行政院國家科學委員會專題研究計畫 成果報告
研究登革病毒第三型專一性抗體的 B 細胞抗原決定位及評估
其為偵檢試劑之可行性
計畫類別: 個別型計畫 計畫編號: NSC92-2320-B-002-198- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學醫學院醫事技術學系暨研究所 計畫主持人: 高全良 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 11 月 29 日
行政院國家科學委員會專題研究計畫成果報告
計畫編號:NSC 92-2320-B-002-198
執行期限:92 年 8 月 1 日至 93 年 7 月 31 日
主持人:高 全 良 臺大醫學院醫事技術系
共同主持人:吳 漢 忠 臺大醫學院口腔生物研究所
計畫參與人員:陳韻晶
一、中文摘要 本計畫主要是要研究登革病毒的 B 細胞抗 原決定位。經由 ELISA 和 immunoblotting 方法,我們找出具專一性的第三型登革病 毒單株抗體。此抗體之抗原決定位的確 認,有助於病毒感染機制的暸解,疫苗及 偵檢試劑的研發。在此研究中,我們以噬 菌體顯現法來研究第二型登革病毒的抗原 決定位。我們也已經篩選出多株的噬菌體 可以專一性的與第二型登革病毒抗體結 合。引發出血性登革熱的登革病毒,持續 地在亞洲和南美洲的公共衛生上的構成嚴 重的問題。在此研究中,我們利用登革第 三型的病毒對 BALB/c 老鼠進行免疫,並且 以細胞融合技術產生對抗和第三型登革病 毒的單株抗體。這些單株抗體在 ELISA 測 試中對登革病毒有專一性的反應。一些單 株抗體對抗病毒的非結構性蛋白,另一些 對抗病毒的套膜蛋白。西方墨點分析法顯 示 DC7-33 和 DC14-33 與四型登革病毒的套 膜蛋白都有所反應。DC12-33 除了第四型登 革病毒以外對所有血清型的登革套膜病毒 DC36-3 同樣的只對抗第三型登革套膜病而 不會與其他血清型的登革病毒交叉反應。 其中有三株單株抗體在 PRNT 和免疫螢光染 色的測試中被證實對登革病毒的感染途徑 發揮中和性效用。我們相信這些登革的單 株抗體能夠爲偵檢試劑和疫苗的研發提供 一些有用的資訊。 關鍵詞:第二型登革病毒,噬菌體顯現法, 抗原決定位,登革偵檢試劑ABSTRACT
Dengue virus (DEN), causing dengue hemorrhagic fever (DHF), still present a public health problem in Asia and Southern America. In this study, monoclonal antibodies (mAbs) against DEN-3 were generated by fusing P3-NS1/-Ag4-1 mouse myeloma cells with lymphocytes from BALB/c mice immunized with purified DEN-3. MAbs were identified to react specifically to the DENs by ELISA. Some mAbs reacted to nonstructured protein 1 (NS1) and the others reacted to envelope proteins (E proteins). Immunoblotting analysis showed that DC7-33 and DC14-33 reacted to envelope proteins of all dengue serotypes. DC12-33 reacted to envelope proteins of all dengue serotypes except DEN-4. DC36-3 reacted only to envelope proteins of DEN-3. Three mAbs were further demonstrated to neutralize DEN infection by plaque reduction neutralization test (PRNT) assay. We believe that these mAbs and epitopes of DENs will provide information for development of virus-specific serologic diagnostic reagents and vaccines.
KEY WORDS: monoclonal antibody;
neutralization; vaccines; serologic diagnosis
INTRODUCTION
Dengue virus (DEN) causes serious febrile illness in humans, including dengue hemorrhagic fever (DHF) and dengue shock syndrome (DSS) (Henchal, et al. 1990; Halstead, 1988). Primary dengue virus infection often results in a painful, debilitating, but nonfatal dengue fever and
appears to against reinfection with the same serotype. However, the more severe and sometimes fatal forms DHF and DSS have been frequently seen in region where more than one serotype of dengue virus is circulating (Halstead, 1988; Gubler, 1998). Secondary infection with a different serotype is associated with an increased risk for DHF and maybe caused by the uptake by monocytes/macrophages of virus complexes
to nonneutralizing antibodies, sub-neutralizing cross-reactive antibodies or
low-titer neutralizing antibodies (Halstead, 1988; Bielefeldt-Ohmann, 1997; Halstead, et al. 1984). Viral infection usually results in the production of antibodies directed against the epitopes of the viral proteins through the activation of the host’s humoral immunity. The epitopes have been divided into linear or continuous epitopes and conformational or discontinuous epitopes (Sela, 1969; Barlow et al., 1986). Linear epitopes are short stretches of the primary structure of the protein and are made up of some continuous amino acid residues of the primary sequence. Conformational epitopes consist of several amino acid residues which are discrete in the primary sequence but assemble to form an antigenic determinant on the tertiary structure of the native protein. Identification of these epitopes on viral proteins is important in understanding the pathogenesis of viral infectious diseases as well as in developing effective vaccines and diagnostic reagent.
Recent advance in peptide technology has allowed the development of combinatorial peptide libraries expressed either on a solid phase support or displayed on bacteriophages. The high molecular diversity displayed by these libraries provides the possibility to study B-cell epitope mapping (Scott and Smith 1990; Young et al., 1997; Wu et al., 2001)
Recently, we have identified serotype-specific B-cell epitope of DEN-1 (Wu et al., 2001). In this study, we used a phage-displayed peptide library to identify the serotype-specific B-cell epitope for DEN-2. Up to now, it is still not clear whether DHF/DSS is due to a primary or secondary infection of DEN or other
immunopathologic mechanisms (Halstead, 1988; Gubler, 1998). Therefore, the identification of B-cell epitopes for DEN can provide important information for the development of a safe and effective dengue vaccine and contribute to the understanding of the pathogenesis and immunological responses in DEN infection.
RESULTS
Generation and identification of mAbs against DEN-1 and DEN-3
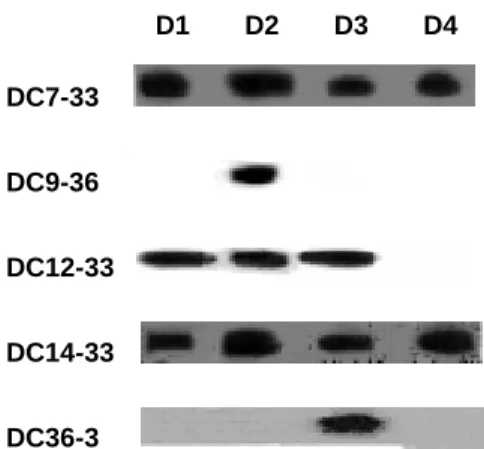
In this study, we generated serotype-specific and cross-reactive mAb against DEN-3. Immunoblotting and ELISA assays determined the reactivity of mAbs with all DEN serotypes. All of these mAbs reacted to envelope proteins (Tables 1; Figs. 1). Immunoblotting analysis showed that DC7-33 and DC14-33 reacted to envelope proteins of all dengue serotypes. DC12-33 reacted to envelope proteins of all dengue serotypes except DEN-4 (Fig. 2). DC36-3 reacted only to envelope proteins of DEN-3 (Fig.2). ELISA further confirmed the specificity of these antibodies (Fig. 1). C6/36 cells were infected with DEN-1, -2, -3, and -4. Cells were then fixed for ELISA analysis using the mAbs against DEN, normal mouse IgG (NM-IgG), and normal mouse serum (NMS). To confirm the binding specificity of mAbs, we also performed an indirect immunofluorescence assay. BHK-21 cells were infected with DEN-1, -2, -3, or -4.
The effect of dengue virus-specific neutralizing antibody in inhibition of dengue virus entry into BHK21 cells
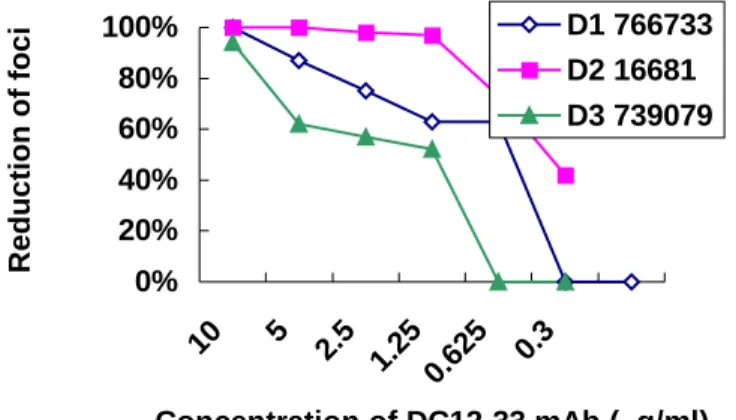
Figures 3 and 4 shows the effects of the mAbs in neutralizing dengue-1, -2, -3, and -4 virus with plaque formation. DC7-33 mAb showed a neutralizing effect on DEN-2 (50% reduction of plaque formation at 2.5 µg/ml), and low neutralizing effect on DEN-1 (50% reduction of plaque formation at 1 µg/ml). DC12-33 mAb showed a high neutralizing
effect on DEN-2 (50% reduction of plaque formation at 0.625 µg/ml), and low neutralizing effect on DEN-1 and DEN-3 (50% reduction of plaque formation at 5
µg/ml in DEN-1 and 2.5 µg/ml in DEN-3). In contrast, DC14-33 and the normal mouse IgG did not show any inhibitory effect.
Comparison of the results obtained in IgG-ELISA using human IgG and mAbs
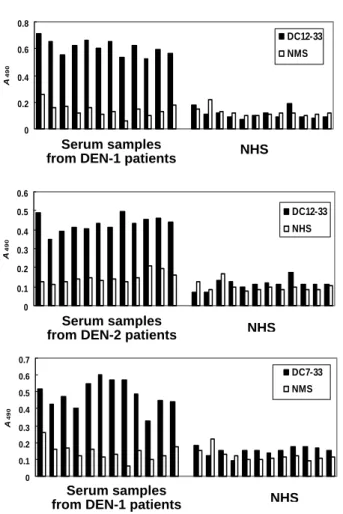
Twelve serum samples of DEN-1 patients and twelve serum samples of DEN-2 were tested in ELISA with mAbs (Figs. 5). All of DEN-1 patients could be detected by DA11-13, DC7-33, and DC12-33 mAbs. All of DEN-2 cases were positive by the use of DC12-33 as detector antibody. Eight of twelve DEN-2 cases were positive by the use of DC7-33 as detector antibody.
DISCUSSION
In this study, we have generation mAbs against E proteins of DEN-3. These mAbs will be useful to study neutralizing epitopes and develop diagnostic reagents for DEN. Our developed method can also be applied to detect future DF and DHF patients who had secondary infection with a heterologous serotype of DEN, which would minimize possible morbidity and mortality. Finally, our test will be very valuable for further development of a serotype-specific diagnostic reagent that can be used to serological distinguish four serotypes dengue patients and thus help combat dengue diseases.
REFERENCES
1. Barlow, D. J., M. S. Edwards, and J. M. Thornton. (1986). Continuous and
discontinuous protein antigenic determinants. Nature 322: 747-748.
2. Bielefeldt-Ohmann, H. (1997).
Pathogenesis of dengue virus diseases:
missing pieces in the jigsaw. Trends Microbiol. 5: 409-413.
3. Folgori, A., R. Tafi, A. Meola, F. Felici, G. Galfre, R. Cortese, P. Monaci, and N. Alfredo. 1994. A general strategy to
identify mimotopes of pathological antigens using only random peptide libraries and human sera. EMBO J. 13: 2236-2243.
4. Gubler D. J. 1998. Dengue and dengue
hemorrhagic fever. Clin. Microbiol. Rev.
11: 480-496.
5. Gubler, D. J., and G. G. Clark. 1995.
Dengue/dengue hemorrhagic fever: the emergence of a global health problem. Emerg. Infect. Dis. 1: 55-57.
6. Halstead, S. B. 1989. Antibody,
macrophages, dengue virus infection, shock, and hemorrhage: a pathogenetic cascade. Rev. Infect. Dis. 11: S830-839.
7. Halstead, S. B. 1988. Pathogenesis of
dengue: challenges to molecular biology. Science 239: 476-481.
8. Halstead, S. B., C. N. Venkateshan; M. K. Gentry; and L. K. Larsen. 1984.
Heterogeneity of infection enhancement of dengue 2 strains by monoclonal antibodies. J. Immunol. 312, 1529-1532.
9. Henchal, E. A., and J. R. Putnak. 1990.
The dengue viruses. Clin. Microbiol. Rev.
3: 376-396.
10. Lam, S. K., and P. L. Devine. 1998.
Evaluation of capture ELISA and rapid immunochromatographic test for the determination of IgM and IgG antibodies produced during dengue infection. Clin. Diagnostic Virol. 10: 75-81.
11. Scott, J. K., and G. P. Smith. 1990.
Searching for peptide ligands with an epitope library. Science 249: 386-390.
12. Scott, J. K., and G. P. Smith. 1990.
Searching for peptide ligands with an epitope library. Science 249: 386-390.
13. Sela, M. 1969. Antigenicity: Some
molecular aspects. Science 166: 1365-1374.
14. Vaughn, D. W., A. Nisalak, S. Kalayanarooj, T. Solomon, N. M. Dung, A. Cuzzubbo, and P. L. Devine. 1998.
immunochromatographic test for diagnosis of dengue virus infection. J. Clin. Microbiol. 36: 234-238.
15. Vaughn, D. W., A. Nisalak, T. Solomon, S. Kalayanarooj, M. D. Nguyen, R. Kneen, A. Cuzzubbo, and P. L. Devine.
1999. Rapid serologic diagnosis of dengue virus infection using a commercial capture ELISA that distinguishes primary and secondary infections. Am. J. Trop. Med. Hyg. 60: 693-698.
16. Young, A. C., P. Valadon, A. Casadevall, M. D. Scharff, and J. C. Sacchettini. 1997. The three-dimensional
structures of a polysaccharide binding antibody to Cryptococcus neoformans and its complex with a peptide from a phage display library: implications for the identification of peptide mimotopes. J. Mol. Biol. 274: 622-634.
Table 1. Generation and characterization of monoclonal antibodies against DEN-3
Western blotting ELISA PRNT50 (µg/ml) mAb D1 D2 D3 D4 D1 D3 D1 D2 D3 D4 Specificity DC7-33 + + + + + + < 1 < 0.25 - - E DC9-36 - + - - + + - < 0.625 - - E DC12-33 + + + - + + < 5 < 0.625 < 2.5 - E DC14-33 + + + + + + - - - - E DC36-3 - - + - - + ND ND ND ND E
ND = not determined E = envelope proteins NS-1 = non-structure proteins 1
Figure 1. Comparison of reciprocal ELISA titers of anti-DEN-3 monoclonal antibodies against DEN-1 and DEN-3 infected C3/36 cells. 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 DC12-33 DC9-36 DC7-33 DC14-33 DC136-3 Pre-immune serum A 490 D1 766733 D3 739079 MOCK VB
Figure 2. Identification of mAbs against E proteins of DENs by immunoblot analysis. Four serotypes of DEN antigens from DEN-infected C6/36 cell lysates were size-fractionated in polyacrylamide gels. The blots were incubated with mAbs. E proteins (55kDa) of DEN-3 were identified by immunoblot analysis using non-reducing gel.
0% 20% 40% 60% 80% 100% 10 5 2.5 1.25 0.63 0.3 Concentration of DC7-33 mAb ( g/ml) R e duc tion of foc i D2 16681 D1 766733
Figure 3. In vitro neutralization of DEN strains by neutralizing mAb DC7-33. The ascitic fluid of DC7-33 was purified by a protein G sepharose column. The neutralizing activity of the mAb was tested by PRNT against DEN-1 strain 766733 and DEN-2
strain 16681. µ DC7-33 DC9-36 DC12-33 DC14-33 DC36-3 D1 D2 D3 D4
0% 20% 40% 60% 80% 100% 10 5 2.5 1.25 0.625 0. 3 Concentration of DC12-33 mAb ( g/ml) R e duc tion of foc i D1 766733 D2 16681 D3 739079
Figure 4. In vitro neutralization of DEN strains by neutralizing mAb DC12-33. The ascitic fluid of DC12-33 was purified by a protein G sepharose column. The neutralizing activity of the mAb was tested by PRNT against DEN-1 strain 766733, DEN-2 strain 16681 and DEN-3 739079.
Figure 5. Capture ELISA for serum samples from patients with DEN infection. The serum samples from patients with DEN-1 and -2 infections were analyzed, and representative data are shown to illustrate the mAbs responses.
Serum samples from DEN-1 patients
NHS Serum samples
from DEN-2 patients
NHS 0 0.1 0.2 0.3 0.4 0.5 0.6 A 490 DC12-33 NHS Serum samples
from DEN-1 patients NHS
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 A 490 DC7-33 NMS 0 0.2 0.4 0.6 0.8 A 490 DC12-33 NMS