行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※
※ 整合型總計畫名稱:溫度逆境下植物基因產物
※
※ 蛋白質的分子研究
※
※ 本計畫名稱:水稻低溫耐性基因構造與功能之研究※
※ The gene structure and function of rice in chilling
※
※ tolerance.
※
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:□個別型計畫 ▓整合型計畫
計畫編號:NSC 89-2313-B-002-113
執行期間:88 年 8 月 1 日至 89 年 7 月 31 日
計畫主持人:朱鈞
共同主持人:盧虎生
協同主持人:陳勝利
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立台灣大學農藝學系
中 華 民 國 89 年 10 月 30 日
行政院國家科學委員會專題研究計畫成果報告 計畫編號:NSC 89-2313-B-002-113 執行期限:88 年 8 月 1 日至 89 年 7 月 31 日 主持人:朱鈞 執行機構及單位名稱:國立台灣大學農藝學系 共同主持人:盧虎生 執行機構及單位名稱:國立台灣大學農藝學系 計畫參與人員:陳勝利 執行機構及單位名稱:國立台灣大學農藝學系 一、中文摘要 水稻(Oryza sativa L.)在台灣及其他亞 熱帶地區早春的幼苗時期經常遭受冷害, 此冷害可能導致作物生長受阻甚至降低產 量。為了進一步探究調控冷害的分子機 制,本研究以差異表現法(differential display)並配合銀染的技術來選殖與水稻幼 苗低溫耐性表現相關的 cDNAs。以低溫敏 感的印度型水稻台中在來 1 號(TN 1)及低 溫抗性的日本型台農 67 號(TNG 67)幼苗, 經過(1)5℃;(2)25℃(對照組);(3) 10 µM ABA,25℃;或(4)10µM methyl jasmonate (MeJa),25℃;等四項各 24 小時處理後, 取其根部之 total RNA 供進行差異表現法 試驗。結果由 600 個差異表現片段中,成 功再擴增出 362 個差異表現之 cDNA 片 段,經基因庫比對結果,發現有 153 個(42.3 %) cDNA 與已知基因有極高的相似性;其 中 61 個 cDNA 分別與 rab16D、heat shock protein、SalT、Zn-induced protein… … 等 29
個已知功能基因有極高的相似性。
另外以 RNA slot blot 分析結果發現, 有 58 個 cDNA clones 在低溫或 ABA 處理 下有極高的表現。再利用北方墨點分析法 (northern-blot analysis)自 58 個差異表現的 片段中,選出 28 個在低溫、ABA 及其它 逆境處理下的表現與對照組有明顯差異的 cDNA clones。其中有部分 cDNA 分別在低 溫、ABA、MeJa、缺水、鹽害及熱休克(heat shock)處理下的水稻根部或地上部具專一 性表現。以 11 個分別具有不同低溫耐性及 ABA 內生量的水稻品種,來測試這 28 個 cDNA clones 的表現,發現有 6 個基因在根 部或地上部的表現量與電解質滲漏量呈顯 著負相關,顯示這 6 個基因與低溫耐性相 關。 低溫逆境下 ABA 前處理可導致部分 低溫耐性基因在低溫敏感的水稻台中在來 1 號幼苗中大量表現,顯示這些基因可能參 與水稻幼苗受 ABA 調節所產生的低溫耐 性。另外,部分基因在未經 ABA 前處理的 水稻幼苗中大量表現,顯然它們的表現不 受 ABA 的影響。 關鍵詞:植物荷爾蒙,差異表現法,水稻, 低溫耐性,離層酸,基因表現 ABSTRACT
Rice (Oryza sativa L.) seedlings
frequently suffer from chilling injury during early spring in Taiwan as well as in other subtropical areas. The chilling injury may cause severe growth retardation and yield reduction. To gain more insights of molecular mechanisms of chilling tolerance, cDNAs associated with chilling tolerance in rice seedlings were isolated using differential display of mRNA. Seedlings of a chilling-sensitive indica type cultivar (cv. Taichung Native 1, TN 1), and a chilling-tolerant japonica type cultivar (cv. Tainung 67, TNG 67) were treated for 24 h with either: (1) 5o
C; (2) 25oC (control); (3) 10 µM ABA at 25oC; or (4) 10 µM methyl jasmonate (MeJa) at 25o
C. The later two treatments were
previously found that could induce chilling-resistance in TN1 seedlings. Total RNA were isolated from the roots of seedlings and used for differential display. Clones of 362 cDNA were re-amplified and isolated from 600 differentially expressed cDNA fragments. Sequences of 153 cDNA clones had significant similarities to the sequences known in the GenBank. Sixty-one of these clones, which could be separated to 29 groups, showed high homology to the sequences with known functions, i.e.,
rab16D, heat shock protein, SalT, and
Zn-induced protein.
clones could be regulated by ABA, MeJA or chilling treatments. Among these 58
differentially expressed cDNA clones, 28 cDNA clones could be enhanced by ABA or chilling using northern blot analysis. In addition, 6 clones of the 28 clones showed differential expressions under stresses including chilling, osmotic, salt, heat shock, and ABA or MeJa treatments. By testing their expressions in 11 cultivars, which have different chilling tolerance and different chilling-induced ABA levels, six cDNA clones showed positively correlated with chilling tolerance. Some of these cDNA clones showed much higher transcript levels after exposure to chilling stress in TN 1 seedlings that pre-treated with ABA.
Therefore, these clones might be involved in the ABA-mediated acclimation of chilling resistance in TN 1 seedlings. In contrast, transcript levels of other cDNA clones were up-regulated in seedlings without ABA pre-treatment, and their expressions seemed not to be related to endogenous ABA levels.
Keywor ds:: plant hormone, differential
display, gene expression, rice, chilling tolerance, abscisic acid.
二、計畫緣由與目的 稻是世界上最重要的糧食作物之一, 1997年全世界稻穀總產量達57326.3萬公 噸,僅次於小麥、玉米,居所有栽培作物 生產的第三位(FAO, 1998)。其中亞洲栽培 的面積佔總栽培面積的90 %以上,稻米為 亞洲及台灣多數人賴以為生的主食。 稻自種子萌芽至開花成熟的過程 中,受到各種環境因子所影響,其中溫度 為一重要環境因子之一,亦為影響作物生 產的重要因素。當水稻生育期間遭受極端 溫度(過高或過低的溫度)時,會破壞其生 理代謝、延緩其生長,嚴重者可導致植株 死亡,因此,溫度對於水稻產量及品質的 影響,受到相當的重視。 植物透過基因的表現的改變,來調節 參與低溫適應複雜生理生化變化 (Thomashow, 1994)。在多數情形下,低溫 適應的過程有許多基因參與(Thomashow, 1990)。一般而言,低溫耐性的遺傳係由量 的性狀所控制(Sasaki, 1997)。分子生物學 上,水稻(Aguan et al., 1991)已由基因表現 的觀點探討植物對低溫反應的機制與 ABA 調控之關係,並已獲得多個與低溫、ABA 有關的 cDNA (Wolfraim and Dhindsa, 1993)。
水稻幼苗耐冷性遺傳的研究中發現, 溫帶日本型的水稻幼苗較熱帶日本型及印 度型水稻抗低溫(Mackill and Lei, 1997)。 Binh and Oono (1992)以水稻品種日本晴 ( 稻)為材料,選殖到對低溫具特殊表現的 clones。Yoshida and Kato (1994)以 稻 Hayayuki 為材料,選殖到 6 個cor genes,
並對 22 個耐冷程度不同的品種進行篩選, 其中有四個基因在乾旱、ABA 及鹽處理下 亦會表現,另二個基因僅在低溫處理下有 表現,顯示這二個基因和水稻的耐冷性有 密切相關。Choi et al. (1997)也以差異表現 法研究水稻低溫逆境,找到四個有差異表 現的 cDNA 片段,其中有一個分子量大約 在 200 bp 左右的 cDNA,以北方墨點雜交 分析試驗,証實的確為低溫所誘導的基 因。Aguan et al. (1991) 以水稻懸浮細胞為 材料,所選殖的三個基因中,ABA 僅稍微 增加 lip5 和 lip19 的表現,而 lip9 則完全不 受 ABA 所影響。
ESTs (expression sequence tags)是對 cDNA 片段的部分序列進行定序,來建立 大量表現基因庫的快速方法(Bouchez and Hoffe, 1998)。經由大量 ESTs 的建立 ,可提 供預測基因產物會在何時及那個部位表 現,進而瞭解基因生物功能的重要線索 (Bouchez and Hoffe, 1998)。Adams et al.
(1991)最早以人體腦組織為材料,隨機篩選 大量的 cDNA clones 加以定序,來進行基 因組結構的分析。隨後這種利用隨機篩選 大量的 cDNA clones 加以定序,來進行基 因組結構的分析的方法,很快的被應用在 高等植物上。Uchimiya et al. (1992)即利用 相同方法對水稻懸浮細胞進行篩選及基因 比對的研究;隨後 Sasaki et al. (1994), Umeda et al. (1994)及 Liu et al. (1995)也曾
進行類似的研究。Yamamoto and Sasaki (1997)並將所有水稻大量 ESTs 定序的結果 做一整理,然而至今並未有學者針對水稻 特定器官,經低溫逆境處理後,來探討低 溫逆境下水稻基因表現的關係。 本研究差異表現法選殖與低溫耐性相 關的 cDNA 以瞭解水稻對低溫的反應機 制,進而來探討水稻如何在低溫下調節其 生理反應與基因表現以增強適應能力,以 提供作物耐冷性改良及選評育種材料之基 礎。 三、結果與討論 前 幾 年 本 研 究 室 以 差 異 表 現 法 (differential display)( Liang and Pardee 1992; Liang et al., 1992),並配合銀染的技術來選 殖 與 水 稻 幼 苗 低 溫 耐 性 表 現 相 關 的 cDNAs。以低溫敏感的印度型水稻台中在 來 1 號(TN 1)及低溫抗性的日本型台農 67 號(TNG 67)幼苗,經過(1)5℃;(2)25℃(對 照組);(3) 10 µM ABA,25℃;或(4)10µM methyl jasmonate (MeJa),25℃;等四項, 各 24 小時處理後,取其根部之 total RNA 供進行差異表現法試驗。結果由 600 個差 異表現片段中,成功再擴增出 362 個差異 表現之 cDNA 片段,經基因庫比對結果, 發現有 153 個(42.3 %) cDNA 與已知基因有 極高的相似性;其中 61 個 cDNA 分別與
rab16D 、 heat shock protein 、 SalT 、
Zn-induced protein… … 等 29 個已知功能基因 有極高的相似性。
另外以 RNA slot blot 分析結果發現, 有 58 個 cDNA clones 在低溫或 ABA 處理 下有極高的表現。再利用北方墨點分析法 (northern-blot analysis)自 58 個差異表現的 片段中,選出 28 個在低溫、ABA 及其它 逆境處理下的表現與對照組有明顯差異的 cDNA clones (表 1)。其中有部分 cDNA 分 別在低溫、ABA、MeJa、缺水、鹽害及熱 休克(heat shock)處理下的水稻根部或地上 部具專一性表現(圖 1)。以 11 個分別具有 不同低溫耐性及 ABA 內生量的水稻品 種,來測試這 28 個 cDNA clones 的表現, 發現有 6 個基因在根部或地上部的表現量 與電解質滲漏量呈顯著負相關,顯示這 6 個基因與低溫耐性相關(圖 2)。 低溫逆境下 ABA 前處理可導致部分 低溫耐性基因在低溫敏感的水稻台中在 來 1 號幼苗中大量表現,顯示這些基因可 能參與水稻幼苗受 ABA 調節所產生的低 溫耐性。另外,部分基因在未經 ABA 前 處理的水稻幼苗中大量表現,顯然它們的 表現不受 ABA 的影響。 四、計畫成果自評 本研究內容與原計畫相符,目前已順 利完成本年度計畫之預期目標,並獲得362 個有差異表現的cDNA,經基因庫比對結 果,發現其中有部分cDNA與已知逆境所誘 導的基因有極高的相似性,目前大部分 cDNA均已完成序列登的工作。 此外,利用北方墨點分析法選出 28 個 在低溫、ABA 及其它逆境處理下的表現與 對照組有明顯差異的 cDNA clones。其中有 部分 cDNA 分別在低溫、ABA、MeJa、缺 水、鹽害及熱休克(heat shock)處理下的水 稻根部或地上部具專一性表現。以 11 個分 別具有不同低溫耐性及 ABA 內生量的水 稻品種,來測試這 28 個 cDNA clones 的表 現,發現有 6 個基因在根部或地上部的表 現量與電解質滲漏量呈顯著負相關,顯示 這 6 個基因與低溫耐性相關。 低溫逆境下ABA前處理可導致部分 低溫耐性基因在低溫敏感的水稻台中在來 1號幼苗中大量表現,顯示這些基因可能參 與水稻幼苗受ABA調節所產生的低溫耐 性。另外,部分基因在未經ABA前處理的 水稻幼苗中大量表現,顯然它們的表現不 受ABA的影響。 目前的結果顯示,本試驗所得到的 cDNA在水稻低溫耐性反應研究上應該具 有相當潛力,也應可以找到一些新的基 因。 本研究室目前正針對上述結果進行整 理,以發表在相關期刊上;基因序列比對 結果中已知功能的cDNA,除可供本研究室 進一步研究其與低溫耐性的相關性外,也 可提供給相關研究人員做進一步的研究。 五、參考文獻
1 Adams, M. D., J. M. Kelley, J. D. Gocayne, M. Dubnick, M. H.
Polymeropoulos, H. Xiao, C. R. Merril, A. Wu, B. Olde, R. F. Moreno, A. R.
Kerlavage, McCombie and J. C. Venter (1991) Complementary DNA sequencing: expressed sequence tags and human genome project. Science 252: 1651-1656 2 Aguan, K., K. Sugawara, N. Suzuki and
T. Kusano (1991) Isolation of genes for low-temperature-induced proteins in rice by a simple subtractive method. Plant Cell Physiol. 32: 1285-1289
3 Binh, L. T. and K. Oono (1992)
Molecular cloning and characterization of genes related to chilling tolerance in rice. Plant Physiol. 99: 1146-1150
4 Bouchez, D. and H. Hofte (1998) Functional genomics in plants. Plant Physiol. 118: 725-732
5 Liu, J., C. Hara, M. Umeda, Y. Zhao, T. W. Okta and H. Uchimiya (1995) Analysis of randomly isolated cDNAs from developing endosperm of rice (Oryza sativa L.): evaluation of expressed
sequence tags and expression levels of mRNAs. Plant Mol. Biol. 29: 685-689 6 Mackill, D. J. and X. Lei (1997) Genetic
variation for traits related to temperature adaptation of rice cultivars. Crop Sci. 37: 1340-1346
7 Sasaki, T., J. Song, Y. Kong-Ban, E. Mastsui, F. Fang, F. Higo, H. Nagasaki, M. Hori, M. Miya, E. Murayama-Kayano, T. Takiguchi, A. Takasuga, T. Niki, K. Ishimaru, H. Ikeda, Y. Yamamoto, Y. Mukai, I. Ohta, N. Miyadera, I. Havukkula and Y. Minobe (1994) Toward cataloguing all rice genes: large-scale sequencing of randomly chosen rice cDNAs from a callus cDNA library. Plant J. 6: 615-624
8 Thomashow, M. F. (1994) Arabidopsis thaliana as a model for studying
mechanisms of plant cold tolerance. In E Meyerowitz, C Somerville, eds,
Arabidopsis. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp 807-834
9 Uchimiya, H., S. Kidou, T. Shimazaki, S. Aotsuka, S. Takamatsu, R. Nishi, H. Hashimoto, Y. Matsubayashi, N. Kidou, M. Umeda and A. Kato (1992) Random sequencing of cDNA libraries reveals a variety of expressed genes in cultured cells of rice (Oryza sativa L.). Plant J. 2:
1005-1009
10 Umeda, M., C. Hara, Y. Matsubayashi, H. H. Li, Q. Liu, F. Tadokoro, S. Aotsuka and H. Uchimiya (1994) Expressed sequence tags from cultured cells of rice (Oryza sativa L.) under stressed
conditions: analysis of transcripts of genes engaged in ATP-generating pathways. Plant Mol. Biol. 25: 469-478 11 Wolfraim, L. A. and Dhindsa R. S. (1993)
Cloning and sequencing of the cDNA for
cas17, a cold acclimation-specific gene of
alfalfa. Plant Physiol. 103: 667-668 12 Yamamoto, K. and T. Sasaki (1997)
Large-scale EST sequencing in rice. Plant Mol. Biol. 35: 135-144
13 Yoshida, H. and A. Kato (1994) Cold-induced accumulation of RNAs and cloning of cDNAs related to chilling injury in rice. Breeding Sci. 44: 361-365
Figure 1. Transcript accumulation in seedlings of 11 rice cultivars, exposed to chilling (5°C) at 85% relative humidity for 24 h. Total RNA (20 µg per lane) was fractionated by denaturing agarose gel electrophoresis, then blotted to nylon
membranes, and hybridized with radiolabeled cDNA probes. Fragments used as probes were Osic1, Osja2, Osjc3, Osjm3, Osjc4, and Osjm5. As a control, the blots were also hybrizied with radioactively labeled probes
encoding the rice 17S and 25S rRNA.
Figure 2. Northern blot analysis of mRNAs from roots and shoots of chilling-tolerant rice (TNG 67) seedlings. HS (heat shock),
incubated at 38°Cfor 1 h; NaCl, incubated with 200 mM NaCl for 24 h; Man, incubated with 0.5 M mannitol for 24 h; MeJa,
incubated with 10 µM methyl jasmonate for 24 h; ABA, incubated with 10 µM ABA for 24 h; LT, 5 °C treatment for 24 h; CK, control. Fragments used as probes were Osic1, Osja2, Osjc3, Osjm3, Osjc4, and Osjm5. Analysis was conducted as described in Figure 1.
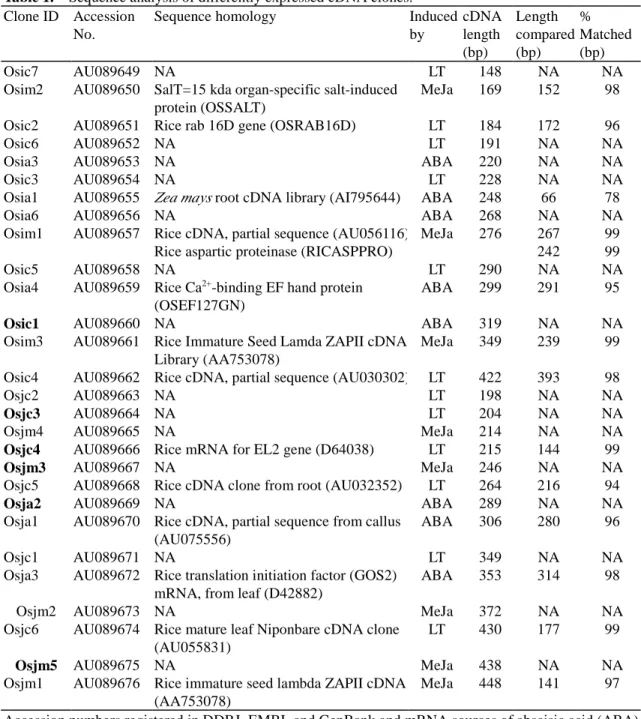
Table 1. Sequence analysis of differently expressed cDNA clones.
Clone ID Accession No.
Sequence homology Induced by cDNA length (bp) Length compared (bp) % Matched (bp) Osic7 AU089649 NA LT 148 NA NA
Osim2 AU089650 SalT=15 kda organ-specific salt-induced protein (OSSALT)
MeJa 169 152 98 Osic2 AU089651 Rice rab 16D gene (OSRAB16D) LT 184 172 96
Osic6 AU089652 NA LT 191 NA NA
Osia3 AU089653 NA ABA 220 NA NA
Osic3 AU089654 NA LT 228 NA NA
Osia1 AU089655 Zea mays root cDNA library (AI795644) ABA 248 66 78
Osia6 AU089656 NA ABA 268 NA NA
Osim1 AU089657 Rice cDNA, partial sequence (AU056116) Rice aspartic proteinase (RICASPPRO)
MeJa 276 267 242
99 99
Osic5 AU089658 NA LT 290 NA NA
Osia4 AU089659 Rice Ca2+-binding EF hand protein
(OSEF127GN)
ABA 299 291 95
Osic1 AU089660 NA ABA 319 NA NA
Osim3 AU089661 Rice Immature Seed Lamda ZAPII cDNA Library (AA753078)
MeJa 349 239 99 Osic4 AU089662 Rice cDNA, partial sequence (AU030302) LT 422 393 98
Osjc2 AU089663 NA LT 198 NA NA
Osjc3 AU089664 NA LT 204 NA NA
Osjm4 AU089665 NA MeJa 214 NA NA
Osjc4 AU089666 Rice mRNA for EL2 gene (D64038) LT 215 144 99
Osjm3 AU089667 NA MeJa 246 NA NA
Osjc5 AU089668 Rice cDNA clone from root (AU032352) LT 264 216 94
Osja2 AU089669 NA ABA 289 NA NA
Osja1 AU089670 Rice cDNA, partial sequence from callus (AU075556)
ABA 306 280 96
Osjc1 AU089671 NA LT 349 NA NA
Osja3 AU089672 Rice translation initiation factor (GOS2) mRNA, from leaf (D42882)
ABA 353 314 98
Osjm2 AU089673 NA MeJa 372 NA NA
Osjc6 AU089674 Rice mature leaf Niponbare cDNA clone (AU055831)
LT 430 177 99
Osjm5 AU089675 NA MeJa 438 NA NA
Osjm1 AU089676 Rice immature seed lambda ZAPII cDNA (AA753078)
MeJa 448 141 97
Accession numbers registered in DDBJ, EMBL and GenBank and mRNA sources of abscisic acid (ABA) treated, methyl jasmonate (MeJa) treated, or controls (CK) are indicated. Homology is given as percent matched and length in base pairs (bp) of nucleotide matched. Sequences showing a similarity score over 200 were considered homologous to the cDNA clones were presented. NA: data are not available.