人體病毒在不同器官中的延遲感染模型
35
0
0
全文
(2)
(3) 摘要 部分病毒會引起人類疾病,雖然針對特定疾病已被大量研發藥物,用以治療部分病 毒所引起的疾病,甚至有疫苗可用以預防疾病之發生,但我們仍應了解確切的治療機制 與效果。因此對病毒感染模型進行研究,將在病毒的治療與疫苗的適時使用上能有所貢 獻。首先藉由基礎的病毒感染模型,提出病毒在不同器官中進行病毒感染及細胞感染的 延遲感染模型,之後我們藉由討論系統的平衡解存在性、局部穩定性分析、全域穩定性 分析來推論解的行為,並以 Maxima 在大部分參數不變時,經由增加某些感染情形測詴 對平衡解的影響,再透過 Matlab 進行數值模擬分析,藉此證實解的行為與我們的推論 是否一致,並希望從上述分析中,更進一步認識病毒的傳染特性。 關鍵字:病毒傳染模型、多器官感染、延遲反應。. i.
(4) Abstract Most viruses can cause human diseases. Although a large number of drugs have been developed for specific diseases to treat diseases caused by some viruses, and even vaccines can be used to prevent diseases, we should understand the mechanism of treatment and the drug effect. Therefore, research on the viral infection model will contribute to the timely treatment of the virus and the timely use of the vaccine. Firstly, based on the concept of viral infection, the delayed model of viral infection and cell infection in different organs was proposed. Then we deduced the behavior of solution by discussing the existence of equilibrium solution, local stability analysis. Maxima was used to explore the effect of equilibrium on certain parameters, and Matlab was used to simulate the viral dynamics confirming whether our theoretical results consist with the behavior of the solution. It helps further understanding the characteristics of viral infection. Keywords:Viral infection、Multiple-organ infection、Intracellular delay。. ii.
(5) 目錄 摘要……………………………………………………………………………………………i. Abstract……………………………………………………………………………………ii 目錄……………………………………………………………………………………………iii 表次……………………………………………………………………………………………iv 圖次……………………………………………………………………………………………v 第一章 前言………………………………………………………………………………… 1 1.1 簡介………………………………………………………………………………………1 1.2 研究動機…………………………………………………………………………………1 第二章 文獻導讀及問題建模……………………………………………………………… 2 2.1 基本數學建模介紹…………………………………………………………………… 2 2.2 多重器官數學建模介紹……………………………………………………………… 3 (1)雙區感染模型……………………………………………………………………… 4 (2)基本再生數………………………………………………………………………… 4 (3)穩定性……………………………………………………………………………… 4 2.3 研究模型的建構……………………………………………………………………… 5 2.4 研究方法……………………………………………………………………………… 6 第三章 研究及分析………………………………………………………………………… 7 3.1 模型平衡點分析……………………………………………………………………… 7 3.1.1 模型生物性質………………………………………………………………………7 定理一……………………………………………………………………………… 7 定理二……………………………………………………………………………… 8 3.1.2 平衡解…………………………………………………………………………… 10 3.1.3 平衡點存在條件………………………………………………………………… 13 定理三……………………………………………………………………………… 15 3.2 反向分歧(Backward bifurcation)…………………………………………………22 第四章 結論………………………………………………………………………………… 27 參考文獻…………………………………………………………………………………… 28. iii.
(6) 表次 表1 表2 表3 表4 表5 表6 表7 表8. 平衡點的存在條件…………………………………………………………………15 系統參數-1…………………………………………………………………………19 系統參數-2…………………………………………………………………………19 系統參數-3…………………………………………………………………………20 系統參數-4…………………………………………………………………………21 系統參數-5…………………………………………………………………………21 系統參數-6…………………………………………………………………………22 系統參數-7…………………………………………………………………………23. iv.
(7) 圖次 圖1 圖2 圖3 圖4 圖5 圖6 圖7 圖8 圖9. 一般的分歧圖……………………………………………………………………………5 向後分歧(Backward bifurcation)的分歧圖…………………………………………5 滿足A < 0且𝐶 ≥ 0對稱軸偏左的情形,病毒數量 V 的函數圖………………………13 滿足A < 0且𝐶 ≥ 0對稱軸偏右的情形,病毒數量 V 的函數圖………………………13 表 2 參數時I1 隨時間的變化關係圖…………………………………………………… 19 表 2 參數時I2 隨時間的變化關係圖…………………………………………………… 19 表 3 參數時I1 隨時間的變化關係圖…………………………………………………… 20 表 3 參數時I2 隨時間的變化關係圖…………………………………………………… 20 表 4 參數時I1 隨時間的變化關係圖…………………………………………………… 20. 圖 10 圖 11 圖 12 圖 13 圖 14 圖 15 圖 16. 表 4 參數時I2 隨時間的變化關係圖…………………………………………………… 20 表 5 參數時I1 隨時間的變化關係圖…………………………………………………… 21 表 5 參數時I2 隨時間的變化關係圖…………………………………………………… 21 表 6 參數時I1 隨時間的變化關係圖…………………………………………………… 21 表 6 參數時I2 隨時間的變化關係圖…………………………………………………… 21 表 7 參數時I1 隨時間的變化關係圖…………………………………………………… 22 表 7 參數時I2 隨時間的變化關係圖…………………………………………………… 22. 圖 17 在表 8 參數下,當β3 = 2 × 10;11 時,V 隨時間變化圖………………………………23 圖 18 在表 8 參數下,當β3 = 2 × 10;10 時,V 隨時間變化圖………………………………24 圖 19 在表 8 參數下,當β3 = 5 × 10;10 時,V 隨時間變化圖……………………………… 24 圖 20 當β3 = β̃2 = 2 × 10;11時,V 隨時間變化圖………………………………………… 25 圖 21 當β3 = β̃2 = 2 × 10;10時,V 隨時間變化圖………………………………………… 25 圖 22 當β3 = β̃2 = 2 × 10;9 時,V 隨時間變化圖……………………………………………26. v.
(8) 第一章 前言 1.1 簡介 病毒有相當多的種類,對人體的影響也各有不同,除了最常見的流感病毒使感 染而引發感冒外,有些病毒還能影響到人的免疫系統就像人類免疫缺陷病毒對人體 的感染,而且大部分病毒感染沒經過妥善治療的話,時常會引發更嚴重的症狀,如: 流感病毒感染沒治療好很容易引起急性肺炎、肝炎病毒感染沒治療好有可能會演變 成肝癌。 病毒主要是由飛沫、血液及體液來進行傳染的,但感染後不一定會立刻發病, 而這感染到發病的期間稱作潛伏期或延遲。在疾病的傳染上,又會因為不同器官或 部位其溫度或酸鹼值不同,進而影響病毒感染的係數。病毒大多經由血液來傳染, 因此除了病灶外其他部位中也會有病毒的存在,例如,有肝炎病毒的患者,病毒除 了在肝細胞中,還會存在於血液裡,因此肝臟移植與輸血都可能使病毒傳播出去。 因此,探討病毒感染過程的延遲效應和多重器官結構對感染的影響是個有趣的 問題。. 1.2 研究動機 相關研究藉由建立數學模型探討病毒的感染,例如,早期 M.A. Nowak et al.(1996)[4]及 M.A. Nowak, R.M. May(2000)[5]建立了 HBV 和 HCV(Hepatitis C Virus,C 型肝炎病毒)傳染的數學模型,之後 J.M. Heffernan, L.M. Wahl(2005)[2] 考慮病毒感染時的消耗率,也就是病毒與細胞連結並把基因輸入細胞複製後,原本 的病毒細胞就剩下不具感染力的空殼,因此微幅修改了模型,使病毒變化量上減掉 消耗率。 Redouane Qesmi et al.(2010)[6]考慮在不同部位的感染情形有所差異,而將 模型再細分成肝臟和血液兩器官中的健康細胞與感染細胞。這是在人體內考慮不同 器官對病毒感染的影響,是我們的主要研究動機。再者,Michael Y. Li · Hongying Shu(2012)[3]的文章考慮了病毒傳染時的延遲反應,也就是醫療上常用的「潛伏期」 一詞,由此引發動機在研究人體多個器官中的病毒動態時,再考慮加入病毒感染健 康細胞、受感染細胞感染健康細胞以及病毒從受感染細胞迸發而出的延遲反應分別 有不同的延遲時間,期望發現延遲反應如何影響病毒的存在與滅絕,進而發現週期 解的存在性。因此,我將致力於同時考慮多重器官結構和感染的延遲效應對整體系 統的感染影響。. 1.
(9) 第二章 文獻導讀及問題建模 2.1 基本數學建模介紹 M.A. Nowak et al.(1996)[4]及 M.A. Nowak, R.M. May(2000)[5] 建立 HBV 和 HCV 的基礎感染動態,如下: dT dt dI dt dV dt. = 𝑎 − βTV − bI = βTV − 𝜇𝐼 = 𝛾𝐼 − 𝜇̃V,. (2.1). 其中未受感染的 T 細胞(T)的產生率為常數𝑎,b為其自然死亡率;當健康 T 細胞與 HBV 或 HCV 接觸後,受到病毒感染其感染率為βxv;受感染的 T 細胞(I)的產生率βxv 為健 康 T 細胞受感染轉化而成,𝜇為其死亡率; HBV 或 HCV(V) 的產生率𝛾y 為在受感染細胞 中進行複製然後迸裂而出,因此一個細胞會產出大量病毒,所以參數𝜇 ≪ 𝛾;𝜇̃代表了病 毒的死亡率。 在病毒的感染上,當一個病毒要侵入健康細胞時,他只能透過表面蛋白配對成功後, 經由打開的通路將自己的核酸輸入細胞,在裡面進行複製與轉錄的工作,而原始的那個 病毒就會變成空殼並被代謝掉,因此 J.M. Heffernan, L.M. Wahl(2005)[2]就提出病毒 感染有消耗率的存在,並以−βxv表示,所以將模型微調如下: dT dt dI dt dV dt. = 𝑎 − βTV − bT = βTV − 𝜇I = 𝛾I − 𝜇̃V − βTV,. (2.2). 這模型與(2.1)的差別只在於病毒單位時間的變化量上,考慮了感染時所需的消耗率。 在常見情況中,當健康細胞與病毒接觸到變成受感染細胞,當中必須經歷一段化學 反應時間,而非瞬間就可達成。在醫療上,我們稱這種情形為此種病毒的潛伏期,因此 Michael Y. Li · Hongying Shu (2012)[3]就提出這種含有病毒感染延遲期間的模型, 如下: dT dt dI dt dV dt. T(𝑡). = 𝑎 − bT(𝑡) + 𝑘T(𝑡)(1 − T. 𝑚𝑎𝑥. ) − βT(t)V(t). = βe;𝑠𝜏 T(t − τ)V(t − τ) − 𝜇I(𝑡) = 𝛾I(𝑡) − 𝜇̃V(t), 2. (2.3).
(10) 在未受感染的 T 細胞(T)中,除了線性的出生率與死亡率分別為𝑎及b外,並且健康 T 細 胞中會行有絲分裂,其成長量因為考慮到環境養育量,所以採取 logistic 模式呈現為 T(𝑡). 健康 T 細胞的環境乘載率=𝑘T(𝑡)(1 − T. 𝑚𝑎𝑥. );. 受感染 T 細胞(I)中,出生率為τ時間前病毒與健康 T 細胞的感染而生,其值原本應為 βT(t − τ)V(t − τ),但轉變的過程中有些細胞的死亡使得轉換效率使其變為 受感染 T 細胞的生成率=βe;𝑠𝜏 T(t − τ)V(t − τ); 受感染細胞的死亡率為𝑑2;最後關於病毒細胞(V),其產生率𝛾I 為在受感染細胞中進行 複製而後迸裂產出,因此一個細胞會產出大量病毒,因此參數𝜇 ≪ 𝛾,而𝜇̃代表了病毒的 死亡率。. 2.2 多重器官數學建模介紹 Redouane Qesmi et al.(2010)[6]認為在不同部位的感染情形也會有所差異(如: 不同部位的 PH 值、血液的流通量都會有些差異),因而將模型再細分成肝臟和血液兩器 官中的健康細胞與感染細胞如下: dT1 dt dI1. = β1 T1 V − 𝜇1 I1. dt dV dt. = 𝛾1 I1 + 𝛾2 I2 − 𝜇̃V − β1 T1 V − β2 T2 𝑉. dT2 dt dI2 dt. = 𝑎1 − β1 T1 V − b1 T1. = 𝑎2 − β2 T2 v − b2 T2 = β2 T2 v − 𝜇2 I2 ,. (2.4). 其中未受感染的肝臟細胞(T1 ) 的產生率為常數𝑎1 ,b1 為其自然死亡率;當健康肝臟細 胞與 HBV 或 HCV 接觸後,受到病毒感染其感染率為β1 T1 V;受感染的肝臟細胞(I1 )的產生 率β1 T1 V為健康肝臟細胞受感染轉化而成,𝜇1 為其死亡率;未受感染的血液細胞(T2 ) 的 產生率為常數𝑎2,b2 為其自然死亡率;當健康血液細胞與 HBV 或 HCV 接觸後,受到病毒 感染其感染率為β2 T2 𝑉;受感染的血液細胞(I2 )的產生率β2 T2 𝑉為健康血液細胞受感染轉 化而成,𝜇2 為其死亡率;HBV 或 HCV(V) 從肝臟及血液的產生率分別為𝛾1 I1及𝛾2 I2 ;𝜇̃代 表了病毒的死亡率;最後,扣除病毒傳染肝臟細胞及血液細胞的代謝率分別是β1 T1 V和 β2 T2 𝑉。 本篇提出的模型與 Redouane Qesmi et al.(2010)[6]提出的雙區域 HBV 或 HCV 感染 模型有很多相關之處,因此對文獻[4]進行觀察與介紹,其中包含平衡點與系統的關係, 並且分析了基本再生數對病毒傳播的影響。. 3.
(11) (1)雙區感染模型 在該篇論文中,其模型分為 HBV 對肝臟細胞的感染及對血液細胞的感染的系統。在 這系統中,我們共使用五條常微分方程來表示,並且我們假設個區域內的細胞其出生、 死亡、受感染及病毒產生率皆不一定相同。 dT1 dt dI1. = β1 T1 V − 𝜇1 I1. dt dV dt. = 𝛾1 I1 + 𝛾2 I2 − 𝜇̃V − β1 T1 V − β2 T2 𝑉. dT2 dt dI2 dt. = 𝑎1 − β1 T1 V − b1 T1. = 𝑎2 − β2 T2 v − b2 T2 = β2 T2 v − 𝜇2 I2 ,. (2) 基本再生數 基本再生數(R 0 )是由分析病毒細胞及受感染細胞的生成率與死亡率的搭配組合,藉 此來說明該疾病的傳染趨勢。當R 0 >1 時,病毒細胞及受感染細胞的生成率大於死亡率, 因此疾病將會開始擴散;反之,當R 0 <1 時,病毒細胞及受感染細胞的生成率小於死亡 率,因此疾病將會開始消失。 此感染模型(2.4)的基本再生數如下: R0 =. 由受感染肝細胞產生病毒的生成率 受感染肝細胞的死亡率 +. 由受感染血液細胞產生病毒的生成率. ×. 受感染血液細胞的死亡率. ×. 肝細胞受感染率 病毒的死亡率. 血液細胞受感染率 病毒的死亡率. = 肝細胞內病毒轉換效率 × 肝細胞內病毒傳染率 +血液細胞內病毒轉換效率 × 血液細胞內病毒傳染率 𝛾1. =𝜇 × 1. 𝑎 β1 1. b1 𝑎 𝑎 ̃ :β2 2 :β1 1 𝜇 b2 b1. 𝛾2. +𝜇 × 2. 𝑎 β2 2 b2. 𝑎 𝑎 ̃ :β2 2 :β1 1 𝜇 b2. (3)穩定性 文獻[6]的穩定性可由下面分岐圖來觀察:. 4. b1. 。. (2.5).
(12) 圖 1:一般的分歧圖. 圖 2;向後分歧(Backward bifurcation) 的分歧圖. (a)圖 1 在R 0 < 1時,只存在無病平衡點其穩定性為穩定的;在R 0 ≥ 1時,存在無病平 衡點跟一個共生平衡點,此時共生平衡點為穩定的,無病平衡點為非穩定的。 (b)圖 2 是滿足 Backward bifurcation 的情形,其中二次函數頂點的R 0 值為R1 ,在 R 0 < R1 時,只存在無病平衡點其穩定性為穩定的;在R1 < R 0 < 1時,存在無病平衡點 跟兩個共生平衡點,此時上共生平衡點和無病平衡點為穩定的,下共生平衡點為非穩定 的;在R 0 ≥ 1時,存在無病平衡點跟一個共生平衡點,此時共生平衡點為穩定的,無病 平衡點為非穩定的。. 2.3 研究模型的建構 在參考文獻[6]以及[3]中分別對病毒感染的兩個影響因素做考慮,前者考慮不同部 位的受感染率,後者思考病毒傳染存在的潛伏期。 病毒的感染主要是有兩種途徑,第一種是病毒對人體細胞的直接感染,第二種為受 感染的人體細胞將病毒傳染給其他體內健康的細胞使其受到感染,這兩種不同的感染途 徑會使細胞受感染率會有些不同。 因此本研究將考慮同時結合這兩種感染的因素以及同時結合這兩種感染的途徑,但 其中假設器官 1 為病毒直接接觸感染的身體細胞,其感染途徑為病毒對細胞的感染,還 有器官 2 為主要受病毒影響的目標細胞,其感染途徑為受感染細胞對健康細胞間感染, 從中找尋該模型動態行為。 不同於文獻[6] 以及[3]只考慮病毒對細胞的感染,我們在兩種器官中各別考慮病 毒對健康細胞感染以及受感染細胞對健康細胞感染,建模如下:. 5.
(13) dT1 dt dI1. = β1 𝑒 ;𝑎1 𝜏1 𝑇1 (𝑡 − 𝜏1 )𝑉(𝑡 − 𝜏1 ) − 𝜇1 I1. dt dT2 dt dI2 dt dV dt. = 𝑎 − bT1 − β1 𝑇1 𝑉. = 𝑎 − bT2 − β2 𝑇2 𝐼2 = β2 𝑒 ;𝑎2 𝜏2 𝑇2 (𝑡 − 𝜏2 )𝐼2 (𝑡 − 𝜏2 ) − 𝜇2 I2. = 𝛾1 𝑒 ;𝑎3 𝜏3 I1 (𝑡 − 𝜏3 ) + 𝛾2 𝑒 ;𝑎3 𝜏3 I2 (𝑡 − 𝜏3 ) − 𝜇̃V − β̃𝑇1 𝑉,. T1 :器官 1 之中健康細胞的數量. I1 :器官 1 之中受感染細胞的數量. T2 :器官 2 之中健康細胞的數量 𝑉:病毒細胞的數量 𝑏: 健康細胞的死亡率 𝑒 ;𝑎𝑖 𝜏𝑖 :受感染細胞經𝜏𝑖 期間的存活率 𝛾𝑖 : 受感染細胞的病毒產生率 β̃:病毒感染時的病毒消耗率. I2 :器官 2 之中受感染細胞的數量 𝑎:健康細胞的生成率 β𝑖 :病毒感染率 𝜇𝑖 :受感染細胞的死亡率 𝜇̃: 病毒的死亡率. (2.6). 其中我們考慮了,病毒在感染細胞後,受感染的細胞對健康的細胞也具有感染力,所以 添加了β2 𝑇2 𝐼2 ,並且病毒的傳染常常存在潛伏期,所以我們分別假設器官 1 上、器官 2 上以及病毒上的感染延遲時間分別為𝜏1 、𝜏2 和𝜏3 。 由於存在感染延遲的時間𝜏𝑖,所以細胞從t − 𝜏𝑖 到 t 的作用時間,其中就存在這段期 間的存活率𝑒 ;𝑎𝑖 𝜏𝑖 ,其中𝑎𝑖 為受感染但還未能產生病毒時的細胞死亡率。 以肝炎病毒的感染過程為例,我們可以知道,病毒會先找到與其外部醣蛋白能相接 收的受體進行連接,藉此進入宿主體內進行轉譯及複製等流程,由此可知病毒感染時的 消耗率β̃近似於病毒感染健康身體細胞的感染率β1 。當病毒進入細胞後會藉由宿主的細 胞進行複製病毒,最後會產非常大量的病毒,因此可知受感染細胞的病毒產生率𝛾𝑖 會遠 大於病毒感染率β𝑖 。最後我們看到健康細胞與受感染細胞的死亡率,因為受感染細胞受 病毒的影響,因此健康細胞的死亡率𝑏會小於受感染細胞的死亡率𝜇𝑖 。. 2.4 研究方法 分析(2.6)平衡點的存在性並使用 maxima 以及 matlab,藉由系統在平衡點的線性化 (linearization)分析系統的局部穩定性(local stability)。再透過 matlab 的 dde23 程式對(2.6)進行數值模擬,藉由觀察延遲對系統的影響,進而應用到更多相關的模型 當中。 其中,以比較𝜏1、𝜏2 和𝜏3 對系統局部穩定性的影響為重點。因為不同病毒感染階段、 不同部位以及不同感染方式的延遲反應時間會有差異,因此藉由調整延遲反應時間𝜏1 、 𝜏2 和𝜏3 ,觀察系統局部穩定性是否會受到延遲反應的變化影響。. 6.
(14) 第三章 研究及分析 3.1 模型平衡點分析 3.1.1 模型生物性質 dT1. = 𝑎 − bT1 − β1 𝑇1 𝑉. dt dI1. = β1 𝑒 ;𝑎1 𝜏1 𝑇1 (𝑡 − 𝜏1 )𝑉(𝑡 − 𝜏1 ) − 𝜇1 I1. dt dT2. = 𝑎 − bT2 − β2 𝑇2 𝐼2. dt dI2 dt dV dt. = β2 𝑒 ;𝑎2 𝜏2 𝑇2 (𝑡 − 𝜏2 )𝐼2 (𝑡 − 𝜏2 ) − 𝜇2 I2 = 𝛾1 𝑒 ;𝑎3 𝜏3 I1 (𝑡 − 𝜏3 ) + 𝛾2 𝑒 ;𝑎3 𝜏3 I2 (𝑡 − 𝜏3 ) − 𝜇̃V − β̃𝑇1 𝑉. 。. 定理一: (2.6)的模型是屬於正常生物系統,其中各種細胞的數量皆為非負,因此 T1 、I1 、T2 、I2 、V ≥ 0。 證明: dT1 (𝑡) dt. = 𝑎 − bT1 (t) − β1 𝑇1 (𝑡)𝑉(t) = 𝑎 − (b + β1 𝑉(t))T1 (t),. 則兩邊積分後可得, 𝑡. 𝑡. 𝑡. T1 (𝑡) = 𝑒 ∫0 ;(b:β1 𝑣(𝜃))𝑑𝜃 T1 (0) + ∫0 𝑒 ∫𝜃 ;(b:β1 𝑣(𝜂))𝑑𝜂 𝑎𝑑𝜃 ≥ 0。 同理, dT2 (𝑡) dt. = 𝑎 − bT2 (t) − β2 𝑇2 (𝑡)I2 (t) = 𝑎 − (b + β2 I2 (t))T2 (t),. 則兩邊積分後可得, 𝑡. 𝑡. 𝑡. T2 (𝑡) = 𝑒 ∫0 ;(b:β2 I2(𝜃))𝑑𝜃 T2 (0) + ∫0 𝑒 ∫𝜃 ;(b:β2 I2 (𝜂))𝑑𝜂 𝑎𝑑𝜃 ≥ 0。 我們接下來要證明在t ≥ 0時I1 (𝑡)、I2 (𝑡)、V(t) ≥ 0,因此我們假設存在一個t1 ≥ 0,使 得0 ≤ t ≤ t1 時min*I1 (t)、I2 (t)、V(t)+ ≥ 0,而且會發生下列其中一種情況: dI1 (t1 :𝜉). (1). I1 (t1 + 𝜉) < 0且. (2). I2 (t1 + 𝜉) < 0且. dt dI2 (t1 :𝜉) dt. ≤ 0, ≤ 0, 7.
(15) 𝑉(t1 + 𝜉) < 0且. (3). d𝑉(t1 :𝜉) dt. ≤ 0,. 其中min*𝜏1 、𝜏2 、𝜏3 + > 𝜉 > 0。 dI1 (t1 :𝜉). (1)首先取I1 (t1 + 𝜉) < 0且. ≤ 0,所以−𝜇1 I1 (t1 + 𝜉) > 0,又因為0 < t1 + 𝜉 − 𝜏1 <. dt. t1 ,所以β1 𝑒 ;𝑎1𝜏1 𝑇1 (t1 + 𝜉 − 𝜏1 )𝑉(t1 + 𝜉 − 𝜏1 ) ≥ 0,因此 dI1 (t1 :𝜉) dt. = β1 𝑒 ;𝑎1 𝜏1 𝑇1 (t1 + 𝜉 − 𝜏1 )𝑉(t1 + 𝜉 − 𝜏1 ) − 𝜇1 I1 (t1 + 𝜉) > 0, dI1 (t1 :𝜉). 由此可觀察到假設的. dt. dI1 (t1 :𝜉). ≤ 0與實際的. dI2 (t1 :𝜉). (2)同理取 I2 (t1 + 𝜉) < 0且. dt. dt. > 0互相矛盾。. ≤ 0,所以−𝜇2 I2 (t1 + 𝜉) > 0,又因為0 < t1 + 𝜉 −. 𝜏2 < t1 ,所以β2 𝑒 ;𝑎2 𝜏2 𝑇2 (t1 + 𝜉 − 𝜏2 )I2 (t1 + 𝜉 − 𝜏2 ) ≥ 0,因此 dI2 (t1 :𝜉) dt. = β2 𝑒 ;𝑎2 𝜏2 𝑇2 (t1 + 𝜉 − 𝜏2 )I2 (t1 + 𝜉 − 𝜏2 ) − 𝜇2 I2 (t1 + 𝜉) > 0, dI2 (t1 :𝜉). 由此可觀察到假設的. dt. dI2 (t1 :𝜉). ≤ 0與實際的. d𝑉(t1 :𝜉). (3)同理取𝑉(t1 + 𝜉) < 0且. dt. dt. > 0互相矛盾。. ≤ 0,所以−𝜇̃𝑉(t1 + 𝜉) > 0以及−β̃𝑇1 (t1 + 𝜉)𝑉(t1 +. 𝜉) > 0,又因為0 < t1 + 𝜉 − 𝜏3 < t1,所以 𝛾1 𝑒 ;𝑎3𝜏3 I1 (t1 + 𝜉 − 𝜏3 ) + 𝛾2 𝑒 ;𝑎3𝜏3 I2 (t1 + 𝜉 − 𝜏3 ) ≥ 0,因此 dV(t1 :𝜉) dt. = 𝛾1 𝑒 ;𝑎3 𝜏3 I1 (t1 + 𝜉 − 𝜏3 ) + 𝛾2 𝑒 ;𝑎3 𝜏3 I2 (t1 + 𝜉 − 𝜏3 ) −𝜇̃𝑉(t1 + 𝜉) − β̃𝑇1 (t1 + 𝜉)𝑉(t1 + 𝜉) > 0, d𝑉(t1 :𝜉). 由此可觀察到假設的. dt. d𝑉(t1 :𝜉). ≤ 0與實際的. dt. > 0互相矛盾。. 由此可證明在𝑡 ≥ 0時T1 (𝑡)、I1 (𝑡)、T2 (𝑡)、I2 (𝑡)、V(𝑡) ≥ 0。. □. 定理二: (2.6)的模型為有界的,因此只要其落在區域Ω內的情形皆會符合這模型, 𝑎. 𝑎. 𝑎. 𝑎. 1. 2. Ω = *(T1 、I1 、T2 、I2 、V) ∈ ℝ5: :T1 ≤ b ,T2 ≤ b ,T1 + I1 ≤ 𝑘 ,T2 + I2 ≤ 𝑘 ,V ≤ 𝑉𝑐 +,. 其中 1 𝑎𝛾1 𝑒 −𝑎3 𝜏3. 𝑉𝑐 = 𝜇̃ (. 𝑘1. +. 𝑎𝛾2 𝑒 −𝑎3 𝜏3 𝑘2. )。. 證明: 從(2.6)的第一式中,因為T1 、V ≥ 0且β1為身體細胞感染率非負,所以−β1 𝑇1 𝑉 ≤ 0, 因此 8.
(16) dT1. ≤ 𝑎 − bT1 ,. dt. 𝑎. limt→∞ 𝑠𝑢𝑝(T1 )(t) ≤ b。 (2.6)的第三式同理,因為T2 、I2 ≥ 0且β2 為肝臟細胞感染率非負,所以−β2 𝑇2 I2 ≤ 0, 因此 dT2 dt. ≤ 𝑎 − bT2 , 𝑎. limt→∞ 𝑠𝑢𝑝(T2 )(t) ≤ b。 令 𝑘1 = min*𝑏、𝜇1 + > 0, 𝑘2 = min*𝑏、𝜇2 + > 0, 由(2.6)的第一式加上以t = t + 𝜏1 帶入的第二式可得: d,T1 (t):I1 (t:𝜏1 )dt. = 𝑎 − bT1 (𝑡) − 𝜇1 I1 (t + 𝜏1 ) + β1 𝑇1 (𝑡)𝑉(𝑡)(𝑒 ;𝑎1 𝜏1 − 1) ,. 因為T1 、𝑉 ≥ 0且0 < 𝑒 ;𝑎1 𝜏1 ≤ 1,所以β1 𝑇1 (𝑡)𝑉(𝑡)(𝑒 ;𝑎1 𝜏1 − 1) ≤ 0,使得 d,T1 (t):I1 (t:𝜏1 )dt. ≤ 𝑎 − bT1 (𝑡) − 𝜇1 I1 (t + 𝜏1 ) ≤ 𝑎 − 𝑘1 (T1 (t) + I1 (t + 𝜏1 )),. 因此 𝑎. limt→∞ 𝑠𝑢𝑝,T1 (t) + I1 (t + 𝜏1 )- ≤ 𝑘 。. (3.1). 1. 同理(2.6)的第三式加上以t = t + 𝜏2 帶入的第四式可得: d,T2 (t):I2 (t:𝜏2 )dt. = 𝑎 − bT2 (𝑡) − 𝜇2 I2 (t + 𝜏2 ) + β2 𝑇2 (𝑡)I2 (𝑡)(𝑒 ;𝑎2𝜏2 − 1) ,. 因為T2 、I2 ≥ 0且0 < 𝑒 ;𝑎2 𝜏2 ≤ 1,所以β2 𝑇2 (𝑡)I2 (𝑡)(𝑒 ;𝑎2 𝜏2 − 1) ≤ 0,使得 d,T2 (t):I2 (t:𝜏2 )dt. ≤ 𝑎 − bT2 (𝑡) − 𝜇2 I2 (t + 𝜏2 ) ≤ 𝑎 − 𝑘2 (T2 (t) + I2 (t + 𝜏2 )),. 因此 𝑎. limt→∞ 𝑠𝑢𝑝,T2 (t) + I2 (t + 𝜏2 )- ≤ 𝑘 。. (3.2). 2. 𝑎. 𝑎. 1. 2. 由(3.1)和(3.2)可知0 ≤ I1 (t) ≤ 𝑘 以及0 ≤ I2 (t) ≤ 𝑘 ,因此將此結果代入(2.6)的第五 式中,可得如下: dV dt. ≤. 𝑎𝛾1 𝑒 −𝑎3 𝜏3 𝑘1. +. 𝑎𝛾2 𝑒 −𝑎3 𝜏3 𝑘2. − 𝜇̃V − β̃𝑇1 𝑉, 9.
(17) 又因為𝑇1 、𝑇2 、𝑉 ≥ 0,所以−β̃𝑇1 𝑉、 − β̃𝑇2 𝑉 ≤ 0,因此 dV dt. ≤. 𝑎𝛾1 𝑒 −𝑎3 𝜏3 𝑘1. +. 𝑎𝛾2 𝑒 −𝑎3 𝜏3 𝑘2. − 𝜇̃V,. 1 𝑎𝛾1 𝑒 −𝑎3 𝜏3. limt→∞ 𝑠𝑢𝑝,V(t)- ≤ 𝜇̃ (. 𝑘1. +. 𝑎𝛾2 𝑒 −𝑎3 𝜏3 𝑘2. )。. 因此可找到一區域Ω,使得(2.6)中解的向量場,在這有界的區域Ω中時,皆不指向外, 區域如下: 𝑎. 𝑎. 𝑎. 𝑎. 1. 2. Ω = *(T1 、I1 、T2 、I2 、V) ∈ ℝ5: :T1 ≤ b ,T2 ≤ b ,T1 + I1 ≤ 𝑘 ,T2 + I2 ≤ 𝑘 ,V ≤ 𝑉𝑐 +,. 其中 1 𝑎𝛾1 𝑒 −𝑎3 𝜏3. 𝑉𝑐 = ̃ ( 𝜇. 𝑘1. +. 𝑎𝛾2 𝑒 −𝑎3 𝜏3 𝑘2. )。. □. 3.1.2 平衡解 為了求上面感染模型(2.6)的平衡解,我們將解下列代數方程式: 0 = 𝑎 − bT1 − β1 𝑇1 𝑉 0 = β1 𝑒 ;𝑎1 𝜏1 𝑇1 𝑉 − 𝜇1 I1 0 = 𝑎 − bT2 − β2 𝑇2 𝐼2 0 = β2 𝑒 ;𝑎2 𝜏2 𝑇2 𝐼2 − 𝜇2 I2 0 = 𝛾1 𝑒 ;𝑎3 𝜏3 I1 + 𝛾2 𝑒 ;𝑎3 𝜏3 I2 − 𝜇̃V − β̃𝑇1 𝑉。. (3.3) (3.4) (3.5) (3.6) (3.7). 我們可由(3.3)得到: 𝑎. T1 = b:β. 𝑎. 1𝑉. ≤ b,. (3.8). 因為生物數量為非負數。 由(3.3)乘上𝑒 ;𝑎1 𝜏1 與(3.4)相加可得: 0 = 𝑒 ;𝑎1 𝜏1 (𝑎 − bT1 ) − 𝜇1 I1 , I1 =. 𝑒 −𝑎1 𝜏1 (𝑎;bT1 ) 𝜇1. ≥ 0,. (3.9). 𝑎. 如果𝑉 ≥ 0時T1 ≤ 𝑏,因此將T1 代入到(3.9)可得: I1 =. 𝑒 −𝑎1 𝜏1 (𝑎;b. 𝑎 ) b+β1 𝑉. 𝜇1. 。. (3.10). 我們可由(3.5)得到: 𝑎. T2 = b:β. 2 I2. 𝑎. ≤ b,. 因為生活的環境中,不會存在負數個病毒或受感染肝細胞。 由(3.5)乘上𝑒 ;𝑎2 𝜏2 與(3.6)相加可得: 0 = 𝑒 ;𝑎2 𝜏2 (𝑎 − bT2 ) − 𝜇I2 , 10. (3.11).
(18) I2 =. 𝑒 −𝑎2 𝜏2 (𝑎;bT2 ) 𝜇2. ≥ 0,. (3.12). 𝑎. 如果I2 ≥ 0時T2 ≤ 𝑏,因此將(3.11)代入到(3.12)可得: I2 =. 𝑒 −𝑎2 𝜏2 (𝑎;b. 𝑎 ) b+β2 I2. ,. 𝜇2. (3.13). 將(3.13)整理後可得: I2 (I2 +. 𝜇2 b;𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. ) = 0,. (3.14). 因此, I2 = 0 或. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β2. 𝑎. (T2 ,I2 ) = .b ,0/ 或(𝑎β. 𝜇2 𝑎 −𝑎 𝜏 2𝑒 2 2. ,. ,. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. ) 。. 𝑎. (I)當(T2 ,I2 ) = .b ,0/時,把(3.8)與(3.10)代入(3.7)得到: 0= 0=. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 (𝑎;b. 𝑎 ) b+β1 𝑉. 𝑎 − 𝜇̃V − β̃ b:β. 𝜇1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 𝑎β1 𝜇1. 𝑉 =0或𝑉 =𝑉. (0). 1𝑉. 𝑉,. 𝑉 − b𝜇̃V − β1 𝜇̃𝑉 2 − β̃𝑎𝑉, 𝛾 𝑒−𝑎3 𝜏3 𝑒−𝑎1 𝜏1 𝑎β1 ̃𝑎) ̃ ;β ( 1 ;b𝜇 𝜇1. =. ̃) (β1 𝜇. 。. 將 V 代回(3.8)與(3.10)可得: 𝑉 = 0時, 𝑎. T1 = b:β I1 =. = b,. 𝑒 −𝑎1 𝜏1 (𝑎;b. 𝑎 ) b+β1 ×0. 𝜇1. 𝑎. (0). 𝑎. 1 ×0. = 0,. 𝑎. (0). 得到一個平衡點E0 = (b ,0, b ,0,0),又因為E0 中病毒及受感染細胞的量皆為 0, (0). 因此可稱E0 為一無病平衡點。 當𝑉 = 𝑉. (0). =. 𝛾 𝑒−𝑎3 𝜏3 𝑒−𝑎1 𝜏1 𝑎β1 ̃𝑎) ̃ ;β ( 1 ;b𝜇 𝜇1. 時,我們可得. ̃) (β1 𝜇 (0). T1 = 𝑇1. = b:β. 𝑎 1 ×𝑉. (0). 11.
(19) 𝑎. =. ̃ 𝑎𝜇. =. (0). I1 = 𝐼1. 𝛾 𝑒−𝑎3 𝜏3 𝑒−𝑎1 𝜏1 𝑎β1 ̃ ( 1 ;β𝑎) 𝜇1. =. (0). (0). 𝑒 −𝑎1 𝜏1 𝜇1. 𝑎𝑒 −𝑎1 𝜏1. =. 得一平衡點𝐸1. 𝛾 𝑒−𝑎3 𝜏3 𝑒−𝑎1 𝜏1 𝑎β1 ̃ 𝑎) ̃ −β ( 1 −b𝜇 𝜇1 b:β1 × ̃) (β1 𝜇. 𝜇1. (𝑎 − b. (1 −. ,. 𝑎. 𝛾 𝑒−𝑎3 𝜏3 𝑒−𝑎1 𝜏1 𝑎β1 ̃ 𝑎) ̃ −β ( 1 −b𝜇 𝜇1 b:β1 × ̃) (β1 𝜇. ̃ b𝜇. ). ),. 𝛾 𝑒−𝑎3 𝜏3 𝑒−𝑎1 𝜏1 𝑎β1 ̃ ( 1 ;β𝑎) 𝜇1. 𝑎. (0). = (𝑇1 ,𝐼1 , ,0,𝑉 (0) )。 b. (II)當−𝜇2 b + 𝑎β2 𝑒 ;𝑎2 𝜏2 > 0時,( T2 ,I2 ) = ( 𝑇2∗ ,𝐼2∗ ) = (β. 𝜇2. 2. , 𝑒 −𝑎2 𝜏2. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. )時,. 把(3.8)與(3.10)代入(3.7)得到: 0= 0=. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 .𝑎;b. 𝑎 / b+β1 𝑉. 𝜇1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 𝑎β1 𝜇1. + 𝛾2 𝑒 ;𝑎3 𝜏3. 𝑉 + b𝛾2 𝑒 ;𝑎3 𝜏3. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2. +β1 𝛾2 𝑒 ;𝑎3 𝜏3. 𝜇2 β 2. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. 𝑎 − 𝜇̃V − β̃ b:β. 1𝑉. 𝑉,. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. 𝑉 − b𝜇̃V − β1 𝜇̃V 2 − β̃𝑎𝑉。. (3.15). 上式(3.15)為病毒數量 V 的二次方程式,我們將(3.15)轉換為函數來觀察其圖形的特性 如下: 𝑓(𝑉) =. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 𝑎β1 𝜇1. +β1 𝛾2 𝑒 ;𝑎3 𝜏3. 𝑉 + b𝛾2 𝑒 ;𝑎3 𝜏3. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. 𝑉 − b𝜇̃V − β1 𝜇̃V 2 − β̃𝑎𝑉. = 𝐴𝑉 2 + 𝐵𝑉 + 𝐶 。. (3.16). 其中假設 A = −β1 𝜇̃, B=. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 𝑎β1 𝜇1. C = b𝛾2 𝑒 ;𝑎3 𝜏3. + β1 𝛾2 𝑒 ;𝑎3 𝜏3. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. 。. 觀察其二次項係數可發現 A = −β1 𝜇̃ < 0, 因此可知其為開口向下的二次方程式。 12. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. − b𝜇̃ − β̃𝑎,.
(20) 然後當𝑉 = 0時,因為. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. 𝑓(0) = 𝐶 = b𝛾2 𝑒 ;𝑎3 𝜏3. = I2 ≥ 0,所以 ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. ≥ 0,. 因此可知(3.13)的圖形呈現如下兩種情形: 4. f(v). 2. 0. -2. -4. -4. -2. 0. 2. 4. v. 圖 3:滿足A < 0且𝐶 ≥ 0對稱軸偏左的情形,V 只有一組正解 4. f(v). 2. 0. -2. -4. -4. -2. 0. 2. 4. v. 圖 4:滿足A < 0且𝐶 ≥ 0對稱軸偏右的情形,V 只有一組正解 由上圖可知不管是哪種情形,系統中的病毒數量 V 都恰只有一正實根V ∗,因此我們可得 一個共生平衡點如下: (2). E1 = (𝑇1∗ ,𝐼1∗ ,𝑇2∗ ,𝐼2∗ ,V ∗ ) = (𝑇1∗ ,𝐼1∗ , β. 𝜇2 −𝑎 𝜏 2𝑒 2 2. ,. ;𝜇2 b:𝑎β2 𝑒 −𝑎2 𝜏2 𝜇2 β 2. 3.1.3 平衡點存在條件分析 13. ,V ∗ )。.
(21) (0). (2). 我們可以從上面推論𝐸1 以及 E1 時,可觀察到以下幾個條件: (0). (1)推論𝐸1 時,為了滿足細胞數量為非負的,因此 (0). 𝐼1 (0). 則可知𝑇1. 𝑒 −𝑎1 𝜏1. =. 𝜇1. (0). .𝑎 − b𝑇1 / ≥ 0,. 𝑎. ≤ 𝑏才會成立,所以 (0). 𝑇1. = b:β. 𝑎 1 ×𝑉. 𝑎. (0). ≤ 𝑏,. 則可知𝑉 (0) ≥ 0才會成立,所以 𝑉. (0). =. 𝛾 𝑒−𝑎3 𝜏3 𝑒−𝑎1 𝜏1 𝑎β1 ̃𝑎) ̃ ;β ( 1 ;b𝜇 𝜇1. ̃) (β1 𝜇. ≥ 0,. 則可整理得到下方關係式, 𝑏. ≤ 𝑎. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. ̃ β. − 𝜇̃,. (0). 當滿足此條件時平衡點𝐸1 才可能存在。 (2). (2) 推論E1 時,為了要讓各細胞數量為非負的,因此可發現 −𝜇2 b + 𝑎β2 𝑒 ;𝑎2 𝜏2 > 0, 此平衡點的各細胞數量就會為非負的,因此把上是做整理後可得到 𝑏. < 𝑎. β2 𝑒 −𝑎2 𝜏2 𝜇2. ,. (2). 當滿足此條件時平衡點E1 才可能存在。 β2 𝑒 −𝑎2 𝜏2. 令. 𝜇2. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1. = 𝜉1 、. 條件. ̃ 𝜇1 𝜇. ̃ β. − 𝜇̃ = 𝜉2 ,則可將上述狀況整理如下表:. 平衡點. 條件. 𝜉1 > 𝜉2. 𝜉2 > 𝜉1. (0). (0). E0 存在且穩定 𝑏 > 𝜉1 𝑎. 平衡點. E0 存在且穩定 𝑏 > 𝜉2 𝑎. (0). E1 不存在. (0). E1 不存在. (2). E1 不存在. (2). (0). E0 存在且不穩定. E1 不存在. (0). E0 存在且不穩定 14.
(22) 𝑏 < 𝜉2 𝑎. 𝑏 < 𝜉1 𝑎. (0). E1 存在 (2). E1 存在. (2). (0). E0 存在且不穩定. E1 存在. (0). E0 存在且部分穩定 𝜉1 >. 𝑏 > 𝜉2 𝑎. (0). E1 存在. (0). E1 不存在. 𝜉2 >. 𝑏 > 𝜉1 𝑎. (2). (0). E1 存在 (2). E1 存在. E1 不存在 表 1:平衡點的存在條件. 由上述的條件我們可以得到以下的定理: 定理三: 𝑏. (0). (0). (1) 𝜉1 > 𝜉2 且𝑎 > 𝜉1時,平衡點只存在E0 ,且E0 為穩定的平衡點。 𝑏. (0). (0). (2). (2) 𝜉1 > 𝜉2 且𝑎 < 𝜉2時,存在平衡點E0 、E1 、E1 。 𝑏. (0). (2). (3) 𝜉1 > 𝜉2 且𝜉1 > 𝑎 > 𝜉2 時,存在平衡點E0 、E1 。 𝑏. (0). (0). (4) 𝜉2 > 𝜉1 且𝑎 > 𝜉2時,平衡點只存在E0 ,且E0 在這條件下為穩定的平衡點。 𝑏. (0). (0). (2). (5) 𝜉2 > 𝜉1 且𝑎 < 𝜉1時,存在平衡點E0 、E1 、E1 。 𝑏. (0). (0). (6) 𝜉2 > 𝜉1 且𝜉2 > 𝑎 > 𝜉1 時,存在平衡點E0 、E1 。 證明: ̃2 ,Ĩ2 ,V ̃) 為 3.1 中求得的任一平衡點 令 E=(T̃1 ,Ĩ1 ,T 原模型為非線性但要求特徵方程需要其為線性,因此我們就將其線性化成以下樣貌: dT1 dt dI1. ̃ + β1 𝑒 ;𝑎1 𝜏1 T̃1 𝑉(𝑡 − 𝜏1 ) − 𝜇1 I1 = β1 𝑒 ;𝑎1 𝜏1 𝑇1 (𝑡 − 𝜏1 )V. dt dT2 dt dI2 dt dV dt. ̃ − β1 T̃1 𝑉 = −bT1 − β1 𝑇1 V. ̃ − β1 ̃ = −bT2 − β1 𝑇2 V T2 𝑉 − β2 𝑇2 Ĩ2 − β2 ̃ T2 𝐼2 = β2 𝑒 ;𝑎2 𝜏2 𝑇2 (𝑡 − 𝜏2 )Ĩ2 + β2 𝑒 ;𝑎2 𝜏2 ̃ T2 𝐼2 (𝑡 − 𝜏2 ) − 𝜇2 I2. ̃ − β̃T̃1 𝑉。 = 𝛾1 𝑒 ;𝑎3 𝜏3 I1 (𝑡 − 𝜏3 ) + 𝛾2 𝑒 ;𝑎3 𝜏3 I2 (𝑡 − 𝜏3 ) − 𝜇̃V − β̃𝑇1 V 15. (3.17).
(23) 為了求的系統的特徵函數,因此我們將先求出系統的 Jacobian 矩陣如下 𝜕T1. 𝜕T1. 𝜕T1. 𝜕T1. 𝜕T1. 𝜕T1 𝜕T2. 𝜕 I1 𝜕T2. 𝜕T2 𝜕T2. 𝜕 I2 𝜕T2. 𝜕𝑉 𝜕T2. 𝜕T1 𝜕𝑉. 𝜕 I1 𝜕𝑉. 𝜕T2 𝜕𝑉. 𝜕 I2 𝜕𝑉. 𝜕𝑉 𝜕𝑉. 𝜕T1 𝜕 I1. J(E) =. 𝜕 I1 𝜕 I1. 𝜕T1 𝜕 I2. 𝜕T2 𝜕 I1. 𝜕 I1 𝜕 I2. (𝜕T1. 𝜕T2 𝜕 I2. 𝜕 I1. 𝜕T2. 𝜕 I2 𝜕 I1. 𝜕 I2 𝜕 I2. 𝜕 I2. ̃ −b − β1 V β1 𝑒. −(𝑎1 +𝜆)𝜏1 ̃. V. =. 0. (. 𝜕𝑉 𝜕 I1. 𝜕𝑉 𝜕 I2. 𝜕𝑉 ). −β1 T̃1 −(𝑎1 +𝜆)𝜏1. 0. 0. 0. −𝜇1. 0. 0. −b − β2 Ĩ2. −β2 T̃2. 0. 0. 0. ̃ −β̃ V. 𝛾1 𝑒−(𝑎3 +𝜆)𝜏3. β2 𝑒−(𝑎2 +𝜆)𝜏2 Ĩ2. β2 𝑒. 0. −(𝑎2 +𝜆)𝜏2. β1 𝑒. T̃1. 0. T̃2 − 𝜇2. 0 −𝜇̃ − β̃ T̃1 ). 𝛾2 𝑒−(𝑎3+𝜆)𝜏3. 藉由上面推導出的 Jacobian 矩陣可得到本系統在點 E 的特徵方程式如下: ̃−𝜆 −b − β1 V 0 ;(𝑎1 :𝜆)𝜏1 ̃ V −𝜇1 − 𝜆 |β1 𝑒 0 0 | 0 0 ;(𝑎3 :𝜆)𝜏3 ̃ ̃ −βV 𝛾1 𝑒. 0 0 −b − β2 Ĩ2 − 𝜆 β2 𝑒 ;(𝑎2:𝜆)𝜏2 Ĩ2 0. 0 0 −β2 ̃ T2 ;(𝑎2 :𝜆)𝜏2 ̃ β2 𝑒 T2 − 𝜇2 − 𝜆 𝛾2 𝑒 ;(𝑎3:𝜆)𝜏3. (0). (0). ̃. β1 𝑒. −β1 T1 ;(𝑎1 :𝜆)𝜏1. T̃1 |. =0 0 | 0 −𝜇̃ − β̃T̃1 − 𝜆. 𝑎. 𝑎. 以下我們針對平衡點E0 做局部穩定性分析,當E = E0 = (b ,0, b ,0,0)時,. |. |. −b − 𝜆. 0. 0. 0. −𝜇1 − 𝜆. 0. 0. 0. −b − 𝜆. 0. 0. 0. | 0. 𝛾1 𝑒. ;(𝑎3 :𝜆)𝜏3. 0. 0 0 −. 𝑎β2 b. 𝑎β2 𝑒 ;(𝑎2 :𝜆)𝜏2 − 𝜇2 − 𝜆 b 𝛾2 𝑒. 16. ;(𝑎3 :𝜆)𝜏3. −. 𝑎β1 b. |. 0. |=0. 𝑎β1 𝑒 ;(𝑎1 :𝜆)𝜏1 b. 0 | 𝑎β̃ −𝜇̃ − −𝜆 b.
(24) |. |. −b − 𝜆. 0. 0. 0. 0. −𝜇1 − 𝜆. 0. 0. 0. −b − 𝜆. 0. 0. 0. −. 0. 𝛾1 𝑒. 𝑎β2 b. 𝑎β2 𝑒 ;(𝑎2 :𝜆)𝜏2 − 𝜇2 − 𝜆 b. 0. 𝛾2 𝑒. ;(𝑎3 :𝜆)𝜏3. −𝜇1 − 𝜆 | = (−b − 𝜆)(−b − 𝜆) |. 0 𝛾1 𝑒. b. ;(𝑎3 :𝜆)𝜏3. = (−b − 𝜆)(−b − 𝜆) *(−𝜇1 − 𝜆) (. |. 0. |. 0 | 𝑎β̃ −𝜇̃ − −𝜆 b 𝑎β1 𝑒 −(𝑎1 +𝜆)𝜏1. 0 𝑎β2 𝑒 −(𝑎2 +𝜆)𝜏2. 𝑎β1 b. 𝑎β1 𝑒 ;(𝑎1 :𝜆)𝜏1 b. 0. | ;(𝑎3 :𝜆)𝜏3. −. b. − 𝜇2 − 𝜆. 𝛾2 𝑒 ;(𝑎3 :𝜆)𝜏3. | |. 0 −𝜇̃ −. ̃ 𝑎β b. −𝜆. 𝑎β2 𝑒 ;(𝑎2 :𝜆)𝜏2 𝑎β̃ − 𝜇2 − 𝜆) (−𝜇̃ − − 𝜆) b b. 𝑎β2 𝑒 ;(𝑎2 :𝜆)𝜏2 𝑎β1 𝑒 ;(𝑎1 :𝜆)𝜏1 − (𝛾1 𝑒 ;(𝑎3 :𝜆)𝜏3 )( − 𝜇2 − 𝜆)( )+ b b 𝑎β2 𝑒 −(𝑎2 +𝜆)𝜏2. = (−b − 𝜆)(−b − 𝜆) . 0(−𝜇1 − 𝜆) .−𝜇̃ −. b. ̃ 𝑎β b. 𝑎β1 𝑒 −(𝑎1 +𝜆)𝜏1. − 𝜆/ − (𝛾1 𝑒 ;(𝑎3 :𝜆)𝜏3 )(. 𝑎β2 𝑒 −(𝑎2 +𝜆)𝜏2. = (−b − 𝜆)(−b − 𝜆) . 0𝜆2 + .𝜇1 + 𝜇̃ +. b. ̃ 𝑎β. / 𝜆 + (𝜇1 𝜇̃ + b 𝑎β2 𝑒 −𝑎2 𝜏2. = (−b − 𝜆)(−b − 𝜆) . 0𝜆2 + .𝜇1 + 𝜇̃ +. − 𝜇2 − 𝜆/. b. b. )1. − 𝜇2 − 𝜆/ ̃ 𝜇1 𝑎β b. −. 𝑎β1 𝛾1 𝑒 −(𝑎3 +𝜆)𝜏3 𝑒 −(𝑎1 +𝜆)𝜏1 b. )1. 𝑒 ;𝜆𝜏2 − 𝜇2 − 𝜆/. ̃ 𝑎β. / 𝜆 + (𝜇1 𝜇̃ + b. ̃ 𝜇1 𝑎β b. )−. 𝑎β1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 b. 𝑒 ;(𝜏3 :𝜏1 )𝜆 1. =0 。 (−b − 𝜆) = 0中可以很明顯地看到當𝜆 = −𝑏 < 0時,整個特徵函數就會為 0。接著觀察 𝑎β2 𝑒 −𝑎2 𝜏2 b. 𝑒 ;𝜆𝜏2 − 𝜇2 − 𝜆,我們可透過文獻[1]提到的判別法,在𝜆 − 𝐴1 − 𝐵1 𝑒 ;𝜏𝜆 = 0中,. 當𝐵1 > 0且𝐴1 + 𝐵1 < 0,則𝜆的實部為負的,因此 𝑎β2 𝑒 −𝑎2 𝜏2 b. > 0,. 且 17.
(25) 𝑎β2 𝑒 −𝑎2 𝜏2. − 𝜇2 < 0,. b 𝑎β2 𝑒 −𝑎2 𝜏2. < 𝜇2 ,. b β2 𝑒 −𝑎2 𝜏2 𝜇2 𝑎β2 𝑒 −𝑎2 𝜏2. 可以得到. b. b. < 𝑎, β2 𝑒 −𝑎2 𝜏2. 𝑒 ;𝜆𝜏2 − 𝜇2 − 𝜆 = 0在. 𝜇2. b. < 𝑎時,其𝜆的實部為負的。. 再來觀察 𝜆2 + .𝜇1 + 𝜇̃ +. ̃ 𝑎β. ̃ 𝜇1 𝑎β. / 𝜆 + .𝜇1 𝜇̃ + b. b. 𝑎β1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1. /−. b. 𝑒 ;(𝜏3 :𝜏1 )𝜆 = 0,. 我們可透過文獻[1]提到的判別法,在𝜆2 + 𝐴2 𝜆 + 𝐵2 − 𝐶2 𝑒 ;𝜏𝜆 = 0中,當2𝐵2 − 𝐴2 2 < 0且 𝐵2 2 > 𝐶2 2 時,其𝜆的實部為非正的,因此 2 .𝜇1 𝜇̃ + 2𝜇1 𝜇̃ +. ̃ 𝜇1 𝑎β b. ̃ 𝜇1 2𝑎β b. / − .𝜇1 + 𝜇̃ +. ̃ 2 𝑎β b. / < 0,. ̃ 𝑎β. < 𝜇1 2 + 𝜇̃ 2 + ( b )2 + 2𝜇1 𝜇̃ + ̃ 𝑎β. 0 < 𝜇1 2 + 𝜇̃2 + ( b )2 +. ̃𝜇 ̃ 2𝑎β b. ̃ 𝜇1 2𝑎β b. +. ̃𝜇 ̃ 2𝑎β b. ,. ,. 且 (𝜇1 𝜇̃ +. ̃ 𝜇1 𝑎β )2 b. ̃ β. 𝑏. (𝑎 + 𝜇̃)2 > ( 𝑏. > 𝑎 可以得到𝜆2 + .𝜇1 + 𝜇̃ + 𝑏. > 𝑎. β1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 ̃ 𝜇1 𝜇. > (−. β1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 2 ) , ̃ 𝜇1 𝜇. β1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1. ̃ 𝑎β. ̃ 𝜇1 𝜇. / 𝜆 + (𝜇1 𝜇̃ + b. 𝑎β1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 2 ) , b. ̃ 𝜇1 𝑎β b. )−. ̃ β. − 𝜇̃,. 𝑎β1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 b. 𝑒 ;(𝜏3 :𝜏1)𝜆 = 0在. ̃ β. − 𝜇̃時,其𝜆的實部為非正的。 𝑏. 由上面的推導可知其呼應我們上面的定理三,在𝑎 >. β1 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 ̃ 𝜇1 𝜇. ̃ β. β2 𝑒 −𝑎2 𝜏2. − 𝜇̃且. 𝜇2. b. < 𝑎時,. (0). 平衡點E0 為局部穩定的平衡點。 此時我們想觀察平衡點的穩定情形,因此透過數值模擬,對兩種情形中的三個區段 進行觀察,如下: (I). 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. ̃ β. − 𝜇̃ <. β2 𝑒 −𝑎2 𝜏2 𝜇2. 18.
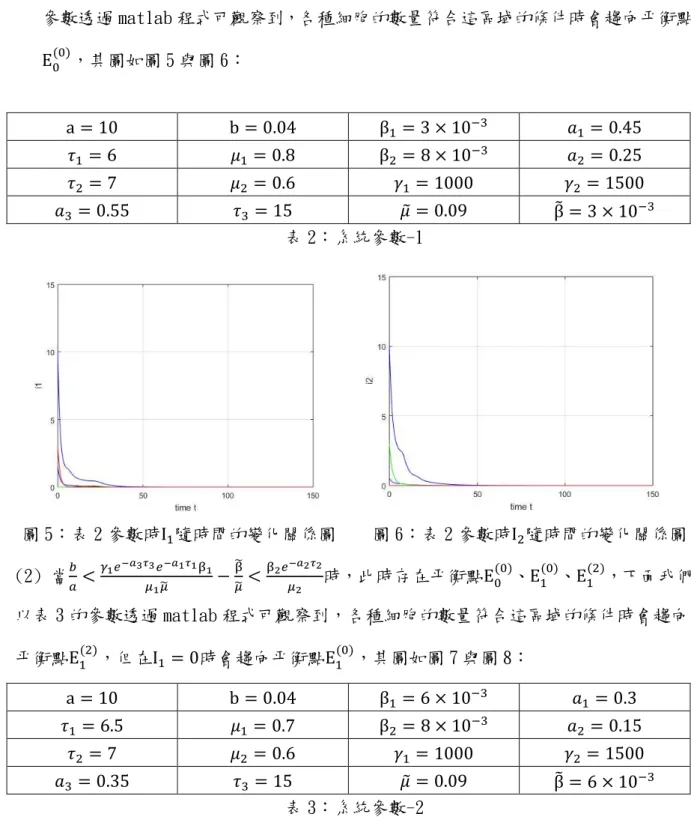
(26) 𝑏. (1)當𝑎 >. β2 𝑒 −𝑎2 𝜏2 𝜇2. >. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. ̃ β. (0). − 𝜇̃時,此時只存在平衡點E0 ,下面我們以表 2 的. 參數透過 matlab 程式可觀察到,各種細胞的數量符合這區域的條件時會趨向平衡點 (0). E0 ,其圖如圖 5 與圖 6: b = 0.04. β1 = 3 × 10;3. 𝑎1 = 0.45. 𝜏1 = 6. 𝜇1 = 0.8. β2 = 8 × 10. ;3. 𝑎2 = 0.25. 𝜏2 = 7. 𝜇2 = 0.6. 𝛾1 = 1000. a = 10. 𝑎3 = 0.55. 𝑏. β̃ = 3 × 10;3. 𝜏3 = 15 𝜇̃ = 0.09 表 2:系統參數-1. 圖 5:表 2 參數時I1 隨時間的變化關係圖 (2) 當𝑎 <. 𝛾2 = 1500. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. ̃ β. − 𝜇̃ <. β2 𝑒 −𝑎2 𝜏2 𝜇2. 圖 6:表 2 參數時I2 隨時間的變化關係圖 (0). (0). (2). 時,此時存在平衡點E0 、E1 、E1 ,下面我們. 以表 3 的參數透過 matlab 程式可觀察到,各種細胞的數量符合這區域的條件時會趨向 (2). (0). 平衡點E1 ,但在I1 = 0時會趨向平衡點E1 ,其圖如圖 7 與圖 8: b = 0.04. β1 = 6 × 10;3. 𝜏1 = 6.5. 𝜇1 = 0.7. β2 = 8 × 10. ;3. 𝜏2 = 7. 𝜇2 = 0.6. 𝛾1 = 1000. a = 10. 𝑎3 = 0.35. 𝜏3 = 15 𝜇̃ = 0.09 表 3:系統參數-2. 19. 𝑎1 = 0.3 𝑎2 = 0.15 𝛾2 = 1500 β̃ = 6 × 10;3.
(27) 圖 7:表 3 參數時I1 隨時間的變化關係圖 β2 𝑒 −𝑎2 𝜏2. (3) 當. 𝜇2. 𝑏. >𝑎≥. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. 圖 8:表 3 參數時I2 隨時間的變化關係圖. ̃ β. (0). (2). − 𝜇̃時,此時存在平衡點E0 、E1 ,下面我們以表 4. 的參數透過 matlab 程式可觀察到,各種細胞的數量符合這區域的條件時會趨向平衡點 (2). (0). E1 ,但在I1 = 0時會趨向平衡點E0 ,其圖如圖 9 與圖 10: b = 0.04. β1 = 3 × 10;3. 𝑎1 = 0.45. 𝜏1 = 6. 𝜇1 = 0.8. β2 = 8 × 10. ;3. 𝑎2 = 0.15. 𝜏2 = 5. 𝜇2 = 0.6. 𝛾1 = 1000. a = 10. 𝑎3 = 0.55. β2 𝑒 −𝑎2 𝜏2. 𝑏. (1)當𝑎 ≥. 𝜇2. β̃ = 3 × 10;3. 𝜏3 = 15 𝜇̃ = 0.09 表 4:系統參數-3. 圖 9:表 4 參數時I1 隨時間的變化關係圖 (II) (. 𝛾2 = 1500. <. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. ̃ β. − 𝜇̃ >. 圖 10:表 4 參數時I2 隨時間的變化關係圖. ̃ β. − 𝜇̃). β2 𝑒 −𝑎2 𝜏2 𝜇2. (0). 時,此時只存在平衡點E0 ,下面我們以表 5 的 (0). 參數透過 matlab 程式可觀察到,各種細胞的數量符合這區域的條件時會趨向平衡點E0 , 20.
(28) 其圖如圖 11 與圖 12: a = 10. b = 0.04. β1 = 5 × 10;3. 𝑎1 = 0.1. 𝜏1 = 10. 𝜇1 = 0.3. β2 = 6 × 10;3. 𝑎2 = 0.25. 𝜏2 = 7. 𝜇2 = 0.6. 𝛾1 = 1000. 𝛾2 = 1500. 𝑎3 = 0.65. 圖 11:表 5 參數時I1 隨時間的變化關係圖 𝑏. (2) 當𝑎 <. β2 𝑒 −𝑎2 𝜏2 𝜇2. β̃ = 5 × 10;3. 𝜏3 = 15 𝜇̃ = 0.09 表 5:系統參數-4. <. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. 圖 12:表 5 參數時I2 隨時間的變化關係圖. ̃ β. (0). (0). (2). − 𝜇̃時,此時存在平衡點E0 、E1 、E1 ,下面我們. 以表 6 的參數透過 matlab 程式可觀察到,各種細胞的數量符合這區域的條件時會趨向 (2). (0). 平衡點E1 ,但在I1 = 0時會趨向平衡點E1 ,其圖如圖 13 與圖 14: a = 10. b = 0.04. β1 = 5 × 10;3. 𝑎1 = 0.1. 𝜏1 = 10. 𝜇1 = 0.3. β2 = 6 × 10;3. 𝑎2 = 0.15. 𝜏2 = 5. 𝜇2 = 0.6. 𝛾1 = 1000. 𝛾2 = 1500. 𝑎3 = 0.45. 𝜏3 = 15 𝜇̃ = 0.09 表 6:系統參數-5. 圖 13:表 6 參數時I1 隨時間的變化關係圖. β̃ = 5 × 10;3. 圖 14:表 6 參數時I2 隨時間的變化關係圖 21.
(29) β2 𝑒 −𝑎2 𝜏2. (2)當. 𝜇2. 𝑏. <𝑎≤. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. ̃ β. (0). (0). − 𝜇̃時,此時存在平衡點E0 、E1 ,下面我們以表 7. 的參數透過 matlab 程式可觀察到,各種細胞的數量符合這區域的條件時會趨向平衡 (0). 點E1 ,其圖如圖 15 與圖 16: b = 0.04. β1 = 5 × 10;3. 𝜏1 = 10. 𝜇1 = 0.3. β2 = 6 × 10. ;3. 𝜏2 = 7. 𝜇2 = 0.6. 𝛾1 = 1000. a = 10. 𝑎3 = 0.45. 𝜏3 = 15 𝜇̃ = 0.09 表 7:系統參數-6. 𝑎1 = 0.1 𝑎2 = 0.25 𝛾2 = 1500 β̃ = 5 × 10;3. 圖 15:表 7 參數時I1 隨時間的變化關係圖 圖 16:表 7 參數時I2 隨時間的變化關係圖 由上面各個數值模擬的圖,我們可看到各個起始值對平衡點的趨近情形,並可發現 其與定理三的推論結果是一致的。. 3.2 反向分歧(Backward bifurcation) 此小節中,我們將對模型進行更多感染面向的探討,藉此希望能找到如文獻[6]中 Backward bifurcation 的情境,我們對感染模型(2.6)中器官 1 新增了細胞跟細胞間的 感染途徑、器官 2 新增了病毒對細胞的感染途徑,以及對不同器官的參數做更詳細的區 分,其模型如下: dT1 dt dI1. = β1 𝑒 ;𝑎1 𝜏1 𝑇1 (𝑡 − 𝜏1 )𝑉(𝑡 − 𝜏1 ) − 𝜇1 I1 + β4 𝑒 ;𝑎1 𝜏1 𝑇1 (𝑡 − 𝜏1 )I1 (𝑡 − 𝜏1 ). dt dT2 dt dI2 dt dV dt. = a1 − b1 T1 − β1 𝑇1 𝑉 − β4 𝑇1 I1. = a2 − b2 T2 − β2 𝑇2 𝐼2 − β3 𝑇2 𝑉 = β2 𝑒 ;𝑎2 𝜏2 𝑇2 (𝑡 − 𝜏2 )𝐼2 (𝑡 − 𝜏2 ) − 𝜇2 I2 + β3 𝑒 ;𝑎2 𝜏2 𝑇2 (𝑡 − 𝜏2 )𝑉(𝑡 − 𝜏2 ). = 𝛾1 𝑒 ;𝑎3 𝜏3 I1 (𝑡 − 𝜏3 ) + 𝛾2 𝑒 ;𝑎3 𝜏3 I2 (𝑡 − 𝜏3 ) − 𝜇̃V − β̃1 𝑇1 𝑉 − β̃2 𝑇2 𝑉, 22. (3.18).
(30) T1 :器官 1 之中健康身體細胞的數量. I1 :器官 1 之中受感染身體細胞的數量. T2 :器官 2 之中健康肝臟細胞的數量 𝑉:HBV 細胞的數量 𝑏𝑖 : 健康細胞的死亡率 𝑒 ;𝑎𝑖 𝜏𝑖 :受感染細胞經𝜏𝑖 期間的存活率 𝛾𝑖 : 受感染細胞的病毒產生率 β̃𝑖 :病毒感染時的病毒消耗率. I2 :器官 2 之中受感染肝臟細胞的數量 a𝑖 :健康細胞的生成率 β𝑖 :病毒感染率 𝜇𝑖 :受感染細胞的死亡率 𝜇̃: 病毒的死亡率. 此處我們透過數值模擬觀察器官 2 中,透過病毒對細胞的感染率β3 的改變,觀察各種病 毒的起始數量隨時間的趨近情形,可得以下 3 種情境: a1 = 105. a2 = 108. b1 = 0.03. b2 = 0.003. β1 = 2.001 × 10;10 β̃2 = 2 × 10;10. β2 = 2.2 × 10;15. β4 = 2 × 10;15. β̃1 = 2.001 × 10;10. 𝜇1 = 1.000000001. 𝜇2 = 1. 𝜇̃ = 1. 𝑎1 = 0.1. 𝑎2 = 0.15. 𝑎3 = 0.2. 𝛾1 = 2000. 𝛾2 = 0.94. 𝜏1 = 10;20 𝜏2 = 1.5 × 10;20 表 8:系統參數-7. 𝜏3 = 1.3 × 10;20. (1) 當β3 = 2 × 10;11 時,. 圖 17:在表 8 參數下,當β3 = 2 × 10;11 時,V 隨時間變化圖 此圖可觀察到,當器官 2 的病毒感染率很小時,疾病的感染會漸漸消退,使得感染 模型的病毒數量趨近到零,也就是會趨近於無病平衡點。 此情境就像是一種沒什麼感染力的病毒,不管空間中有再多病毒,其對器官的影響 幾乎微乎其微。. 23.
(31) (2) 當β3 = 2 × 10;10 時,. 圖 18:在表 8 參數下,當β3 = 2 × 10;10 時,V 隨時間變化圖 此圖可觀察到,當器官 2 的病毒感染率略為增加但還沒過大時,此時有兩種趨近情 形,其一為感染模型的病毒起始數量夠多時,使得感染模型的病毒數量趨近到一非零的 數值,也就是會趨近於共生平衡點,另一種為感染模型的病毒起始數量不多時,使得感 染模型的病毒數量趨近到零,也就是會趨近於無病平衡點。 此情境為感染率再增強一些的病毒,此時當環境中病毒量多於某數值時,對器官就 會產生影響,但病毒數量不夠時,對器官的影響就不顯著。 (3) 當β3 = 5 × 10;10 時,. 圖 19:在表 8 參數下,當β3 = 5 × 10;10 時,V 隨時間變化圖 此圖可觀察到,當器官 2 的病毒感染率很大時,疾病變會開始蔓延,使得感染模型 的病毒數量趨近到一非零的數值,也就是會趨近於共生平衡點。 此情境就像是一種感染力很強的病毒,不管空間中有多少病毒,其對器官都能產生 影響。 由此三種情境中,我們可觀察到器官 2 的病毒感染率β3 對此模型的影響,隨著β3 的 24.
(32) 增加,從全部趨近到無病平衡點變成有部份趨近到共存平衡點再變成全部趨近到共存平 衡點,因此可發現如同文獻[6]的反向分歧(Backward bifurcation)現象存在。 由於我們前面有提到病毒感染率會跟病毒感染時的消耗率可能會相似,因此以下我 們將會對上面數值模擬再加上此條件觀察,此結果如下: (1) 當β3 = β̃2 = 2 × 10;11時,. 圖 20:當β3 = β̃2 = 2 × 10;11時,V 隨時間變化圖 (2) 當β3 = β̃2 = 2 × 10;10時,. 圖 21:當β3 = β̃2 = 2 × 10;10時,V 隨時間變化圖. 25.
(33) (3) 當β3 = β̃2 = 2 × 10;9 時,. 圖 22:當β3 = β̃2 = 2 × 10;9 時,V 隨時間變化圖 以上三種情境中,我們可觀察到器官 2 的病毒感染率β3 以及感染器官 2 時的病毒消 耗率β̃2 對此模型的影響,其結果與上一個結果有些不同,可能原因為病毒感染消耗率對 病毒數量有一定程度的影響,隨著β3 與β̃2 的增加,從全部趨近到共存平衡點變成有部份 趨近到無病平衡點再變成全部趨近到無病平衡點,但還是能得到如同文獻[6]的反向分 歧(Backward bifurcation)現象存在。. 26.
(34) 第四章 結論 本篇文章中,我們對病毒感染模型加了延遲反應,並對感染方式進行修改,其中器官 1 的健康細胞會受到病毒直接感染,器官 2 的健康細胞則是受到被感染後的器官 2 的細胞 感染,藉此觀察是否會有與參考文獻中產生不一樣的動態情形。 𝑏. 從 3.1.3 章節的推倒中可得到,在𝑎 < 𝜉2 時,這模型有一個器官 2 的細胞沒受到感染 𝑏. (0). (2). 的平衡點E1 ,以及在𝑎 < 𝜉1 時,這模型有一個全部細胞都同時存在的平衡點E1 。此時可 𝑏. 𝑏. 觀察到系統中平衡點的穩定性與存在性會受到𝑎 、𝜉1 、𝜉2的影響,如定理三。當𝑎最大時, (0). 𝑏. (2). 只存在無病平衡點E0 且穩定;當𝑎最小時,三個平衡點都存在,但只有共生平衡點E1 穩 𝑏. (0). (2). (0). (0). 定;當𝑎介於𝜉1 、𝜉2 之間時,此時會受𝜉1 、𝜉2 的大小影響,存在E0 與E1 或存在E0 與E1 , 皆非無病平衡點才為穩定的。 由𝜉1 、𝜉2 我們可觀察到,𝜉1 =. β2 𝑒 −𝑎2 𝜏2 𝜇2. 所以當其越大時,對器官-2 的感染率就會越高, (2). 也就代表越容易被感染,因此當𝜉1 越大才能使平衡點E1 存在;𝜉2 =. 𝛾1 𝑒 −𝑎3 𝜏3 𝑒 −𝑎1 𝜏1 β1 ̃ 𝜇1 𝜇. ̃ β. − 𝜇̃所. 以當其越大時,對器官 1 的感染率就會越高且病毒的生成率也會越高,也就代表越容易被 (0). 感染且病毒在器官 1 越容易繁衍,因此當𝜉2 越大就能使平衡點E1 存在。 從本篇的模型可觀察到幾個現象,第一,從 3.1.3 的數值模擬中,當器官 2 只有很 少受感染的細胞時,器官 2 就不會被感染而發病產生病毒,但只要有病毒細胞存在器官 1 就會受影響,因此如果假設其病毒為肝炎病毒的話,可推得病毒細胞會呈現先感染最 外層的血液細胞 (器官 1),然後再透過受感染的細胞感染更深處的肝臟細胞 (器官 2); 第二,本篇的器官 2 的細胞受到的感染是由受感染的器官 2 的細胞影響,但人體中各器 官間其實也緊密相連,就像血液與其他各器官間都有流通,因此未來可以增加考慮受感 染血液細胞感染其他器官細胞的機制,以便進一步了解更真實的病毒動態。. 27.
(35) 文獻參考 [1] Cooke, K. L., & Grossman, Z. (1982). Discrete Delay, Distributed Delay and Stability Switches. Journal of Mathematical Analysis and Applications 86,592-627 . [2] Heffernan, J. M., & Wahl, L. M. (2005). Monte Carlo estimates of natural variation in HIV infection, Journal of Theoretical Biology 236,137-153. [3] Li, M. Y., & Shu, H. (2012). Joint effects of mitosis and intracellular delay on viral dynamics:two-parameter bifurcation analysis. Journal of Mathematical Biology 64,1005– 1020 . [4] Nowak, M. A., Bonhoeffer, S., Hill, A. M., Boehme, R., Thomas, H. C., & McDade, H. (1996). Viraldynamics in Hepatitis B Virus infection, Proc. Natl. Acad. Sci. USA 93,4398-4402. [5] Nowak, M. A., & May, R. M. (2000). Virus Dynamics:Mathematical Principles of Immunology and Virology, Oxford University Press, Oxford . [6] Qesmi, R., Wu, J., Wu, J., & Heffernan, J. M. (2010). Influence of backward bifurcation in a model of hepatitis B and C viruses. Mathematical Biosciences 224,118–125.. 28.
(36)
數據
Outline
相關文件
Baoshan sand and Xiangshan sand were first conducted in the laboratory, and TDR was used to measure the volumetric water content in order to obtain the
This study aims to explore whether the service quality and customer satisfaction have a positive impact on the organizational performance of the services and whether the
Therefore, in this project, to explore whether using Perusall combined with Zuvio IRS can help to strengthen the learning effect of flipped teaching and improve students'
In our research, four operational parameters, included flow velocity, intensity of voltage, volume of water sample, and concentration of effluent, were used to
The DEMATEL was used to analyze the key factors on elementary school parents' school choice.. According to analysis result, the most influencing parents to choose school factor is
The DEMATEL was used to analyze expert questionnaires, and found the crucial factors of electronic schoolbag-assisted instruction.. According to the analysis
Therefore, E-commerce system success model proposed by Molla and Licker (2001) was used to investigate the frequency of negative incidents (NCIs), quality
The purpose of this study was to explore the knowledge management status of elementary excellent teachers, strategy used and promoting factors.. A questionnaire
