國立臺灣大學生物資源暨農學院動物科學技術學系 碩士論文
Department of Animal Science and Technology College of Bioresources and Agriculture
National Taiwan University Master Thesis
預先皮膚擴張於豬模式中用以準備自體脂肪移植之潛能
The Potential of Pre-Expansion of Skin Preparing for Autologous Fat Grafting in a Swine Model
陳昇志
Sheng-Chih Chen
指導教授:吳信志 博士 Advisors: Shinn-Chih Wu, Ph. D.
中華民國 106 年 1 月
January, 2017
誌謝
本論文得以順利完成,承蒙恩師 吳信志老師之悉心指導,吳老師不僅是學業 上更是我人生的導師,總是不斷給予機會讓我勇於克服自己的弱點,從每次專討報 告中吸取經驗並改正缺點,學習獨立思考和解決問題的能力,讓我深刻體會到自己 確實在一點一滴進步當中;亦感謝長庚醫院 高煌凱醫師在研究過程中於試驗設計 之建議以及實驗方向之引導,並且於口試時對此論文之試驗結果提出獨到之解析,
值得發人省思;感謝 鄭登貴老師撥冗擔任此次口試委員,並給予此論文之指導和 建議,使本論文得以漸趨完整。
碩士班求學之路雖然坎坷,所幸受到許多人的幫助,皆銘記於心。特此感謝 莊志遠先生提供必要之實驗器材以及魏恒巍老師提供實驗用豬欄,讓我的研究得 以順利進行。感謝系辦奕雯學姊和位育學長的關心,以及遇到挫折時的鼓勵。感謝 動物生殖技術研究室的學長姐們:李妍樺學姊、周志任學長、林育聖學長、張乃云 學姐、張家駿學長、張廉本學長、許慈宇學姊、陳碩甫學長、彭紹于學長、曾哲揚 學長、葛亦辰學長、鄭淳淳學姊、蕭冠宇學長於實驗技術之指導以及學習經驗分享;
感謝曾經與我共同奮鬥的同學們:王浩、林書毅、高藝榕、歐陽桓、魏可軒於實驗 和生活上互相幫忙和照顧。
最後,感謝我最摯愛的家人:爸爸、媽媽、弟弟,你們一直是我最忠實的聽眾,
共同分享且分擔著我的所有喜怒哀樂,謝謝你們的陪伴、支持和包容,讓我有繼續 堅持到最後的動力。另外感謝為此研究而犧牲奉獻的所有小豬。
謹誌于 國立臺灣大學動物科學技術學系 中華民國一百零六年一月
摘要
自體脂肪移植 (autologous fat grafting, AFG) 為採集自身的脂肪組織進行注射,
目前已經廣泛用於整形外科中之乳房重建;然而,AFG 之手術成果通常被視為不 可預測,因長期存留容積通常在 20%和 90%之間變化,其原因一般認為是移植後 的早期階段缺乏血液供應所導致的結果。外部體積擴張 (external volume expansion, EVE) 的負壓處理,長期以來被認為具有預先處理受體部位之潛能。此外,機械力 已被證實對於間葉幹細胞增殖和分化具有關鍵的作用。因此,為了探討 EVE 之體 內機械力作用對於皮下脂肪中的細胞增殖,血管生長和成熟以及脂肪幹細胞 (adipose stem cells, ASCs) 的細胞標記,增殖能力和分化潛能的影響,利用豬的皮 膚和皮下組織與人類之間在解剖和生理上相似性設計為動物模式。
本研究使用負壓 50 毫米汞柱,在每天相同 8 小時 (上午 9 點到下午 5 點) 的 時間間隔連續作用於豬隻側腹表皮直到 10 或 21 天。在第 11 或 22 天進行採樣之 前,使用超音波測量軟組織厚度,結果顯示 EVE 誘導軟組織擴增是屬於一種暫時 性 效 應 。 接 著 將 試 驗 組 和 對 照 組 所 取 得 之 樣 本 進 行 各 項 分 析 。 H&E 染 色 (hematoxylin and eosin staining) 結果顯示,EVE 可以促進血管重塑的過程,但是對 脂肪細胞的大小和數量並沒有顯著影響。Ki67 免疫組織化學染色的結果顯示,基 底角質細胞和脂肪細胞之細胞增殖相較於對照組沒有出現顯著差異;相反地,通過 α-SMA 染色得到證實,具有平滑肌細胞的血管網絡在 EVE 處理組中有明顯增加。
另外,使用 image J 測量表皮厚度的結果顯示各組之間並沒有觀察到顯著的差異。
CD31 免疫螢光染色結果顯示,血管密度會隨著 EVE 的作用時間而逐漸增加。
基質血管細胞 (stromal vascular fraction, SVF) 和 ASCs 分別從脂肪組織中分 離和純化。ASCs 的增殖能力透過倍增時間和集落形成分析進行檢定,結果卻顯示 對照組和 EVE 處理組之間沒有發現統計上的差異。ASCs 進行脂肪分化誘導 21 天,
隨後進行油紅染色 (Oil-Red O staining),結果 ASCs 的脂肪分化潛能在各組之中也 沒有顯著差異。流式細胞術分析結果顯示,無論處理時間長短,ASCs 皆會表現諸 如 CD29, CD44, CD90 與 CD105 等多種間葉細胞標記,惟缺乏表現造血細胞標記 如 CD34 之能力。多色流式細胞儀分析 SVF 細胞組成,結果顯示各組之間 ASCs
的比例沒有顯著差異;相較之下,在 EVE 處理組中的內皮細胞,其百分比,顯著 高於對照組者,且亦會隨著其作用時間之延長而顯著增加。
綜合上述,EVE 之主要作用機制,在於調節新血管網絡之形成、生長和功能
化血管的成熟。EVE 的預先處理效果已經在李宋豬模式中得到證實,此結果可以 輕易地轉化為臨床實踐,以提高細胞和組織移植存活率。期望本研究成果可以幫助 臨床醫生在手術前優化受體組織的血管分布,以進一步改善脂肪移植的存留容積。
關鍵詞:自體脂肪移植、外部體積擴張、脂肪幹細胞、基質血管細胞、脂肪容積 存留
ABSTRACT
Autologous fat grafting (AFG) has been widely used as an injectable substance for breast reconstruction in cosmetic surgery; however, the clinical outcome of AFG is generally considered an unpredictable procedure, with long-term retention commonly varying between 20 and 90%, which is believed to be caused by poor blood supply in the early after grafting. Negative pressure using external volume expansion (EVE) has long been theorized as a potential means to precondition the recipient bed. In addition, the mechanical force has been confirmed to play a pivotal role in mesenchymal stem cells proliferation and differentiation. Accordingly, to investigate the effects of in vivo mechanical loading of EVE on cell proliferation, vascular growth and subsequent maturation as well as cellular markers, proliferation capacity and differentiation potential of adipose stem cells (ASCs) in subcutaneous fat, a swine model was devised to take advantage of anatomical and physiological similarities in skin and subcutaneous tissue between pigs and human.
In this study, pigs were treated with continuous suction at -50 mmHg during the same eight-hour (9:00-17:00) interval each day until 10 or 21 days. Before sampling on day-11 or 22, an ultrasonography was performed to study the soft tissue thickness and results revealed that EVE-induced soft tissue enlargement is a transient effect. Specimens from control and treated groups conducted a various analysis. The result of H&E staining showed that EVE can enhance the process of vascular remodeling but has no significant effect on adipocytes size and numbers. IHC stain with Ki67 showed cell proliferation in basal keratinocytes and adipocytes did not appear significant difference as compared with the non-treated group; in contrast, vascular networks layered with smooth muscle cells
increased in EVE treated groups as evident by the α-SMA staining. On the other hand, the epidermal thickness was measured by image J but no significant difference was observed across the groups. Immunofluorescence stain with CD31 suggested that blood vessel density would gradually increase with the loading time of EVE.
Stromal vascular fraction (SVF) cells and ASCs were isolated and purified from fat tissue, respectively. Proliferation capacity of ASCs was measured by doubling time and colony-forming assay but no statistical difference was found between the control and EVE treated groups. ASCs were subjected to adipogenic induction for 21 days followed by Oil-Red O staining and adipogenic differentiation potential of ASCs had no significant difference across the groups. Flow cytometry analysis showed regardless of treatment interval, ASCs expressed mesenchymal markers such as CD29, CD44, CD90, CD105 while lacking expression of hematopoietic marker such as CD34. Multicolor flow cytometric analysis of SVF cells revealed no significant difference in the ratio of ASCs across the groups; in contrast, the percentage of endothelial cells of EVE treated groups significantly increased as treatment lengthened when compared with the control group.
In conclusion, the predominant mechanism of action of EVE, which would modulate neovascular network formation, growth and maturation of functional blood vessels. The preconditioning effect of EVE has been demonstrated in the swine model, which may be easily translated into clinical practices to enhance cell and tissue engraftment. It is expected that this understanding may help clinicians to optimize the vascularity of the recipient bed to further improve fat volume retention before the operation.
Key words: Autologous fat grafting, External volume expansion, Adipose stem cells, Stromal vascular fraction, Fat volume retention
目 次
頁次 口試委員審定書 ··· Ι 誌謝 ··· Ⅱ 摘要 ··· Ⅲ ABSTRACT ··· Ⅴ 目次 ··· Ⅶ 圖次 ··· Ⅹ 表次 ··· Ⅻ
第一章 緒論 ··· 1
第二章 文獻探討 ··· 4
2.1 皮膚的解剖構造 ··· 4
2.1.1 皮膚結構簡介 ··· 4
2.1.2 皮下組織 ··· 4
2.1.3 結締組織 ··· 5
2.2 脂肪組織 ··· 7
2.2.1 脂肪組織之簡介 ··· 7
2.2.2 脂肪組織之分類 ··· 7
2.2.3 脂肪組織之生理功能 ··· 9
2.2.4 脂肪組織之擴增方式 ··· 9
2.2.5 脂肪組織之細胞組成 ··· 11
2.3 自體脂肪移植 ··· 13
2.3.1 簡介 ··· 13
2.3.2 非血管化脂肪移植 ··· 13
2.3.3 脂肪細胞移植後之命運 ··· 15
2.3.4 改善脂肪移植存留容積之策略 ··· 18
2.4 力學生物學 ··· 21
2.4.1 簡介 ··· 21
2.4.2 機械力與細胞增殖 ··· 22
2.4.3 機械力與間葉幹細胞分化 ··· 25
2.4.4 機械力與血管形成 ··· 34
2.5 豬為動物模式於生物醫學之研究 ··· 40
2.5.1 動物模式之簡介 ··· 40
2.5.2 小型豬為動物模式之優點 ··· 40
第三章 試驗研究 ··· 43
3.1 外部體積擴張對軟組織厚度、細胞增殖和血管重塑之影響 ··· 43
3.1.1 前言 ··· 43
3.1.2 材料與方法 ··· 44
3.1.3 結果與討論 ··· 51
3.2 外部體積擴張對脂肪幹細胞增殖、分化潛能及其細胞標記之探討 ··· 59
3.2.1 前言 ··· 59
3.2.2 材料與方法 ··· 61
3.2.3 結果與討論 ··· 64
第四章 綜合討論 ··· 70
第五章 結論 ··· 73
第六章 未來展望 ··· 74
REFERENCE ··· 75
圖 次
頁次
圖 1. 皮膚層之解剖構造 ... 6
圖 2. 固有結締組織之細胞和纖維 ... 6
圖 3. 棕色脂肪組織與白色脂肪組織 ... 8
圖 4. 肥厚型與增生型脂肪細胞之特徵 ... 10
圖 5. 脂肪組織和基質血管細胞之細胞組成 ... 12
圖 6. 脂肪組織於缺血時之細胞活動 ... 14
圖 7. 脂肪組織移植後之細胞和分子事件 ... 15
圖 8. 移植物的三個區域和脂肪細胞在移植脂肪中的命運之結論模式 ... 17
圖 9. EVE 允許大容積脂肪移植之原理 ... 20
圖 10. EVE 建立有利於自體脂肪移植生存之受體環境 ... 20
圖 11. 機械力調節內皮細胞之功能 ... 21
圖 12. 細胞之張拉整體結構 ... 22
圖 13. 整聯蛋白協同生長因子受體調節細胞週期 ... 24
圖 14. 細胞微環境調控細胞之命運 ... 26
圖 15. 培養基質剛度影響間葉幹細胞分化之機制 ... 28
圖 16. 整合素內部化經由 BMP – Smad 路徑調節間葉幹細胞分化 ... 29
圖 17. 胞外基質的物理性質調節間葉幹細胞分化 ... 31
圖 18. 胞外基質的剛度經由改變細胞與核骨架的張力影響基因表現 ... 32
圖 19. 胞外基質的物理因素調節間葉幹細胞之分化命運 ... 32
圖 20. 內在與外在機械信號調節間葉幹細胞分化 ... 34
圖 21. 封閉式負壓引流主要作用機制 ... 37
圖 22. 封閉式負壓引流造成氧氣和 VEGF 濃度梯度引導血管生長 ... 38
圖 23. HIF-1α-VEGF 訊息路徑... 39
圖 24. 外部體積擴張裝置及其作用期間之示意圖 ... 45
圖 25. EVE 處理之試驗設計和流程圖 ... 46
圖 26. 免疫組織染色觀察血管重塑 ... 52
圖 27. 皮下脂肪中脂肪細胞大小對脂肪細胞數之標繪圖 ... 53
圖 28. 細胞增殖和表皮厚度之定量分析結果 ... 55
圖 29. 血管密度之定量分析結果 ... 56
圖 30. 血管成熟之定量分析結果 ... 58
圖 31. 成體幹細胞不同類型的分支 ... 60
圖 32. EVE 處理組與對照組中脂肪幹細胞增殖能力分析 ... 64
圖 33. EVE 處理組與對照組中脂肪幹細胞分化潛能分析 ... 65
圖 34. 流式細胞技術分析脂肪幹細胞之表面標記 ... 67
圖 35. 多色流式細胞儀分析基質血管細胞之細胞組成 ... 69
表 次
頁次 表 1. 豬作為生物醫學模式動物之優點 ··· 42 表 2. 超音波測量軟組織厚度 ··· 51
第一章 緒論
根據國際美容整形外科學會 (International Society of Aesthetic Plastic Surgery, ISAPS) 於 2016 年 7 月 25 日所發布之統計數據表示,2015 年全世界進行了超過 2100 萬次手術和非手術整形,其中女性進行了超過 1800 萬次整形,占整形總數 的 85.6%。最常見的女性手術整形項目之前五名分別為:豐胸 (breast augmentation)、
抽脂 (liposuction)、眼瞼手術 (eyelid surgery)、腹壁整形術 (abdominoplasty) 和隆 鼻手術 (rhinoplasty),因此就手術整形而言,豐胸在女性中相當受歡迎。
目前在醫學美容整形手術中,以自體脂肪移植 (autologous fat grafting, AFG) 進行豐胸或隆乳手術愈來愈受到歡迎。AFG 顧名思義就是從身上多餘的脂肪處,
例如腹部,側腰,臀部,大腿等處,以吸取的方式取得小直徑的脂肪球,經過離心 及純化後,再用針頭注入欠缺脂肪的部位,術後不易留下疤痕,且不會有交叉感染 和異物反應等問題。而自體脂肪豐胸,就是將處理過的脂肪均勻地注入到乳房的肌 肉層以及皮下層,從而創造出自然的乳房 (Schweizer et al., 2015)。
應用自體脂肪移植於整形手術最早在 1893 年 Neuber 已有文獻報告,1911 年 Bruning 首次將自體脂肪注射到皮下組織進行軟組織擴增,1950 年 Peer 的研究發 現脂肪移植的存活率可達 50%。1980 年代,抽脂手術 (liposuction) 開始盛行,當 時檢驗技術還未成熟,抽脂豐胸手術可能會妨礙到乳癌之診斷,因此 1987 年遭美 國整形外科醫師學會禁止。隨著新的診斷工具的發明和技術的進步,以及缺乏科學 文獻上的證據,直到 2009 年才解禁 (Khouri et al., 2012)。近年來應用自體脂肪移 植進行乳房重建和擴增以及軟組織修復等方面已被廣泛的運用,儘管脂肪組織取 得容易,可經由抽脂等低侵入性的方式大量取得,但植入至皮下的脂肪組織被吸收 率介於 20%至 90%,且長期生存率約 30%至 50%,其主要原因為受體部位缺乏足 夠的血液供應造成細胞處於局部缺血 (ischemia) 的情況 (Brucker et al., 2008;
Zhong et al., 2009)。而脂肪存活率的高低會影響手術的結果和後遺症的多少,若存 活率太低,壞死的脂肪細胞數太多,可能會造成鈣化和硬塊等問題。
以負壓進行外部體積擴張長久以來被視為組織再生與重建的一種方式,早於 1997 年 Argent 和 Morykwas 所設計的真空輔助閉合裝置 (vacuum-assisted closure device, VAC) 用於治療糖尿病患之創傷修復,並證實可有效刺激血管的再生以及 細胞的增殖 (Huang et al., 2014)。2000 年 Khouri 使用外部體積擴張 (external volume expansion, EVE) 的方式用於非手術性乳房擴增 (nonsurgical breast enlargement),
稱之為 Brava,其為一種外部負壓組織擴張裝置,標準使用方式為手術前 4~6 週每 天連續使用 8~12 小時。隨著 Brava 的改良,目前可根據患者的需求調整不同的負 壓強度和自動循環模式,大幅縮短整體作用時間並達成相同之預先處理效果 (Khouri et al., 2014)。經過 EVE 作用之後,可以促進血管新生,增加組織空間,減 少脂肪移植體的擁擠和注射時的壓力,使結果較為理想 (Khouri et al., 2012)。此外,
利用 EVE 於創傷修復和組織再生的優點,在於組織經負壓處理之後能夠接受比預 期更多的脂肪移植容積。當大量的脂肪移植到一個狹窄的空間時,組織間隙液壓 (interstitial fluid pressure) 的增加會減少微血管血流量和脂肪移植物與周圍組織的 接觸面積,導致脂肪細胞因無法獲取足夠的氧氣和養分而壞死。而 EVE 所產生的 機械力可在手術前用以增加移植部位的組織間隙以及血管新生,因此可一次移植 大量脂肪 (~250 ml) 並改善長期移植容積因缺血而流失的情況 (Khouri et al., 2014)。此脂肪移植大量容積 (mega-volume) 的成功使臨床試驗上可引用另一種新 的方法於乳房重建和擴增 (Khouri et al., 2015)。
當一項新技術首次被使用時,對於其結果的定量方式以及潛在的生理過程通 常有許多爭議 (Gundeslioglu, 2012; Herold et al., 2012)。以小鼠為動物模式所做的 研究顯示,在沒有脂肪移植的情況下,EVE 作用一段時間後能夠增加皮下組織的 血管密度以及脂肪細胞數 (Heit et al., 2012; Lancerotto et al., 2013)。然而從囓齒動 物模式所得到的實驗結果於臨床的實用性一直以來時常爭議不斷,並且在沒有脂 肪移植的情況下進行外部負壓處理上缺乏臨床經驗。而從人類和小鼠所得到的實 驗結果的差異,一直被歸因於兩者間先天上解剖構造,生理情況以及作用機轉的不 同所使然 (Swanson, 2014)。
新血管網路的形成對於脂肪移植的存活和組織再生而言十分重要。然而脂肪 移植的生存能力依賴於周圍組織養分的擴散和血管之新生 (neovascularization),特 別是在移植後之早期階段。而外部機械力的刺激對於血管網路的形成和血管重塑 的影響,顯示出血管新生的過程取決於周圍的環境和機械力刺激的強度 (Yung et al., 2009),因此使用負壓進行外部體積擴張對於提升移植部位的血管形成而言可以 說是相當具有潛力的方式。
先前已有研究證實,機械力會影響多種類型細胞之細胞表型 (phenotype)、生 長速率以及訊息傳遞;並且由這些結果也顯示機械力可以誘導間葉幹細胞分化為 成骨細胞以及平滑肌細胞 (Kurpinski et al., 2006; Park et al., 2004)。然而,經過 EVE 作用後的組織所分離出的脂肪幹細胞,迄今尚未對其增殖、分化能力以及表面標記 進行評估。
基於上述,本研究以小型豬為模式動物,探討 EVE 之機械力作用在生物體內 對於皮下脂肪之血管生成以及進一步成熟為具功能性血管的影響。首先假設機械 力作用在生物體內所調節之新血管生成和血管的重塑的成效,在於作用時間長短 之差異。因此,將 EVE 作用時間分為短時間 (10 天) 和長時間 (21 天),再對皮下 脂肪的血管生成和重塑進行定量評估。此外,比較 EVE 不同的作用時間對於小型 豬的脂肪幹細胞其細胞表型,增殖能力,脂肪分化潛能以及幹性 (stemness) 的影 響進行定量分析。
第二章 文獻探討
2.1 皮膚的解剖構造 2.1.1 皮膚結構簡介
皮膚結構在人類和大多數哺乳動物非常相似,雖然人類皮膚都覆蓋著毛囊,但 一般可將皮膚分為有毛和無毛兩種類型 (圖 1)。皮膚是由表皮和真皮所組成,表皮 (epidermis) 位於皮膚最外層,是由多層排列緊密的上皮細胞所組成,形成保護屏 障,可防止水分散失與細菌的侵入。真皮 (dermis) 位於表皮下方,由含有膠原纖 維 (collagenous fiber) 及彈性纖維 (elastic fiber) 的緻密結締組織組成,富含血管、
淋巴管、神經末梢、接受器、腺體和毛囊等。皮膚底下的皮下組織稱為皮下層 (hypodermis),是由疏鬆結締組織和脂肪組織所組成 (許, 2008)。
2.1.2 皮下組織的結構
皮下組織又稱為“皮下脂肪組織”,位於真皮下方,與真皮無明顯的界限,解剖 學上稱為淺筋膜 (superficial fascia),臨床上稱為蜂窩組織 (areolar tissue)。皮下脂 肪組織是一層比較疏鬆的組織,它是一個天然的緩衝墊,能緩衝外來壓力,同時也 是熱的絕緣體,能夠儲存能量。除脂肪外,皮下脂肪組織也含有豐富的血管、淋巴 管、神經、汗腺和毛囊。
皮下組織屬於間葉組織 (mesenchymal tissue),主要組成成分為脂肪細胞、纖 維間隔 (fibrous septa) 和血管。此外,皮下組織內尚分佈有淋巴管、神經、汗腺體 以及毛囊 (乳頭部)。脂肪細胞呈圓形或卵圓形,平均直徑約為 94 微米,大者可達 120 微米。細胞質內充滿脂質、少數粒線體和較多游離核醣體,細胞核擠向邊緣呈 扁平。脂肪細胞聚集,形成大小不一的脂肪小葉 (fat lobules),其間以纖維間隔為 界 (脂肪小葉間隔)。皮下組織內富有血管,由小葉間隔小動脈分支形成微血管,
伸入脂肪小葉並圍繞著每個脂肪細胞。微血管基底膜與脂肪細胞之細胞膜緊密接 觸,有助於血液迴圈和脂質的輸送。皮下組織分布於真皮和肌膜,上方與真皮、下
方與肌膜密接,廣布於體表,形成脂肪層,約占人類體重 18%。其厚度因體表部 位、年齡、性別、內分泌、營養和健康狀態等,而有明顯差異 (Marieb, 2009)。
2.1.3 結締組織 (connective tissue)
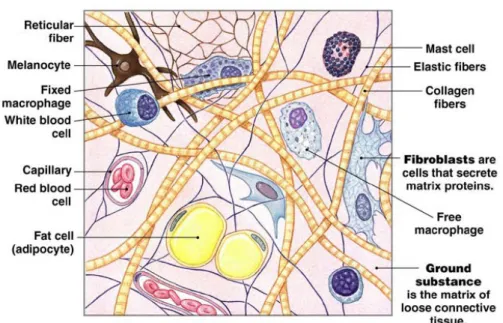
結締組織是體內含量最多,分布最廣的組織,外觀因種類不同而有很大差異,
但均有細胞、細胞基質 (ground substance) 及纖維 (fiber),所含的細胞均由胚胎期 之間葉幹細胞 (mesenchymal stem cells) 分化而來,依所含細胞和基質的種類或比 例可分為四大類:(1) 固有結締組織 (connective tissue proper):分為疏鬆結締組織、
緻密結締組織、脂肪組織、彈性結締組織和網狀結締組織,所含的主要細胞有纖維 母細胞 (fibroblasts)、巨噬細胞 (macrophages)、脂肪細胞 (adipose cells) 和肥胖細 胞 (mast cells) 等 (圖 2);(2) 軟骨 (cartilage):分為透明軟骨、纖維軟骨和彈性軟 骨;(3) 硬骨 (bone):其基質的密度和硬度均較高,支撐力強,是身體的主要的支 撐架構;(4) 血液 (blood):屬一種液狀的結締組織,由定型成分 (如紅血球、白血 球和血小板) 和血漿 (plasma) 所組成。其中除了軟骨不含血管外,結締組織均富 含血管且有充足的血液供應 (許, 2008)。
圖 1. 皮膚層之解剖構造。 (https://en.wikipedia.org/wiki/File:Skin_layers.png)
Figure 1. Anatomic structure of skin layers.
(http://asavory.edublogs.org/2012/11/05/connective-tissues/)
圖 2. 固有結締組織之細胞和纖維。
Figure 2. Cells and fibers of connective tissue proper.
2.2 脂肪組織 (adipose tissue) 2.2.1 脂肪組織之簡介
脂肪組織在組織學上屬於一種結締組織,主要由許多能儲存三酸甘油脂 (triglyceride) 的脂肪細胞 (adipocytes) 所構成,並被疏鬆結締組織分隔成許多脂肪 小葉。由於細胞質與核被大脂肪油滴擠到細胞邊緣,因此形成形狀像印章戒指 (signet ring)。脂肪細胞常與疏鬆結締組織在一起,尤其是在皮下層、腎周圍、心臟 底部與表面、長骨骨隨、關節周圍、眼眶內的眼球後等處,提供器官一般性的保護 和支持,同時也是良好的絕緣體,可減少體內熱量由皮膚散失,並有儲存脂肪,緩 衝機械性壓力,參與脂肪代謝等功能。脂肪組織能夠以旁泌、自泌和內分泌等方式 產生生物活性因子,稱為脂肪因子,例如肥胖賀爾蒙 (leptin)、抵抗素 (resistin)、
細胞激素 (cytokine) 及腫瘤壞死因子 (TNFα) 等,影響胰島素敏感性、血壓、內皮 功能、纖溶活動及發炎反應,參與多種重要病理與生理過程,因此脂肪組織不僅是 最大的能量儲存庫,並且是重要的內分泌系統 (Illouz and Sterodimas, 2011)。
2.2.2 脂肪組織之分類
脂肪組織是一種高度特殊的結締組織,根據脂肪細胞結構和功能不同,可分為 兩類:
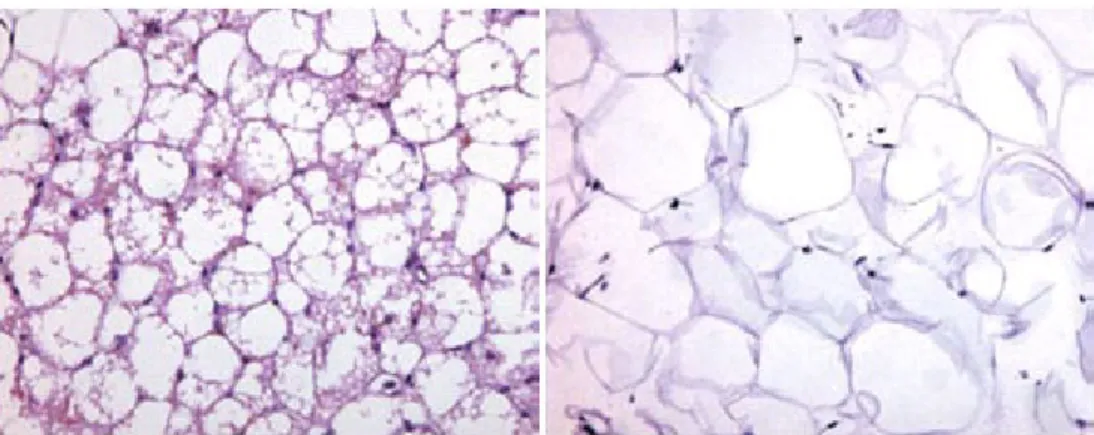
(1) 白色脂肪組織 (white adipose tissue, WAT),即通常所說的脂肪組織,由大量單 泡脂肪細胞聚集而成,脂肪細胞呈圓形或多邊形,細胞中央有一大脂滴,胞質呈薄 層,位於細胞邊緣,包繞脂滴。在 hematoxylin-eosin 切片上,脂滴被溶解成一大空 泡。細胞核呈扁圓形,被脂滴推擠到細胞一側,連同部分細胞質呈新月形 (圖 3)。
白色脂肪組織主要分布在皮下組織、網膜和腸系膜等處,在正常成年男子一般約占 體重的 10%~20%,女性通常更多一些。WAT 為體內最大的能源庫,具有貯存脂 肪、保持體溫、參與脂肪和能量代謝之功能,並具有產生熱能、維持體溫、緩衝保 護和支持填充等作用。白色脂肪組織之運作可受神經與內分泌系統之調控,以調節 其釋出或儲存之脂肪酸及甘油的速率與方向。
(2) 棕色脂肪組織 (brown adipose tissue, BAT) 之特點,乃其組織中含有豐富的微 血管;脂肪細胞內儲存許多個小脂滴,其細胞核呈圓形位於細胞之中央,且細胞質 內富含粒線體,此類脂肪細胞稱之為“多泡脂肪細胞”(圖 3)。棕色脂肪組織在成 年者極少,新生兒和冬眠動物者則較多;在新生兒者主要分布於肩胛間區、腋窩及 頸後部等處。棕色脂肪細胞在脂質氧化過程中不形成 ATP,而是將所含的能量以 熱能形式釋放,提供動物體溫與熱能之需求;特別對小型哺乳動物體溫之維持與動 物冬眠之甦醒分別扮演有極重要之角色。棕色脂肪組織的主要功能在於確保在寒 冷刺激下,棕色脂肪細胞內的脂類得以被分解及氧化,從而散發出大量能量而不轉 變為化學能,此功能受交感神經調節。人類之棕色脂肪組織從出生後到生命期之前 十年內會逐漸減少,而白色脂肪組織則相反,因此成為成人之主要脂肪組織 (Albright and Stern, 1998)。
(Virtanen et al., 2009) 圖 3. 棕色脂肪組織 (左) 與白色脂肪組織 (右)。
Figure 3. Brown adipose tissue (left) and white adipose tissue (right).
2.2.3 脂肪組織之生理功能
脂肪組織不僅一直被視為動物體之能量儲存站,且兼具絕緣功能與動物體重 要結構之機械支撐;直到 1990 年 Friedman 之研究團隊發現脂肪細胞會根據生物 體的營養狀態分泌一種蛋白質類細胞激素稱為 leptin。此後,更多研究所發現由脂 肪細胞所分泌的細胞激素、賀爾蒙和胜肽 (peptides),例如 adiponectin, resistin, retinol-binding protein 4, omentin 和 nesfatin 等由胜類組成的激素,統稱為脂肪因子 (adipokines);另由細胞中脂質的合成和降解之脂質代謝物 (lipid metabolites) 則稱 為 lipokines。因此,脂肪組織也被視為一種內分泌器,可參與調節止血、血壓、免 疫功能、血管生成的過程和生物體內能量平衡 (Rosen and MacDougald, 2006)。
2.2.4 脂肪組織之擴增方式
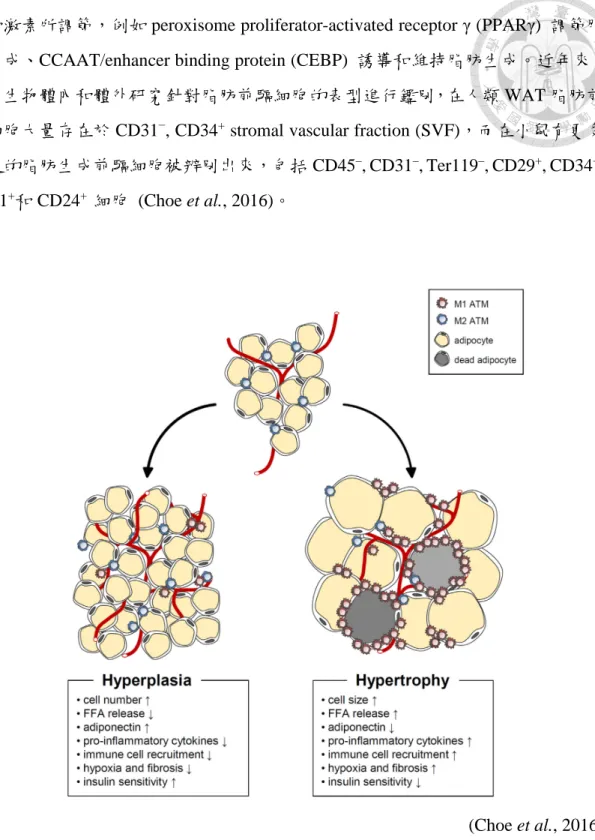
脂肪組織的擴增可經由脂肪細胞體積的增加 (hypertrophy) 以及脂肪細胞數 量的增加 (hyperplasia) (圖 4)。肥厚性脂肪細胞 (hypertrophic adipocytes) 具有下列 特點:(1) 在過度肥胖的脂肪組織中,肥厚的脂肪細胞具有壞死樣異常 (necrotic- like abnormalities),進而導致細胞的壞死。(2) 肥厚的脂肪細胞會增加促發炎反應 細胞激素 (pro-inflammatory cytokines) 的表現和分泌,包括腫瘤壞死因子 (tumor necrosis factor α, TNFα), interleukin (IL)-6, IL-8 以及單核細胞趨化蛋白 (monocyte chemoattractant protein-1, MCP-1),這些激素會經由 nuclear factor κB 和 Jun N- terminal kinase signaling,使 insulin receptor substrate-1 中的絲胺酸發生磷酸化 (serine phosphorylation),導致胰島素阻抗性 (insulin resistance) 的發展。此等促進 發炎反應細胞激素且會吸引各種免疫細胞,包括巨噬細胞和 T 細胞,導致脂肪組 織產生發炎反應。(3) 脂肪細胞的肥厚會造成相對的血液供應減少,而導致局部性 脂肪組織缺氧;當組織發生缺氧,血管生成因子以及發炎反應相關的基因表現量會 增加。此外脂質分解作用會提高,使大量的游離脂肪酸從脂肪組織中釋放出來,而 被 其 他 組 織 例 如 肝 臟 和 肌 肉 所 吸 收 , 導 致 異 位 性 脂 肪 堆 積 (ectopic lipid accumulation) 和 脂 肪 中 毒 (lipotoxicity) 。 相 對 而 言 , 脂 肪 組 織 的 增 殖 性 擴 增 (hyperplasic expansion),需由脂肪前驅細胞分化成脂肪細胞,此過程受許多轉錄因
子和激素所調節,例如 peroxisome proliferator-activated receptor γ (PPARγ) 調節脂 肪生成、CCAAT/enhancer binding protein (CEBP) 誘導和維持脂肪生成。近年來,
許多生物體內和體外研究針對脂肪前驅細胞的表型進行鑑別,在人類 WAT 脂肪前 驅細胞大量存在於 CD31−, CD34+ stromal vascular fraction (SVF),而在小鼠有更多 特定的脂肪生成前驅細胞被辨別出來,包括 CD45–, CD31–, Ter119–, CD29+, CD34+, Sca-1+和 CD24+ 細胞 (Choe et al., 2016)。
(Choe et al., 2016) 圖 4. 肥厚型與增生型脂肪細胞之特徵。
Figure 4. Characteristics of hypertrophic and hyperplasic adipocytes.
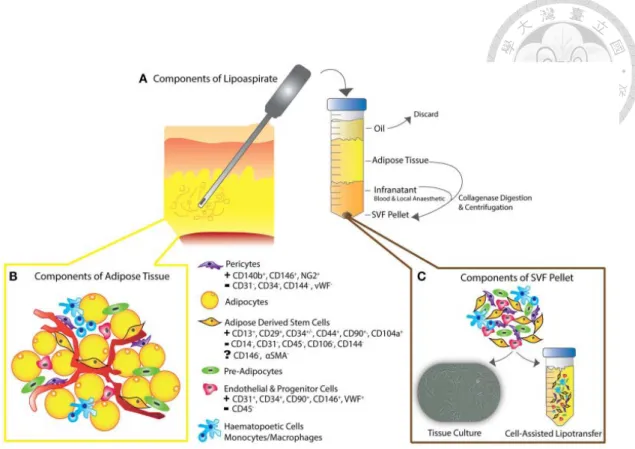
2.2.5 脂肪組織之細胞組成
脂肪組織主要由脂肪細胞、脂肪幹細胞、血管內皮細胞、血管外膜細胞 (pericytes)、纖維母細胞、巨噬細胞和胞外基質等所組成 (Yoshimura et al., 2006)。
脂肪細胞由於直徑介於 50~130 μm,故佔組織體積 90%以上,但以細胞數量而言卻 低於 50% (Suga et al., 2008)。脂肪組織中富含微血管網,脂肪細胞由膠原蛋白固定 在基質 (stroma) 中,每一個脂肪細胞至少和一條微血管接觸,儘管脂肪細胞具有 較大的體積,使得微血管彼此相距相當大的距離 (Gomillion and Burg, 2006)。
將取出的脂肪組織去除血水後,再經由膠原蛋白酶 (collagenase) 進行分解後,
可以將脂肪細胞從脂肪組織中與其它類型的細胞分離出來形成細胞團塊 (cell pellet),經此過程脂肪細胞被瓦解到油脂中,然後經過離心後將懸浮的組織包含脂 肪細胞和油液去除。沉澱於離心管底部的細胞團塊稱為基質血管細胞 (stromal vascular fraction, SVF),其中主要包含血管內皮伴隨著基質細胞 (stromal cells) 和 壁細胞 (mural cells)。存在於 SVF 中的細胞為中胚層或間葉細胞來源的細胞,包 括:脂肪前驅細胞 (pre-adipocytes)、纖維母細胞、內皮細胞、血管平滑肌細胞 (vascular smooth muscle cells)、免疫細胞 (immune cells) 和脂肪幹細胞 (Shukla et al., 2015) (圖 5)。在 基質血管細胞中,同 時也包含大量源自循環血液之細胞 (circulating blood-derived cells) 例如白血球和紅血球,雖然大部分之紅血球可經由 低滲溶液進行破壞。剛分離出來的脂肪幹細胞可經由表現出 CD31- CD34+ CD45- CD90+ CD105- CD146- 的細胞進行鑑定,但培養時會另外表現 CD105+ (Yoshimura, 2006)。而經由抽脂所取得之 SVF 中的有核細胞包含 37%的白血球 (CD45+),35%
脂肪幹細胞 (CD31- CD34+ CD45-),15%的內皮細胞 (CD31+ CD34+ CD45-) 和其它 細胞 (CD31- CD34- CD45-),儘管這些血液來源的細胞所佔的百分比會根據個體出 血情況而有明顯的不同 (Suga et al., 2008; Yoshimura et al., 2009)。
(Shukla et al., 2015) 圖 5. 脂肪組織和基質血管細胞之細胞組成。
Figure 5. Cellular compositions of adipose tissue and stromal vascular cells.
2.3 自體脂肪移植 2.3.1 簡介
脂肪組織可視為一種自體填充物 (autologous filler),目前已廣泛運用於整形、
美容和重建手術 (Mailey et al., 2013)。相較於其它人工填充材質,自體脂肪具有取 得容易、高生物相容性、可根據需求重複採集及外觀呈現自然等優點 (Khouri et al., 2014)。脂肪組織具有其代謝活性,由異質細胞群 (heterogeneous cell population) 組 成,可分泌細胞因子、激素和生長因子,故脂肪移植可通過添加複能性幹細胞 (multipotent stem cells) 和刺激因子來改變組織的微環境 (Largo et al., 2014)。脂肪 組 織 可 視 為 一 種 動 態 填 料 (dynamic filler) , 其 具 有 兩 種 不 同 的 補 充 效 應 (supplementary effects) , 包 括 : 體 積 效 應 (volumetric effect) 和 再 生 效 應 (regenerative effect);前者用於修復軟組織的缺陷,後者則能促進受體部位令其逐 漸回復自然的外觀 (Mojallal et al., 2009; Rigotti et al., 2016)。
脂肪組織具有高度血管化之能力,遂能促使脂肪細胞周遭被圍繞形成高密度 之微細管網絡。因此,脂肪移植最重要的是受體組織需有充足之血液供應。然而,
移植的脂肪組織由於缺乏血管形成,導致組織的再吸收,正因如此,臨床上自體脂 肪移植之手術結果非常難以預測,其長期存留容積一般在 20~90%之間變化 (Cherubino and Marra, 2009)。
此外,自體脂肪移植於臨床應用尚有許多問題需要克服,例如脂肪細胞無法忍 受低氧環境 (Rueda-Clausen et al., 2011);移植的脂肪組織容易液化 (liquefied),其 原因可能係由於缺氧或環境變化所使然 (Eirin et al., 2012);鑒於脂肪移植物常由 於被組織吸收,遂會隨著時間之進展而失去其體積約 20%到 90%之譜 (Cherubino and Marra, 2009);即使大部分脂肪組織或大量脂肪細胞用於移植,但業經老化之脂 肪細胞已不具再生能力遂會被逐漸吸收而消失 (Chien et al., 2012; Huang et al., 2016)。
2.3.2 非血管化脂肪移植
非血管化脂肪移植 (nonvascularized fat grafts, nVFGs) 乃指從供體部位採集獲 得之脂肪,經離心去除其上層之油脂及下層之紅血球細胞後,即可將剩餘之脂肪組
織進行注射到受體部位 (Serra-Renom et al., 2010)。鑒於移植之脂肪組織並未添加 任 何 生 長 因 子 或 組 織 細 胞 協 助 其 建 立 血 管 系 統 , 故 又 稱 傳 統 的 脂 肪 移 植 (conventional fat grafting) (Mailey et al., 2013)。
脂肪細胞對局部缺血非常敏感,在移植後的早期階段,氧氣是細胞存活所需的 關鍵分子;特別在低氧分壓 (pO2 = 15 mmHg) 將導致脂肪細胞於 24 小時內死亡。
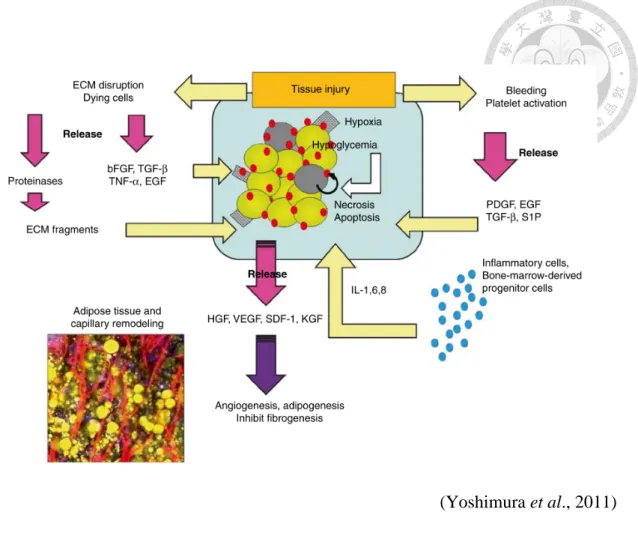
隨著缺血的時間延長,內皮細胞也開始死亡。相較之下,脂肪幹細胞可存活長達 3 天,在該 3 天之內,脂肪幹細胞會被活化,並通過其再生能力協助其組織之再修 復,且可協同免疫細胞促進其血管和脂肪之生成 (Suga et al., 2010) (圖 6)。
脂肪移植會導致受體組織的損傷,因此受體組織的出血會激活血小板釋放各 種生長因子,進而激活幹細胞或前驅細胞。同時,受傷組織以及死亡細胞會釋放 bFGF 和其它因子,來刺激脂肪幹細胞釋放 HGF 等生長因子,進而促進血管和脂 肪的生成,並且抑制纖維化產生。而組織損傷會進一步誘導發炎細胞的浸潤,並且 釋放各種炎性細胞因子 (圖 7)。大多數的已分化細胞會在移植後死亡,但駐留的幹 細胞和前驅細胞會活化,或從宿主的骨髓聚集過來。部分死亡細胞在血管成功形成 後會被幹細胞或前驅細胞所分化的下一代細胞替換 (Yoshimura et al., 2011)。
圖 6. 脂肪組織於缺血時之細胞活動。 (Suga et al., 2010) Figure 6. Cellular events in ischemia tissue.
(Yoshimura et al., 2011) 圖 7. 脂肪組織移植後之細胞和分子事件。
Figure 7. Cellular and molecular events after adipose-tissue grafting.
2.3.3 脂肪細胞移植後之命運
移植後之脂肪細胞,其存活率,主要係取決於受體組織之微環境,包括:血管 分布和氧氣供應;另外脂肪油滴的大小會影響移植物和周邊組織的接觸面積,因為 移植的脂肪在早期缺乏血管的支持,因此必須通過附近的微血管藉由擴散作用來 取得氧氣和養分,直到新生血管之形成,所以移植物的整體表面積大小勢必會影響 細胞的生存 (Yazawa et al., 2006)。
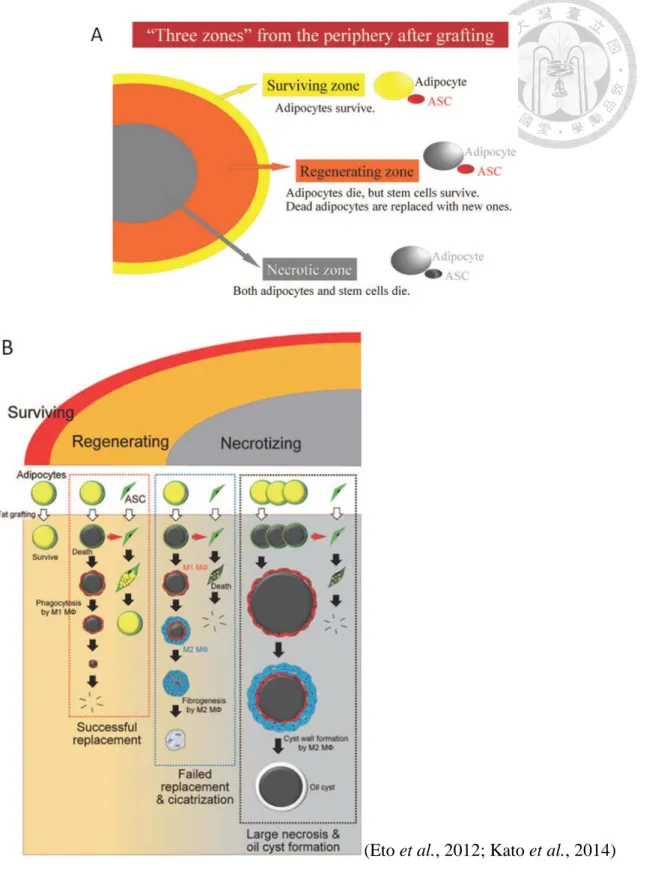
根據移植組織中的氧氣濃度是否足以供應脂肪細胞和脂肪幹細胞 (adipose derived stem cells, ASCs) 的存活,可將移植組織分為三個區域:存活區、再生區和 壞死區 (圖 8)。最表面的區域為存活區,厚度約 300 μm;其中位於存活區者最容
易獲得氧氣,因此脂肪細胞和 ASCs 皆可存活;第二區為再生區,其厚度根據微環 境條件,例如周圍組織的血管分布和氧氣供應而變化,在此區域,脂肪細胞早在第 1 天就已死亡,而 ASCs 會被鄰近的死亡細胞激活並產生新的脂肪細胞,死亡的細 胞最終被 M1 macrophage 所吞噬,並成功被新細胞替換;最中心的區域為壞死區,
其中脂肪細胞和 ASCs 皆死亡,最終停止其再生之過程。彼等死亡之細胞中殘存之 油滴,經由 M1 macrophage 吸收並且由 M2 macrophage 作用形成纖維化而替換之 (Eto et al., 2012)。
關於脂肪移植後之長期容積存留有兩個主要論點,包括:(1) The cell survival theory:係假設移植初期之脂肪細胞由受體部位組織經擴散作用取得血液供應直到 血管生成,故最終存活的脂肪細胞為最初移植之細胞 (Peer, 1950);與 (2) The host replacement theory:係假設移植之脂肪細胞全部死亡,作為供體與受體組織中與血 管生成相關之前驅細胞的基質,並誘導 ASCs 增殖和分化,故最終存活的脂肪細胞 為 ASCs 產生之新細胞 (Dolderer et al., 2011; Eto et al., 2012; Kato et al., 2014)。
Kato et al. (2014) 曾基於小鼠研究成果為脂肪移植後的續發事件做一總結,指 稱:除了位於組織表面300 μm 內的脂肪細胞外,幾乎所有脂肪細胞在脂肪移植後 的最初幾天內死亡。脂肪生成由活化的脂肪幹細胞和前驅細胞開始,並在 3 個月 內完成,而死亡脂肪細胞殘餘的脂肪油滴在接下來的 9 個月內被吸收。部分死亡 的脂肪細胞在前 3 個月會被新的脂肪細胞替換。脂肪油滴 (lipid droplets) 會被巨 噬細胞經吞噬作用吸收,但是非常緩慢,且吸收時間取決於脂肪油滴的直徑。若脂 肪油滴直徑過大,會在完全吸收之前形成囊壁,並且隨時間而鈣化。在移植物的中 心區域,因脂肪幹細胞和前驅細胞死亡而不形成新的脂肪細胞,導致瘢痕組織或囊 腫形成。脂肪移植後的最終存留容積由脂肪細胞的成功替換速率決定,若僅移植具 有小脂滴的脂肪,則可在 3 個月內完成吸收,且體積在 3 個月後基本上不會改變。
如果 3 個月後仍存在許多大脂滴,則脂肪組織會在 3 到 12 個月之內萎縮 (Kato et al., 2014) (圖 8)。
(Eto et al., 2012; Kato et al., 2014) 圖 8. 移植物的三個區域和脂肪細胞在移植脂肪中的命運之結論模式。
Figure 8. (A) Conclusive schema for three zones of the grafts. (B) Conclusive schema for the fate of adipocytes in grafted fat.
2.3.4 改善脂肪移植存留容積之策略
為了減少壞死區,可以經由製備具有更好活力和適當大小的移植物,通過理想 分布使移植物之接觸面積最大化,以及將移植物置於具有高血管分布之受體組織 中,因此提高脂肪生存主要有兩種策略:
一、脂肪移植物的優化 (optimization of the fat grafts),常見的有兩種方式:
(1) 細胞輔助脂肪移植 (cell-assisted lipotransfer, CAL),其利用 ASCs 具有分 化為脂肪細胞和血管內皮細胞的再生能力,以及 ASCs 因組織損傷或缺氧誘導分泌 血管生成因子例如 HGF 和 SDF-1 (stromal derived factor-1) 幫助血管生成,所以科 學家額外添加 ASCs 或 SVF 來充實脂肪組織,並以脂肪組織作為移植後支持 ASCs 生存之生物骨架 (Yoshimura et al., 2009)。但是,最近在臨床應用上卻發現幾個問 題:ASCs 缺乏端粒酶,且其端粒較短,因此只能在體外的有限時間內增殖 (Zhao et al., 2012);從老年患者分離的 ASCs 具有較低的增殖能力和穩定性,因此需要使 用年輕個體的 ASCs 進行同種異體移植 (Pandey et al., 2011; Sharpless and DePinho, 2007)。另外,如何在體外和體內調節 ASCs 的增殖和分化過程、哪些因子調控 ASCs 的增殖和分化、調節 ASCs 的增殖和分化過程的主要因素、哪些因子刺激 ASCs 分 泌旁分泌因子、移植的 ASCs 是否致瘤、什麼原因導致 ASCs 在體內液化,這些問 題都需要在 CAL 廣泛應用於臨床之前更深入探討 (Huang et al., 2016)。
(2) 高濃度血小板血漿 (platelet-rich plasma, PRP),其再生潛能主要取決於血 小 板 活 化 時 分 泌 之 蛋 白 , 包 括 血 管 內 皮 生 長 因 子 (VEGF) , 表 皮 生 長 因 子 (epidermal growth factor, EGF),血小板衍生生長因子 (PDGF),轉化生長因子 (TGF- 1),TGFβ2 和類胰島素生長因子 (IGF-1) (Nakamura et al., 2010; Oh et al., 2011)。
PRP 的取得方式簡單來說,是通過全血 (未除去任何成分的血液) 離心之後獲得高 於正常值 5~7 倍濃度的自體血小板,懸浮在小體積血漿中。PRP 在使用前,必須 先用氯化鈣(CaCl2)和血漿凝血酶 (plasma thrombin) 來激活血小板觸發α- granules 的去顆粒作用 (degranulation),然後釋放各種生長因子,包括血管相關的生長因子 來促進血管生成 (James et al., 2016)。但是,最近的試驗卻發現,使用添加 PRP 的 脂肪會導致顯著的發炎反應以及不良的血管變化,例如血管基底膜會重複複製
(Rigotti et al., 2016)。因此,儘管有研究表示利用 EGF,纖維母細胞生長因子 (fibroblast growth factor, FGF),以及 PRP 來改善脂肪移植的存活率並可達到理想 的效果 (Oh et al., 2011; Park et al., 2011; Tamura et al., 2007),但是這些方法的缺點 在於需要有專業的實驗室以及繁複的處理步驟,且目前對於臨床試驗以外的採用,
尚未充分對其利益和風險進行評估。因此,PRP 在大規模使用在人體試驗之前,其 中安全性和功效還需要進一步證實和確認。
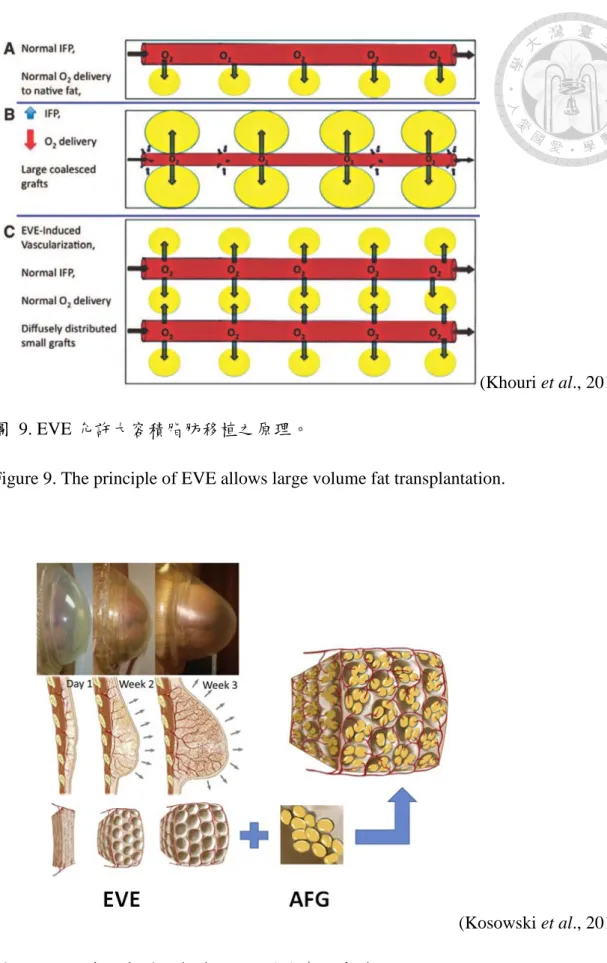
二、手術前受體組織的預先處理 (preconditioning the recipient bed),做法是利用外 部擴張裝置 (EVE) 的負壓處理作用在受體部位。目前已經有許多研究證實機械力 具有刺激細胞再生的能力,因此可利用外部機械力的作用來增加受體部位的組織 間隙空間以及血管生成,進而允許大量體積的脂肪移植,而不會減少移植物與受體 交界面積或增加組織間隙的液體壓力 (interstitial fluid pressure, IFP)。如果沒有經過 術前 EVE 之處理,而移植大量體積的脂肪到緊密的組織空間當中,IFP 會上升而 減弱微血管的血流量,進而減少氧氣輸送,導致新血管形成之前移植組織發生中心 壞死 (Khouri et al., 2014) (圖 9)。所以在臨床應用上,經過 3 週的外部體積擴張之 後,可以產生具有豐富微血管網絡所形成的生物骨架,以及充分的組織空間,提供 自體脂肪移植良好的受體環境,來提高細胞的存活 (Kosowski et al., 2015) (圖 10)。
綜觀上述,EVE 簡單執行就能達到良好的效果。研究表明負壓療法 (negative pressure therapy) 可產生許多效果,其中最吸引人的就是能促進血管生成 (Heit et al., 2012)。負壓作用時導致組織暫時性缺氧,可刺激 HIF-1α/VEGF 分子路徑,進 而促進血管生成 (Erba et al., 2011; Lancerotto et al., 2013; Lee et al., 2015)。
(Khouri et al., 2014) 圖 9. EVE 允許大容積脂肪移植之原理。
Figure 9. The principle of EVE allows large volume fat transplantation.
(Kosowski et al., 2015) 圖 10. EVE 建立有利於自體脂肪移植生存之受體環境。
Figure 10. EVE creates a larger and more favorable recipient site for AFG surviving.
2.4 力學生物學 (mechanobiology) 2.4.1 簡介
生物力學 (biomechanics) 泛指研究生物體的力學現象,例如肢體運動、神 經肌肉控制、呼吸運動等。隨著科學技術的進步逐漸深入到細胞分子層次,形 成新的學科稱為力學生物學 (mechanobiology),其主要於探討受力環境下細胞 的 感 知 與 細 胞 內 力 學 訊 息 傳 遞 之 分 子 機 制 , 稱 為 機 械 力 傳 導 (mechanotransduction),對於生物體生理、發育和疾病的影響 (David Merryman and Engler, 2010)。例如內皮細胞的力學感應器 (mechanosensors) 受機械力刺激,
以及細胞受體和化學配體結合,皆可經由訊息傳遞路徑活化下游的轉錄因子導 致基因和蛋白質的表現,進而調控細胞的功能包括遷移、增殖、分化、凋亡及 代謝等 (Chien, 2007) (圖 11)。
(Chien, 2007) 圖 11. 機械力調節內皮細胞之功能。
Figure 11. Mechanical forces modulate the functions of endothelial cells.
2.4.2 機械力與細胞增殖
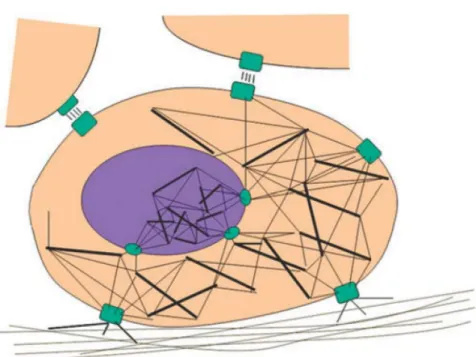
早 期 Folkman 和 Moscona 觀察 到 細胞形 狀 是決定細胞 功 能的關鍵因 素 (Folkman and Moscona, 1978)。細胞膜上的黏著斑 (focal adhesions, FAs) 主要由穿 膜蛋白組成例如整聯蛋白 (integrins),將細胞內的細胞骨架和胞外基質以及鄰近細 胞作連接,為機械力傳遞的主要路徑。其中細胞骨架的微小管為堅硬的支架 (rigid struts),微細絲和中間細絲為彈性元件 (elastic elements),這些組件之間拉伸和壓縮 力的平衡用以維持細胞完整的立體結構,稱為張拉整體 (tensional integrity or tensegrity) (圖 12),一旦有外力干擾此平衡使細胞內與細胞骨架連結的胞器和分子 移動,就會改變細胞的型態,進而影響細胞的功能 (Ingber, 2003)。此外細胞核的 結構同樣根據 tensegrity 原理,其內部架構經由核膜與細胞骨架緊密連結,機械力 的刺激會直接傳遞到 DNA 影響基因的轉錄 (Buxboim et al., 2010)。
(Ingber, 2003) 圖 12. 細胞之張拉整體結構。
Figure 12. The tensegrity structures of cells.
Integrins 是細胞膜上的醣蛋白受體,由 18 個α 和 8 個 β 次單位所組成的 24 種 heterodimer receptors,調節細胞與胞外基質的黏附與溝通 (Legate et al., 2009)。在 訊息傳遞中,integrins 能感知周圍環境的化學成分和力學狀態,並將訊息傳遞至細 胞內細胞骨架之組成蛋白 (e.g. actomyosin)、酵素 (e.g. tyrosine kinase) 和銜接蛋白 (adaptor protein) 而形成 integrin adhesion complex (IACs),進而調節細胞的功能特 性,包括型態、運動、增殖、分化等 (Ross et al., 2013)。近十年研究發現細胞週期 除了經由生長因子調節啟動細胞增殖,整合素黏附 (integrin adhesion) 也是多細胞 動物細胞增殖的必要條件,若細胞與胞外基質分離或基因剔除 integrin 次單位,即 使有生長因子的存在仍無法進入細胞週期 (Jeanes et al., 2012; Nikolopoulos et al., 2005; Streuli and Akhtar, 2009)。Integrins 協同生長因子受體 (growth factor receptors, GFRs),經由 PI3K/Akt 和 Mek/Erk 路徑促進 S 期所需 cyclin/Cdk complexes 形成,
並抑制及降解 cyclin-dependent kinase inhibitors (CKIs) 例如 p27 和 p21;另外 integrins 經由 Rac 活化的下游路徑調節細胞的增殖 (Moreno-Layseca and Streuli, 2014) (圖 13)。
體外培養的細胞,常在細胞膜與細胞質之特定區域,以及胞外基質之間形成緊 密黏附之黏著斑 (focal adhesions, FAs),在 FAs 的細胞質內側常有許多機動蛋白絲 (actin filaments) 排列成束,稱為應力纖維 (stress fibers, SFs) (Tojkander et al., 2012)。
SFs 的結構組成除微細絲之外,還包括肌動球蛋白 (actomyosin)、肌球蛋白 (myosin II)、輔肌動蛋白 (α-actinin) 和其它細胞骨架之組成蛋白 (Burridge and Wittchen, 2013)。在應力纖維中,機械力傳遞發生在細胞與胞外基質以及鄰近細胞之黏合處,
分別主要由 integrins 和 cadherins 組成。當機械力由細胞外往細胞內移動時,可經 由活化黏合處之組成蛋白,進而啟動細胞內的訊息傳遞改變細胞行為例如型態發 生 、 分 化 和 胞 質 分 裂 (cytokinesis) 等 (Chen et al., 2004; Horton et al., 2016;
Tojkander et al., 2012)。在黏著斑的組成蛋白中,p130Cas (Crk-associated substrate) 會直接受機械力影響發生磷酸化進而活化下游因子 Rap1 (Sawada et al., 2006);talin 則會改變構型而揭露出其它蛋白之接合位例如 vinculin,從而將機械作用力轉變為 化學訊息 (Kanchanawong et al., 2010)。另外有某些 actin filaments 從黏著斑連接至 細胞核周圍形成 actin cap associated focal adhesion (ACAFAs),由此形成之 actin cap
fibers 藉由核骨架和細胞骨架連接的複合體 (LINC complexes) 直接與核膜連結,
說明機械力可能經由此物理路徑 (physical pathway) 調節基因的表現 (Kim et al., 2012)。
(Moreno-Layseca and Streuli, 2014)
圖 13. 整聯蛋白協同生長因子受體調節細胞週期。
Figure 13. Integrins cooperate with growth factor receptors in cell cycle regulation.
2.4.3 機械力與間葉幹細胞分化
間葉幹細胞 (mesenchymal stem cells, MSCs) 屬複能性幹細胞 (multipotent stem cells),具自我更新 (self-renew) 能力,可分離自骨髓、脂肪、肌腱、周邊血 液、骨骼肌等組織,並可體外誘導分化為源自中胚層的細胞系包括脂肪細胞、成骨 細胞、軟骨細胞、心肌細胞和平滑肌細胞等,此外也可轉分化為其它胚層細胞,例 如源自外胚層的神經細胞以及源自內胚層的肝細胞和胰臟細胞 (Zhang et al., 2012)。
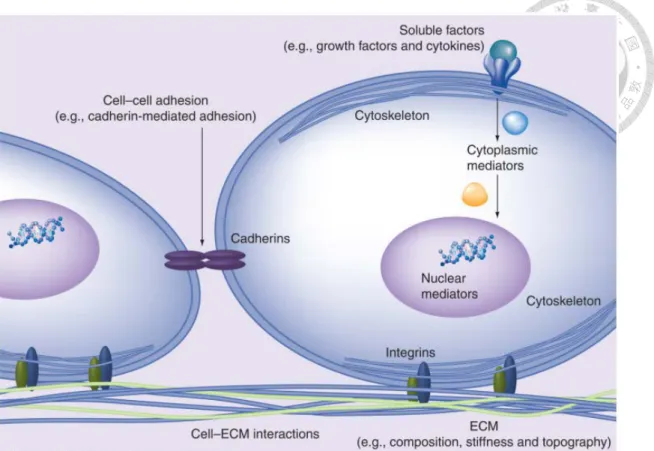
幹細胞之分化命運決定於細胞所存在之微環境,包括可溶性因子、細胞外基質 (extracellular matrix, ECM) 以及相鄰細胞之間的相互協調作用,可提供細胞能夠增 殖、存活、遷移或分化的生物化學和機械訊號。此外,表面粘附受體,例如整聯蛋 白 (integrins) 和鈣粘素 (cadherins),可分別調節細胞黏附於 ECM 骨架與鄰近細胞 (Li et al., 2011) (圖 14)。在可溶性因子當中,transforming growth factor-β (TGF-β)、
platelet-derived growth factor (PDGF) 和 fibroblast growth factor (FGF) 作為激活素 (activin) 所調節的三種分子訊號參與 MSCs 分化之關鍵途徑 (Ng et al., 2008)。ECM 物理性質包括剛度 (stiffness)、構型 (topography) 和組成 (composition) 可調節細 胞行為。此外,細胞會因應來自微環境中的訊息而採取不同的形狀,進而將產生的 牽引應力傳遞到相鄰細胞。若施加外部之機械力刺激例如流體剪切應力,可激活細 胞表面的離子通道、異源三聚體 G 蛋白 (heterotrimeric G proteins)、蛋白激酶和其 它膜上之訊息傳遞分子,進而觸發下游之訊息傳遞,導致機械力依賴性 (force- dependent) 之基因表現的變化 (Wang et al, 2009)。
(Li et al., 2011) 圖 14. 細胞微環境調控細胞之命運。
Figure 14. The cellular microenvironment control cell fate.
2.4.3.1 基質剛度引導間葉幹細胞之分化命運 (Substrate stiffness directs MSC fate specification)
從纖維母細胞和其它細胞之力學研究已顯示出細胞感測到 ECM 彈性性質的 重要性 (Paszek et al., 2005)。Engler 研究團隊第一次嘗試評估基質剛度 (stiffness) 對於調節人類間葉幹細胞 (hMSCs) 命運的重要性 (Engler et al., 2006)。他們生成 塗有膠原蛋白之聚丙烯酰胺凝膠 (polyacrylamide gel, PAAG) 作為體外細胞附著 之人工基質,而基質的彈性性質根據化學交聯的程度,從軟性到相對剛性。當培養 基保持相同時,測試各種彈性對 hMSCs 的影響。結果發現細胞命運由基質剛度決 定:當細胞培養於模擬腦組織彈性 (0.1-1 kPa) 的基質時,細胞會表現出神經元表
型;於模擬肌肉之中等剛度 (8-17 kPa) 的基質會表現出肌原性抗原;而於模擬膠 原骨 (collagenous bone) 之相對高剛性 (25-40 kPa) 的基質會表現成骨細胞之抗原 (Engler et al., 2006)。另以微陣列分析 (microarray analysis) 結果顯示各細胞在相應 之基質上,神經元,肌肉或骨骼之表面抗原的表現可增加四至六倍。然而,相較於 分化誘導培養基,此三種細胞之特殊標記表現水平於人工基質僅有其 50%。因此,
儘管基質彈性不足以誘導終末分化,但對於發育早期引導 MSCs 之細胞特化相當 重要。關於分子機制,Engler 等人表示非肌肉肌球蛋白 (non-muscle myosin II, NMMII) 的抑制會阻斷基質彈性所誘導之細胞特化,但不影響細胞其它正常功能,
表示基質剛度可透過不同機制調控定向分化 (directed differentiation) (Engler et al., 2006)。另有研究證實當 MSCs 和基質剛度相似性達最大值時,NMMII 於應力纖維 中的排列非專一性依賴於基質剛度,說明機械耦合 (mechanical coupling) 存在於 外部環境和細胞內部骨架組織之間 (Zemel et al., 2010)。因此微環境剛度調控細胞 特化之分子訊息傳遞尚需更深入探討。
ECM 剛度對 MSCs 細胞特化的影響在更多生理相關之 3D 培養系統進行評估。
Huebsch 等人將小鼠 MSCs 封裝培養於 3D 水凝膠合成之 ECM (Huebsch et al., 2010)。
在此培養模式中,基質剛度對 MSCs 表型具有較顯著之影響,其中成骨分化主要 發生在中等彈性 (11-30 kPa),而脂肪分化主要在較柔軟 (2.5-5.0 kPa) 微環境中。
雖然基質剛度對 MSCs 的效應大致上與先前所提及之 2D 培養模式一致,但是 3D 培養基質剛度可調節整合素 (integrins) 的結合以及粘附配體的重組,兩者對於 MSCs 都具有牽引依賴性以及成骨分化相關性 (Engler et al., 2006)。更重要的是,
基質剛度不僅調節細胞特化,並且可調節成體幹細胞之自我更新。舉例而言,在高 剛度培養基 (〜106 kPa) 所培養的肌肉幹細胞會失去其幹性 (stemness),導致前驅 細胞的再生能力大幅下降;相較之下,將模擬肌肉自身彈性 (12 kPa) 的柔軟基質 上培養的肌肉幹細胞隨後移植到小鼠,其自我更新和肌肉再生能力大幅提升 (Gilbert et al., 2010)。此項研究結果再次證實,基質剛度可調控各種幹細胞之分化 命運。
基質彈性影響 MSCs 成骨分化是經由 integrin-α2/ROCK/FAK/ERK1/2 路徑 (Shih et al., 2011) (圖 15)。基質上 ECM 彈性 (elasticity) 對 MSCs 產生機械性刺激,
從而誘導 FAs 蛋白活性和重塑之變化 (Du et al., 2011)。FAs 的生長和延長取決於 培養基質的剛度,表示 ECM 彈性可調節 FAs 聚集 (assembly)。Integrins 為細胞重 要之力學感應器 (mechanical sensors),MSCs 於柔軟培養基誘導β1 integrin 活化的 程度明顯高於堅硬培養基,但是位於細胞膜上的 integrins 卻明顯低於堅硬培養基,
其原因是柔軟的 ECM 會明顯提高 integrin 之內部化 (internalization),此現象主要 經由 caveolae/raft-dependent 內吞作用 (endocytosis)。Integrin 內部化會抑制 bone morphogenetic protein (BMP) − Smad pathway 而誘導 MSCs 往神經細胞分化;若使 用 caveolae/raft 的抑制劑:methyl- β- cyclodextrin,將會抑制 integrin 內部化進而阻 礙 MSCs 於柔軟基質上分化為神經細胞。柔軟基質抑制 BMP − Smad pathway 部分 是經由 integrins 調節之 BMP 受體的內吞作用 (Higuchi et al., 2013) (圖 16)。
(Shih et al., 2011) 圖 15. 培養基質剛度影響間葉幹細胞分化之機制。
Figure 15. The mechanism of the effect of culture substrate stiffness on MSCs differentiation.
(Higuchi et al., 2013) 圖 16.整合素內部化經由 BMP – Smad 路徑調節間葉幹細胞分化。
Figure 16. Integrin internalization regulates MSCs differentiation via the BMP − Smad pathway.
2.4.3.2 細胞形狀調節間葉幹細胞之分化方向 (Cell shape regulates commitment of MSCs)
根據先前研究證實,細胞形狀對於細胞的增殖,存活和分化的調節至關重要 (Chen et al., 1997; Roskelley et al., 1994)。為了探討細胞形狀的變化是否可以調節 hMSCs 細胞特化,McBeath 等人通過微縮成像技術 (micropatterning technique) 來 控制細胞形狀和細胞擴散程度:當允許 hMSCs 粘附、扁平和延展時,將進行成骨 分化;反之,未延展且呈現圓細胞型的 hMSCs 將分化為脂肪細胞 (McBeath et al., 2004)。另有研究表示細胞形狀會影響 Rho 家族中 GTPases 的活性 (Ren et al., 1999)。
為了證實 RhoA 是否涉及細胞特化,McBeath 等人研究發現:ECM 調控之細胞形
狀會通過 RhoA 以及 Rho 相關蛋白激酶 (Rock) 之分子路徑決定 MSCs 分化命運 (McBeath et al., 2004)。RhoA 的活化將誘導 MSCs 之成骨分化,反之則分化為脂肪 細胞。此等藉由 RhoA 調控之細胞特化訊息需要肌動蛋白—肌球蛋白產生的張力,
而不需要可溶性分化因子參與 (Arnsdorf et al., 2009)。此項研究顯示細胞形狀,細 胞骨架力學和 RhoA 訊息傳遞是決定幹細胞分化命運的組成要素。
2.4.3.3 構型變化影響間葉幹細胞之命運 (Topographic changes influence MSC fate)
ECM 是由許多孔隙、脊梁和不同大小的纖維所組成奈米級網絡結構,研究表 示 ECM 物理構型會影響細胞行為 (Curtis and Wilkinson, 2001)。隨著電子束光刻 (electron beam lithography, EBL) 技術之發展,已可用於製造極精密之奈米形貌,
大幅地促進細胞與奈米微環境相互作用之研究。通過 EBL 技術,Dalby 等人證實 MSCs 如何響應奈米級地形特徵 (Dalby et al., 2007)。然而 EBL 所創造之奈米構型 還不足以代表性的呈現生物體內之組織水平,因此 Dalby 研究團隊不僅利用 EBL 產生高度有序的對稱性,並產生具有不同水平的奈米無序和隨機表面,其中包括 (1) 正方形陣列 (square array, SQ):基底從原始圖案壓印 120 nm 直徑,100 nm 深 的奈米比特 (nanopits),平均間隔皆為 300 nm;(2) 無序正方形陣列 (disordered square array, DSQ):nanopits 從原本 SQ 位置在兩軸上隨機位移至多 50 nm (DSQ50) 或 20 nm (DSQ20);(3) RAND:nanopits 隨機在150 μm × 150 μm 的區域,並重複 以填充 1cm2面積。經培養 21 天後,DSQ50 上的 MSCs 表現骨骼專一性 ECM 蛋 白質例如骨橋蛋白 (osteopontin) 和骨鈣蛋白 (osteocalcin) 以及早期結節形成 (Dalby et al., 2007)。
因此 ECM 具有較複雜之幾何結構以及相對高剛度可促使 MSCs 產生 SFs 和 FAs 而往成骨細胞分化,若 SFs 和 FAs 形成受到抑制則往脂肪細胞分化 (圖 17)。
此結果可能因柔軟的 ECM 無法使 integrins 和 ECM 穩定結合造成整合素內化 (integrin internalization),進而使 FAs 無法形成,細胞無法感知並傳遞足夠的機械信 號 (Du et al., 2011)。ECM 物理性質可藉由改變細胞以及核骨架之張力影響基因的
表現,當 ECM 具有較高之剛度可增加細胞內的張力,導致細胞核變形並且增加 lamin A 和成骨細胞分化相關基因之表現;反之,低剛度 ECM 會降低細胞內張力,
而增加脂肪分化相關基因之表現 (Higuchi et al., 2013)。細胞黏著過程中藉由建立 細胞骨架的張力調節 MSCs 的分化方向,ECM 剛度可穩定核骨架蛋白–lamin A,
經由 serum response factor (SRF) 與 YAP1 轉而提高 SFs 相關基因之表現 (Swift et al., 2013)。lamin A 靜默 (silencing) 會促使 MSCs 在柔軟的 ECM 分化為脂肪細胞,
而 lamin A 增加則促使 MSCs 在堅硬的 ECM 分化為成骨細胞。ECM 的物理訊號 經 retinoic acid (RA) 路徑轉換至細胞核調節 lamin A 的轉錄 (Huang et al., 2015;
Swift et al., 2013) (圖 18)。
綜合上述,ECM 的物理性質決定 MSCs 的分化命運。當 MSCs 培養在具有高 剛度、尖銳邊緣或大尺寸固定區域的 ECM 基質上,會誘導成骨細胞分化;反之,
當 MSCs 培養在具有低剛度、平滑邊緣或小尺寸固定區域的 ECM 基質上,則會誘 導脂肪細胞分化 (Huang et al., 2015) (圖 19)。
(Huang et al., 2015) 圖 17. 胞外基質的物理性質調節間葉幹細胞分化。
Figure 17. Physical properties of ECM regulate MSCs fate and differentiation.
(Huang et al., 2015) 圖 18. 胞外基質的剛度經由改變細胞與核骨架的張力影響基因表現。
Figure 18. ECM stiffness affect gene expresion by altering cytoskeletal and nuclearskeletal tension.
圖 19. 胞外基質的物理因素調節間葉幹細胞之分化命運。 (Huang et al., 2015) Figure 19. Physical parameters of ECM regulate MSC fate and differentiation.
2.4.3.4 機械力調控間葉幹細胞的基因表現
MSCs 的分化除了受基質剛度 (stiffness) 和構型 (topography) 等內在機械信 號 (intrinsic mechanical cues) 所影響,外在的機械信號例如流體剪力、壓縮力、靜 水壓力和張力等,可經由細胞膜上的離子通道、細胞之間的 cadherins、細胞和胞外 基質的 focal adhesion 或細胞內的細胞骨架轉換為化學訊息,從而活化不同之分子 路徑進而調節 MSCs 的分化 (Steward and Kelly, 2015) (圖 20)。先前已有許多研究 將機械力刺激視為調節細胞結構和功能的關鍵因素。細胞將感測到的機械力傳遞 到本身或其它細胞內部,並將其轉換為化學訊號進而調節細胞之運動、分化和增殖 (Discher et al., 2009; Engler et al., 2004; Janmey and McCulloch, 2007; Yeung et al., 2005)。Kurpinski 等人使用微化條帶 (micropattened strip) 使 MSCs 的排列沿著單 軸應力的方向。結果顯示平滑肌細胞的標記 calponin 1 的表現增加,而軟骨基質標 記表現降低。然而,當細胞排列垂直於應力方向時,會減少基因表現的變化 (Kurpinski et al., 2006)。此結果表示機械力對 MSCs 的基因表現具有深遠的影響。
而另一項研究發現,若同一組中 TGF-β1 和循環之機械應力同時作用時,可協同 calponin1 基因表現增加,說明可溶性因子和機械應力可協同調控 MSCs 的基因表 現 (Kurpinski et al., 2009)。此外,流體剪力會經由活化 RhoA − ROCKII 路徑提高 actin filaments 的張力,誘導成骨細胞分化相關基因 Runx2, Wnt, Ror2 的表現。若 使用 LPA (lysophosphatidic acid) 激活 RhoA 則會抑制脂肪細胞分化相關基因 PPARγ 的表現;而使用 cytochalasin D 抑制機動蛋白聚合可增加 PPARγ 並抑制 Runx2 的表現 (Arnsdorf et al., 2009)。
(Steward and Kelly, 2015) 圖 20. 內在與外在機械信號調節間葉幹細胞之分化。
Figure 20. The intrinsic and extrinsic mechanical cues that regulate the differentiation of MSCs.
2.4.4 機械力與血管形成
新血管形成 (neovascularization) 根據血管生長的機制不同主要分為三種:(1) 血 管 新 生 (vasculogenesis) 由 中 胚 層 來 源之內 皮 前 驅細 胞 或稱 為血 管 母 細 胞 (angioblasts) 分泌內皮細胞遷移所需的生長因子和趨化因子 (chemokines) 重新形 成新的血管,負責胚胎時期原始血管之形成。(2) 血管生成 (angiogenesis) 為低氧 (hypoxia) 所誘導之內皮細胞 (endothelial cells, ECs) 的活化,沿著血管生成訊號的
方向形成小口徑的血管。(3) 動脈生成 (arteriogenesis) 為循環中的白血球被活化的 血管內皮所吸引,形成成熟具功能的側支動脈 (collateral arteries) 以繞開血管阻塞 的部位 (van Oostrom et al., 2008)。在 angiogenesis 過程當中,ECs 從組織中現有之 血管滲入到無血管的空間稱為血管萌芽 (vascular sprouting) (Song and Munn, 2011)。
血管內皮生成因子 (vascular endothelial growth factor, VEGF) 為血管生成主要的調 節因子,從無血管空間的細胞分泌出的 VEGF 形成 VEGF 濃度梯度,提供附近的 血管具有方向性和趨化性的訊號,進而刺激並引導血管萌芽 (Eilken and Adams, 2010)。有研究表示流動血液產生的流體剪力 (fluid shear stress) 會以一氧化氮依賴 的方式 (nitric oxide-dependent manner) 減弱 ECs 的萌芽;另外從血管滲出之血漿 產生的間隙滲流 (interstitial flow) 會引導 ECs 型態發生和萌芽。因此,流體剪力和 VEGF 濃度梯度使 ECs 整合達到血管擴張和萌芽之目的 (Song and Munn, 2011)。
流體剪力 (fluid shear stress, FSS) 是指血液或組織液流動時對組織或細胞所造 成的摩擦力,可促進骨髓來源之內皮前驅細胞 (endothelial progenitor cells, EPCs) 分化為成熟之內皮細胞。EPCs 於體外經 FSS 作用後其黏附、遷移、增殖和管狀形 成能力明顯增強,且內皮細胞特有標記之蛋白質如 Tie2, VEGF-R2, VE-cadherin 等 及其 mRNA 表現量皆明顯上升,而 PI3K/Akt/mTOR 訊息傳遞路徑涉及 EPCs 的增 殖和黏附並與內皮細胞特有標記之蛋白質表現有關 (Obi et al., 2012)。另外抑制分 析顯示 FSS 作用於integrin β1 後會經由 Ras/ERK 或 FAK/Paxillin 兩種訊息傳遞路 徑調節細胞骨架的重新組織 (cytoskeletal rearrangement) 進而導致 EPCs 的分化 (Cheng et al., 2013)。
機械性拉力 (mechanical tension force) 施於大鼠耳部可觀察到真皮層血管的 直徑以及內皮細胞的標記 PECAM-1 都有明顯增加,而且拉力作用後的血管口徑呈 現異質性 (heterogeneity) 和高度彎曲,這些皆為血管重塑 (vascular remodeling) 的 早期特徵 (Pietramaggiori et al., 2007)。此外拉力作用的過程中會造成組織短暫性缺 氧,RT-PCR 分析結果顯示週期性拉力可明顯提升 hypoxia-inducible factor 1-alpha (HIF-1α) 的基因表現,其為低氧環境所活化之血管生成轉錄因子 (Chin et al., 2010)。