國立交通大學
分子醫學與生物工程研究所
碩士論文
三 葉 型 扭 結 蛋 白 -YbeA 之摺 疊機 制 與功 能分 析
Folding and function analysis of trefoil knot
protein-YbeA
研 究 生 : 張 毓 娟
指 導 教 授 : 張 家 靖 教 授
三 葉 型 扭 結 蛋 白 -YbeA 之 摺 疊 機 制 與 功 能 分 析
Fol di ng and fun ct i on anal ys i s of t re foi l knot prot ei n -YbeA
研 究 生 : 張 毓 娟 Student:Yu-Chuan Chang
指 導 教 授 : 張 家 靖 Advisor: Chia-Ching Chang
國 立 交 通 大 學
分 子 醫 學 與 生 物 工 程 所
碩 士 論 文
A Thes i sS ubm i tt ed t o Ins t i t ut e of Mol ecul ar M edi ci ne and Bi oen gi ne e ri ng C ol l ege of Bi ol ogi ca l S ci ence and Te chnol og y
Nat i onal C hi ao Tun g Uni vers i t y
In pa rt i al Ful fi l l m ent of R equi rem ent for t he Degree o f M as t e r i n M ol ecul ar M edi ci ne and Bi oen gi nee ri ng
J ul y 2011
Hs i nchu,Tai wan,R ep ubli c of C hi na
誌 謝
碩 班 生 活 終 於 告 了 段 落 , 感 謝 一 路 上 許 多 人 的 支 持 , 首 先 最 先 感 謝 的 是 我 的 家 人 們 , 他 們 支 持 與 鼓 勵 , 是 我 努 力 前 進 的 動 力 , 此 外 十 分 感 謝 老 師 這 兩 年 來 的 悉 心 指 導 , 教 會 我 許 多 知 識 以 及 讓 我 學 習 到 追 求 知 識 的 該 有 的 態 度 , 另 外 , 感 謝 實 驗 室 同 伴 的 陪 伴 , 讓 我 的 研 究 生 活 更 加 有 趣 , 我 會 永 遠 記 得 大 伙 們 一 起 在 實 驗 室 裡 說 著 非 常 有 慧 根 的 奇 怪 笑 話 , 談 論 的 最 新 的 社 會 八 卦 新 聞 , 以 及 在 實 驗 室 裡 頭 做 的 所 有 蠢 事 。 在 這 裡 首 先 感 謝 喜 歡 請 我 喝 咖 啡 的 鄭 博 , 感 謝 您 非 常 耐 心 地 教 我 實 驗 的 技 術 , 不 過 不 要 再 給 你 兒 子 推 薦 媳 婦 了 , 這 樣 他 會 很 困 擾 的 。 再 來 是 長 得 很 像 壞 人 , 不 過 實 際 上 卻 是 個 大 好 人 的 學 亮 學 長 , 常 常 在 實 驗 上 麻 煩 到 你 , 還 讓 你 要 非 常 有 耐 心 的 跟 我 解 釋 實 驗 , 而 延 誤 你 回 家 的 時 間 , 真 是 歹 勢 , 不 過 念 在 我 常 常 跟 你 說 最 新 八 卦 消 息 的 份 上 , 學 長 你 以 後 成 功 應 該 還 是 會 好 好 提 拔 我 吧 。 還 有 總 是 忙 進 忙 出 、 跑 來 跑 去 的 憲 儀 姊 , 雖 然 很 忙 卻 還 是 常 常 幫 我 們 想 中 午 要 吃 什 麼,還 常 開 車 帶 我 們 出 門,真 是 太 感 謝 你 了, 還 有 我 不 得 不 對 你 的 開 車 技 術 加 以 讚 賞 , 真 是 好 到 沒 話 說 , 真 是 太 令 我 佩 服 了 。 感 謝 我 的 最 佳 飯 友 虹 瑋 , 跟 你 吃 飯 真 是 最 快 樂 的 時 候 了 , 害 我 不 知 不 覺 胖 了 好 幾 斤 , 我 一 定 要 好 好 瘦 下 來 。 感 謝 偉 賢 的 專 業 電 腦 教 學 , 讓 我 在 碩 班 生 涯 的 電 腦 功 力 成 為 人 生 中 最 巔 峰 的 時 刻 了 , 雖 然 常 常 你 講 一 講 我 就 忘 記 了 , 不 過 我 想 你 已 經 習 慣 了 。 還 有 昌 翰 , 雖 然 我 常 嗆 你 , 不 過 說 實 在 如 果 不 嗆 你 還 真 是 對 不 起 我 的 良 心 。 學 弟 維 邦 , 你 真 的 是 一 個 很 有 趣 的 人 , 我 覺 得 我 的 搞 笑 功 力 真 的 差 你 一 大 截 喔 , 希 望 你 可 以 繼 續 保 持 下 去 , 實 驗 室 有 你 應 該 會 越 來 越 熱 鬧 , 還 有 不 要 再 說 學 姊 很 想 跟 你 說 話 , 或 是 你 唱 歌 超 級 好 聽 。 最 後 希 望 在 未 來 的 日 子 裡 , 大 家 都 能 夠 朝 著 自 己 的 目 標 , 逐 漸 一 一 實 現 自 己 的 夢 想 。三葉型扭結蛋白-YbeA 之摺疊機制與功能分析
學生:張毓娟 指導教授:張家靖 教授 國立交通大學生分子醫學與生物工程研究所 碩士班摘要
近年來隨著蛋白結構隨著X光晶體繞射以及核磁共振技術的進步,有越來越多的蛋 白質結構逐漸被解析與發表,造成蛋白資料庫越來越完整。此外人們發現在自然環境底 下,某些蛋白胜肽鏈會自我交錯產生結。在這裡我們以YbeA為對象來研究扭結蛋白摺 疊機制,其中YbeA本身為大腸桿菌內的核醣體的核醣核酸甲基化酶,其本身蛋白胜肽 鏈會自我交錯產生三葉型扭結,其扭結型式較為簡單,且分子量較小。然而細胞內蛋白 質生成的過程本身是一個非常複雜的過程,因此我們希望透過體外重新摺疊的方式來探 討扭結的生成。在這裡我們透過圓二色光譜以及螢光光譜儀分析蛋白質的二級結構以及 疏水性核心在摺疊過程中的變化。此外我們透過胰蛋白酶分別對YbeA的摺疊態以及未 摺疊態反應,依據不同時間點收集產物,然後透過MALDI-TOF分析水解產物序列,結 果顯示,其結構中最晚被切下來的部份與pKNOT-環結蛋白質結構資料庫及分析工具所 模擬的扭結區域重疊,表示扭結部分在整個蛋白裡頭是最穩定,就算在高濃度尿素環境 底下,也無法使其裸露出來被酵素水解。此外我們透過螢光能量共振轉移來分析扭結區 位分子間的螢光能量轉移現象,結果顯示不論是在摺疊態或是未摺疊態其扭結區域的距 離都沒有改變,根據MALDI-TOF以及螢光能量共振轉移的結果,我們推測YbeA得扭結 部分為一個無法打開的結,此外我們透過動力學截流儀結合圓二色光譜來分析YbeA的 摺疊機制,結果顯示,不論是在摺疊或是變性的過程的速率都是相當緩慢的,且整個變 化都是相當緩慢的,我們推測可能扭結區域穩定蛋白構性變化所造成。Folding and function analysis of trefoil knot protein-YbeA
Advisee: Yu-Chuan Chang Adviser: Dr. Chia-Ching ChangInstitute of Molecular Medicine and Bioengineering National Chiao Tung University
Abstract
Knotted proteins are more commonly observed in recent years due to the growing number of structures dissolved by x-ray and NMR in the Protein Data Bank(PDB).A small number of natural proteins have knot configurations in its polypeptide backbone. We select YbeA as a model for its small size and simple knot configurations.YbeA is a RNA methyltransferase that contains a deep embedded trefoil knot in its backbone structure. It is a challenge to reveal the knot formation mechanism during the protein biosynthesis process. For exploring those questions,we use trypsin digested matrix-assisted laser desorption/ ionization time of flight (MALDI-TOF) mass spectroscopy technique to analyze the digested intermediates of YbeA indicated the knotted region is the most compact in overall structure, both in unfolded and folded state. That revealed the knotted structure can’t be opened even in high concentration denaturant environment. The similar result was observed by fluorescence resonance energy transfer (FRET) measurement. Meanwhile,folding kinetic of YbeA analyszed by stopped -flow assistted with CD demonstrate it gently changes both in the folding and unfolding process .It reveals that the knot structure may stabilize the protein comformation.
目錄
摘要 ... i
Abstract ... ii
表目錄 ... viii
圖目錄 ... ix
縮寫表 ... xii
第一章
緒論 ... 1
1-1 蛋白質結構與性質 ... 1 1-2 蛋白摺疊 ... 1 1-3 蛋白質摺疊的相關問題 ... 1 1-4 蛋白質錯誤摺疊相關疾病 ... 1 1-5 蛋白質摺疊相關研究 ... 1 1-6 蛋白質的變性方法 ... 2 1-7 蛋白質的摺疊方法 ... 2 1-7-1 直接稀釋法(direct dilution) ... 21-7-2 階段性熱平衡透析法(stepwisethermal equilibrium dialysis) ... 2
1-8 蛋白質的摺疊速率 ... 2
1-9 蛋白質摺疊理論模型 ... 3
1-9-1 Framework model ... 3
1-9-2 The nucleation and growth mechanism ... 3
1-9-3 The diffution-collision model ... 3
1-9-4 Hydrophobic collapse model ... 3
1-9-6 First-order-like phase transition ... 4 1-10 研究動機 ... 4 1-11 YbeA 蛋白的簡介 ... 5 1-11-1 YbeA 的結構 ... 5 1-11-2 YbeA 的功能 ... 5 1-12 23S rRNA 的功能 ... 6 1-13 扭結蛋白(knotted protein)簡介 ... 7 1-13-1 扭結蛋白的分類 ... 7 1-13-2 扭結蛋白生物物理性質研究 ... 8 1-13-3 扭結蛋白質結構對蛋白穩定性的影響 ... 8 1-13-4 扭結蛋白之摺疊研究 ... 9 1-14 實驗策略 ... 10
第二章
實驗原理與方法 ... 11
2-1 基因構築 ... 112-1-1 ybeA wild type 基因構築 ... 12
2-1-2 ybeA△73-75 基因構築 ... 13 2-1-3 ybeA△1-8 基因構築 ... 15 2-1-4 ybeA△1-70 基因構築 ... 15 2-1-5 ybeA△118-155 基因構築 ... 16 2-2 蛋白的製備 ... 17 2-2-1 蛋白質小量表現 ... 17 2-2-2 蛋白質大量表現 ... 17 2-2-3 蛋白摺疊 ... 17
2-3 圓二色光譜(Circular Dichroism Spectroscopy) ... 18
2-4-1 螢光原理 ... 18
2-4-2 蛋白質螢光原理 ... 19
2-5 恆溫滴定微卡計(Isothermal Titration Calorimetry) ... 19
2-6 介質輔助雷射脫附游離飛行式質譜儀 (MALDI-TOF-MS) ... 20
2-7 螢光共振能量轉移(Förster resonance energy transfer) ... 20
2-8 力學截流儀(stopped -flow)結合圓二色光譜(CD Spectroscopy) ... 21
第三章
實驗結果 ... 22
3-1 基因構築結果 ... 22 3-1-1 pET30a-ybeA 質體構築 ... 22 3-1-2 pET30a-ybeA△73-75 質體構築 ... 24 3-1-3 pET30a -ybeA△1-8 質體構築 ... 26 3-1-4 pET30a-ybeA△1-70 質體構築 ... 28 3-1-5 pET30a-ybeA△118-155 質體構築 ... 30 3-2 蛋白質表現結果 ... 32 3-3 螢光分析摺疊中間產物的疏水性核心變化 ... 33 3-3-1 YbeA 摺疊中間產物疏水性核心分析 ... 33 3-3-2 YbeA△73-75 摺疊中間產物疏水性核心分析 ... 33 3-3-3 YbeA△1-8 摺疊中間產物疏水性核心分析 ... 33 3-3-4 YbeA△1-70 摺疊中間產物疏水性核心分析 ... 33 3-3-5 YbeA△118-155 摺疊中間產物疏水性核心分析 ... 33 3-4 圓二色光譜分析 YbeA 與其他突變株蛋白摺疊中間產物的二級結構變化 ... 38 3-4-1 YbeA 摺疊中間產物二級結構分析 ... 38 3-4-2 YbeA△73-75 摺疊中間產物二級結構分析 ... 38 3-4-3 YbeA△1-8 摺疊中間產物二級結構分析 ... 38 3-4-4 YbeA△1-70 摺疊中間產物二級結構分析 ... 383-4-5 YbeA△118-155 摺疊中間產物二級結構分析 ... 39 3-4-6 YbeA 與其他突變株蛋白摺疊態之二級結構比較 ... 39 3-5 YbeA 摺疊產物功能分析 ... 48 3-6 YbeA 酵素水解穩定性分析 ... 49 3-6-1 YbeA 摺疊態酵素水解穩定性分析 ... 49 3-6-2 YbeA 非摺疊態酵素水解穩定性分析 ... 51 3-7 YbeA 扭結區域分子間距離分析 ... 53 3-8 蛋白熱穩定性分析 ... 54 3-8-1 螢光光譜儀分析熱穩定性 ... 54 3-8-1-1 YbeA 蛋白螢光變溫 ... 55 3-8-1-2 YbeAΔ73-75 蛋白螢光變溫 ... 56 3-8-1-3 YbeAΔ1-8 蛋白螢光變溫 ... 57 3-8-1-4 YbeAΔ1-70 蛋白螢光變溫 ... 58 3-8-1-5 YbeAΔ118-155 蛋白螢光變溫 ... 59 3-8-2 圓二色光譜儀分析熱穩定性 ... 60 3-8-2-1 YbeA 二級結構熱穩定性分析 ... 61 3-8-2-2 YbeA△73-75 二級結構熱穩定性分析 ... 62 3-8-2-3 YbeA△1-8 二級結構熱穩定性分析 ... 63 3-8-2-4 YbeAΔ1-70 二級結構熱穩定性分析 ... 64 3-8-2-5 YbeAΔ118-155 二級結構熱穩定性分析 ... 65 3-9 摺疊速率分析 ... 66 3-9-1 靜態平衡摺疊及變性分析 ... 66 3-9-2 動態摺疊速率分析 ... 68
第四章
討論 ... 70
4-1 基因構築 ... 704-2 蛋白質定性分析 ... 70 4-2-2 摺疊產物結構及功能分析 ... 71 4-2-3 熱穩定性分析 ... 73 4-2-4 摺疊速率分析 ... 74 4-2-5 酵素結構穩定性分析 ... 75 4-2-6 扭結的距離分析 ... 75
第五章
結論 ... 76
第六章
參考文獻 ... 77
表目錄
表 1 E. coli strain. K-12 substn. DH10B 的 ybeA 基因序列 ... 12
表 2 重組 ybeA 基因序列 ... 14 表 3 蛋白質二級結構特徵鋒 ... 18 表 4 YbeA 及其突變株之螢光圖譜數據 ... 36 表 5 SELCON3 分析 YbeA 摺疊中間產物二級結構數據 ... 46 表 6 SELCON3 分析 YbeA△73-75 摺疊中間產物二級結構數據 ... 46 表 7 SELCON3 分析 YbeA△1-8 摺疊中間產物二級結構數據 ... 46 表 8 SELCON3 分析 YbeA△1-70 摺疊中間產物二級結構數據 ... 46 表 9 SELCON3 分析 YbeA△118-155 摺疊中間產物二級結構數據 ... 47 表 10 SELCON3 分析 YbeA 及其突變株摺疊態之二級結構數據 ... 47 表 11YbeaA 蛋白分子內部螢光能量轉移 ... 53 表 12 螢光光譜儀分析熱相變數據 ... 59 表 13 YbeA 變性摺疊平衡熱力學參數 ... 66 表 14 YbeA 及 Δ1-8 變性摺疊動力學參數 ... 69 表 15 ITC 實驗數據表 ... 73 表 16 靜態平衡摺疊及變性比較表 ... 74 表 17 動態速率分析比較 ... 75
圖目錄
圖 1 蛋白質摺疊自由能漏斗型模型 ... 3
圖 2 蛋白質摺疊似一階轉變示意圖 ... 4
圖 3 YbeA 結構模型 ... 5
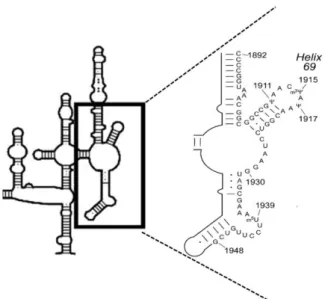
圖 4 23S rRNA 二級結構上 helix loop 69(H69)的 m3Ψ1915 示意圖 ... 6
圖 5 m3Ψ1915 的甲基化示意圖 ... 6 圖 6 23S rRNA 甲基化示意圖 ... 6 圖 7 扭結蛋白種類 ... 7 圖 8 基因構築實驗流程圖 ... 11 圖 9 ybeA△73-75 基因 megaprimer PCR 流程圖 ... 13 圖 10 螢光原理 ... 19 圖 11 動力學截流儀結合圓二色光譜儀裝置 ... 21 圖 12 質體 pET30a-ybeA 的建構圖... 22 圖 13 pET30a-ybeA 質體構築結果 ... 23
圖 14 重組 ybeA 基因與 NCBI 公布之 ybeA 基因的比較結果。 ... 23
圖 15 質體 pET30a-ybeA△(73-75)的建構圖... 24 圖 16 pET30a-ybeA△(73-75)基因建構結果 ... 25 圖 17 重組 ybeA 與重組 ybeA△(73-75)基因序列比較結果。 ... 25 圖 18 質體 pET30-ybeA△1-8 的建構 ... 26 圖 19 ybeA△1-8 基因建構結果。 ... 27 圖 20 重組 ybeA 與重組 ybeA△(1-8)基因序列比較結果。 ... 27 圖 21 質體 pET30-ybeA△1-70 的建構圖 ... 28 圖 22 ybeA△1-70 基因建構結果。 ... 29 圖 23 重組 ybeA△1-70 與重組 ybeA 基因序列比較結果 ... 29 圖 24 質體 pET30-ybeA△118-155 的建構圖 ... 30 圖 25 ybeA△118-155 基因建構結果。 ... 31 圖 26 重組 ybeA 與重組 ybeA△118-155 基因序列比較結果。 ... 31
圖 27 YbeA 以及其突變株 IPTG 誘導後 SDS PAGE 分析結果 ... 32
圖 28 YbeA 摺疊中間物螢光圖譜 ... 34
圖 29 YbeA△73-75 摺疊中間物螢光圖譜 ... 34
圖 30 YbeA△1-8 摺疊中間物螢光圖譜 ... 35
圖 31 YbeA△1-70 摺疊中間物螢光圖譜 ... 35
圖 33 YbeA 摺疊中間產物圓二色圖譜 ... 40 圖 34 YbeA 摺疊中間產物圓二色圖譜 SELCON3 分析結果 ... 40 圖 35 YbeAΔ73-75 摺疊中間產物圓二色圖譜 ... 41 圖 36 YbeAΔ73-75 摺疊中間產物圓二色圖譜 SELCON3 分析結果 ... 41 圖 37 YbeAΔ1-8 摺疊中間產物圓二色圖譜 ... 42 圖 38 YbeAΔ1-8 摺疊中間產物圓二色圖譜 SELCON3 分析結果 ... 42 圖 39 YbeAΔ1-70 摺疊中間產物圓二色圖譜 ... 43 圖 40 YbeAΔ1-70 摺疊中間產物圓二色圖譜 SELCON3 分析結果 ... 43 圖 41 YbeAΔ118-155 摺疊中間產物圓二色圖譜 ... 44 圖 42 YbeAΔ118-155 摺疊中間產物圓二色圖譜 SELCON3 分析結果 ... 44 圖 43 YbeA 與其他突變株蛋白的 CD 圖譜 ... 45 圖 44 SELCON3 軟體分析 YbeA 及其突變株之二級結構比例 ... 45
圖 45 ITC 之 AdoMet 滴定 YbeA 的原始圖 ... 48
圖 46 AdoMet 滴定 YbeA 之作用力圖... 48 圖 47 YbeA 摺疊態胰蛋白酶水解反應圖 ... 50 圖 48 YbeA 非摺疊態胰蛋白酶水解反應圖 ... 52 圖 49 YbeA 摺疊態以及非摺疊態和 AMCA 的螢光能量轉移圖 ... 53 圖 50 YbeA 蛋白變溫螢光光譜 ... 55 圖 51 YbeA 蛋白螢光變溫波峰變化圖 ... 55 圖 52 YbeA△73-75 蛋白變溫螢光光譜 ... 56 圖 53 YbeA△73-75 蛋白螢光變溫波峰變化圖 ... 56 圖 54 YbeA△1-8 蛋白變溫螢光光譜 ... 57 圖 55 YbeA△1-8 蛋白螢光變溫波峰變化圖 ... 57 圖 56 YbeA△1-70 蛋白變溫螢光光譜 ... 58 圖 57 YbeA△1-70 蛋白螢光變溫波峰變化圖 ... 58 圖 58 YbeA△118-155 蛋白變溫螢光光譜 ... 59 圖 59 YbeA△118-155 蛋白螢光變溫波峰變化圖 ... 59 圖 60 YbeA 蛋白 α 螺旋二級結構熱變化 ... 61 圖 61 YbeA 蛋白 β 平板二級結構熱變化 ... 61 圖 62 YbeAΔ73-75 蛋白 α 螺旋二級結構熱變化 ... 62 圖 63 YbeAΔ73-75 蛋白 β 平板二級結構熱變化 ... 62 圖 64 YbeAΔ1-8 蛋白 α 螺旋二級結構熱變化 ... 63 圖 65 YbeAΔ1-8 蛋白 β 平板二級結構熱變化 ... 63
圖 66 YbeAΔ1-70 蛋白 α 螺旋二級結構熱變化 ... 64 圖 67 YbeAΔ1-70 蛋白 β 平板二級結構熱變化 ... 64 圖 68 YbeAΔ118-155 蛋白 α 螺旋二級結構熱變化 ... 65 圖 69 YbeAΔ118-155 蛋白 β 平板二級結構熱變化 ... 65 圖 70 螢光光譜儀分析 YbeA 靜態平衡之摺疊及變性 ... 67 圖 71 圓二色光譜儀分析 YbeA 靜態平衡之摺疊及變性 ... 67
圖 72 Chevelon plot of YbeA ... 69
圖 73 Chevelon plot of YbeAΔ1-8 ... 69
圖 74 YbeA 摺疊螢光圖譜 ... 71
圖 75 YbeA 摺疊圓二色圖譜 ... 72
縮寫表
縮寫 英文全寫 中文
ATP Adenosine Triphosphate 三磷酸腺苷
CD Circular Dichroism 圓二色光譜
DTT Dithiothreitol 2-4 二硫代蘇糖醇
ETTA Ethylenediaminetetraacetic Acid 乙烯二胺四乙酸
FRET Förster Resonance Energy Transfer 螢光共振能量轉移
ITC Isothermal Titration Calorimetry 恆溫滴定熱卡計
MALDI Matrix-Assisted Laser Desorption Inoization 基質輔助雷射去吸
附離子化
MTase Methylase 甲基化酶
PCR Polymerase Chain Reaction 聚合酶連鎖反應
SAM S-Adenosyl Methionine S-腺苷甲硫氨酸
SAM S-Adenosyl-L-Homocysteine S -腺苷- L -半胱氨
酸
SDS Sodium Dodecyl Sulfate 十二烷基硫酸鈉
第一章 緒論 1-1 蛋白質結構與性質 蛋白質主要成分為碳、氫、氧、氮這四種元素,而胺基酸為蛋白質的組成的基本單 位,其中每一個胺基酸分子都有一個羧基(-COOH)與胺基(-NH2),蛋白質可由一條或多 條多肽鏈組成,因此其結構可依次分為四級,蛋白質一級結構指的是組成蛋白質胜肽鏈 的線性胺基酸序列。而二級結構依據不同胺基酸之間的 C=O 和 N-H 基團間的胜肽鍵形 成的穩定結構,主要為α 螺旋和 β 平板。三級結構指的是多個二級結構在三維空間的排 列所形成的一個蛋白質分子的三維結構。四級結構指的是由不同多肽鏈間相互作用形成 具有功能的蛋白質複合物分子。 1-2 蛋白摺疊 從多肽鏈的線性胺基酸序列到具有三維空間構型的過程就被稱為蛋白質摺疊。特定 序列在蛋白摺疊時,有時可以摺疊為一種以上的構型。在真核細胞內,許多蛋白質的正 確摺疊往往需要分子輔佐子的幫助摺疊或是將錯誤摺疊的蛋白重新變性後再摺疊以防 止錯誤發生。細胞處在逆境環境下時,一種叫做熱休克因子(heat shock factor)的蛋白 質會察覺到環境的改變,然後啟動分子輔佐蛋白(molecular chaperone)的製造。分子 輔佐蛋白在身體內的工作是負責監督蛋白質的摺疊,防止摺疊失敗的蛋白質在體內堆積 (1)。 1-3 蛋白質摺疊的相關問題 蛋白質摺疊問題在於蛋白質如何從一條胜肽鏈,摺疊回具有三維結構的蛋白構型﹖ 以及蛋白質啟動摺疊的關鍵在於什麼?另外蛋白質真的在摺疊過程不會出錯嗎?這些問 題成為科學界欲探討的重要課題,為了解釋這些問題,陸陸續續有許多理論先後被提 出,這些理論中包含了蛋白質透過分子間交互作用來進行摺疊的理論。 1-4 蛋白質錯誤摺疊相關疾病 蛋白質在摺疊過程之中可能會產生錯誤摺疊或是沉澱聚集的現象,如狂牛症以及阿 茲海默症。Prion為1997年諾貝爾醫學獎的得主Stanley Prusiner所致力研究的主體,他在 人類以及動物身上發現傳染性海綿狀腦病變(transmissible spongiform encephalopathy), 例如從牛身上的狂牛症(BSE)(2, 3)、羊的搔癢病(scrapie)(4, 5)、人類的庫賈氏症(CJD) (6)都是由於體內Prion錯誤摺疊所導致,腦組織從電腦斷層上看起來就會有一個一個空 洞,所以稱之為海綿腦。PrP有兩種isoform,一種是正常的PrPc(cellular PrP),一種為 不正常PrPsc ( PrP scrapie ),PrPc與PrPsc兩者最大的差異就是在結構上,而不是在共價鍵 的修飾上。PrPc主要以α-螺璇結構為主,幾乎沒有β-板狀結構,而PrPsc則是以β-板狀結構 為主,此形式蛋白會有自動聚集的傾向。 阿茲海默症(Alzheimer's disease)俗稱為老年癡呆症(7, 8),是一個神經退化性的疾 病,由阿滋海默醫生在1907年發表的。這種病會造成記憶的失去,認知的失調,還有不 正常的行為,他在阿茲海默症病患腦內觀察到斑塊(plaques)現象,斑塊主要由名為乙 型-澱粉樣蛋白(beta amyloid)的蛋白質所組成,此種蛋白質很容易聚集形成沈澱物, 阻斷神經彼此溝通和傳遞訊息的功能。 1-5 蛋白質摺疊相關研究
1972年諾貝爾化學獎得主Christian Boehmer Anfinsen等人(9)以核糖核酸水解脢 (RNase)為研究對象,利用還原劑破壞雙硫鍵以及尿素破壞分子間共價作用力讓RNase
變性後失去水解RNA的能力,然而當環境中的還原劑以及變性劑移除時,RNase即可恢 復其活性,且物理化學性質均與未變性前相同,不論試驗幾次,蛋白質都僅此一種立體 結構。由於實驗中並無破壞胺基酸間的共價鍵結,因此可說蛋白質胺基酸序列決定其特 定結構。 1-6 蛋白質的變性方法 蛋白質的變性方法可以分成物理:如加熱(10),加壓(11),冷凍(12)或化學:極端 pH 變化(13),有機溶劑(11)方法,以及生物方法:如酵素水解(14)……等。 一般常用的化學變性劑有尿素(urea)(15)、鹽酸胍(Guanidine HCl)(15, 16)、硫氫乙 醇、(Mercaptoethanol)(17)、十二烷基硫酸鈉(SDS)(18) 二硫蘇糖醇(DTT)……等。其中 尿素以及鹽酸胍的作用在於破壞蛋白內氫鍵,以及疏水性作用力等一些非共價還原力。 而硫氫乙醇以及二硫蘇糖醇為有效的還原劑可以將蛋白質內雙硫鍵打斷。而十二烷基硫 酸鈉能在蛋白表面包覆上一層負電荷。 1-7 蛋白質的摺疊方法 1-7-1 直接稀釋法(direct dilution) 就是單一步驟溶液置換或所謂之直接反應模式(off-path) (19),將含有高濃度變性劑 的蛋白質直接置換至去除變性劑的環境底下,使蛋白質進行摺疊。此外根據 1996 年 Rudolph 等人的實驗結果,蛋白質濃度須維持在 0.01-0.05 mg/ml 才有較佳的摺疊效果 (20),蛋白質的摺疊效率會隨著蛋白質的濃度增加而大幅降低。
1-7-2 階段性熱平衡透析法(stepwisethermal equilibrium dialysis)
透過逐步透析去除變性劑,整體溶液交換擴散反應是緩慢的,蛋白質摺疊可以視為 近乎靜止的狀態,在這種透析方法中,蛋白每一階段均遵守著準靜熱力學平衡原理,蛋 白質的構型能量都會達到最低態,最後達到自然構型的能量,階段性溶液置換準靜過 程,蛋白質摺疊反應中並未碰觸到態轉變區間,讓蛋白可以順利摺疊回摺疊態(21, 22)。 1-8 蛋白質的摺疊速率 根據實驗技術的進步,可以測到不同範圍的摺疊速率,目前針對蛋白質摺疊速率大 多利用 stopped-flow 或 continuous-flow 技術,透過改變蛋白質外在環境溶液,測量其光 學訊號如螢光光譜儀或圓二色光譜儀變化來決定蛋白質的摺疊速率。其中 stopped-flow 偵測極限在 10-3秒左右(23),contiuous-flow(24)偵測速率提升至 10-5秒左右。此外 laser flash photolysis(25)新穎技術可測到 ns 等級的解析度。根據文獻指出二級結構 α 螺旋的 形成時間約 100ns(26),β 平板的形成時間約 6 秒(27),而三級結構約需要毫秒的時間(23)。 蛋白質摺疊從變性狀態到自然狀態,所需的時間依照序列長短而有所不同,其中越 長的序列需要越長的時間,根據前人的研究指出蛋白質的摺疊時間約需 10-9 ~10-3秒(26, 28)。蛋白質在未摺疊態有多種構型,如果蛋白質在摺疊的過程為系統性地搜尋所有可 能構形,這樣一來需要花費的時間是相當驚人的。1995 年 Peter Wolynes(29)所提出的蛋 白摺疊漏斗模型(funnel model)解釋了這個現象,這為蛋白質摺疊提出新的見解,利用蛋 白質的自由能與其構型之間的關係,透過 energy landscape 的方式解釋蛋白質如何從未 摺疊態到自然態的過程,構型能量變化往能量譜中能量最低處進行尋找其最穩定的狀 態,如圖 1 所示(30) 。
圖 1 蛋白質摺疊自由能漏斗型模型 1-9 蛋白質摺疊理論模型 1-9-1 Framework model Framework model 指出蛋白質在摺疊初期會因為氫鍵的交互作用力先形成二級結 構,而三級結構再由這些二級結構組合而成(31),並指出若是在二級結構形成前即產生 三級結構,如蛋白質內雙硫鍵錯誤鍵結,便會造成蛋白質無法摺疊回自然態。
1-9-2 The nucleation and growth mechanism
在蛋白質摺疊的過程之中,某些特定胺基酸之間的殘基會產生交互作用,形成類似 核(nucleus)的結構,然後再逐漸形成穩定的結構,蛋白質其他部分再將之前的核結構包 覆起來,最後完成摺疊形成三級結構。這些核形成時常常伴隨著二級結構的產生(32, 33),因此若是在摺疊過程中核結構不穩定,或是缺乏長距離交互作用力時,其摺疊反 應會較緩慢(33)。
1-9-3 The diffution-collision model
1976 年由 Karplus 以及 Weaver 提出(34, 35),他們將蛋白質視為許多微小區間 (micro-domain)的組合。在摺疊初期為混亂的結構,許多微小區間快速擴散碰撞尋找最 合適的結構,最後在慢慢調整至自然態的構型。
1-9-4 Hydrophobic collapse model
1954 年由 Kauzmann 提出(36),認為在蛋白質摺疊過程中,非極性胺基酸受到疏水 性效應(hydrophobic effect)後,造成彼此之間互相吸引聚集。疏水作用力是蛋白質摺疊過 程中的強作用力,蛋白質受到如此強的作用力後會快速崩縮至近似自然態大小,才有其 他分子內交互作用力而形成二級結構(37, 38)。
1-9-5 The hierarchical model
1981 年 G.D. Rose 提出在蛋白質摺疊過程中,蛋白質由區塊間的交互作用力形成一 些三級結構的區塊,然後在這些區塊包含了疏水效應崩縮形成部分的結構,這些結構在 彼此間產生交互作用(如靜電作用、凡得瓦力……)組成蛋白質的自然結構。其中區塊分 子之間的交互作用力包含了疏水性作用力、靜電作用力以及凡得瓦力等等(39)。
1-9-6 First-order-like phase transition
由 Chang,et al 於 2002,2004 年提出的蛋白摺疊過程之模型(40, 41),主要研究蛋白 質在準靜態熱力學平衡過程中之摺疊模型,此模型與 Landau 的一階相變(firt-order phase transition)相似,一階相變指出系統相圖(phase diagram )中的相變線(transition line)為有限 長的線段,如果此一線段出現一次微分不連續,此相變我們稱之為一階相變,通常一次 微分不連續出現在其端點,此點稱為特異點(critical point)。 一階相變將線兩旁的相區分開來,在此線上處於兩相共存的狀態,如果繞過此特異 點(over-critical point),儘管相已經改變,仍然不會觀察到相轉變現象。蛋白質摺疊視為 一階相變轉變,未摺疊態與摺疊態中間存在著相轉變線或區間,此區間為一相轉變區 間,在此轉變區間可以看到摺疊態以及未摺疊態共存的現象,透過 stepwise thermal equilibrium dialysis (TED)的摺疊方法,蛋白質摺疊時可以繞過此有限區間到達自然態。 圖 2 為 stepwise thermal equilibrium dialysis (TED)摺疊方法,透過此一方法可以繞過轉
變有限區得到不同中間體 M1、M2、M3、M4、M5,若是直接穿過相轉變區,蛋白質會 發生沉澱或是產生玻璃態的未摺疊態。 圖 2 蛋白質摺疊似一階轉變示意圖 1-10 研究動機 蛋白質的結構與其功能息息相關,然而蛋白質如何從一條胜肽鏈摺疊成具有立體結 構的蛋白質,一直是科學家們想要想要了解的課題。然而近年來隨著許多蛋白質分子結 構逐漸解析與發表,其中某些蛋白質的胜肽骨架結構會自我交錯產生扭結。蛋白質摺疊 本來就是一個複雜的過程,然而扭結又是如何在摺疊過程中形成呢?成為了一個相當有 趣的課題。因此我們透過 pKNOT 扭結蛋白資料庫(42)找到一個大腸桿菌所擁有的扭結 蛋白 YbeA(PDB:1NS5),其本身結的形式較簡單且分子較小,基因來源取得方便且容易 表現取得,為研究扭結蛋白摺疊機制極佳的研究對象。本研究希望透過基因重組蛋白重 新摺疊的方式,了解此一扭結蛋白結構的變化並嘗試解析其摺疊機制,並嘗試建立數個 扭結結構相關突變株以瞭解扭結結構對蛋白摺疊的影響,並和已知之蛋白質摺疊機制比 對,以求建立普適性蛋白摺疊機制模型。
1-11 YbeA 蛋白的簡介 1-11-1 YbeA 的結構 YbeA 本身大腸桿菌裡頭的蛋白質,為雙聚體的結構,根據 YbeA(PDB 1NS5)的電 腦模擬圖(圖 3),每個單體由內部五個 β 平板,以及外部的五個 α 螺旋所組成,兩個單 體間的α1 螺旋以及 α5 螺旋以反相平行方式緊密靠在一起(43, 44)。其中每個單體由 155 個胺基酸所組成,其 C 端的 35 個胺基酸(121-155)肽鏈穿過中間 51 個胺基酸構成的肽鏈 環(70-120)形成了一個三葉型扭結(trefoil knot) (43),為一個 α 螺旋、β 平板交錯排列的 蛋白,因此又稱為α/β knot。 圖 3 YbeA 結構模型
圖(a)為 YbeA 單體 圖(b)、(c)為 YbeA 雙體,兩個單體互相反平行(43),其中 α 代表 α 螺旋,β 代表 β 平 板,C 代表蛋白胜肽鏈之羧酸端,而 C’為另一個單體的羧酸端。
1-11-2 YbeA 的功能
YbeA 屬於大腸桿菌內核醣體的核醣核酸甲基化轉移酶,當缺乏 ybeA 基因時會影 響大腸桿菌生長,其作用主要在對 70S 核醣體複合體內的 50S 大次單元體的 23S rRNA
螺旋環 69(helix loop 69)的 Ψ1915 位置甲基化(圖 4 圖 5)當 70S 核醣體複合體解離時,
YbeA 即 失 去 甲 基 化 的 功 能 。 於 是 YbeA 蛋 白 又 叫 做 RlmH(rRNA large subunit methyltransferase H)(45, 46),是目前為止發現唯一能針對 pseudouridine 甲基化(45-47)的 甲 基化 酶 ,若是 1915 位置的 U 沒有被 RluD pseudouridine sythase(48) 作用生成 pseudouridine,YbeA 即無法對 1915 這個位置甲基化(45, 46)。
圖 4 23S rRNA 二級結構上 helix loop 69(H69)的 m3Ψ1915 示意圖
其中插圖為 H69 的放大圖
圖 5 m3Ψ1915 的甲基化示意圖
圖中 RluD 為 pseudouridine sythase,負責催化 uridine 的 pseudo 化,而 RlmH 即為 YbeA 負責 pseudouridine 的甲基化
1-12 23S rRNA 的功能
原核生物的 70S 核醣體由 30S 以及 50S 次單位所組成,其成分主要為蛋白質以及核 醣體 rRNA。30S 次單元體由 16S rRNA 以及 21 種蛋白質所組成,50S 次單元體由 23S 核 醣體 rRNA 以及 34 種蛋白質所組成。其中 23S rRNA 本身由 2904 個核甘酸所組成,23S rRNA 具有核糖酶(ribozyme) 的能力,能催化胺基酸之間肽鍵(peptide bond)生成,因此 稱之為胜肽轉移酶(peptidyl transferase),(49)其作用機制如圖 6 所示。
圖 6 23S rRNA 甲基化示意圖 圖中 A site 代表核醣體上胺基酸進入位置,P site 代表核醣體上肽鏈轉移位
1-13 扭結蛋白(knotted protein)簡介 扭結在日常生活中雖然很常見(50, 51),甚至在 1976 年,人們在 DNA (52)、RNA(53) 上都發現了打結的構型,近年來隨著許多蛋白質分子結構逐漸解析與發表,人們發現某 些蛋白質的胜肽骨架結構會自我交錯產生扭結,於是確認確實有扭結蛋白的存在。從第 一個扭結蛋白被發現到現在,結構資料庫中約有 273 個扭結蛋白,佔已知結構蛋白中的 百分之一(54),此外,扭結蛋白廣泛分布於各物種,從原核生物、古菌一直到真核生物 以至於到人類(55, 56)身上也都有發現,然而蛋白質為何發展出這樣的構型,是否具有演 化上的意義,目前仍然不是很清楚,有說法指出扭結可能對結構的穩定性以及功能上有 助益(57)。 扭結蛋白的深度隨著蛋白的種類而有所不同,有些扭結非常淺就像是蛋白質肽鏈末 端晃動到附近的肽鏈,只要移除兩三個胺基酸即失去扭結的構型,或是必須在特定的角 度才看得到(58)。隨著電腦科技的進步,讓許多一剛開始被歸類為非扭結蛋白可以正名 為扭結蛋白,讓扭結蛋白資料庫越來越完整。 1-13-1 扭結蛋白的分類
根據相同平面上多肽鏈的接觸點數目分為 31 knot 、41 knot、52 knot,以及 61 knot。
圖 7 蛋白依序分別為 31 knot 、41 knot、52 knot,以及 61 knot。以下將分別對其做介紹 (59)。
圖 7 扭結蛋白種類
§ 31 knot
31 knot 又稱為三葉型扭結(trefoil knot),其同一平面上的接觸點數目為三,佔扭結蛋
白裡頭數目的 最大宗 ,其 例子有 甲 基化酶 (Methyltrasferase)(44, 60)、( 甲醯基轉移 酶)Transcarbamylase(42, 60)、碳酸酐酶(Carbonic anhydrase)(61, 62)……等等。
S-腺苷甲硫氨酸合成酶(S-adenosylmenthione synthase)為第一個被發現的三葉型扭 結 蛋 白 , 其 作 用 在 於 負 責 催 化 蛋 胺 酸 (methione) 以 及 三 磷 酸 腺 苷 (ATP) 合 成 S-adenosylmenthione(63, 64)。而碳酸酐酶(Carbonic anhydrase)(64)是最早被證實的三葉型
扭結蛋白,作用在於催化 CO2+H2O→HCO3-+H+ 。而 α/β 扭結摺疊(α/βknot fold)為最常
被研究的三葉型扭結蛋白,是由一群核醣核酸甲基化轉移酶(RNA methyltrasferase )所組 成,如大腸桿菌(Escherichia coli)的 YbeA(43)以及流感嗜血桿菌(Haemophilus influenza) 的 Yibk 都屬於這一類(43, 44, 65)。此外早年由於儀器以及技術上的限制,讓許多扭結蛋 白 被 誤 判 為 非 扭 結 蛋 白 , 像 是 在 2000 年 發 現 的 N- 琥 珀 鳥 氨 酸 轉 胺 甲 酰 酶 (N-succinylornithine transcarbamylase)(66)在剛開始並不被認為是扭結蛋白,一直到後來
分析程式發展進步,才確定其三葉型扭結的存在。
§ 41 knot
41 knot 又稱為八字結(figure eight knot),其同一平面上的接觸點數目為四(67),如植
物的乙酰羥酸還原異構酶(ace-tohydroxy acid isomeroreductas)就屬於這一類,其功能為合 成胺基側鏈,為目前為止發現扭結深度最深的蛋白,當同時從其 N 端移除 300 個胺基酸 以及從 C 端移除 60 個胺基酸後並不會破壞扭結(68)。而另外一個例子是耐輻射奇異球 菌 野 生 型 菌 株 (Deinococcus radioduran) 內 光 敏 色 素 (phytochrome) 的 發 光 團 接 合 區 (chromophore binding region)(67),phytochrome 扮演了感應紅光與紅外線的雙重角色。
§ 52 knot
52 knot 其同一平面上的接觸點數目為五。其例子有泛素羧基末端水解酶(ubiquitin
carboxyl-terminal hydroxylase,UCH-L)中的 UCH-L1(56, 69)以及 UCH-L3(66, 70, 71),其
中泛素羧基末端水解酶在泛素-降解體路徑(ubiquitin proteasome)反應中負責水解泛素 (ubiquitin)與被標記蛋白之間的鍵結,協助泛素的回收利用,維持生理系統的平衡。UHL 成員中 UCH-L1 在腦內約佔重量比(1-2%),此外有研究指出 UCH-L1 與腦內一些疾病如 阿茲海默症以及帕金森氏症有關,他們在病患腦內發現到有大量的 UCH-L1 蛋白,另外 在某些癌細胞上常會發現 UCH-L1 蛋白過度表現增生的現象(72)。 § 61 knot
61 knot又稱為雙八字結(stevedire knot),其中唯一的例子為α-haloacid dehalogenase, 他是目前為止發現到最複雜的結,科學家發現土壤中戀臭假單胞菌(Pseudomonas putida) 的α-haloacid dehalogenase可以將水解haloacid的碳鹵鍵(73)。
(S)-2-haloacid + H2O (R)-2-hydroxyacid + halide
1-13-2 扭結蛋白生物物理性質研究
2002 年 Mohammad Taufiq Alam 透過原子力顯微鏡觀察三葉型扭結蛋白-牛碳酸酐 酶 B(bovine carbonic anhydrase B)受機械力後情形,透過固定蛋白兩端後用力往兩邊拉 扯,發現扭結蛋白被拉扯後的實際距離來得比預估距離要來的短(74)。此外在 2009 年 Thomas Bornschlögl 等 人 也 同 樣 利 用 原 子 力 顯 微 鏡 來 拉 扯 八 字 結 蛋 白 - 光 敏 色 素 (phytochrome) (75),其實驗結果和 Bargia A 相輔,八字結在被拉扯後的距離比預估距離 來的短。 1-13-3 扭結蛋白質結構對蛋白穩定性的影響 蛋白質的降解是細胞運作中重要的一環,主要在於清除過多或者錯誤的蛋白,避免 在真核生物中造成細胞的毒性,首先有待降解的蛋白質被一種多泛素(ubiqutin)所標記, 接著這些蛋白質進入細胞的蛋白酶複合體進行水解。其中蛋白酶體是在真核生物和古菌 中普遍存在,在真核生物中,蛋白酶體位於細胞核和細胞質中,蛋白酶複合體是一個上 下有蓋的圓桶狀酵素,其結構上包括一個由四個堆積在一起的環所組成的核心(nuclus), 其中空直徑約為 13nm,如同細胞的垃圾桶,專門負責蛋白質的分解及再循環利用,泛 素在這一過程中釋出訊號,讓蛋白酶複合體分辨出有待降解的蛋白質,一旦蛋白質被標 記上一個泛素分子,就會引發接上更多的泛素分子,從而標記的蛋白質帶到蛋白酶體開 始降解。因此結構穩定的蛋白就算泛素標定,再送往蛋白酶體水解過程其立體上的障 礙,使得很難被蛋白酶體拉進去阻礙水解作用,挽救蛋白被降解的命運(56)。
1-13-4 扭結蛋白之摺疊研究
2007 年 Anna L. Mallam 以及 Sophie Jackson 以三葉型扭結蛋白 Yibk、YbeA 以及八 字結蛋白 UCH-3 為對象,透過化學變性後將這些扭結蛋白構過透析重新摺疊,然後透 過螢光光譜儀以及圓二色光譜譜儀分別觀察其變性前以及復性後的疏水性結構以及二 級結構差異,結果顯示扭結蛋白在化學變性後可以重新摺疊回原來的構型,屬於可逆的 反應(43, 76)。
此外 2008 年他們在 Yibk 以及 YbeA 的 N 端、C 端以及兩端分別連接 ThiS 小蛋白 試圖阻礙結的生成。其中 ThiS 為嗜高溫古生菌(Archaeoglobus fulgidus)的蛋白質,本身 由 92 個胺基酸所構成,在這裡 ThiS 做為一個阻止蛋白肽鏈穿過環生成扭結的立體障 礙,根據 x 光立體繞射結晶以及受質接合力分析結果指 ThiS–Yibk/ThiS–Yibk-ThiS/Yibk-ThiS 以及 ThiS–YbeA/ThiS–YbeA-ThiS/YbeA–ThiS 均有 扭結的構型,表示接合 ThiS 不阻礙扭結生成,於是他們推測扭結可能在蛋白摺疊生成 二、三級結構初期就形成(44, 77)。 因此我們希望透過基因重組蛋白摺疊的方式,了解此一扭結的結構的變化,並嘗試 解析其摺疊機制並嘗試建立數個扭結相關突變株,以了解扭結結構對蛋白摺疊的影響。 因此本研究特別針對扭結蛋白的熱穩定性、酵素水解穩定性以及扭結在摺疊過程的變 化,以了解扭結區域對整體蛋白結構影響。
1-14 實驗策略
第二章 實驗原理與方法
2-1 基因構築
PCR 技術是一個普遍的不可缺少的技術,目前廣泛運用於醫學和生物學(78)。是由 1983 年科學家 Kary Mullis 發明,其做法是在欲擴增的 DNA 片段兩端分別設計一個前置 引子(forward primer)和反置引子(reverse primer)透過高溫使雙股 DNA 打開雙股,再透過 降溫讓單股 DNA 和設計的單股 primer 黏合配對(annealing)後,利用 DNA 聚合酵素(DNA polymerase),以目標 DNA 的兩股分別做為模板(template)來合成新的 DNA 股。經由變 性反應(denaturation),使 DNA 的兩股分離。緩冷配對反應(annealing),使引子與目標 DNA 配對。延長反應(extension),合成新的 DNA 股。
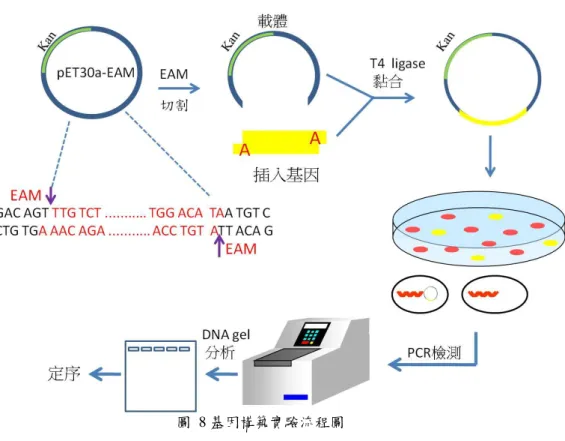
在這裡我們以 PCR 的方式放大目標基因後,然後和表現載體 pET30a EAM 進行黏 合,在將黏合後基因送入大腸桿菌 BL21 DE3 宿主細胞後,透過 PCR 以及定序做最後基 因確認(圖 8)。
2-1-1 ybeA wild type 基因構築
YbeA 本身為 E coli 中的蛋白質。我們選擇使用 E. coli strain K-12 substn DH10B (GI code,169887498)的染色體作為模版,該基因序列如
表 1所示,灰底部分為 ybeA 基因序列。在這裡我們以引子 ybeA 4 fp 及 ybeA 4 rp
擴大 ybeA 基因,再透過 ybeA start fp 以及 ybeA 4 進行 nest PCR,其中在 nest PCR 時使 用 pro-taq DNA polymerase(PROTECH® ; Miao-Li, TW),利用 dATP 在 ybeA 的 3’端接上 A,然後將 PCR 產物 clean up 後接進載體 pET30a EAM 內,送入大腸桿菌 BL21 DE3 宿 主細胞後,透過 PCR 以及定序做最後基因確認。其中在 pET30a EAM 切位有多一個 T 可以和 TA clony 的 ybeA 的 3’A 末端做黏合,因此重組後的 ybeA 基因除了原本的基因 以外,還多了載體本身 150 個核甘酸。 PCR 實驗條件: Primer Sequence Tm bp YbeA 4 fp 5’-GGATTGTCGTGGATTTGG-3’ 42.9°C 18 YbeA 4 rp 5’-CTCTGCCGTATAGTCGC-3’ 44.3°C 19 YbeA start fp (1~24) 5’- GTGAAGCTGCAACTTGTCGC -3’ 60.7°C 20 PCR 條件
Buffer Volume(μl) Temperature Time Cycle
dd H2O 40.5 94°C 2 mins 1 1 mM dNTP 1 94°C 30 secs 30 YbeA 4 fp 1 42°C 30 secs YbeA 4 rp 1 72°C 30 secs 10x buffer 5 72°C 5 mins 1 pfx 0.5 Template DH10B 1 Total volume 50
表 1 E. coli strain. K-12 substn. DH10B 的 ybeA 基因序列 1 c c a g c a t c a c c g a c t g c a t g a t c a t c t g t a c g g g t a c g t c c a g c c g t c a t g t t a t g t c c a 6 1 t t g c t g a c c a c g t t g t g c a g g a g t c t c g c g c a g c g g g c c t g t t a c c g c t c g g c g t a g a a g 1 2 1 g t g a a a a c a g c g c c g a c t g g a t t g t c g t g g a t t t g g g c g a t g t g a t t g t c c a t g t c a t g c 1 8 1 a g g a a g a g a g c c g t c g c c t g t a t g a a c t g g a a a a a c t c t g g a g t t a a t g c g t g a a g c t g c 2 4 1 a a c t t g t c g c c g t g g g a a c g a a a a t g c c g g a c t g g g t a c a a a c c g g t t t t a c c g a g t a c c 3 0 1 t g c g t c g t t t t c c g a a a g a t a t g c c c t t c g a g c t g a t t g a a a t t c c g g c c g g a a a a c g c g 3 6 1 g c a a g a a t g c g g a c a t c a a g c g c a t a c t c g a c a a a g a g g g t g a g c a g a t g t t g g c g g c c g 4 2 1 c a g g c a a a a a c c g c a t t g t c a c c c t c g a t a t t c c a g g c a a g c c c t g g g a t a c g c c g c a g t 4 8 1 t a g c c g c t g a g c t g g a a c g c t g g a a g c t g g a t g g t c g c g a c g t c a g t c t a c t g a t t g g c g 5 4 1 g g c c t g a a g g g t t g t c g c c t g c c t g t a a a g c g g c g g c t g a g c a g a g c t g g t c g c t g t c g g 6 0 1 c g c t t a c c c t c c c c c a t c c g c t g g t t c g c g t g c t g g t c g c a g a g a g t c t g t a c c g g g c g t 6 6 1 g g a g c a t c a c c a c c a a c c a t c c t t a t c a c c g t g a g t g a t a a g g g a g c t t t g a g t a g a a a a
2-1-2 ybeA△73-75 基因構築
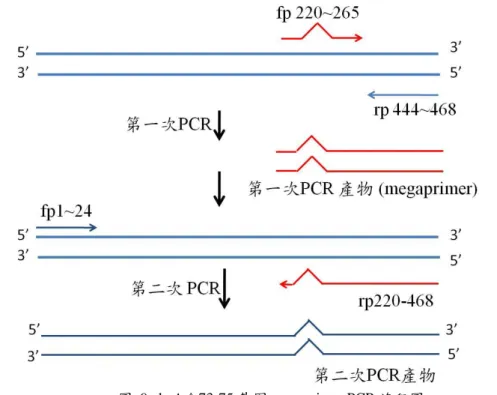
ybeA△73-75 突變株基因構築是以先前構築好的 ybeA 基因為模板,其中 ybeA 的基 因序列如表 2 所示,灰底的部分為 fusion 片段,ATG 為起始密碼子,TGA 為終止密碼 子。在這裡我們利用 megaprimer 方式將 ybeA 的 73~75 號(DIP)胺基酸所對應的 DNA 序 列去除掉,其實驗流程如圖 9 所示。其中 megaprimer PCR 擴增可分為兩部分,先以 asymmetry PCR 方式擴增包含突變位置的 megaprimer (220~468),以作為下一步 PCR 的 primer,其中 megaprimer 的 5’端至 3’端的單股 DNA 必須較多,以作為第二次 PCR 的 reverse primer。
因此 asymmetry PCR 的步驟時,我們將 ybeA DIP forward primer (220~240 bp)濃度 增加,讓 forward primer 的濃度高於 reverse primer(444~468 bp)20 倍,第一次 PCR 完後, 須先 cleanup 後去除多餘的 oligonucleotide,再進行第二次 PCR,利用 TA clony 方式將 ybeA DIP del gene 接進 pET30a EAM 載體然後利用 PCR 進行篩選,所選用之 primer 為 ybeA DIP del fp(220~248)及 T7 rp。我們之所以選擇 ybeA DIP del fp(220~248)作為 forward primer,是因為該 primer 可以鍵結在 ybeA DIP del gene 上卻無法鍵結在 ybeA wt gene 上, 故可以利用此點來區別 ybeA DIP del gene 與 wt gene 之間的區別。
圖 9 ybeA△73-75 基因 megaprimer PCR 流程圖
圖中 fp(220-265)為第一次 PCR 的 forward primer,rp444-468 為第一次 PCR 的 reverse primer,而第二次 PCR 的 forward primer 為第一次 PCR 產物 megaprimer 而 reverse primer 和第一次 PCR 的 reverse primer 相 同。 PCR 實驗條件: Primer Sequence Tm bp YbeA DIP fp (220~248) 5’-ACCCTCGGCAAGCCCTGGGAT-3’ 60°C 21 YbeA DIP rp (440~468) 5’-TCACTCACGGTGATAAGGATGGTTG-3’ 57°C 25 YbeA start fp (1~24) 5’- GTGAAGCTGCAACTTGTCGC -3’ 60.7°C 20
第一次 PCR 條件
Buffer Volume(μl) Temperature Time Cycle
d.d H2O 40.5 94°C 1 min 1 1 mM dNTP 1 94°C 30 secs 25 YbeA 4 fp 1 57°C 30 secs YbeA 4 rp 1 72°C 30 secs 10x buffer 5 72°C 7 mins 1 pfx 0.5 Template DH10B 1 Total volume 50 第二次 PCR 條件
Buffer Volume(μl) Temperature Time Cycle
d.d H2O 23 94°C 1 min 94℃ 1 mM dNTP 1 94℃ 30 secs 94℃ 60℃ 72℃ 5 μM fp1~24 1 60℃ 30 secs 220~468 bp megaprimer 18 72℃ 30 secs 10x buffer 5 72℃ 7 mins 72℃ Pro-taq 1 Template (ybeA pET 30a) 1 Total vol 50 表 2 重組 ybeA 基因序列
1atgcacc a tc a tca tc a tca ttcttc tg gt c tgg tgc cac g cg gttc tgg ta tgaaa gaa 61accgc tgc tg c taaa ttc ga a c gc cagca c a tg ga cagc c ca gatc tg gg tac cga cga c 121gac gacaag g cc a tg gc tga tta cga cag t g tgaag c tgc a acttg tc gc c gtgg gaacg
181aaaatgcc gg ac tg gg tac a aaccgg tttt ac cga gta cc tg cg tcg ttt tc cgaaa gat 241atgcc c ttcg ag ctga ttga aattcc ggc c ggaaaa cgc g gc aagaatg c gga catcaag 301cgcata c tcg a caaagag g g tga gcaga tg ttg gcg g c cg c aggcaaaaa c cgca ttg tc 361accctcga ta ttccag gcaa gccc tggga t ac gcc gca gt ta gcc gc tga g ctggaa cgc
421tggaag c tgg a tg gtcg cga cgtcag tc ta c tga ttgg cg ggc ctgaagg g ttg tc gc ct 481gcc tg taaag c ggc gg ctg a gcagag ctgg tc gc tg tc gg c gc tta ccc t c cc cca tcc g 541ctg gttc gc g tgc tgg tcg c agagag tc tg tac cgg gc gt gg agcatcac ca ccaa cca t 601c ctta tca cc gtgagtga
2-1-3 ybeA△1-8 基因構築
ybeA△1-8 突變株構築是以先前構築好的 YbeA 基因為模板,透過引子 ybeA(175) fp 及 ybeA (627) rp 進行 TA cloning 來擴增 ybeAΔ1-8 基因,然後將 PCR 產物 clean up 與 pET 30a EAM 做黏合,再利用 primer T7 fp 及 ybeA (627) rp 進行 PCR 篩選。
PCR 實驗條件:
Primer Sequence Tm bp
ybeA (175)fp 5’-GGAACTAAAATGCCTGACTGG-’ 52.3oC 21
ybea (627)rp 5’-GCTCCCTTATCACTCACGG-3’ 53.2oC 19
PCR 條件
Buffer Volume(μl) Temperature Time Cycle
dd H2O 20 94°C 1 min 1 1 mM dNTP 0.5 94°C 30 secs 30 ybeA (175)fp 0.5 53°C 30 secs ybea (627)rp 0.5 72°C 30 secs 10x buffer 2.5 72°C 5mins 1 Pro-taq 0.5
Template(pET 30a- ybeA) 1
Total volume 25
2-1-4 ybeA△1-70 基因構築
以先前實驗室建構好的 ybeA 基因為模板,透過 ybeA(361)fp 及 ybeA(627)rp primer 近進行 TA cloning 擴增 ybeA Δ1-70 基因,然後將 PCR 產物經過 clean up 與 pET30a EAM 進行黏合,再利用 primer 為 T7 fp 及 ybeA (627)rp 利用進行 PCR 篩選。 PCR 實驗條件: Primer Sequence Tm bp ybeA (361)fp 5’-ACACTAGATATTCCAGGCAAGC-3’ 53oC 22 ybea (627)rp 5’-GCTCCCTTATCACTCACGG-3’ 53.2oC 19 PCR 條件
Buffer Volume(μl) Temperature Time Cycle
dd H2O 20 94℃ 1 min 1 1 mM dNTP 0.5 94℃ 30 secs 30 ybeA (361)fp 0.5 53℃ 30 secs ybea (627)rp 0.5 72℃ 30 secs 10x buffer 2.5 72℃ 5mins 1 Pro-taq 0.5 Template (ybeA
pET 30a EAM) 1
2-1-5 ybeA△118-155 基因構築
以先前實驗室建構好的 ybeA 基因為模板,透過 ybeA(151)fp 及 ybeA(501)rp 進行 TA cloning,將 PCR 產物經過 clean up 與載體 pET30a EAM 進行黏合再利用 T7 fp 與 ybeA (627) rp 進行篩選。 PCR 實驗條件: Primer Sequence Tm bp ybeA (151)fp 5'-GTGAAACTGCAACTTGTAGCC-3' 52.4oC 21 ybea (504)rp 5’-CGACAAGTTGCAGCTTCAC-3’ 54.5oC 19 PCR 條件
Buffer Volume(μl) Temperature Time Cycle
dd H2O 20 94℃ 1 min 1 1 mM dNTP 0.5 94℃ 30 secs 30 ybeA (151)fp 0.5 53℃ 30 secs ybea (501)rp 0.5 72℃ 30 secs 10x buffer 2.5 72℃ 5mins 1 Pro-taq 0.5 Template (ybeA pET 30a
EAM)
1
2-2 蛋白的製備
將所需的基因接合在 pET30a EAM 載體後,送入大腸桿菌 BL21 DE3 中以乳糖異構物 IPTG 誘導 16 小時以求獲得大量表現的蛋白質內含體。其中 IPTG 可以誘導乳糖對照組 (lac peron)活化表現下游基因。其表現原理在於在大腸桿菌 BL21 DE3 的乳糖對照組可以 產生大量的 T7 RNA polymerase 表現,然後 T7 RNA polymerase 再去活化 pET30a 上的 T7 promoter,使得我們建構的基因得以大量表現,而誘導物-乳糖會被乳糖對照組所產
生的β-galatosidase 會分解終止反應;然而 IPTG 卻不會被分解,因此可以不斷的產生大
量的蛋白。
2-2-1 蛋白質小量表現
實驗步驟:
(1)從-80oC 取 BL21 DE3 菌種含有 pET30a 帶有 ybeA 基因或突變株基因的菌,待溶化後
取出 60μl 加入 3 ml 的 LB 培養液(含 20 ng/mL Kanamycin),於 37℃,200 rpm 震盪隔天 培養活化菌株。 (2)取活化菌液 3μl 加入 3 mL LB 培養液(含抗生素 Kanamycin 20 ng/mL),37℃,200rpm 震盪培養至 OD 0.6-0.8 後加入 IPTG,使 IPTG 最終濃度為 1 mM,持續震盪培養 12-16 小時。 (3)抽取 1 ml 菌液,以 12000 rpm 離心 1 分鐘,將沉澱的菌體以 100μl 的 sample buffer(10 mM Tris-HCl,pH6.8,4 % SDS,0.2 % Bromophenol Blue,20 %Glycerol,200 mM DTT)回 溶。以超音波震盪器(UP200s,Hielsher,Teltow,Germany)打破細胞壁,透過 15 % SDS-PAGE 確認是否有目標蛋白大量表現。
2-2-2 蛋白質大量表現
實驗步驟:
(1)從-80oC 取 BL21 DE3 菌種含有 pET30a 帶有 ybeA 基因或突變株基因的菌,待溶化後
取出 60μl 加入 3 ml 的 LB 培養液(含 20 ng/mL Kanamycin),於 37℃、200 rpm 震盪隔天 培養活化菌株。 (2)取活化菌液 250μl 加入 250 mL LB 培養液(含抗生素 Kanamycin 20 ng/mL),37℃、200 rpm 震盪培養至 OD 0.6-0.8 後加入 IPTG,IPTG 最終濃度為 1 mM,持續震盪培養 12-16 小時,離心獲得菌體,使用二次水回溶菌體,利用破菌機打破細胞壁。 (3)將破碎菌液以(10000 rpm 4 oC 離心 10 分鐘)後,移除上清液,收集蛋白沉澱物持續以 二次去離子水回溶、離心、去除上清液,清洗至上清液完全澄清。 2-2-3 蛋白摺疊
在這裡我們透過實驗室建構的準靜態摺疊法(Quasi-static-like thermal equilibrium dialysis)將蛋白質摺疊回自然態。
實驗步驟:
(1)將內含體 (Inclusion body)透過 denature buffer 回溶,再將回溶的蛋白透過離心移除沉
澱物,再將離心後蛋白溶液,透過孔徑為 0.22μm PVDF 濾膜進行過濾。然後將過濾好
的蛋白溶液置入孔徑為 3.5 kDa 的透析膜後,置入 R1緩衝液做透析,此時摺疊中間物稱
為 M1。
(2)將摺疊中間物 M1置入 R2緩衝液透析,此時摺疊中間物稱為 M2。
(4)將摺疊中間物 M3置入 R4 緩衝液透析,此時摺疊中間物稱為 M4。
(5)將摺疊中間物 M4置入 R5 緩衝液透析,此時摺疊中間物稱為 M5或摺疊態。
2-3 圓二色光譜(Circular Dichroism Spectroscopy)
圓二色光譜是一種特殊的吸收光譜,它對手相分子的構型相當敏感,其中手相分子 具有光學活性,當單色左旋光及右旋光偏振光通過樣品時,對左旋光及右旋光偏振光有 不同吸收就叫做圓二色性。圓二色光譜技術(Circular Dichroism),早在 1969 年就由 Greenfield 利用來估計蛋白質的二級結構,是一種快速簡單且相當準確的方法。圓二色 光譜可以用來分析生物樣品的結構像是蛋白質以及核酸分子,在遠紫外線旋光光譜波段 (180-250 nm)主要用於偵測蛋白質二級結構,而在近紫外線旋光光譜波段(250-350 nm) 主要偵測蛋白質胺基酸的殘基,蛋白質二級結構特徵鋒如表 3 所示。 表 3 蛋白質二級結構特徵鋒 -band(nm) +band(nm) α 螺旋 222 192 208 β sheet 216 195 220(week) 203(strong ) 205 Polypro II 螺旋 190 210-230 Random coil 200 212 實驗步驟: (1)調整蛋白質濃度為 0.5 mg/ml。 (2)打開氮氣持續吹 20 分鐘以上,設定水浴槽溫度為 20 oC,開啟 Xe LAMP power 以及 CPU/INST power (3)打開 AVIV 程式,設定實驗參數 (3)取體積 250 μl 的樣品加入光徑為 1 mm 的石英樣品槽(quarz cell)
實驗參數:掃描波長為 200~260 nm,band width 是 0.5 nm,interval time 是 10 秒。
2-4 螢光光譜儀(Fluorescence Spectroscopy)
2-4-1 螢光原理
螢光分子吸收外界光原能量 hvEX 使其能量跳至 S1'後,受激螢光分子產生一些變
化,並與外在環境產生許多化學反應,使能量從 S1'掉到 S1 再掉到 S0,放出能量 hvEM,
圖 10 螢光原理 2-4-2 蛋白質螢光原理 蛋白質內部主要由疏水性胺基酸所構成,在這些胺基酸中的芳香族類胺基酸 Trp、 Tye、Phe 有著特殊的光學性質可以吸收波長為 280 nm 的光,其中 Trp 更可以吸收 295 nm 的光,屬於蛋白質分子內的螢光基團。當這些螢光基團分子受到光激發後會造成內部電 子躍遷釋放出螢光;倘若這些螢光基團處於疏水性環境底下,其分子內部共振結構相當 穩定,在受到光激發後能量不容易以熱能型式發散;然而當處於親水性環境底下,其螢 光基團結構相當不穩定,在受到光激發之後,分子內部共振相當明顯,造成釋放出的螢 光相當微弱。根據這些螢光基團的螢光強度可以用來分析其所處的蛋白質的摺疊狀態, 當蛋白質內部核心於疏水性環境底下環境底下,這些螢光基團受光激發後仍相當穩定; 然而若是受到一些變性因子如高溫、酸鹼等外在因素影響,造成螢光基團暴露在水溶性 環境,則會導致讓螢光能量下降,以及波峰發生紅位移現象。 實驗步驟: (1)調整蛋白質濃度為 0.1 mg/ml。 (2)水浴槽溫度為 25 oC。
(3) 打開程式連結電腦與儀器(Hitachi F-4000, Scientific Equipment Source Inc , Canada), 設定實驗參數。
(4)取體積 700-800 μl 的樣品加入光徑為 1 cm 的石英樣品槽(quarz cell)
實驗參數:掃描波長為 310~450 nm,延遲時間(delay time)為 10 秒,EX Slit:5.0 nm,EM Slit: 5.0 nm,波長間隔為 0.2 nm,掃瞄速度是 1200 nm/min。
2-5 恆溫滴定微卡計(Isothermal Titration Calorimetry)
透過恆溫滴定微卡計可以得知生化反應或分子間交互作用力的大小,如酵素以及受 質間的交互作用力以及核酸分子間的交互作用力。恆溫滴定微卡計其原理是在恆溫環境 下觀察生化反應或分子間交互作用,將欲觀察的反應物透過滴定的方式滴入另一欲觀察 反應的反應系統中,分析在恆溫環境底下反應熱的變化。在這裡我們利用 ITC 偵測 YbeA 蛋白與其受質 S-Adenosyl methionine(AdoMet)之間的交互作用力。 實驗步驟: (1)樣品偵測前要先用孔徑為 0.22μm 過濾膜過濾樣品,並透過抽真空幫浦以及超音波震 盪器去除氣泡。 (2)在樣品端加入 2.8 ml 的樣品溶液,而參考端的反應槽加入 3 ml 和樣品端相同的緩衝 液,緩慢將參考端以及樣品端放進反應槽內。
(3)在外部注射器加入約為 300 μl 的反應物,將前端細管插入反應槽內,打開攪拌器, 等待平衡穩定後開始做測定。
(4)設定實驗參數(注射器以每個十分鐘滴入 10 μl 的樣品,共二十次,最終滴入 200 μl, 最後樣品槽的體積達 3ml 和參考端相同)
2-6 介質輔助雷射脫附游離飛行式質譜儀 (MALDI-TOF-MS)
基質輔助雷射脫附-飛行時間質譜法(MALDI-time of flight, TOF),即為結合基質輔助 雷射脫附離子化技術與飛行時間質譜分析的應用。 MALDI 採用短的脈衝鐳射 (1~10 ns)與高電壓使樣品分子離子化並加速之後進入 飛行管後以進行質譜儀分析。將處於固相或粘稠的液相狀態樣品與基質的混和物經由雷 射脈衝的撞擊,激發基質分子並將能量與電荷轉移至樣品分子,樣品分子接受能量與電 荷之後使得接近表面的分子相斥分離,釋出帶電的氣態離子,然後藉由電場加速後進入 飛行管中進行自由飛行(free flight),透過其飛行時間的長短可得到其精確的質荷比 (m/z)。最常被使用的基質(matrix)有三種:α-氰基-4 -羥基肉桂
(α-cyano-4-hydroxycinnamicacid)、2,5-二羥基苯甲酸 2,5-dihydroxybenzoic acid (DHB)與 芥子酸 sinapinic acid (SA)。
在這裡我們透過胰蛋白酶分別對 YbeA 摺疊態以及未摺疊態進行水解作用,然後將 水解產物透過(MALDI-TOF-MS)來進行分析。
樣品前處理
(1)取適量 YbeA 樣品於 1.5 ml eppendorf 中後加入 trypsin(promega) 使之莫耳比例為 25:1 及 50:1。
(2)置於 37 ℃恆溫水槽中進行酵素消化反應。
(3)每 1、2、3……分鐘取一次 (約取 5 μl) 開始消化之 YebA,將其注入裝有等體積之 50 % ACN+ 0.1 % TFA 的 eppendorf 中終止消化反應。(各取出每一個時間點取出的消化 產物 0.5 μl 混合 0.5 μl 基質點於 384 well 的 ground target 上。
(4)待乾燥結晶後,推入 MALDI-TOF-MS 分析。
(5)根據質譜圖,使用 MASCOT 軟體線上搜尋 YebA,根據其及格分數及 cover 到之片 段進行結果判斷。
2-7 螢光共振能量轉移(Förster resonance energy transfer)
螢光共振能量轉移可以用來比較分子距離以及分子內直徑的有效工具,廣泛的應用 於涉及分子間距離變化的生物現象,可以用來測量兩個發光基團間的距離及蛋白質蛋白 質間的交互作用關係以及核酸分子及蛋白間的交互作用。當螢光基團被光照時,被激發 的分子可以透過散發能量回到基態,其中釋出的光能可被發光基團在 10-15 秒內吸收而 在 10-9秒內再發射出來。然而也有可能被激發分子並不發光,而將能量傳遞另外的螢光 基團,這些螢光基團可以在相同的時間內發螢光,稱為螢光共振能量轉移(FRET)。其 中產生螢光共振能量轉移(FRET)要具備三個要素,以下將分別做說明。 (1) 供體和受體的距離必須在 10-100 Å 之間 (2) 受體的吸收光譜與供體的發射光譜重疊 (3) 供體和受體能量轉移偶極子的方向必須近似地平行 在這裡我們透過 FRET 的方式分析 YbeA 扭結區域在摺疊過程中的距離的變化。其 中 YbeA 扭結區域分子內有三個 Trp 以及一個 cysteine 分子,透過在 cysteine 接上螢光 基團 AMCA 以波長 295 nm 的激發光來激發 Trp 分子,然後計算 Trp 與 AMCA 的螢光 能量轉移率進而推得螢光基團在摺疊過程中距離的變化。
其中 AMCA 的激發波長在 340-350 nm 而發射波長在 440-460 nm。而 Trp 的激發光 波長在 295 nm,而發射光波長為 345-350 nm 左右。
實驗步驟:
(1)取適量 YbeA 樣品溶於 10mM Tris–HCl (pH 8.8)置於 1.5 ml eppendorf 中,後加入 AMCA-HPDH 使之莫耳比例為 10:1,於室溫底下避光靜置 1 個半鐘頭,待其反應完全。 將 YbeA 及 AMCA-HPDH 混和物置入 3.5 kDa 的透析膜,置入 10mM Tris -HCl (pH 8.8) 做透析,去除多餘未接合的 AMCA-HPDH 分子。
(2)藉由紫外光可見光譜儀來計算 YbeA 以及 AMCA-HPDH 的濃度來計算接和率。 (3)利用波長 295 nm 的光源激發 Trp 分子,分析 Trp 以及 AMCA 之螢光能量轉移效率來 分析扭結區域的距離變化。
2-8 力學截流儀(stopped -flow)結合圓二色光譜(CD Spectroscopy)
圖 11 動力學截流儀結合圓二色光譜儀裝置 動力學截流儀可以偵測溶液中非常快速的反應,將兩種溶液經注射流入混合室中迅 速地混合,透過活塞來控制流入的體積,結合光學儀器來分析反應速率的變化。在這裡 我們透過在這裡我們透過動力學截流儀結合圓二色光譜儀快速混合,來分析蛋白在摺疊 以及變性的反應速率變化。 實驗步驟: (1)配置蛋白質濃度為 22 mg/ml。 (2)樣品偵測前用孔徑為 0.22 μm 過濾膜過濾樣品。 (3)開啟液態氮氣鋼瓶,打開水浴槽設定溫度 19 oC,待液態氮開啟後 20 分鐘,開啟 Xe LAMP power。
(4)開啟 CPU 控制閥,打開電腦,開啟 AVIV stopped-fow CD 程式。
(5)開啟 stopped-flow 程式,將注射器歸位(home syringe),設定 syringe 體積(syringe1:1 ml,syringe 2:2 ml, syringe 3: 5 ml)。
(6)利用二次去離子水清洗樣品注射器。
(7)將蛋白溶液放入 syringe 1,而將欲混和之緩衝液置入 syringe 3。
(8)設定實驗參數(平衡 200 秒,混和時間 50 秒,偵測時間 2 分鐘,偵測波長 222 nm,掃 描次數四次,系統溫度 25 度)
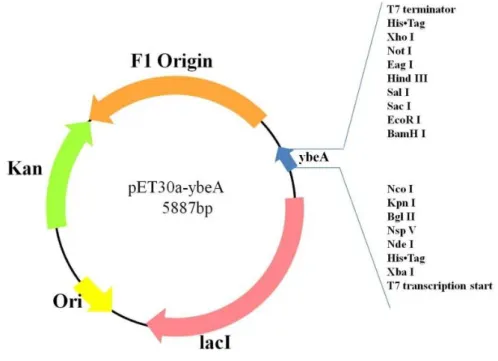
第三章 實驗結果 3-1 基因構築結果 3-1-1 pET30a-ybeA 質體構築 此基因由本實驗驗室所構築,圖 12 為質體 pET30a-ybeA 的建構圖,在這裡我們選 用大腸桿菌 K-12 DH10B 的染色體為模板來擴增 ybeA 基因,DH10B 染色體本身有 4686137 bp,而 ybeA 基因則位在 DH10B 染色體基因 607270 bp 之間,我們透過 nest PCR 方式來擴增 ybeA 基因,圖 13(A)為 DH10B 染色體為模板,透過引子 ybeA 4 fp 及 ybeA 4 rp 擴增含 ybeA 基因的結果,在 lane1 部分 600~700bp 位置有單一片段,其大小與 ybeA 4 fp 及 ybeA 4 rp PCR 擴增預計長度 592bp 符合;圖 13(B)為 ybeA start fp 以及 ybeA rp 進行 PCR 結果,lane1 在 400-500bp 位置有單一片段其大小與預期片段大小 461 bp 吻合。
再將我們擴增好的 ybeA 基因與 pET30a EAM 載體進行黏合,送入大腸桿菌 BL21
DE3 菌株,圖 13(C) 為透過 T7 fp 以及 ybeA 4 rp PCR 篩選挑選出來的菌株凝膠電泳結
果,在 lane 3 部分 800~900 bp 位置有單一片段,其大小與預計片段 811 bp 吻合,因此 菌落 3 可能有 ybeA 基因,於是我們將菌落 3 送定序,而圖 14 為定序結果與 NCBI 上 公布的 ybeA 基因比對,其序列相符,我們確定菌落 3 確實含 pET30a-ybeA 基因。
圖 13 pET30a-ybeA 質體構築結果
圖中以菌株 DH10B 的染色體作為模版,利用引子 ybeA 4 fp 及 ybeA 4 rp 來擴增 ybeA 基因結果,lane M 為 DNA ladder,lane N 為負對照組,lane 1 為 DH10B 的染色體,利用引子 ybeA 4 fp 及 ybeA 4 rp 擴增結 果,箭頭所指為預期片段位置。圖(b)是以 start fp 以及 ybeA 4 rp 擴增結果,箭頭所指為預期片段位置。 圖(c)為透過 T7 fp 及 ybeA 4 rp 引子來 PCR 檢測挑出來的菌落是否有 ybeA 基因,lane M 為 DNA ladder, lane N 為 nagative control ,lane1-3 是挑出來菌落, 箭頭所指為預期片段位置,DNA 電泳使用 1.5% Agarose gel 及 0.5x TBE buffer。
圖 14 重組 ybeA 基因與 NCBI 公布之 ybeA 基因的比較結果。
圖中 NCBI -ybeA 為 NCBI 上所登錄的 ybeA DNA 序列,而 r-ybeA 為我們所構築的 ybeA 基因定序結果, 其中星號部份為有對應到的基因序列,我們所擴增的 ybeA 除了前面多了載體的 his-taq 融和片段外,其 餘序列完全符合 NCBI 所登錄的資料。
3-1-2 pET30a-ybeA△73-75 質體構築
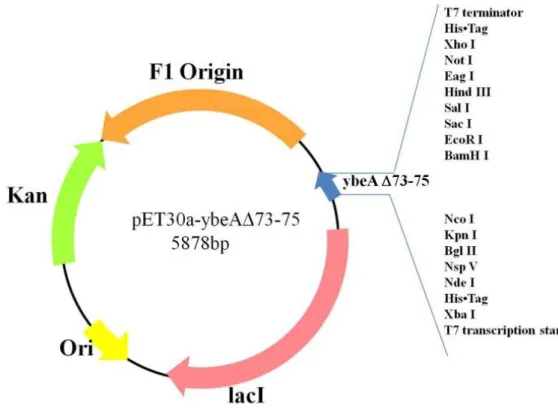
此基因由本實驗室所構築,圖 15 為質體 pET30a-ybeA△(73-75)的建構圖,我們以 pET30a-ybeA 為模板,透過 megaprimer PCR 方式來去除 DIP 胺基酸對應核甘酸,其中 ybeA△73-75 的 PCR 主要分成兩部分,第一個部份我們先以 ybeA DIP del fp (220~248) 以及 ybeA DIP del rp (440~468)為引子,擴增含突變位置的 megaprimer (220~468)以作為 下一步 PCR 的 reverse primer。而第二次 PCR 時以 pET30a-ybeA 為模板以 ybeA start fp(1~24)做為 forward primer 而第一次 PCR 的 megaprimer (220~468)做為 reverse primer, 擴增 ybeA △(73-75)基因。
圖 16(a)為 ybeA △(73-75)基因擴增結果,lane1 在 400~500bp 位置有單一片段其大
小與預計長度 492bp 符合,將我們擴增好的 ybeA△(73-75)基因與 pET30a EAM 載體進
行黏合,再送入大腸桿菌 BL21 DE3 菌株,圖 16(b)為利用 ybeA DIP del fp (220~248)及
T7 rp 塞選挑出來菌落,菌落 7、8、17 在 400-500bp 位置有單一片段其大小與預期片段 大小 485 bp 吻合,因此菌落 7、8、17 可能有 ybeA△(73-75)基因,於是我們將菌落送定 序,而圖 17 為重組 YbeA 與重組 ybeA△(73-75) 基因序列比較結果,綜合以上結果, 我們確定菌落確實含 pET30a-ybeA△(73-75)基因。
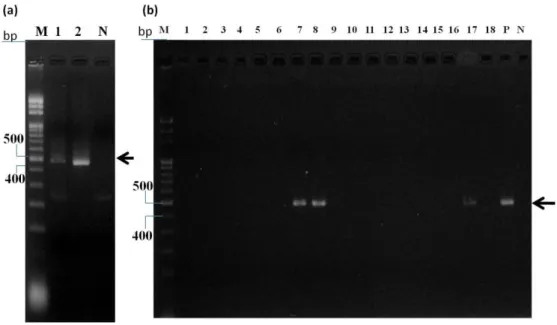
圖 16 pET30a-ybeA△(73-75)基因建構結果
(a)圖為利用 megaprimer 方式擴增 ybeA 去除胺基酸 DIP 相對基因結果,lane M 為 DNA ladder,lane 1 為 ybeA △(73-75),lane P 為正對照組,lane N 為負對照組,箭頭所指處為預計片段大小位置,在這裡 DNA 電泳使用 1.5 % 瓊脂膠體及 0.5 x TBE buffer;圖(b)為透過引子 ybeA DIP del fp(220~248)以及 T7 rp PCR 篩選挑選出來的 17 個菌落,lane M 為 DNA ladder,lane 1-17 為挑選出來的挑選 17 顆菌落,lane P 為正 對照組,而 lane N 為負對照組,箭頭所指處為預計片段大小位置,在這裡 DNA 電泳使用 1.5 % 瓊脂膠 體及 0.5 x TBE buffer。 圖 17 重組 ybeA 與重組 ybeA△(73-75)基因序列比較結果。 星號部份為有對應到的基因序列,方塊圈選位置為基因序列上不同處,在第一個方塊中的密碼子 CTT 靜 默突變為 CTA,此突變不影響轉譯結果,而在第二個方塊部分,ybeA△(73-75)相較於 ybeA 少了 3 個密 碼子,其相對應的胺基酸分別為 D.I.P,綜合以上結果,得知獲得 ybeA△(73-75)基因。
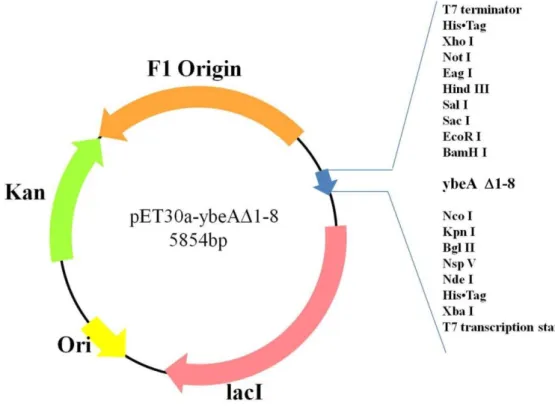
3-1-3 pET30a -ybeA△1-8 質體構築
圖 18 為質體 pET30 -ybeA△1-8 的建構圖,我們以 pET30a-ybeA 為模板,ybeA(175) fp 以及 ybeA (627) rp 做為引子擴增 ybeA△1-8 基因,圖 19(a)為 ybeA△1-8 基因擴增的 凝膠電泳結果,lane1 在 400~500 bp 位置有單一片段,其大小與 ybeA (175)fp 以及 ybea (627)rp 預計擴增長度 477 bp 符合,將我們擴增 ybeA△1-8 基因與 pET30a EAM 載體進 行黏合,再送入大腸桿菌 BL21 DE3 菌株。圖 19(b)為利用引子為 T7 Fp 以及 ybeA (627) rp PCR 篩選挑出來的菌落,其中預計擴增片段大小為 811 bp,而我們挑選出來的菌落 1、 2、4、5、6、7、8、9 在 700~800 bp 位置都有單一片段,其大小與 T7 fp 及 ybeA (627)rp 預計擴增片段大小吻合,綜合以上結果,菌落 1、2、4、5、6、7、8、9 可能有 ybeA△(1-8) 基因。於是我們將菌落送定序。圖 20 為重組 ybeA 與重組 ybeA△(1-8)基因序列比較結 果,綜合以上結果,我們確定挑出來的菌落確實含 pET30a -ybeA△1-8 基因。 圖 18 質體 pET30-ybeA△1-8 的建構
圖 19 ybeA△1-8 基因建構結果。
圖(a)為利用引子 ybeA (175) fp 以及 ybeA(627) rp 擴增 ybeAΔ1-8 結果,lane M 為 DNA ladder,lane1 為 以 pET30a-ybeA 質體為模板 PCR 擴增結果,而箭頭所指處為預計片段位置,在這裡 DNA 電泳使用 1.5 % 瓊脂膠體及 0.5 x TBE buffer;圖(b)為利用引子以 T7 fp 及 ybeA (501) rp 引子來 PCR 檢測挑出來的 10 顆 菌落,laneM:DNA ladder,lane1-10 是挑出來的 10 顆菌落,lane P 為正對照組,lane N 為負對照組。箭頭 所指處為預計片段大小位置,在這裡 DNA 電泳使用 1.5 % 瓊脂膠體及 0.5 x TBE buffer。
圖 20 重組 ybeA 與重組 ybeA△(1-8)基因序列比較結果。
其中星號部份為有對應到的基因序列,方塊部分為靜默突變不影響轉譯結果,此外重組 ybeA△(1-8)除了 最前端少 8 個密碼子以外其他序列和 ybeA 完全相同,而不見的基因的對應胺基酸分別為 V.K.L.Q.L.V.A.V. 這八個胺基酸,這與我們預期要去除的胺基酸相同,綜合以上結果,得知我們得到所需的 ybeA△(1-8)基 因。
3-1-4 pET30a-ybeA△1-70 質體構築
圖 21 為質體 pET30 -ybeA△1-70 的建構圖,ybeA (361) fp 以及 ybeA (627) rp 做為 引子擴增 ybeA△1-70 基因,圖 22(a)為 ybeA△1-70 基因擴增的凝膠電泳結果,lane1 在 200-300 bp 位置有單一片段,其大小與 ybeA(361) fp 以及 ybeA(627) rp 擴增預計長度 267 bp 符合,再將我們所擴增 ybeA△1-70 基因與 pET30a EAM 載體進行黏合,再送入大腸 桿菌 BL21 DE3 菌株。圖 22(b)為利用引子為 T7 fp 以及 ybeA (627) rp PCR 篩選挑出來 的菌落,利用 PCR 篩選挑出來的菌落,菌落 2、3、4、6。7、8、9 在 500~600 bp 位置 有單一片段,其大小與 T7 fp 及 ybeA (627) rp 預計擴增片段 514 bp 吻合,菌落 2、3、4、 6。7、8、9 可能有 ybeA△(1-70)基因,所選用之 primer 為 T7 fp 以及 ybeA (627) rp,預 計擴增片段大小為 514 bp,而我們的 PCR 擴增結果的片段大小與預期大小符合,於是 我們將菌落送定序,而圖 23 重組 ybeA△1-70 與重組 ybeA 基因序列比較結果,綜合以 上結果,我們確定挑出來的菌落確實含 ybeA△1-70 基因。