國立交通大學
分子醫學與生物工程研究所
碩士論文
新穎的小分子化合物之抗癌機制研究
Synthetic small molecular inhibitors of the
transcription factor NF-кB suppress cell viability and
induce apoptosis in cancer cells
研究生
:
羅福轅
指導教授 :
梁美智 博士
新穎的小分子化合物之抗癌機制研究
Synthetic small molecular inhibitors of the
transcription factor NF-кB suppress cell viability and
induce apoptosis in cancer cells
研究生
:
羅福轅
Student
:
Lo Fu Yuan
指導教授 :
梁美智 博士
Advisor
: Dr. Mei-Chih Liang
國立交通大學
分子醫學與生物工程研究所碩士班
碩士論文
A Thesis
Submitted to Institute of Molecular Medicine and Bioengineering
National Chiao Tung University
in partial Fulfillment of the Requirements
for the Degree of
Master
in
Molecular Medicine and Bioengineering
August 2012
Hsinchu, Taiwan, Republic of China
中華民國一零一年八月
I
新穎的小分子化合物之抗癌機制研究
摘要
轉譯因子 NF-кB (nuclear factor kappa-light-chain-enhancer of activated B cells) 在許多細胞反應 (如細胞增生、細胞凋亡、免疫及發炎反應) 的調控過程中扮演 重要的角色。研究顯示 NF-кB 調控失常與部分腫瘤細胞的增生有關,且證實透 過抑制 NF-кB 的過度表現,能夠有效地抑制腫瘤細胞的生長,並促進細胞凋亡。 因此,轉譯因子 NF-кB 是一個具潛力的抗癌標靶。交通大學應用化學系孫仲銘 教授的實驗室,以 2-aminofuran-linked-benzimidazole 為核心結構,透過修飾周邊 的官能基而衍生出一系列新穎的小分子化合物。經過篩選實驗,我們發現化合物 #1412 (與其衍生物# 21026),在微摩爾濃度範圍內,具有抗癌的生物活性,且能 夠有效地抑制多種癌症細胞株內 NF-кB 的訊息活化與傳遞。例如,在人類 T-淋 巴癌細胞株 Jurkat 與人類多發性骨髓瘤細胞株 RPMI-8226,#1412 (與其衍生物# 21026)藉由抑制 TNF-所活化的 IкBα 的磷酸化反應及降解反應,進而抑制 NF-кB 成員 p65 的細胞核遷移活動 ( nuclear translocation ),使 NF-кB 無法從細 胞質進入細胞核內進行目標基因的轉錄反應。另外,#1412 (與其衍生物# 21026) 也能夠有效地抑制此二種血液癌症細胞株的增生,並誘導細胞凋亡。總結,此實 驗結果建議新穎化合物#1412 (與其衍生物# 21026) 的抗癌效果與其抑制轉譯因 子 NF-кB 訊息傳遞的能力有關。
II
Synthetic small molecular inhibitors of the
transcription factor NF-кB suppress cell viability and
induce apoptosis in cancer cells
Abstract
Transcriptional factor NF-кB plays a critical role in mediating cellular processes, including cellular growth control, apoptosis, immune and inflammatory responses. Dysregulation of the NF-кB signaling pathway has been reported in a variety of cancer types and inhibition of the constitutive NF-кB activity may have therapeutic applications. In this study, we have identified the synthetic compounds #1412 and #21026, novel derivatives of the 2-aminofuran linked benzimidazole (synthesized in the laboratory of Prof. C.M. Sun in the Department of Applied Chemistry at National Chiao Tung University), as potent NF-кB inhibitors. Compound #1412 does- and time-dependently inhibited TNF-α induced NF-кB activation in the low micromolar range in cultured cells. In the human leukemic T cell Jurkat cell line and multiple myeloma RPMI-8226 cell line, both compounds blocked TNF-α induced IкBα phosphorylation and degradation and nuclear translocation of the NF-кB subunit p65. In addition, both compounds affected cell viability and induced cell apoptosis. Taken together, these results suggest that the anticancer activity of compound #1412 and #21026 in human cancer cells is related to inhibition of the NF-кB signaling pathway.
III
致謝
碩士生涯在不知不覺中逐漸步入尾聲了,雖然覺得十分高興,但心裡卻仍然 隱隱覺得有些不捨。在兩年的碩士生涯當中,我學到了許多新的知識與實驗技術, 但我覺得我改變最多的是,我由原本一個只會 DNA cloning 的大學生,變成了一 個具有獨立思考能力的研究生。我學會了如何從 papers 當中學習並設計實驗, 也學會了如何從實驗失敗的經驗當中尋找出解決的方法;我覺得這是我在大學時 候所無法做到的。因此,我在此要感謝這兩年所遇到的老師、學長姐、學弟妹們, 謝謝你們給了我這麼多的指導以及特別的回憶。 感謝梁美智老師,總是不厭其煩地給予我一些實驗上的指導與方向,甚至遇 到一些需要較高技術的動物實驗時,還會親自示範給實驗室的大家看,令我們獲 益良多。因此,能夠成為梁老師實驗室的一員以及第一屆的畢業生,真的倍感榮 幸與驕傲。在此也要特別感謝楊昀良老師以及孫仲銘老師的不吝指導,令我學習 到一些我之前沒想過的知識,也讓我的論文能夠更加地完整。 由於我是一個不折不扣的夜貓子,所以十分感謝實驗室的大家能夠體諒我無 法在中午十二點以前進入實驗室的壞習慣,哈哈!謝謝世宏學長,給予我許多實 驗上的指導,希望學長的病能夠早日痊癒;謝謝與我大學同窗四年的黃俊嵐與馬 祖元,每次都會跟我討論 data,讓我能夠更順利地把漂亮的 data 生出來;謝謝逸 翔、千芳與婷媛,能夠幫忙分擔實驗室的一些雜事,好讓我能夠更專心地做實驗; 謝謝大學部的各位專題生們,每當我實驗做得不順時,會陪我去打球紓解壓力。 最後,我要感謝我最愛的雙親,從小到大甚麼事情都讓我自己選擇並給予我 支持及肯定,令我真的覺得十分幸福。IV
目錄
壹、緒論……….1 1.1 Aminofuran-linked-benzimidazole………...………1 1.2 NF-кB 訊息傳遞途徑………..…………2 1.2.1 NF-кB……….………...21.2.2 IкB kinase enzyme complex (IKK)……….……..4
1.2.3 NF-кB inhibitor protein (IкB)………....……6
1.3 NF-кB 訊息傳導途徑的啟動機制………7 1.3.1 致癌蛋白(oncoproteins)……….………7 1.3.2 致癌相關途徑(oncogenic-associated pathways)………...…..8 1.3.3 細胞因子(cytokines、growth factor、ligands)………...9 1.4 NF-кB 過度表現與癌症關係………...……….10 1.4.1 NF-кB 促進腫瘤細胞的生長(proliferation)………...10 1.4.2 NF-кB 抑制腫瘤細胞的凋亡(apoptosis)………...11 1.4.3 NF-кB 促進腫瘤細胞的血管新生(angiogenesis)、侵襲(invasion)、 轉移(metastasis)……….………11 1.5 探討 NF-кB 在人類癌症(human cancer)內的表現……….………….12 1.5.1 NF-кB 與人類淋巴癌(human lymphoma)……….……12 1.5.2 NF-кB 與人類多發性骨髓瘤(multiple myeloma)………...….14 1.5.3 NF-кB 與其他人類癌症類型………...………….15 貳、材料………..16 2.1 培養之細胞株………...……….16 2.2 一級抗體(primary antibody)………...………...16 2.3 二級抗體(secondary antibody)………..……17
V 2.4 藥品………....17 2.5 緩衝液………...…….18 2.6 試劑組………19 2.7 血清、抗生素及細胞培養液...19 2.8 儀器設備………....20 2.9 其他………20 參、實驗方法………..……21 3.1 Aminofuran-linked-benzimidazole 與其衍生物的合成………...…….21 3.2 細胞培養………..…..21 3.2.1 細胞培養之環境與方法……….………...21 3.2.2 細胞保存及解凍技術………22 3.3 細胞治療(cell treatment)………..……23
3.4 抽取細胞內蛋白質(whole cell extracts)之技術………23
3.5 抽取細胞核內蛋白質(nuclear extracts)之技術……….24
3.6 聚丙烯醯胺膠體電泳 (SDS-PAGE)………...………..25
3.7 西方墨點法 (western blotting)………..…26
3.8細胞存活檢測實驗(cell viability assay)...27
3.8.1 MTT assay...27 3.8.2 MTS assay...27 肆、結果...28 4.1 Aminofuran-linked-benzimidazole 系列衍生物之 IкBα 降解反應的抑制能 力比較...28 4.2 #1412 抑制 NF- кB 訊息傳遞途徑的主要分子作用機制:...29 4.2.1 #1412 抑制 NIH3T3 細胞內 TNF-α 活化之 p65 蛋白的細胞遷 移(translocation)活動...29
VI 4.2.2 #1412 抑制 NIH3T3 細胞內 TNF-α 活化之 IкBα 蛋白的降解 (degradation)反應...30 4.2.3 #1412 抑制 NIH3T3 細胞內 TNF-α 活化之 IкBα 蛋白的磷酸化 (phosphorylation)反應...31 4.3 #1412 的抗癌效果...32 4.3.1 #1412 對於不同類型腫瘤細胞生長的抑制能力比較...32 4.3.2 #1412 有效地抑制淋巴癌細胞株 Jurkat 細胞的生長...33 4.3.3 #1412 誘導 Jurkat 細胞進行細胞凋亡(apoptosis)活動...34 4.3.4 #1412 抑制 Jurkat 細胞內 TNF-α 活化之 IкBα 蛋白的降解 (degradation)反應...35 4.4 #21026 的抗癌效果...36 4.4.1 #1412 與其進階衍生物 #21026 在 NIH3T3 細胞之 IкBα 降解反 應的抑制能力比較………...36 4.4.2 #21026 抑制 Jurkat 細胞內 TNF-α 活化之 IкBα 蛋白的降解 (degradation)反應...37 4.4.3 #21026 有效地抑制淋巴癌細胞株 Jurkat 細胞的生長...37 4.4.4 #21026 誘導 Jurkat 細胞進行細胞凋亡(apoptosis)活動...38 4.4.5 #1412 與其進階衍生物 #21026 在 RPMI-8226 細胞之 IкBα 降解 反應的抑制能力比較...39 4.4.6 #21026 有效地抑制多發性骨髓瘤細胞株 RPMI-8226 細胞的生長 ………..40 4.4.7 #21026 誘導 RPMI-8226 細胞進行細胞凋亡(apoptosis)活動...40 伍、討論...41 陸、參考文獻...44
VII
圖目錄
圖 1A #1412 及#21026 的主要結構-- Aminofuran-linked-benzimidazole………54 圖 1B Aminofuran-linked-benzimidazole 系列衍生物 #1412、 #1410、#1411、 #1413 在 NIH3T3 細胞之 IкBα 降解反應的抑制能力比較………55 圖 1C Aminofuran-linked-benzimidazole 系列衍生物 #1412、 #1422、#1423、 #1424 在 NIH3T3 細胞之 IкBα 降解反應的抑制能力比較…………..…56 圖 1D Aminofuran-linked-benzimidazol 系列衍生物 #1412、 #1425、#1426、 #1427 在 NIH3T3 細胞之 IкBα 降解反應的抑制能力比較…………..…57 圖 2.1 #1412 抑制 NIH3T3 細胞內 TNF-α 活化之 p65 蛋白的細胞遷移 (translocation)活動………...…58圖 2.2 #1412 抑制 NIH3T3 細胞內 TNF-α 活化之 IкBα 蛋白的降解(degradation) 反應...59 圖 2.3 #1412 抑制 NIH3T3 細胞內 TNF-α 活化之 IкBα 蛋白的磷酸化 (phosphorylation)反應...60 圖 3.1 #1412 對於不同類型腫瘤細胞生長的抑制能力比較……….……..62 圖 3.2 #1412 有效地抑制淋巴癌細胞株 Jurkat 細胞的生長……….63 圖 3.3 #1412 誘導 Jurkat 細胞進行細胞凋亡(apoptosis)活動………..…64 圖 3.4 #1412 抑制 Jurkat 細胞內 TNF-α 活化之 IкBα 蛋白的降解反應…….…65 圖 4.1 #1412 與其進階衍生物 #21026 在 NIH3T3 細胞之 IкBα 降解反應的抑制 能力比較………...66
圖 4.2 #21026 抑制 Jurkat 細胞內 TNF-α 活化之 IкBα 蛋白的降解(degradation) 反應……….….67
圖 4.3 #21026 有效地抑制淋巴癌細胞株 Jurkat 細胞的生長……….…68
圖 4.4 #21026 誘導 Jurkat 細胞進行細胞凋亡(apoptosis)活動………..…..69 圖 4.5 #1412 與其進階衍生物 #21026 在 RPMI-8226 細胞之 IкBα 降解反應的
VIII
抑制能力比較………..70 圖 4.6 #21026 有效地抑制多發性骨髓瘤細胞株 RPMI-8226 細胞的生長……71 圖 4.7 #21026 誘導 RPMI-8226 細胞進行細胞凋亡(apoptosis)活動...72
IX
附錄
附圖 S1 Canonical NF-кB pathway 的主要機制……….….73 附圖 S2 Alternative NF-кB pathway 的主要機制………...….74 附圖 S3 #1411 對於不同類型腫瘤細胞生長的抑制能力比較………...…75 附圖 S4 #1413 對於不同類型腫瘤細胞生長的抑制能力比較………...76 附圖 S5 Proximicin A, B 以及 C 的結構圖………...77 附圖 S6 Bisbenzimidazoles (Hoechst 33258) 的結構圖...………...78X
縮寫表 (Abbreviation)
aa amino acids
APS ammonium persulfate
β-TrCP β-transducin repeat-containing protein DMEM Dulbecco’s Modified Eagle’s Medium
DMSO dimethyl sulfoxide DNA deoxyribonucleic acid
DTT dithiothreitol
EDTA ethylenediamine tetraacetic acid disodium salt EGTA ethylene glycol tetraacetic acid disodium salt
FBS fetal bovine serum
g gram(s)
h hour(s)
HLH helix-loop-helix
IC50 the half maximal inhibitory concentration
ICAM-1 intracellular adhesion molecule-1 IKK IкB kinase enzyme complex
IL interleukin
IL-1 interleukin-1
IL-6 interleukin-6
XI
M molar
mA milliamp (current)
MAP3K mitogen-activated protein kinase kinase kinase MEKK mitogen activated protein/ERK kinase kinase
min minute(s)
ml milliliter(s)
MTS 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H- tetrazolium
MTT 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
NAK NF-kappa-B-activating kinase NEMO NF-kappa-B essential modulator
NF-кB nuclear factor kappa-light-chain-enhancer of activated B cells NHL non-Hodgkin’s lymphoma
NIK NF-kappa-B-inducing kinase NLS nuclear localization signal PAGE polyacrylamide gel electrophoresis PARP poly (ADP-ribose) polymerase
PBS phosphate-buffered saline P-IкBα phosphorylation form of IкBα
PMSF phenylmethylsulfonyl fluoride RHD Rel homology domain
XII
RIP1 receptor-interacting protein 1 rpm revolutions per minute SDS sodium dodecyl sulfate
sec second(s)
Ser serine
TAK1 TGF-β activating kinase 1 TBS tris-buffered saline
TBST tris-buffered saline with tween-20 TD transactivation domain
TEMED N,N,N’,N’-tetramethylethelenediamine TFIID transcription factor II D
TGF-α transforming growth factor-alpha TNFR1 TNF-α receptor type 1
TNF-α tumor necrosis factor-alpha TRADD TNFR1-associated death domain protein
TRAF2 TNF-receptor-associated factor 2
µg microgram(s)
µl microliter(s)
µM micromolar
VCAM-1 vascular cell adhesion molecule-1 VEGF vascular endothelial growth factor
1
壹、緒論
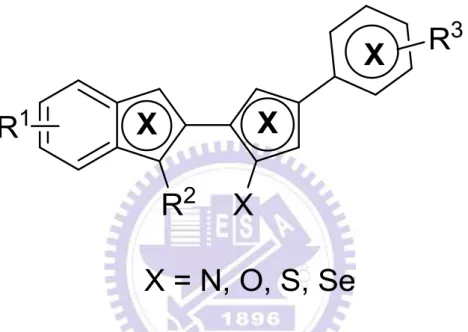
1.1 Aminofuran-linked-benzimidazole
Aminofuran-linked-benzimidazole 是透過有機化學合成的方式,將 aminofuran 與 benzimidazole 這兩種化合物結合而產生的新穎有機化合物。在這之前, aminofuran 與 benzimidazole 在許多研究已被證實具有抗癌 (anticancer) 的生物活 性。舉例來說,Proximicins (附圖 S5) 是一種新穎的 aminofuran 抗生素(antibiotics)。 Proximicins 被證實能夠有效地抑制人類胃腺癌 (gastric adenocarcinoma) 細胞株 AGS、人類肝癌 (hepatocellular carcinoma) 細胞株 Hep G2 以及人類乳癌 (breast carcinoma)細胞株 MCF7 的生長 (Fiedler et al., 2008)。除此之外,一些研究也顯 示由 benzimidazole 所衍生出來的化合物具有抗高血壓、抗病毒以及抗腫瘤的功 效 (Spasov AA et al., 1999)。 例如 bisbenzimidazoles (附圖 S6),為benzimidazole 的衍生物,已被證實能夠有效地抑制腎癌 (renal cancer)、中樞神經細胞癌 (CNS cancer)、結腸癌 (colon cancer)、皮膚黑色素癌 (melanoma),以及乳癌 (breast cancer)細胞的生長 (Baraldi et al., 2004)。
雖然 aminofuran 與 benzimidazole 這兩種化合物皆有抗腫瘤的功效,但其在 腫瘤細胞內的分子抗癌機制卻還未明瞭。因此,交通大學應用化學系孫仲銘教授 實 驗 室 將 aminofuran 與 benzimidazole 結 合 衍 生 出 一 系 列 以 aminofuran-linked-benzimidazole 為主要結構的衍生物,再由我們深入研究這些衍 生 物 的 生 物 活 性 及 其 分 子 抗 癌 機 制 。 我 們 希 望 能 夠 透 過 將 aminofuran 與 benzimidazole 的結合來增強這些化合物的抗癌功效,並且探討其在腫瘤細胞內的 抗癌機制及其作用。
2
1.2 NF-кB 訊息傳遞途徑:
1.2.1 NF-кB
NF-кB ( nuclear factor kappa-light-chain-enhancer of activated B cells)是動物 細胞內非常重要的蛋白轉錄因子。NF-кB 參與了許多基因的轉錄反應過程 ( transcription activity );因此 NF-кB 在許多細胞反應 (如細胞生長、細胞凋亡、免 疫及發炎反應) 的調控過程中扮演著非常重要的角色 (Loop et al., 2003)。 一旦 NF-кB 的調控失常,NF-кB 被活化會誘使其下游的一些炎症調控因子【如 細胞因子 (cytokines), 趨化因子 (chemokines), 及附著因子 (adhesion molecules)】 表現,造成發炎反應,進而導致癌症產生 (Rothwarf and Karin, 1999)。
NF-кB 是由五種不同的亞基體 (subunits) 透過不同的排列組合而成的一種 異源 (hetero-) 或同源二聚體 (homodimers)。這五種亞基體 (NF-кB 家族成員) 分別是:c-Rel , p65(Rel-A) , Rel-B , NF-кB 1(p50 及先驅基因 p105) , 及 NF-кB 2 (p52 及先驅基因 p100)。其中 c-Rel、p65(Rel-A)、Rel-B 具有激活區 (transactivation domain) (TDs) , 能夠透過與其他轉錄因子接觸而協助轉錄反應進行。每個 NF-кB 二聚體皆具有一個 N 末端活化區域 (N-terminal region),稱為 Rel Homology Domain (RHD) 。 RHD 內 包 含 了 基 因 結 合 (DNA binding) 與 聚 合 區 域 (dimerization domains)
,
以及細胞核的定位訊息 (nuclear localization signal) (NLS)。 RHD 主要負責進行 NF-кB 的聚合反應以及專一性地結合在特定的基因序列上以 起始基因轉錄反應 (reviewed in Jost and Ruland, 2007)。在大部分細胞種類,未活化的 NF-кB 皆存在於細胞質內。在細胞質內,未 活化的 NF-кB 首先會與其特定的抑制蛋白 IкB 相結合。IкB 能夠捉住 NF-кB,抑 制其轉變成活化狀態並遷移 (translocation) 至細胞核內以進行目標基因的轉錄
3
反應。因此,NF-кB 的活化可說是起始於抑制蛋白 IкB 的降解反應 (degradation) (Rothwarf and Karin, 1999)。
目前已被證實共有兩種 NF-кB 訊息傳遞途徑,分別是 canonocal pathway 及
alternative pathway (Bonizzi and Karin, 2004)。
在 canonocal pathway,當外部訊息傳遞至接收器上時 (如某些細胞因子: TNF-α , IL-6),會活化其下游的 IкB 激酶 (IKK),以進行磷酸化反應。一旦 IKK 被 活化,IкB 激酶便會磷酸化位於 IкB 調節區域 ( regulatory domain) 上的兩個絲氨 酸 (serine) 序列片段: Ser 32 及 Ser 36。磷酸化後的 IкB 會透過泛素蛋白酶體途 徑 (ubiquitin pathway) 進行泛素蛋白酶反應 (ubiquitination),進而最終被蛋白酶 (26S 蛋白酶) 給降解掉。在失去了 IкB 的抑制後,NF-кB 二聚體 (主要是 p65/p50) 便能夠遷移至細胞核內以結合目標基因序列,並起始目標基因的轉錄 反應。在這整個循環結束前,NF-кB 通常會在細胞質內被 IкB 再合成 (re-synthesis), 並與 IкB 相結合 (DiDonato et al., 1996) (附圖 S1)。
在 alternative pathway , 外 部 傳 導 的 訊 息 則 會 活 化 IKKα 同 源 二 聚 體 (homodimers),使其直接磷酸化 NF-кB2/p100,誘使 p100 進行蛋白水解作用 (proteolysis),餘下 p52。之後,p52 會與 Rel-B 相結合形成異源二聚體(heterodimers), 再遷移至細胞核內 (Dejardin et al., 2002) (附圖 S2)。
4
1.2.2 IкB kinase enzyme complex (IKK)
IκB kinase enzyme complex (IKK)主要是由 IKKα 及 IKKβ 所構成的異源二聚 體 (hetero-dimer) 以 及 調 控 蛋 白 IKKγ (NEMO) 所 共 同 組 合 而 成 的 複 合 體 (reviewed Hayden and Ghosh , 2004)。 除了以上這三種亞基體 (subunits),整個 IKK 複 合 體 還 包 含 了 一 些 成 員 如 ELKS (Ducut Sigala et al., 2004) 以 及 HSP-90/Cdc37 (Chen et al., 2002)。
IKKα 及 IKKβ 皆為絲氨酸專一激酶 (serine-specific kinases),且彼此共享 52%的同源氨基酸 (Mercurio et al., 1997)。IKKα 或 IKKβ 皆具有一個 N 末端激酶 活化區 (N-terminal kinase domain),一個白氨酸拉鍊結構 (leucine zipper),以及 一個 helix-loop-helix (HLH) motif (Delhase et al., 1999)。
一旦受到刺激時,IKKβ 的激酶活化區 (aa15-300) 上的活化環 (activation loop) (aa166-192) 便會被活化,誘使活化環上的兩個絲氨酸序列片段 (Ser-177 以 及 Ser-181) 進行磷酸化反應 (Delhase et al., 1999)。
IKKβ 的活化環負責保護一些激酶如 IKKα,MEK1 及 MEK2,故其在調控 激酶反應的過程中扮演著非常重要的角色。當IKKβ 的活化環因受到細胞因子如 TNF-α 或 IL-1 的刺激而活化時,會造成 Ser-177 與 Ser-181 進行磷酸化反應。然 而,在同樣存在 TNF-α 或 IL-1 活化的情況下,若把這兩個重要的絲氨酸序列片 段 (serine residues) 與丙氨酸序列片段 (alanine residues) 相置換,則 IKKβ 的激 酶反應 (kinase activity) 將會被終止。相反,若利用谷氨酸序列片段 (glutamic acid residues) 與這兩個絲氨酸序列片段 (serine residues) 相置換,則會造成 IKKβ 的過度活化異常 (constitutively active form) (Delhase et al., 1999)。
5
目前為止,誘使 IKK 活化的主要機制還未完全明瞭,但初步認為 IKK 的活 化與 mitogen-activated protein kinase kinase kinase (MAP3K) 有關。MAP3 kinase 成員包括了 NIK, mitogen activated protein/ERK kinase kinase (MEKK)1, MEKK3, NF-кB-activating kinase (NAK), 以及 TGF-β activating kinase 1 (TAK1)。 MAP3K 已被證實能夠直接磷酸化 IKK (reviewed in Hayden and Ghosh, 2004; Yamamoto and Gaynor, 2004)。
IKKγ 是 NF-кB 最主要的調控蛋白。IKKγ 在哺乳動物細胞的 IKK 活化過程 中是不可或缺的 (Yamaoka et al., 1998)。IKKγ 能夠協助 IKKα、IKKβ 與其他成 員匯集成 IKK 蛋白複合體 (Li et al., 2001; Yamamoto et al., 2001),且同時也能夠 增 強 IKK 蛋 白 複 合 體 與 IкB 之 間 的 聯 繫 , 以 促 進 IкB 的 磷 酸 化 反 應 (phosphorylation) 及降解反應 (degradation) (Yamamoto et al., 2001)。IKKγ 的磷酸 化反應在 NF-кB 的調控上扮演著非常重要的角色。有趣的是,IKKβ 能夠直接參 與IKKγ 的磷酸化反應。IKKβ 主要是刺激 IKKγ 的兩個絲氨酸序列片段 ( Ser-369 與 Ser-375) 進行磷酸化反應。IKKβ 與 IKKγ 之間的聯繫 (interaction) 減弱,將 降低 IKKγ 的磷酸化反應,並進而導致 IKKβ 的活化能力下降 (Prajapati and Gaynor, 2002)。
IKK 複合體位於 NF-кB 訊息傳遞途徑的上游,主要負責調控 NF-кB 的抑制 蛋白 IкBα 的磷酸化反應。一旦 IKK 複合體被活化,IKKα 或 IKKβ 便會磷酸化 IкBα,進而導致 NF-кB 得以遷移至細胞核內。雖然 IKKα 及 IKKβ 皆能直接磷酸 化 IкBα 的 N 末端上的 Ser-32 及 Ser-36 序列片段,但 IKKβ 已被證實它才是造成 IкBα 的 Ser-32、Ser-36 序列片段磷酸化的最主要的激酶 (Li et al., 1999a; Li et al., 1999b) 。
6
1.2.3 NF-кB inhibitor protein (IкB)
IкB 是 NF-кB 主要的抑制蛋白,其在 NF-кB 訊息傳遞途徑的調控上扮演著 非常重要的角色。目前,已有七種 IкB 成員被證實,分別是:IкB-α, IкB-β, IкB-γ, IкB-ε, Bcl-3, p100 及 p105,其中 IкB-α 為最主要的 NF-кB 抑制蛋白 (reviewed in Basseres and Baldwin, 2006)。
每個 IкBs 皆具有多段重複性的 30-33 aa 基因序列,稱為 ankyrin repeats,可 協助調控 IкB 與 NF-кB dimers 之間的聯繫。Ankyrin repeats 透過與 NF-кB 複合 蛋白的 Rel Homology Domain (RHD) 相結合,藉此遮蔽其細胞核定位訊息 (nuclear localization signal) (NLS),以防止 NF-кB 的遷移活動 (translocation activity) (reviewed in Basseres and Baldwin, 2006)。
一旦 IкBs 接收到上游的活化訊息時,其調控區域(regulatory domain)上的兩 個絲氨酸序列片段(serine residues) : Ser 32 及 Ser 36 便會被磷酸化。磷酸化後的 IкBs 會與 β-TrCP 相結合,並進而誘發泛素連接酶複合體蛋白(ubiquitin-ligase complex)的合成,起始泛素蛋白酶體途徑 (ubiquitin pathway)。透過泛素蛋白酶
體途徑,IкBs 最終會被蛋白酶 ( 26S proteasome ) 給降解掉,並讓 NF-кB 得以遷 移至細胞核內 (Maniatis, 1999)。
Bcl-3 是當中唯一比較特殊的 IкB 家族成員,Bcl-3 並不會抑制 NF-кB 的遷 移活動 (translocation activity),反而會輔助 NF-кB p50 或 p52 同源複合蛋白進行 往細胞核的遷移 (Fujita et al., 1993;Westerheide et al., 2001)。
7
1.3 NF-кB 訊息傳導途徑的啟動機制
1.3.1 致癌蛋白(oncoproteins)
透過觀察癌細胞內 NF-кB 的基因結合活動 (DNA-binding activity),證實了 許多致癌蛋白 (oncoproteins) 也參與了 NF-кB 的活化 (Karin et al., 2002)。舉例 來說,致癌基因 Her-2/Neu (ErbB2) 或 H-Ras 的過度表現能夠有效地啟動 NF-кB 訊息傳遞途徑,使到 NF-кB 活化 (Finco et al., 1997;Pianetti et al., 2001)。
在小鼠纖維細胞株 (immortalized murine fibroblasts) 內,H-Ras 被發現能夠 誘導 NF-кB 亞基體 (subunits) p65 (RelA)及 c-Rel 進行細胞核的遷移活動 (translocation)。除此之外,H-Ras 也被證實參與了 IKK 複合蛋白的活化 (Arsura et al., 2000)。
另外一個致癌蛋白--Bcr-Abl,也被發現與 chronic myelogenous leukemia(CML) 的 NF-кB 活動異常有關 (Reuther et al., 1998)。 藉由抑制 Bcr-Abl 所引發的 CML 內的 NF-кB 活動,被證實能夠有效地抑制腫瘤細胞的生長 (Reuther et al., 1998)。 在這之後,Bcr-Abl 被發現是藉由活化 MEK kinase 1(MEKK1),進而再活化其下 游的 NF-кB (Nawata et al., 2003)。除此之外,在 Bcr-Abl 表現異常的 acute lymphocytic leukemias (ALL) 內,也發現其 NF-кB 表現過度活化 (reviewed in Basseres and Baldwin, 2006)。
在一些淋巴癌疾病, NF-кB 也會被 MALT/c-IAP2 所構成的複合蛋白體活化 而造成 NF-кB 過度表現。MALT/c-IAP2 是藉由誘使 IKKγ (NEMO)進行泛素蛋白 酶體途徑(ubiquitin pathway),從而使 NF-кB 的調控失常 (Zhou et al., 2005)。
8
1.3.2 致癌相關途徑(oncogenic-associated pathways)
許多研究顯示,一些重要的訊息傳導途徑 (signaling pathway) 也參與了細胞 內 NF-кB 的活化,例如:P13K/Akt-dependent signaling。 Akt 被發現在 primary acute myeloid leukemia 有過度活化的跡象,且 Akt 的過度活化會影響其下游的 NF-кB 表現以及促進癌細胞的生長 (Grandage et al., 2005)。 除此之外,在人類 皮膚黑色素瘤 (human melanoma) 細胞,Akt 的活化也會造成 NF-кB 的表現異常 以 及 促 進 腫 瘤 的 生 成 (Dhawan and Richmond, 2002) 。 P13K/Akt-dependent signaling 也會促進 Her-2/ErbB2 基因表現,進而導致處於其下游的 NF-кB 也會隨 之被活化 (Pianetti et al., 2001)。
在一些肺腺癌細胞,Fas-associated death domain 的磷酸化反應被證實會造成 IKK 的活化,並間接導致了 NF-кB 的活化 (Chen and Goeddel, 2002)。 除此之外, 在胰臟癌細胞 (pancreatic cancer cells),E3-ubiquitin ligase receptor--βTRCP1 (beta-transducin repeat-containing protein 1) 的過度表現也被發現與 NF-кB 的活化 有關 (Muerkoster et al., 2005)。最後,在一些乳癌 (breast cancer) 細胞,CK2 的 表現也證實了能夠誘導 NF-кB 的活化 (Eddy et al., 2005)。
9
1.3.3 細胞因子(cytokines、growth factor、ligands)
當細胞質內的轉錄因子 NF-кB 受到外來刺激時,便會被活化並遷移至細胞 核內進行目標基因的轉錄反因。這些外來刺激包括了生長因子(如 TGF-α、EGF)、 細胞因子(如 TNF-α, IL-6)及干擾因子 (如 CD40-ligand) (reviewed in Pahl, 1999)。
在一些細胞種類,生長因子 EGF 會促使 NF-кB 的活化 (Biswas et al., 2000)。 EGF 是藉由造成 IкB 的降解,而促使 NF-кB 得以進行細胞核的遷移活動 (translocation activity) (Sitcheran et al., 2005)。
在一些細胞因子當中,tumor necrosis factor-α (TNF-α)是最為常見、也是最為 重要的 NF-кB 訊息傳導途徑的起始媒介。因此,許多相關的 NF-кB 標靶治療研 究皆選擇採用 TNF-α 作為實驗反應的起始因子。當 TNF-α 與其接受器
TNF-α receptor type 1 (TNFR1)相結合時,便會導致 TNFR1 進行三聚反應 (trimerization),並誘使特定的酪氨酸序列片段 (tyrosine residues) 進行自體磷酸 化反應 (autophosphorylation)。接著,磷酸化後的 TNFR1 會釋出一些調控蛋白如 TNFR1-associated death domain protein (TRADD), receptor-interacting protein 1 (RIP1) 以及 TNF-receptor-associated factor 2 (TRAF2)。這些調控蛋白會刺激一些 IKK 激酶如 mitogen-activated protein kinase kinase kinase (MAP3K),並導致這些 IKK 激酶活化其下游的 IKK 複合蛋白體 (reviewed in Chen and Goeddel, 2002; Hayden and Ghosh, 2004)。 IKK 復合蛋白體被活化便會導致其下游的 NF-кB 抑 制蛋白--IкB 進行磷酸化反應。
10
1.4 NF-кB 過度表現與癌症關係
癌症 (human cancer) 是一種由於體內細胞的基因表現異常而所引發的慢性 疾病。構成細胞癌化的主要原因大致上可以分為以下幾點: 1. 細胞生長異常或失去抑制細胞生長(cell growth)的機制 2. 失去促進細胞凋亡(apoptosis)的機制 3. 血管新生(angiogenesis)的能力增強 4. 細胞轉移(metastasis)的能力增強 有趣的是,這些構成細胞癌化的主要原因皆與 NF-кB 訊息傳遞途徑有關。 (reviewed in Hanahan and Weinberg, 2000;Hanahan and Weinberg, 2011)。1.4.1 NF-кB 促進腫瘤細胞的生長(proliferation)
NF-кB 能夠藉由調控特定的目標基因,而促進腫瘤細胞的生長 (cellular proliferation)。例如 NF-кB 能夠誘使 cyclin D1 進行活化,藉此導致腫瘤抑制基因 (tumor suppressor gene)--Rb 進行高度磷酸化反應 (hyperphosphorylation) (Karin, 2006)。除此之外,IкB 成員之一的 Bcl-3 聯合 p52 同源二聚體被發現也同樣具有 活化 cyclin D1 的能力 (Westerheide et al., 2001)。 除了 NF-кB 及 Bcl-3,由於 IKKα 具有調控 β-catenin 磷酸化的能力,因此 IKKα 在 cyclin D1 的轉錄過程中也 扮演著非常重要的角色 (Albanese et al., 2003)。最後,NF-к B 也被發現能夠藉由 調控 H1F-1α 以及 c-myc 的表現而促進腫瘤細胞的生長 (reviewed in Basseres and Baldwin, 2006)。
11
1.4.2 NF-кB 抑制腫瘤細胞的凋亡(apoptosis)
NF-кB 在一些抑制凋亡蛋白(anti-apoptotic proteins) (如 c-IAP1, c-IAP2, IXAP, TRAF1, TRAF2, Bcl-XL, Bcl-2 and Bfl1/A1) 的調控上扮演著非常重要的角色,因
此透過抑制 NF-кB 的過度表現,已被證實能夠促使大部份的腫瘤細胞進行細胞 凋亡的動作 (Ni et al., 2001; Oya et al., 2001; Gasparian et al., 2002; Mathas et al., 2003) 。舉例來說,PS-1145 為一種 IKKβ 抑制劑,它已被證實能藉由抑制 NF-кB 訊息傳遞途徑而導致多發性骨髓瘤細胞 (multiple myeloma) 進行細胞凋亡 (Hideshima et al., 2002)。
1.4.3 NF-кB 促進腫瘤細胞的血管新生(angiogenesis)、侵襲(invasion)、
轉移(metastasis)
NF-кB 已被證實能夠藉由調控血管內皮生長因子(vascular endothelial growth factor) (VEGF)以及 MMPs 的表現,促進腫瘤細胞的血管新生 (Karin et al., 2002)。 除此之外,透過抑制 NF-кB 訊息傳遞途徑也能夠很有效地抑制腫瘤細胞的侵襲 與轉移。舉例來說,藉由加強在人類皮膚黑色素癌(melanoma)以及卵巢癌(ovarian cancer)細胞內的 IкBα 表現,能夠使 VEGF 及 IL-8 的表現下降,並進而抑制腫瘤 細胞的生長、血管新生以及轉移 (Huang et al., 2000b)。此外,透過 IкBβ 的過度 表現,也能夠有效地抑制肺癌細胞內的 NF-кB 表現,並進而抑制肺癌細胞的轉 移(metastasis) (reviewed in Basseres and Baldwin, 2006)。
12
1.5 探討 NF-кB 在人類癌症(human cancer)內的表現
1.5.1 NF-кB 與人類淋巴癌(human lymphoma)
許多研究顯示,NF-кB 過度活化被發現與 Hodgkin lymphoma (HL)的病變 有關。與 B 細胞、T 細胞、及 NK 細胞這三種淋巴球細胞突變所引發的 non-Hodgkin lymphoma (NHL) 不同,由 Hodgkin/Reed-Sternberg (H/RS) 細胞所引發的淋巴癌 疾病類型,稱為 Hodgkin lymphoma (Kuppers et al., 2002)。目前,有許多關於造 成 NF-кB 在 HL 過度表現的機制已被證實。舉例來說,在 H/RS 細胞內的 CD30 接收器分子的自行聚合反應 (oligomerization) 會釋出 TRAF2 及 TRAF5,並導致 這兩個 TNF-α receptor-associated factor (TRAF) 進而刺激其下游的 IKK 活化 (Horie et al., 2002)。除此之外,在 H/RS 細胞也發現 RANK 與其 ligand (RANK-L) 會共同表現而導致 RANK 訊息傳遞途徑表現異常,並進而透過釋出的 TRAF2、 TRAF5 及 TRAF6 去活化下游的 IKK (Darnay et al., 1998) (RANK 是 TNF 接收器 的其中一個家族成員)。第三個在 H/RS 細胞表現異常的 TNF 接收器為 CD40。在 遭受到外部訊息的不斷刺激下,CD40 同樣會進行自行聚合反應 (oligomerization) 並釋出 TRAF2 以及 TRAF5 去活化下游的 IKK (Grech et al., 2004)。除 了 Hodgkin lymphoma(HL) , NF-кB 的 過 度 活 化 也 被 證 實 與 mucosa-associated lymphoid tissue (MALT lymphoma) 有 關 。 數 個 致 癌 基 因 (oncogene) 被發現在 MALT 淋巴瘤表現異常,這當中包括了 MALT1 以及 BCL10, 而這兩個致癌基因皆與 NF-кB 的活化有關。在 MALT 淋巴瘤,MALT1 從染色體 18 (chromosome 18) 遷移 (translocate) 至染色體 11,並與在染色體 11 的 IAP2 相結合,形成新的 IAP2-MALT1 蛋白複合體。IAP2-MALT1 會促使 IKKγ (NEMO) 進行泛素蛋白酶反應 (ubiquitination),進而導致 NF-кB 的活化 (Zhou et al., 2005)。 除此之外,BCL10 及 MALT1 也能互相結合形成 BCL10/MALT1,且同樣能夠藉 由 IKKγ (NEMO)進行泛素蛋白酶反應來造成NF-кB 的活化 (Zhou et al., 2004)。
13
Diffuse large B-cell lymphoma (DLBCL)是最常見的 non-Hodgkin lymphoma (NHL) 類型,其中 DLBCL 大致上又可分為三種類型:分別為 activated B-cell-like DLBCL (ABC-DLBCL)、germinal center-like DLBCL (GC-DLBCL)以及一些未明 確分類的 DLBCLs 另成一組 (Wright et al., 2003;Alizadeh et al., 2000; Rosenwald et al., 2002)。有趣的是,透過觀察在 DLBCL 內的 NF-кB 訊息傳遞途徑,發現在 ABC-DLBCL 內的 IKK 會產生異常活化並造成 IкBα 的降解,進而導致 NF-кB 的 高度基因結合反應 (DNA binding activity);但是在 GC-DLBCL 的 NF-кB 訊息傳遞 途徑卻沒有發現有任何基因表現異常的現象 (Davis et al., 2001)。CARMA1、 BCL10 以及 MALT1 這三種位於 NF-кB 上游傳遞途徑的蛋白分子,相信與 ABC-DLBCL 的 NF-кB 活化有關。然而,關於這三種蛋白分子對於調控 NF-кB 活化的機制則目前還未完全明瞭 (Ruland and Mak, 2003)。除此之外,在少數的 DLBCL 案例,也發現透過 alternative pathway 的 NF-кB 表現異常。那是由於 NF-кB2 進行基因重組,而造成 p100 缺失所引起的 (Ishikawa et al., 1997)。
除了體內的致癌蛋白,一些淋巴癌細胞的 NF-кB 也同樣會被一些帶有活化 NF-кB 活性蛋白的病毒所影響而異常表現,例如:Kaposi sarcoma-associated herpesvirus (KSHV)以及 human T-lymphotropic virus type I (HTLV-1)。KSHV 具有 一個 FLIP 蛋白稱為 vFLIP,能夠活化在 primary effusion lymphoma (PEL) 的 NF-кB 訊息傳遞途徑 (Chaudhary et al., 1999;Guasparri et al., 2004)。HTLV-1 具有 一個 Tax 磷酸致癌蛋白 (phospho-oncoprotein),Tax 能夠直接連接 adult T-cell lymphoma/leukemia (ATL) 的 IKKγ 並起始 IKK 的活動,增加 IкBα 及 IкBβ 的磷 酸化反應,進而造成 NF-кB 被活化 (Sun and Ballard, 1999; Xiao et al., 2001)。除 此之外,Tax 也能夠與 NF-кB2 的抑制蛋白 (inhibitory precursor) p100 相結合, 並廢除 p100 的抑制作用,誘使替代型 NF-кB 傳導途徑 (alternative NF-кB pathway) 的活化 (Beraud et al., 1994; Yin et al., 1998)。
14
1.5.2 NF-кB 與人類多發性骨髓瘤(Multiple myeloma)
在人類多發性骨髓瘤 (multiple myeloma) (MM),同樣發現有 NF-кB 過度活 化的現象。盡管如此,但卻沒有任何研究顯示在 MM 裡的 NF-кB 或者 IкB 的基 因有突變的跡象。但根據 Annunziata 等人在 2007 年的研究,NF-кB inducing kinase--NIK 的穩定化與大量堆積,是造成 MM 的 NF-кB 過度表現的主因。NIK 是 MAPK kinase kinase (MAP3K) 的 其 中 一 個 成 員 。 在 正 常 情 況 下 , TNF receptor-associated factor 3 (TRAF3) 會連接 NIK,接著由泛素連接酶 (ubiquitin ligases) cIAP1 或 cIAP2 聯合 TRAF2 所構成的蛋白複合體會誘導 NIK 進行泛素蛋白酶反應 (ubiquitination),進而造成 NIK 的降解 (degradation) (Vallabhapurapu et al., 2008)。因此,當在這過程中發生基因突變而造成 NIK 的降解反應被抑制,便 會導致其下游的 NF-кB 過度活化。這些基因突變包括了泛素連接酶蛋白複合體 或 NIK 基因本身發生突變 (Annunziata et al., 2007; Keats et al., 2007)。除此之外, cIAP1 或 cIAP2 的基因缺失也是造成 NIK 無法進行泛素蛋白酶反應的原因之一
(Annunziata et al., 2007; Keats et al., 2007)。最後,在某些人類多發性骨髓瘤病例, 也發現 TRAF2 突變而造成 NF-кB 過度活化。但由於 TRAF2 並不直接參與 NIK 的泛素蛋白酶反應,其作用只是加強泛素連接酶 cIAP1 或 cIAP2 的能力,故 TRAF2 基因突變只是少例 (Vallabhapurapu et al., 2008)。
15
1.5.3 NF-кB 與其他人類癌症類型
除了人類淋巴癌 (lymphoma) 與多發性骨髓瘤 (multiple myeloma),NF-кB 的活化異常也與其他癌症類型的引發有關,這包括了乳癌 (breast cancer)、皮膚 黑 色 素 癌 (melanoma) 、 肺 癌 (lung cancer) 以 及 胰 臟 癌 (pancreatic cancer) (reviewed in Basseres and Baldwin, 2006)。舉例來說,約有 86%的 ER-,ErbB2/Her2+ 乳癌細胞內有 p50/p65 異源二聚體 (heterodimer) 的異常活化現象 (Biswas et al., 2004)。 除此之外,NF-кB 也參與了一些趨化因子 (chemokines) 的調控,例如 IL-8。這些趨化因子皆被證實在人類皮膚黑色素癌有異常表現的跡象 (reviewed in Basseres and Baldwin, 2006)。另外,在動物以及人類胰臟癌組織細胞裡,也發 現有 NF-кB 表現異常的現象 (Liptay et al., 2003;Garcea et al., 2005)。
以上的研究結果一再地顯示 NF-кB 的過度活化與癌症的引發有莫大的關係。 因此,我們認為 aminofuran 與 benzimidazole 這兩種化合物的抗腫瘤機制或許也 與 NF-кB 訊息傳遞途徑的抑制有關。為了證明我們這一假設,我們利用小鼠胚 胎 纖 維 細 胞 株 NIH3T3 細 胞 做 為 篩 選 平 台 (platform) , 藉 此 深 入 觀 察 aminofuran-linked-benzimidazole 系列衍生物對於 NF-кB 訊息傳遞途徑的分子作 用機制。除此之外,我們也同時利用不同的腫瘤細胞株來進行細胞存活檢測實驗 (cell viability assay),以觀察 aminofuran-linked-benzimidazole 系列衍生物是否同 樣具有抗癌的效果。
16
貳、材料
2.1 培養之細胞株
:
這次的論文題目總共採用了六株不同的細胞株,分別是:
名稱 來源
i NIH 3T3 mouse fibroblast
ii HeLa human cervical adenocarcinoma
iii A549 human alveolar adenocarcinoma
iv H1299 human non-small cell lung carcinoma
v Jurkat human acute T-cell leukemia
vi RPMI 8226 human multiple myeloma
2.2 一級抗體 (Primary antibody) :
pAb : polyclonal antibody
名稱 種類 稀釋倍數 宿主 來源
i IкBα pAb 1:500 rabbit Santa Cruz Biotechnology #sc-371
ii Phospho - IкBα pAb 1:500 rabbit Cell Signaling Technology # 2859S
iii α-Tubulin pAb 1:1000 rabbit Santa Cruz Biotechnology #sc-12462-R
iv p65 pAb 1:500 rabbit Santa Cruz Biotechnology #sc-372
v TFIID pAb 1:500 rabbit Santa Cruz Biotechnology #sc-204
vi PARP-1 pAb 1:500 rabbit Santa Cruz Biotechnology #sc-7150
vii Caspase-3 pAb 1:500 rabbit Santa Cruz Biotechnology #sc-7148
17
2.3 二級抗體 (Secondary antibody) :
2.4 藥品 :
2-Mercaptoethanol (2-ME) (Sigma) 30% acrylamide/Bis-acrylamide (Serva) Ammonium persulfate (APS) (J.T.Baker) Bromophenol blue (Sigma)
DMSO (Sigma) DTT (Sigma) EDTA (Sigma) EGTA (Sigma) Glycerol (J.T.Baker) Glycine (J.T.Baker) Hepes (Sigma) Isopropanol (J.T.Baker) KCl (Sigma) Leupeptin (Sigma)
N,N,N’,N’-tetramethylethylenediamine (TEMED) (J.T.Baker) Na3VO4 (Sigma) Na4P2O7 (Sigma) NaCl (J.T.Baker) NaF (Sigma) Pepstatin (Sigma) PMSF (Sigma) SDS (Bio-Rad) TNF-α (R&D Systems) Tris-Base (J.T.Baker) Tris-HCl (J.T.Baker) Triton X-100 (J.T.Baker) Trypsin-EDTA (Biowest) Tween-20 (Sigma) 名稱 種類 稀釋倍數 宿主 來源
i Goat anti-Rabbit IgG (H+L), HRP conjugate
pAb 1:10000 goat Thermo Scientific Pierce #31460
18
2.5 緩衝液:
AT buffer
[20 mM Hepes , pH 7.9, 1% (w/v) Triton X-100, 20% (w/v) glycerol, 1 mM EDTA, 1 mM EGTA, 20 mM NaF, 1 mM Na4P2O7, 1 mM DTT, 1 mM Na3VO4,
1 µg/ml PMSF, 1 µg/ml leupeptin, 1 µg/ml pepstatin] Buffer E
【(10 mM HEPES, pH 7.9, 10 mM KCl, 0.1 mM EDTA, pH 8.0, 0.1 mM EGTA, pH 8.0) supplemented with 1 mM DTT, 5 µg/ml leupeptin, 10 µg/ml aprotinin, 1 µg/ml pepstatin and 1 mM PMSF】
Buffer F
【(20 mM HEPES, pH 7.9, 400 mM NaCl, 1 mM EDTA, 1 mM EGTA)
supplemented with 1 mM DTT, 5 µg/ml leupeptin, 10 µg/ml aprotinin, 1 µg/ml pepstatin and 1 mM PMSF】
1X resolving buffer (1.5 M Tris-HCl, pH 8.8) 1X stacking buffer (1 M Tris-HCl, pH 6.8) 10X SDS-running buffer
(0.25 M Tris Base, 1.92 M glycine, 1% SDS) 5X SDS loading buffer
【0.5 M Tris-HCl (pH 6.8), 12.5% 2-ME, 100% glycerol, 5% SDS, 0.25% bromophenol blue】
1X transfer buffer
(0.2 M Tris-HCl, 150 mM glycine, 20% (w/w) methanol ) 10X TBS buffer
(0.2 M Tris-HCl, 0.02 M KCl, 1.5 M NaCl, pH 7.5) TBST buffer (0.05% Tween-20 in 1X TBS)
19
2.6 試劑組:
Protein Assay Kit (Bio-Rad)
Supersignal Dura West Chemiluminescence Kit ( Pierce) MTT reagent (Sigma)
MTS one solution reagent (Promega)
2.7 血清、抗生素及細胞培養液:
10X PBS (Biowest)
Fetal bovine serum (FBS) (Biowest) Penicillin (Biowest)
Streptomycin (Biowest)
RPMI-1640 medium (Biowest)
20
2.8 儀器設備:
震盪器 (TEHTNICA model VORTEX-VIBROMIX 104 EV) 加熱攪拌器 (FISHER SCIENTIFIC model 210N3807) 可震盪乾浴槽 (EPPENDORF model F1.6A/250V) 酸鹼值檢測計 (METTLER TOLEDO model FE20/EL20) 搖晃器 (Shaker) (MAJOR SCIENCE model MS-NRK) 酵素免疫分析測讀儀 (BIO-RAD model 550)
PCR 溫度控制儀 (BIO-RAD model My CyclerTM
) 微量離心機 (CUBEE model A00691)
桌上型小型高速離心機 (DENVILLE SCIENTIFIC model 260D) 桌上型高速離心機 (EPPENDORF model 5702R)
桌上型低溫高速離心機 (BECKMAN model CS-15R Centrifuge) 電泳系統 (BIO-RAD model Mini-Protean@
Tetra System) 電子天秤 (DENVER INSTRUMENT model TP-3102) 微電腦溫控培養箱 (NUAIRE model NU-5500) 電子倒立顯微鏡 (OLYMPUS model CKX41)
2.9 其他:
21
參、實驗方法
3.1 Aminofuran-linked-benzimidazole 與其衍生物的合成:
本次實驗所用的 aminofuran-linked-benzimidazole 系列衍生物 (#1410、#1411、 #1412、#1413、#1422、#1423、#1424、#1425、#1426、#1427、#21026) 皆是由 交通大學應用化學系孫仲銘教授實驗室合成與提供。在用於細胞治療 (cell treatment) 前,所有化合物皆是溶於 100% DMSO 並儲存於 -20℃以做長期保 存。3.2 細胞培養:
3.2.1 細胞培養之環境與方法
:
Hela、A549、H1299、Jurkat 及 RPMI 8226 皆使用 RPMI-1640 medium (Biowest) 培養,而 NIH 3T3 則使用 Dulbecco’s modified Eagle’s medium (DMEM) (Biowest) 培養。 RPMI-1640 medium 及 DMEM 皆內含 10% 的胎牛血清 (fetal bovine serum) (FBS) (Biowest) , 50 units/ml 青 黴 素 (penicillin) 以 及 50 µg/ml 鏈 黴 素 (streptomycin) 。 以 上 這 些 細 胞 株 都 放 置 於 5% CO2、 37 ℃ 的 細 胞 培 養 箱
(NuAire,Inc.)內培養。
所有細胞株皆培養於 100-mm 組織培養皿上。當進行貼附型細胞 (NIH 3T3、 Hela、A549、H1299) 的分盤時,去掉原培養液後利用 1X PBS (Biowest) 稍微沖 洗細胞表面以去掉懸浮的死細胞殘骸,之後加上約 1 ml 的 trypsin-EDTA (0.25% trypsin, 0.02% EDTA; Biowest),並在 37℃靜置 1~2min。待原本貼附的細胞皆懸 浮起來後,加上約 10 ml 的培養液以中止胰蛋白酶反應 (trypsinization),并將懸
22 浮的細胞連同培養液移至 15-ml 離心管,之後在室溫下 1500 rpm 離心 5 分鐘 (eppendorf 離心機 , rotor HE 054)。在去掉上清液後,加入 10 ml 的培養液並再懸 浮 (resuspend) 細胞沉澱 (pellet) 數次,之後將約 1/10 的細胞移至內含 9 ml 新 鮮培養液的培養皿內培養。 懸浮式細胞 (Jurkat、RPMI 8226) 的分盤則是把內含懸浮細胞的培養液的 1/10 移至內含 9 ml 新鮮培養液的培養皿培養。
3.2.2 細胞保存及解凍技術 :
當保存貼附型細胞時,是把細胞養至 80~90% 滿後利用 trypsin-EDTA 來進 行胰蛋白酶反應(方法步驟如 3.2.1)。需收集 1.5~2 盤 100mm 的細胞量於同一管 50-mL 的離心管內去離心,以確保往後解凍時有足夠的細胞量存活下來。離心結 束後,利用不含抗生素的 0.9 ml 的培養液去再懸浮細胞沉澱數次。之後,緩慢加 入 0.1 ml 的細胞抗凍劑--DMSO (sigma)。反覆再懸浮細胞至融合均勻後,將內含 細胞與 DMSO 的培養液移至細胞冷凍管,並迅速將細胞冷凍管移動至 -20℃保 存兩小時。兩小時後,將冷凍管移動至 -80℃隔夜保存。第二天,再將細胞冷凍 管從 -80℃移動至液態氮桶做永久之保存。至於懸浮式細胞之保存,則省略胰蛋 白酶反應(trypsinization)這一步驟。 當解凍細胞冷凍管時,從液態氮桶取出後迅速移至 37℃水浴槽並將凍管底 部在水浴槽內緩慢攪拌以加速細胞之解凍。之後,將解凍完成的細胞溶液加至內 含 20 ml 新鮮培養液的 50-mL 離心管,在室溫下 1500 rpm 離心 5 分鐘 (eppendorf 離心機 , rotor HE 054)。離心結束後,去掉上清液並利用 10 ml 的新鮮培養液將 細胞沉澱再懸浮至均勻。最後,將內含細胞的培養液移至 100-mm 培養皿培養。23
3.3 細胞治療(Cell Treatment):
在進行細胞治療前,將原本養在 10% FBS 培養液內 70~80% 滿的
NIH 3T3、Jurkat 及 RPMI 8226 細胞移至 0.5% FBS 的培養液培養 24 小時,以進 行細胞飢餓(cell starving)的動作,讓該細胞對於藥物更加地敏感。24 小時後, 加入依據實驗所需之濃度的化合物至細胞並在 37.℃培養 2 小時。經過 2 小時的
培養後,加入 5 ng/ml 腫瘤壞死因子 TNF-α ( R&D Systems),並在 37℃培養 20 分鐘(或依據實驗所需之時間),以活化 NF-кB 活動。
3.4 抽取細胞內蛋白質(whole cell extracts)之技術:
抽取細胞內蛋白質時,先使用預冷的 1X PBS (Biowest)沖洗細胞表面兩次, 之後加上 1 ml 的 1X PBS 並使用細胞刮勺(scraper)把貼附型細胞連同 1X PBS 刮 收至 1.5 mL 微量離心管(eppendorf)內。接著,利用高速離心機 (Beckman Inc., rotor F2402H) 在 4℃、10000 rpm 下離心 5 分鐘。去掉上清液後,加入 80~100 µl AT 緩衝液[20 mM Hepes, pH 7.9, 1% (w/v) Triton X-100, 20% (w/v) glycerol, 1 mM EDTA, 1 mM EGTA, 20 mM NaF, 1 mM Na4P2O7, 1 mM DTT, 1 mM Na3VO4,
1 µg/ml PMSF, 1 µg /ml leupeptin, 1 µg /ml pepstatin],並利用 1 mL 型號的針筒抽 取細胞沉澱約 20 次,以確保細胞破裂釋出細胞內之蛋白質。確保細胞沉澱已完 全溶入 AT 緩衝液後,加入 5.6 µl 的 4 M 氯化鈉(最終濃度為 150 mM Nacl),之 後於 4℃、13000 rpm 下離心 30 分鐘。最後收集上清液(細胞內蛋白質)並保存 於 -80℃。 抽取懸浮型細胞內之蛋白質時省略刮收這一步驟,但須先將培養液去掉後方 可使用 1 ml 的 1X PBS 去再懸浮細胞沉澱,之後再重新離心一次。
24
3.5 抽取細胞核內蛋白質(nuclear extracts)之技術:
抽取細胞核內蛋白質時,前面如同 3.4 的方法,細胞在刮收、離心結束後, 將細胞沉澱溶解於 400 µl 的緩衝液 E 內。【(10 mM HEPES, pH 7.9, 10 mM KCl, 0.1 mM EDTA, pH 8.0, 0.1 mM EGTA, pH 8.0) supplemented with 1 mM DTT, 5 µg/ml leupeptin, 10 µg/ml aprotinin, 1 µg/mlpepstatin and 1 mM PMSF】接著,在4℃下靜置 15 分鐘。15 分鐘後,加入 25 µl 的 10% NP-40 並高速震盪(vortex)約 10 秒。之後, 將細胞混合物利用高速離心機 (Beckman Inc. , rotor F2402H) 在 4℃、10000 rpm 下離心 1 分鐘。離心結束後,去掉上清液只留下細胞沉澱。這時的細胞沉澱會發 現比原本的細胞沉澱小得多,即為細胞核之沉澱。接著,利用 300 µl 的 1XPBS 沖洗細胞核沉澱數次後,在 4℃、10000 rpm 下離心 1 分鐘。接著,去掉上清液 後加入 50~80 µl 的緩衝液 F 【(20 mM HEPES, pH 7.9, 400 mM NaCl, 1 mM EDTA, 1 mM EGTA) supplemented with 1 mM DTT, 5 µg/ml leupeptin, 10 µg/ml aprotinin, 1
µg/ml pepstatin and 1 mM PMSF】,並再懸浮細胞核沉澱數次。之後,將細胞混合 物置於 -80℃ 20 分鐘。20 分鐘後,將細胞混合物轉移至室溫下讓其解凍,以利 用結凍解凍所造成的物理傷害來把細胞核打破。重複結凍解凍兩次後,再把細胞 混合物在 4℃、14000 rpm 下高速離心 5 分鐘。最後,收集上清液(細胞核內蛋 白質),並保存於 -80℃。 抽取懸浮型細胞核內之蛋白質時,則與 3.4 之方法相同,省略刮收步驟,但 須先將培養液去掉,再將細胞沉澱溶解於 400 µl 的緩衝液 E 內。
*注:收集到的細胞及核內蛋白質,皆是透過使用 Protein Assay Kit (Bio-Rad)來 進行定量,而蛋白質定量的步驟與方法則參閱 Bio-Rad 所附之實驗步驟。
25
3.6 聚丙烯醯胺膠體電泳(SDS-PAGE):
聚丙烯醯胺膠體可分為上下層膠體兩部份。下層膠體為 resolving gel,而上 層膠體為 stacking gel。當配製 resolving gel 時,會依據樣品蛋白分子量的大小而 決定要配製多少% 的 resolving gel。(蛋白分子量越大,所需要的膠體% 數越低) 配製 resolving gel 時,需先配製 1X resolving buffer (1.5 M Tris-HCl, pH 8.8),接 著加入 30% acrylamide (Serva)以及 10% SDS,最後才加入 10% ammonium persulfate (APS) 及 N,N,N’,N’-tetramethylethylenediamine (TEMED) 以 加 速 resolving gel 的凝聚(polymerization)反應。當將尚未凝固的膠體溶液加入已架好 的注膠器內時,需在膠體表面加入約 500 µl 的異丙醇 (isopropanol) 以確保膠體 凝固後表面的平整。當 resolving gel 凝固後,去掉表面的異丙醇並加入 5% 的 stacking gel。配製 stacking gel 時,需先配製 1X stacking buffer (1 M Tris-HCl, pH 6.8),接著加入 30% acrylamide/bis acrylamide (Serva)及 10% SDS,最後才加 入 10% APS 及 TEMED。待上層的 stacking gel 凝固後,則可開始電泳實驗。
在開始電泳實驗前,將電泳膠體浸泡在 800 ml 的 1X SDS-running buffer 內 ( 10X SDS-running buffer: 0.25 M Tris Base, 1.92 M glycine, 1% SDS in 1L ddH2O)。
在這之前,可預先將 5X SDS loading buffer【0.5 M Tris-HCl (pH 6.8), 12.5% 2-ME, 100% glycerol, 5% SDS, 0.25% bromophenol blue】稀釋至 1 倍,並混入 20 µg 樣 品蛋白後,在 98℃下加熱 10 分鐘,之後才開始填充(loading)至電泳膠體內。待 樣品蛋白填充結束後,則可把電壓調至 80 伏特並開始跑電泳。直到樣品蛋白跑 到 resolving gel 時,則可把電壓上升到 120 伏特。
#注:在聚丙烯醯胺膠體電泳實驗裡所使用的電泳系統為 Mini-PROTEAN®Tetra System (Bio-Rad)。
26
3.7 西方墨點法(Western blotting):
當樣品蛋白成功透過 SDS-PAGE 分解並嵌在聚丙烯醯胺膠體上時,可利用 西方墨點法(蛋白轉漬實驗)把膠上之蛋白轉移至 PVDF 轉漬膜(Millipore)上。當 進行蛋白轉漬實驗時,需先將聚丙烯醯胺膠體與轉漬膜組裝在印跡轉移裝置 (transfer apparatus) (Bio-Rad)上,然後浸泡在 1X transfer buffer (0.2 M Tris-HCl,150 mM glycine, 20% methanol)內。之後,把電壓調至 250 微安培開始進行蛋白轉漬 實驗。大約 70~80 分鐘後,膠體上之蛋白便會完全轉移至轉漬膜上。接著,把該 轉漬膜浸泡在含 5% 脫脂奶粉的 TBST 內(TBS 加 0.05% Tween-20 配置成 TBST), 並放置於室溫 1 小時或 4℃一個晚上以進行阻斷(blocking)的動作。之後,將阻斷 (blocking)結束後的轉漬膜浸泡在一抗溶液內室溫培養 1 小時。一抗溶液的配法 為利用含 5% 脫脂奶粉的 TBST 將一抗稀釋至 1:500 倍。之後利用 TBST 沖洗膜 3 次,每次 10 分鐘。之後,再將轉漬膜浸泡在二抗溶液內室溫培養 1 小時。二 抗溶液的配法是利用含 5% 脫脂奶粉的 TBST 去稀釋二抗至 1:10000 倍。最後, 再利用 TBST 沖洗膜 3 次,每次 15 分鐘。接著,透過 Supersignal Dura West Chemiluminescence Kit (Pierce)來偵測膜上貼附之蛋白。
在這次論文題目裡,西方墨點法實驗所使用之抗體請參閱第二章節材料的部份。 (2.2、2.3)
27
3.8
細胞存活檢測實驗(
Cell viability assay) (MTT/MTS):
3.8.1 MTT assay :
MTT 實驗只適用於貼附型細胞。當進行 MTT 實驗時,將細胞數量稀釋至 50,000~75,000 cells/mL(依該細胞株之生長速度判斷)。之後,將一定數量之細胞 種在 96 孔盤隔夜培養。(Hela、A549、H1299 每孔約 7500 cells/100 µL)。 第二天,加入 1 µl 化合物(依據實驗所需之濃度,且每種濃度皆需要重複三 次)至細胞並在 37.℃培養箱培養 24 小時。 第三天,每孔加入 20 µl 的 5 mg/mL MTT 試劑 (Sigma)。接著,將 96 孔盤 重新放入 37℃.培養箱培養 3.5~4 小時。3.5 小時後,小心抽取內含 MTT 試劑的 培養液(切記不能用 PBS 去沖洗)。之後,每孔加入 200 µl 的 DMSO。接著, 將 96 孔盤在室溫下搖晃 15 分鐘。最後,利用酵素免疫分析測讀儀 (Bio-Rad) 在 540 nm 波長下讀取數值。3.8.2 MTS assay :
MTS 實驗一般使用於懸浮型細胞。與 MTT 實驗一樣,在進行 MTS 實驗前 需 先 將 細 胞 數 稀 釋 至 特 定 的 數 量 , 但 懸 浮 型 細 胞 需 要 的 細 胞 數 較 多 , 約 250,000~2,500,000 cells/mL (依該細胞株之生長速度判斷)。之後,將一定數量之 細胞種在 96 孔盤隔夜培養。(Jurkat 及 RPMI 8226 每孔約 75,000 cells/100 µl)。第二天,加入 1 µl 化合物(依據實驗所需之濃度,且每種濃度皆需要重複三 次)至細胞並在 37.℃培養箱培養 24 小時。第三天,每孔加入 20 µl 的 MTS 試劑
(Promega)。接著,將 96 孔盤重新放入 37.℃培養箱培養 4 小時。4 小時後,直接 利用酵素免疫分析測讀儀 (Bio-Rad) 在 490 nm 波長下讀取數值。
28
肆、結果
4.1 Aminofuran-linked-benzimidazole 系列衍生物之 IкBα 降
解反應的抑制能力比較
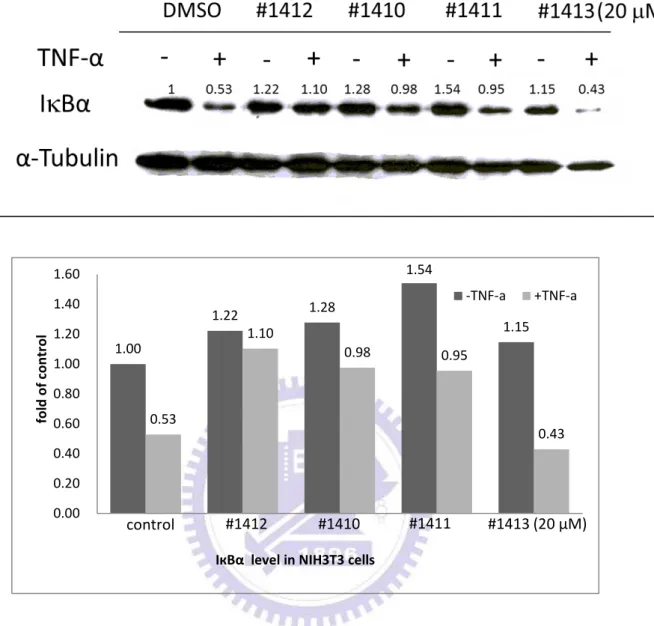
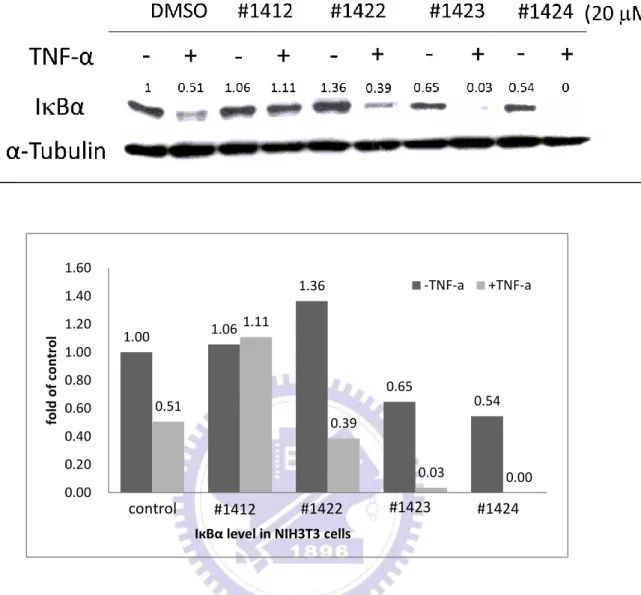
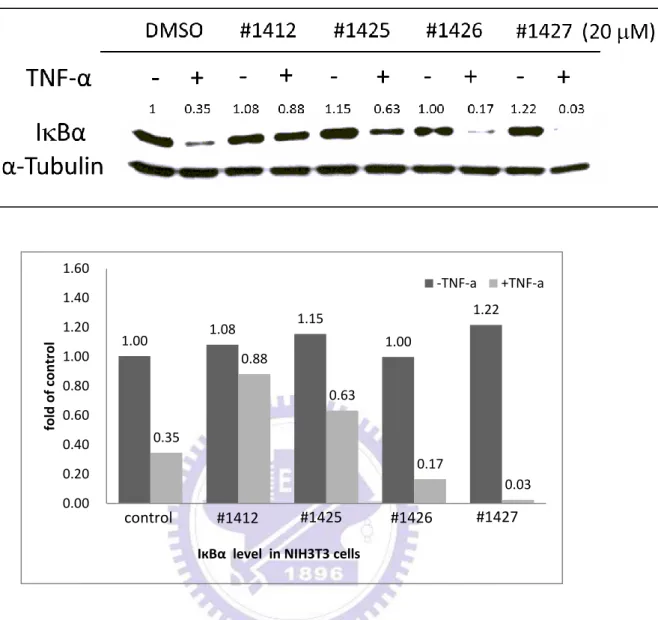
IкBα 是 NF-кB 訊息傳遞途徑裡非常重要的抑制蛋白,其主要的功能是捉住 NF-кB,使其無法遷移 (translocate) 至細胞核內並促使目標基因的轉錄。因此, 抑制 IкBα 的降解反應將是個不錯的藥物標靶目標。我們希望藉由西方墨點法觀 察各 aminofuran-linked-benzimidazole 系列衍生物抑制 IкBα 降解的能力,藉此比 較並分辨出能夠最有效地抑制 IкBα 進行降解反應的衍生物。在藥物篩選的過程 中,我們利用小鼠的胚胎纖維細胞株 NIH3T3 細胞做為篩選平台 (platform)。 NIH3T3 細胞在經過 24 小時的細胞飢餓 (cell starving) 動作後,加入 DMSO (控 制組) 或 20 M 的 aminofuran-linked-benzimidazole 系列衍生物 (#1410、#1411、 #1412、 #1413、 #1422 、 #1423 、#1424 、#1425 、 #1426 、 #1427) 至細胞並在 37.℃培養 2 小時。之後,加入 5 ng/ml 腫瘤壞死因子 TNF-α,並再培養 20 分鐘 以活化 NF-кB 活動。最後,利用西方墨點法偵測細胞內的 IкBα 與 α-tubulin 蛋白 質含量。圖 1B 的結果顯示,加入 5 ng/ml TNF-α 活化的細胞內的 IкBα (lanes 2、 4、6、8、10)因為會進行降解 (degradation) 反應,因此 IкBα 蛋白質含量會比沒 加入 5 ng/ml TNF-α 活化的細胞內的 IкBα (lanes 1、3、5、7、9) 蛋白質含量來得 少。但是同樣是在 TNF-α 活化的情況下,在加了 20 µM #1412 後的細胞內的 IкBα 蛋白質含量與控制組及其他衍生物 (#1410、 #1411、 #1413) 比較有顯著的較 多,這結果證明 #1412 能夠更有效地抑制 IкBα 進行降解反應。圖 1C、1D 的結 果與圖 1B 相似,證明了 20 μM 的 #1412 相較於其他相同濃度的同系列衍生物 (#1422、 #1423、 #1424、 #1425、 #1426、 #1427) 有較好的抑制 IкBα 降解 (degradation) 的能力。29
4.2 #1412 抑制 NF- кB 訊息傳遞途徑的主要分子作用機制:
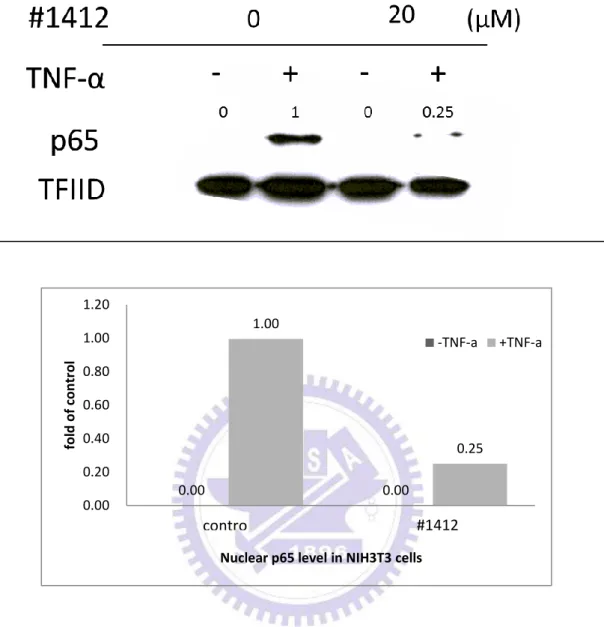
4.2.1 #1412 抑制 NIH3T3 細胞內 TNF-α 活化之 p65 蛋白的細胞遷
移(translocation)活動
在初步篩選出 #1412 具有最好的抑制 IкBα 降解的能力後,我們希望能夠更 深入地探討 #1412 對於抑制 NF-кB 訊息傳遞途徑的分子作用機制。NF-кB 的細 胞核遷移 (translocation) 活動是 NF-кB 訊息傳遞途徑裡非常重要的過程。因此, 我們接下來的實驗便是要觀察 #1412 是否也能夠抑制 NF-кB 的細胞核遷移活動。 NIH3T3 細胞在經過 24 小時的細胞飢餓 (cell starving) 動作後,加入 DMSO (控 制組) 或 20 M 的 #1412 至細胞並在 37.℃培養 2 小時。之後,加入 5 ng/ml TNF-α 並培養 20 分鐘以活化 NF-кB 活動。為了證實 #1412 能夠抑制 NF-кB 從細胞質 遷移至細胞核內,我們藉由抽取細胞核內蛋白,之後再透過西方墨點法去鑑定加 了#1412 後細胞核內的 p65 蛋白質含量 (p65 為 NF-кB 的其中一個成員)。根據圖 2.1,加入 5 ng/ml TNF-α 活化的細胞內的 p65 蛋白 (lanes 2、4 ) 因為會進行細胞 核內遷移活動,因此收集到的核內的 p65 蛋白質含量會比沒加入 5 ng/ml TNF-α 活化的細胞核內的 p65 蛋白質含量 (lanes 1、3) 來得多。但是同樣是在加入 5 ng/ml TNF-α 活化的情況下,在加了 20 µM #1412 後的細胞核內蛋白的 p65 量 與控制組比較有顯著的減少,這結果證明了#1412 能夠有效的抑制 NF-кB 的細 胞核內遷移,使其無法從細胞質進入細胞核內,因此細胞核內的 p65 量會比控制 組的 p65 量來得少。30
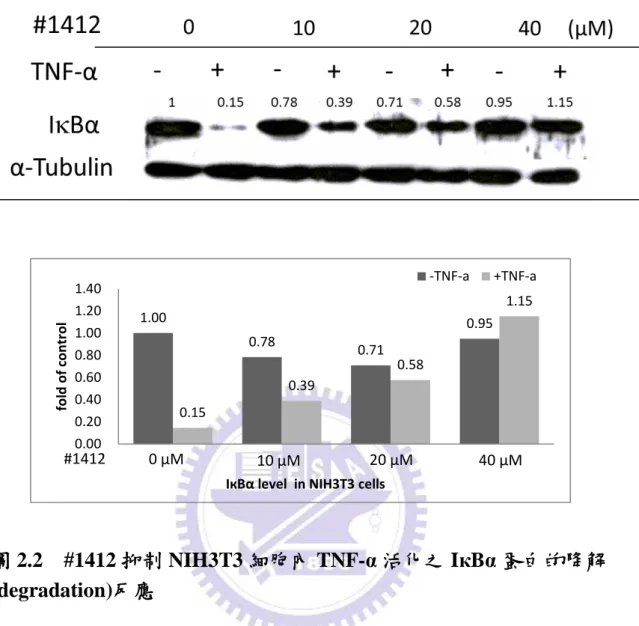
4.2.2
#1412 抑制 NIH3T3 細胞內 TNF-α 活化之 IкBα 蛋白的降解
(degradation)反應
為了進一步證實#1412 能夠直接參與 IкBα 的降解反應,所以我們利用不同 濃度的 #1412 進行了一個劑量相關 (dose-dependent) 的實驗,藉此觀察在不同 濃度 #1412 的影響下,NIH3T3 細胞內的 IкBα 表現差異。NIH3T3 細胞在經過 24 小時的細胞飢餓 (cell starving) 動作後,分別加入 DMSO (0 µM)或不同濃度的 #1412 (10 , 20 , 40 µM) 至細胞並在 37.℃培養 2 小時。之後,加入 5 ng/ml 腫瘤 壞死因子 TNF-α,並再培養 20 分鐘以活化 NF-кB 活動。最後,再利用西方墨點 法偵測細胞內的 IкBα 與 α-tubulin 蛋白質含量。圖 2.2 的結果顯示,在同樣加入 5 ng/ml TNF-α 活化的情況下 (lanes 2、4、6、8),控制組的 IкBα (lanes 2) 會進 行磷酸化反應而被降解掉。但隨著#1412 的濃度增加 (lanes 4、6、8),IкBα 降解 所受到的抑制越強,因此細胞內的 IкBα 蛋白質含量也越高。這個結果證明了 #1412 與 IкBα 降解反應有直接的抑制關係。
31
4.2.3
#1412 抑制 NIH3T3 細胞內 TNF-α 活化之 IкBα 蛋白的磷酸
化(phosphorylation)反應
IкBα 的磷酸化反應在 NF-кB 訊息傳遞途徑裡扮演著非常重要的角色。IкBα 的磷酸化反應是構成 IкBα 進行泛素蛋白酶反應 (ubiquitination),並進而導致 IкBα 被 26S 蛋白酶降解的主要原因。因此,我們接下來的實驗便是要觀察#1412 是 否 也 同 樣 能 夠 抑 制 IкBα 的 磷 酸 化 反 應 , 所 以 進 行 了 一 個 時 間 相 關 (time-dependent) 的實驗。透過利用不同時間點的 TNF-α ( 0 , 5 , 10 , 20 min ) 去 誘導 NF-кB 活動的進行,並藉此來觀察磷酸化後的 IкBα 在 NIH3T3 細胞內的表 現量。NIH3T3 細胞在經過 24 小時的細胞飢餓 (cell starving) 動作後,加入 DMSO (控制組) 或 20 M 的#1412 至細胞並在 37.℃培養 2 小時。之後,分別加入 0 , 5 , 10, 20 min 的 5 ng/ml TNF-α 去活化 NF-кB 活動。最後,再利用西方墨點法偵測 細胞內 phospho-IкBα , IкBα 與 α-tubulin 蛋白質含量。根據圖 2.3,在還沒加入 20 µM #1412 前 (lanes 1、2、3、4),在 5 分鐘的時候仍然還可以看到 IкBα 磷酸化 所產生的 phospho-IкBα,然後 phospho-IкBα 在 10、20 分鐘後就會開始被蛋白酶 降解掉而逐漸消失。但是在加上 20 µM #1412 後 (lanes 5、6、7、8),可以發現 所產生的 phospho-IкBα 與加藥前比較明顯的減少了許多。這個結果證明了 #1412 能夠有效地抑制住 IкBα 的磷酸化反應。除此之外,隨著 TNF-α 的活化,會誘使 IкBα 開始進行降解反應並導致 IкBα 蛋白質含量逐漸減少 (lanes 1、2、3、4), 但是在加入 20 µM 的 #1412 後,IкBα 蛋白質含量與加藥前比較卻沒有明顯的減 少 (lanes 5、6、7、8)。這個結果也進一步證實了 #1412 能夠藉由抑制 IкBα 的 磷酸化反應,並進而抑制 IкBα 進行降解反應。結論:從以上的實驗結果來看,我們認為 #1412 可能是透過抑制 NF-кB 的抑制 蛋白--IкBα 的降解反應,進而達到影響 NF-кB 訊息傳遞途徑的效果。
32
4.3 #1412 的抗癌效果:
4.3.1
#1412 對於不同類型腫瘤細胞生長的抑制能力比較
許多研究證實 NF-кB 的過度活化與很多癌症的引發有關,例如:子宮頸癌、 肺癌、淋巴癌等等。因此,為了證明#1412 具有抗癌的效果,我們接下來利用不 同的腫瘤細胞株來進行細胞存活檢測實驗 (cell viability assay)。在細胞存活檢測 實驗裡,我們總共使用了 HeLa、A549、H1299、以及 Jurkat 這四種不同的腫瘤 細胞株來進行 #1412 的抗癌測試。其中 HeLa 是子宮頸癌細胞、A549 與 H1299 皆是肺癌細胞、而 Jurkat 則是一種 T 淋巴癌細胞。在進行細胞治療 (cell treatment) 的時候,將不同濃度的#1412 (0 , 5 , 10 , 20 µM) 分別加入不同類型的腫瘤細胞株 內 ( HeLa , A549 , H1299, Jurkat ) 並在 37.℃培養二十四小時。二十四小時後,
利用 MTT 或 MTS 檢測各腫瘤細胞株的細胞存活率。其中 MTT 的檢測波長為 540 nm 而 MTS 的檢測波長為 490 nm。每個樣品皆重複測定 3 次以增加實驗準 確率。圖 3.1 的結果顯示在加了#1412 二十四小時後,不同濃度的 #1412 對於各 種癌細胞的生長所帶來的影響。#1412 並無法抑制 A549 細胞的生長,但卻能夠 十分有效地抑制 H1299 細胞以及 Jurkat 細胞的生長;而相較於 H1299 細胞以及 Jurkat 細胞,#1412 只能稍微抑制 HeLa 細胞的生長。 #1412 對於 HeLa 細胞的 IC50大約為 18 M;而對於 H1299 細胞的 IC50大約為 4 M 以及對於 Jurkat 細胞
的 IC50大約為 3.5 M。根據實驗結果,#1412 對於淋巴癌細胞的生長抑制能力最
33
4.3.2
#1412 有效地抑制淋巴癌細胞株 Jurkat 細胞的生長
由於#1412 對於淋巴癌細胞株 Jurkat 細胞的生長抑制效果最好,因此我們接 下來的實驗主要會集中在淋巴癌細胞上做更深入地探討。為了進一步探討 #1412 對於 Jurkat 細胞的抗癌效果,我們分別觀察 Jurkat 細胞在加入不同濃度的 #1412 ( 0, 2 , 4 , 8 µM) 十二小時、二十四小時、以及四十八小時後的生長情況。之後, 利用 MTS 檢測 Jurkat 細胞的細胞存活率。MTS 的檢測波長為 490 nm。每個樣 品皆重複測定 3 次以增加實驗準確率。根據圖 3.2,在加入 #1412 十二小時後 , Jurkat 細胞的生長在 4 µM 的濃度下便會開始受到抑制;並在 8 µM 的時候,其 細胞存活率只剩下大約 20%。在加入#1412 二十四小時後 , Jurkat 細胞的生長只 需在 2 µM 的濃度下便會開始受到抑制;並在 8 µM 的時候,其細胞存活率只剩 下大約 10%。在加入#1412 四十八小時後, 只需要 4 µM 的濃度,便能夠完全地 抑制住 Jurkat 細胞的生長。34
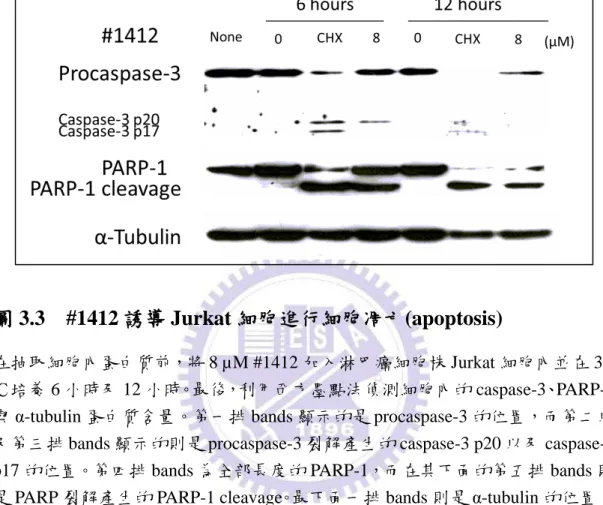
4.3.3
#1412 誘導 Jurkat 細胞進行細胞凋亡(apoptosis)活動
造成細胞停止生長並死亡的方式主要有兩種: 分別為細胞凋亡 (apoptosis) 以及細胞壞死 (necrosis)。細胞凋亡是生物體維持正常生理功能的一種行為,由 外接收死亡訊號,或由內自發性的死亡;而細胞壞死則是細胞遭受物理性或生物 毒物的剌激而導致細胞膜破裂,且通常會附隨引發發炎反應 (Kanduc et al., 2002; Mullen, 2004)。因此,在證明了#1412 能夠有效地抑制 Jurkat 細胞生長後,接下 來我們想要觀察#1412 主要是透過哪個途徑而導致 Jurkat 細胞死亡的。在抽取細 胞內蛋白質前,將 8 µM #1412 加入淋巴癌細胞株 Jurkat 細胞內並在 37.℃培養 6 小時及 12 小時。之後,再利用西方墨點法偵測細胞內的 caspase-3、PARP-1 與 α-tubulin 蛋白質含量。當細胞進行凋亡時,procaspase-3 以及 PARP-1 便會裂解 產生 caspase-3 以及 PARP-1 cleavage。Procaspase-3 以及 PARP-1 的這種蛋白裂解 反應目前已被許多研究當作細胞凋亡時很重要的指標。根據圖 3.3,第一排 bands 顯示的是 procaspase-3 的位置,而第二以及第三排 bands 顯示的則是 procaspase-3 裂解產生的 caspase-3 p20 以及 caspase-3 p17 的位置。第四排 bands 為全部長度的 PARP-1,而在其下面的第五排 bands 則是 PARP 裂解產生的 PARP-1 cleavage。 最下面一排 bands 則是 α-tubulin 的位置。cycloheximide 為一種蛋白合成抑制劑, 已被證實能夠有效地造成細胞凋亡,故在這個實驗裡採用了 cycloheximide 作為 正極的對照組 (positive control) 來比較。根據結果,在加了#1412 六小時後,皆 可發現有 caspase-3 以及 PARP-1 cleavage 的產生;但在加了#1412 十二小時後, 由於#1412 的毒性過於強烈,procaspase-3 及 PARP-1 幾乎已經被切光了,只能偵 測到少許的 caspase-3 以及 PARP-1 cleavage。這個結果證明了#1412 能夠有效地 誘導 Jurkat 細胞進行細胞凋亡活動。35