行政院國家科學委員會專題研究計畫 成果報告
綠竹筍老化相關酵素 pheylalanine-ammoinalyase 基因及酵
素功能探討(2/2)
計畫類別: 整合型計畫 計畫編號: NSC92-2317-B-002-016- 執行期間: 92 年 08 月 01 日至 93 年 10 月 31 日 執行單位: 國立臺灣大學生化科技學系 計畫主持人: 李平篤 共同主持人: 楊健志 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 5 月 12 日
第一章 緒論
第一章 緒論
1.1 竹
竹 類 屬 於 單 子 葉 植 物 (monocotyledon) 禾 本 科 (Gramineae) 之 竹 亞 科 (Bambusoideae) 或獨立成為竹科 (Bambusaceae),共計有 62 屬,約 1250 種,主 要分布在熱帶及亞熱帶等溫暖地區,生長適應性強。亞洲37 屬 700 餘種,非洲 5 屬約 50 餘種,大洋洲 7 屬 20 餘種,北美洲只有一種。一般認為竹類源於亞洲。 亞洲之竹類與非洲及大洋洲之竹類間親緣關係較為密切,與美洲竹類較為疏遠。 報告指出中國有26 屬 300 種,其中 22 屬 200 種以上具有經濟價值,而台灣目前 生 長 約 有 79 種 (Chang, 1991) 。 主 要 是 蓬 萊 竹 屬 (Bambusa) 、 麻 竹 屬 (Dendrocalamus) 及孟宗竹屬 (Phyllostachys)。 自然界中,竹類通常以種子或地下根莖為其繁殖方法。由於竹類的花期不穩 定且具有結實率低及發芽不易等問題,繁殖以分株、竹稈扦插及種子播種為主, 但這些繁殖方法效率都不高、不便於運輸,在大量繁殖及造林上造成莫大的困難 (Prutpongse and Gavinlertvatana, 1992)。1.2 植物二級代謝
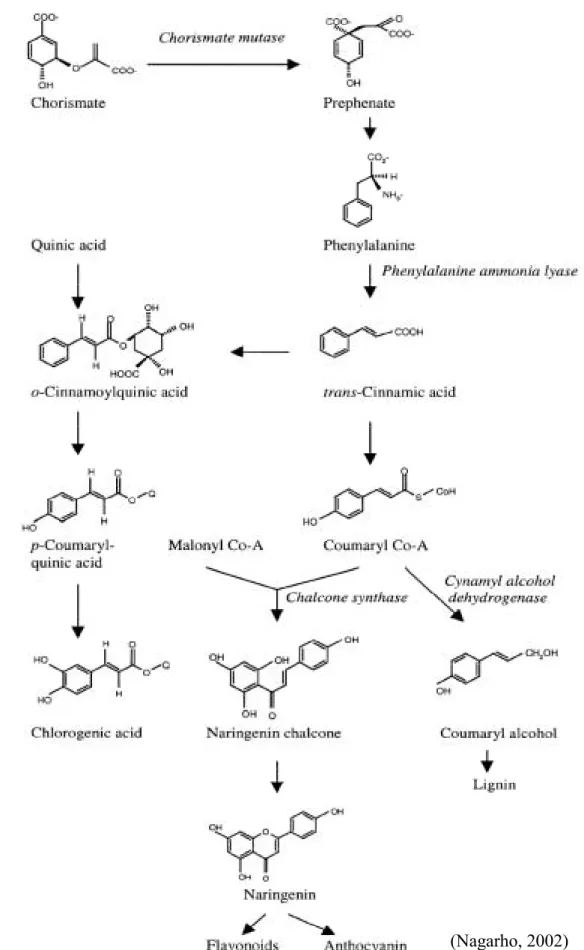
植物由 shikimate pathway 合成芳香族胺酸 (aromatic amino acids)。酚類化 合物 (phenolics) 是植物體內芳香環胺酸經過二級代謝 (secondary metabolism) 所生成之次級產物 (Figure 1.)。其中的 general phenyl- propanoid pathway 主要 由 三 個 酵 素所 組 成,包 括 phenylalanine ammonia-lyase (PAL, EC 4.3.1.5)、 cinnamic acid 4-hydroxylase (C4H, EC 1.14.13.11) 和 4-coumarate:CoA ligase (4CL, EC 6.2.1.12)。這是植物二級代謝中的 core-mechanism,之後所參與之反
應有不同的分支,可以衍生出許多的二級代謝產物,例如:flavonoid、anthocyanin
等,目前至少有 3,000 種以上的 flavonids 被發表。因此這三種酵素中只要任一
個受到干擾,就會影響整個二級代謝。第一個關鍵步驟是將 L-phenylalanine 藉
由 PAL 轉變成 trans-cinnamic acid。第二個步驟是將 trans-cinnamic acid 進行 hydroxylation 轉變成 4-coumaric acid;催化此反應的酵素為 cytochrome P450 monooxygenase,也就是 C4H,會受到光照、誘導物 (elicitor)、創傷等原因而誘 導 (Fahrendorf and Dixon, 1993; Bell- Lelong et al., 1997),而且誘導的機制與 PAL 相關 (Mizutani et al., 1997)。
第一章 緒論
Figure 1. Pathways for biosynthesis of phenylpropanoid productsderived from phenylalanine ammonia-lyase.
第一章 緒論
1.3 苯丙胺酸脫氨裂解酶
在 1961 年時,Koukol 和 Conn 最早從大麥苗中發現苯丙胺酸脫氨裂解酶
(phenylalanine ammonia-lyase, EC 4. 3. 1. 5,以下簡稱 PAL) 的存在,經過四十多
年的相關研究,PAL 成為二級代謝中最熱門的研究酵素之ㄧ。植物二級代謝是
被高度調控的,PAL 是 phenylpropanoids 生合成之第一個酵素,也是類黃酮
(flavonoids) 和木質素 (lignin) 與其他化合物生合成的關鍵酵素。在植物體中的
有機成分,大約有 30-45% 是衍生自 L-phenylalanine。PAL 的活性會被光照、
受傷 (wounding)、植物生長調節劑 (growth regulator) 和生物性或非生物性刺激
物 (elicitor) 所 誘 導 。 所 催 化 的 反 應 是 將 L-phenylalanine 經 非 氧 化 型 (non-oxidative) 脫氨反應 (deamination),產生 trans-cinnamic acid 和 ammonia。 目前發表的 PAL 大多是四元體 (tetramer) 形式存在,分子量約介於 250- 360 kD,具有一至兩個活性催化區 (catalytic site);植物中通常都不只有一個 PAL 存 在。換言之是 multigene family (Cramer et al., 1989; Fuka- sawa-Akada et al.,
1996)。植物 PAL 基因會因為微生物感染或是植物細胞直接處理 elicitor,會造
成 PAL 轉錄量增加 (Edward et al., 1985; Lowton and Lamb, 1987)。PAL 在植物
生長與發育扮演了決定性的角色,包括表皮組織對 UV-B 的抵抗,合成訊息代
謝物 (signal metabolites),和細胞壁中結構性大分子,維持細胞堅硬與保護的功
能。PAL 也是植物對抗病原菌入侵的因子之一,主要是 phenylalanine 的衍生
物,包括持續性或誘導性的抗菌素與殺菌素 (phytoalexin)、系統性防禦訊息
(systemic resistance signals) 與使細胞壁堅硬之聚合物。利用基因轉殖方法降低
PAL 活性,會使植物的形態與代謝受到影響,更會增加疾病的發生。
PAL 也存在部分真菌 (fungi) 以及酵母菌 (yeast) 中,在原核生物中只有放 線菌 Streptomyces、Basidiomycetes 和 Ascomycetes 被發表有 PAL 存在,真細
菌 (eubacteria)、古細菌 (archaea) 與動物細胞則是缺乏此酵素。植物中 PAL 所
扮演的角色已經相當清楚,但是在真菌中 PAL 所扮演的生理意義卻仍是未知。
這 些 真 菌 產 生 的 PAL 可 將 L-phenylalanine 代 謝 為 與 氧 結 合 的 衍 生 物 (oxygenated derivatives),例如:hydroxyphenylacetic acid、benzoic acid derivatives (Wat et. al., 1979) 或是完全降解該胺酸 (Moore et. al., 1979)。目前已經有證據顯
示,如果破壞 PAL 基因,在實驗室的培養環境下,對真菌的形態與生長不會有
明顯影響;在非植物材料中 PAL 的相關研究仍無法為 PAL 的功能下定義。
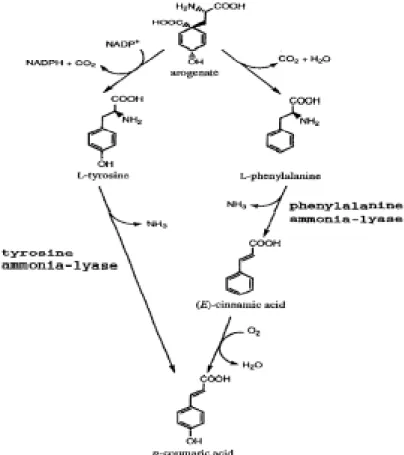
Tyrosine ammonia-lyase (TAL) 將L-tyrosine催化產生 trans-p-hydroxy- cinnamic acid (p-coumaric acid) ( Figure 2. )。Rosler 等人指出 (1997),在單子葉禾本科植
第一章 緒論 中該酵素大多只能利用 Phe 當作催化基質。許多研究指出 (Neish, 1961; Young et al., 1966;Haviy et al., 1971; Jangaard, 1974),單子葉植物利用 PAL 酵素具
有 TAL 活性,可以代謝 tyrosine;然而在雙子葉植物只對 Phe 有專一性。
Figure 2. Two alternative pathways of p-coumaric acid formation.
荷 蘭 芹 (parsley) 有 四 個 同 質 四 元 體 (homotetramer) 之 PAL 同 功 酶 (isoforms),對 Phe 的 Km 值範圍為 15~25 µM,對 Tyr 則為 2~8 mM (Appert et al., 1994)。以同功酶 PAL-1 而言,對 Phe 的 turnover number 為 22 s-1,但是
對 Tyr 則為 0.3 s-1。這樣的結果顯示,在生理狀況下 Tyr 極度不可能為該酵素
之基質。另外,從玉米中純化到的 PAL 對 Phe 的 Km 值為 270 µM,對 Tyr 的 Km 值為 29 µM (Havir et al., 1971),這樣的結果顯示玉米中的 PAL 具有 TAL 活性。參考荷蘭芹 PAL cDNAs之表現 (expression) 實驗 (Schulz et al., 1989; Appert et al., 1994);Whetten 與 Sederoff 在1995年指出:可利用同樣的實驗來
檢驗玉米之 cDNA 中,同一條胜肽鏈是否同時具有 PAL 與 TAL 活性。將玉
米中 PAL cDNA 在 E. coli 中表現,得到的結果可以證明 PAL 與 TAL 活性是
由同一條 polypeptide 所提供。玉米與一般單子葉植物可以利用 Phe 或 Tyr 來
產生 p-coumaric acid;目前這兩個代謝路徑在植物中之生理意義以及對於
phenylpropanoid pathway 的影響仍不清楚。另外還有許多因素會影響此兩種胺酸
在細胞質中的濃度變化,例如 arogenate dehydrase 和 arogenate dehydrogenase 活性及不同的表現量,在 chorismate 代謝成 Phe 或 Tyr 時之調控,及此兩種
第一章 緒論 胺酸運輸出質體 (plastid) 膜的不同形式。並且,Phe 和 Tyr 也會參與蛋白質的
生合成,同樣會影響細胞質內的濃度。所以只有 in vivo 實驗可以解釋此兩種路
徑對 p-coumaric acid 生合成所造成的影響。先前的實驗指出,在小麥及大麥葉
處理 PAL 和 TAL 的抑制劑 L-α-amino-β-phenylpropionic acid,會造成 Phe 和 Tyr 量 的 增 加 , 因 此 知 道 此 兩 種 芳 香 族 胺 酸 都 會 參 與 脫 氨 反 應 。 但 是 L-α-amino-β-phenylpropionic acid 同時也是 tyrosine decarboxylase 的抑制劑,仍
然存在另一個酵素被抑制,造成 Tyr 在植物體內累積的可能性。
1.4 轉殖菸草中 PAL 之相關研究
目前對二級代謝研究的重心擺在與代謝相關酵素基因轉錄的調控上,並且萃 取酵素分析活性在植物發育與環境影響下的變化 (Cramer et al., 1985; Lowton and Lamb, 1987; Hahbrock and Scheel, 1989)。這套系統必須要能在植物體內外操
作與分析 phenylpropanoids 化合物增加與減少的量,研究胞器隔間代謝關係 (metabolic compartmentalization),且要能對基因產物進行分子層次的研究。目前 菸草有很好的系統,已經證明在菸草中有一特定 PAL 會與菸草 microsomes 結 合。在 PAL 基因大量表現的菸草轉殖株中,PAL 活性的增加是不需要伴隨著 C4H 活性的增加 (Howels et al., 1999)。相對地,如果受到 elicitor 的誘導,C4H 和 PAL 的活性都會增加 (Howels et al., 1999)。 當植物 PAL 活性被壓抑會造成表現型不正常,並且較容易受病原菌的攻
擊,對於菸草鑲嵌病毒 (tobacco mosaic virus) 的感染沒有系統性防禦 (systemic acquired resistant) 機制 (Maher et al., 1994)。菸草中 PAL 被壓抑 (suppress) 的
現象是非常不穩定。將轉殖大豆 PAL2 基因之菸草,經自花授粉後可得到許多
不同表現型之子代。很有趣地,在菸草 PAL 壓抑的轉殖株,經自花授粉後的 T1
子代約有超過 60% 會轉變成 PAL 大量表現的性狀 (Howles et al., 1996)。PAL 基因靜默的植株也有可能穩定地遺傳好幾代或是表現型回復成類似野生型。在第 一 次 轉 殖 時 所 發 生 的 PAL 表 現 量 減 少 的 現 象 , 稱 之 為 epigenetic gene silencing,也就是 co- suppression 或 sense suppression (Flavell, 1994; Meyer,
1996)。PAL 活性在植物中會受到發育與廣泛環境因素所誘導。控制 PAL 表現 是非常複雜的,會在轉錄層次、轉譯層次、蛋白質穩定性,以及受到內生性與外 生行性因子影響,對該酵素去活化或活化 (Bolwell, 1992)。甲基化的現象已經證 明會在 PAL 轉殖時發生,但是對內生性 PAL 沒有影響,被認為是減少基因表 現的原因之一 (Dieguez et al., 1997)。然而目前仍然不清楚甲基化是否為基因靜 默的主因。植株矮化、葉片捲曲以及花型態的不正常,可能都是因為 phenyl-
propanoid pathway 受到影響。在轉殖的菸草中利用改變 PAL 基因的表現,來干
第一章 緒論 phenylpropanoid compounds。
1.5 苯丙胺酸裂解酶同功酶
PAL 為廣泛研究之二級代謝關鍵酵素,在 ExPasy (SwissPort) 資料庫中,
目前共有 129,768 個序列資料,其中有 52 個為 PAL 不同物種之序列,不同物
種 PAL 具有 isoforms 存在,核苷酸序列全長為 1,863~2,265 bp。換言之,蛋白
質胺酸序列介於 620~754 a.a.,推測之單元體分子量約為 67.8~82.6 kD。大多數
植物中 PAL 都有 isoforms 存在,在馬鈴薯中甚至含有 30 種以上。以菜豆
(french bean) 為例,在受到逆境時,至少有四種 PAL 基因產物被調控,這些被
誘導的蛋白質分子量都大約 77 kD。另外有一持續表現的 PAL,分子量約為 83
kD。目前對於 isoforms 和經誘導後的基因產物間之關係並不清楚。目前發現高
等植物中 PAL 基因大多由兩個 exons 和一個 intron 所組成。在真菌中的 PAL
則多由一整段 exon 所組成,並沒有 intron 存在。在 植 物 界 中 大 部 分 的 物 種 所 含 的 PAL 基 因 家 族 大 約 有 2 到 6 個 成 員 。
PAL 屬於多基因家族 (multigene family),阿拉伯芥中至少有四個 PAL 基
因,其中 PAL1、PAL2 的核苷酸序列與其他植物中的 PAL 基因序列相似度很
高。在基因表現方面,不同物種間對於創傷等不同的誘導因子,PAL 基因表現
在值與量方面有類似的結果。然而在第三種 PAL 基因 – PAL3,其核苷酸序列
與 PAL1、PAL2 和其他植物中的 PAL 核苷酸序列有明顯的差異;PAL3 有兩
個 introns,且所轉錄轉譯出的 PAL 蛋白質胺酸序列和其他的 PAL 蛋白質同源
性也不高,而且 PAL3 和其它 PAL promoter 比較,缺少某些保守性區域。
荷蘭芹之 PAL 的基因家族已經被完全發表,主要有四個 PAL 基因。此基
因家族可依照其相似度分成兩類;其中一類的組成為 PAL1 和 PAL2,它的
TATA-proximal promoter 與 coding region 非常相似;另外一類也有兩個成員,
PAL3 和 PAL4 彼此的相似度不高,且與第一類的 PAL1 與 PAL2 相似度低。
使用 PAL1/2、PAL3、PAL4 在 5,
untranslated regions 來設計專一性探針可以用
來 偵 測 荷 蘭 芹 葉 子 與 組 織 培 養 的 細 胞 在 經 過 UV 照 射 後 與 真 菌 感 染 之 後
mRNA 的變化量;甚至在受傷的葉或根中進行。
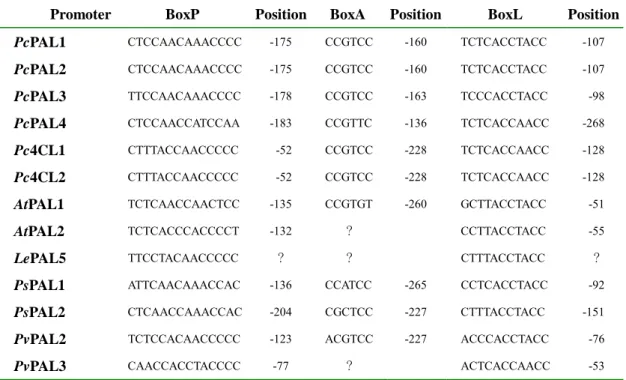
比對荷蘭芹四個 PAL 基因的 promoter 區域,有三個 boxes 是由 PAL1 gene promoter 經過 in vivo 和 in vitro 的實驗證明具有 DNA-protein interaction
功能,可能的功能為 cis-acting element,經過序列比對之後,可以推測所有荷蘭
芹的 PAL 基因 promoter 區域都含有這三個 boxes。分析結果參考 Table 2.。
在荷蘭芹的四個 PAL 基因 promoter 位置有相似序列,包括 TATA box 以及三 個 putative cis-acting element (boxes P, A, L)。利用報導基因來進行短暫表現分析
第一章 緒論 受光照或 elicitor 誘導的功能;類似報導也在 4CL 發現。荷蘭芹根部 PAL4 對 創傷有強烈的反應,但是在葉中則沒有;這樣的結果顯示:組織特異性與 PAL 基 因表現有關。類似的結果也在大豆 PAL2 和 PAL3 被觀察到。在荷蘭芹中,未 來的研究方向在細胞層次,包括 PAL 組織特異性、胞內定位等,將可以釐清 multiple PAL 基因的功能。
Table 2. Putative cis-acting elements and their positions on various PAL and 4CL gene promoters.
At, Arabidopsis thaliana; Le, Lycopersicon esculentum; Pc, Petroselinum crispum; Ps, Pisum sativum; Pv, Phaseolus vulgaris.
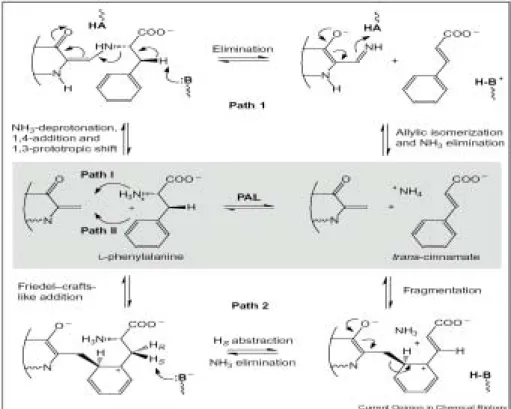
1.6 推測 PAL 可能催化機制
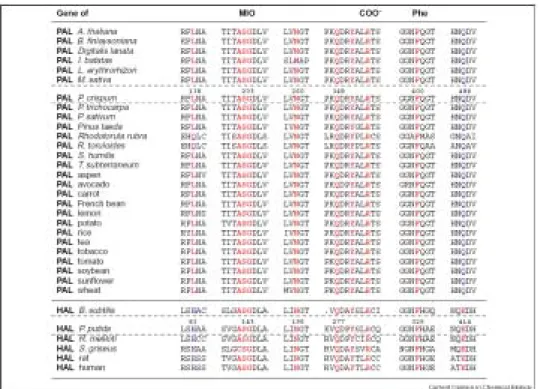
雖然目前沒有 PAL 的結晶學結構,但是在1999年 Pseudomonas putida 內 histidine ammonia-lyase (HAL) 已經有 X-ray 結晶結構,發現含有 3,5-dihydro- 5-methylidene-4H-imidazol-one (MIO) 環,主要由三個胺酸 Ala-Ser143-Gly 可以 形 成 自 體 催 化 形 式 (auto-catalytically form), 這 是 一 種 新 發 現 的 prosthetic group,並且也存在 PAL 中,以親電性反應 (electrophilic interaction) 來活化基 質。MIO 結構在1999年被發現,之後就立刻被用來討論 HAL (EC 4.3.1.3) 和 PAL 的反應機制。裂解酶一般會使用的 prosthetic group 包括 thiamine pyro- phosphate、iron-sulfur cluster、pyridoxal 5,
-phosphate、NAD(P)(H)、dihydroalanine (DHA) 和 MIO。在 P. putida 中 HAL 利用定點突變將 Ser143 換成 alanine, 會造成活性喪失,也確定了 Ser143 為 prosthetic group 之前驅物。同樣的實驗 也可以在荷蘭芹 PAL 中發現 Ser202 有相同的現象,由 Figure 3. 可以發現: 胺酸序列 Ala-Ser-Gly-Asp-Leu 為 HAL 和 PAL 高度保守的區域。Hanson 和
Promoter BoxP Position BoxA Position BoxL Position
PcPAL1 CTCCAACAAACCCC -175 CCGTCC -160 TCTCACCTACC -107 PcPAL2 CTCCAACAAACCCC -175 CCGTCC -160 TCTCACCTACC -107 PcPAL3 TTCCAACAAACCCC -178 CCGTCC -163 TCCCACCTACC -98 PcPAL4 CTCCAACCATCCAA -183 CCGTTC -136 TCTCACCAACC -268 Pc4CL1 Pc4CL2 CTTTACCAACCCCC CTTTACCAACCCCC -52 -52 CCGTCC CCGTCC -228 -228 TCTCACCAACC TCTCACCAACC -128 -128 AtPAL1 TCTCAACCAACTCC -135 CCGTGT -260 GCTTACCTACC -51 AtPAL2 TCTCACCCACCCCT -132 ? CCTTACCTACC -55 LePAL5 TTCCTACAACCCCC ? ? CTTTACCTACC ? PsPAL1 ATTCAACAAACCAC -136 CCATCC -265 CCTCACCTACC -92 PsPAL2 CTCAACCAAACCAC -204 CGCTCC -227 CTTTACCTACC -151 PvPAL2 TCTCCACAACCCCC -123 ACGTCC -227 ACCCACCTACC -76
第一章 緒論 Havir (1970) 假設了 PAL 的反應機制 (Path 1, Figure 4.),這個假設指出,親電 性催化會增加基質之胺基 (amino) 部分離開能力 (leaving ability)。雖然在親電基 加上胺基會促進離去基 (leaving group) 的特性,並且解釋產物釋放的順序 (NH3
會最後離開),可是沒有解釋 non-acidic β-proton 為了移除也被活化。雖然沒有 清楚的證據可以解釋這個假設的正確性,但是大多數之前的作用結果都以這個假 設來解釋。
Figure 3. Comparison of the amino acid sequences around the active sites of PAL and HAL of different origins.
而根據 Rétey 等人在1995年的文獻中指出 (Path 2, Figure 4.),基質帶有 多電子之芳香環,親電基會以類似 Fieldel-Crafts addition 方式來進行攻擊。陽離 子性中間產物在 β-hydrogens 非常鄰近的正電荷會增加其酸度。脫氨完成催化反 應之後適當的路易士鹼 (base) 會移除 β-proton。在這個機制下,基質在陽離子 性中間產物遺失芳香性 (aromaticity) 的能量需求是可以被補償的。有兩個可能 性:第一,形成 MIO 芳香環需要將 N 原子由 sp3 轉變成 sp2,因此控制了酵 素在催化循環中結構的改變。第二,為了要穩定陽離子性中間產物,MIO 中的
羰基 (carbonyl group) 會與一帶正電基團有作用,可能部份質子化 (protonation)。
以 [β-2H2]phenylalanine 當作基質並無法觀察到 kinetic isotope effect,解釋
為質子被帶走並非整個反應之速率決定步驟。以 [2H5]phenylalanine 和 [3H5] phenylalanine 當作基質,可以觀察到 kinetic isotope effect,解釋為基質之苯環要 先形成陽離子性中間產物。更進一步證明這個機制,是 4- hydroxy-phenylalanine (tyrosine) 為 PAL 很差之基質,而 3-hydroxyphenyl- alanine (m-tyrosine) 的作用
第一章 緒論 速率比 L-phenylalanine 還要快。
Figure 4. Predicted mechanisms for the PAL reaction. (Popple, 2001)
1.7 影響 PAL 活性之因素
1.7.1 光照
白光、紅光、遠紅光、藍光以及 UV 都會誘導 PAL 活性,白光是最早發
現可以誘導馬鈴薯塊莖 PAL 活性,最終產物是 chlorogenic acid。植物色素
(phytochrome) 會牽涉到 PAL 活性變化,紅光會誘導 PAL 活性,而遠紅光則會
阻止這個現象。荷蘭芹懸浮細胞之 flavonoid 生合成是需要依賴 UV 之 PAL 反
應,顯示轉錄或市後轉錄修飾會受到 UV receptor 的影響。小黃瓜幼苗會因為藍
光刺激而誘導 PAL 活性,藉由與抑制性蛋白質分離來活化 PAL,phenolic
compound/lipoprotein 複合體可能是這個抑制現象的主因。
1.7.2 創傷與感染
許多植物會因為創傷而誘導增加 PAL 活性,這樣的結果類似光照誘導的現
象。甘藷塊根經過切片後,會同時伴隨著 PAL 的合成與降解,乙烯也會牽涉到
整個反應的發生。當植物受到病原菌感染時也會誘導 PAL 反應。菸草鑲嵌病毒
會造成 PAL 重新合成 (de novo synthesis)。大多數研究集中在真菌或多醣類衍生
第一章 緒論 質素物質、植物殺菌素以及類黃酮醇衍生物。快速增加 PAL 活性通常是非常大 量但是短暫的,目前認為當受到真菌或 elicitor 刺激後,PAL 重新合成是一般的 通則。在高濃度刺激物存在下對 PAL 活性誘導是比較有效率的,顯示可能還有 其他後轉譯修飾現象存在。PAL 在十幾年前被認為是植物對病原菌防禦與逆境
調節的感應器 (sensor or receptor),一度受到大家極為重視。因為 PAL 在許多 情況下都會被活化,合成許多植物殺菌素等物質,參與抗病與逆境下的反應。可 是在近年的研究,已經發現許多 PR 蛋白質 (pathogen related protein),包括
chitinase、peroxidase 等等,漸漸地 PAL 在植物病原菌防禦上,研究就不是那
麼重要。目前植物 systemic acquire resistance (SAR) 是非常重要的研究領域,這
是屬於全株植物系統 性的抗病機制。SAR 是使用可移動訊息分 子 (mobile
molecule),當病原菌感染後,這些訊息分子會運送到其他植物組織,增加抗病能
力。目前最重要的兩種訊息分子是 salicylic acid (SA, 水楊酸,柳酸) 和 jasmonic
acid (JA, 茉莉酸)。SA 是 PAL 的下游產物,抗病植物中 SA 在沒有病原存在
下也有非常高的累積量。
另 外 在 植 物 中 過 氧 化 氫 (H2O2) 是 極 為 重 要 的 AOS (active oxygen species)。H2O2 本身當為訊息分子 (signal molecule),一般認為 H2O2 是造成防禦
基因活化的訊息 (Bolwell, 1999)。PAL 受 H2O2 所誘導是非常重要的,因為 PAL
是 SA 生合成的第一個酵素 (Dong, 1998)。藉由提高 H2O2 的量,來使得水楊
酸鹽作用,放大外來刺激的訊息。植物訊息傳導 (signal transduction) 在近十年 間,成為非常熱門的研究領域,其中 MAPK (mitogen activate protein kinase)
cascade 在植物防禦訊息上扮演了非常重要的角色。受到病原菌感染或逆境活化
此 cascade 後,經過一連串磷酸化與去磷酸化反應來放大訊號強度,活化下游
防禦基因,使得 HR (hypersensitive response) 細胞死亡以及系統性防禦機制啟 動,來達到防禦的最終目標。
1.7.3 植物生長調節劑
植物生長調節劑 (plant growth regulators) 對植物體內 PAL 活性影響機制
至今仍未明瞭。在組織培養環境下,減少內生性 auxin 與增加內生性 cytokinin
會造成 PAL 活性在數日內增加。暗培養之菜豆癒創組織 (callus) 在固體培養基
含有 2 mg/L 2,4-dichlorophenoxyacetic acid (2,4-D),繼代培養到含有 1 mg/L 1-naphthylacetic acid (NAA) 與 0.2 mg/L kinetin 培養基六天後,隨著發育,增加
PAL 活性;在光照培養之菜豆懸浮細胞也可以觀察到類似的反應。濃度 1 mg/L
NAA 處理兩天會增加 PAL 活性,kinetin 直到活性增加前 24 小時都是不需要
的;2 mg/L NAA 會對 PAL 誘導產生抑制效果,會因為 kinetin 濃度增加而被
克服,光照培養之菜豆懸浮細胞在低濃度 auxin 處理下會增加 PAL 比活性。這
第一章 緒論 光照培養之菜豆懸浮細胞加入適量轉錄及轉譯之抑制劑,來研究當細胞在增
加 cytokinin 及減少 auxin 濃度時之反應。發現 PAL 活性增加現象被抑制了,
顯示轉錄及轉譯都有牽涉到 PAL 活性。當 PAL 活性下降期,會被轉錄抑制劑
actinomycin D 所大量誘導,但是並不會受到轉譯抑制劑 propanemide 衍生物影
響,PAL-mRNA 是此反應之重要調節因子。荷蘭芹懸浮培養細胞在生長曲線之
stationary phase 用盡培養基中之氮源時,PAL 活性受光照誘導最為明顯。吉貝
素 (gibberellic acid, GA) 和離層酸 (abscisic acid, ABA) 也會對 PAL 活性有影 響。菜豆癒創組織在低 auxin 與高 cytokinin 濃度下會誘導 PAL 活性,此現象
會被 ABA 抑制,GA 會延遲此現象。豌豆與其他植物發現 GA 會促進 PAL 活
性與木質化現象 (liginfication) 的發生;菠菜懸浮細胞 GA3 會抑制 PAL 活性。
1.7.4 產物調控
許多實驗證明 cinnamate 與 hydroxycinnamate 衍生物會影響 PAL 之反
應。利用核苷酸與蛋白質抑制劑來研究 PAL 活性受光照與內生性 cinnamate 與
p-coumarate 之調控。光照似乎會刺激 PAL-mRNA 之轉錄,而後轉錄影響較為
快速,也許 cinnamate 會影響到 PAL-mRNA 之轉譯現象。切除豌豆表皮組織會
造成短暫 PAL 活性上升,內生性之中間代謝產物會快速減少酵素活性。研究指
出 cinnamate 會同時抑制酵素之重新合成與刺激移除已經存在之酵素。這種雙
重影響不能歸因於 cinnamate 之細胞毒性,因為 acid phosphatase 在實驗中並沒
有受到影響,cinnamate 可能是影響在後轉錄層次。
PAL 之生合成與被移除之速率會受到 cinnamate 的調節,在 in vivo 形況下
可以用 D-phenylalanine 與 L-α-amino-oxy-β-phenylpropanoid acid (L-AOPP) 之 影響來解釋。L-AOPP 為 PAL 之競爭型抑制劑,會抑制內生性 cinnamate 之生
成,破壞產物回饋作用,會限制酵素之合成與促進降解。當加入 L-AOPP 會造
成 PAL 重新合成,當經過 L-AOPP 處理後會促進 PAL 活性明顯上升且累積
phenylalanine。然而部分組織對內生性 L-AOPP 或 cinnamate 沒有反應,準確
測定 PAL 活性需要將 L-AOPP 與 PAL 確實地分離,才不會造成假象。在部份
植物如大豆或菜豆,L-AOPP 可能會與 PAL 緊密結合,即使經過透析也無法測
定準確之 PAL 活性。
1.8 磷酸化與後轉譯修飾
PAL 是病原菌防禦、逆境反應與二級代謝的關鍵酵素。原本以為二級代謝
酵 素 一 般 只 會 在 genernal phenylpropanoid pathway 以 及 代 謝 路 徑 分 支 位 置 (branching site) 進 行 調 控 。 近 年 來 發 現 PAL 會 受 到 後 轉 譯 磷 酸 化
(posttranslational phosphorylation) 作用的影響,這也是二級代謝中第一個發現有
第一章 緒論
磷酸化的可逆反應,是真核生物廣泛存在的調控機制。 為了強調這個現象的顯
著性,需要去確認負責進行磷酸化的蛋白質激酶 (protein kinase) 並且瞭解調控
的環節。AtCPK1 (CDPK, calcium-dependent protein kinase) 既可磷酸化重組的
PAL蛋白質又可以對在植物體內可被磷酸化的胜肽 (peptide, SRVAKTRTLTTA)
進行磷酸化反應 (Chang et al., 2001)。針對 PAL 重要的磷酸化現象,確認此專
一 CDPK 為 PAL 蛋白激酶,也開啟了鈣離子連結到生物性逆境的訊息傳導與
水 楊 酸 、 植 物 殺 菌 素 產 生 的 探 討 。 在 受 刺 激 的 細 胞 中 可 以 部 分 純 化 出 Ca2+-stimulated protein kinase,分子量約為 55 kD。
PAL kinase 是 ser/thr kinase,磷酸化位置已經被確認是 threonine,磷酸化 後會降低 Vmax 並且可能造成次單元體 (subunit) 的降解,PAL 的降解路徑會有
70、53、4 kD 的中間產物被觀察到。菜豆 PAL 蛋白質磷酸化位置應為 Thr545,
附近序列為 VAKRTLTT (539~546)。當菜豆受到刺激後,PAL mRNA 量會迅速 提 高 , 經 過 轉 譯 後 , 蛋 白 質 的 去 活 性 與 轉 錄 量 的 回 饋 調 控 , 可 能 是 因 為 phenylpropanoids 量來進行控制。CDPKs 為具有 calmodulin-like domain 之蛋白 激酶,在蛋白質 N-端為 kinase 催化區,C-端有 calmodulin-like domain,具有
四個 EF-hand 之 calcium 結合區,與鈣離子有高親和力,磷酸化之保守序列為 R/K-X-X-S/T。PAL 為植物二級代謝關鍵酵素,已知經過 elicitor 處理之菜豆懸 浮細胞會受到磷酸化調控 (Bolwell, 1992)。目前為止,在植物二級代謝中,此蛋 白激酶是第一個被證明藉由共價鍵的磷酸化作用來調控酵素活性,然而磷酸化可 能還具有其他功能。PAL 經磷酸化後,經由放射線標定追蹤,約有 20﹪PAL subunits 會在細胞膜上被發現。這也代表磷酸化與去磷酸化可能調控 PAL 往膜 上運輸。在不同的物種中 PAL 磷酸化位置有保守性,因此 PAL 磷酸化作用可 能是高等植物中廣泛存在的調控機制。
1.9 實驗緣起
綠竹筍 (Bambusa oldhamii Munro) 適宜種植在海拔 500 公尺以下砂質壤 土或壤質砂土之土壤,台灣各地均有零星栽培,在北部地區最大栽培地區為台北 縣五股鄉。綠竹筍是國內夏季生產的重要蔬菜之一,其生長一般為4月至9月, 而南部由於地理環境影響,會比北部早大約2週到1個月上市,而6月至8月為 颱風季節,綠竹筍因為生長於地下,根莖堅實,颱風災害較少;但是竹筍幼嫩的 生長組織,在採收後旺盛的生理變化,使其品質極易發生劣質,如褐化、纖維化 或木質化,嚴重影響竹筍的品質與價值,因此筍農會將剛採收的竹筍泡於水中、 冷藏或是以開水殺青的方式來降低劣質程度。Neish (1964) 就發現 PAL 與木質 素生合成有關,有研究發現 PAL 活性的上升與細胞壁木質化有關,且此酵素活 性在蘆筍嫩莖中以基部的含量最高,然後越往頂端部分酵素活性有遞減的現象。 除此之外,在儲藏期間蘆筍嫩莖中 PAL 活性會隨著時間而上升。儲存的時間和
第一章 緒論 溫度對木質化有密切的關係,且實驗證明將蘆筍放在水中可以抑制木質化現象。 綠竹多以地下根莖進行無性繁殖,繁殖以分株、竹稈扦插為主。竹之花期不 穩定,數十年才開一次花,且結實率低及種子發芽不易,開花後常伴隨著植株死 亡。竹為自我保護 (selfish) 之植物,以群落 (community) 方式進行繁殖,一般 竹林中不易有其他植物生長,可能原因為:竹子生長旺盛,對養分吸收能力強; 竹林一但茂密會遮蔽陽光,造成底下植物無法進行光合作用;另外竹會分泌
phenolics 類化合物等相剋物質 (allelopathic agent),對其他植物造成毒害。一般
生物選擇無性繁殖的最主要原因是躲避天敵。以植物而言,開花結果需要消耗許 多能量,而且易受到天敵攻擊,竹選擇以地下根莖繁殖的原因可能也是為了躲避 天敵,數十年才開花結果一次,使得天敵無法攝食。然而用進廢退,長久演化下 來選擇了無性繁殖,連帶地也將有性繁殖能力幾乎都喪失了。竹子生長旺盛,植
物荷爾蒙含量高,尤其是 cytokinins 的含量高,並有實驗證實竹筍含量豐富之
tRNA 具有 cytokinesis 活性,其活性當量(activity equivalent) 高達1.8 µg kinetin/ mg (Lee et al., 1970),也是雨後春筍的主因;而 cytokinin 也是植物延遲老化的 重要因素之一。中國人以松、竹、梅當作歲寒三友,另有松竹常青這個成語,代 表終年常綠,同時也代表竹之老化現象並不明顯。竹子一但開花常常會伴隨著全 植株死亡,目前仍不知其真正原因,但是可以推測的是植物荷爾蒙一定扮演重要 角色,當植物體內荷爾蒙比例改變,來當作老化的訊息,使全植株或整個群落同 時老化甚至死亡。 竹筍,由於澱粉含量低,因此質地脆硬,而久煮不爛;且炊熟後不油膩,除 因竹筍 Lipid 含量低之外,並散發出具有清香口感的天然物—Phenolics( 其中最 主要的是 p-Hydroxybenzoic acid )所致。台大農化系生化研究室蘇教授在七十年 代起就使用綠竹筍當作實驗材料,進行竹細胞壁構成分及竹筍與醣代謝相關酵素 之研究 (Su, 1965; Sung, et al., 1971),也曾經嘗試進行組織培養;先前研究發現, 在進行綠竹組織培養時,可以觀察到 callus 生長,但是無法長久保存 callus 與
進行懸浮細胞(suspension cell)培養,褐化 (browning) 或黑化的發生是實驗最大 之干擾因素;造成此褐化現象之主要原因為 phenolics 累積,phenolics 為木質 素之前驅物 (Su, unpublished data),為二級代謝衍生物。然而 PAL 為二級代謝
關鍵酵素,抑制 PAL 活性,可能可以減緩褐化程度。研究 PAL 活性變化與褐
化發生之關係,可能有助於克服懸浮細胞誘導時所遇到之困難。
為了防止採收後的綠竹筍生理代謝依舊旺盛且迅速,會造成快速劣變,包括
木質化、纖維化等,使其品質和價值受到影響。本實驗將綠竹分成竹筍 (bamboo
shoot)、竹葉 (bamboo leaf) 以及竹籜 (bamboo shell),來分別探討 PAL 的生化 性質以及對綠竹木質化的重要性。其中竹籜為介於竹筍與竹葉之部位,從未出土 到出土過程間,會開始受到光照影響,進行光合作用,伴隨著葉綠體的發育,因
第一章 緒論
此推測竹籜 PAL 之生化性質會介於竹筍與竹葉 PAL 之間。為 了 防 止 採 收 後
的 綠 竹 筍 因 快 速 劣 變 , 使其品質和價值受到影響 , 本 研 究 計 畫 , 將 分 離 並 純 化 出 綠 竹 筍 的 PAL 或 其 isoforms,來 研 究 PAL 的 生 化 學 特 性;以 及 PAL 對 綠 竹 筍 木 質 化 的 關 連 性 ; 並 以 純 化 的 酶 蛋 白 製 備 專 一 性 抗 體 , 同 以 測 定 未 採 收 及 採 收 後 , 不 同 部 位 和 不 同 儲 藏 條 件 , 及 不 同 儲 藏 時 間 與 溫 度 下 PAL 的 活 性。 此 外,擬 藉 由 本 實 驗 所 得 酶 蛋 白 資 料 , 比 較 不 同 高 等 植 物 已 發 表 的 pal cDNA 與 序 列 分 析 , 進 行 竹 筍 pal-cDNA 及 基 因 庫 的 建 構 與 選 殖 ; 研 究 竹 筍 pal 基 因 結 構 與 調 控 ; 利 用 Southern、Northern bloting 與 Western bloting 技 術 , 比 較 綠 竹 筍
中 pal gene 表 現;以 及 pal 基 因 表 現 調 節 機 制。除 進 行 綠 竹 PAL 之
蛋 白 生 化 學 , 及 其 基 因 之 分 子 生 物 學 之 基 礎 研 究 外 , 期 能 以 植 物 生 化 代 謝 的 觀 念 ,探 討 較 實 用 的 物 理 或 化 學 方 法 降 低 竹 筍 PAL 表 現 , 延 緩 竹 筍 因 PAL 所 導 致 之 褐 變 、 木 質 化(老 化), 而 使 品 質 劣 變 , 達 到 保 鮮 目 的 , 以 增 加 農 民 收 益 為 目 的 。
第二章 材料與方法
2.1 材料
綠竹 (Bambusa oldhamii):樣本採集自台北縣五股鄉與台北市文山區筍農之竹 林。採收後要清洗乾淨後,低溫保存運回實驗室,先以液態氮 (liquid nitrogen, N2) 處理以保持低溫。運回實驗室後,立刻處理竹筍,將竹筍剝殼,依照所需不 同的部位,分別儲存於 -20℃ 及 -80℃ 冰箱。葉子的樣本為了新鮮,一般都是 要現採現處理,無法在冰箱中長久的儲存。2.2 蛋白質定量法
將牛血清蛋白 (bovine serum albumin) 標準液 (200 µg/mL) 分別配成各種不同 濃度之牛血清白蛋白標準液。取 50 µL BSA 標準液或樣品液加入 ELISA 微量 滴定盤中。加入 200 µL 染料結合試劑,混合均勻後,在室溫下靜置反應 5 分 鐘,測 570 nm 吸光值。以不同濃度之牛血清白蛋白標準液及其吸光值,繪製用 來定量蛋白質的標準曲線,以定量樣品液中的總蛋白質量。 於微量管中,加入 50 µL 之適當濃度酵素液。加入 950 µL 混合後之基質液, 最終之 L-phenylalanine 濃度為 12.1 mM,加蓋後於 37℃ 下,反應 30 分鐘。 加入 100 µL 6 N HCl,終止反應。以分光光度計 (Hitachi U-1100) 用石英材質 之測光管測定 290 nm 吸光值變化。以 12.1 mM D-phenylalanine(已證明PAL對 2.3 苯丙胺酸脫氨裂解酶活性分析法
第一章 緒論 D-phe無反應) 作空白對照組 (blank)。
2.4. 電泳檢定系統
2.4.1 原態膠體電泳 (native-PAGE)
樣品與追蹤染料以 5:1 的比例混合,將10∼20 µL之樣品注入槽中。以 120 伏 特進行電泳,當追縱染料通過焦集膠體進入分離膠體時,將電壓改成 150 伏特, 追縱染料跑出膠體後,即可停止電泳。在4℃冷房中進行電泳以保持酵素活性。 電泳後進行膠體染色、活性染色及免疫轉印等實驗。2.4.2 SDS-膠體電泳 (SDS-PAGE)
電泳前樣品需加入同體積的 SDS 膠體電泳樣品緩衝液。以 100℃ 水浴加 熱 10 分鐘。鑄膠、電泳、染色、脫色等步驟同 2.4.1 原態膠體電泳,但電 泳緩衝液中需含 0.1% SDS。2.4.3 膠體染色法
2.4.3.1 CBR 蛋白質染色法
將膠片放入染缸中,倒入 CBR 染色液。在旋轉器上輕搖約 20~30 分鐘後, 倒出染色液 (含 10% 冰醋酸及 20% 甲醇)。以一次水洗過後,再加入脫色液進 行脫色,直至背景透明為止。2.4.3.2 醣蛋白質染色法 (過碘酸-硝酸銀法)
電泳後,膠片以固定液A(10% 醋酸及25%異丙醇的混合液體)固定過夜。隔日 改以固定液B(7.5% 醋酸溶液)浸泡30分鐘。倒去固定液 B,加入過碘酸溶液於 4℃反應1小時。以水洗 3-5 次。加入硝酸銀溶液,反應10分鐘。倒去硝酸銀溶 液,用水洗3∼5次,每次10分鐘。加入還原液乙(0.05% citric acid, 0.1%甲醛, 15% 甲醇),開始呈色,觀察 negative control (BSA) 的蛋白質是否呈色,在其未呈色前倒去還原液乙。用水洗過膠片後,馬上加入反應中止液(0.05% citric acid, 0.001% ethylenediamine)中止呈色。1 小時後,即可進行膠片乾燥。
2.5 苯丙胺酸裂解酶的純化
2.5.1 粗抽及硫酸銨分劃
取約50-100 g之樣品,在冷房中操作,用液氮研磨成細粉末。加入 200-400 mL 之萃取液,以四層紗布過濾,將濾液離心30分鐘。取上清液,於冰浴下緩慢加入0.5 %魚精蛋白硫酸鹽(protamine sulfate, Sigma, P-4380),攪拌均勻,持續攪拌平
第一章 緒論
和度,持續攪拌平衡30分鐘,離心30分鐘。取上清液,於冰浴下繼續加入硫酸銨
直到 60% 飽和度,攪拌平衡30分鐘,離心30分鐘。取沉澱,以少量緩衝液A(50
mM Tris – HCl, 10 mM MgCl2, 1 mM EDTA•2Na)溶之,對5000 mL緩衝液A透
析,至少換三次緩衝液A。隔日取透析液,離心30分鐘(Hitachi RPR20,10,000 rpm) 取上清液進行下一步純化。
2.5.2 膠體過濾管柱層析
在冷房中,將平衡後適量之Sephacryl S-200膠體與緩衝液A(50 mM Tris-HCl,pH
7.5;10 mM MgCl2,1 mM EDTA)在燒杯中混合均勻,將燒杯中的膠體攪拌均勻, 沿著管柱緩緩倒入膠體。以緩衝液A連續流洗(30 mL/hr),一般需要 2~5 倍膠體 體積平衡。取適量樣品,離心30分鐘去沉澱。然後關掉幫浦將輸送管從緩衝液A 中取出,插入樣品中,以30 mL/hr方式注入層析管柱中。樣品之溶離(elution): 當樣品注入完全後,關幫浦並將輸送管改接入1,000 mL 溶離緩衝液。溶離流速 約30 mL/hr,並以分劃收集器收集每管5.0 mL。當溶離完後以1.0 M NaCl 流洗, 將其餘蛋白流洗下來。當溶離完成後以ELISA reader偵測每一分劃的蛋白質濃度
及活性測定A290。收集具PAL活性部份,以限外過濾(Amicon, YM-10)濃縮,進行
下一步純化。
2.5.3 疏水性作用管柱層析(Phenyl-Sepharose)
Phenyl- Sepharose 膠體以緩衝液B連續流洗至pH值及離子強度不變為止。取前 一步適量樣品,離心30分鐘去沉澱。注入樣品保持正常流速(60 mL/hr),並進行 分劃收集(5 mL/tube)。注入樣品完畢後,以2~5倍管柱體積之緩衝液B,流洗至沒 有蛋白質流出後,行直線梯度溶離,高限緩衝液為緩衝液B (125 mL),低限緩衝 液為酵素緩衝液A(125 mL)。溶離速度約為60 mL/hr,並以分劃收集器每管收集5 mL。當鹽濃度梯度跑完之後,又補充酵素緩衝液A到mixer中,將其餘未溶離的 蛋白質沖下來。當溶離完成後以ELISA reader偵測每一分劃的蛋白質濃度,及活 性測定吸光值。收集具 PAL 活性部份,離心濃縮至 5~10 mL,置於-20℃冰箱 中保存,待下一步純化。2.5.4 陰離子交換層析
DEAE Sepharose 膠體同膠體過濾法。以緩衝液A連續流洗至 pH 值及離子 強度不變為止。取適量樣品,離心30分鐘去沉澱。然後關掉幫浦將輸送管從緩 衝液A中取出,插入樣品中,以30 mL/hr方式注入層析管柱中,小心不要有氣泡。 當樣品注入後,關幫浦並將輸送管改接入緩衝液A。溶離流速約為50 mL/hr,並 以分劃收集器收集每管5 mL行直線梯度溶離。梯度由0到0.5 M NaCl,當溶離完 後以含有1.0 M NaCl 緩衝液流洗,將其餘蛋白流洗下來。當溶離完成後以
第一章 緒論
ELISA reader 偵測每一分劃的蛋白質濃度及測定酵素活性變化。收集具PAL活
性部份,以限外過濾(Amicon, YM-10)濃縮,進行下一步純化。
2.5.5 快速蛋白質液相管柱(FPLC)層析
以Mono Q HR 5/5 (50 mM Tris-HCl,15 mM MgCl2,1 mM EDTA,pH 7.5) 的,
以緩衝液A流洗平衡2~3小時(流速 0.5 mL/min)。樣品先離心10分鐘 (Hitachi RPR20,10,000 rpm)去沉澱。取1~2 mL樣品注入樣品環 (sample loop)。執行程式, 以 0~100% 溶離緩衝液(50 mM Tris-HCl ,15 mM MgCl2,1 mM EDTA,pH 7.5, 1 M NaCl)梯度溶離。每個分劃管收集0.5 mL 溶離液,共收31管。當溶離完成後, 立刻取下放入冰中保存。以ELISAreader偵測每一分劃(0.5 mL)的蛋白質,及活性 測定290 nm 吸光值,收集具PAL活性部份,進行下一步分析。
2.6. 蛋白質胺酸定序分析
取部分樣本,進行native-PAGE或SDS-PAGE分析,在電泳上層緩衝液加入 2 mM thioglycolic acid,去除自由基。將膠片浸在轉印緩衝液中平衡30分鐘。將 轉印膜浸入100%甲醇中1-3秒。再以轉印緩衝液平衡 15 min。取出轉印膜,以 1/5 濃度的CBR 染液短暫染1 分鐘,或等到色帶出現時,即以50%甲醇脫去背 景值;若蛋白質濃度低,脫色時間須較久,若背景顏色太深,可用 100% 甲醇 脫色。染色脫色後,將轉印膜放入二次水中清洗,再放入烘箱中乾燥。將轉印膜 上的目標蛋白質色帶切下,以封口袋保存送定序。2.6.2 Q-TOF (ESI-MS-MS)
將欲檢視的蛋白質從已染色的聚丙烯醯胺膠體切下。利用解剖刀將膠體切成約 1 mm2 的顆粒,裝入 1.5 mL 的微量管內。加入 100 µL 的 25 mM ammonium bicarbonate/50﹪acetonitrile,以vortex振盪10分鐘。利用gel-loading pippet tips將溶液部分移除並且丟棄。重複清洗、脫水步驟三次。使用離心真空乾燥機30 分
鐘,使膠片顆粒乾燥成白色顆粒。加入10 mM DTT溶液,使之足以淹沒整個膠
片顆粒,在 56℃下還原 1 小時。將溫度降至室溫並移除 DTT 溶液,這時所有
CBR染劑顏色都應該被去除;之後加入相同體積之55 mM iodoacetamide。在室
溫 下 反 應 45 分 鐘 , 避 光 並 不 時 振 盪 。 亦 可 加 入 2%vinylpyridine 來 取 代 iodoacetamide 的功能。清洗膠片薄片以100 µL之 25 mM ammonium bicarbonate
(pH 8.0),反應 10 分鐘,並振盪,之後使用約 100 µL 之 25 mM ammonium
bicarbonate/50﹪acetonitrile 脫水。重複水合與脫水的動作至少三次。將水相移
除,用離心真空乾燥機乾燥。再將膠體顆粒以一倍體積0.5-0.1 mg/mL trypsin 溶 液進行水合,並vortex 5分鐘。在 37℃下反應 12-16 小時。回收 peptides 加入 2 倍體積的水並且vortex 5 分鐘,然後超音波震盪5 分鐘。使用gel-loading tip
第一章 緒論 將peptide 溶液移到新的微量管。利用2倍體積的5% formic acid/50% acetonitrile
再進行兩次萃取。將收集到的peptide 溶液濃縮,使用離心真空乾燥機來將總體
積降到約 10 µL。以 5% formic acid /50% acetonitrile 將體積調整到25 µL。將
收集到的 peptide 溶液儲存在 -20 冰箱,等待 Q-TOF–MS 使用。
2.7 蛋白質電泳轉印法
將4℃冷卻系統將轉印緩衝液(10 mM CAPS, pH11)倒入轉印槽內,並攪拌使溫度 均勻。電泳後的膠片在轉印緩衝液約15 min 平衡。將轉印膜裁成略大於膠片的 方形,以少許甲醇浸溼數秒鐘,再置入轉印槽中的緩衝液10 min後使用。另取兩 張濾紙,以及兩片方形海綿墊放在轉印槽的緩衝液中備用。取出轉印夾打開平 放,先墊一張方形海棉墊,舖上一張溼濾紙,小心疊上已潤溼的轉印紙,其間勿 陷入氣泡;轉印紙再滴上數滴緩衝液後,小心平舖膠片上去,加蓋一層濾紙,及 另一張海綿,也都不可陷入氣泡,即可把整個轉印三明治卡夾裝好。將轉印三明 治置入已經放有一半轉印緩衝液的轉印槽中,注意有轉印膜的那一面向正極 (紅 色),膠片那面向負極 (黑色)。接好電源以400 mA 開始轉印,1.5 h 後中止。轉 印後的膠片可繼續進行 CBR染色,看有無蛋白質殘留。若電泳時加有藍色標準 蛋白質 marker (SeeBlue),則可在轉印紙上看到藍色的色帶,確定轉印成功,並 可評估轉印效率。轉印紙浸在約15 mL尿素洗液中浸洗過夜,期間換三次尿素洗 液並溫和搖盪之。2.8 免疫染色法
尿素浸洗過夜的轉印膜置於塑膠容器中,再以10 mL PBST (phosphate bufferedsaline & Tween)洗三次,每次約10 min,以下的步驟均在此容器中進行。加入明
膠-NET (0.25% gelatin, 0.15 M NaCl, 0.05% Tween, 5 mM EDTA•2Na, 50 mM
Tris)溶液置室溫中反應1 h。倒出明膠,加入一次抗體溶液,置室溫反應1 h。反 應後以PBST洗三次,每次10 min。改用二次抗體連結體溶液反應,在室溫下反 應1 h。以 PBST 洗四至五次,每次10 min。以 PBS 洗二次,每次10 min。加入 DAB (diaminobenzidine) 基質溶液約10 mL呈色,儘量避光,並不時搖動呈色液。 數分鐘內可呈色,在背景加深前倒去 DAB,以水沖數次後晾乾。
2.9 分子量測定
2.9.1 Superose 6 和 Superose 12 (FPLC) 膠體過濾層析
以緩衝液流洗(50 mM Tris buffer, pH 7.5)平衡2~3小時(流速0.3 mL/min)。取100
第一章 緒論 ELISA reader偵測每一分劃(0.3 mL)的蛋白質及酵素活性。另外以100 µL標準蛋 白組製作分子量對數值的校正曲線,算出酵素分子量。
2.9.2 SDS-PAGE
酵素經由SDS-PAGE電泳分離後,算出其相對移動距離(Rf)值,利用標準蛋白質 的相對移動距離值和其分子量對數值求出標準曲線後,算出酵素單元體分子量。2.10 酵素生化性質
2.10.1 酵素反應最適 pH 值
以不同pH值緩衝液配製基質液(Universal buffer,pH 3-10),將酵素和不同pH 值 基質液混合,於 37℃下反應30分鐘。加入100 µL 6 N HCl 終止反應,測量A290。2.10.2 最適反應溫度
將酵素與基質液混合後,分別置於不同溫度下反應 30 分鐘,測量酵素活性的 變化。2.10.3 活化能 (Activation energy, Ea)
先以logkcat (turnover number)對1/T(絕度溫度)作圖,再利用Arrehenius equation:
dlog kcat = (-Ea/2.303R) d(1/T),求活化能 Ea;在實際計算時,使用 log initial
velocity 對 1/T 作圖。
2.10.4 熱安性性
將酵素分別置於不同溫度下保溫 5、10、15、30、45、60 分鐘後,立刻移入 0℃ 冰浴 5 分鐘,再加入基質液,於 37℃下反應30分鐘,測量酵素活性的變化。2.10.5 Km 值
以不同濃度的L-phenylalanine為基質,測定PAL催化反應之反應速率。測得之酵 素反應速率與phenylalanine 濃度作成雙倒數圖,由線性迴歸方程式求Km 值。2.10.6 金屬離子對活性影響
選用 K+、Na+、Ca2+、Mg2+、Mn2+、Hg2+ 及 Co2+ 來探討金屬離子對 PAL 活 性的影響。2.10.7 二級代謝物對活性影響
利用PAL的代謝衍生物,觀察活性變化。原則上,二級代謝物一般帶有苯環,
第一章 緒論 可以當作產物的回饋抑制,亦可當成反應時基質的類似物。依照實驗需要,配製 成不同濃度之酵素基質液:o-coumaric acid, m-coumaric acid, p-coumaric acid, trans-cinnamic acid, ferulic acid, p-amino-benzonic acid, caffeic acid, tannic acid.
2.10.8 基質專一性
使用L-phenylalanine結構類似物,包括D-phenylalanine、L-tyrosine 及
L-tryptophan為基質,測定PAL活性。
2.10.9 化學修飾物對活性影響
選用以下幾種蛋白質化學修飾劑來探討PAL活性會因為何種胺酸被修飾而有 所改變。依照實驗需要,配製成不同濃度之酵素基質液:β-Mercaptoethanol , DTT, PMSF, DEPC, Vinylpyridine.2.11 綠竹中 PAL 基因的選殖
2.11.1 植物材料
使用綠竹筍 (Bambusa oldhamii) 之 cDNA 基因庫 (林, 2001)及染色體DNA 基因庫 (邱, 2000)
2.11.2 載體
Lambda ZAP II : 綠竹筍cDNA基因庫構築時所採用的噬菌體載體,可利用大腸
桿菌XL1-Blue MRF’菌株作為宿主,進行重組噬菌體的放大與增殖。
pBluescript SK(-):Lambda ZAP II載體經由胞內裁切後所得到的載體,作為cDNA
保存與定序之載體。
2.11.3 菌種
XL1-Blue MRF’ 品系: Lambda ZAP II 保存之宿主菌。
SOLR品系: Lambda ZAP II載體經由胞內裁切後所得到的載體保存菌。
MRA 品系: Genomic DNA 庫篩選用宿主菌。
2.11.4 培養基
LB medium (Luria-Bertani medium), LB-Ampicillin Agar (LB/amp plate), LB-Tetra- cycline Agar, LB-Kanamycine Agar, LB Top Agar, NZY Agar, NZY Top Agar
2.11.5 實驗藥品
第一章 緒論 0.5X wash buffer。10 mM MgCl2 buffer: 用以稀釋XL1-Blue MRF’ 菌體。NBT/ BCIP substrate solution:用於呈色反應。Hybridization buffer: 用於雜合反應。 Blocking reagent : 減少非專一性結合反應。Detection buffer: 呈色反應前平衡用。
2.11.6 PAL 專一性探針製備
針對甘藷、玉米、萵苣等植物的phenylalanine ammonia lyase 之 cDNA 序列進 行排列比對,選取數段具高度保留區來設計degenerate primer,將不同degenerate
primer以分別配對方式進行degenerate PCR,將得到的DNA片段進行長度預測
與判斷,經由將第一次得到的PCR產物稀釋後作為模版,再利用nested PCR確 認第一次PCR所得產物的正確性。 PCR反應條件如下: 94 ℃ 5 min 1次循環 95 ℃ 45 sec 52 ℃ 45 sec 40次循環 72 ℃ 1 min 72 ℃ 7 min 1次循環 反應完成後,以膠體純化系統進行PCR產物純化,純化後的PCR產物直接以T-A
clone的方式接入pGEM-T 載體,轉殖於competent cell 並進行定序;定序後所
得之 DNA 序列進行網路核酸資料庫的比對工作,確認該序列與其他物種 PAL 基因極為相似;該段DNA序列有0.8 Kb,可作為我們的專一性探針,進行綠竹 筍 cDNA 基因庫的篩選工作。探針標定方法乃利用 PCR 的方式將 DIG-labeled dNTP嵌入所合成的PCR產物中,作為PCR模版DNA為pGEM-T/PAL0.8kb 。
2.11.7 綠竹筍 cDNA 庫之篩選
2.11.7.1 溶菌斑的轉印
λ 噬菌體cDNA庫的感染力價經測試後約為1x106與1x107 pfu之間 ,取相當 於力價50000 pfu/plate 的λ噬菌體cDNA庫,與 600 µL、A600 = 0.5 的XL1-BlueMRF’ 大腸桿菌細胞混合後,加入 6.5 ml NZY top agar(充分溶解並降溫為
48℃),迅速倒入 NZY agar plate。等待NZY top agar 凝固後放入 37℃ 培養箱, 培養6~8 小時;8小時後將NZY agar plate放於4℃冰箱保存2小時,(防止top agar 沾黏到 nitrocellulose 薄膜上);2 小時後將NZY agar plate由4℃冰箱取出,覆
蓋上 nitrocellulose薄膜5分鐘,並利用無菌針頭作出標記(使用打洞的方式,至
少需要打3 個洞以釐清方向);將nitrocellulose薄膜浸泡於1.5 M NaCl 與0.5 M
NaOH 的混合液中 10 分鐘,使 DNA 分子變性(denature),放於濾紙上 15 分鐘
第一章 緒論
混合液中 10 分鐘,使 DNA 分子復性,放於濾紙 15 分鐘使其風乾。再將
nitrocellulose薄膜浸泡於2 x SSC buffer中10分鐘,放於濾紙上15到20分鐘使 其完全風乾。風乾後將 nitrocellulose 薄膜利用能量達12,000 µJ 的UV光源照射 約2 到3 次,使DNA分子與nitro- cellulose薄膜形成共價鍵結(crosslink)。並妥
善保存NZY agar plate於4℃,待雜合反應完成後進行正反應株確認與挑選的工
作。
2.11.7.2 雜合反應
將 轉 印 完 成 的 nitrocellulose 薄 膜 放 入 洗 淨 後 的 保 鮮 盒 當 中 , 加 入 pre- hybridization buffer/blocking reagent以進行預先雜合反應,於 60℃下在恆溫反應 箱中反應 8小時以上。待 pre-hybridization完成後,加入DIG 標定完成後的PAL
專一性探針進行雜合反應,反應溫度為60℃,進行至少16小時。
2.11.7.3 呈色反應
待雜合反應完成後取出 nitrocellulose薄膜,放入 2x SSC buffer 於室溫下清洗 10 分鐘,重複進行共兩次,隨即放入 0.5x SSC buffer 於 60℃下清洗10分鐘, 重複進行共兩次,取出nitrocellulose薄膜並滴乾洗液,放入washing buffer中平
衡 2 分鐘,由 washing buffer 中取出 nitrocellulose 薄膜並稍微滴乾水分,放入
blocking solution中60分鐘並在室溫下輕微搖晃;加入相對於blocking solution 體 積五千分之一(1/5000)的anti-DIG antibody,室溫下反應60分並緩和的搖晃,使 反應能完全且均勻。反應完成後將 nitrocellulose 薄膜取出並以 washing buffer
在室溫下潤洗15分鐘,共進行兩次;潤洗完成後將 nitrocellulose薄膜取出,輕
微瀝乾之後放入 detection buffer 中平衡約 5 分鐘,加入適量 substrate solution
(NBT/BCIP)進行呈色反應,於 37℃下極輕微的搖晃混合,於3小時後觀察情形。
2.11.7.4 挑選正反應之溶菌斑
將呈色後的 nitrocellulose 薄膜放入乾淨的塑膠袋中並固定之,再以色筆將正反
應溶菌斑 的位置 上標 示記號; 取出 4℃ 保 存下的 NZY agar plate,對 照於
nitrocellulose 薄膜的正確位置後,以 yellow tip 將該位置的溶菌斑挖下懸浮於
0.5 mL 的 SM-buffer中,再次測定力價並進行第二次獲第三次的篩選,以挑選
出正確的正反應溶菌斑。
2.11.7.5
λ
噬菌質體的胞外切除 (in vitro excision)
以 LB/maltose/MgSO4 培養XL1-blue MRF’ 品系大腸桿菌與SOLR品系之大腸
桿菌;以2000g 離心1 分鐘後,去除LB medium而加入10 mM MgSO4 水溶液
將兩種品系的大腸桿菌細胞調整濃度達到A600 = 1.0;將大腸桿菌與λ噬菌體以下
第一章 緒論 250 µL的λ噬菌體(約包含1 x 105 力價的噬菌體),1 µL的輔助噬菌體(ExAssist helper phage) (約包含1 x 106 pfu/µL)。混合後將無菌塑膠試管置於37℃下,放置 15 分鐘;隨即加入3.0 mL 的 LB medium,放置於37℃ shaker上搖晃8 小時後, 將試管於70℃下放置20 min,離心15分鐘(1000 x g),取上清液100 µL 與SOLR 品系大腸桿菌200 µL (濃度為 A600 = 1.0)混合,於 37℃下放置15分鐘後,取出 200 µL塗抹接種於LB/amp plate,放置37℃下16 小時以上;能在 LB/amp plate
上生長之大腸桿菌即為轉殖成功的菌體,隨即進行大量培養,分離質體 DNA 後 進行 DNA 定序。
2.12 綠竹筍 PAL 的定序
將轉殖成功的SOLR品系大腸桿菌送定序;一共送出8個不同的正反應株,第 一次採用 T7 primer 定序,將得到的結果進行網路資料庫比對,若為 PAL 的部 份序列,則再用T3 primer 定序,可快速確認所篩選的PAL基因是否為包含全長的序列(包含 start codon 與 poly-A tail),確認可能得到全長 PAL基因後,則由
定序結果再設計專一性引子,進行 5’-端或 3’-端的定序,每次定序的結果約 600
個核苷酸序列,以 PAL 基因的長度而言,約需要定序4次才能得到全長的核酸
序列。
2.13 染色體 DNA 庫之篩選
將含有約5 ×104 pfu之噬菌體溶液加入600 µL上述菌液,置於37℃ 15 分鐘後,
加入以50℃預熱之8~9 mL NZY top agar,並快速倒入直徑為15 cm之plate 中,
待其凝固後,於 37℃ 培養過夜,使噬菌體之溶菌斑生長至全滿。隔日,加入 6-9 mL SM buffer,於室溫中緩慢震盪3小時,使得噬菌體懸浮在SM buffer中。吸 出 plate上的噬菌體懸浮液,加入5% 體積的氯仿,劇烈震盪後以14,000 rpm 離 心10分鐘,取出上清,加入DNase與 RNase至濃度分別為20 g/mL 與 8 g/mL, 於 37℃ 下作用30分鐘。加入氯化鈉固體使濃度成為1 M,混合溶解後置於冰 上1 小時,此高鹽情況下可使噬菌體與宿主細菌分離。在4℃下以 11,000g,離 心 10 分鐘,取上清液到乾淨離心管中。再加入PEG 8000 使濃度為10% (w/v) 用以沈澱噬菌體,溶解後於室溫下放置1小時。4℃下以 11,000g 離心10 分鐘, 沈澱物以 1-3 mL SM buffer 溶解。加入等體積之氯仿萃取 PEG 8000,於 4℃ 以 3,000g 離心15 分鐘,收集上清液,並於4℃下以25,000 rpm 之超高速離心 2 小 時,可得到半透明之噬菌體沈澱。半透明之噬菌體以 1-2 mL SM buffer 溶解, 於 4℃下放置過夜以幫助回溶。隔日確定沈澱已完全回溶後,加入 EDTA 至最 終濃度為 20 mM,加入 proteinase K 至最終濃度為 50g/mL,再加入 SDS 至 0.5% (w/v),混合均勻,於 37℃下反應1小時。在室溫下以等體積PCI 萃取一
第一章 緒論 次,再用 CI 萃取一次,回收水層。加入1/10 體積之3 M sodium acetate (pH 7.0), 與 2 倍體積之酒精,於室溫下放置 30 分鐘,即可看到絲狀物出現,直接以 12,000 rpm 離心分鐘,沈澱再以70% 酒精清洗,乾燥後,回溶於40~50 mL 無 菌水中,置於 -20℃ 保存。
2.14 大腸桿菌之表現載體
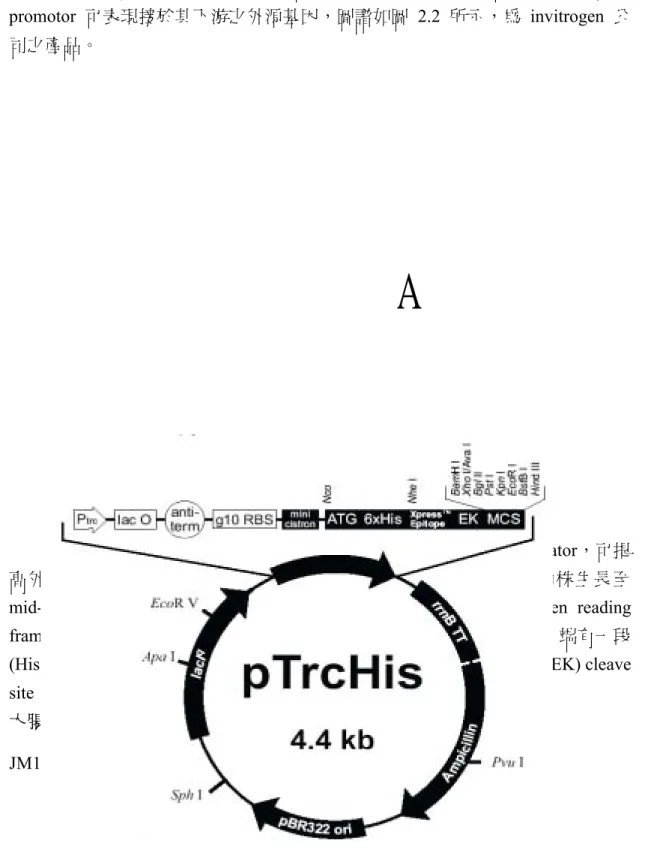
PAL 1/pBluescript SK(-)、PAL 2/pBluescript SK(-):為本實驗室保存綠竹 PAL 1 與 PAL 2 基因全長序列所用的質體。pTrcHisA:為表現載體,其上具有 trc
promotor 可表現接於其下游之外源基因,圖譜如圖 2.2 所示,為 invitrogen 公
司之產品。
圖 2.2、表現載體 pTrcHisA 之圖譜 Fig.2.2 Map of the pTrcHisA expression vector
表現載體 pTrcHisA 具有 trp (trp-lac) promoter與rrnB antiterminator,可提
高外來基因之表現,同時可以表現 lac repressor protein,當轉形菌株生長至
mid-log phase 時,添加適量的 IPTG 誘導表現外源基因;而在 open reading
frame 前有 minicistron 序列,可以增加外源基因進行轉譯的效率;N 端有一段
(His)6-tag的序列有利所表現蛋白之純化工作,並且帶有 enterokinase (EK) cleave
site 可以去除N 端融合約3 kDa 的片段。其宿主為大腸桿菌Top 10。
大腸桿菌
JM109:作為質體保存、增殖之宿主。
第一章 緒論
Top10:為表現載體 pTrcHisA 之宿主,用於大量表現重組蛋白之菌株。
上述菌株之基因型 (genotype) 表示如下:
菌株 基因型
JM109 e14-(McrA-) recA1 endA1 gyrA96 thi-1 hsdR17(rK- mK+) supE44 relA1 (lac-proAB) F’ traD36 proAB lacIqZM15
Top10 F-, mcrA .(mrr-hsdRMS-mcrBC) Φ80lacZ.M15 .lacX74 deoR recA1 araD139(ara-leu)7697 galU galK rpsL endA1 nupG
2.15 表現載體之建構
選用pTrcHisA載體系統(Invitrogene 公司之產品)作為苯丙胺酸脫氨裂解酶基
因的表現載體,於大腸桿菌Top10中表現。建構表現系統時,以帶有限制酶切位
(BamHI 與 EcoRI)的引子經聚合酶鏈鎖(polymerase chain reaction)反應產生大量 PAL 1 與 PAL 2 DNA 片段,接著以限制酶 BamHⅠ與EcoRⅠ分別處理表現載體
pTrcHisA、PAL 1 與 PAL 2 序列,再將處理過的PAL1與PAL2分別與處理過的表
現載體接和、轉形進入表現宿主大腸桿菌Top 10,最後以限制酶及定序分析所接
入的序列無誤後,再進行重組蛋白最佳誘導條件之探討
2.15.1.苯丙胺酸裂解酶基因接入表現載體
表現載體建構策略,設計四段分別包含BamHI 與EcoRI 限制酶切位的專一性引
子 (圖 2.3),利用PCR反應增殖放大出的PAL 1 與PAL 2 全長片段,以BamHI 與 EcoRI 限制酶分別切割PAL1 、PAL2 及 pTrcHisA,再將處理過的DNA片段分別 接合(ligation)到pTrcHisA載體的His•Tag序列的下游,形成表現載體 pTrcHisA /PAL1 與 pTrcHisA /PAL 2,經誘導可產生在N 端具有連續6 個Histidine的 PAL 融合蛋白質。
For PAL1
第一章 緒論
P5 P6
5' CGC GGA TCC ATG CCG CGC GAG GAT3‘ 5' CCG GAA TTC GCA GAT GGG CAG TGG CTC 3'
For PAL2
P7 P8
5’ CGC GGA TCC ATG GAG TGC GAG AAT G 3’ 5’ CCG GAA TTC ATT AGT TGA TGG GCA 3’
圖 2.3 含有限制酶切位的專一性引子與 pTrcHisA 部分載體
2.15.2 聚合酶鏈鎖反應
聚 合 酶 鏈 鎖 反 應(polymerase chain reaction, 簡 稱 PCR)是 利 用 pfu DNA
polymerase之耐高溫及PCR溫度控制器,在微量離心管中,將DNA template 高
溫加熱打開雙股螺旋後,兩個互補股末端接合上設計之primers,進行n 次之反 應就可得到 2n 倍之 DNA,於短時間內獲得大量欲增生之 DNA 片段。將離心 管置入 PCR 溫度控制器中進行聚合鏈鎖反應,其反應程式如下:聚合酶鏈鎖 反應完成後,將離心管移置4 ℃暫存或進行電泳分析。 30 cycle 95℃ 94℃ 61℃ 74℃ 74℃ 4℃ 5分鐘 1分鐘 0.5分鐘 3.5分鐘 1分鐘 60分鐘
2.15.3 質體 DNA 之小量分離
取1.5 mL 菌液,加於微量離心管中,以6,000 rpm 離心2 min後,倒去上清, 以微量吸管頭儘可能吸去殘留液體。沉澱加入0.1 mL MP I (25 mM Tris-HCl, pH 8.0; 10 mM EDTA, pH 8.0; 50 mM glucose),劇烈震盪,將沉澱完全打散。慢慢加 入 0.2 mL MP II (0.2 N NaOH; 1% SDS; 使用前以 10 N NaOH, 10% SDS 稀釋混 合配製),蓋上管蓋後將管子反覆上下搖動,注意不可震盪!此時溶液應漸澄清 且黏度增加;將離心管置冰浴中。加入 0.15 mL MP III (3 M 醋酸鉀溶液,pH 5.2,置冰浴中),混合均勻,亦不可劇烈震盪,置冰浴中 5 min。以12,000 rpm 離 心5 min。小心吸出上清至另一離心管,加入2 倍體積之純酒精 (或 0.6 倍體積之isopropanol),混合均勻,置室溫 2 min。以12,000 rpm 離心10 min。倒去上 清液,沉澱以70% 酒精洗二次,每次各離心 1 min。沉澱以SpeedVac 乾燥後, 以 30 µL TE-8.0 (10 mM Tris-HCl, pH 8.0; 1 mM EDTA, pH 8.0)溶解。進行洋菜膠
體電泳,視色帶位置,檢測DNA片段大小是否正確。
2.15.4 DNA 片段之分離純化
A. Gel Extraction Kit (Viogene)
將PCR產物進行洋菜膠體電泳。將DNA片段以小刀自洋菜膠體割下。秤膠體
重量,並加入膠體三倍體積之Buffer GEX (100 mg 膠體視為100 µL)。置於50 ℃
第一章 緒論
水浴10 分鐘至膠體完全溶解。可每隔2-3分鐘震盪。膠體完全溶解之後,將其
加入spin column,13,000 rpm 離心1 分鐘。倒掉濾液,加入500 uL Buffer WF 至
管柱中,13,000 rpm 離心1 分鐘。倒掉濾液,加入700 uL Buffer WS 至管柱中,
13,000 rpm 離心1 分鐘,倒掉濾液,再次13,000 rpm 離心1分鐘。以36 µL 無 菌水加入管柱中,室溫靜置1分鐘後,13,000 rpm離心1 分鐘將DNA 流洗出來。
反應完進行洋菜膠體電泳。檢測DNA片段大小是否正確。
B. PCR Clean-up Kit (viogene)
將限制酶切割後的產物加入 500 µLBufferPS,混合均勻。將溶液加入 spin column,13,000 rpm 離心1 分鐘。倒掉濾液,加入500 µL Buffer WF 至管柱中, 13,000 rpm 離心1 分鐘。倒掉濾液,加入700 uL Buffer WS 至管柱中,13,000 rpm 離心1 分鐘,倒掉濾液,再次13,000 rpm 離心1 分鐘。以36 µL 無菌水加入管 柱中,室溫靜置1 分鐘後,13,000 rpm 離心1 分鐘將 DNA 流洗出來,收集濾 液。反應完進行洋菜膠體電泳,視色帶位置,檢測DNA 片段大小是否正確
2.15.5 質體與 DNA 片段之限制酶切割
質體 DNA (pTrcHisA vector)1.0 µL,加入限制酶0.5 µL BamHⅠ 及 0.5 µL EcoRⅠ,1.0 µL之10×EcoRⅠ Buffer,加水至總體積為10 µL,37 ℃ 下反應 1~2 小時。DNA 片段 (PAL 1 或 PAL 2) 10 µL,加入限制酶 0.5 µL BamHⅠ,0.5 µL EcoRⅠ,2 µL 之 10× NE Buffer,加水至總體積為20 µL,37 ℃ 反應 1~2 小時。 反應完後,以Viogene PCR clean-up kit純化DNA片段。
2.15.6 接合反應 (Ligation)
將分別經限制酶(BamHⅠ 與 EcoRⅠ)切割的 pTrcHisA 與欲接入表現載體的 DNA 片段(PAL 1 與 PAL 2),利用 T4 DNA ligase 連接起來,再度形成完整的表 現質體。添加試劑至 1.5 mL 微量離心管: 1 µL pTrcHisA vector, 16 µL insert DNA (PAL 1or PAL 2), 1 µL T4 DNA ligase, 2 µL Ligation 10× buffer。12~14℃ 反應 16~20 小時。
2.15.7 Competent cell 製備
進行轉形實驗前16~20 h,自-70℃取出貯存的菌種,以劃單一菌落方式將菌株
Top10 接種在SOB(2% Bacto tryptone, 0.5% yeast extract, 10 mM NaCl, 2.5 mM
KCl, 1.5% Bacto agar)固體培養基上,在37℃培養。取40 mL SOB 培養基裝入
125 mL 已滅過菌之三角瓶。另取1 mL SOB加於微量離心管中。由SOB 固體培
養基上選4~6 個約2~3 mm 大小的單一菌落接入1 mL SOB中,震盪將菌體打
散後,加入40 mL SOB 中,直至A600 約0.5~0.6。收集菌液於離心管中,置冰
第一章 緒論
體倒乾,或以微量吸管頭吸乾淨。沉澱加入1/3 菌液體積冰冷的0.1 M CaCl2,
稍加震盪,混合均勻,置冰浴中 10~15 min。加入 1/25 菌液體積冰冷的 0.1 M
CaCl2,稍加震盪使混合均勻,competent cells 製備完成。
2.15.8 質體之轉形
將表現質體DNA (<10 µL)先裝於微量離心管中,置冰浴中預冷,每管加入 200
µL competent cell,震盪混合後,置冰浴中20~40 min。42℃加熱90秒。置冰浴
中2 min。加入800 µL SOC,混合後,於37℃ 震盪培養30~60 min。將各管轉
形液上下搖動混合均勻,取20 µL 至另一離心管中,加入180 µL SOC,混合均 勻後,塗佈於 SOC/Amp plate。剩餘菌液以6,000 rpm 離心1 min,倒去上清,
沉澱加入200 µL SOC,均勻打散後塗抹在另一個SOC/Amp plate 上。倒置培養
皿於 37℃ 培養16~20 h 後,觀察各個plate上菌落的形狀並計算菌落數,抽取 質體以限制酶及定序分析所建構的表現載體,確認 insert DNA。
2.16 表現蛋白之誘導
2.16.1 最適 IPTG 濃度與最適溫度探討
挑單一菌落培養於3 mL LB medium(50µg / mL ampicillin),37 ℃震盪培養過 夜。隔天,取100 µL菌液加入 3 mL LB medium (50µg / mL ampicillin),震盪培 養至 A600 達到0.6~0.8。加入不同量的IPTG,使最終濃度分為 0、0.5、1、1.5、 2 mM,誘導表現,以120 rpm 震盪培養在 30℃、37℃ 二種不同的溫度。誘導 4 h 後,收菌置於 4℃,以8000 rpm 離心15-20 min,倒去上清液,存於-80℃中 保存。以 10% SDS-PAGE 檢測誘導情形,並測活性。2.16.2 最適時間探討
挑單一菌落培養於3 mL LB medium(50 µg/mL ampicillin),37℃震盪培養過夜。 隔天,取100 µL 菌液加入3 mL LB medium (50 µg/mL ampicillin),置於37℃, 以120 rpm 震盪培養至A600 達到0.6~0.8。加入2 mM IPTG,置於30℃,以120 rpm 震盪培養,分別誘導2、4、6、8、10、12 小時。收菌置於4℃,以 8000 rpm 離心15-20 min,倒去上清液,存於-80℃ 中保存。2.16.3 表現蛋白之純化
重組蛋白質在 N 端帶有由六個 Histidine 標籤,可用鎳離子親和管柱進行純化。其原理為利用Sepharose 膠體上的基質iminodiacetic acid 螯合鎳離子可與表
現蛋白 N 端 6 個 histidine 結合,可用 imidazole 競爭結合鎳離子而溶離出目
標蛋白,達到純化效果。大量培養表現菌株,離心破菌取上清,得到粗抽液,再 以 Ni2+ chealting 親和管柱純化帶有His6-tagged 的PAL,純化流程如下:
第一章 緒論 Crude lysate(from 200 mL liquid culture)
lysis buffer, pH 8.0
Ni-IDA column PBS buffer, pH 8.0
Imidazole gradient ( 10~500 mM )
10 % SDS-PAGE & Activity assay
2.16.4 粗抽液之取得
大量培養所得菌體,加入25 mL lysis buffer (50 mM NaH2PO4, 100 mM NaCl,
10 mM imidazole)重新懸浮菌體,並加入lysozyme (40 mg/mL) 使最終濃度為 1
mg/mL,靜置冰上1 h。以超音波細菌粉碎機打破細胞,每打一分鐘,休息一分
鐘,共打15 次。以4℃, 12000 rpm 離心20 min,上清液即為粗抽液。
2.16.5 批次鎳離子親和管柱層析
Hi-Trap column 以緩衝液流洗二倍體積,注入 10 mL 之 10 mM NiSO4, 將
chelating sepharose 螯合上鎳離子。再以緩衝液流洗兩倍體積。將25 mL 粗抽液
以蠕動幫浦直接注入已螯合Ni2+ 的Hi-Trap column,並且以50 mL離心管開始 收集第一個分劃(Flow-through)。接著以25 mL wash buffer (100 mM imidazole) 流 洗,並收集第二個分劃(Wash)。再以25 mL elution buffer1 (250 mM imidazole) 流 洗,並收集第三個分劃(Elution 1)。最後以25 mL elution buffer1 (500 mM imidazole) 流洗,收集第四個分劃(Elution 2)。以10% SDS-PAGE 檢測純化情形,並測活性。