行政院國家科學委員會專題研究計畫 期中進度報告
(子計畫三)利用核糖核酸結構預測與核糖核酸-蛋白質互
動關係分析推論蛋白質結構(1/3)
計畫類別: 整合型計畫
計畫編號: NSC94-2627-B-009-004-
執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日
執行單位: 國立交通大學資訊科學學系(所)
計畫主持人: 胡毓志
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 95 年 5 月 12 日
行政院國家科學委員會補助專題研究計畫期中報告
※※※※※※※※※※※※※※※※※※※※※※※※※
※
※
※
利用核糖核酸結構預測與核糖核酸-蛋白質互動關係分析推論
※
※
蛋白質結構(1/3)
※
※
※
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:整合型計畫
計畫編號:NSC 94-2627-B-009 -004-
執行期間:94 年 8 月 1 日至
95 年 7 月 31 日
計畫主持人:國立交通大學資訊工程系 胡毓志
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:交通大學資訊工程系
中
華
民
國
95 年
5
月
6
日
行政院國家科學委員會專題研究計畫期中報告
國科會專題研究計畫成果報告撰寫格式說明
Preparation of NSC Project Reports
計畫編號:NSC
94-2627-B-009
-004-執行期限:94 年 8 月 1 日至 95 年 7 月 31 日
主持人:胡毓志
交通大學資訊工程系
計畫參與人員:許登貴 賴昀君 黃子緯 交通大學資訊科學系
中文摘要
在本計劃執行的第一年中,我們針對多數已 常被應用於核糖核酸結構預測的方法做評估與分 析,此外,我們將搜尋有實用價值的屬性作為檢測 核糖核酸與蛋白質交互關聯的基礎,由於以往所提 出的方法有其個別的應用目的與設計理念,因此必 須先分析其優缺點。我們計畫研發新的結構預測系 統,同時計劃將結構預測與分群合而為一,提升系 統的使用彈性。關鍵詞: 核糖核酸、二級結構、分群
Abstract
The goal of the first-year period for the project is to investigate those various methods for RNA secondary structure prediction. In addition, we planed to find useful features that can be applied to identifying the correlation between RNA and proteins. These previously proposed approaches are based on different design philosophies as well as with quite different purposes. We were aimed to evaluate the pros and cons of these systems, and planed to develop a novel prediction system that is flexible enough to deal with a wide variety of RNA secondary structures. After a thorough analysis and comparison of the systems available, we have decided to design a new RNA structural motif predictor combined with RNA clustering. With this new approach, we expect to perform RNA clustering and secondary structure prediction simultaneously.
Keywords: RNA, secondary structures, clustering
Introduction
RNA molecules are the key players in the biochemistry of the cell, playing many important roles in regulation, catalysis and structural support. Like proteins, their functions generally depend on their structures. Although structural genomics, the systematic study of all macro-molecular structures in a genome, is currently focused more on proteins, thousands of genes produce transcripts exerting their functions without ever producing protein products [1]. It can be easily argued that the comprehensive understanding of the biology of a cell requires the knowledge of identity of all functional RNAs (both
non-coding and protein-coding) and their molecular structures. Since it is often difficult to acquire the 3D spectrum data of RNA molecules for structure determination, versatile and reliable computational methods that can predict RNA structures are highly desirable.
Many functional RNAs have evolutionarily conserved secondary structures in order to fulfill their roles in a cell. For protein-coding RNAs, some of the functions can be presented by functional motifs. For example, several best-understood structurally conserved RNA motifs are found in viral RNAs, such as the TAR and RRE structures in HIV and the IRES regions in Picornaviridae. Apparently, structural information is very useful in characterizing a class of functional RNAs. Based on characteristic structures, we can likely identify novel functional RNAs or partition given RNAs into biologically meaningful families. Several systems have been developed to find consensus structural elements within a family of functionally related RNAs; however, there is little work on clustering of unaligned RNAs based on characteristic secondary structures. Given a set of unaligned RNA sequences without prior knowledge of the number or identity of families in the set, our goal is to automate both clustering and secondary structure prediction simultaneously. In this report, we propose an adaptive approximation approach combined with a genetic programming-based structure prediction method to identify from unaligned RNAs reasonable clusters associated with characteristic secondary structure elements.
Material and Methods
There are currently a lot of various approaches to RNA secondary structure prediction; nevertheless, few of them address the issue of simultaneous RNA clustering and structure prediction. According to the system log of our earlier system, GPRM, we found that those RNAs submitted by the users (possibly biologists) were not necessarily within a single family per se. This gives us the idea that biologists generally do not know a specific RNA family in advance. Thus we aimed to develop a new system to find meaningful clusters and their characteristic structure motifs at the same time.
set of unaligned RNAs without knowing beforehand how many clusters actually existing in this set, we assume that each cluster is likely a functional family that contains characteristic structure elements. Based on this assumption, our new method is focused on finding significant consensus structure elements that can be used to characterize the families of RNAs. Since the number of clusters and its size are not known in advance, we adapt a generate-and-test strategy that iteratively adjusts the hypothesized cluster size until some significant consensus structure elements can be found associated with this cluster. After a cluster is obtained, all its members are then removed from the given set of RNAs. We can repeat the same separate-and-conquer strategy to identify other clusters until the set of RNAs is emptied.
The generate-and-test strategy we use is an adaptive approximation approach that systematically revises the hypothesized range of the cluster size. The cluster size is defined by a range between an upper bound U and a lower bound L. Without sufficient prior information of clusters, an initial hypothesis about the cluster size is generated for a given set of n RNA sequences. The cluster size is initialized within a range between an upper bound U=n and a lower bound L=0, that is, we first assume that all the given RNA sequences may form an entire family. To the entire family, a genetic programming-based structure prediction method is applied to look for the fittest consensus structure elements. The definition of fitness is detailed later. If the specificity of the structure elements associated with a cluster exceeds or equals some pre-specified threshold, the hypothesis is considered verified, and the cluster along with the associated structure elements will be reported. On the other hand, low specificity suggests that the current hypothesized cluster size is too big to be real and needs to be decreased. In this case, we reduce the current hypothesized cluster by updating U to the median (i.e. n/2) of the current U(=n) and L(=0), and L remains the same. With the cluster sized fixed in the new range [0,n/2], we search the fittest consensus structure elements and evaluate their specificity again. If the specificity is still lower than the threshold, we modify the cluster size by updating U in the same way as above. The same process for cluster reduction can be repeated till we find a cluster with high-specificity structure elements. On the contrary, if the specificity is over or equal to the threshold, it can mean one of the two possibilities: (1) the current cluster is likely real, and any more sequences added will be harmful to the specificity of consensus structures, or (2) the current cluster found is only a subset of a bigger real cluster. To verify which event actually happens, we increase the cluster size. We set L to the current U’s value, and change U to the median of its current value and its last. After the update, the new range for cluster size becomes [n/2,3n/4], and a new search for the fittest consensus structure elements constrained by the new range is conducted. Likewise, we have two possible outcomes from the examination of consensus structure
specificity, better (or equal to) or worse than the threshold. Based on the outcome, we can either increase or decrease the cluster size by revising its range. As each update generates a tighter range for cluster size, we expect the range will eventually converge to the appropriate cluster size.
Consensus structure element prediction can be considered a supervised learning problem which involves both positive and negative examples. Positive examples are a given set of RNA sequences; negative examples are some number of sequences randomly generated based on the observed frequencies of sequence alphabet in positive examples. The objective here is to learn the structure elements that can be used to distinguish the given functionally related sequences from the random sequences.
We modify our previous system, GPRM, an RNA consensus secondary structure prediction tool, to find significant structure elements from a dataset that may contain multiple variable-sized clusters of unaligned sequences. GPRM has been tested on several real RNA families, including pseudoknots, and shown its effectiveness in predicting conserved structure elements in a given RNA family. To describe the characteristic structure elements for a cluster, we adapt the same representation that is expressive enough to even represent pseudoknots. We also apply the same genetic operators to optimize candidate structure elements during evolutionary process. What is different from the previous work is the fitness function.
The fitness function is used to measure the quality of individuals (i.e. candidate structure elements) in a population. The higher the fitness of an individual, the better its chances of survival to the next generation. In the previous work, the input dataset was assumed to be a single class of functionally related RNA sequences. We were interested in those structure elements that can reflect the characteristics conserved in a family, e.g. the RNA protein binding sites. Derived from the F-score, the fitness function was aimed to balance the importance of two measures, recall (i.e. sensitivity) and precision (i.e. positive predictive value). It assigns higher values to those structural motifs commonly shared by the given family of RNAs, and rarely contained in random sequences. For a given set of RNA sequences that form a single family only, the fitness function used in can effectively guide the evolutionary process in genetic programming. Nevertheless, when the input dataset contains multiple functional classes, the recall measure may dominate the calculation of F-score if the fitness function treats the entire dataset as a single class. This will mislead the system to find over-general elements shared by most sequences. To alleviate the bias, we define a new measure of recall, and present the fitness function by taking into account the hypothesized cluster sizes. In that case, we can better constrain the search space, and accelerate the convergence of the optimization process.
Lessons Learned and Future Work
Without the prior knowledge of the number of clusters, it is computationally difficult to simultaneously perform clustering and structure prediction. According to our preliminary study, the simple generate-and-test approach combined with the revised fitness measure in our earlier system can effectively estimate the family size, and consequently identify RNA families. With this new combinatorial approach, we expect to deal with a wider variety of RNAs even without knowing their family identity. Furthermore, the common conserved structure motifs can be considered useful features used to characterize RNA families. This sheds some light on the possibility of classifying RNA-protein interactions based on these features.
In the following year, we plan to design more thorough tests on the new approach as well to collect RNA-protein interaction literature and resource of sorts. For the tests, several objectives are considered. Firstly, we will test the accuracy of the structure prediction. Secondly, we will evaluate how well the clusters predicted correspond to real RNA families. Thirdly, we will investigate the robustness of the new approach in terms of various RNA family sizes and motif forms. Lastly, in addition to structural motifs, based on the domain knowledge obtained from the literature available, we plan to derive more useful features to describe RNA-protein interactions.
References
1.
The Genome Sequencing Consortium (2001) “Gene content of the human genome”, Nature, 409, p860-921.2.
Zuker, M and Stiegler, P. (1981) ”Optimal computer folding of larger RNA sequences using therdynamics and auxiliary information”, Nucleic Acids Res., 9, p133-148.3. Zuker, M. (1989) “On finding all suboptimal foldings of an RNA molecule”, Science, 244, p48-52.
4. Hofacker, I., Fontana, W., Stadler, P., Bonhoeffer, L., Tacker, M. And Schuster, P. (1994) “Fast folding and comparison of RNA secondary structures”, Monatsh. Chem., 125, p167-188. 5. Chiu, D. and Kolodziejczak, T. (1991) “Inferring
consensus structure from nucleic acid sequences”, Comput. Appl. Biosci., 7, p347-352.
6. Gutell, R., Power, A., Hertz, G., Putz, E. and Stormo, G. (1992) “Identifying constraints on the higher-order structure of RNA: continued development and application of comparative sequence analysis methods”, Nucleic Acids Res., 20, p5785-5795.
7. Gulko, B. and Haussler, D. (1996) “Using multiple alignments and phylogenetic trees to detect RNA secondary structure”, Proc. Pac. Symp. Biocompt., p350-367.
8. Akmaev. V., Kelley, S. and Stormo, G. (1999) “A
phylogenetic approach to RNA structure prediction”, Proc. Int. Conf. Intell. Syst. Mol. Biol., p10-17.
9. Eddy, S. and Durbin, R. (1994) “RNA sequence analysis using covariance models”, Nucleic Acids Res., 22, p2079-2088.
10. Sakakibara, Y., Brown, M., Hughey, R., Mian, I., Sjolander, K., Underwood, R. and Haussler, D. (1994) “Stochastic context-free grammars for tRNA modeling”, Nucleic Acids Res., 22, p5112-5120.
11. Durbin, R., Eddy, S., Krogh, A. and Mitchison, G. (1998) “Biological sequence analysis”, Cambridge University Press.
12. Abraham, J., van denBerg, M., van Batenburg, F. and Pleij, C. (1990) ”Prediction of RNA secondary structure, including pseudoknotting, by computer simulation”, Nucleic Acids Res., 18, p3035-3044.
13. Gultyaev, A., van Batenburg, F. and Pleij, C. (1995) “The computer simulation of RNA folding pathways using a genetic algorithm”, J. Mol. Biol., 250, p37-51.
14. van Batenburg F., Gultyaev, A. and Pleij, C. (1995) “An APL-programmed genetic algorithm for the prediction of RNA secondary structure”, J. Theor. Biol., 174, p269-280.
15. Hu, Y. (2002) "Prediction of consensus structural motifs in a family of coregulated RNA sequences", Nucleic Acids Res., 30, p3886-3893.
16. Luck, R., Graf, S. and Steger, G. (1999) “ConStruct: a tool for thermodynamic controlled prediction of conserved secondary structure”, Nucleic Acids Res., 27, p4208-4217.
17. Hofacker, I., Fekete, M. and Stadler, P. (2002) “Secondary structure prediction for aligned RNA sequences”, J. Mol. Biol., 319, p1059-1066. 18. Juan, V. and Wilson, C. (1999) “RNA secondary
structure prediction based on free energy and phylogenetic analysis”, J. Mol. Biol., 289, p935-947.
19. Gorodkin, J., Stricklin, S. L. and Stormo, G. D. (2001) Discovering common stem-loop motifs in unaligned RNA sequences. Nucleic Acids Res., 29, 2135-2144.
20. Rivas, E. and Eddy, S. (1999) “A dynamic programming algorithm for RNA structure prediction including pseudoknots”, J. Mol. Biol., 285, p2053-2068.
21. Mathews, D. and Turner, D. (2002) “Dynalign: an algorithm for finding the secondary structure common to two RNA sequences”, J. Mol. Biol., 317, p191-203.
22. Hu, Y. (2003) "GPRM: a genetic programming approach to finding common RNA secondary structure elements", Nucleic Acids Res., 31, p3446-3449.
23. Hofacker, I., Priwitzer, B. and Stadler, P. (2004) “Prediction of locally stable RNA secondary structures for enome-wide surveys”, Bioinformatics, 20, p186-190.
24. Clark, P and Boswell, R. (1991) “Rule Induction with CN2: some recent improvements”, in Proceedings of the Fifth European Conference on Machine Learning, p151-163.
25. Cheeseman, P. and Stutz. J. (1996) “Bayesian Classification (AUTOCLASS): Theory and Results”, in Advances in Knowledge Discovery and Data Mining, p153-180, AAAI.Batenburg, F.H.D. van, Gultyaev, A.P. and Pleij, C.W.A. (2001) “PseudoBase: structural information on RNA pseudoknots”, Nucleic Acids Res., 28, 1, 201-204.
26. Matthews, B.W. (1975) Comparison of the predicted and observed secondary structure of T4 phage lysozyme. Biochem. Biophys. Acta, 405, 442-451.
27.
Rand, W. M. (1971) Objective criteria for the evaluation of clustering methods. Journal of American Statistical Association, 66, 846–850.行政院國家科學委員會補助國內專家學者出席國際學術會議報告
95 年 4 月 27 日報告人姓
名
胡
毓
志
服務機構
及職稱
交通大學資訊工程系
副教授
時間
會議
地點
06/26/2006-06/29/2006 Las Vegas, USA本會核定
補助文號
NSC
94-2213-E-009-143-會議
名稱
(中文) 生物資訊暨計算生物學國際研討會
(英文)2006 International Conference on Bioinformatics and
Computational Biology
發表
論文
題目
1.(中文) 以多策略方法設計蛋白質結構字元
(英文)
A Multi-Strategy Approach to Protein Structural Alphabet Design因期中進度報告繳交期限在研討會舉行之前,因此,僅附上會議論文,請參考。
附
件
A Multi-strategy Approach to Protein Structural Alphabet Design
Shih-Yen Ku and Yuh-Jyh Hu
Department of Computer Science
National Chiao Tung University
{gis92622,yhu}@cis.nctu.edu.tw
Abstract
The search for structural similarity among proteins can provide valuable insights into their functional mechanisms and their functional relationships. Though the protein 1D sequence contains the information of protein folding, the performance of predicting the 3D-structure directly from the sequence is still limited. As the increase of available protein structures, we can now conduct more precise and thorough studies of protein structures. Among many is the design of protein structural alphabet that can characterize protein local structures. We use the self-organizing map combined with the minimum spanning tree algorithm for visualization to determine the alphabet size and then apply the k-means algorithm to group protein fragments into clusters corresponding to the structural alphabet. The intra-cluster and inter-cluster analyses show the significant structural cohesiveness. A comparative study of our alphabet with one of the recently developed structural alphabets also demonstrated a competitive result.
1. Introduction
Various genome sequencing projects have been producing numerous linear amino acid sequences; however, complete understanding of the biological roles played by these proteins requires knowledge of their structures and functions [1]. Despite that experimental structure determination methods provide reasonable structure information regarding subsets of proteins, computational methods are still required to provide valuable information for a large fraction of proteins whose statures may not be experimentally determined. Even though the primary sequence implies the whole information guiding the protein folding, yet the performance of predicting the 3D-structure directly from the sequence is still limited. The complexity and the number of physicochemical, kinetic and dynamic parameters involved in protein folding prohibit an efficient 3D-structure prediction without first knowing the 3D-structures of closely related proteins [2]. Some
ab initio methods do not directly use 3D-structures, but their applications are often limited to small proteins [3].
Early analysis of protein structures has shown the importance of repetitive secondary structures, i.e. -helix and -sheet. With variable coils, they constituted a basic standard 3-letter alphabet, and this has led to early secondary structure prediction algorithms, e.g. GOR [4], and more recent ones that apply neural networks and homology sequences [5-8] with prediction accuracy approaching 80%. In spite of the increase of predictive accuracy, the approximation of 3D-structures with only a 3-letter alphabet is apparently too crude for meaningful 3D reconstruction. All the predictions are highly dependent on the definitions of periodic structures, but unfortunately the structure description is incomplete. As the increase of available protein structures, it allows more precise and thorough studies of protein structures. Various more complex structural alphabets have been developed by taking into account the heterogeneity of backbone protein structures through sets of small protein fragments frequently observed in different protein structure databases [2][9]. The alphabet size can vary from several to around 100. For example, Unger et al. [10] and Schuchhardt et al. [11] used k-means method and self-organizing maps respectively to identify the most common folds, but the large number of clusters (about 100) is not appropriate for prediction. Rooman et al. found 16 recurrent folding motifs, ranging from 4 to 7 residues and categorized into four classes corresponding to-helix, -strand, turn and coil [12]. By applying autoassociative neural networks, Fetrow et al. defined six clusters representing supersecondary structures that subsume the classic secondary structures [13]. Bystroff and Baker produced similar short folds of different lengths and grouped them into 13 clusters for prediction [14]. Taking into account the Markovian dependence, Camproux et al. developed an HMM approach to lean the geometry of the structural alphabet letters and the local rules for assembly process [15].
In this paper, we propose a multi-strategy approach to identifying structural alphabet that can characterize protein local structures. Instead of applying
cross-validation [14] or shrinking procedures [16] to refine the clusters directly, we use self-organizing maps as a visualization tool to determine the size of structural alphabet. Given the alphabet size, we later apply the k-means algorithm [17] to group protein fragments into clusters that correspond to a structural alphabet. The analysis of structural similarities between proteins not only provides significant insight into functional mechanisms and biological relationships, but also offers the basis for protein fold classification. An expressive structural alphabet can allow us to quantify the similarities among proteins encoded in appropriate letters. It also enables us to work with a primary representation of 3D structures, simply using standard 1D amino acid sequence alignment methods. To demonstrate the performance of our new method, we tested it on the all- proteins in SCOP. The experimental results show that using our structural alphabet rather than the standard amino acid letters can outperform BLAST in finding the best hit for a protein query. This suggests that our structural alphabet can successfully reflect protein structural characteristics which are implied in protein fragments. Besides, in order to make a consistent and fair comparison, we also compared our alphabet with others that are also developed by the SOM, but in a different design methodology [9][19]. Our structural alphabet shows competitive performance in protein matching.
2. Material and methods
The use of frequent local structural motifs embedded in polypeptide backbone has recently shown improvement in protein structure prediction [1][14][18]. Its success has shed some light on further studies of structural alphabet. We used the proteins classified to all-fold within the SCOP database (version 1.65) in our study with the aim to build the structural alphabet suitable for all-proteins. The same approach can be easily applied to other databanks as well.
There are three issues addressed in our study. They are: (1) protein fragment representation, (2) alphabet size determination and (3) structural alphabet definition. Like others, we transform each protein backbone into a series of the dihedral angles (and , neglecting ) [14][16]. Adapted from [16], the analysis is limited to fragments of five residues since they are adequate for describing a short helix and a minimal structure. With the fixed window size of five residues, we slid the window along each all-protein in SCOP, advancing one position in the sequence for each fragment, and collected a set of overlapped 5-residue fragments. As
the relation between two successive carbons,
i
C
and1 i
C
, located at the ith and (i+1)th positions, can be defined by the dihedral angles iofC
i andi+1 of1 i
C
, a fragment of L residues can then be defined as a vector of 2(L-1) elements. Thus, in our study, each protein fragment, associated with -carbons2 i
C
, 1 iC
, iC
, 1 iC
and 2 iC
, is represented by a vector of eight dihedral angles, i.e.].
,
,
,
,
,
,
,
[
i2
i1
i1
i
i
i1
i1
i2 Based onthis representation, we totally gathered 1,143,072 fragment vectors.
Self-organizing maps (SOM) are widely used as a data mining and visualization tool for complex data sets. A self-organizing map usually consists of a regular 2D grid of so-called map units, each of which is described by a reference vector mi= [mi1, mi2, mi3,…, mid], where d is the input vector dimension, e.g., d = 8, in our case of fragment vectors. The map units are usually arranged in a rectangular or hexagonal configuration. The number of units affects the generalization capabilities of the SOM, and thus is often specified by the researcher/user. It can vary from a few dozen to several thousands. An SOM is a mapping from the ensemble of input data vectors (Xi=[xi1, xi2, xi3,…, xid]Rd) to a 2D array of map units. During training, data points near each other in input space are mapped onto nearby map units to preserve the topology of the input space [19][20]. The SOM is trained iteratively. In each training step t, distances between a randomly picked input vector xjand all the reference vectors are computed. The unit with the least distance is then selected as the winner unit and denoted by w. The winner unit and its topological neighbors are updated to move closer to input vector xjin the input space by the following rule:
)
(
)
(
)
(
)
(
)
1
(
t
m
t
t
h
t
x
m
t
m
i
i
wi j
iwhere t is time, (t) is the adaptation coefficient, |xj-mi(t)| is the component-wise difference between the input vector and the ith reference vector, and hwi(t) is the neighborhood function acting on the array of units, whose form includes bubble kernel, Gaussian kernel and other more complicated ones. In our study, we used the bubble kernel [20][21]. Unlike previous works that directly apply SOM to obtain clusters of backbone fragments as the basis to define the structural alphabet, our approach instead uses SOM only for the visualization purpose to predetermine the number of letters in the alphabet.
By visual inspection of the trained SOM, we can get a preliminary idea of the number of clusters on the map. The unified distance matrix (U-matrix) is one of the most widely used methods for visualizing the clustering result on the SOM. It shows distances between neighboring reference vectors, and can be efficiently visualized using grey shade [22], as shown in Figure 1(a). In spite of the initial idea of the cluster structure provided by the U-matrix, a systematic method to determine the number of clusters on the map is still desired. We implement a post-process on the U-matrix that is based on the minimum-spanning-tree algorithm. Given the grey levels in the U-matrix, we can build the minimum spanning tree for all the map units, e.g., in Figure 1(b), all map unit are linked in the spanning tree. Based on a threshold of the grey level, we can partition the entire tree into several disconnected subtrees, by removing the links between map units with grey levels below the threshold, as shown in Figure 1(c). Conceptually, it means that we break the links of a distance longer than some threshold. Furthermore, those relatively smaller subtrees left can be also deleted later such that the remaining clusters can maintain a reasonable size, as presented in Figure 1(d). The number of the subtrees finally kept becomes the structural alphabet size. As the SOM can be viewed as a topology preserving mapping from input space onto the 2D grid of map units [19], the number of map units can affect the clustering result. We systematically increase the number of units, and repeat the above process till the alphabet size stabilizes.
Rather than adapt the two-level approach that first trains the SOM, then performs clustering of the trained SOM [19], after determining the alphabet size, we apply the k-means algorithm to the input data vectors directly to obtain the clusters. The SOM established a local order among the set of reference vectors in such a way that the closeness between two reference vectors in the Rd space is dependent on how close the corresponding map units are in the 2D array. Nevertheless, an inductive bias of this kind may not be appropriate for structural alphabets since the local order does not always faithfully characterize the relation between structural building blocks, and can sometimes be misleading, e.g. forcing the topology to preserve mapping from the input space of-helix and -strand to a 2D grid of units could be harmful to clustering. As a result, we use the SOM only for visualization the alphabet size, and rely on the k-mean algorithm to extract the local features from the input data that can actually reflect the characteristics of the clusters respectively. The centroid of each cluster forms the prototypical representation of each alphabet
letter. Given the clustering result by the k-means algorithm as the basis of the structural alphabet, we can transform a protein into a series of the alphabet letters by matching each of its fragments against our alphabet prototypes. The control flow of our system named SMK is illustrated in Figure 2.
(a) (b)
(c) (d)
Figure 1. Visualization of the trained SOM. (a) the grey shade of the trained SOM, where darker areas mean larger distances, (b) the minimum spanning tree for the map units, (c) the disconnected subtrees after removing the links below some threshold and (d) the final disconnected subtrees after discarding those relatively small ones.
3. Experimental results
We tested our approach on the all-proteins in SCOP. By this experiment, we show that our method can produce an appropriate structural alphabet for describing these all- proteins. After transforming protein backbones into dihedral angles and extracting protein fragments, we trained the SOM on these dihedral angle vectors.
Three issues were addressed in the experiments. First, the meaningfulness of the structural alphabet size in terms of the number of clusters was presented by showing the size stability given various parameters. Second, we demonstrated cluster cohesiveness by visual superimpositions of protein fragments as well as computed the intra-cluster and inter-cluster distance. Third, we proved the fragment clusters found were not arbitrary by comparing our result with that from a random background.
Figure 2. The system control flow of SMK
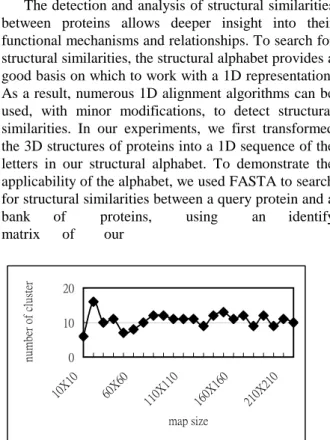
Since the number of map units has influence over the SOM’s clustering behavior, to obtain the optimal number of clusters, we varied the number of units on the map until the number of clusters found became steady. The results are shown in Figure 3, which indicates a distinctive plateau within the range between nine and twelve. Because eleven is the most frequent number of clusters on the plateau, as shown in Figure 4, it is designated as the structural alphabet size.
To further confirm the general geometric regularities characterized by the structural alphabet, we also built a negative all- protein fragment set for comparison. The negative set was derived from the real all- protein fragment vectors prepared earlier by rotating the dihedral angles at random (increase or decrease) within a certain degree, e.g. 30in our analysis. We compared the clusters produced by
clustering on the real vector set and on the negative control set. Insignificant difference suggests that the alphabet we found could be arbitrary. Our experiments (see Figure 5) show that clustering on the negative control set cannot even produce consistent clusters, which supports our hypothesis that the clusters found from the real fragment vectors reflect the classes of local protein structures; otherwise, these clustering results would have been similar.
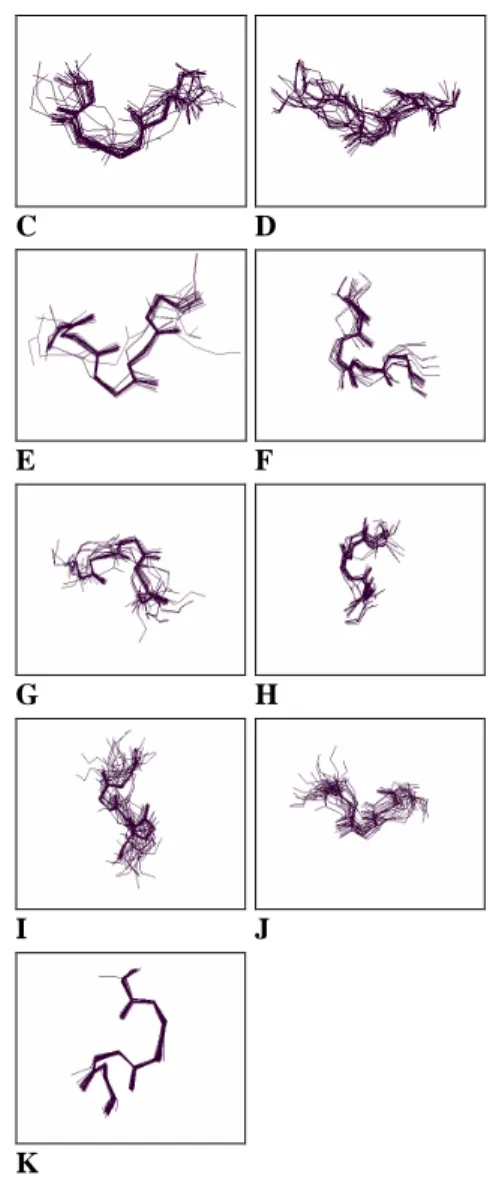
Given the size, we ran the k-means algorithm on the input fragment vectors to find the twelve clusters by which to define the structural alphabet. Figure 6(a) and (b) shows the fragment superimpositions for the alphabet. Even though the fragment structures do not superimpose perfectly, yet the general structural cohesiveness of each category is quite evident. In addition, we computed the Euclidean distances from each fragment in a given cluster to its centroid. The average of these within-cluster distances was then compared with the center-to-center distances between clusters as presented in Table 1. It shows that in most cases, the center-to-center distance between any two clusters is greater than the mean distance of all vectors in that cluster from its center plus one standard deviation. The result indicates that the individual clusters are fairly well separated from each other.
The detection and analysis of structural similarities between proteins allows deeper insight into their functional mechanisms and relationships. To search for structural similarities, the structural alphabet provides a good basis on which to work with a 1D representation. As a result, numerous 1D alignment algorithms can be used, with minor modifications, to detect structural similarities. In our experiments, we first transformed the 3D structures of proteins into a 1D sequence of the letters in our structural alphabet. To demonstrate the applicability of the alphabet, we used FASTA to search for structural similarities between a query protein and a bank of proteins, using an identify matrix of our 0 10 20 10X1 0 60X6 0 110X 110 160X 160 210X 210 map size nu m be r of cl us te r
Figure 3. The variance in the number of clusters produced by the SOMs of varying sizes. There exists a Backbone Transformation into
Dihedral Angles
Protein Fragment Vectors Extraction as Input to SOM, i.e.
].
,
,
,
,
,
,
,
[
i2
i1
i1
i
i
i1
i1
i2Train SOM on Protein Fragment Vectors
Visualizing trained
Build Minimum Spanning Tree from U-matrix
Partition Minimum Spanning Tree into Disconnected Subtrees
Use number of subtrees as K and Run K-mean Algorithm on Input Vectors
Define Structural Alphabet based on K-means Clusters
Transform Proteins into Structural Alphabet
distinctive plateau that suggests the cluster number has stabilized. 0 2 4 6 8 1 6 11 16 21 number of cluster fr en qu en cy
Figure 4. The frequencies of cluster numbers. It shows 11 is the most frequent number of clusters.
0 10000 20000 30000 40000 10X1 0 60X6 0 110X 110 160X 160 map size nu m be r of cl us te r
Figure 5. The variance in the number of clusters produced by the SOMs of varying sizes trained on a negative fragment set. It shows no sign of convergent cluster number.
alphabet letters to find maximal exact matches. For comparison, we also conducted the same tests also using FASTA but based on different structural alphabets, one developed by de Brevern et al. [9], the other by the two-level SOM approach [19]. As the baseline reference, we used BLAST with the standard 20 amino acid letters to find the best sequence hit.
The proteins used in the experiments were selected from the all-proteins in SCOP. After filtering out those with more than 30% sequence similarity, we have totally 1055 proteins. For each run of the experiment, we randomly picked one protein as the query, and then matched it against the rest, using FASTA or BLAST with different alphabets. Given the best hit, we computed the RMSD between the query and the hit,
Table 1. Summary of within-cluster distances and center-to-center distances. K J I H G F E D C B A 4 3 .1 5 ± 5 0 .1 3 8 8 .7 7 ± 5 3 .3 3 1 9 6 .7 5 ± 9 7 .2 1 5 5 .0 2 ± 7 7 .8 2 2 0 .5 2 ± 8 7 .7 9 1 4 3 .6 2 ± 9 0 .8 4 1 5 0 .4 1 ± 7 1 .5 3 1 9 3 .6 7 ± 6 9 .1 9 1 7 3 .5 8 ± 7 7 .5 9 1 9 2 .8 4 ± 7 4 .9 7 1 8 6 .1 0 ± 6 8 .0 7 m ea n ± sd w it h in -c lu st er 0 A 0 2 8 2 .3 B 0 28 4 .5 9 2 0 5 .2 7 C 0 25 0 .3 1 2 0 3 .4 1 2 1 6 .7 5 D 0 2 3 4 .3 1 2 5 1 .6 2 0 2 .8 2 2 6 .9 3 E 0 30 2 .9 3 2 5 2 .0 5 1 9 7 .7 6 2 7 5 .0 8 2 3 6 .7 2 F 0 3 4 6 .1 4 5 1 1 .0 4 3 8 8 .9 3 8 3 .8 6 4 1 4 .9 9 3 9 9 .5 3 G 0 3 4 3 .0 7 2 2 0 .6 3 2 8 4 .5 1 2 6 1 .9 2 4 3 .0 2 1 6 9 .3 2 4 6 .5 H 0 33 5 .8 4 2 7 6 .0 3 3 4 6 .9 8 3 4 3 .0 2 3 2 3 .8 1 3 3 3 .4 1 3 2 1 .0 3 3 2 5 .9 4 I 0 3 5 8 .5 8 1 3 6 .6 3 3 4 1 .2 2 1 6 1 .1 1 2 8 2 .1 9 1 8 3 .7 7 1 8 8 2 0 8 .2 8 1 9 7 .4 4 J 0 8 6 .7 1 1 3 6 0 .9 5 1 6 4 .8 7 2 7 8 .5 1 7 7 .4 8 3 5 8 .4 8 2 3 3 .3 3 2 2 6 .5 2 2 6 4 .6 9 2 4 5 .8 1 K ce n te r-to -c en te r
and recorded the lowest level in the SCOP hierarchy at which the query and the hit are both located, i.e. class, fold, superfamily or family. Smaller RMSD and lower common level in SCOP hierarchy indicates higher structural similarity. We repeated the same experiment for 100 times and the results are summarized in Table 2 and 3. According to Table 2, we notice that our method SMK and de Brevern et al.’s both produced higher frequencies at lower common levels than the other two methods. This suggests that our structural alphabet and de Brevern et al.’s can better characterize the SCOP hierarchy. Table 3 shows that SMK has the lowest mean RMSD and standard deviation among all.
Table 2. Summary of frequencies at the lowest common level. The first column shows the methods used in the experiments. The remaining columns present the frequency for different levels at which the query and the best hit are both located.
frequency at different level
Method class fold super family family BLAST 71 4 5 20 SMK 55 11 5 29 de Brevern 58 4 11 27 2-level SOM 73 6 14 7
Table 3. Summary of average RMSD and standard deviation between the queries and the best hits.
method mean (RMSD) sd (RMSD) BLAST 8.953744 4.764597 SMK 7.290972 3.934283 de Brevern 8.076746 4.819178 2-level SOM 10.38624 5.217078
4. Discussion
In this paper, we propose a multi-strategy approach to designing the structural alphabet which allows local approximation of protein 3D structures as well as enables the applications of 1D alignment algorithms to search for 3D structural similarities. The success of the alphabet design depends on three crucial factors. First, it is the protein fragment representation, which determines what and how 3D structural characteristics to be approximated, e.g. thermodynamic stability, amino acid physicochemical properties, amino acid usage in known proteins, distances, dihedral angles, bond lengths, bond angles, etc.
A B C D E F G H I J K
Figure 6(a). The superimposition in wireframe format for the structures of each structural cluster found by SMK.
A B
E F
G H
I J
K
Figure 6(b). The superimposition of the structures of each structural cluster found by SMK in the ball-and-stick form.
The effects of the representation selected are entangled with the performance of the learning approach we apply to develop the structural alphabet. Overcomplicated representations can sometimes lead to overfitting. To avoid this problem, we currently focus on the dihedral angles. Other features can be easily included in the representation if proved necessary.
The second factor is the size of the alphabet. We took advantage of the SOM as a visualization tool that helps determine the alphabet size. By systematically varying the number of map units on the map, we visualized the clustering behavior of the SOM. Our experiments showed a distinct plateau corresponding to the convergent number of clusters, compared with the increasing number of clusters in the results of clustering on the random negative control dataset. This suggests that the structural alphabet size we found is not arbitrary.
Various types of algorithms have been applied to clustering local protein 3D fragments into a limited set of fold patters, e.g. self-organizing maps (SOM), hidden Markov models (HMM), neural networks, hierarchical clustering, k-means clustering, etc. Each
has its own learning bias and inherent limitations. For example, the topology (e.g. number of layers or map units) of neural networks, the SOM and the HMM strongly affect the performance. The value of k in k-means algorithm determines the clusters. As a consequence, the third factor is the learning algorithm. In our study, we took a multi-strategy approach. We first used the SOM and the minimum-spanning tree algorithm to determine the alphabet size, and then applied the k-means algorithm to group fragments into meaningful clusters. The number of map units in the SOM and the value of k in k-means are not pre-specified in advance, but instead determined systematically. To verify the correspondence of our structural alphabet letter to the fold patterns, we computed the average within-cluster distance for each alphabet cluster as well as the distance across clusters. The small average within-cluster distance and the relatively large between-cluster distance demonstrate the significance of the structural alphabet we found. Furthermore, the visualized superimposition of protein fragments in each cluster also justifies the structural cohesiveness.
The objective of the paper is to propose a new approach to developing the structural alphabet. To verify its usefulness, we tested it on the all-proteins in SCOP, and the experimental results show its promising applicability. After the success on the all- proteins in SCOP, we plan to test our method on different data banks to further verify its feasibility and generality. Also as mentioned above, the representation is a crucial factor in the alphabet design. We will consider other structural features besides dihedral angles, add more useful features to enhance our structural alphabet, and test the new approach on other families in SCOP.
5. References
[1] D. Baker and A. Sali, “Protein Structure Prediction and Structural Genomics”, Science, vol. 294, 2001, pp. 93-96.
[2] A.G. de Brevern and S.A. Hazout, “Hybrid Protein Model(HPM): a method to compact protein 3D-structure information and physicochemical properties”, IEEE Comp. Soc. S1, 2000, pp. 49-54. [3] C.A. Orengo, J.E. Bray, T. Hubbard, L. LoConte and I. Sillitoe “Analysis and assessment of ab initio three-dimensional prediction, secondary structure, and contacts prediction”, Protein, vol. 37, 1999, pp. 149-170.
“Analysisoftheaccuracy and implicationsofsimple methods for predicting the secondary structure of globularprotein”, Journal of Molecular Biology, vol. 120, 1978, pp. 97-120.
[5] B. Rost and C. Snader, “Prediction of protein secondary structure at better than 70% accuracy”, Journal of Molecular Biology, vol. 232, 1993, pp. 584-599.
[6] A. Salamov and V. Solovyev, “Protein secondary structureprediction using localalignments”, Journal of Molecular Biology, vol. 268, 1997, pp. 31-36.
[7] T.N. Petersen, C. Lundegaard, M. Nielsen, H. Bohr, J. Bohr, S. Brunak, G.P. Gippert and O. Lund, ”Prediction of protein secondary structure at 80% accuracy”, Proteins, vol. 41, 2000, pp. 17-20. [8] B. Rost, “Review: Protein secondary structure prediction continues to rise,” Journal of Structural Biology, vol. 134, 2001, pp. 204-218.
[9] A.G. de Brevern, H. Valadie, S.A. Hazout and C. Etchebest,“Extension ofalocalbackbonedescription using a structural alphabet: A new approach to the sequence-structure relationship,”Protein Science, vol. 11, 2002, pp. 2871-2886.
[10] R. Unger, D. Harel, S. Wherland and J.L. Sussman, “A 3D building blocks approach to analyzing and predicting structure of proteins”, Proteins, vol. 5, 1989, pp. 355-373.
[11] J. Schuchhardt, G. Schneider, J. Reichelt, D. Schomburg and P. Wrede, “Local structural motifs of protein backbones are classified by self-organizing neural networks”, Protein Engineering, vol. 9, 1996, pp. 833-842.
[12] M.J. Rooman, J. Rodriguez and S.J. Wodak, “Automatic definition of recurrent local structure motifs in proteins”, Journal of Molecular Biology, vol. 213, 1990, pp. 327-336.
[13] J.S. Fetrow, M.J. Palumbo and G. Berg, “Patterns, structures, and amino acid frequencies in structural building blocks, a protein secondary structure classification scheme”, Proteins, vol. 27, 1997, pp. 249-271.
[14] C. Bystroff and D. Baker, “Prediction of local structure in proteins using a library of sequence-structure motif”, Journal of Molecular Biology, vol. 281, 1998, pp. 565-577.
[15] A.C. Camproux, R. Gautier and P. Tuffery, “A hidden Markov model derived structural alphabet for proteins”, Journal of Molecular Biology, doi: 10.1016/j.jmb.2004.04.005.
[16] A.G. de Brevern, C. Etchebest and S. Hazout, “Bayesian probabilistic approach for predicting backbone structures in terms of protein blocks”, Proteins, vol. 41, 2000, pp. 271-287.
[17] J.A. Hartigam and M.A. Wong, “A k-means clustering algorithm”, Applied Statistics, vol. 28, 1975, pp. 100-108.
[18] K.T. Simons, I. Ruczinski, C. Kooperberg, B.A. Fox, C. Bystroff and D. Baker, “Improved recognition of native-like protein structures using a combination of sequence-dependent and sequence-independent features of proteins”, Proteins, vol. 34, 1999, pp. 82-95.
[19] J. Vesanto and E. Alhoniemi, “Cluster of the self-organizing map”, IEEE trans. Neural Networks, vol. 11, 2000, pp. 586-600.
[20] T. Kohonen, “Self-organizing Maps”, Berlin/Heidelberg, Germany; Springer, Vol. 30, 1995. [21] P. Tamayo, D. Slonim, J. Mesirov, Q. Zhu, S. Kitareewan, E. Dmitrovsky, E.S. Lander and T.R. Golub, “Interpreting patterns of gene expression with self-organizing maps: Methods and applications to hematopoietic differentiation”, PNAS, vol. 96, 1999, pp. 2907-2912.
[22] J. Iivarinen, T. Kohonen, J. Kangas and S. Kaski, ”Visualizing theclusters on the self-organizing map”, in Proc. Conf. Artificial Intelligence Research Finland, 1994, pp. 122-126.