜與 GFP 的螢 光激發波長的位置很接近,根據螢光共振能量轉移 (Fluorescence Resonance Energy Transfer, FRET) 理論(圖2) [16],推測 MT 及 GFP 蛋白之間可能會有 FRET 現象, 於是我們合成金屬硫蛋白融合綠色螢光蛋白 (Fusion protein of Metallothionein and green fluorescence protein, MTGFP) 做為實驗材料。觀察及探討 MTGFP 蛋白質在摺 疊過程越接近自然態 (native state) 時,MT 及 GFP 兩蛋白質分子間距離越接近,是 否GFP 產生的螢光能量會有轉移的現象。 1-2 螢光原理介紹 螢光物質受到特定能量的光源激發後,可以產生高或低於原本能量光源的光的 物質。在平常的狀況下,這些螢光物質的電子大多會處於最低能量的狀態,稱為基 態。當分子吸收了與它的特徵頻率一致的特定光源時,其電子便由基態躍遷至能量
2 較高的激發態 (此激發態又依能量的高低區分為各階的激發態)。經過一段時間,處 於不同能階激發態的電子會降至最低能量的激發態,在這一過程中所消耗的能量, 並不以光的形式釋放,這是因為這些螢光分子會和周圍的同類分子或其他分子經歷 震動鬆弛 (vibrational relaxation) 將能量散失掉。當降至較低能量激發態的電子又下 降至基態時,此時會以光的形式釋放出多餘的能量,所產生的光即是螢光(圖 1) 。
基態
激發光 放射光 震動鬆弛 熱激發態
基態
激發光 放射光 震動鬆弛 熱激發態
圖1 螢光能階示意圖 分子受到激發光刺激,由基態上升到激發態,因為電子處於激發態時能量較高且不穩定,所以 分子間會產生震動鬆弛(Vibrational relaxation)會先以熱的形式釋放能量,最後以螢光的形式放出 能量,回到能量最低的基態。1-3 螢光共振能量轉移(Fluorescence Resonance Energy Transfer, FRET)
廣義的來說,FRET 指的是有螢光能量的改變造成螢光共振能量的轉移。但一般 而言,FRET 較常用來探討兩螢光分子間的作用因距離的關係,使被激發的螢光基團 (能量提供者, Donor , D) 能量在回到基態所放出的能量被轉移到鄰近的螢光基團 (能量接受者, Acceptor, A)的現象(圖2、圖3)。FRET 發生的主要條件:1. Donor (D)
和Acceptor (A)之間必須相互靠近小於 100 Å[17]2.Acceptor(A)的激發(Excitation,Ex) 光譜與Donor(D) 螢光放射(Emission, Em)光譜必須有重疊(圖4)。3.Donor(D)和 Acceptor(A)躍遷偶極方向(transition dipole orientations)並非垂直[14,18]。在本研究中 將MT 當作 donor,而 GFP 當作 Acceptor,倘若有 FRET 的現象產生時,會發生在蛋
3 白質結構接近自然態的狀況下,使兩分子的距離彼此接近,並推測此時GFP 的螢光 能量可能會增加。
能
量
10-100 Å 距離 FRET 能量轉移 Donor 供體 Acceptor 受體 D激發光 D放射光 A激發光 A放射光 震動鬆弛 熱 螢光能量轉移能
量
10-100 Å 距離 FRET 能量轉移 Donor 供體 Acceptor 受體 D激發光 D放射光 A激發光 A放射光 震動鬆弛 熱 螢光能量轉移 10-100 Å 距離 FRET 能量轉移 Donor 供體 Acceptor 受體 D激發光 D放射光 A激發光 A放射光 震動鬆弛 熱 螢光能量轉移基態
激發態
能
量
10-100 Å 距離 FRET 能量轉移 Donor 供體 Acceptor 受體 D激發光 D放射光 A激發光 A放射光 震動鬆弛 熱 螢光能量轉移能
量
10-100 Å 距離 FRET 能量轉移 Donor 供體 Acceptor 受體 D激發光 D放射光 A激發光 A放射光 震動鬆弛 熱 螢光能量轉移 10-100 Å 距離 FRET 能量轉移 Donor 供體 Acceptor 受體 D激發光 D放射光 A激發光 A放射光 震動鬆弛 熱 螢光能量轉移基態
激發態
圖2 螢光能量共振轉移 FRET 效應能階圖 Donner 受到激發光激發的 donor,能量會由基態上升到激發態,因為電子處於激發態時能量較 高且不穩定,所以分子間會產生震動鬆弛 (Vibrational relaxation) 會先以熱的形式釋放能量,最 後以螢光的形式放出能量,回到能量最低的基態。 FRET 效應發生在 Donor 與 Acceptor 兩螢 光分子距離範圍約 10~100 Å 時,Donor 釋放的能量不會以螢光的方式釋出,而是轉移能量給 Acceptor,造成 Donor 螢光能量減弱而 Acceptor 螢光能量增加。4 (a)
D
A
D激 發 光距離
Donor AcceptorR>>1.5
R
0D
A
D激 發 光距離
Donor AcceptorR>>1.5
0R
0D
A
D激 發 光距離
Donor AcceptorR>>1.5
R
0D
A
D激 發 光距離
Donor AcceptorR>>1.5
0R
0D
A
D激 發 光距離
Donor AcceptorR>>1.5
R
0D
A
D激 發 光距離
Donor AcceptorR>>1.5
0R
0 (b)D
A
D激發光
A放
射光
距離
Donor Acceptor能量轉移
R<1.5
R
0D
A
D激發光
A放
射光
距離
Donor Acceptor能量轉移
R<1.5
0R
0D
A
D激發光
A放
射光
距離
Donor Acceptor能量轉移
R<1.5
R
0D
A
D激發光
A放
射光
距離
Donor Acceptor能量轉移
R<1.5
0R
0D
A
D激發光
A放
射光
距離
Donor Acceptor能量轉移
R<1.5
R
0D
A
D激發光
A放
射光
距離
Donor Acceptor能量轉移
R<1.5
0R
0 圖3 螢光能量共振轉移 FRET 與距離示意圖FRET 現象的發生與兩螢光分子基團的距離是有強烈相關性。D: Donor ; A: Acceptor。R:兩螢光 分子之間的距離。R0 : Donor 因 FRET 效應而有 50 %能量被轉移至 Acceptor 的距離,一般最 R0
5 螢光吸 收強度 波長(λ) nm 光譜重疊 激發 Ex 釋放 Em CFP 供體, (D) 激發 Ex 釋放 Em YFP 受體,(Y) 螢光吸 收強度 波長(λ) nm 光譜重疊 激發 Ex 釋放 Em CFP 供體, (D) 激發 Ex 釋放 Em YFP 受體,(Y) Donor,(D) Acceptor,(A) 螢光吸 收強度 波長(λ) nm 光譜重疊 激發 Ex 釋放 Em CFP 供體, (D) 激發 Ex 釋放 Em YFP 受體,(Y) 螢光吸 收強度 波長(λ) nm 光譜重疊 激發 Ex 釋放 Em CFP 供體, (D) 激發 Ex 釋放 Em YFP 受體,(Y) Donor,(D) Acceptor,(A) 圖4 CFP 螢光蛋白及 YFP 螢光蛋白的 FRET 螢光光譜
以 CFP 螢光蛋白做為 Donor (D) , YFP 螢光蛋白做為 Acceptor (A) 的螢光光譜。Donor: CFP 螢 光蛋白質,深藍色波形為其激發光 ;淡藍色波形為其放射光。Acceptor: YFP 螢光蛋白質,深綠 色波形為其激發光Excitation ;淡黃色波形為其放射光 Emission 。綠色區塊為 Donor 釋放光與 Acceptor 激發光的光譜重疊的位置。 Ex 指 Excitation 激發波, Em 指 Emission 釋放光。產生 FRET 現象必須有以下條件:1.Donor (D) 必須能釋放螢光 2.Donor 及 Acceptor 間必須要有光譜重 疊[20]。 1-4 淬熄效應 (Quenching) 淬熄效應 (quenching) 屬於能量轉移效應的一種,原理為分子受到光刺激後, 電子從基態躍遷到激發態,然而從激發態回到基態的過程中,與處於基態的淬熄分 子 (quencher) 結合,導致部分分子能量喪失使總光能強度下降[21]。 影響淬熄效應的因素:(1) fluorophore 與 quencher 彼此距離;(2)溶劑的極性或溶 液的pH[22-24] ; (3)所用緩衝液的離子強度 [25,26] ; (4) 色胺酸(Trytophane)的殘基 (Residue)暴露或埋入的狀態[27,28]; (5) 淬熄分子的特性[29,30];(6) 淬熄分子的濃 度。 1-5 生物體發光 生物冷光(Bioluminescence)屬於化學發光(Chemiluminescence,簡稱 CL)的一種 [31,32]。通常化學反應都以熱能的形式釋放能量,但化學發光是在低溫、無光的環 境下,仍然可藉由化學反應產生激發能而釋放光能且不會產生高熱。生物發光分為 直接發光及間接發光兩種。直接發光指分子本身容易達到電子基態且又能產生有效 光的雙重官能基。也就是說由A、B 兩物質發生化學反應生成 P 物質,被激發光 (Excitation)刺激,造成反應能量被 P 物質分子吸收並躍遷到激發態 P*當 P*回到基態 的過程中釋出能量並產生放射光(Emission)。而在此過程中由於 P 物質直接參與反應
6 並產生P*的放射光,故稱直接化學發光,如 GFP 螢光蛋白發出螢光;間接發光又稱 能量轉移化學發光,因為在化學發光的反應中,提供能量的發光劑與負責產生冷光 的冷光劑不是同一個分子。也就是說由反應物A 和 B 反應產生提供能量的激發態中 間體 C*,當 C*反應時釋出能量,使 D 被激發而躍遷到激發態 D*,當 D*躍遷回基 態過程中,放出能量並產生放射光。負責提供能量指C*和負責發出螢光指 D 是不同 分子,其彼此藉由能量的轉移而發光,主要的例子如讓螢火蟲發光的發光素-發光酵 素系統[33]。
1-6 綠色螢光蛋白(Green Fluorescence Protein,GFP)介紹 綠色螢光蛋白起源及發展 2008 年諾貝爾化學獎頒發給研究螢光蛋白的三位科學家,因為 GFP 使原本看 不見的東西,變成看得見,也變成更容易觀察活生物體基因表現。在1960 年日本化 學家下村脩(Osamu Shimomura) 在太平洋東岸北美海域的深海中發現具有生物螢光 的水母(Aequorea Victoria),具有一般水母的外型,但在其傘狀頂部邊緣,可發現綠 色的螢光。在螢光水母內,Aequorin 為一種可將儲存的 ATP 的化學能轉換成光能的 冷光蛋白chemilluminescent protein),後來又發現純化 Aequorin 蛋白質可得到 GFP 此種綠色螢光蛋白。也就是說Aequorin 和 GFP 這兩種蛋白質藉由不同的發光機制產 生螢光 [9,34-36] 。
O
2 CaO
Ca GFPO
Ca*
O
CaCO
2 hv(508nm) hv(470nm) Aequorin (excited chromophore) Aequorin (active) Aequorin (inactive)O
2 CaO
Ca GFPO
Ca*
O
Ca*
O
CaO
CaCO
2 hv(508nm) hv(470nm) Aequorin (excited chromophore) Aequorin (active) Aequorin (inactive) 圖5 Aequorin 的發光機制示意圖 Aequorin"在經過鈣離子的催化後,經過氧化反應,會釋放波長 470nm 的藍色螢光。此色螢光 會傳遞能量激發GFP,使 GFP 釋放波長 508nm 的綠色螢光。7
Aequorin 這種會將化學能轉換形成光能的冷光蛋白(Chemilluminescent protein) 可與鈣離子結合,將儲存於Aequori 中的 ATP 化學能進行發生化學反應產生光能。 此過程為Aequorin 中的 coelenterazine 氧化成 coelentetramide,並釋出 470 nm 的藍 光,而此時發現GFP 吸收 Aequorin 釋出的藍光後,放出 508 nm 的綠光螢光[37-39] (圖 5),後來發展應用於偵測細胞體內鈣離子的濃度[40]。 然而從水母發現的綠色螢光蛋白,其發螢光的機制是建立在螢光蛋白質的結 構,其內可形成共振的環狀發光基團[9]。另外後來陸續被 Morin 等人從腔腸動物的 某些物種中如水螅(Obelia)及海葵(Renilla)、珊瑚蟲(Discosoma)等發現會發螢光的蛋 白[41],因為這些會發螢光的蛋白,具有類似的螢光基團結構,所以有 GFP-like protein 的名稱[42]。同時間 Morin 等人提出能量轉移理論,指出由化學發光的 aequorin 所 發出的藍光會刺激 GFP,將光能轉換成綠色螢光[20,43,44]。1974 年 Morise 純化出 GFP 蛋白並獲得蛋白的質結晶[45,46];1978 年 Prendergast 和 Mann 將 GFP 的單一 分子量明確算出來約30kDa[47,48];1992 年由 Cubitt 等人選殖出來,1994 年 Chalfie 等成功將綠色螢光蛋白的基因轉移 (transgene) 到大腸桿菌 E.Coli 和線蟲 C.elegans 中,並且表現出穩定的螢光[36,47,48];1980 年華人化學家錢永健(Roger Y. Tsien)發 展的能發出各種顏色的螢光蛋白質[36,49]。 綠色螢光蛋白結構及發光機制 GFP 蛋白是由 238 個胺基酸所組成的單一蛋白質鏈 (single polypeptide) [50] ,大部分的胺基酸形成 11 條的 ß-sheets (ß-摺版) 的二級結構緊密的聚在一起, 形成筒狀三級結構,稱為ß-barrel,其 3D 立體結構為圓面直徑為 30 nm、高度 40 nm。 另外桶狀蛋白(ß-barrel)中軸上有一條α-helix,而參與激發螢光的發光基團 (chromophore)位於這條α-helix ,將發光基團包圍在 ß-can 的內部,成一個不受外 界環境影響的穩定空間[43,51] (圖6)。
GFP 的發光基團主要由 Ser65-Tyr66-Gly67 三個胺基酸組成[52] (圖 6),經由環
化、脫氫、氧化三步驟形成多重的共軛雙鍵的共振系統而發出螢光。詳細介紹發光 的機制如下,1.環化作用:藉由單鍵旋轉,使 Tyr66 兩側的醯胺鍵逐漸靠近,此時
Ser65-Tyr66醯胺鍵上氫氧基的氧原子,與 Tyr66-Gly67醯胺鍵上的 N-H 非常接近,使
的陰電性極大的氮原子,與部分帶正電的C=O 雙鍵中的碳原子形成親核作用,產生 新的鍵結。使原本兩個醯胺鍵結的位置產生一個五元環(Imidazolidone ring),且碳的 旁邊新接一個氫氧基,提供脫水位置。2.脫水:在五元環上的氫氧基和鄰近氮上的氫 原子脫出,形成一分子水持留下一個雙鍵結構。3.氧化:在鄰近 Glu222協助下,氧分
8 的雙鍵可以互相溝通,形成p-hydroxybenzyliden-imidazolidinone 的共軛(conjugate)結 構[53],這個結構形成π共振系統 (π-resonance system),當受到適當波長的光線激 發時,此系統上的電子便會被激發而放射出螢光[42,43,54]。形成這步驟需要吸收可 見光並放出螢光[55,56](圖7)。 圖6 GFP 的發光基團結構
GFP 是一個筒狀結構,由 Ser65-Tyr66-Gly67三個胺基酸組成,鄰近Glu222 及 Arg96 可以協助穩
9 (a) (b) 4-pHBI 4-pHBI 圖7 GFP 發光機團與其機制 (a)發螢光的共振結構示意圖:p-hydroxybenzyliden-imidazolidinone 的共軛(conjugate)結構[57]。 (b)為 Ser65-Tyr66-Gly67經由環化、脫氫、氧化產生螢光的步驟[57]。
GFP 的激發光及放射光各有兩個波段,當發光基團為電中性時其激發光為 470 到475 nm,放射光為 503 nm,而發光基團帶負電時其激發光為 395 到 397 nm,放 射光為508 nm,這是因為原本是電中性的發光基團會因為 Tyr66 上的酚基失去一個
10 氫離子而帶負電,使得激發π電子所需要的能量降低,而一般狀況下這兩種構型都 會存在 [9,49,54,58,59]。 綠色螢光蛋白的穩定性 GFP 本身對溫度具有高穩定性,在溫度達到 65℃時,仍可保持其結構並具有活性 [60,61];並且 GFP 在 pH5.5 到 pH12 的範圍均可保有較佳螢光強度,小於 pH4 及高 於pH12 會有螢光失活現象發生[61];GFP 對蛋白質變性試劑耐受性很高,如:8M urea、6M guanidine hydrochloride、1% SDS 中皆非常穩定,但若將 GFP 置於 6M guanidine hydrochloride 並加溫到 96℃,則蛋白質就會失去活性[61]。 螢光蛋白應用及種類 突變GFP 可得到更多類型的螢光蛋白可以應用在生物技術上,做為一個良好的 標記材料,可廣泛應用在偵測基因在細胞體內的表現、蛋白質交互作用的研究。其 最大特色是不需要任何酵素及輔酶的作用,自行進行蛋白質摺疊,隨後利用UV 燈 即可觀察其表現,另外其他突變的螢光蛋白的簡介請參考(表1) [9,43,49,55,57,62-64]。 表1 突變 GFP 螢光蛋白種類 名稱 激發光/放射光 突變位置 螢光變化
EBFP 380-383 nm /440-447 nm Tyr66 ->His66 藍光 ECFP 433 nm/475 nm Tyr66 ->Trps66 藍綠光 EYFP 513 nm / 527 nm Ser65 ->Gly65
Val68-> Leu68 黃光 EGFP 488 nm / 507 nm Ser65-> Thr65 更易更容易離子化 (Ionization),增加螢光 強度。 1-7金屬硫蛋白(Metallothionein, MT)介紹 金屬硫蛋白(圖8)為一種熱穩定且非酵素性之低分子量(約 6-7 kDa)蛋白質,最早 於馬腎臟中純化得到的鎘結合蛋白質[5,7],普遍存在於原核與真核生物中。 金屬硫蛋白(Metallothionein, MT)為熱穩定蛋白質, 無苯環(Aromatic)胺基酸、疏 水性(hydrophobic )氨基酸、無α-helix 或β-sheet 等結構,含有 61-68 個氨基酸(amino acid),內具有 20 個高度保留的半胱胺酸(cysteine, Cys)約佔 23-33%,其保守序列為 Cys-Cys, Cys-X-Cys 或 Cys-X-Y-Cys ( X 為任意的胺基酸),半胱氨酸硫氫基與一價 或二價的過渡金屬元素形成硫金屬鍵結(對金屬親合性為 Zn2+ <Pb2+ <Cd2+ <Cu+ <Ag+ =Hg2+ <Bi3+ [65]),形成高金屬含量(4-12 atoms/mole)的金屬硫氫基團(metal
11
thiolate cluster) [3-6],造成兩個金屬鍵結團簇(α-、β-Cluster)。MT 與體內必須重金 屬(鋅、銅)之調節、毒性重金屬解毒作用以及活性氧化物質清除中扮演相當重要 之角色[66-69]。 S S SS S S Zn Zn S S S S S S S S S S Zn Zn Zn Zn NH3 OOC Zn Zn Zn Zn S S S S SS S S S S S S S S S S Zn Zn Zn Zn
β-cluster
α- cluster
Metallothionein
Metallothionein
protein (MT)
protein (MT)
S S S S S S S S S S SS S S Zn Zn S S S S S S S S S S Zn Zn Zn Zn NH3 OOC Zn Zn Zn Zn S S S S SS S S S S S S S S S S Zn Zn Zn Zn
β-cluster
α- cluster
Metallothionein
Metallothionein
protein (MT)
protein (MT)
S S S S S S S S 圖8 金屬硫蛋白(Metallothionein, MT)結構示意圖 金屬硫蛋白(Metallothionein, MT)形成兩個金屬鍵結團簇(α-、β-Cluster),含 20 個高度保留的 半胱胺酸(cysteine,Cys)與重金屬形成硫金屬鍵結。MT 的 C 端 (Carboxyl terminal) 由 31 到 61 個氨基酸組成含11 個半胱胺酸,形成α-金屬鍵結團簇(α-Cluster),鍵結 4 個二價金屬離子的 (M4S11)3-的團簇;MT 的 N 端(amino terminal)由 1 到 30 個氨基酸組成含 9 個半胱胺酸,形成 β-金屬鍵結團簇(β-Cluster),鍵結 3 個二價金屬離子,形成了(M3S9)3-的團簇。M 指二價金屬 離子(Zn2+, Cd2+,或其它金屬離子)。這兩個離子結合的團簇整體的大小約 3-4nm,其結構如 “metal semiconductor, MS”的複合結構。[70-73] MT 依結構特性分為三類[74],ClassⅠ:半胱胺酸結構位置與馬的 MT 類似,如哺 乳類動物的 MT [75-78];ClassⅡ:半胱胺酸結構與來自馬的 MT 相似度低,例如酵母 菌 的 copper-thionein [79,80];Class Ⅲ : 大 部 分 存 在 植 物 或 微 生 物 體 如 植 物 的 phytometallothionein[75,81-84]。 Binz 和 Kagi 在 1999 年提出新的分類方式[75,85],新分類系統是依據序列的相 似度以及親緣關係進行分類,分類包括16 個 MT 家族(families)[86]、10 個 MT 亞 家族subfamilies)、MT 群(groups)及 MT 同分異構物(isoforms)[87]。舉例來說, MT 家族 (families)分出的 16 個 MT 家族,其中 MT 家族 (Families)I 為脊椎
12 動物的 MTs ,其親源遺傳特徵再分為 10 個亞家族 (subfamilies) ,分別是哺乳類 MT-1、 MT-2 、MT-3、MT-4[88] [89,90]及哺乳類未區分之 MT ;鳥類 MT-1、 MT-2 及鳥類未區分之 MT ;兩棲類 MT 及硬骨魚類 MT [91,92]。 而 MT 家族 Ⅱ 為軟體動物類、 MT 家族 Ⅲ 為甲殼類、 MT 家族 Ⅳ 為棘皮動物類、 MT 家 族 Ⅴ為雙翅類、 MT 家族 Ⅵ 為線蟲類、 MT 家族 Ⅷ 為纖毛蟲類、 MT 家族 Ⅸ ~ⅩⅢ 為黴菌類 MT 家族、 MT 家族 ⅩⅣ 為原核動物類、 MT 家族 ⅩⅤ 為植 物類[87]。 哺乳類動物具有多 MT 基因家族,泛稱 MT 異構型(iso-metallothionein),可分 成四個次群: MT-1, MT-2, MT-3[77], MT-4 [88,93-96]; MT-1 及 MT-2 是最 主要的普遍存在各種組織中,例如肝、腎、胰臟、腸道等組織[93],被認為可能分別 各在重金屬的代謝與在細胞生長中扮演重要之生理角色。因為 MT-1 及 MT-2 受到 如重金屬、醣腎上線皮質素、氧化壓力及細胞激素以及冷、熱的生理性緊迫因子等 刺激會被誘導而表現,這兩型金屬硫蛋白其主要功能為[97]:(1)減低重金屬毒性。(2) 維持必須重金屬的體內平衡。(3)自由基的清除。(4)細胞的生長及增殖。[97,98];而 MT-3 及 MT-4 對生理性的緊迫因子的反應較不敏感,這兩種主要分佈位置, MT-3 主要存在神經系統中,由人類的由腦皮層星狀細胞所分泌,原本命名為生長抑制因 子(growth inhibitory factor) 且通常[66,90,99-102]; MT-4 存在鱗狀上皮組織中 [103]。最近研究指出腦受到一些刺激時會誘導 MT-3 的表現,並在阿茲海默症患者 腦中有MT-3 耗盡的現象[104,105]。
1-7.1蛋白質結構與摺疊的重要性
1-7.2 蛋白質的變性 (denaturation) 與沈澱
蛋白質的結構與蛋白質的生物功能有相關性[106],將具有功能性的蛋白質結構 定義為自然結構 (Native structure) ,又稱此蛋白質為自然態蛋白質 (Native protein)。
在水溶液中,具自然結構的蛋白質分子表面,除了N 端的胺基 (-NH3+)、C 端的 羧基 (-COOH-) 外,其他殘基之側鏈基團會彼此鍵結,或和游離離子、水鍵結,因 此形成水層及雙電層成為穩定的膠體顆粒。 當蛋白質結構受到外在環境的劇烈改變時,蛋白質內穩定結構完全被破壞,而 彼此聚集(aggregation)沉澱,且失去生物活性,此狀態的蛋白質稱為變性蛋白質 denaturation protein)。讓蛋白質變性的方法大致可分三大類:物理法、化學法、生 物法等,作用原理是使蛋白質結構改變、降低溶解度、失去生物活性[107]及結晶能 力[108]。而其他變性方法參閱(表 2) [106,109-111]。
13 表2 蛋白質變性的方法 方法 例子 物理法 高溫、冷凍、壓力、攪拌、超音波 化學法 1 極端 pH 變化 (強酸、強鹼);2.有機溶劑 (酒精、丙酮);2.鹽 析法 (硫酸胺);3.化學變性藥劑 (尿素(urea)、胍鹽酸 (guanidine hydrochloride)、二硫代蘇糖醇 (sodium dodecylsulfate) ;4.重金 屬 生物法 蛋白質水解酵素 (proteolytic enzyme) 1-7.3 蛋白質的復性 (renaturation) 與摺疊 (fold) 蛋白質復性 探討蛋白質從解構(unfold)到摺疊(fold)的過程,將未摺疊或摺疊錯誤的蛋白質, 重新展開摺疊到擁有最小自由能(free energy)的特殊三級結構,使蛋白質回復活性, 此過程稱為復性 (renaturation) 。 蛋白質來源 本實驗是利用細菌產生產量高、純度高、不可溶、不具生物活性的的內涵體 (inclusion body)為蛋白質來源,其內帶有實驗所需的重組目標蛋白 (MTGFP 及 MTekGFP)。為得到具活性的重組目標蛋白質,需經三個步驟包括破菌、清洗內涵體 及溶解內涵體,再藉由蛋白質復性與摺疊後,才能得到高純度且具生物活性的蛋白 質[112]。 利用內涵體進行復性時,要注意當蛋白質結構被展開後,使疏水性區域的胺基 酸鏈大量暴露在蛋白質的外部容易形成聚集體(aggregation)。在蛋白質復性的過程 中,因為結構正確的多胜肽鏈內 (intrachain )與結構錯誤多胜肽鏈間(interchain)產生 交互作用(interaction),進而糾結在一起形成錯誤的雙硫鍵鍵結 (misfolding)[113]。所 以最好在低蛋白質濃度下進行復性的過程,避免因聚集體產生而減少自然態蛋白質 的產率[114]。
14 蛋白質摺疊 有研究顯示蛋白質摺疊過程中,不是只有摺疊態與未摺疊態兩種形式。同時也 包括了摺疊過程中的摺疊中間體 (folding intermediate) [106,111,115-118]。而蛋白質 的結構會受到許多環境因素影響,包括壓力、PH 值、離子強度[119]、溫度[120]等, 所以將蛋白質溶液的環境改變時,蛋白質為適應新環境會改變結構,使高能量趨向 低能量,當達到自然態時,就是處於最低能量態,維持熱力學上的平衡。另外在蛋 白質摺疊的過程中,通常也會添加一些幫助摺疊的試劑,減少聚集的分子間錯誤鍵 結[121]及維持適合的氧化還原環境[122]。減少蛋白質分子間錯誤鍵結的添加劑簡介 請參考( 表3)[112,113,121,123]。 表3 減少蛋白質分子間錯誤鍵結的添加物 添加物 建議濃度 Urea 2 M
Guanidinium chloride (GuHCl) 1 M
Arginine 0.5 M Glycerol 0.4 M Sucrose 0.4 M Lauryl maltoside 0.3 M Polyethylene glycerol (3350MW) 0.05 % W/V Tris buffer 0.5 M Triton X-100 10 mM Acetamide 2 M Acetone 1.4M n-hexanol 5 mM Salts (NaCl、Na2SO4、K2SO4) 0.5M 蛋白質摺疊的方式
摺疊蛋白質主要分為兩種:直接稀釋法 (Direct Dilution)及透析法 (Dialysis)(表 4),這兩種摺疊方法的目的都是將蛋白質周圍環境的變性藥劑濃度降低或是移除,使
變性藥劑其無法破壞蛋白質結構,最終使蛋白質能重新摺疊到正確結構並具生物活 性的型態[124]。在本實驗中所使用的方法為兩種方法的改良,稱階段性熱平衡法 (Stepwise Thermodynamics Equilibrium Dialysis)。介紹如下:
15 直接稀釋法 (Direct Dilution):將未摺疊的蛋白質,直接從含有高濃度的變性試 劑緩衝液的環境,直接稀釋到低濃度變性試劑或無變性試劑的緩衝液[125]。雖降低 變性試劑濃度就無法破壞穩定蛋白質結構的力量,蛋白質即可自行摺疊,但用直接 稀釋法進行復性的缺點為增加蛋白質溶液的體積,蛋白質濃度會因此降低許多,因 此不利於蛋白質回收和純化。 透析法 (Dialysis):變性溶液的蛋白質利用透析膜的特性,將其置於透析袋內, 透析袋外放置大量要置換的復性溶液。由於透析膜只允許比孔徑小的分子通過,蛋 白質無法穿透透析膜。藉由膜內外鹽濃度的不一致,造成滲透壓的差異,使溶液中 的離子可藉由布朗運動而自由移動,最終使得膜內外離子濃度達平衡,達到將變性 藥劑完全移除的目的[126,127]。使用透析法做摺疊,蛋白質最終的回收濃度高,但 也很容易形成結構摺疊錯誤的蛋白質及聚集體[128]。於是在 1996 年 Rudolph 等人 提出蛋白質濃度維持 0.01~0.05 mg/ml 下有最佳的摺疊效果[129]。
階段性熱平衡透析法 (Stepwise Thermodynamics Equilibrium Dialysis):階段性 熱平衡法準靜態是遵守熱力學平衡的理論,可以將高濃度蛋白質進行蛋白質摺疊, 且可避免形成聚集體的方法。實驗利用透析膜的特性,將變性溶液的蛋白質置於透 析袋內,透析袋外放置大量要置換的復性溶液,不同的是逐步改變變性藥劑到要置 換的復性溶液中的濃度,最終使得膜內外離子濃度達平衡,達到緩慢的將變性藥劑 完全移除的目的[126,127]。因為蛋白質在摺疊每一階段都符合準靜態熱力學平衡的 定律,因此可避免蛋白質因為周圍環境的劇烈變化而造成沈澱,另外蛋白質有自組 織的能力,所以其構形會自發性的摺疊,使能量能連續的改變反應位能,最後達到 能量最低態(local minima)也就是自然結構[115]。 表 4 直接透析與階段性熱平衡透析法比較 名稱 特色 蛋白質影響 透析法 透析膜只允許比孔徑小的分子通 過,蛋白質無法穿透透析膜,藉由 布朗運動而自由移動,最終使得膜 內外離子濃度達平衡。 蛋白質最終的回收濃度高,但 也很容易形成結構摺疊錯誤的 蛋白質及聚集體, 直接透析法 降低蛋白質濃度,減少聚集。 高濃度蛋白質環境下,易造成 蛋白質聚集而沈澱。 階段性熱平衡 透析法 逐步透析降低變性試劑的濃度到 完全移除。 避免因蛋白質周圍環境變化劇 烈,造成蛋白質聚集而沈澱。
16
第二章實驗原理:
2-1 DNA 聚合酶連鎖反應 (Polymerase Chain Reaction,PCR)
1983 年 Kary B. Mullis 博士則想出 PCR 技術-現今所發展出來的 PCR 技術放大 選殖DNA 的片段,並且在 1993 年獲得諾貝爾化學獎[130]。PCR 是放大特定 DNA 片段的實驗技術,使欲放大的目標DNA 序列兩端分別合成能與其形成緩冷配對反應 (annealing) 的前置引子 (forward primer) 和反置引子 (reverse primer),再利用 DNA 聚合酵素(DNA polymerase)以目標 DNA 的兩股分別做為模板(template)來合成新的 DNA 股。
在理想的PCR 反應條件下,每執行 PCR 一次其 DNA 產物增加為原來的兩倍, 故PCR 公式為 DNA 增加的量將會是 2n,n 是代表 PCR 執行的次數。在本實驗中採 用的是Tag DNA 聚合酶(Taq DNA polymerase)是個耐高溫的酵素,在 95℃ 中其活性 的半衰期 (half life) 長達 40 分鐘,故可供 PCR 操作使用。其酵素有效作用溫度為 72℃,此溫度下分鐘可合成 2000-4000 個核甘酸(nucleotidees)。但 Taq 聚合酵素因為 缺乏3'至 5'端外切酵素 (exonuclease),所以在 DNA 合成時沒有校對(proofreading) 的 功能,在每一個循環中錯誤配對的頻率可高達1/6000 個核甘酸。 影響PCR 的 DNA 合成時的精確性之因素:1.核甘酸在 PCR 合成時的濃度,核甘 酸的量應控制在 10pg~1ug 之間,過多或過少都會影響 PCR 之精確性; 2.DNA 的長 度:以1kb 以下為宜; 3.循環數:越多時精確度越低; 4.DNA 聚合酶的種類:有校對能力 者為佳; 5. Mg2+ 的含量:建議在0.5-2.5 mM 之間,其會影響引子黏合、DNA 雙股打 開時所需之溫度、引子形成二聚物等 。 2-2 DNA 定序
DNA 定序有兩種方法,分別是 Sanger 法及 Maxam-Gilbert 法。而 Sanger 法 是目前市面上所用的定序法,以 DNA 為模板,加入四種不同螢光標示的核甘酸的 類似物ddNTPs (ddATP、ddTTP、 ddCTP、 ddGTP)及適當比例的 dNTPs 和聚合酵 素,同時進行聚合酵素鏈鎖反應。因為核甘酸類似物缺少3'-hydroxyl group,因此無 法與下一個核甘酸的5'-Phosphate group 形成 phosphodiester bond,使得 DNA 複製終 止在這些類似核甘酸的位置上,造成各種長短不一的DNA 片段。再以高解析度的毛 細管電泳技術將這些長短不一的DNA 片段 分離,最後透過雷射光激發,並利用 CCD 偵測每一片段不同螢光的組合來判讀DNA 序列。
17
2-3誘導劑 IPTG
養菌時使用IPTG 誘導目標蛋白質產生,且 IPTG 其結構式類似乳糖 (lactose) 的 誘導劑,其誘導的目標是乳糖操作因子(lac promotor)的抑制蛋白(repressor),所以 IPTG 或乳糖會使抑制蛋白失活,達到誘導基因的目的。在細菌(E.coli) 培養中,添 加 IPTG 可有效地在短時間內快速誘導外來重組基因,產生目標蛋白質,但會對細 胞新陳代謝能力的產生影響,相對於不帶有質體的菌體,對帶有質體的菌體有較大 的抑制作用,使得帶有質體的菌體比生長速率降低,即使沒帶有質體的菌體也會受 到影響[131]。一般使用方式多半為先培養菌體到高濃度,再利用誘導劑誘導外來蛋 白質,以取得最大量的目標蛋白質。改變 IPTG 的濃度也會使得內涵體形成的位置 產生改變,在IPTG 濃度較低(0.025 ~ 0.1 mM)時內涵體(inclusion body) 會只在細胞 間質(periplasm)形成,但如果 IPTG 濃度提高(0.1 mM),則在細胞間質和細胞質 (cytoplasm) 內都會產生[132]。 2-4內涵體 (Inclusion body) 在本實驗中用MTGFP 基因或 MTekGFP 基因以 E.coli 為宿主,大量表現目標重 組蛋白,使蛋白質過量生產產生內涵體。內涵體是一種不可溶、密度較大、顆粒狀 的無活性蛋白質[133],內涵體不一定只在受損、突變、生長溫度偏高才會出現在細 胞內,當表現蛋白質為有毒蛋白或微生物在細胞內大量表現重組蛋白質時,也會在 其內形成不可溶的蛋白質堆積,即為內涵體[134,135]。 形成內涵體的原因有兩種,一種是因為高濃度的重組聚胺基酸鏈(recombinant polypeptide chains) 因疏水性作用力而發生聚集(aggration)且形成無定型的聚合物在 細胞內或細胞間質[136] ;另一種則為原本應在蛋白質分子內形成的雙硫鍵(disulfide bond) ,卻形成在兩個蛋白質分子間,使得蛋白質分子串聯形成密度較原本單一蛋 白質分子為大的巨型分子。然而內涵體的形成不一定都是雙硫鍵的原因,但是經過 研究內涵體之所以不能行逆反應變回原來的蛋白質分子,應該與雙硫鍵形成後增加 分子的穩定度有關[137]。 以 產 量 及 純 化 的 觀 點 看 內 涵 體 , 其 實 利 用 內 涵 體 獲 得 蛋 白 質 有 許 多 好 處 [138-140]:1.可獲高產量的蛋白質,因為內涵體佔全部細胞蛋白質 30% 以上,且內 涵體內有 95 % 皆是重組的目標蛋白質,或是每公升菌液可產 8.5 克蛋白質 [141,142]2.內涵體因為在分子間有雙硫鍵形成,且可被分解的胺基酸序列都被包埋在 內涵體內部,所以不易被蛋白質水解脢 (protease) 水解,3.內涵體為不可溶的蛋白 質,可藉由此特性將破菌後的內涵體,利用離心、重複洗滌、膜過濾等方式[143], 分離可溶性蛋白(例如:核酸、磷脂、及其他水溶性蛋白質),減少純化步驟[144]。
18 有研究建議洗滌內涵體時,可依狀況添加低濃度的尿酸或胍鹽酸或 EDTA 等,可移 除可能會吸附在內涵體表面或疏水性區域上的雜蛋白,增加復性的效率[145]。 2-5 蛋白質變性與沈澱 蛋白質發生變性時,其多胜鏈的共價鍵並沒有斷裂,被破壞的只有蛋白質的二 級、三級、四級結構。然而蛋白質變性時,常會造成蛋白質溶解度降低而產生沉澱。 常見的蛋白質沉澱法︰ 強酸(強鹼)沉澱法:在中性蛋白質溶液中加入強酸(或強鹼),使羧基(或胺基) 變成未解離之羧基 (或已質子化之胺基),而可接電子之氮原子,因質子化造成蛋 白質帶有正電荷(或去質子化而不帶電荷),導致蛋白質受到離子鍵(鹽橋鍵) 及 氫鍵等破壞,使蛋白質溶水性變差而沉澱。 有機溶劑沉澱法:加入水溶性有機溶劑 (如: 酒精、丙酮等) 因為其具有低介電常 數,所以會造成蛋白質間正負電荷結合力增加,形成厭水性膠體使水與蛋白質間作 用力減低,而蛋白質溶解度下降隨即發生沉澱而沉澱。 重金屬離子沉澱法:蛋白質在水溶液中是酸鹼兩性電解質,在 pH 7.0 或以上溶 液中,蛋白質分子大部分帶負電荷,故能與正電荷的重金屬離子結合,中和蛋白質 其負電荷,形成蛋白質鹽沉澱出來,此種沉澱稱為蛋白化重金屬。 植物鹼沉澱法:利用生物鹼之酸根與蛋白質之陽離子電荷結合形成難溶性鹽類 而沉澱。常用的生物鹼有鞣酸、三氯醋酸(trichloroacetic acid) 、磷鉬酸、苦味酸及 sulfosalicylic acid 等。當 pH 小於蛋白質等電點時,沉澱最明顯。 鹽析法(salting out):於蛋白質溶液中加入多量輕金屬鹽類,改變溶劑介電常數, 使蛋白質間作用力增加而沉澱。一般常用硫酸銨鹽((NH4)2SO4)沉澱蛋白質。 加熱沉澱法:幾乎所有的蛋白質都會因為加熱變性而凝固,產生不可逆的沈澱。 蛋白質在等電點時,加熱凝固最完全且迅速。但在酸性或鹼性溶液中,蛋白質帶有 正電或負電時,雖加熱蛋白質也不會凝固,除非同時有足量的中性鹽存在,則蛋白 質即可因加熱而凝固。 2-6 各種常見的化學變性劑 一般常用的化學變性劑溶解內涵體,或將糾結聚集在一起的蛋白質質結構解 開,其作用原理簡單介紹如下: 尿素 (Urea): 一般建議使用高濃度的尿素 9 M,破壞分子內氫鍵及疏水性作用力,而在本實 驗中,我們使用4.5M 的濃度來達到蛋白質的變性[146]。 胍鹽酸 (Guanidine hydrochloride,GuHCl):
19 一般建議濃度 6 M,主要可破壞分子內疏水性作用力及離子性作用力。和尿素 一樣是和蛋白質分子間形成氫鍵鍵結且其作用力大於穩定蛋白質結構之力量,因而 將蛋白質拉開至鬆散的構形。此外亦可伴隨著添加少量氯化鋰(LiCl)或氯化鈉 (NaCl)等鹽類,以加強破壞離子性作用力[146]。 鹽類: 常用氯化鈉(NaCl)、氯化鉀(KCl)、氯化鋰(NaLi),主要破壞離子性作用力[147]。 還原劑 (reducing agent): 一般常用二硫代酥糖醇 (dithiothreitol,DTT)[148]、DTE (dithioerythritol)、半胱 胺酸(cysteine)、β-氫硫基乙醇(β- Mercaptoethanol)[110]等,主要作用是破壞雙硫 鍵,並加入還原劑將雙硫鍵還原成氫硫鍵。由於雙硫鍵為共價鍵結,為穩定蛋白質 結構最主要力量,因此要完整破壞其結構還需要尿素或胍鹽酸的協助方可達成。另 外無雙硫鍵的蛋白質,像 Bacillus subtilus α-amylase 則不需添加此種還原劑。
清潔劑 (detergents):
常用的清潔劑有包括十二烷基硫酸鈉 (sodium dodecylsulfate, SDS)[149]、n-cetyl trimethlammonium chloride[150]、sarkosy[151]、sodium n-laurosyl sarcosine[152]等, 利用清潔劑本身親水性基將蛋白質的立體結構解開成直鏈,使蛋白質疏水性區域裸 露出來,並帶負電荷。可將蛋白質立體結構拉開並帶負電荷[110]。最有名的例子, 就是用在跑蛋白質變性電泳時,用SDS 使蛋白質變性。 pH 值: 胺基酸的組成中含有氨基(-NH3+)與羧基(-COO-)這兩種基團,在高 pH 值的環境 中氨基會失去一個氫原子而變成 NH2,使得胺基酸帶負電,在低 pH 值的環境中羧 基會接上一個氫原子形成 COOH 而使胺基酸帶正電。因此在高、低 pH 值的環境 中,蛋白質會因為胺基酸之間有庫倫作用力互相相斥,而無法正常的排列、組織成 自然的結構。 金屬螯和劑 一般常用乙二胺四乙酸 (ethylenediaminetetraacetic acid,EDTA) 及 2-氨乙基醚 (ethylene glycol tetraacetic acid,EGTA) 等金屬螯合劑,其會與溶液中的金屬結合,形 成穩的螯合物,避免金屬影響化學反應的進行。
20
2-7 蛋白質摺疊 (Protein Refolding)
探討蛋白質由一無特定構形的鬆散結構摺疊到具有特殊構形的過程,其分子內 結構變化及分子間交互作用是目前熱門的研究蛋。白質摺疊的機制在真核細胞與原 核細胞中可分為Cha peronin-dependent 與 Chaperonin-independent 兩種[153]。對於小 分子蛋白質而言是不需要chaperones 即可自行摺疊到自然態;但大分子蛋白質在摺 疊過程中,需要chperones 的協助才能順利摺疊到自然態,因為 chaperones 可結合到 暴露在親水環境中的疏水性的胺基酸鍊上,避免蛋白質分子產生交互作用造成聚集 而影響摺疊效率[154,155]。
本實驗用階段性熱力學平衡透析法來做蛋白質摺疊,這個透析法的理論基礎是 建立在2002 年提出的 First – order like state transition model[115],這是一個首先探討 蛋白質在凖靜態熱力學平衡的過程摺疊的模型。並且此理論與 1937 年的相轉換理論 相符合[156]。而 First – order like state transition model 可以有效的降低摺疊過程中蛋 白質分子的交互作用,避免聚集現象產生[157]。另外根據這理論所衍生出的階段性 熱力學平衡透析法,可以使蛋白質摺疊路徑有效率的繞過相轉變區間 (即
over-critical point reaction path) 達到自然態。已經有許多研究是利用階段性熱力學平 衡透析法成功的將蛋白質復性到自然態,且添加一些化學物質幫助蛋白質摺疊,就 像chaperons 協助蛋白摺疊的角色一樣[111,115,157,158]。由於階段性熱力學平衡透 析法,其整體溶液交換擴散反應是緩慢進行的,再加上此透析方法縮小溶液中起始 狀態及末狀態的差異 ,因此整體溶液置換的速率是非常緩慢的,約需 8 到 48 小時 才能達到平衡[159]。且對於利用階段性熱平衡透析法摺疊的蛋白質來說,因為其摺 疊速率在10-9~103秒之間,所以在整個摺疊過程幾乎一直處於平衡狀態,所以把這 種溶液置換的過程稱為凖靜態過程 (quasi-static process)(圖 9)。 實驗步驟內的詳細原理記錄於下方: 蛋白質變性(denature):第一步就是要讓本實驗中目標蛋白 (MTGFP 及 MTekGFP)的蛋白質變性,本實驗當中所使用的變性緩衝溶液(Denature Buffer),包 含有 4.5M 的尿素(Urea)用來破壞蛋白質內氫鍵與疏水作用力等,一些非共價鍵結 力;0.1M β-ME (β-Mercaptoethanol)是抗氧化劑,也可防止雙硫鍵形成,且本實驗的 蛋白質,具有含硫的氨基酸半胱胺酸)之間容易形成雙硫鍵,藉由高濃度的β-ME 可 以有效的將雙硫鍵(S-S)還原成硫氫鍵(S-H)。而一般常用為緩衝溶液的有磷酸 (phosphate)與三羥甲基氨基甲烷(Tris),phosphate 為酸性的緩衝液,而 Tris 則是鹼性 的緩衝液。本實驗的蛋白質,是在鹼性環境 Tris buffer 下進行摺疊。除了變性劑和 還原劑之外,還有加入0.5%的甘露醇(Mannitol) 和 0.1mM 的 Pefabloc,這兩種藥劑
21
的功能主要是用來保護在水溶液中的蛋白質。甘露醇能保護蛋白質的側鏈分子,避 免在高度氧化、還原的環境下發生一些修飾(modify)作用。而 Pefabloc 是一種可抑制 蛋白質分解酶的化合物(synthetic protease inhibitors),也能避免蛋白質被分解。它能 和蛋白質分解酶產生不可逆的共價鍵結,使分解酶失去活性,就無法分解蛋白質。 因此在摺疊的過程中,都會加入 Pefabloc 來保護蛋白質不會被分解。另外低濃度 (0.1mM)的 β-ME 可以當做抗氧化劑,其作用和甘油(glycerol)相似。
階段性熱平衡透析法 (Stepwise Thermodynamics Equilibrium Dialysis)的過 程:第二部取得變性蛋白質後,用0.22μM 過濾蛋白質樣品,再將蛋白質樣品裝入只 允許小於 3kD 的蛋白質通過的透析袋內,並將半透膜置放於 2.5L 的摺疊緩衝溶液 ( Refolding Buffer)中,使高濃度的變性劑及還原劑可緩慢擴散到外面低濃度的摺疊 緩衝溶液中,使膜內變性劑及還原劑濃度降低,因此可避免環境劇烈變化而使蛋白 質沈澱的現象,使蛋白質順利的進行摺疊。 在本實驗中設計5 個的階段性的摺疊步驟,透過 5 種摺疊緩衝溶液使蛋白質摺 疊的環境緩慢的改變到接近自然態的環境。這五種階段的緩衝溶液,分別稱為R1、 R2、R3、R4、R5 的摺疊中間體,則稱 M1、M2、M3、M4、M5。而五個階段中, 前三階段(R1~R3)將變性劑尿素從 4.5M 依次減為 2M、1M,然後移除;而還原劑 β-ME 則從R1 開始就降為 0.1mM 當做保護劑;而摺疊緩衝液的 pH 值從 R4 開始調到接近 中性(pH=8.8);而 R5 在最後一個階段移除保護劑 Mannitol,使環境接近自然狀態, 讓蛋白質能摺疊至自然結構。另外收集每個階段蛋白質所形成較穩定的摺疊中間體 (folding intermediates)的結構,以提供後續蛋白質結構分析的量測。
22 圖9 能量改變示意圖 階段性熱平衡透析法的過程符合熱力學之準竟過程,在每一個位能改變的過程都能達到熱力學 的平衡狀態,也就是局部能量的最低態。所以使蛋白質從未摺疊到摺疊完全的自然態,可以逐 步緩慢的進行摺疊,避免蛋白質因外在環境劇烈變化而產生沈澱現象[160]。 2-8 圓二色光譜儀 (Circlular Dichrorism, CD) CD 光譜可分析蛋白質的二級結構比例:α-螺旋 (α-helix ) 、β-板狀 (β -sheet )、迴路 (turn)、及無序纏捲 (random coil)[161],因此探討蛋白質摺疊過程中 其二級結構的變化。胺基酸分子大部分都具有光學活性,除甘胺酸 (glycine) 以外, 其餘的氨基酸從胺基酸的結構來看,皆是一個四面體的形狀,且四個角都帶有不同 的基團 (碳基、胺基、羧基、R 基),且各基團在空間中的不同排列,而形成兩種不 同構形的胺基酸。而稱這樣不同的構形為鏡像異構物 (enantiomer),或是生物分子的 手性 (chirality)。而這類分子光學活性的來源是由於能產生左旋或右旋偏極光,能產 生左旋偏極光稱為左旋異構物 (levorotatory isomer,L-form);而能產生右旋偏極光 則為右旋異構物 (dextrorotatory isomer,D-form)。因此胺基酸具有光學活性的特色, 所以由胺基酸組成的蛋白質其立體結構也具有光學活性,尤其以二及結構最具有特 徵。故當圓形入射光視為左右旋光向量之合通過水溶液中的蛋白質時,將會吸收與 電子雲方向相同之電場,而使入射光產生偏轉角度,最後最後利用圓二色旋光光譜 儀就是透過計算左旋偏極光 (LCPL) 與右旋偏極光之差異 (RCPL) 吸收,得到蛋白 質吸收橢圓率,可畫出個吸收特徵圖形[162]。 一般測量蛋白質其CD 波長範圍 260-195 nm,典型的 α-helix 在 222nm、208nm 擁有兩個負波,但在192nm 左右有一個正波;而 β-sheet 在 210nm、222nm 有兩個 負波,但在195nm 有一正波;而 turn 的結構是在 190nm 有一波,但 208nm 左右 有一正波[163]。
23
2-9 動靜態光散射儀 (Dynamic Light Scattering,DLS)
用來分析蛋白質在摺疊過程中,各摺疊階段的溶液內蛋白質大小的分佈情形。 利用雷射光射入蛋白質溶液內,因為雷射光撞擊到溶液中的蛋白質後會產生散射 光,而散射光會依著時間產生變化。所以可由測量散射光計算出粒子之粒徑分佈情 形。由於蛋白質粒子處於溶液中,因本身具有動能所以會產生不規則的擴散運動 (Diffusion motion),另外粒子的大小也會影響粒子之擴散速度與布朗運動,加上雷射 光照射的區域粒子數量和位置不同。使得訊號到達偵測器時會產生光程差,產生干 涉影響光線強度,因粒子位置會隨脽間改變,因此散射光亦會隨時間改變。而透過 這樣的變化,可以得到所謂的擴散係數,而根據Stockes-Einstein 方程式: H R kT D 6 D 是擴散係數,k 是波茲曼常數,T 是絕對溫度,η 為溶液之黏滯係數,R 為水 合半徑。我們就可測量到在溶液中蛋白質的粒徑大小。 2-10 螢光儀(Fluoresce) 大部分的蛋白質中都具有芳香族的胺基酸,分別是酪胺酸(tyrosine)、色胺酸 (tryptophan)和苯丙胺酸(phenylalanine),這些芳香族的胺基酸是一種螢光發色團 (fluorophore),所以蛋白質是一種螢光物質。然而蛋白質內的這些螢光基團對其四周 環境的改變極敏感。當蛋白質的立體結構出現變化時會影響其螢光光譜,當螢光基 團在蛋白質中被其他胺基酸基團所包覆而遠離水相時其螢光光譜會有所謂的藍位移 (blue shift); 而螢光基團是暴露在親水性環境中時,其螢光最強波長則會向紅光移 動,稱為紅位移 (red shift)[164]。而螢光強度也是一種蛋白質結構穩定的特徵,當蛋 白質結構穩定時,其螢光基團的共振結構也是相對比較穩定,所以螢光強度也會相 對比較高;若是結構較為鬆散,其共振結構也會不穩定,如此一來螢光的強度會跟著 變弱。而透過螢光光譜來分析我們的摺疊中間物可以瞭解蛋白質在摺疊過程中一些 結構的變化及其結構的穩定性。另外也可分析蛋白質是否有交互作用,蛋白質間彼 此有交互作用則會影響螢光的變化,產生如FRET 及 Quenching 等現象。
2-10.1 螢光共振能量轉移(Fluorescence Resonance Energy Transfer, FRET)
能量轉移是藉由螢光團之間的偶極-偶極作用“dipole-dipole interaction", 因此 分子間距離與兩螢光團間有極大關係,FRET 的有效範圍約從 10~100 Å,其能量轉 移效率EFRET的強度與兩螢光分子距離(r)的六次方成反比[17](圖 10、圖 11)。
24 6 0 1 1 R r
E
FRET (1) 圖10 FRET 效應與距離的關係R0 指 Förster 半徑(Förster distance)發生在 FRET 效應轉移 50 % 能量的距離。 FRET 效應
的強弱與兩分子之間的距離有關,若兩分子之間距離大於 1.5 倍 R0,則不會產生 FRET 效應。
R0指 Förster 半徑(Förster distance),發生 FRET 效應時,Donor 將 50 % 螢光
共振能量轉移到Acceptor 的距離,其計算公式如下:
17k
2Q
D AJ
16nm
02
.
8
10
R
(2) 2k :偶極方向因子 (dipole orientation factor),依兩分子的偏振情況,數值範圍從 0 到 4 ,如果兩分子是隨機方向其數值則為 2/3 ;QD: Donor 在沒有 Acceptor 的情
況下,其螢光量子產率; :Acceptor 最大吸光係數 (MolA -1.cm-1) ;J(λ):Donor 激發
光及Acceptor 釋放光的光譜重疊處的積分計算如下:
F
E
d
J
D A
4
(3)A
E :Acceptor 的消光係數 (Extinction coefficient of acceptor);FD:Donor 在 FRET 時的螢光放射強度 (Extinction coefficient of acceptor ) 佔總面積的分率。
25 綜合以上幾點,再次強調當Donor 激發光譜與 Acceptor 放射光譜有重疊時,表 示它們的能階有重疊,故能使共振 (resonance) 效率提高,導致有螢光共振能量 轉移的現象產生[20,91,165-167] D A B A A 放射 光 B A D A D A D B D D B D D B A A D A D A D B A A 放射 光 激 發 光 距離 長度 距離 長度 距離 長度 D A B A A 放射 光 B A D A D A D B D D B D D B A A D A D A D B A D A B A A 放射 光 B A D A D A D B D D B D A B A A 放射 光 B A D A D A D B D D B D D B A A D A D A D B A A 放射 光 激 發 光 距離 長度 距離 長度 距離 長度 圖 11 FRET 強度與距離的示意圖 Donor 及 Acceptor 之間要有光譜重疊才能產生螢光,若兩分子間距離過長,造成光譜無法重疊 則不會有螢光轉移效應產生。 2-11 淬熄效應 (Quenching) Quenching 為能量轉移效應的一種,其種類分為三種:1.碰撞淬熄 (collisional quenching)、2.靜態淬熄 (static quenching)、3.能量轉移 (energy transfer)[24] 。當螢 光分子 (fluorophore) 與 quencher 彼此接觸 (contact) ,才會有碰撞淬熄與靜態淬熄 的發生。常用淬熄分子:氧分子 (O2) 、丙烯醯胺 (Acylamide)、碘離子 (I-)、氨基
化合物 (Amide)、四氧化溴離子 (BrO4-)等。
碰撞淬熄效應 (collisional quenching) :研究指出螢光強度與 quencher 濃度呈反 比[168,169];當 fluorophore 處於激發態時,會與 quencher 碰撞而產生碰撞效應。另 外處於自然態(Native state)的蛋白質其 Tryptophan 處於分子內,其擴散係數(diffusion coefficient)較自由狀態下的 Tryptophan 小,所以不容易與 quencher 接觸產生較小 的淬熄效應。
靜態淬熄效應 (static quenching) :Fluorophore 處於激發態時,會與 quencher 形 成錯合物(Complex)[22,170,171],錯合物的形成易造成螢光強度降低。然而在靜態效 應中,未與quencher 結合的 fluorophore 仍然會放出螢光。
26
2-12 感應耦合電漿原子發射光譜法(Inductively Coupled Plasma Atomic Emission Spectrometry, ICP - AES)
ICP-AES 的基本原理乃是利用 ICP 產生之高溫可達 6000K,破壞吸入樣品 使其產生離子狀態,該離子裂碎物再進入質譜儀偵測。由於高溫,形成離子主要為原 子態之離子,或與基質結合之離子。 ICP-AES 為一種以感應耦合高頻電漿為光源之原 子發射光譜裝置(圖 12),其感應耦合高頻電漿激發源(圖 13),由三支同心軸石英管所 組成。 高頻電漿激發源向耦合線圈提供高頻能量,等離子炬管置於耦合線圈中 心,內通冷卻氣、輔助氣與載氣,於炬管中產生高頻電磁場。以微電火花引燃,讓部 分氬氣電離產生電子與離子。電子於高頻電磁場中獲得高能量,經碰撞將能量轉移給 氬原子,使之進一步電離產生更多電子與離子。該過程像雪崩似進行時,導電氣體受 高頻電磁場作用,形成與耦合線圈同心之渦流區。強大之電流產生高熱將氣體加熱, 從而形成火炬形狀之可自持電漿。試樣氣溶膠通過溫度較低之中心通道,故不易擴散 至 ICP 火焰周遭而產生自吸收冷蒸氣層。試樣由載氣引入霧化系統進行霧化,以氣溶 膠形式進入軸向通道,於高溫與惰性氬氣氣氛中,氣溶膠微粒被充分蒸發、原子化、 激發與電離。被激發原子與離子發射原子譜線與離子譜線,分光檢測系統將各待測元 素發射之特徵譜線經分光、光電轉換與檢測,經數據處理系統對實驗數據進行處理打 印輸出。 圖 12 ICP-AES 架構原理圖 ICP-AES 為一種以感應耦合高頻電漿為光源之原子發射光譜裝置。由高頻電漿激發源、等離子 炬管、進樣系統、分光系統、測光系統與數據處理系統組成。1.進樣系統裝置:利用氬氣進入
27 霧化器中產生交錯的氣流,將樣品霧化成微細分散液滴,再帶入電漿中,另外也可藉由超音波 霧化器將液體變成氣懸膠。2.高頻電漿激發源:由內中外三層同心圓石英管組成,分別氬器分 別由三層石英管下端輸入,外層石英管以切線方向通入,這些氬氣盤旋而上,一方面冷卻外層 石英管的內壁及將電將限制在管中心,另一方面也是主要的支撐氣流,中層石英管為輔助氣流, 中心則為載送霧狀試樣、粉末或氣體的載送氬氣流。電將在石英管上方開口處形成,這些開口 處繞以感應線圈,當感應線圈通入電流時,線圈周圍產生感應磁場,其磁力線垂直於石英管。 使石英管內充滿氬氣,再以一個Tesla Coil 在氬氣內引發電子或離子,便會使得氬氣在一瞬間大 量游離,產生高溫電漿。3.光柵掃描器:測量大量元素的發射譜線強度。4.光電轉換器:光電倍 增管。 圖 13 感應耦合電漿激發源之構造示意圖
依序分為外層管(outer tube)、中層管(intermediate tube)與內層管(inner tube);並於電漿火炬(torch) 之外圍環繞與高頻產生器(high-frequency generator)連接之感應線圈(induction coil)。