電場對蛋白質(Hen Egg White Lysozyme)晶體成長機制以及形貌的影響
Crystal Growth Mechanism and Morphology of Hen Egg White Lysozyme
under Electric Field.
研 究 生:李姿玲 Student:Tzu-Ling Lee 指導教授:吳耀銓 Advisor:YewChung Sermon Wu 國 立 交 通 大 學 材料科學與工程研究所 碩 士 論 文 A Thesis
Submitted to Department of Materials Science and Engineering College of Engineering
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Materials Science and Engineering July 2004
電場對蛋白質(Hen Egg White Lysozyme)晶體成長機制以及形貌的影響
研究生:李姿玲 指導教授:吳耀銓博士 國立交通大學材料科學與工程研究所碩士班摘
要
為了瞭解蛋白質結構與功能的對應關係,以及根據蛋白質結構設計新藥物的開發,以 X-ray 繞射方法得到其結構是達成此目的的第一步。而其中 X-ray 繞射的瓶頸又在於難以得 到足夠大且結構完美的生物巨分子晶體。各類關於生物巨分子結晶的研究-包括實驗操作以 及原理推論兩部分-也因應而生。使生物巨分子晶體學從80 年代早期起成為獨立的研究學 門。 1999 年始有團隊開始研究電場對於生物巨分子結晶的影響。本實驗利用微影製程於矽晶 圓上設計一小尺寸系統,觀察Hen Egg White Lysozyme (HEWL)分子在外加電場下的結晶狀 況,以光學顯微鏡(Optical Microscopy)和原子力顯微鏡(AFM),分別得到巨觀形狀和微觀形貌 的影像,與一般無電場條件下所得之結晶不同之處如下所列,並於文中嘗試推測所列結果的 形成機制。
整體溶液未達飽和,也有結晶在陰極析出。2.外加電場系統中所得之 HEWL 晶體形狀上較一 般Tetragonal 形式之 HEWL 結晶多出一未知平面。3.外加電場系統中 HEWL 晶體之[-110]方 向有沿著電場方向排列的趨勢。
在微觀上,相較於一般無電場條件下所得之結晶,外加電場下所得之HEWL 結晶{110}面上
Crystal Growth Mechanism and Morphology of Hen Egg White Lysozyme
under Electric Field
Student : Tzu-Ling Lee Adviser: Dr. Yew-Chung SermonWu
Department of
Materials Science and EngineeringNational Chiao Tung University
ABSTRACT
This experiment use lithography and etching to fabricate a device on Si wafer to manipulate and observe the crystal growth of Hen Egg White Lysozyme under external electric field. Image and morphology were taken by both optical microscopy and atomic force microscope. The difference with respect to normal growth is list below and possible mechanisms are proposed in the text.
Macroscopically, the crystal growth in external electric field shows the following characteristics: 1. At pH~4.6, crystals precipitate near cathode even when the solution is undersaturated.
2. An unknown face appears on the crystal of the tetragonal form HEWL crystal. 3. The [-110] direction of the HEWL crystal appears to be align with the electric field.
Microscopically, besides steps parallel to [-110] direction that is also observable on the crystal without external electric field, there exist steps parallel to [001] direction under electric field.
第二章 前言
首先要感謝指導教授吳耀銓老師,給我機會挑戰這樣新興的題目。一個人要從選定題目、 實驗設計到整個實驗的執行和討論委實不易,但因此碩士班所學得的東西比我想像多很多。 實驗過程之中要感謝楊裕雄老師在實驗設計之前提供建議;李志偉老師、黃仁清老師提供原 子力顯微鏡的協助;呂宗育、周重光兩位同學在他們自己課餘的時間能夠撥空跟我討論;還 有林其慶學弟在專題期間跟著我忙東忙西;以及實驗室的諸位學長同學們,平常的照顧和幫 忙,才得以讓我得到現有的結果。 當然最重要還是要感謝我親愛的爹娘,扶養他們女兒到二十幾歲,還在我最後寫論文時 期提供生活上完全的支持。 一個實驗的開始實在很不容易,在這期間受到許多人的幫忙也讓我學到許多處理事情的 方法。目前實驗已有階段性的成果,歸功於大家的協助,不管這個實驗能否繼續承傳,我想 開始一項新研究的好奇心和做實驗應俱備的嚴謹性都更應該承傳下去。目錄
第一章 摘要……..………...…………1 中文摘要………...1 英文摘要……..……… ..3 第二章 前言……….4 第三章 文獻回顧………...12 3-1 蛋白質結晶……….………....12 3-1-1 簡介……….………….…....12 3-1-2 常用方法簡介……….…………...15 3-1-2-1 Vapor Diffusion………...16 3-1-2-2 Batch Method………..………….18 3-2 蛋白質結晶的成長機制以及其他物理參數導入的影響………...19 3-2-1 成核………..………....20 3-2-1-1 量測儀器………..……….……...21 3-2-1-2 成核機制………..………...21 3-2-2 成長……….………...22 3-2-2-1 量測儀器………..…...22 3-2-2-2 成長機制……….243-2-2-3 導入物理參數的影響……….32 3-2-2-4 關於本文實驗………...37 第四章 實驗設計及流程...38 4-1 結晶……….………38 4-2 原子力顯微鏡(AFM)檢測………….………...………...40 第五章 實驗結果………...44 5-1 溶解度………...………..44 5-2 由光學顯微鏡(OM)所得之照片………...……….45 5-2-1 一般狀態下所得之晶體巨觀形狀….……….………..45 5-2-2 外加電場下所得之晶體巨觀形狀…..………..46 5-2-3 電流導通時所得之晶體巨觀形狀 Æ 意外的發現..………...47 5-3 由原子力顯微鏡(AFM)取得之數據………..……….………...50 5-3-1 一般狀態下所得之晶體微觀形貌………..………..50 5-3-2 外加電場下所得之晶體微觀形貌………..………..51 5-4 小結……….…………..………..53 第六章 結果討論………..54 6-1 電位及電場在水溶液中之分布情形………...………..54 6-2 成核……….………..57 6-2-1 陰極附近溶液與氧化層的界面(50~100nm),直接受電場影響
的分子………...57 6-2-2 遠離溶液與氧化層的界面(距界面 100nm 以上)的 HEWL 分 子………...59 6-2-3 小結……..………...…59 6-3 成長………..62 6-3-1 由 OM 所觀察得到 HEWL 晶體巨觀形狀上的變…………...62 6-3-2 由 AFM 所觀察得到 HEWL 晶體微觀形貌上的變化………...71 第七章 結論……….73 第八章 展望……….74 參考資料……….75
圖目錄
圖3-1 hanging drop 示意圖……….………..16 圖3-2 Linbro plate……….……….17 圖3-3 過飽和度與巨分子晶體成長的關係圖………...20 圖3-4 成核數目與驅動力關係圖………...21 圖3-5 dense fluid 示意圖………...………22 圖3-6 原子力顯微鏡裝置示意圖………...23 圖3-7 原子力顯微鏡操作模式………...24 圖3-8 晶體成長模式整理………...24圖3-9 Growth Sites(step 以及 screw dislocation 上的 kink 位置)…………...25
圖3-10HEWL 晶體巨觀形狀………...27
圖3-11 過飽和度對 HEWL 晶體各 facet 成長速度的影響………...28
圖3-12 HEWL 晶體表面微觀形貌………...………...29
圖3-13 HEWL 晶體{110}分子結構圖……….………...30
圖3-14 以蛋白質分子表示 HEWL 晶體{110}鍵結圖………...31
圖3-15 HEWL 表面 dangling bond 分佈示意圖…………...………...31
圖3-16 晶體成長反應步驟圖…..………...32
圖3-17 反應決定步驟與晶體品質的影響………...33
圖3-19 電場下析出 HEWL 結晶在溶液中之分佈圖(光學顯微鏡)……...36 圖4-1 實驗流程圖………..………..39-40 圖4-2 AFM 試片架設裝置示意圖………...………...41 圖5-1 不同析出劑濃度下所對應的溶解度………...44 圖5-2 無外加電場下所得 HEWL 之 OM 影像………...45 圖5-3 有無外加電場 HEWL 晶體巨觀形狀比較圖………...………47 圖5-4 外加電流時所得 HEWL 晶體之 OM 影像……….48 圖5-5 外加電流時所得 HEWL 晶體之連續 OM 影像……….49 圖5-6 無外加電場時 HEWL{110}上之 AFM 影像………50 圖5-7 外加電場時 HEWL{110}上之 AFM 影像(1)……….51 圖5-8 外加電場時 HEWL{110}上之 AFM 影像(1)局部放大……….51 圖5-9 外加電場時 HEWL{110}上之 AFM 影像(2)………...52 圖6-1 35V,鄰近界面部份,HEWL 溶液中電位的變化……….55 圖6-2 35V,鄰近界面部份,HEWL 溶液中電場的變化……….55 圖6-3 100V,鄰近界面部份,HEWL 溶液中電位及電場的變化……….56 圖6-4 電場(電位)在溶液中的分佈示意圖……….56 圖6-5 有無外加電場下,擴散之化學位能示意圖……….57 圖6-6 隨距離改變,電場誘導 HEWL 產生偶極的能力……….58
圖6-8 有無外加電場時,HEWL 晶體巨觀形狀比較示意圖……….62 圖6-9 未知平面(x0z)示意圖………...………63 圖6-10 外加電場下 HEWL 晶體成長過程示意圖.………...66 圖6-11 高過飽和度之 HEWL 晶體之巨觀形狀 OM 影像以及示意圖…….68 圖6-12 有無外加電場時,各平面形成之能障以及自由能差比較示意 圖………..…………...69 圖6-13 HEWL 晶體{110}面之分子鍵結以及有無外加電場時表面形貌比 較示意圖………..………..71
表目錄
表5-1 不同外加電場下所得 HEWL 之 OM 影像………..46
表5-2 有無外加電場與電流所得之 HEWL 晶體,巨觀和微觀上的特徵比
第三章 文獻回顧
3-1 蛋白質結晶
3-1-1 簡介
蛋白質是建構所有生物體的基本單位,而且是活體系統中能量和訊息傳遞過
程中(information and energy processing)必備的構成要素。為了瞭解基因組(genome)對應於蛋白質 (可能是未知性質的一連串胺基酸或是核苷酸(nucleotide)序列)的 “結構-功能"關係、探討巨分子如何在一個生物系統中作用,甚至是新藥物(抑制劑)的開發, 而開啟了近年來在蛋白質結晶這領域的研究123 。 不同蛋白質、核酸(nucleic acid)、以及蛋白質-核酸複合物(complex)的數量,相較於已知無 機物可能組成的相(phase),可能是數十萬倍以上。此種情況造成我們至今無法以定量的方式 預測一種新的生物巨分子的結構或是結晶狀況2 。另一方面,雖然逐漸進步的核磁共振(NMR) 技術正提高可接受蛋白質分子尺寸的上限;以繞射方法,包括:X-ray、電子以及中子繞射, 仍然是現今檢測蛋白質結構的主流 4 。 與一般無機材料不同的是蛋白質的結晶中還納入了溶液中大量的水分子、離子以及少量 的中性分子2 ,而且晶體中蛋白質巨分子互相接觸用以穩定晶體結構的鍵結,比起一般無機物 或是簡單有機物複合物的結晶顯得相對脆弱且為數稀少,且晶體之中存在大量不規則排列的 溶質通道,由於以上原因,使得在 X-ray 繞射圖形下才會發現,即使巨觀上看起來是各晶面 整齊漂亮的蛋白質晶體,實際上並不一定是高度組成以及結構統一之巨分子秩序排列的晶體
3 。為了提高解析度至 1.5~2Å4 ,X-ray 等各類繞射方法需要足夠大(通常是 0.3~1.3mm)、低缺陷、 高組成統一,以及結構統一的單晶。日前蛋白質的表達(expression)、純化、檢測儀器發展以及 繞射數據的電腦分析系統,都有快速的成長以提高解(solve)蛋白質結構的速度。最大的瓶頸惟 獨存在於:製備高品質的蛋白質晶體以提供 X-ray 繞射的素材 3 4 。 在結晶的過程之外,許多生物化學或是生物醫學方面的研究,常常受限於因為無法完全 透視蛋白質成核成長的機制而窒礙難行。例如在藥物釋放方面,因為晶體較為緩慢的溶解速 率,可以達成較穩定長時間藥物治療的目的,已被使用在胰島素和其他蛋白質藥物上4 。在這 方面的需求上,體積大且等體積而數量少的晶體能夠達成緩慢且穩定之溶解速率的要求,當 然遠遠勝過體積較小而數量多的晶體。為了達成這樣的目的,需要縮短成核時間,並使所有 晶體都於相同遞減的過飽和度下成長4 。 其他結晶在生物醫學上相關的研究包括:在人體內因為蛋白質結晶或是其他規則性的固 體聚集造成的疾病,例如鐮刀型貧血症以及白內障4 ;蛋白質工程以及合成疫苗的設計3 。 蛋白質的分子大(幾個 nm)、成長速度慢(成長速率可以秒計算),滿足於現今表面檢測 工具的限制,可以直接檢測出成長實況。因此蛋白質晶體的成長,也為傳統基礎的晶體學和 成長機制的研究提供理論或是實證上的貢獻4 。 綜合以上各項因素,蛋白質結晶大約於 80 年代早期成為一門獨立的研究領域4 。如前所 述,受限於此系統龐大的複雜性,目前尚無方法可以預測一未知蛋白質可能的結構或是可能
得蛋白質結晶逐漸成為一門較偏向經驗性(empirical search)的學門,眾多回顧蛋白質結晶的文 獻,常常以“蛋白質結晶是一門藝術而非科學…"作為開端3 。而此經驗性的研究方法大致可 分為兩步驟 5 : 1. 從錯誤中嘗試找到適當的溶液,使蛋白質能夠結晶而不是非晶質的析出物。 2. 調整溶液的比例和濃度,使晶體長到適合的大小,足以提供 X-ray 繞射檢測 結構。 雖然以目前的狀況看來,從嘗試錯誤中個別找出適合不同蛋白質的結晶條件,似乎能達 到較佳的結晶效率。但是如何將蛋白質結晶在藝術之外,真正納入科學可以接受的領域?部 分相信,實驗結果顯示,由於蛋白質結晶是一種過於龐雜的過程和系統,很難歸納出有系統 的基本原則;然而近十年來,眾多研究開始著重於各情況下成核以及成長機制的推演。在眾 多研究中,推衍出一個合理的規則系統,才是這些方法背後迫切需要3 而且最終應該達成的目 的。深度的理論研究和實驗技術上操作的同時並進,才能真正促成一門學問的發展與演進2 。 在蛋白質成長機制的研究中,以Hen Egg White Lysozyme (HEWL)作為實驗材料者,佔 有相當顯著的比例 4 6 。HEWL 這種醣類的水解酵素,是最早利用 X-ray 解出結構的蛋白質之 一7 。除了因為長期的研究使得這個蛋白質累積相較於其他種類蛋白質為多的實驗經驗和參數 外,不少實驗已證實以HEWL 為基礎所推衍出的成長機制適用於其他廣大範圍的蛋白質4 。 HEWL 也常使用於蛋白質 folding dynamic 以及其他蛋白質相關的基礎研究中4
。
本實驗的目的是觀察在外加電場環境下,蛋白質晶體成長機制的變化-以HEWL 為例。
構、成長機制和檢測技術,接著導入本實驗進行的流程設計。 3-1-2 常用方法簡介 與一般傳統的結晶方式相同,要開始蛋白質結晶的第一步驟即是使溶液中的溶質濃度達 到足夠的過飽和度。當溫度、pH 值或其他因素改變時,巨分子靜電特性、表面型態以及結構 都可能隨之改變,同時也影響蛋白質溶液的溶解度。除了溫度和pH 值,蛋白質結晶系統中 另外一個較為常用的參數是析出劑(precipitant)。析出劑通常是鹽類或高分子:鹽類會跟蛋白 質競爭水分子;高分子,如PEG (polyethylene glycol)會在溶液中與水作用,逐漸擴大體積而 迫使水中的巨分子擠在一起,在其他條件相同的狀況下,使蛋白質溶液的濃度提高。基本的 巨分子結晶方法就是在定溫下調控pH 值以及嘗試找出可能的析出劑,不斷的試驗之中,慢 慢降低溶解度,提高巨分子濃度,以達到足夠的過飽和度。 目前可用以結晶巨分子的方法如下所列: 1. Bulk Crystallization 2. Batch Method 3. Evaporation 4. Bulk Dialysis 5. Concentration Dialysis 6. Microdialysis 7. Liquid Bridge
8. Free Interface Diffusion 9. Vapor Diffusion 10. Sequential Extraction 11. pH-Induce Crystallization
一般最常使用且成功率最高的兩者為2. Batch Method 以及 9. Vapor Diffusion 。在本小節中 僅就此兩方法的原理、操作方法以及優缺點作一介紹。
3-1-2-1 Vapor Diffusion
Vapor Diffusion 是目前最廣為使用的方法,適合用來篩檢(screen)大量、不同溶液的各類
結晶情形,從中選取適合的溶液,再加以調整濃度,成長出可以用來做X-ray 繞射試驗大小
的晶體。
Vapor Diffusion 依支撐液體方式的不同可分為 hanging drop、sitting drop 以及 sandwich。右圖 3-1 所示為 hanging drop。
圖3-1 Cdrop (圖中 drop 的濃度)不能達到足以析出結晶的過飽和度。且 C析出劑indrop< C析出劑in reservoir。因蒸汽壓隨鹽類析出劑的濃度增加而降低, 所以液滴中的水氣會逐漸揮發(液滴中的析出劑濃度一定相對 reservoir 為高),直到最終系統內液滴和 reservoir 的蒸汽壓達到平衡。驅使液
滴發生過飽和現象。(C: concentration)
雖然支撐方法不同,但這三種方式的基本原理是類似的。首先必須將固定pH 值之緩衝溶液
(buffer solution)內加入析出劑。析出劑和緩衝溶液的混和溶液稱為 reservoir。如上圖 3-1 所示。 希望結晶出現的液滴(drop)中可能由極小量的蛋白質溶液,reservoir,或是其他析出劑所組成, 液滴內析出劑的量必須少於可以在溶液中析出蛋白質的量,也就是液滴濃度不能達到足以析
許reservoir 和液滴中的水或其他揮發性溶劑進行交換。因蒸汽壓隨鹽類析出劑的濃度增加而 降低,所以液滴中的水氣會逐漸揮發(液滴中的析出劑濃度一定相對 reservoir 為高),直到最 終系統內液滴和reservoir 的蒸汽壓達到平衡。這樣的過程激發了至少兩項行為驅使液滴發生 過飽和現象: 1. 提高液滴中析出劑的濃度。 2. 提高液滴中蛋白質的濃度。 以此原理在可商業取得的 Linbro plate (如下圖所示)的每個凹槽中,設計不
圖3-2 Linbro plate。每個凹槽中,設計不同種類溶液混合的 reservoir,即可初步檢索出最適合於某種巨分子晶體生長的容易
條件
同種類溶液混合的reservoir,進行養晶實驗,即可初步檢索出最適合於某種巨分子晶體生長
的容易條件。完成初步的檢索後,再微調溶液濃度,培養體積和品質足以使用於X-ray 繞射
的巨分子晶體。
但是在絕大多數的 Vapor diffusion 系統中,液滴和 reservoir 之間必定存在空氣,而當操 作的液滴體積在幾個微升(micro Liter、uL)時,即使是少量的空氣也會使液滴中的水氣揮發,
3-1-2-2 Batch Method
Batch Method 是結晶和蛋白質晶體最古老的結晶法。此種方法的基本原理即是:直接把 未飽和的蛋白質溶液混入析出劑。析出劑加入後會調整原來蛋白質溶液的濃度使巨分子達到 足夠的過飽和度而析出成核。因為他的方便性,常用於設計觀察蛋白質晶體成長過程的影像 攝錄裝置中,或是用於粒徑分析(light scattering)、干涉式光學顯微鏡(interferometric optical microscopy)、原子力顯微鏡(atomic force microscopy)等各類分析時的操作5。此法中所需的蛋 白質溶液少則幾微升(uL)多則可達幾百毫升(mL)。小體積的 Micro Batch Method 為了避免水
分揮發,常搭配礦物油的使用5。近期也發展出利用不同密度的礦物油將小體積液滴置於兩種
不同密度的油之間,以懸浮不碰觸器壁的方式結晶。但此種方法不適用於需要緩慢地增加濃 度以達到足量過飽和度的某些蛋白質。
此外,micro batch method 由於析出劑和未飽和之蛋白質溶液接觸時,立即造成蛋白質溶 液濃度提高,故無論是將析出劑注入蛋白質溶液或是將後者注入前者混合時,微量分注器 (micropipette)滴入的速度以及注入溶液分佈都有可能造成不同的局部過飽和狀況,導致同樣 變因的不同組別實驗可能發現不同的成核數目或是晶體數目,此現象容易發生於低濃度的蛋 白質溶液2 ,造成定量再現的困難。 本實驗中為了方便原子力顯微鏡的觀察,以及維持系統的密閉性,使用 Micro Batch Method 養晶,並配合礦物油的使用。 以上介紹兩種方法是目前最廣為使用且最有效率能取得蛋白質晶體的養晶方式。但是如 前所述,此兩法成功地培養出蛋白質晶體基礎均憑依於實驗經驗,以及由嘗試錯誤所得到的
資訊之上。一門較為成熟的科學領域中,深度的理論探討和實驗技術理應是並進前趨的。另 一方面,近十年來也有許多研究開始著眼於各物理條件對於巨分子晶體成長的影響,得利於 其他現有的量測儀器的發展,同時觀察巨分子晶體的成長並歸納其機制。在下一節中,本文 將回顧現今已建構之巨分子晶體成長機制,以及部分物理參數對巨分子晶體成長系統所帶來 的影響。
3-2 蛋白質結晶的成長機制以及其他物理參數導入的影響
過飽和 溶質加入可使之溶解的溶劑之中,溶質即會在此環境下開始溶解,直到溶解進入溶劑之 溶質的速率與由溶液析出之溶質速率相等。系統達到平衡的狀態,此時之濃度稱為此種溶質 在此溶液以及其他此時之物理參數下的溶解度。溶質濃度低於溶解度稱為未飽和溶液 (undersaturated);若溶質濃度高於溶解度則稱為過飽和溶液(supersaturated)。本文定義之過飽 和度為:溶液濃度(C)/此條件下溶解度(Cs) 巨分子結晶學與一般傳統結晶學仍存在許多相似的地方:首先必須要達成過飽和的條 件,使得成核得以發生,成核之後接著則是晶體的成長。下文將分節討論成核以及成長。圖3-3 過飽和度與巨分子晶體成長的關係圖5。橫軸為析出劑濃度,縱軸為飽和度。當溶液於未飽和狀態時 (C/Cs<1),不會有結晶析出;隨溶液濃度提高逐漸達到過飽和的狀態,始有成核與晶體成長的現象發生。 3-2-1 成核 溶質濃度必須超過飽和度為成核的必要條件之一。當驅動力(driving force,此時為過飽 和度)大到足以讓溶質析出而聚集,並可以繼續存在、或是成長成晶體,而不溶解回溶液中(成 長所降低的自由能可以克服成長時體積增加造成表面增加所引發的自由能提高),此時析出聚 集之溶質稱為“核(nucleus)",溶質凝聚的過程稱為“成核(nucleation)"。 因為能取得高純度的生物巨分子材料數量較少,也較難成長出結構完美缺陷低的晶種 (seed)。與一般成長半導體、金屬或是其他傳統晶體不同的是,巨分子結晶是由自發性 (spontaneous)成核開始,而不是藉由晶種的引導成長2 。又由前述架構於經驗法則上的養晶方 式已可以得到好的晶體,目前的研究現況多專注於巨分子自動成核的事實,而不是如何造成 這樣成核結晶的過程。如此對成核系統薄弱的了解與控制,也削弱了對整個結晶系統的控制: 成核數目對應最後晶體的大小(核數目多,晶體較小);又因為巨分子晶體的塑性(plasticity)較
差,成核時造成的缺陷將無法在續成長過成中彌補2 。
3-2-1-1 量測儀器
成核時的動態分析多使用粒徑分析儀(static or dynamic light scattering)來進行5 8 9 10 。其 原理為利用雷射光行經溶液中,粒子產生之散射造成的波程差,求得平均粒徑11 。應用在此, 可以測得核的數目及平均大小。 3-2-1-2 成核機制 另外1999 年 Galkin 和 Vekilov 發表的論文中12 ,將量測傳統成核速率的方法移植於蛋白 質的成核過程。此種稱為“development”的方式,是將同條件的溶液至於同樣足夠大的過冷度 (相關於過飽和度)下,持續不同時間(Δt1,下圖 3-4 之橫軸),再將溶液升高至某一特定溫度(此 溫度因過冷度過小,以不足以誘發新的核產生),使在方才高過冷度時產生的核尺寸得以生長 至可觀察的大小。如下圖所示 圖 3-4 成核數目 v.s. 在大過冷度下提供之成核時間(驅動力)。斜率為成核速率。(ref 11) 斜率即為成核速率(J)。在均質成核(homogeneous nucleation)的區域,成核速率與過飽和程度 (σ=(C-S)/S)的關係為 ~exp( 2) 3 σ γ const J − ,其中C 為溶液濃度,S 為溶解度,γ 為晶體與溶液 間之表面能(約0.6mJ/m2)。此與傳統之晶體學中成核速率相較13 ,得到類似的結果,均與正比
2 2 2 3 * hom * hom 0 0 hom ) ( 1 ) 3 16 ( ) exp( T L T G KT G C f N v m SL ∆ = ∆ ∆ − = πγ 於exp( 2) 3 σ γ :其中N hom為均質成核速率,∆Ghom* 為成核時所需越過的臨界自由能能障。 與傳統小分子結晶學不同的是,蛋白質或是其他巨分子溶液在成核前,液體中存有相分離 (liquid-liquid phase separation)的現象,如圖 3-514
所示,
(a) (b)
圖 3-5(a)圖中若有一固定溫度 T 橫過 fluid-fluid 線,交於兩點,三部份,此時溶液分為 protein-depleted 和 protein-enriched 兩種, 即 liquid-liquid separation。(b)表示 dense fluid 的現象出現於成核前,並有助於晶體成長。
近年來的研究顯示,相分離中較溶質密集的溶液(dense fluid)可能在成核之前先形成,並 有助於成核過程的發生3 14 15 16 。 3-2-2 成長 溶液中的溶質按一定秩序吸附於已穩定存在的核上,使晶體尺寸變大的過程稱為成長 (growth)。 3-2-2-1 量測儀器
相較於成核,成長時的晶體尺寸較大,影像也可利用現有的儀器觀察。由早期的光學顯 微鏡(optical microscopy)17 18 19,電子顯微鏡(electric microscopy)20 21開始,至晚近被廣為使 用的原子力顯微鏡(atomic microscopy)8 22 23 24 25和干涉式顯微鏡(interferometric
microscopy)26 27 28 29逐漸累積豐富的資訊可供參考。 干涉式顯微鏡 干涉式顯微鏡是利用表面高低不同,光程差可能造成相消或相加的效果成象。由於其可 觀察範圍較大,遍及整顆晶體(數百um),常使用於觀察晶體成長速度(step rate,下文將作較 詳細解釋)以及成長動力學上的探討。 原子力顯微鏡 原子力顯微鏡利用試片表面高低不同時,探針尖端與試片表面原子間凡得瓦力作用也隨 之改變的原理成像,使探針與試片表面距離(const. force mode / contact mode) 改變,
圖 3-6 原子力顯微鏡(AFM)在液態中檢測生物巨分子的裝置(ref.8)。當圖中探針掃過表面高低不同的區域時,打在探針上的雷 射光束會隨之偏移,再由系統轉換成表面形貌的圖像。
更有敲拍法(tapping mode)的發明,盡量減低了探針對試片物理性的傷害,又由於可在常溫常 壓的空氣或是液態中進行觀察,更使原子力顯微鏡成為生物性樣本檢測上的利器8 。 圖 3-7 三種原子力顯微鏡(AFM)的操作模式30 。接觸法時,探針直接接觸試片表面並施予一固定力,故探針將隨 表面高低起伏。非接觸法時,探針與試片表面固定高度,依探針與試片表面作用力間接得到表面形貌。敲拍法 時,探針以一固定頻率敲拍試片表面,得到表面形貌資訊。 3-2-2-2 成長機制 由目前的文獻資料得知,所有的晶體成長模式約可以下圖表示: 圖 3-8 晶體成長模式整理。右側照片以及左側微觀 2 圖片部分參照 Ref 8 Tangential Layer growth normal growth 2D-nucleation (巨分子晶體:發生於中高過飽和) screw dislocation (巨分子晶體:發生於低過飽和) 3D-nucleation (只在巨分子晶體中發現) Layer growth 微觀 Normal growth 微觀
無論是巨分子或是一般小分子晶體,當驅動力(例:過飽和度)小時,分子傾向以減少表 面能的方式附著在step 上 kink,step 的成長形成成整層 terrace/layer,表面相較於 normal growth 顯得相當平滑。
而一層 terrace/layer 可填充的位置若結束時,必須產生新的 step,其上有可讓分子繼續加
入的位置才能讓晶體繼續成長,產生新的step 方式有兩種:一是以 screw 的方式;另一則是
2D-nucleation 在拓展成新的 layer。實際上由原子力顯微鏡(AFM)所拍攝的照片如圖 3-6 圖最 右所示。Step 上 kink 的位置以及 screw 上可提供的空缺如下圖 3-8 所示5 13
: (a)
(b)
在尚未利用原子力顯微鏡拍攝微觀形貌的時期,即有文獻以光學顯微鏡(OM)所測得之巨 觀成長速度推算蛋白質晶體之成長機制,以lysozyme 17
為例:低過飽和度時的成長機制為依 靠screw dislocation 產生新的 layer;中高過飽和度時則以 2-D nucleation 的方式成長。類似的 推算也運用於canavalin18
晶體。後來AFM 的引入為前人提供了實證上的圖像資訊,而此種低
濃度以screw dislocation,高濃度憑藉 2-D nucleation 的成長模式亦可延伸套用於多數其他生
物巨分子晶體上5
。而所謂的3-D nucleation 目前僅發現存在於少數生物巨分子晶體上,多為 已有層狀(layer)結構的粒子直接附著於大的晶體上,而吸附上已有層狀結構(layer)的粒子將引 導大晶體上新的 layer 的產生5 8
。
若驅動力大時,表面能容易被克服,晶體則以normal growth 的方式成長,不需要憑藉 step 或是kink。相較於 layer growth,這種有位置即可吸附的方式將使晶體表面顯得非常粗糙,成 長速度也較快。但此種成長方式在目前關於生物巨分子晶體成長的研究中仍相當少見。
{101} top side {110} a b c
3-2-2-3 成長機制 - 以 hen egg white lysozyme 為例
前文中曾提到,由於hen egg white lysozyme(HEWL)的取得和結晶容易,所以常用來做成長機 制推演的模型,由此種酵素所推衍出的成長機制也廣為適用於其他種類的生物巨分子。本小
節則以HEWL 為範例由巨觀和微觀兩方面說明從低濃度到高濃度其成長機制的變化。
過飽和度(C/Cs)
根據文獻21,以micro batch 方式要產生 HEWL 晶體之最低 C/Cs =1.25。
HEWL 晶體巨觀形狀
以熱力學觀點,由各個 miller index 平面表面能所繪出之γ-plot 可以知道:晶體最後的巨 觀形狀將以表面能最低的面決定:動力學上,生長速度較緩慢的面將佔較大的存在面積,而 生長較快速的面則會因逐漸消失而佔有較小比例的面積13 。 由文獻可知,HEWL 晶體的{101}及{110}擁有相對較小的表面能2 ,於是光學顯微鏡(OM) 可見之HEWL 巨觀形狀如下圖 3-10 示,左側圖示為從{110}之俯視圖,右側為從{101}面之俯 視圖,也就是左圖之側視圖。 圖 3-10 HEWL 晶體巨觀形狀。{101}、{110}具有較低之表面能,為最後穩定存在的面。左側圖式為從{110}之俯視圖,右側 為從{101}面之俯視圖,也就是左圖之側視圖。 a b c
由於{101}、{110}面受濃度影響程度不同,導致過飽和度變化時,晶體整體的巨觀形狀也有所 不同。下圖為文獻中依過飽和度不同時,對HEWL 晶體巨觀形狀變化所做的觀測17。 圖 3-11 {101}、{110}成長速度受過飽和度影響情形不同,濃度不同時,HEWL 巨觀形狀的變化。過飽和度低時,{101} 成長速率較快;過飽和度高時,{110}成長速度較快(ref 16)。約在過飽和度 = 4.3 時 {101}與{110}之成長速率相同。 已知生長速度較緩慢的面將佔較大的存在面積,而生長較快速的面則會因逐漸消失而佔 有較小比例的面積13 。由圖中的資訊可以得知當過飽和度低時,{101}成長速率較快,由光學 顯微鏡(OM)所觀察得到的晶體形狀,成長速率慢的{110}佔了大部份,而成長速率較快的{101} 則佔有較小比例;過飽和度高時,{110}成長速度較快,成長速率慢的{101}佔有比例提高, 而成長速率較快的{110}則變小。約在過飽和度,σ = 4.3 時 {101}與{110}之成長速率相同。 HEWL 晶體表面微觀形貌
在微觀形貌(morphology)方面:低過飽和度時,成長依賴 screw dislocation 產生新的位置 可供溶液中的HEWL 分子附著5 8
,在較高的過飽和度時,2-D nucleation 的方式取而代之成為 kink 的提供者。如下頁圖示:
[001] [001]
[-110]
[-110]
圖 3-12 左圖為低濃度時於HEWL 晶體之 AFM 照片,以 screw dislocation 的方式成長(ref8);中間及右邊則為較高過
飽和度時{110}面的 TEM (ref20)與 AFM (ref 21)照片,晶體以 2D-nucleation 的方式成長,且 2D-nuclei 有沿著[-110]方向延長 的現象(ref20)。
1. 圖 3-12 所示左圖為低濃度時於HEWL 晶體之 AFM 照片,以 screw dislocation 的方 式成長8
;
2. 中間及右邊則為較高過飽和度時{110}面的 TEM21與AFM 22照片,晶體以 2D-nucleation 的方式成長。
3. 2D-nuclei 的高度是晶格常數的倍數通常為一個晶格常數高度,
(此處之 HEWL 皆視為 tetragonal 形式。space group : P43212 ; lattice parameter:a=b=7.91nm , c=3.79nm 。
{101}上可約略測得一層 step 高度為沿 a、b 軸(垂直 43軸)之晶格間距=7.91nm;{110}上可約略測得一層 step 高度為沿 c 軸(沿 43軸)之晶格間距=3.79nm 21), 4. 但如中間圖示,有時可見重疊的雙層2D-nuclei,或是 layer 結構 5. 且值得注意的是{110}上之 2D-nuclei 和 layer 有沿著[-110]方向延長的現象21。如中 間圖和右圖所示。 為什麼{110}上之 2D-nuclei 和 layer 有沿著[-110]方向延長的現象?根據文獻推論20 ,依 據Black 等人解出 tetragonal 形式的 HEWL 晶體結構7
[001] [-110] {110} X Y Z 分別定義如下:
X:為靜電力構成之鍵結,一個分子的 Lys 13 和另一個分子終端之 carboxyl group 產生
鍵結。分子間彼此以2-fold rotation axis 對應。
Y:第一個分子的 Asn39 and Gln41 與另一個分子之 Asp66, Gly67 以及
Cys80 形成氫鍵 。分子間彼此以 2-fold 對應。
Z:Arg114 被同一個分子中之 Phe 34 與另一分子中之 Tyr23 包夾。此種鍵
結因附近存在氫鍵而更穩定。 兩分子間以 4-fold screw axis4 對應。 下圖為參考ref20 所繪之 HEWL 於{110}之分子鍵結情況:
圖 3-13 左圖為參考ref20 所繪之 HEWL 於{110}之分子鍵結情況。右圖為對應的平面以及方向。
根據上圖之兩層分子結構表示,淺藍色為下層,深藍色為上層,鍵結如圖右側所示。在 {110}上若有往[-110]方向之 step,如圖中所示之 A-B-C-D;且沿[001]方向之 step 則為 a-b-c-d 由X,Y 鍵結交替形成之螺旋20
。
依Black 等人發表之結構,將是 X<Y;X<Z 20
。又依照前述圖 3-917
所表示{101}與{110}依過 飽和度不同生長速度受影響不同的結果,其人利用Monte Carlo Method 模擬,得到結果如下, 當鍵結能量: X<Y<Z 時,最能符合圖 3-12 中所顯示在{110}上之 step 有沿[-110]延伸,以及雙層之 2-D nuclei 以及 layer 結構20 。下圖為以蛋白質分子所表示之{110}鍵結情形,以不同 symmetry operation 對應之分子以不同顏色表示,圖中之 BCD 對應上圖 3-13 中之 BCD 分子21。 圖 3-14 蛋白質分子所表示之{110}鍵結情形,以不同 symmetry operation 對應之分子以不同顏色表示,圖中之 BCD 對應上圖3-11 中之 BCD 分子21。W 為較弱之鍵結,以 2-fold 對應兩分子。
另外值得注意的是,{110}上層與層交替之 dangling bond 是 X+Y 與 Z 交替出現:一層全 部都是Z,或是一層之中兩列 X 兩列 Y,如右圖 3-15 所示。但由 Monte Carlo Method 模擬結 果得知X<Y<Z,故可以預測{110}上之 dangling bond 為兩列 X 兩列 Y,較符合實際可能發生 之情形25。
Diffusion/Transport Kinetic 表面
溶液 結束以HEWL 為例的晶體成長方面研究,回到一般狀況下,下一節將繼續討論當其他物理參 數加入生物巨分子結晶系統時,對於晶體成長方面的影響。 3-2-2-3 導入物理參數的影響 反應決定步驟 前文提到,成長是溶液中的溶質分子加入已存在的核或是小晶體,使晶體體積擴展的過 程。其中溶液中之溶質加入到已存在的核或晶體上時,大致可分為兩步驟,如下圖 3-16 示: 1. Transport : 由溶液至接近晶體表面的區域,主要與擴散有關。 2. Kinetic : 由接近晶體表面的區域加入到晶體上,調整適當的方位,成為晶體的一部 份。主要與表面之化學反應有關,即與溶質在晶體中和溶液的化學位能差有關。 已存在之核 或小晶體 圖 3-16 溶質分子加入已存在的晶體,成長過程可分兩步驟:1. transport 2.kinetic 。速率慢的為反應決定 步驟 成長過程中,速率慢的為反應決定步驟(control step)13 。
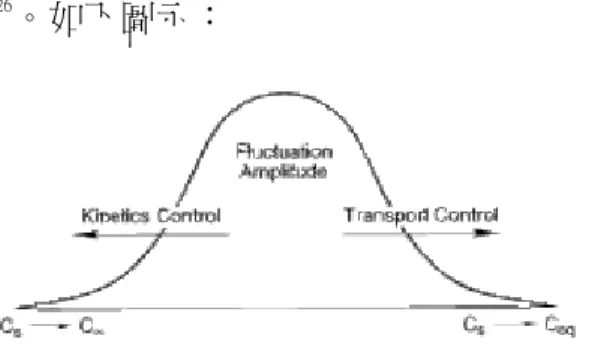
由文獻可知,當反應趨向於某一種單一步驟控制時(transport control 或是 kinetic control ), 能夠得到較好的結晶品質4 5 26
。如下圖示:
圖 3-17 溶液中之擴散步驟和靠近表面的反應,交互影響晶體成長的品質。當成長過程接近極右(完全是transport
/diffusion control)或是極左(完全 kinetic control)時,能夠達到最好的結晶品質。
當反應接近以下兩個狀態時,能夠得到品質較好的晶體:
1. 完全的kinetic control,接近晶體表面的溶質濃度幾乎等於遠處溶液中的溶質濃度,意 即擴散速度非常快,或是,
2. 完全的 transport/diffusion control,接近晶體表面的溶質濃度幾乎等於反應平衡濃度, 意即表面反應速度快,一有溶質擴散過來即可加入晶體到達正確的方位和位置 換言之,若反應愈偏向於中間transport / diffusion 和 kinetic mix control 時,得到晶體品
質越差26 。 改變物理參數對反應決定步驟的影響 由希望得到較好品質的晶體,而將反應推向某單一的決定步驟來看,物理參數的改變, 對此造成顯著的影響。 例如,在養晶系統外部加上幫浦,強制循環濃度固定的溶液17 ,即是將整個結晶矽統推 向kinetic control 的方向。另外,近年來相當熱門的研究方向:將整個養晶系統送上太空站。
少重力而減少溶液中擴散的現象26 29
,因此把整個系統的反應推向transport / diffusion control 的一端。
但是由實驗結果顯示,相較於在一般重力狀態下,不同種類的生物巨分子晶體在外太空 進行養晶實驗所得到的晶體品質優劣未趨向於一定。根據文獻推論,要解釋這樣的情形,首 先必須計算在一般狀態下,某特定晶體是偏向由kinetic control 或是 transport / diffusion control 26 28
。
假若某生物巨分子 A 在一般重力,且無其他特殊物理參數介入的狀況下,其晶體成長反 應是transport / diffusion control,那麼,將此系統移至外太空進行實驗,將會得到更佳品質的 晶體,因為外太空無重力系統使整個反應更推向 transport / diffusion control 的一端。
反之若某生物巨分子 B 在一般重力,且無其他特殊物理參數介入的狀況下,其晶體成長 反應是kinetic control,那麼,將此系統移至外太空進行實驗,則會得到相較於地球上品質為 差的晶體 !! 因為外太空無重力系統將使整個反應更推向 transport / diffusion control,對於 B 系統而言,就是把整個反應推向圖 3-17 中所示之mix control 的部份,反而使晶體品質變差。 其他物理參數的導入 除了上一節所提到的:強制溶液循環,或是減少重力影響之外,尚有許多針對改變物理 參數所做的研究,例如:控制溫度,以達到小區域的局部過飽和18 ,觀察蛋白質的成核成長; 或是研究磁場對於生物巨分子結晶所造成的影響,此項研究也促成了核磁共振(NMR)技術的 進步5 ,發展出可在液態中量測蛋白質結構的方法。 在本節的最後將討論的物理參數是電場的加入。雖然 80 年代已有學者提出,利用電場穩
定提供小於解離能的能量,得到品質較佳之蛋白質晶體的想法31 ;也有文獻以靜電力的觀點討 論生物巨分子間的作用力32 。但一直到晚近才有研究團隊開始真正著手實驗電場對生物巨分子 結晶的影響。下一節本文將針對目前電場影響蛋白質結晶系統的結果做一介紹。 電場對蛋白質結晶系統的影響 由於蛋白質在不同pH 值時,residue 解離程度不同,會帶有不同的電性。當 pH=pI 正負 總帶電量相消,不帶電。pH<pI 時蛋白質分子總帶電量為正,反之則為負。 1999 年,M. Taleb 的團隊始發表第一篇關於電場對蛋白質結晶的研究成果33 。並以下圖的 裝置進行外加電場的蛋白質結晶實驗:
圖 3-18 所示為兩片相隔 5 毫米(mm)之平行金屬版,以 micro batch 的方式混合 pH=4.5,10ul 的 HEWL 溶液,且與金屬 版間隔有空氣。施與系統 1000 伏特(V)的直流電壓
圖 3-18 所示為兩片相隔 5 毫米(mm)之平行金屬版,以 micro batch 的方式於 pH=4.5 混合 10ul 的 HEWL 溶液,且與金屬版間隔有空氣。施予系統 1000 伏特(V),的直流電壓,意即造
5mm
λ L x
e
− 上述文獻所得之重要結論可分下列兩點:1. 由於 HEWL 的 pI~10.5,故 pH=4.5 時,HEWL 分子上約帶有 10~12 個正
電,造成 HEWL 晶體陰極析出。以下為有外加電場與一般狀況下的光學顯微鏡(OM) 照片比較: 圖 3-19 左圖為無外加電場的正常狀況,HEWL 晶體散亂分佈於溶液中;右圖為有外加電場的系統,帶正電的 HEWL 晶體於陰極析出。 圖 3-19 左圖為無外加電場的正常狀況,HEWL 晶體散亂分佈於溶液中;右圖為有外 加電場的系統,帶正電的 HEWL 晶體於陰極析出。 2. 另一個重要結論則是:上述文獻由含有自由離子的介電材料遵守的 Poission-Boltzmann 方程式 ( kT fix πρ φ ελ φ ε ) sinh( ) 4 ( ∇ − 2 =− ∇ ),計算出因為電雙層的屏蔽效應,電 位能在水溶液中幾乎完全集中在靠近水溶液的邊緣,急速以 的形式衰退。以 一維的觀點,水溶液中央為圓點,其中 x 為水溶液中之任意位置,L 為水溶液和空 氣接處的邊界,λ為 HEWL 分子之德拜長度(Debye length )。
兩年後,同一組團隊又發表另一實驗結果:在外加電場下,pH=4.5,HEWL 尚未成核前, 帶正電的 HEWL 分子將在陰極聚集,使得陰極之蛋白質濃度高於陽極,此實驗也說明了前述
3-2-2-4關於本文實驗
本實驗是利用黃光微影製程,於矽晶圓上設計小尺寸的溶液儲存槽,以及各式金屬電極, 觀察在電場作用下,HEWL 晶體巨觀形狀以及微觀形貌的變化,以及不同電力線方向分布下, 對於晶體成長方向的影響。
第四章 實驗設計及流程
4-1 結晶
結晶部分的實驗採用前述 micro batch 結晶法。分外加電場的實驗組以及不加電場之對照 組,結果分別以光學顯微鏡觀察。實驗使用之溶液、裝置以及流程如下文敘述。實驗全程於 室溫下進行。 溶液 A. 0.1M PH=4.6 醋酸鈉水溶液 B. 10% 氯化鈉溶於 0.1M PH=4.6 之醋酸鈉水溶液中 C. 8% 及 20% HEWL 溶於 0.1M PH=4.6 之醋酸鈉水溶液中 裝置 為觀察電場對於HEWL 晶體成長的影響,本實驗利用微影技術設計一可施加電場並可承載液 體之裝置於矽晶圓上。並設計各類形狀的電極以觀察在不同電場分佈下,HEWL 晶體成長的 變化情形。實驗裝置及流程如下頁圖示。實驗流程 矽晶圓 以乾氧方式成長約 3000Å 之氧化矽薄膜 利用微影技術在氧化矽薄膜上定義溶液承載器皿 的位置及大小(1.5mm 平方)。並利用乾式蝕刻法 完成此蝕刻部分。 再次利用微影技術以及乾式蝕刻法定義金屬電極 的位置 利用電子槍鍍覆不同形狀之鈦-金電極。且金屬電極與矽晶 圓、與先前定義之溶液承載器皿之間皆留有氧化矽薄膜作為 絕緣之用。 利用濕式蝕刻法完成一可施加電場並可承載液體之裝置的製 作 採用micro batch 結晶法。混合適量之醋酸鈉以及氯化鈉水溶 Ti-Au 電極 Si-溶液槽 氧化矽上設計16 組不同形狀的電極,用來測驗不同電力線分布時,對晶體成長的影響 Top view
將蓋玻片切成小塊並以丙酮清洗,蓋住溶液。 用礦物油封住玻璃片,避免水蒸氣揮發。 (不外加電場的對照組,結晶部分實驗到此告一段落,等待結 晶。) 施加直流電場,等待結晶。 圖4-1 實驗流程
4-2 原子力顯微鏡(AFM)檢測
目前雖有人提出生物巨分子成核機制與傳統結晶學之成核理論有所差異15,但是因為此 理論尚未獲得廣泛的證實,在本文中不列為討論範圍。故不論一般傳統無機物或是生物巨分 子的結晶過程,大致皆可分為成核和成長兩部分。成核部分難以由影像直接觀察,目前多以 小角度 X-ray 散射8 或是 Light scattering5 8 9 15 作為檢測方法。成長部分由於可在晶體表面可預測 的位置進行觀察,原子力顯微鏡(AFM)在近年來已逐漸成為直接觀察生物巨分子成長及相關 現象的有力工具8 。關於原子力顯微鏡(AFM)在生物巨分子晶體檢測方面的文獻已於前一章節 介紹,本小節將著重於本實驗中原子力顯微鏡(AFM)使用時有別於文獻的樣品準備方式以及 操作。 Top view樣品準備及數據收集 1. 加入礦物油於上述裝置中覆蓋第一部分取得的已存在結晶之溶液 雖然如 HEWL 一般的標準品已可由商業管道取得,但是一般蛋白質的取得仍須自經 過自行純化的步驟,實屬得來不易。另外由文獻資料可以得知利用原子力顯微鏡作為檢測 工具時,樣品準備多依據以下兩種方法 : 其一,先長出晶種在移至蓋玻片上以適當濃度 的蛋白質溶液潤濕22 23 24 ,或是直接於蓋玻片上直接成長晶體,然後挑選一方位適當的晶體 作為檢測之用8 25 ( 各約 50uL )23 。為節省蛋白質溶液的使用量,本實驗僅使用第一部分結 晶所需的溶液量(約 0.4uL),實驗至結晶出現之後即利用礦物油(密度~0.8g/㎝3 )覆蓋於溶液 上,避免溶液在檢測時蒸發。由於 AFM 檢測可在液態中進行,不若掃描式顯微鏡(SEM) 須經過脫水以及鍍金的處理21 。此舉不但可如文獻中所期待在不使晶體表面接觸到空氣, 盡量維持晶體自然的狀況下檢測8 22 23 24 25 ,相較於文獻中兩法(各約 50uL),皆減少了蛋白質 溶液的使用量。
樣品準備好之後即置於 AFM 所配備之 standard liquid cell 之下,利用氣壓吸附上述裝 置於 AFM 之壓電平台,配置如下圖 4-2 所示。
2. 原子力顯微鏡(AFM)之數據收集
本實驗收集數據時所使用的AFM 機型為 Digital Nanoscope Ⅲa ( Digital Instruments ) 。
難題 由於生物巨分子易碎和柔軟的特性,在利用 AFM 檢測時,常常遭遇一些實際操作上的困 難。 i. 第一個難處是因為檢測必須在液態中進行,首先要克服的就是如何將待測的 晶體固定於基材上,避免因為晶體漂浮在溶液之中造成滑動而無法檢測的情 形8 。 ii. 其二則為前述溶液暴露在空氣造成揮發的問題。 iii. 利用 AFM 探針掃描成像的過程中,最困難的部分則是礙於生物巨分子晶體的 過於柔軟以及對探針的敏感8 。甚至有可能因為探針的掃描而對晶體表面產生 物理性的傷害8 。 解決方案 根據文獻,晶體難以固定的問題可以依賴直接在基材,通常是蓋玻片,上直接成長巨分 子晶體來解決8 25 ;或是在檢測時使用彈性的碳纖維將晶體“鉗"(clamp)住8 。在實作經驗中, 我們也曾經嘗試直接將晶體成長於蓋玻片上,但是並未因此得到良好的固定效果,同時在水 中漂流的晶體嚴重干擾了 AFM 懸臂(cantilever)的移動。但嘗試將晶體成長於上述設計的裝置 中時,由於溶液的體積減少,成長出的晶體減少,而觀察所得全部的晶體都成長於矽晶圓表
面,而且附著良好,不至於因為探針掃描而移動位置,倒是適切地解決了晶體飄動的問題。 對於生物巨分子晶體的過於柔軟,不易檢測,且為了避免探針和晶體表面的其他交互作 用力,懸臂之彈性係數(spring constant)須小於 1N/m8 25
。根據文獻,AFM 的 tapping mode 雖然 可能造成獲得的影像較 contact mode 不清晰,但在某種程度上已經稍微克服了巨分子晶體過於 柔軟不易檢測的困難8 。但在本實驗實際操作過程中,這仍然是一個相當棘手的問題。即使在 以拍敲式掃瞄(tapping mode)的情況下,為了獲得較清晰的影像,我們必須施加一個較大的力 量於 AFM 探針,增加探針對待測物表面高低起伏的敏感度。但是過大的力量即會對生物巨分 子晶體造成物理性的傷害。得到清晰的影像實為不易,在這方面的實驗技術仍需要更加改進。
第五章 實驗結果
5-1 溶解度
本文實驗結果中過計算飽和度所依據的溶解度為:Pascal Retailleau 等人於 1997 年在 Biophysics Journal 上發表的結果。其於室溫,pH~4.6 條件下,依析出劑(此處為 NaCl)濃度不 同,所得之Hen Egg White Lysozyme (HEWL)溶解度如下圖 5-1,右表中即為原始數據35。
圖5-1 中黑色實線為依據文獻中數據所繪之曲線,紅色虛線部份為利用 4 次多項式求得之最 接近曲線。右表中為論文中之原始數據1。 圖5-1 中黑色實線為依據文獻中數據所繪之曲線,紅色虛線部份為利 用4 次多項式求得之最接近曲線。與黑色實線部份已相當接近。下文實驗結果中計算過飽和 度的溶解度依據即為此紅色虛線。(本文所定義之過飽和度為:C/Cs,其中 C 為溶液濃渡; Cs 為此條件下之溶解度) NaCl (%) Solubility (mg/ml) 0 366 0.29 250 0.584 180 0.87 118 1.16 67 1.45 44 1.74 26.4 2.32 19.2 3.48 8 4.64 3.1 6.96 1.5 0 1 2 3 4 0 50 100 150 200 250 300 350 400
Solubility from Pascal Retailleau 4th-order Polynomial Fit
[HE W L ] mg /ml [NaCl] Weight%
5-2 由光學顯微鏡(OM)取得之照片
5-2-1 一般狀態下所得之晶體巨觀形狀
下圖5-2 為一般無外加電場狀況下,OM 所攝得 Hen Egg White Lysozyme (HEWL)晶體的照片。
(a) (b)
圖5-2 為一般無外加電場狀況下,OM 所攝得 Hen Egg White Lysozyme (HEWL)晶體的照片,均可見異質成核的晶體貼近
Si-溶液槽之邊緣成長。(a)C/Cs = 3.5,(b) C/Cs = 5.03 圖(a)為過飽和度 3.5(C/Cs = 3.5),圖(b) C/Cs = 3.5,均為無外加電場下的照片。晶體接近 透明,如圖中所圈起的部份。由於晶體呈現透明,故可見到由下方於TMAH 濕蝕刻不均勻所 造成的塊狀微突物。由 OM 照片可以得知一般無外加電場狀況下,所得之HEWL 晶體巨觀形 狀有下列特徵: 1. 未飽和(C/Cs<1)時,無結晶發生。 2. 圖中可見到如文獻中HEWL 晶體的{101}面及{110}面,如圖中卡通圖所示。 3. 圖中可見,HEWL 在 Si-溶液槽中並無特定分布或秩序,但由於異質成核 (heterogeneous nucleation)所需要越過的能障較低,所以可以見到可能由異質成核成 {110} {101}
5-2-2 外加電場下所得之晶體巨觀形狀
下表5-1 為外加電場狀況下,OM 所攝得 Hen Egg White Lysozyme (HEWL)晶體。
電極圖案 、陰陽極位置
+ -
+ -
+
-+ -
+ -
+ -
+ -
+ -
+
-靠近陰極之 電場模擬(上 圖圓圈部分) 結晶條件 C/Cs= 0.15 35V 5 小時 C/Cs= 0.37 35V 5 小時 C/Cs=5.02 100V 3 小時 OM 照片 陰極附近之 OM 照片: 1. 結晶多 在陰極析出 2. 形狀與 無電場情形 不同 3. [-110]沿 電力線方向 排列 有電場之{110} ---比較---無電場之{110} 陽極附近之 結晶情形 無結晶 無結晶 有結晶但數目較少, 且排列較不規則 [ 110] [001] [-110] [001]由上表5-1 的結果可以將外加電場下,由 OM 所得之 HEWL 晶體成長在巨觀上(形狀、分布、 排列)的特徵,定性陳述如下: 1. 不論溶液整體是否已達飽和(C/Cs>1 以及 C/Cs<1),均有晶體出現。唯當未飽和時,晶體 僅存在於陰極附近;過飽和時,溶液內部均有晶體出現,但仍屬靠近陰極的地方為多。 2. 晶體巨觀形狀從{110}觀之,相較於無電場的情形,有菱形晶體,或多出一未知斜面(下圖 5-3 中 左圖之灰色部份),與已知之{101}在{110}上夾角 160°,將於討論中詳述。 θ 圖5-3 從{110}觀之,有無外加電場時,HEWL 晶體巨觀形狀的比較。有外加電場較無電場時多產生一未知斜面,為圖示之 灰色部份。此現象將於討論章節分析。 3. HEWL 晶體之[-110]方向有沿電力線方向排列的趨勢。 5-2-3 電流導通時所得之晶體巨觀形狀 Æ 意外的發現 施加電流本來並非計畫中的實驗。但在施予外加電場實驗過程中,偶然發現若電流導通 (1~2 秒即切斷電流供應),但施加之能量不至於使蛋白質變質的情形下,所得之 HEWL 晶體, 巨觀上的形狀,與施加電場組別所得之晶體,有某些相似性: 1. 不論溶液整體是否已達飽和(C/Cs>1 以及 C/Cs<1),均有晶體出現。 [ 110] [001] 有電場時之{110} [-110] [001] 無電場時之{110} 多出的未知斜面; θ=160°
色部份)。下圖 5-4 即為一例: 圖5-4 左圖為施加電流 1 秒,靜置約 30 分後的 HEWL 晶體 OM 照片。與上表所示外加電場之晶體同樣擁有無 電場時所沒有的斜面。與已知之{101}夾 160°。將於討論中詳述。中間圖為參照,圖中灰色部份為左圖中所見之 {110}。右圖為一般條件下所得之晶體形狀。 施加電流與施加電場的組別所得到的晶體相異之處則為: 1. 電場下 HEWL 晶體生長時所具有之方向性和分布,在電流導通的狀況下全無。包括: i. 晶體無沿電力線方向排列的趨勢, ii. 以及C/Cs<1 時,OM 可觀察得到的晶體散布於溶液各處。 2. 且晶體產生的速度較施加電場時更為快速。 3. 但移除電流供應之後,晶體繼續成長的速率,巨觀上而言(由 OM 量測晶體大小變化), 與無電場時之成長速率相同。 4. 另一點有趣的則是,由施加電流而快速產生的晶體,經過一段長時間(~12 小時)靜置後, 有些晶體會逐漸消失。 電流和電場影響 HEWL 晶體成長系統之推論與假設 由上述電流與電場對於HEWL 晶體成長系統影響相異之處的 3、4 兩點,本文推測:電 流的能量對於HEWL 晶體成長系統,提供了短時間且動態的變化, (101) (?) 160° (110) (110) (101) 一般條件
導致系統在電流剛移除的時間內仍處於不平衡的狀態。但經過長時間的靜置,系統趨向平衡 態,原先存在的晶體才會消失。 但在本文中暫不討論基於何種原因或參數,導致部份晶體存在,部份晶體消失的結果,以及 存在或消失的選擇性。而是借力於:在電流影響下晶體成長變化的資訊,可以在短時間內取 得,這項優點。又由於所得之晶體巨觀上與施加電場所得之晶體相同,故將電流影響下晶體 巨觀形狀變化的過程與機制,引申為與電場影響下的方式相同。下一章將討論:在電場影響 下,晶體成長之動態變化過程,其推論即是建立於:電場與電流影響下之晶體成長巨觀形狀 上的變化過程是相同的假設之上。 電流影響下, HEWL 晶體動態成長過程 下圖為電流影響下,時間連續之 HEWL 晶體成長過程,參照圖下文字說明: 04 分鐘 06 分鐘 08 分鐘 10 分鐘 圖5-5 本圖所示為導通電流 1 秒即移除電流,錄下晶體成長過程的影像。各圖 下所示為錄影時間。可得到的訊息有: 1. 晶體形狀與前圖-3 所示,電場作用下所得相同。 2. 從 4:00 到 10:00,晶體變大,未知平面所佔比例逐漸下降。 3. 12 小時以後,部分晶體已完全消失,存在者也相對變薄。 表示此一快速結晶現象為不穩定的動態過程。
5-3 由原子力顯微鏡(AFM)取得之數據
以下結果中所說的晶格常數或是晶體結構皆以HEWL 最常見之 Tetragonal 結構為討論對象。 (a=b=7.91nm , c=3.79nm , P43212) 5-3-1 一般狀態下所得之晶體微觀形貌 下圖為C/Cs=2.3 時,於{110}面上得到之 AFM 圖像。 圖 5-6 左圖為C/Cs=2.3,{110}面上得到之 AFM 圖像。2D nuclei、step 平行[-110]方向排列;高度約 0.4 奈米(nm), 約是平行C軸之晶格間距。上述現象與文獻上記載者相同21 。右圖為參照,圖中{110}面上之條紋表示 step 沿[-110] 方向排列 如圖 5-6 所示: 1. {110}面上的 2D nuclei 沿[-110]方向拉長,且 step 平行[-110]方向排列。 2. 一層step 與 2D nucleation 的高度約 0.4 奈米(nm),約是晶格中沿C軸(平行 43)之晶格常 數。 上述現象與文獻上記載者相同21 。 [001] [-110] Step 排列方向 無電場時之{110}形貌(morphology)5-3-2 外加電場下所得之晶體微觀形貌
下圖為C/Cs=0.15,施加 35 伏特(V),5 小時,於{110}面上得到之 AFM 圖像
圖5-7 C/Cs=0.15,35V,5 小時,於{110}面上得到之 AFM 圖像。圖中可見黃色圓圈中之 step 沿[-110]方向排 列;藍色圓圈中之 step 沿[001]方向排列。若此兩方向之 step 交會於同一 layer 上,則將融合為一半月形之 step,
如圖中粉紅色方框所示,放大於下圖 5-8 圖5-8 為上圖 5-7 中粉紅色框之局部放大。如圖 5-8 中白色箭號所示,仍可見到半月形 step 內部有平行[001]方 向之 step,外部則有平行[-110]方向之 step,如圖中黃色箭號所示。 由於不能保證試片在接受AFM 檢測時,放置的平台是完全水平的,所以通常在數據處理時, 會加入一步桿平(flatten)的動作,此動作的目的為假設存在一平面,利用斜面或是曲面方程式 桿平前原本應有的step 桿平後圖中所見step,淺色 為高起部分,深色凹下 桿平時會將圖示斜 面校成回平面 桿平前原本應有的step 桿平後圖中所見step [001] [-110]
數目較多,導致桿平的校正動作 step 本身高低(圖中所見顏色的深淺)受到影響。參照圖 5-7 中文字敘述。圖中可見黃色圓圈中之 step 沿[-110]方向排列;藍色圓圈中之 step 沿[001]方向 排列若此兩方向之step 交會於同一 layer 上,則將融合為一半月形之 step,如圖中粉紅色方框 所示,放大於圖 5-8。如圖 5-8 中白色箭號所示,仍可見到半月形 step 內部有平行[001]方向 之step,外部則有平行[-110]方向之 step,如圖中黃色箭號所示。 下圖 5-9 為另一外加電場下所得之 HEWL 結晶的 AFM 圖。圖中仍可同時見到沿[001]及 [-110]兩方向排列之 step。圖右所示為原本未桿平之前應有之 step。 圖 5-9 圖中白色箭號所示,可見到平行[001]方向之 step;圖中黃色箭號所示,則是平行[-110]方向之 step。圖右 卡通圖為外加電場下{110}面上所觀察得到之 step 方向。 由上面幾張圖片,外加電場下HEWL 晶體{110}面上之形貌特徵可歸納如下: 1. 平行[001]與[-110]方向排列之 step,同時在外加電場下 HEWL 晶體{110}面上共存。 2. 當平行此兩方向之 step 交會於同一層 layer,兩方向之 step 將融為一半月形之 step。 3. 一層 step 的高度約 0.4 奈米(nm),約是晶格中沿C軸(43軸)之晶格常數。 沿白色箭號方向所示, 平行[001]方向之 step 沿黃色箭號方向所示, 平行[-110]方向之 step [001] [-110] 外加電場下{110}面上所觀 察得到之 step 方向 Step 排列方向
5-4 小結
在本章最後,本文將實驗所得無外加電場,外加電場與電流所得之晶體,巨觀和微觀上的特 徵整理如下表 5-2: 無外加電場 ,一般條件 外加電場 外加電流 成核 1. C/Cs<1,無晶體出現 2. 除異質成核於Si-溶 液槽器壁外,晶體無 特殊分布狀況 1. C/Cs<1、C/Cs>1,均有 晶體出現 2. C/Cs<1 時,晶體於陰 極附近析出 3. C/Cs>1 時,溶液中均 有晶體出現,但陰極附 近數目較多 1. C/Cs<1、C/Cs>1,均有 晶體出現 2. 晶體無特殊分布狀況 OM 所得 晶體巨觀形 狀,由{110} 面觀之 方向性 無 [-110]沿電場方向排列 無 AFM 所得 晶體{110}表 面 微觀特徵 僅觀察得到沿[-110]方向排 列之step 觀察得到沿[-110]方向以及 [001]方向排列之 step 無檢測 表5-2 無外加電場,外加電場與電流所得之晶體,巨觀和微觀上的特徵 & & [001] [-110] [001] [-110]第六章 結果討論
本章討論就上一章實驗所得到的結果,並由理論上嘗試推測可能的機制。首先將由計算水溶 液中電位、電場的分布以及大小開始討論。6-1 電位及電場在水溶液中之分布情形
由文獻可知,兩介質間電流量密度連續。 ε1E1 =ε2E2 ………... eq(1) 其中ε為介電常數,E 為電場。 且知在有自由離子之介電材料(本實驗之的 HEWL 溶劑%)中,電位將自邊界以 的形式衰 退。 ………... eq(2) 其中,L 為:一維狀況下,此欲求電位分布之 HEWL 溶液中心設為原點時的邊界座標。(意即 中心為原點,此介電材料(溶液)總長 2L)。x 為此一維狀況下,溶液中欲知電位點的坐標;λ 為HEWL 分子的德拜長度(Debye length)。又知電場為電位的微分,E = dψ/ dx ……….………... eq(3) 今,假設HEWL 與絕緣用氧化層接處之電位為ψ,外加電位 V,氧化層厚度 d,由 eq(2)(3) 可得絕緣氧化層與溶液附近之eq(1)可表示成: ……….………….…... eq(4) 當外加35V 時,代入實際參數: V=35/2 V,
ε
oxide =3.9,ε
HEWL =3.5,d =5⋅10−3㎜,L=0.75㎜,λ=1⋅10−6㎜ λ L xe
−V
−
φ
d
ξ
1ξ
2⋅
x
φ e
x L− λ⎛⎜
⎝
⎞
⎠
⋅
d
d
得到氧化層和HEWL 溶液界面(x~L)附近的電位為(以 V 單位): 鄰近此界面HEWL 溶液中,電位衰退的趨勢以 eq(5)表示: ……….………….………….…... eq(5) 將描述電位衰退的方程式eq(5)在接近界面的 HEWL 溶液中作圖,即可得到鄰近界面部份, HEWL 溶液中電位的變化: 圖6-1 鄰近界面部份,HEWL 溶液中電位的變化。電位在 50~100 nm 的距離內衰退至接近零。 再將電位對距離微分,可得到鄰近界面部份,HEWL 溶液中電場的變化。 f y( ) φ e y L− λ
⎛⎜
⎝
⎞
⎠
⋅ := ξ1⋅λ V ξ1⋅λ exp (x− L) λ⎡⎢
⎣
⎤⎥
⎦
⋅ ξd⋅ 2 +⎡⎢
⎣
⎤⎥
⎦
⋅ = 3.899× 10− 3 0.749993 0.749994 0.749995 0.749996 0.749997 0.749998 0.749999 0.001 0.002 0.003 電位(V) - 距離(mm) 距離(mm) 電位 (V ) f y( ) y 0.749994 0.749996 0.749998 0 2000 h y( ) 電場 (V/mm) 電位 (V) 距離(mm)0.749993 0.749994 0.749995 0.749996 0.749997 0.749998 0.749999 0.002 0.004 0.006 0.008 0.01 電位(V) - 距離(mm) 距離(mm) 電位 (V ) f y( ) y 0.749994 0.749996 0.749998 0 5000 1 .104 h y( ) y 在外加100V 時,也得到類似的情形,電位和電場都在短距離內急速衰減,惟 100V 時的界面 電位比35V 時更高出許多( )。 施加100V 時,界面附近電位和電場的分布分別如下圖所示: (a) (b) 圖6-3 施加 100V 時,界面附近(a)電位和(b)電場的分布。與 35V 時相比,界面電位及電場都較大,但仍都在 50~100 nm 內衰減至接近零。 將上述計算之電位、電場橫跨整個Si-溶液槽的分布狀況整理如下圖所示: 圖6-4 電場橫跨整個 Si-溶液槽的分布狀況,縱軸亦可為電場強度(V/mm) 比較計算所得兩種外加電位下,電位和電場在溶液中大小及分布特徵如下: 1. 外加電位大時,界面電位越大。但無論外加電位為何,溶液中之電位、均集中於與絕緣層的交界, 且於 50~100 nm 的短距離內衰減至幾乎無電場電位效應。 ξ1⋅λ V ξ1⋅λ exp (x L− ) λ ⎡⎢ ⎣ ⎤⎥⎦⋅ ξd⋅ 2 + ⎡⎢ ⎣ ⎤⎥⎦ ⋅ =0.011 電場 (V/mm) 距離(mm)
v
(v)x
(mm) 0.75 -0 7+
-6-2 成核
本節將以化學位能差的觀點,推測以下現象的機制:實驗觀察中發現,在外加電場的狀 況下,即使未達飽和(C/Cs<1),也有 HEWL 晶體集中於陰極析出。 首先,將水溶液中之 HEWL 分子分為:靠近溶液與氧化層的邊界(50~100nm),直接受電 場影響;以及較遠離邊界者(>100nm),未直接受電場影響者,兩部分來討論。 6-2-1 電極附近溶液與氧化層的界面(50~100nm),直接受電場影響的分子 1.F
=
qE
因 pH~4.6 時,HEWL 分子表面帶有約 10~12 個正電。且在接近界面 50~100nm 的範圍內, 有強大的電場,使系統內存在一強作用力,將帶有正電荷之 HEWL 推往陰極。F
=
qE
為造 成部份 HEWL 分子往陰極聚集,且陽極附近的分子將遠離陽極的第一個原因。 2. 電場造成的非等向擴散 一般常態下討論的擴散行為,建立於一物質往各方向擴散所需越過的能障是相等的,即是一 物質往各方向的最鄰近位置跳躍機會相等(random walk)。如下圖 a 示: (a) (b) A X B A C 圖 6-5 (a) 若 A 點有一原子或分子,在一維空間上,往左跳或往右跳至最近位置所須越過的能障均為 Em,機會 相等。(b)由於電場的作用,對化學位能造成梯度。從 A 點跳到 B 點,跟 A 點跳到 C 點之能障和化學位能差均到 B 點,跟 A 點跳到 C 點之能障和化學位能差均不同, i. 就跳躍所需越過的能障(Em)而言,Emc < EmA
ii. 就驅動力(化學位能差:ΔE)而言,ΔEAC >ΔEAB。 以上兩點都有利從A 往 C 跳躍(/擴散),造成非等向性的擴散。 圖中 C 可視為因靠近陰極而較穩定(對於帶正電之 HEWL 分子能量較低)的位置。電場造成非 等向的擴散為 HEWL 分子往陰極聚極,且遠離陽極的第二個原因。 3.誘導偶極(induce dipole) 最後對於直接受電場影響的分子,仍有電場誘導偶極的考量。根據文獻,將帶有正負電 之蛋白質分子誘導產生偶極所需的能量為10-4 kT eV16。以外加 35V 時為例,假設將分子由電 位為零處,移至實驗給予系統中各電位所對應之位置得到的能量(以分子一個電子視之,即為 電位能分布),除以誘導偶極所需之能量,得到下圖: 圖 6-6 隨距離改變,縱軸為電場提供能誘導 HEWL 產生偶極的能力,以誘導偶極所需要的能量的倍數表示。在 接近界面的範圍,確能提供強大的力量誘導 HEWL 產生偶極。 0.749993 0.749994 0.749995 0.749996 0.749997 0.749998 0.749999 500 1000 1500 誘導偶極所需能量的倍數-距離(mm) 距離(mm) 誘導 偶極 所需 能量的 倍數 g y( ) y





![圖 3-12 左圖為低濃度時於 HEWL 晶體之 AFM 照片,以 screw dislocation 的方式成長(ref8);中間及右邊則為較高過 飽和度時{110}面的 TEM (ref20)與 AFM (ref 21)照片,晶體以 2D-nucleation 的方式成長,且 2D-nuclei 有沿著[-110]方向延長 的現象(ref20)。](https://thumb-ap.123doks.com/thumbv2/9libinfo/8753257.206314/30.892.150.764.108.271/照片方式成長中間及右邊則為較高過飽和度時體以方式成長有沿向延.webp)