行政院國家科學委員會專題研究計畫成果報告
長鏈不飽和脂肪酸對轉錄因子 ADD1 和脂肪酸合成之影響機制
The mechanism by which long chain polyunsaturated fatty acids
]affect ADD1 and fatty acid synthesis
計畫編號:NSC 90-2313-B-002-304
執行期限:90 年 8 月 1 日至 91 年 7 月 31 日
主持人:丁詩同 國立台灣大學畜產學系
計畫參與人員:劉秉勳 國立台灣大學畜產學系
一、中文摘要
飼糧添加脂肪除提供能量等基本功能
外,脂肪酸的變化更可以改變組織中脂肪酸
的組成,調節各組織的生理代謝。在豬方面,
其脂肪之生合成與堆積,主要發生在脂肪組
織,並且調控此相關代謝之轉錄因子-脂肪細
胞 分 化 決 定 因 子 一
(ADD1) 也 主 要 表 達 於
此。許多試驗指出,多不飽和脂肪酸(PUFA)
可藉由抑制
ADD1 之表現,減少脂肪細胞堆
積脂肪。本研究試驗中發現,二十二碳六烯
酸(DHA)抑制 ADD1 表現,而抗氧化劑的添
加(400 nM N,N’-diphenyl-1,4-diphenylenediamine, 15
M (+)--tocopherol acid succinate, 600 nM butylated hydroxytoluene, and 160 nM deferoxamine mesylate)可以彌補此抑制效果,顯示
DHA 對 ADD1 表
現的調節與脂肪酸的氧化作用有關。而在抗
氧化基因與類前列腺素的試驗中,我們無法
進一步證明多不飽和脂肪酸抑制
ADD1 之表
現只經由脂肪酸的過氧化機制。簡言之,我
們可知
PUFA 抑制 ADD1 mRNA 的表現,以
達到抑制脂肪細胞內脂肪之堆積,惟其所產
生之類前列腺素代謝產物對
ADD1 影響機制
待尚進一步之試驗證明。
關鍵詞:
ADD1(脂肪細胞分化決定因子 1),
FAS(脂肪酸合成? ),PUFA(多不飽和脂
肪酸)
Abstract
Supplementation of dietary fat provides
energy for animals. Fatty acid composition in
many tissues of animals can be modified by
dietary fat sources that have different fatty acid
(FA) composition. Such a modification may
regulate some physiological functions. In pigs,
the FA de novo synthesis and fat accumulation
is primarily located in adipose tissues. The
adipocyte determination and differentiation
factor 1(ADD1) expressed highly in adipose
tissue. It has been shown that polyunsaturated
fatty acid (PUFA) reduced fat deposition in
adipocyte by inhibiting ADD1 expression.
Current research showed that docosahexaenoic
acid (DHA) inhibited ADD1 expression. The
inhibition could be recovered by adding
antioxidant mixture
( 400 nM N,N’-diphenyl- 1,4-diphenylenediamine, 15 M (+)--tocopherol acid succinate, 600 nM butylated hydroxytoluene, and 160 nM deferoxamine mesylate ), indicating the
inhibitory effect of DHA to the expression of
ADD1 was partially due to PUFA peroxidation,
however. We could not further prove whether
the inhibition of ADD1 expression by PUFA is
solely via fatty acids peroxidation mechanism.
In conclusion, PUFA inhibit fat deposition in
adipocyte by reducing ADD1 mRNA
expression, but the detail mechanism remains
to be determined.
二、緣由與目的
脂肪可被添加於家禽和家畜的飼糧中以
提供能量、減少飛塵和增加適口性,飼糧中
脂肪酸的變化更可以改變組織中脂肪酸的組
成。飼糧中脂肪酸可被動物利用而貯存於各
種組織,餵飼含不同脂肪酸的脂質來源可顯
著改變豬隻脂肪組織、肌肉組織和肝臟中脂
肪酸的組成(Villegas et al., 1973; Otten et al.,
1993; Innis et al., 1996)。鄭等(2000) 發現餵
飼 3 週齡仔豬一含 10 % 牛油或 10 % 魚油
的飼糧可顯著改變豬隻脂肪組織、肌肉組織
和肝臟中脂肪酸的組成,隨著魚油的添加-3
脂肪酸顯著增加。這些結果顯示飼糧中的長
鏈多不飽和脂肪酸(PUFA)可被豬隻利用而貯
存於各種組織,並可能因此而改變各組織的
生理代謝。近年來,許多研究報告指出,
PUFA
的添加處理,會降低細胞內脂肪之堆積,其
主要之作用機制有二。其一,藉由增加轉錄
因子過氧小體增生活化受體
α(PPARα)
之表
現,間接促進細胞內氧化及生熱作用;其二,
PUFA 之添加,抑制細胞內許多脂肪生合成
基因之表達,如:脂肪合成?
(FAS),而其調
控主要是藉由影響轉錄因子脂肪細胞分化決
定因子一 (ADD1)之表現來達成。
在小鼠的試驗中,Xu et al.(1999)發現,
PUFA 藉由抑制肝臟中轉錄因子 SREBP1 之
表現,減少
FAS 之合成,因而降低肝臟中脂
肪堆積。並且更進一步發現,PUFA 並非影
響
SREBP1 之轉錄,而是增加其 mRNA 之降
解,以達到抑制
SREBP1 之表現(Xu et al.,
1999)。 Foretz et al. (1999)則指出,肝細胞
FAS mRNA 之表現,可能是藉由多不飽和脂
肪 酸 過 氧 化 產 物
( 如 類 前 列 腺 素 衍 生 物 .
Mater et al., 1999)所調控的。在豬方面,不同
於小鼠,其脂肪酸之生合成,主要發生在脂
肪組織,並且
ADD1(與 SREBP1 為同一基因)
也大量表達於此。本實驗室初步之試驗發
現,許多
PUFA 確實可以有效降低豬脂肪細
胞
ADD1 蛋白質及 mRNA 之表現 。
本計畫
之 研 究 重 點 包 括 :
1. 探 討 DHA 對
ADD1 基因表達之控制機制,是否與脂肪
酸之過氧化有關;
2. 測試不同
長鏈多不
飽和
脂肪酸(AA 和 DHA)的過氧化作用及
其對
ADD1, FAS, superoxide dismutase,
和
glutathione peroxidase 表達之影響;3.
探討維生素
E 是否可因其抗氧化性而降
低
DHA 對 ADD1 和 FAS 之影響; 4. 研
究花生四烯酸(AA)經由其代謝產物來影
響
ADD1 和 FAS 表達之機制。經由本計
畫之執行,吾人將更瞭解
長鏈多不飽和
脂
肪酸影響脂肪代謝的分子調控機制,並可
提供做為減少豬隻脂肪堆積之策略模式
的參考
三、結果與討論
試驗一:
DHA 對 ADD1 基因表達之
控制機制,是否與脂肪酸之過氧化有關
。
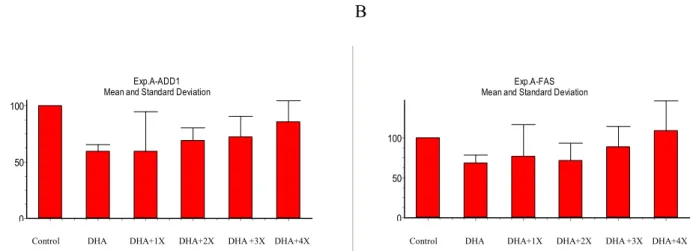
試驗之結果顯示,隨著抗氧化劑(1X:100 nM
N,N’-diphenyl-1,4-diphenylenediamine, 3.75 M (+)-- tocopherol acid succinate, 150 nM butylated hydroxytoluene, and 40 nM deferoxamine mesylate)濃度的增加,部分脂肪細胞基因表現有回復的
趨勢。加入
4X 濃度的抗氧化劑,即可使濃
度
100 μM DHA 所抑制表現的 ADD1 由
59.5%提升至 85.5%,轉錄因子 ADD1 下游
調控基因
FAS 更可由 68.9%回復達 109.2
%,唯再提高抗氧化劑之濃度,亦會影響脂
肪細胞本身之正常生長,顯示細胞仍需要適
度的氧化還原反應,以維持正常細胞生長機
能。ADD1 mRNA 之表現在 DHA 處理組與
DHA + 1X 抗氧化劑處理組各有顯著差異(P
<0.05),顯示添加 2X 以上之抗氧化劑處理
組,確實能彌補
DHA 抑制 ADD1 表現之效
果。前人研究已證明
DHA 在 25μM 時之抑
制程度,甚至低於
AA 及 CLA 於 100μM 時
之影響(NSC 89-2316-B-002- 037),因此本
實驗除了顯示添加
100 μM DHA 可藉由抗
氧化劑的添加來回復
ADD1 的抑制效果,亦
證明
DHA 對 ADD1 基因調控之機制的確與
脂肪酸之過氧化有關。
試 驗 二 :
不 同
長 鏈 多 不 飽 和
脂 肪 酸
( AA 和 DHA ) 的 過 氧 化 作 用 及 其 對
superoxide
dismutase
(SOD),
和
glutathione peroxidase (GPX)表達之影
響
。試驗處理中,使用
100 μM AA 或 100
μM DHA,以充分表現 PUFA 對基因的影響
效果,並參考試驗
A 之經驗,添加抗氧化劑
最高為
4X。
在
此
試驗中發現,不論添加
AA 或 DHA
並不能影響抗氧化基因
SOD1 與 GPX 的表
現, PUFA 的添加似不能增加抗氧化基因之
表現,本試驗無法證明抗氧化劑的添加是否
能緩和抗氧化基因的表達。
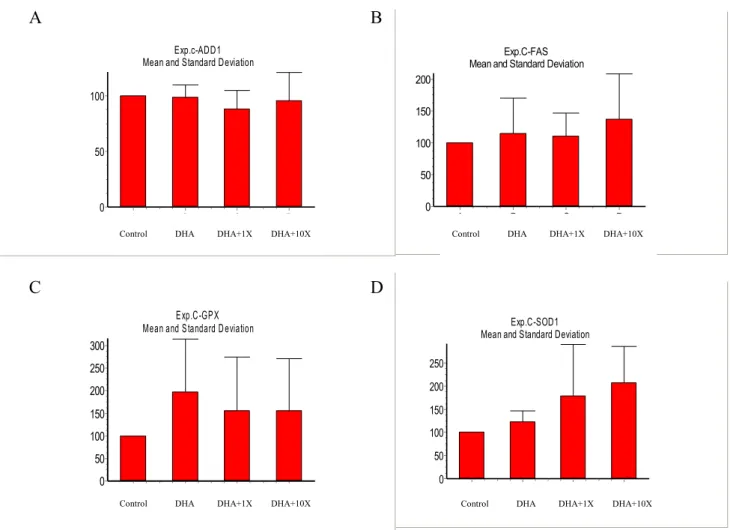
試驗三:
維生素
E 是否可因其抗氧化
性而降低
DHA 對 ADD1 和 FAS 之影響。
此
試驗處理中,主要將抗氧化劑混和物改為
0.75μM 與 7.5μM 的維生素 E。
維生素
E 為豬隻血液中常見之天然抗過
氧化物,本試驗之目的在探索維生素
E 是否
具有減緩
DHA 抑制 ADD1 與 FAS 表現之能
力,並期望抗氧化基因亦能在此試驗中藉由
維生素
E 的添加而減少原本因 DHA 的刺激
效果,以確立
DHA 過氧化機制與 ADD1 和
FAS 的影響。本試驗結果除確定添加正常生
理質之維生素
E(0.75μM)與 10X 生理質
維生素
E 可不影響正常脂肪前身細胞生長
外,高於生理質
100X 之維生素 E 亦如試驗
一添加
20X 抗氧化劑的細胞一般無法正常生
長分化。
試驗四:
AA 經由其代謝產物來影響
ADD1 和 FAS 表達之機制
。本試驗目的在
於添加各種類前列腺素生合成路徑之抑制物
(
cycloogenase; indomethacin, lipoxygenase;
ETYA, monooxygenase; clotrimazole.),以探
討花生四烯酸在脂肪細胞抑制
ADD1 與 FAS
表現之可能過氧化途徑。前人研究指出,
AA
可在細胞內代謝成各種類前列腺素,並可能
以自分泌或旁分泌的方式影響脂質合成基因
之表現,顯示類前列腺素衍生物的生成是
PUFA 抑制脂肪合成基因表達的一種可能機
制,唯其試驗結果諸如試驗二與試驗三,並
無法推論類前列腺素的生成是否對豬隻脂肪
細胞造成影響。
四、計畫成果自評
本計畫之研究結果,證明藉由外源性添
加
DHA 可以顯著地降低豬脂肪細胞 ADD1
的表達,額外加入抗氧化劑則可彌補此抑制
效果,顯示
PUFA 抑制豬隻脂肪細胞 ADD1
之表現的確與脂肪酸過氧化機制有關,後續
詳細機制的探討則需要再次重複試驗證明。
本計畫最後之成果將與前人之相關研究成果
合併發表,這些結果亦將是研究生劉秉勳之
碩士論文的一部份。
五、參考文獻
Foretz, M., F. Foufelle, and P. Ferre. 1999. Polyunsaturated fatty acids inhibit fatty acid synthase and spot-14- protein gene expression in cultured rat hepatocytes by a peroxidative mechanism. Biochem J. 341:371-6.
Innis, S. M., R. Dyer, P. T. Quinlan, and D. Diersen- Schade. 1996. Dietary triacylglycerol structure and saturated fat alter plasma and tissue fatty acids in piglets. Lipids. 31:497-505.
Mater, K.M, D.A. Pan, W.G. Bergin, and D.B. Jump. 1998. Arachidonic acid inhibits of hepatic lipogenic gene expression in 3T3-L1 adipocyte through a prostaglandin pathway. J. Lipid Res. 39:1327-1334. Otten, W., C. Wirth, P. A. Iaizzo, and H. M. Eichinger.
1993. A high omega 3 fatty acid diet alters fatty acid composition of heart, liver, kidney, adipose tissue and skeletal muscle in swine. Ann. Nutr. Metab. 37:134-41.
Villegas, F. J., H. B. Hedrick, T. L. Veum, K. L. McFate, and M. E. Bailey. 1973. Effect of diet and breed on fatty acid composition of porcine adipose tissue. J. Anim. Sci. 36:663-668.
Xu, J., M. T. Nakamura, H. P. Cho, and S. D. Clarke. 1999. Sterol regulatory element binding protein-1 expression is suppressed by dietary polyunsaturated fatty acids. A mechanism for the coordinate suppression of lipogenic genes by polyunsaturated fats. J Biol Chem. 274:23577-83.
Exp.A-FAS Mean and Standard Deviation
Column A B C D E F 100 50 0 Exp.B-FAS Mean and Standard Deviation
Column A B C D E F 150 100 50 0 Exp.B-SOD1 Mean and Standard Deviation
Column A B C D E F 250 200 150 100 50 0 Exp.A-ADD1 Mean and Standard Deviation
Column
A B C D E F
100
50
0
Fig. 1. ADD1 and FAS mRNA expression in pig S/V cells culture with or without antioxidants. First, pig S/V cells cultured to conflunce in 24 h, then treated with differentiation medium for 24 h. After above 48 h,cells treated with differentiation medium containing 100 M of DHA for 24 h. 1X antioxidant mixtures:100 nM N,N’-diphenyl -1,4-diphenylenediamine, 3.75 M (+)-- tocopherol acid succinate, 150 nM butylated hydroxytoluene, and 40 nM deferoxamine mesylate. Total RNA was extracted from cultured cells. ADD1(A) and FAS(B) were determined by Northern analysis and mRNA values were normalized to 18S rRNA. n= 4 replicates/treatment.
Exp.B-ADD1 Mean and Standard Deviation
Column A B C D E F 100 50 0 Exp.B-GPX Mean and Standard Deviation
Column
A B C D E F
100
50
0
Fig. 2. PUFA regulate ADD1, FAS, SOD1 and GPX mRNA expression by fatty acid peroxidation. First, pig S/V cells cultured to conflunce in 24 h, then treated with differentiation medium for 24 h. After 48 h culture, cells were changed to differentiation medium containing control(only 1% BSA), 100 M AA and 100 M DHA. 4X antioxidant mixtures(400 nM N,N’-diphenyl-1,4-diphenylenediamine, 15 M (+)--tocopherol acid succinate, 600 nM butylated hydroxytoluene, and 160 nM deferoxamine mesylate) was added to medium. ADD1, FAS, GPX, SOD1 and 18S were determined by Northern analysis(A), (B), (C), (D).The decay of ADD1 and FAS are depicted in B and C, respectively. The mRNA values are relative to 0 h of control and normalized to 18S rRNA. n= 4 replicates/treatment.
A
A
Control DHA DHA+1X DHA+2X DHA +3X DHA+4X Control DHA DHA+1X DHA+2X DHA +3X DHA+4X
B
Control 4X AA AA+4X DHA DHA+4X Control 4X AA AA+4X DHA DHA+4X
Control 4X AA AA+4X DHA DHA+4X Control 4X AA AA+4X DHA DHA+4X
B
Exp.C-FAS Mean and Standard Deviation
Column A B C D 200 150 100 50 0 Exp.C-SOD1 Mean and Standard Deviation
Column A B C D 250 200 150 100 50 0 Exp.D-ADD1 Mean and Standard Deviation
Column A B C D E F G H 200 150 100 50 0 Exp.c-ADD1
Mean and Standard Deviation
Column A B C D 100 50 0 Exp.C-GPX Mean and Standard Deviation
Column A B C D 300 250 200 150 100 50 0
Fig. 3. Vitamin E and DHA coregulate ADD1, FAS, SOD1 and GPX mRNA expression by fatty acid
peroxidation. First, pig S/V cells cultured to conflunce in 24 h, then treated with differentiation medium for 24 h. After
48 h culture, cells were changed to differentiation medium containing control(only 1% BSA), or 100 M DHA. Vitamin E (1X = 0.75 μM) was added to medium. ADD1, FAS, GPX, SOD1 and 18S were determined by Northern analysis(A), (B), (C), (D).The decay of ADD1 and FAS are depicted in B and C, respectively. The mRNA values are relative to 0 h of control and normalized to 18S rRNA. n= 4 replicates/treatment.
Exp.D-ADD1 Mean and Standard Deviation
Column A B C D E F G H 100 50 0
Fig. 4.AA metabolites have the effect of down regulation the expression of ADD1 and FAS. First,
pig S/V cells cultured to conflunce in 24 h, then treated with differentiation medium for 24 h. After 48 h culture, cells were changed to differentiation medium containing control(only 1% BSA), or 100 M AA. Eicosanoid inhibitors (1C:1 μM clotrimazole, 10C:10 μM clotrimazole, 1i:1 μM indomethacin, 10i:10 μM indomethacin, 1E:1 μM ETYA, 10 μM ETYA.) was added to medium. ADD1, FAS and 18S were determined by Northern analysis(A), (B). The mRNA values are relative to 0 h of control and normalized to 18S rRNA. n= 4 replicates/treatment.
Control DHA DHA+1X DHA+10X Control DHA DHA+1X DHA+10X
Control DHA DHA+1X DHA+10X Control DHA DHA+1X DHA+10X
A
B
C
D
Control AA AA+1C AA+10C AA+1i AA+10i AA+1E AA+10E Control AA AA+1C AA+10C AA+1i AA+10i AA+1E AA+10E