國 立 交 通 大 學
生物科技研究所
碩士論文
利用定點飽和突變研究抹香鯨肌紅蛋白中
Ile-107 位置對其過氧化能力之影響
Site-Saturated Mutational Analysis of Isoleucine 107 from
Sperm Whale Myoglobin on the Effect of Peroxidase
Activity
研究生: 林宏明
指導教授: 吳東昆 博士
利用定點飽和突變研究抹香鯨肌紅蛋白中
Ile-107 位置對其
過氧化能力之影響
Site-Saturated Mutational Analysis of Isoleucine 107 from Sperm Whale Myoglobin on the Effect of Peroxidase Activity
研究生:林宏明 Student: Hung-Ming Lin 指導教授:吳東昆 博士 Advisor: Dr. Tung-Kung Wu
國 立 交 通 大 學
生物科技研究所
碩士論文
A Thesis
Submitted to Department of Biological Science and Technology College of Science
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Biological Science and Technology December, 2006
Hsinchu, Taiwan, Republic of China
利用定點飽和突變研究抹香鯨肌紅蛋白中 Ile-107 位
置對其過氧化能力之影響
研究生:林宏明 指導教授:吳東昆 博士 國立交通大學 生物科技研究所碩士班摘要
肌紅蛋白(myoglobin)為一種血基質蛋白(heme-protein),在脊椎動 物體內具有儲存及攜帶氧氣的功能。本論文的研究主要是將不具酵 素活性的肌紅蛋白突變成具有過氧化酵素(peroxidase)活性的功能。 將 mb 基因殖入 pET28a(+)表現質體中,利用定點突變及定點飽和突 變技術改變 His-64、Val-68、Ile-107 這三個推測可改變活性的胺基 酸。利用 IPTG 誘導蛋白質的大量表現以形成包涵體(inclusionbody),接著以 guanidine hydrochloride 使包涵體變性再復性後,利用 DEAE 管 柱 純 化 , 可 以 得 到 純 度 高 達 90% 的 脫 輔 基 肌 紅 蛋 白 (apo-myoglobin),利用蛋白質輔基重組實驗可將血基質(heme)包入突 變 的 脫 輔 基 肌 紅 蛋 白 中 。 與 過 氧 化 氫 和 2,2’-azinobis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS)反應後可
較文獻的雙重突變 H64D/V68L Mb 以及利用定點飽和突變技術改變 Ile-107 的 三 重 突 變 , 在 經 由 動 力 學 參 數 測 量 後 發 現 H64D/V68L/I107M Mb 在與過氧化氫形成 Compound I 的效率可提高 30%,與 ABTS 的一個電子傳遞效率則提高 60%。另外也發現將 Ile-107 置換成體積較小的 Ala 及 Val 對形成 Compound I 的效率影響
不大,但是卻可以提高一個電子傳遞效率 47%及 36%。因此認為
Ile-107 可能藉由立體效應去影響酵素與受質的一個電子傳遞效率。 未來本實驗室會將各種不同金屬的紫質包入突變的脫輔基肌紅蛋白 中,並研究其特性以期能應用並開發新型的染料敏化生物性太陽能 電池(dye-sensitized biosolar cell)。
Site-Saturated Mutational Analysis of Isoleucine 107
from Sperm Whale Myoglobin on the Effect of
Peroxidase Activity
Student: Hung-Ming Lin Advisor: Dr. Tung-Kung Wu Institute of Biological Science and Technology
National Chiao Tung University
Abstract
Myoglobin is a heme-protein, functioning as oxygen storage/carrier in vertebrates. In this study, the nonenzymatic myoglobin was functionally converted into a heme enzyme with peroxidase activity. Three amino acid residues, His-64/Val-68/Ile-107, located on the putative active site cavity, were subjected to site-directed mutagenesis and cloned into a pET-28a(+) expression vector. Following IPTG induction, the cell cultures were harvested and subjected to protein purification. After guanidine hydrochloride disruption and renaturation, the soluble apo-myoglobin was purified to homogeneity by DEAE chromatography. The heme molecule was reconstituted into these renatured apo-myoglobin mutants. A well established peroxidase activity with one-electron oxidation of 2,2’-azinobis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) was observed by the UV/VIS spectrophotometer from the myoglobin mutants, demonstrating its capability in electron transfer reaction. Among these mutations, as compared with the original MbH64D/V68L double mutant, MbH64D/V68L/I107M triple mutant exhibited a 30% activity increase in the compound I formation, whereas, a 60% activity increase in the one-electron oxidation was observed. Alternativily, the MbH64D/V68L/I107A or MbH64D/V68L/I107V showed the similar compound I formation rate, but with the one-electron oxidation rate of 47% or 36% increment respectively. These results indicated that the Ile-107 position may influence the one-electron oxidation of substrates via steric effect after
the compound I formation. In the future, various metalloporphyrins with different metal ions will be reconstituted into the apo-myoglobin mutants to investigate their potentials as novel dye-sensitizers for biosolar cell application.
謝誌
碩士班的研究生活真的讓我獲益良多,首先當然要感謝我的指導 教授吳東昆 博士,感謝老師能讓我在這實驗室新的題目中學習,並 且在忙碌中也經常的提供我們實驗上的幫助及傳授正確的實驗觀 念。感謝口試委員袁俊傑 教授、刁維光 教授、林敬堯 教授及鄭建 中 教授於百忙中能撥空參加我的口試、審閱我的論文,有了各位寶 貴的建議讓本人及本論文都進步了不少。 實驗室的各位對我的幫助都不少,因此我要感謝程翔學長及媛婷 學姊在我初進實驗室時指導我分生方面的技術以及實驗和生活上許 多的幫助,感謝坐在我旁邊的裕國學長將他許多實驗的經驗及技術 傳授於我,感謝和我同 group 的豪哥在實驗上幫我許許多多的忙, 感謝文鴻、晉源、宗哥、大鳥經常與我討論實驗上問題及週遭的生 活,感謝小妹和大景陪我上英文課、吃宵夜、夾娃娃和 PK 投籃機, 感謝食神皓宇、文暄及新進來的衣娟學姊、Nayak、小高、文祥、采 婷還有接我題目的世勳在許多方面的幫助,感謝已畢業的小宇、宏 城、小八、阿美和希希慷慨的照顧我們學弟妹們。 此外,我也要感謝袁老師實驗室的俊傑之友會成員對我的關心及幫忙,特別感謝威震學長對本論文動力學測定方面的幫助以及感謝 佳穎學姊、弘毅學長及我的好同學彥棋、詩穎對我生活上的關心, 還有 fish、世昌和偉志在各方面的幫忙。 最後我要感謝我的家人,感謝讓我寄宿 2 年的三叔及三嬸,還要 謝謝培養我的母親,有她的支持我才能進到交通大學來完成這篇論 文。雖然碩士班短短的兩年家中卻發生了不少事情,有母親妥善的 處理我才能安心的完成我的學業。感謝所有幫助過我的人,謝謝你 們!!
目錄
頁次
中文摘要………..Ⅰ 英文摘要……….Ⅲ 謝誌………V 目錄………VII 圖目錄………..XI 表目錄………....XVI 第一章 緒論...1 1-1 肌紅蛋白(myoglobin)的簡介 ...1 1-2 紫質(prophyrin)的簡介...4 1-3 血基質蛋白(heme protein) ...5 1-4 過氧化酵素 ...6 1-5 肌紅蛋白經突變後具有過氧化酶酵素活性 ...9 1-6 染料敏化太陽能電池 (DSSC) ...12 1-7 肌紅蛋白在人工光合作用反應中心的應用 ...14 1-8 研究目的 ...20 第二章 實驗材料及方法 ...222-1 實驗材料 ...22 2-1-1 化學藥品與材料 ...22 2-1-2 緩衝液及溶液配製 ...24 2-1-3 實驗儀器 ...28 2-1-4 菌株與載體 ...29 2-2 實驗方法 ...30 2-2-1 目標基因的建構 ...31 2-2-2 重組質體的建構 ...33 2-2-2-1 質體之轉化作用...34 2-2-3 定點突變與飽和定點突變實驗...35 2-2-4 脫輔基(apo-from)蛋白質的表現 ...38 2-2-5 包涵體(inclusion body)粗萃液之製備 ...39 2-2-6 包涵體(inclusion body)的變性與再摺疊 ...39 2-2-7 脫輔基(apo-form)蛋白質的純化 ...40 2-2-8 蛋白質分子量及純度分析...40 2-2-9 蛋白質輔基重組實驗...42 2-2-10 酵素最適化條件之研究...43 2-2-10-1 最適反應環境之酸鹼值測定...43 2-2-10-2 溫度與活性之關係...43
2-2-11 酵素動力學參數測定 ...44 第三章 結果與討論 ...46 3-1 目標基因質體之建構 ...46 3-2 篩選目標 ...46 3-2-1 細胞粗萃液的活性篩選...49 3-3 脫輔基(apo-form)蛋白質表現及純化 ...51 3-4 蛋白質輔基重組 ...53 3-5 過氧化酵素活性最適反應條件 ...57 3-5-1 最適反應之pH值 ...57 3-5-2 溫度與活性之關係 ...59 3-6 酵素動力學參數 ...60 第四章 結論與未來展望 ...82 第五章 參考文獻 ...84 附錄一...87 附錄1-1 建構不同金屬紫質的MbH64D/V68L/I107M三重突變肌紅蛋白 ...87 附錄二...89 附錄2-1 單株抗體的製備 ...89 附錄2-2 免疫動物 ...89 附錄2-3 骨髓癌細胞 (myeloma cell)...90
附錄2-4 細胞融合 ...91 附錄2-5 細胞增殖及取樣篩檢 ...92 附錄2-6 融合細胞之單株化 ...93 附錄2-7 單株抗體之產生 ...94 附錄2-8 利用西方墨點法 (Western blotting)確認單株抗體之專一 性...94
圖目錄
頁次
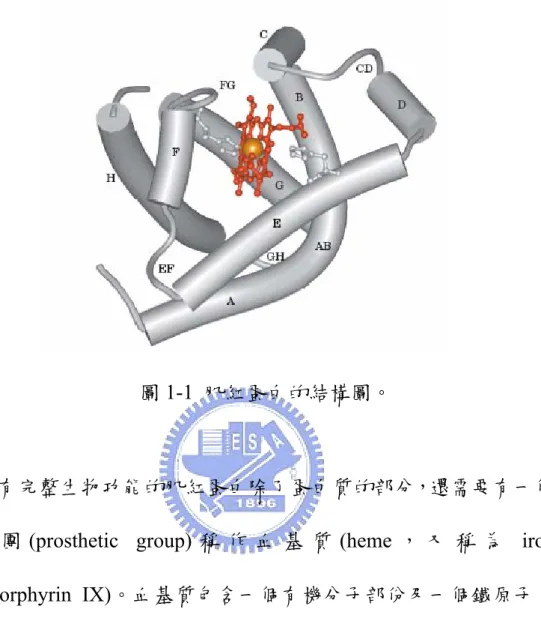
圖1-1 肌紅蛋白的結構圖………2
圖1-2 紫質和血基質的化學結構………5
圖1-3 推測的 Compound I 形成機制……….… 7
圖1-4 兩種酵素 peroxidase 和 cytochrome P450 反應的比較………...8 圖 1-5 在 chloroperoxidase 中 Glu-183 對形成 Compound I 的可能功 能………..11
圖1-6 DSSC 太陽能電池的運作原理……….13
圖1-7 尚未與肌紅蛋白結合的兩種三聚體結構………..15
圖1-8 Mb(FeIIIOH2)-Ru2+-BXV4+在經過激發Ru(bpy)3 後的能階與選擇
性路徑………..16 圖 1-9 (a)為Ru(bpy)3的MLCT band在 460 nm吸收值增加的動力學追
縱;(b)為時間偵測範圍至 2 ms,分別偵測在 426 和 610 nm的瞬態吸 收值動力學(transient absorption kinetics)………..17
圖 1-10 圖示將 1 和 2 與脫輔基肌紅蛋白質包覆後而形成的肌紅蛋白
質三聚體………..19
圖3-1 含相同蛋白質量的三重突變肌紅蛋白H64D/V68L/I107X Mb細
胞粗萃液10 μL與 90 μL 的 1 mM ABTS、5 mM H2O2 反應後偵測 405
圖3-2 20%的 SDS-PAGE 分析………53 圖3-3 UV/Vis 全光譜掃描圖………...55 圖 3-4 相同量蛋白質的野生型(Mbwt)及MbH64D/V68L與Heme進行蛋白 質與輔基的重組後與 1 mM ABTS及 2 mM H2O2反應依時間偵測 ABTS氧化產物 730 nm 吸光值……….56 圖3-5 MbH64D/V68L於不同pH值環境之殘留活性百分率………...58 圖3-6 MbH64D/V68L/I107M於不同溫度下反應之殘留活性百分率……....60 圖 3-7 ABTS濃度對MbH64D/V68L催化ABTS受質的Michaelies-Menten 曲線圖………..61 圖 3-8 Lineweaver-Burk雙倒數圖計算MbH64D/V68L對受質ABTS的K’m 及V’max值………..61 圖 3-9 ABTS 濃 度 對 MbH64D/V68L/I107M 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖……….62 圖3-10 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107M對受質ABTS的 K’m 及V’max值………62 圖 3-11 ABTS 濃 度 對 MbH64D/V68L/I107A 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖……….63 圖 3-12 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107A對受質ABTS的 K’m 及V’max值………63
圖 3-13 ABTS 濃 度 對 MbH64D/V68L/I107V 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖……….64 圖 3-14 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107V對受質ABTS的 K’m 及V’max值………64 圖 3-15 ABTS 濃 度 對 MbH64D/V68L/I107L 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖……….65 圖 3-16 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107L對受質ABTS的 K’m 及V’max值………65 圖 3-17 ABTS 濃 度 對 MbH64D/V68L/I107F 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖……….66 圖 3-18 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I10F對受質ABTS的 K’m 及V’max值………66 圖 3-19 ABTS 濃 度 對 MbH64D/V68L/I107G 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖……….67 圖 3-20 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107G對受質ABTS的 K’m 及V’max值………67 圖 3-21 ABTS濃度對MbWt催化ABTS受質的Michaelies-Menten 曲線 圖……….68 圖 3-22 Lineweaver-Burk雙倒數圖計算MbWt對受質ABTS的K’m 及
V’max值……… 68 圖 3-23 H2O2濃度對MbH64D/V68L催化ABTS受質的Michaelies-Menten 曲線圖………..69 圖 3-24 Lineweaver-Burk雙倒數圖計算MbH64D/V68L對受質H2O2的K’m 及V’max值………..69 圖 3-25 H2O2 濃 度 對 MbH64D/V68L/I107M 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖………...………..70 圖 3-26 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107M對受質H2O2的 K’m 及V’max值………..…..70 圖 3-27 H2O2 濃 度 對 MbH64D/V68L/I107A 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖………..….…..71 圖 3-28 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107A對受質H2O2的 K’m 及V’max值………..…..71 圖 3-29 H2O2 濃 度 對 MbH64D/V68L/I107V 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖………...…..72 圖 3-30 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107V對受質H2O2的 K’m 及V’max值………..……..72 圖 3-31 H2O2 濃 度 對 MbH64D/V68L/I107L 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖………...…..73
圖 3-32 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107L對受質H2O2的 K’m 及V’max值………..……..73 圖 3-33 H2O2 濃 度 對 MbH64D/V68L/I107F 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖………...…..74 圖 3-34 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107F對受質H2O2的 K’m 及V’max值………..……..74 圖 3-35 H2O2 濃 度 對 MbH64D/V68L/I107G 催 化 ABTS 受 質 的 Michaelies-Menten 曲線圖………...…..75 圖 3-36 Lineweaver-Burk雙倒數圖計算MbH64D/V68L/I107G對受質H2O2的 K’m 及V’max值………..……..75 圖 3-37 H2O2濃度對MbWt催化ABTS受質的Michaelies-Menten 曲線 圖………...…..76 圖 3-38 Lineweaver-Burk雙倒數圖計算MbWt對受質H2O2的K’m 及 V’max值………..…….. 76 圖 3-39 過氧化酶 peroxidase 和 peroxygenase 催化反應循環形成 compound I 與 compound II 的步驟………....78
表目錄
頁次
表2-1 設計的 6 條長單股 DNA 引子及兩端的 2 條短引子…………...32 表2-2 rTth polymerase kit 的 PCR 反應之組成……….33 表2-3 PCR program………...…..33 表2-4 設計合成 3 種定點突變的互補引子(Primer) ………...…..37 表 2-5 Quick Change Site-Directed Mutagenesis 聚合酶連鎖反應組 成………..38 表2-6 QuickChange Site-Directed Mutagenesis Kit 使用之聚合酶連鎖 反應放大程式………..38 表2-7 20% separating gel 與 4% stacking gel 的 SDS-PAGE 膠片組成 份………..41
表 3-1 野生型抹香鯨肌紅蛋白中各胺基酸殘基與血基質鐵原子最近
的原子及相距的距離………..49
表3-2 突變後的肌紅蛋白催化受質的動力學參數………..80
表 3-3 突變後的肌紅蛋白催化受質的 turnover number 及 catalytic efficiency………..80
第一章 緒論
1-1 肌紅蛋白(myoglobin)的簡介
肌紅蛋白(Myoglobin)在生理上的功能為與一分子的氧氣結合,在 肌肉組織作為氧氣儲存的蛋白質,以緩衝組織呼吸時對氧氣濃度的 需求。它對氧氣的親和力介於血紅蛋白(Hemoglobin)與細胞色素 (Cytochromes)之間,因此肌紅蛋白在組織中接收從血紅蛋白釋出的 氧氣,並將氧氣供給細胞中粒線體的細胞色素,提供其氧化呼吸作 用所需的氧氣。在海洋哺乳類動物(例如: 海豹和鯨魚)的肌肉中,肌 紅蛋白特別豐富,因此可以延長這些動物在海底潛水的時間。 肌紅蛋白的三級結構為水溶性的橢圓球狀(44 x 44 x 25 Å),以單 體(monomer)形式存在於自然界中,分子量約為 17 kDa。肌紅蛋白的 二級結構含有很高比例 (75%)的 α-螺旋結構(α-helices),由 153 個 胺基酸摺疊成 8 段右手旋的 α-螺旋結構,並分別命名為 A-H 螺旋, 各螺旋的中間由一些非螺旋區連接。疏水性(hydrophobic)的胺基酸 殘基在經過正確摺疊後大部分會處於整個肌紅蛋白分子的內部,而 親水性(hydrophilic)的胺基酸殘基則位於分子的外部,因此使得整個 肌紅蛋白分子相對的具有水溶性。圖1-1 肌紅蛋白的結構圖。
具有完整生物功能的肌紅蛋白除了蛋白質的部分,還需要有一個 輔 基 團 (prosthetic group) 稱 作 血 基 質 (heme , 又 稱 為 iron protoporphyrin IX)。血基質包含一個有機分子部份及一個鐵原子, 有機分子部分稱為原紫質(protoporphyrin)是由四個吡咯環(pyrrole rings)組成。而這四個吡咯環是藉由次甲基橋(methane bridges)相連接 而 構 成 一 個 四 吡 咯 環(tetrapyrrole ring) , 且 有 四 個 甲 基 (methyl groups),二個乙烯基(vinyl groups)及二個丙酸側鏈(propionate side chains)與這個四吡咯環相連。血基質中的鐵原子與原紫質環中心的 四個氮共價結合,鐵原子還能另外形成二個鍵,分別位於血基質平 面的二側,這些結合部位稱為第五及第六配位。
早 在 1960 年代肌紅蛋白的結構已由英國劍橋大學的 John Kendrew 利用 X-ray 結晶學方法解出,此為第一個利用 X-ray 結晶學 解出結構的蛋白質[1]。可顯示出肌紅蛋白各原子間的相對位置。其 結構顯示肌紅蛋白由分別標示為 A-H 的 8 個螺旋片段構成,並在 E 和 F 螺旋片段之間留下一個疏水性的狹縫以供血基質的嵌入。血基 質的四吡咯環與狹縫內的疏水性胺基酸藉由疏水性的交互作用穩定 血基質與肌紅蛋白的連結。血基質中間的鐵原子直接與肌紅蛋白的 近端組胺酸(proximal histidine) (稱為殘基 F8;意即 F 螺旋的第 8 個 胺基酸殘基)即第 93 號組胺酸的 Nε(組胺酸側鏈上 ε 位置的氮原子) 形成共價鍵結。氧氣的結合部位在血基質平面的另一側,即鐵原子 的第六配位上。鐵原子可以處於亞鐵(+2)或高鐵(+3)二種不同的氧化 態 , 分 別 稱 為 亞 鐵 肌 紅 蛋 白(ferromyoglobin) 及 高 鐵 肌 紅 蛋 白 (ferrimyoglobin) , 高 鐵 肌 紅 蛋 白 亦 可 稱 為 正 鐵 肌 紅 蛋 白 (metmyoglobin)。僅有亞鐵肌紅蛋白(+2 氧化態)能夠與氧氣結合。第 64 號組胺酸殘基(殘基 E7)稱為遠端組胺酸(distal histidine),靠近血 基質但不與它結合。含有血基質的肌紅蛋白稱為 holo-form 的肌紅蛋 白,若沒有血基質的結合,此形態的肌紅蛋白通常稱為脫輔基肌紅 蛋白(apomyoglobin)。
1-2 紫質(prophyrin)的簡介
紫質的結構是由四個吡咯環(pyrrole rings)藉由次甲基橋(methane bridges)相連接而構成一個四吡咯環(tetrapyrrole ring)。在自然界中及 人類體內均存在紫質的衍生物,與人類生活有著密不可分的關係。 例如,人體中負責氧氣運輸與儲存的血基質(heme)及自然界中進行 光合作用的葉綠素。近幾年來關於紫質的研究與應用已愈來愈多, 例如,光電元件(optoelectronic device)、分子邏輯元件(molecular logic device)、太陽能的吸收與儲存(artificial solar energy harvesting and storage scheme)以及太陽能電池(solar cell)[2-4]。紫 質 的 中 間 若 嵌 合 一 個 金 屬 原 子 則 此 種 紫 質 稱 為 金 屬 紫 質 (Metalloporphyrin),例如:血基質(heme)就是嵌合入一個鐵原子 [the iron(II) protoporphyrin-IX complex] 在生理上作為氧氣的儲存和運 輸。
含有鎂離子嵌入其中的紫質是光合作用中能將光能轉換成化學 能的關鍵因子。因此不同的金屬嵌入紫質中,使其擁有許多的應用 價值包括應用在光學治療[5],新穎的光化學反應中心[6],生物模擬
模型,催化和感測系統。圖 1-2 為紫質和血基質的化學結構。 圖1-2 紫質和血基質的化學結構。
1-3 血基質蛋白(heme protein)
含 有 血 基 質 的 蛋 白 質 可 被 稱 為 血 基 質 蛋 白(heme protein or hemoprotein),生物中目前已知的血基質蛋白可歸納出四種生物功 能:電子傳遞(如:細胞色素 b5)、傳輸氧氣(如:血紅蛋白,肌紅蛋 白)、氧氣或一氧化碳的感測(如:FixL,CooA)及催化氧化還原反應 (如:horseradish peroxidase,cytochrome P450,catalase,NO synthase, NO reductase)。這些蛋白質雖然功能各異,但是它們都有一個共同 的血基質(iron protoporphyrin IX)當作其輔基團(prosthetic group)。在血基質蛋白中,血基質週邊的胺基酸殘基似乎是控制血基質蛋 白功能的關鍵。帶電的血基質蛋白通常在其鐵原子的軸位(axial)有二 個很強的配位體結合,以防止其它可能的受質進入。如肌紅蛋白的 第六配位處也就是遠端口袋的(distal pocket)位置,經常會被一個水 分子所佔據,以利於其它的配體,像是氧氣來交換。在近端的組胺 酸殘基被當做軸位的配體,而遠端的組胺酸則是藉由氫鍵來穩定氧 氣與血基質的結合。另外在作為感測氧氣的血基質蛋白 FixL 中,其 血基質周圍也顯露出一個可以供配位結合的位置,而有些血基質蛋 白的鐵原子亦可與過氧化氫配位結合並發生反應。同樣的血基質卻 可以造成功能各異的蛋白質[7]。因此研究血基質週邊胺基酸殘基的 改變,使不具酵素功能的肌紅蛋白變成具有過氧化酵素(peroxidase) 活性的研究,可以幫助我們更了解血基質蛋白。
1-4 過氧化酵素
山葵過氧化酵素(horseradish peroxidase;HRP),是常見的過氧化 酵素之一,這類高鐵的過氧化酵素(ferric peroxidase)與過氧化氫(H2O2) 反應時會打斷其氧-氧鍵(HO-OH bond),使血基質中的鐵和氧結合構成特殊形式(oxo-ferryl(O=FeIV)),而紫質部分為帶有自由基的陽離子 (prophyrin radical cation),這樣的血基質稱為Compound I (oxo-ferryl porphyrin radical cation)。在Compound I (圖 1-3)的形成過程中,過氧 化氫結合到血基質的鐵原子上,遠端的組胺酸先是當作general base 進行質子化,遠端質子化的組胺酸再扮演general acid的角色提供質子 氫給氧-氧鍵斷裂的OH– 以形成一分子的水。這過程中帶正電荷的遠 端精胺酸(distal arginine)亦幫助氧-氧鍵的極化(polarization)。因此對於 遠端組胺酸與精胺酸殘基的貢獻稱之為〝拉效應(pull effect)〞。而相 對的,帶陰電的近端組胺酸提供很強的電子去幫助穩定氧-氧鍵斷裂 的過渡狀態(transition state),因此對於近端組胺酸的貢獻稱為〝推效 應(push effect)〞[7]。 圖1-3 推測的 Compound I 形成機制。 N N H Arg+ III N H N Fe N N H Arg+ III N H N Fe O H O H N N H Arg+ III N H N Fe O H O N N H Arg+ N H N Fe O His His H2O2 H2O2 His His His His
General base function histidine General acid function histidine
δ− δ+ His His IV H2O +
.
Compound I另外一些血基質蛋白,如細胞色素 P450 單加氧酶(cytochrome P450 monooxygenases),則是與氧氣而非與過氧化氫結合,並且需要 二個電子以形成 Compound I。圖 1-4 顯示自然界中過氧化酵素與單 加氧酶(monooxygenases)相同反應中間物(reactive intermediates)的部 分,以及單加氧酶與過氧化酵素的不同在於單加氧酶將 Compound I 的氧原子傳遞給受質。 圖1-4 兩種酵素 peroxidase 和 cytochrome P450 反應的比較[7]。
1-5 肌紅蛋白經突變後具有過氧化酶酵素活性
利用蛋白質酵素活性部位的結構可以影響其酵素活性的特性,目 前已有將生理上僅能儲存氧氣的肌紅蛋白變成具有過氧化酵素活性 的研究報導。比較兩種已被結晶出結構的代表性血基質蛋白,肌紅蛋 白及細胞色素 C 過氧化酶(cytochrome c peroxidase;Ccp)這兩種血基 質蛋白,它們具有結構完全相同的血基質,也一樣有近端的咪唑 (imidazole)配體以及其遠端的活性部位皆有組胺酸殘基。但是在已結 合氧氣分子的肌紅蛋白中,遠端的組胺酸可以非常靠近已配位結合鐵 原子的氧氣分子。而在已結合氧氣分子的細胞色素 C 過氧化酶中, 遠端的組胺酸與氧氣分子的距離則相對較肌紅蛋白中的遠。因此 Ozaki 等研究學者推測,由於肌紅蛋白中的遠端組胺酸離血基質的鐵 原子較近,因此使肌紅蛋白與過氧化氫反應形成Compound I 的能力 遠低於細胞色素 C 過氧化酶[7]。 基於遠端組胺酸的位置有決定血基質蛋白的功能的重要性, Ozaki 等研究學者將抹香鯨肌紅蛋白(sperm whale myoglobin)及細胞 色素 C 過氧化酶的結晶結構做活性區域的比較,再改變遠端組胺酸 的位置,希望可以得到具有過氧化酵素活性的突變肌紅蛋白[7]。他們先是單純的將遠端的組胺酸殘基與其它活性區域位置的胺基酸殘
基對調,因此建構 F43H/H64L 和 L29H/H64L 二組胺基酸殘對調的
雙重突變肌紅蛋白,由於是將活性區域的遠端組胺酸位置改變,因此 稱為〝distal histidine relocation mutant〞。而他們實驗的結果顯示 F43H/H64L 的雙重突變肌紅蛋白遠端組胺酸環境較相似於過氧化 酶,此雙重突變肌紅蛋白的過氧化酶(peroxidase activity)與過氧合酶 酵素活性(peroxygenase activity)都優於野生型肌紅蛋白,並且具有 general acid-base 的催化作用。 在野生型肌紅蛋白中Compound I 無法形成的另一個原因可能是 肌紅蛋白遠端組胺酸的迅速氧化作用,因此嘗試飽和突變實驗將遠端 組胺酸置換成各種不同胺基酸的實驗亦被 Ozaki 等研究學者發表 [8]。將肌紅蛋白的遠端組胺酸殘基改變後,Compound I 便可以很快 的 形 成 , 尤 其 是 突 變 成 天 門 冬 胺 酸 (Asp) 後 功 能 便 類 似 chloroperoxidase(CPO)活性區域的麩胺酸(Glu)[9] (圖 1-5)。但是天門 冬胺酸的羧酸氧原子在突變的肌紅蛋白中與血基質鐵原子的距離似 乎太遠,因此推測在突變的肌紅蛋白中第64 號天門冬胺酸(Asp)殘基 的功能可能是增加對過氧化氫的極性與親合性,來改善突變肌紅蛋白 的氧化活性。
N H N H O O H OOH III S Fe NH+ N H O O H III S Fe O OH N H N H O O S Fe O His 105 Glu183 Cys 29 His 105 Glu183 Cys 29 His 105 Glu183 Cys 29 IV +
.
HOH圖1-5 在 chloroperoxidase 中 Glu-183 對形成 Compound I 的可能 功能。 另外,從結構的角度來看肌紅蛋白的Val 68 位置對受質到達活性 部位有很大的重要性,將其置換成各種不同的胺基酸殘基會大大的改 變其活性區域的形狀與體積[10]。因此以 H64D 為模板,將 Val 68 突 變成各種不同胺基酸的雙重突變已被建構出來以測量其過氧化酶 (peroxidase)與過氧合酶(peroxygenase)酵素活性,在這些雙重突變中, H64D/V68L 突變在形成 Compound I 後,與受質傳遞一個電子的效率 最好,即有很好的過氧化酶活性。而 H64D/V68I 雖然形成 Compound I 的效率較差但卻有最好的過氧合酶活性,因此過氧合酶活性的速率 決定步驟可能是在Compound I 形成後與一當量的受質反應。另外也 發現,H64D/V68A 和 H64D/V68S 這二組雙重突變肌紅蛋白在過氧合 酶 酵 素 活 性 (peroxygenase activity) 上 具 有 鏡 像 選 擇 性
(enantioselectivity),再搭配結晶結構的結果發現影響氧化速率和造成
鏡像選擇性的可能原因有二,可能是第 64 號天門冬胺酸(Asp 64)殘基
直接和受質的立體交互作用(steric interaction)或是使受質和遠端口袋 的水分子產生極化的交互作用(polar interaction)。
在 1998 年 Wan 等人也以 PCR 隨意突變(random mutagenesis)的 方法,企圖篩選比野生型肌紅蛋白的過氧化酶活性好的突變株。在經 過四次 PCR 反應後,他們篩選到一株氧化酶活性較野生型有明顯差 別的 T39I/K45D/F46L/I107F 突變株,由其酵素動力學的資料顯示這 個突變株可以穩定Compound I 的形成。此四重突變的肌紅蛋白與野 生型肌紅蛋白的熱穩定性相似,因此其蛋白質整體結構應該沒有太大 的差異[11]。
1-6 染料敏化太陽能電池 (DSSC)
由於全球氣候變遷、空氣污染問題以及資源日趨短缺之故,太陽 能發電是目前廣泛被推薦使用的一種替代能源,將太陽光能轉變成電 能可減少對環境的污染。瑞士科學家Grätzel在 1991 年提出一種新型 的太陽能電池稱為染料敏化太陽能電池(Dye-sensitized solar cell),此型太陽能電池可源自 19 世紀,人們照相技術的理念。其製作是採用 奈米結構的電極材料,以及適切的染料,組成光電效率超過 11﹪的 光電池。此類型光電池的工作原理是藉由染料做為吸光材料。染料中 價電層電子受光激發,躍昇至高能階層,進而傳導至奈米二氧化鈦半 導體的導電層,再經由電極引至外部。失去電子的染料則經由電池中 的電解質得到電子,電解質是由I/I3+溶於有機溶劑中形成。圖1-6 清 楚的標示染料敏化太陽能電池的運作原理[12]。 圖1-6 DSSC 太陽能電池的運作原理。 對於應用於染料敏化太陽能電池中的染料有以下幾項要件:(1) 它必須可以堅固的連接到二氧化鈦半導體的表面,並且可以將躍昇的 電子傳導到導電層中。(2)氧化還原的位能要夠高,並可以迅速的從 電解質中得到電子而還原。(3)它還必須可以忍受至少 108次以上的光
照氧化還原反應[13]。 一 種 編 號 為 N3 或 N-719 染 料 的 有 機 釕 金 屬 化 合 物 cis-di(thiocyanato)bis(2,2’-bipyridyl-4,4’-dicarboxylate) ruthenium(II) 已被用於染料敏化太陽能電池中,並且經由偵測得知,此染料躍昇的 電子傳導到導電層中的時間為 57 飛秒(femtosecond),而在波長 530 nm 的光源激發下,此染料的入射光子對電流轉換效率(incident photon to current conversion efficiency;IPCE)高達 85 %[12, 14]。
1-7 肌紅蛋白在人工光合作用反應中心的應用
在自然界的光合作用包括能量傳遞和電子轉移兩個最基本的過 程︰太陽能首先被天線色素分子(antenna pigment molecules)接收後傳 遞到光合作用的反應中心(reaction center);反應中心接受能量後發生 電荷分離(charge-separate)與電子轉移(electron transfer)繼而進行一系 列的氧化還原反應,將太陽能轉變成可貯存的化學能。人們為了模擬 光合作用的這兩個過程,設計合成了許多比自然界光合作用系統更簡 單的模型分子,研究它們的光物理和光化學性質,探索光合作用中能 量傳遞和電子轉移的機制,最終實現光能轉換的目的。其中一種模擬
光合作用反應中心的模型是利用半合成(semisynthesis)的方法製造一 含有肌紅蛋白當電子提供者的donor-sensitizer-acceptor的三聚體(triad) [15] 。 Mb(FeIIIOH2)-Ru2+-BXV4+,1, 的 acceptor (cyclobis(paraquat -
p-phenylene) BXV4+)是利用catenane非共價連結sensitizer (Ru(bpy)3),
而sensitizer則是與肌紅蛋白的血基質共價結合。另一種相似結構的三 聚體Mb(ZnPP)-Ru2+-BXV4+,2,也被以cofactor reconstitution的方式合成 出來(圖 1-7)。 圖1-7 尚未與肌紅蛋白結合的兩種三聚體結構[15]。 這兩種三聚體在與脫輔基肌紅蛋白質包覆後,形成一種以肌紅蛋 白為光合作用反應中心的全新系統。藉由此系統可以研究光誘導後電 荷 分 離 及 電 子 轉 移 的 過 程 。 實 驗 發 現 , 以 雷 射 激 發 此 系 統 的 sensitizer , 其 激 發 的 電 子 會 被 傳 遞 到 acceptor (BXV4+) 而 形 成
Mb(FeIIIOH2)- Ru3+-BXV3+•的過渡狀態。接下來由於熱力學上的不穩
定會使得一個氧化還原反應發生,即電子由系統中的高鐵肌紅蛋白 (ferric-Mb)轉移到Ru3+(bpy)3上,以形成Mb(FeIV=O)-Ru2+-BXV3+•。詳
細反應步驟如圖1-8。
圖1-8 Mb(FeIIIOH2)-Ru2+-BXV4+在經過激發Ru(bpy)3 後的能階與
選擇性路徑[15]。
當Ru(bpy)3在溶液中時,其UV-Vis光譜圖會呈現兩個主要的吸收
峰,一個為位於284 nm的高能帶的吸收,此為配位基的π→π*的躍
charge-transfer)的躍遷。此種MLCT相當於在激發態時,Ru被氧化, bpy被還原。因次藉由偵測 460 nm吸收峰消長的時間可以得知圖 1-8
中步驟1 與步驟 2 電子傳遞的時間。步驟 1 為sensitizer中激發電子傳
遞到acceptor (BXV4+)的時間由於太快因而無法測得,但在步驟 2,電
子由系統中的高鐵肌紅蛋白(ferric-Mb)轉移到Ru3+(bpy)3上則被偵測
約為1 μs。由Mb(FeIIIOH2)-Ru3+-BXV3+•變成Mb(FeIII•OH2)-Ru2+-BXV3+•
的狀態後很快的會轉變成電荷分離態(charge-separated state)的最低能 階態即Mb(FeIV =O)-Ru2+-BXV3+•。可由 426 nm的吸收峰(圖 1-9)證實 oxoferryl-Mb的產生,而停留在Mb(FeIV =O)-Ru2+-BXV3+•的時間經測量 超過2ms。另外(BXV3+•)的存在可藉由偵測的610 nm吸收峰的消長, 而整個三聚體若沒有肌紅蛋白的部分則(BXV3+•)會在 1 μs內就淬熄 (quenching)消失。可見得肌紅蛋白部分在這整個系統是非常重要的。 (b) 圖1-9 (a)為Ru(bpy)3的MLCT band在 460 nm吸收值增加的動力學 追縱;(b)為時間偵測範圍至 2 ms,分別偵測在 426 和 610 nm的瞬態
吸收值動力學(transient absorption kinetics)。 以鋅取代鐵的Mb(ZnPP)-Ru2+-BXV4+也被用來做比較,其電荷分 離態的能階依次為Mb(Zn)-*Ru2+-BXV4+ > Mb-(Zn)-Ru3+-BXV3+• > Mb(Zn+)-Ru2+-BXV3+•。總體而言由於鋅的氧化數不像鐵如此複雜因 而加快了到達其電荷分離態最低能階Mb(Zn+)-Ru2+-BXV3+•的時間。 相同的,若沒有肌紅蛋白的部分則(BXV3+•)會很快的淬熄而消失。 蛋 白 質 部 分 在 整 個 系 統 中 所 扮 演 的 功 能 可 能 是 固 定 發 射 團 (chromophore)以及調控電子的傳遞並穩定電荷分離態。而在結構方面 可能是穩定或儘可能的延伸整個三聚體的U-shape-like 構型。 比較鐵和鋅二種肌紅蛋白三聚體在電荷分離態最低能階停留的 時 間 , 可 發 現 Mb(FeIV=O)-Ru2+-BXV3+• 超 過 2 ms , 而 Mb(Zn+)-Ru2+-BXV3+•只存在約 100 μs。可能是Mb(FeIV=O)在其血基 質的活性區域缺乏提供質子者以幫助其還原。因此本系統中電子傳遞 的關鍵可能是在遠端組胺酸的質子化與去質子化。
圖 1-10 圖示將 1 和 2 與脫輔基肌紅蛋白質包覆後而形成的肌紅 蛋白質三聚體[15]。
1-8 研究目的
經由文獻膫解肌紅蛋白的活性區域遭受改變後,原本不易形成 compound I 的肌紅蛋白會變成具有過氧化酶酵素活性。因此我們嘗試 從肌紅蛋白的結晶結構中比較各胺基酸殘基與血基質鐵原子的距 離,挑選距離較近且可能影響受質進出活性區域的胺基酸殘基,並藉 由定點突變的技術改變3 個胺基酸殘基,利用過氧化酶之受質反應特 性,篩選最好過氧化酶酵素活性的三重突變肌紅蛋白。 經由先前所述,在前人的研究裡已經發現第 64 號 His 位置的迅速氧化作用,尤其是突變成天門冬胺酸(Aspartic acid, Asp)後,可能會 增加對過氧化氫的極性與親合性來改善突變肌紅蛋白的氧化活性。而 第 68 號 Val 的位置對加速使受質到達活性部位有很大的重要性,其 中雙重突變H64D/V68L 可迅速形成 Compound I 後與受質有效的進行 一個電子的傳遞,即有很好的過氧化酶活性。經由結構比較後,我們 推測與血基質鐵原子距離 6.8 Å 的第 107 號 Ile 位置,對影響受質進 出活性部位也可能具有重要的決定因素,因此我們嘗試利用定點飽和 突變第 107 號 Ile 位置的胺基酸,篩選出過氧化酶酵素活性較好的幾 種三重突變肌紅蛋白。利用酵素動力學的方式,我們可以偵測其動力
學常數,再與先前文獻的H64D/V68L 肌紅蛋白做比較。 擁有較好過氧化酶酵素活性的三重突變肌紅蛋白在電子傳遞上 也具有較好的效率。而含有不同金屬的紫質在自然界各種不同物種中 也扮演著不同的角色。因此,當三重突變的肌紅蛋白在活性中心已創 造了最佳的反應環境,再與不同金屬的紫質結合後是否仍然具有相似 的特性亦引起我們很大的興趣去探討。 我們嘗試將本實驗室自行合成的不同金屬紫質與具有過氧化酶 活性的突變型脫輔基肌紅蛋白進行蛋白質輔基重組實驗,並與野生型 肌紅蛋白做比較,並偵測是否仍然具有過氧化酶的活性。
第二章 實驗材料及方法
2-1 實驗材料
2-1-1 化學藥品與材料
Acetic acid (Merck)
40% Acrylamide (GE Healthcare) Agarose (USB)
APS (GE Healthcare)
2,2,-azino-bis(3-ethybenthiazoline 6-sulfonic acid) (ABTS) (Sigma) BactoTM Agar (DIFCO)
Bromophenol blue (USB) Citric acid (Sigma)
Coomassie® Brilliant blue R 250 (Merck) Dimethyl sulfoxide (MP Biomedicals) DEAE sepherose (Bio-Rad)
Dodecyl sulfate sodium salt (Merck)
dNTP Set, 100 mM Solutions (GE Healthcare) Ethylenediamine-tetraacetic acid (Merck) Glycerol (Merck)
Glycine (Merck)
Guanidine hydrochloride (Gdm-HCl) (Sigma) Hydrogen chloride (Merck)
IPTG (GeneMark, Taiwan) Kanamycin sulfate (USB)
LB Broth, Miller (DIFCO) 2-mercaptoethanol (Merck) Methanol (Merck)
N,N’-methylene-bis-acrylamide (Bis-Acrylamide)(Sigma) Potassium chloride (Merck)
Potassium diphosphate (Merck) Potassium phosphate (Merck) Primers (Bio Basic Inc., Taiwan)
Restriction enzymes (New England Biolabs) Sodium azide (Merck)
Sodium chloride (AMRESCO) Sodium hydroxide (Merck) Sodium diphosphate (Merck) Sodium phosphate (Merck) SYBR® Green I (Roche) T4 DNA ligase (Promega) TEMED (GE Healthcare) Tris base (USB)
BCA Protein Assay Reagent and Albumin Standard (PIERCE) BigDye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems)
GFXTM PCR DNA and Gel Band Purification Kit (GE Healthcare) LMW-SDS Marker Kit (GE Healthcare)
Plasmid Miniprep Purification Kit (GeneMark)
Biosystems)
QuickChange Site-Directed Mutagenesis Kit (Stratagene)
2-1-2 緩衝液及溶液配製
50X TAE Buffer:
242 g Tris base、57.1 mL acetic acid、0.5M EDTA 加水至體積為 1 L,調 pH 至 8.5,儲存於室溫。使用時稀釋成 1 X 並調 pH 至 7.5-7.8。
6X DNA loading dye
0.25% bromophenol blue, 30% glycerol,儲存於-20 ℃。
30% Acrylamide Mix:
取40% acrylamide 750 mL、10 g bis-acrylamide 加二次水至體積 為1 L,避光儲存於 4 ℃。
Separation Gel Buffer:
1M Tris pH 8.8,儲存於 4 ℃。
Stacking Buffer:
20% SDS: 20 g SDS,加水至體積為 100 mL,儲存於室溫。 10X Running Buffer: 144 g glycine、30 g Tris-base、10 g SDS,加水至體積為 1 L,儲 存於4 ℃,使用時稀釋成 1 X。 樣品緩衝液 (5X Sample Buffer): 取1 mL 1M Tris-HCl (pH 6.8),加入 0.8 mL 甘油、1.6 mL 10% SDS、0.4 mL 2-mercaptoethanol、0.05% bromophenol blue,加水至體 積為8 mL。
膠片染色液(0.1% Coomassie blue R-250 Stain Solution):
取1 g 的 Coomassie brilliant blue R-250,溶於 400 mL 的甲醇中,
再加入100 mL 的醋酸,加二次水至體積為 1 L。
脫色溶液 I (Destain Solution I):
將甲醇400 mL 與醋酸 100 mL 混合後,加二次水至體積為 1 L。
將甲醇50 mL 與醋酸 70 mL 混合後,加二次水至體積為 1 L。
溶菌緩衝液(Lysis buffer):
Tris-HCl 100mM, KCl 100mM and EDTA 1mM, pH 8.0。
Solubilization buffer Tris-HCl 100mM, pH 8.0,Gdm-Cl 8M。 Disruption buffer Tris-HCl 100mM, pH 8.0 平衡緩衝液(Equilibration buffer): Sodium phosphate 50mM, pH 7.2 LB 培養液
每升加入10 g Bacto-Tryptone、5 g yeast extract 以及 5 g NaCl
IPTG
將IPTG (isopropyl β-D-thiogalactoside) 溶於二次去離子水,再用 0.22 μm 濾膜過濾,儲存於-20 ℃。
每毫升取Tetracycline 20 mg 以 Ethanol 溶解,避光存於–20°C LB-Tet 培養基 每升LB 培養液加入 15 g Agar 經過高溫滅菌後等冷卻至 70 °C 以下再加入1 mL Tetracycline 避光存放於 4°C Ampicillin
(
100 mg/mL) 將Ampicillin 溶於二次去離子水,再用 0.22 μm 濾膜過濾,儲存 於-20 ℃。 Kanamycin(
25 mg/mL) 將250 mg kanamycin sulfate 溶於 10 ml 二次去離子水,再用 0.22 μm 濾膜過濾,儲存於-20 ℃。 LB-Kan 培養基 每升LB 培養液加入 15 g Agar 經過高溫滅菌後等冷卻至 70 °C 以下再加入1 mL Kanamycin 存放於 4°C2-1-3 實驗儀器
數 位 照 相 系 統 (Kodak, DC120 Kodak Electrophoresis Documentation and Analysis System 120)
DNA定序儀 (Applied Biosystems, ABI PRISM® 3100 Genetic Analyzer)
震 盪 培 養 箱 (Firstek Scientific, orbital shaking incubator Model-S302R)
桌上型高速離心機 (Beckman, AllegraTM 21R Centrifuge) 高速離心機 (Beckman, Avanti® J-E Centrifuge)
微量離心機 (eppendorf, Centrifuges 5415R)
微量旋轉式真空濃縮機 (SAVANT, Spin Vaccum SPD-111V) 紫外光/可見光光譜儀 (Beckman, DU-7500 spectrophotometer) 超過濾裝置 (Ultrafiltration System, Amicon)
微盤光譜分析儀 (Fusion Universal Microplate Analyzer, Packard) PCR (Applied Biosystems, GeneAmp® PCR System 9700)
2-1-4 菌株與載體
菌株 XL1-Blue 大腸桿菌之一株,購自於Stratagene 公司。 BL21(DE3) 大腸桿菌之一株,購自於 Novagen 公司。 載體 pET-28a(+) 購自於Novagen 公司。2-2 實驗方法
實驗流程設計
Nco I
BamH I
Use PCR to get the hole sperm whale myoglobin gene BamH I
Nco I
Nco I / BamH I digest
Mb gene ligated into pET-28a(+) vector pET-28a(+) Kan Nco I Nco I BamH I BamH I pET-28a(+) Mb gene Kan Transform into E. Coli.
E. Coli strain BL21(DE3)
pET28-Mb
2-2-1 目標基因的建構
從NCBI 中搜尋 sperm whale myoglobin 基因的 DNA 序列,再將
此序列平均分成六段來設計六條長約 90 bp 的單股 DNA 引子 (HMLpET28-whaleMb-1~6 見表 2-1),鄰近的引子之間有 12~18 bp 的 互補重複區以供鄰近的引子黏合。引子的 N 端起始序列上游設計有 NcoI 的限制酶切位,C 端終止序列下游則設計有 BamHI 的限制酶切 位,使基因全長約為479 bp。將這 6 條引子利用具有修補功能的 rTth polymerase 聚合酶以 PCR 方式(表 2-2,2-3)讓這 6 條引子利用鄰近引子
的互補重複區黏合並延長,得到完整的sperm whale myoglobin 雙股
DNA 基因序列。最後在 PCR 產物加入 2 µL 的 6 倍 Loading Buffer 及 2 µL 的 SYBR Green I,利用 2 % 洋菜凝膠(Agarose gel)以電壓 120 伏特進行電泳分析,確認產物。
引子名稱 引子 DNA 序列
HMLpET28-whaleMb-1(F1) 5’ TAT ACC ATg gTT CTg TCT gAA ggT gAA Tgg CAg CTg gTT CTg CAT gTT Tgg gCT AAA gTT gAA gCT gAC gTC gCT ggT CAT ggT CAg gAC 3’
HMLpET28-whaleMb-2(R2) 5’AgC TTC AgT TTT CAg ATg TTT gAA ACg ATC gAA TTT TTC CAg AgT TTC Cgg ATg AgA TTT gAA CAg TCg AAT CAA gAT gTC CTg ACC Atg 3’
HMLpET28-whaleMb-3(F3) 5’ AAA ACT gAA gCT gAA ATg AAA gCT TCT gAA gAT CTg AAA AAA CAT ggT gTT ACC gTg TTA ACT gCC CTA ggT gCT ATC CTT AAg AAA AAA 3’
HMLpET28-whaleMb-4(R4) 5’ gAT gAA TTC CAg gTA TTT gAT Cgg gAT CTT ATg TTT AgT AgC ATg CgA TTg CgC AAg Cgg TTT gAg CTC AgC TTC ATg ATg CCC TTT TTT CTT Aag 3’
HMLpET28-whaleMb-5(F5) 5’ AAA TAC CTg gAA TTC ATC TCT gAA gCg ATC ATC CAT gTT CTg CAT TCT AgA CAT CCA ggT AAC TTC ggT gCT gAC gCT CAg ggT gCT ATg 3’
HMLpET28-whaleMb-6(R6) 5’ Cgg gAT CCT TAA CCC Tgg TAA CCC AgT TCT TTg TAC TTA gCA gCg ATA TCT TTA Cgg AAC AgC TCg AgA gCT TTg TTC ATA gCA CCC Tg 3’
HMLpET28-whaleMb-N1 5’ TAT ACC ATg gTT CTg TCT 3’ HMLpET28-whaleMb-C1 5’ Cgg gAT CCT TAA CCC Tg 3’
反應物 體積(單位 µL) Each Primer (100 μM/μl) 1 dNTP (10mM) 4 Mg(OAc)2(1.5 mM ) 3 3x3 XL rTth buffer 15 DDW 21 rTth polymerase 1
Total reaction volume 50
表2-2 rTth polymerase kit 的 PCR 反應之組成。 部分 循環次數 溫度 時間 1 1 94℃ 1 分 94℃ 1 分 55℃ 2 分 2 25 72℃ 2 分 3 1 72℃ 5 分 4 1 4℃ ∞ 表2-3 PCR program。
2-2-2 重組質體的建構
利用在N端及C端設計NcoI及BamHI限制酶切位的mb基因將基因嵌入pET-28a(+)表現載體 (expression vector)中。將載體與欲殖入之 DNA分別利用限制酶作用 3-4 小時,於反應中加入 2 µL的 6 倍 Loading Buffer 及 2 µL的SYBR Green I,利用 1.2 % 洋菜凝膠 (Agarose gel)以電壓 120 伏特進行電泳分析,把經過限制酶作用後之 正確大小的載體與欲接入之DNA質帶切下,以Gel Band Purification Kit(GE Healthcare) 純化凝膠中之DNA,以一定比例(insert:vector=5:
1)混合,將樣品以微量旋轉式真空濃縮機抽乾後,利用接合酵素(T4
DNA ligase)於 16 ℃下作用 16 小時。
2-2-2-1 質體之轉化作用
將XL1-Blue 的 competent cell 拿出至於冰上,將經接合酵素反應
之樣品與之混合,冰浴20 分鐘,於 42 ℃水浴中反應 1 分鐘,隨即置
入冰中1 分鐘之後,將樣品全部置入 1 mL LB 培養液中,於 37℃振
盪培養(200 rpm)1 小時,取 200 μL 塗於含有 kanamycin (25 μg/ml)的 LB 培養皿上,在 37 ℃培養 12-16 小時。挑單一菌落至 3 mL LB/kna 培養液中,37 ℃振盪培養(200 rpm) 16 小時,利用 Plasmid Prep Kit (GeneMark),將建構好之質體抽出,並用原先作接合之限制酶 NcoI
與 BamHI,確認插入之 DNA 片段是否有接入載體之中。最後得到肌
紅蛋白重組質體pET28-Mb。確認無誤的重組質體則再同上述步驟轉
形至 E.coli BL21 (DE3)中表現,挑單一菌落至 3 ml LB/kna 培養液 中,37 ℃振盪培養(200 rpm) 16 小時後,取樣品 500 μl 與 500 μl 的 50 % (v/v) glycerol 混合後冰存於-80℃冰箱作為 stock。
2-2-3 定點突變與飽和定點突變實驗
定點突變實驗策略(Site-Directed mutagenesis strategies)
引子設計 ( primer design ) QuickChange PCR 膠體電泳確認 PCR 產物 DpnI 去除母股 DNA 轉入大腸桿菌 XL1-Blue 挑單一菌落,抽取重組質體DNADNA 膠體電泳
利用設計的限制酵素截切( Mapping )做確認
DNA 定序確認突變序列
設計3 種含有靜默突變(Silent Mutation)及欲突變的 3 個胺基酸殘 基His 64, Val 68, Ile107 位置的定點突變互補引子(Primer) (表 2-4)。利 用 Stratagene (Merck 代理)的 QuickChange Site-Directed Mutagenesis Kit 依序進行突變實驗。以 pET28-Mb 為模板,加入 Pfu polymerase 聚合酶以PCR 方式(表 2-5, 2-6)放大而得到 DNA 產物,利用 DpnI 限
制酶截切甲基化DNA 之特性,去除母股,留下含有定點突變之突變
質體。將產物利用 2-2-2-1 節質體之轉化作用所敘述之方法直接轉形 至 E.coli XL1-Blue 中,利用 Plasmid Prep Kit (GeneMark),將定點突
變之質體抽出後,利用在 primer 上設計的靜默突變,以特定限制酵 素截切確認為突變株後再以 DNA 定序確認 DNA 的序列。以此方式 依 序 建 構 pET28-H64D Mb , pET28-H64D/V68L Mb 及 pET28-H64D/V68L/I107X Mb 突變株。 將突變處設計在 primer 上,利用 PCR 放大可直接得到突變點。 突變處在 primer 上前後約各需 3~5 個胺基酸距離,以利黏合反應
(Annealing)。由於飽和突變實驗是將欲突變處突變成其它 19 種胺基 酸,所以primer 的突變處設計為 NNN,N 代表 A、T、C、G 四種鹼 基,因此 primer 包含了 64 種可能性,即可轉譯(translate)出 20 種不 同的胺基酸。在 primer 上設計靜默突變,可利用設計的特定限制酵 素截切以確認是否為突變株。定序確認後的質體 引子名稱 引子 DNA 序列 HMLwhaleMb-H64D-1 E M K A S E D L K K D G
5’ gAA ATg AAg gCT TCT gAA gAT CTg AAA AAA gAT ggT g 3’
HMLwhaleMb-H64D-2 5’ C ACC ATC TTT TTT CAg ATC TTC AgA AgC CTT CAT TTC 3’ HMLwhaleMb-V68X-1 V T L T A L G A I
5’ gTT ACC (g/T/A)(C/T)(A/T) TTA ACT gCC CTA ggg gCT ATC 3’
Bgl I
HMLwhaleMb-V68X-2 5’ gAT AgC CCC TAg ggC AgT TAA (T/A)(G/A)(C/A/T) ggT AAC 3’ HMLwhaleMb-I107X-1 K Y L E F S E A
5’ C AAA TAC CTg gAg TTC NNN TCT gAA gCg 3’
HMLwhaleMb-I107X-2 5’ CgC TTC AgA NNN GAA CTC CAg gTA TTT g 3’
反應物 體積(單位 µL) Template 1 Primer 1 (10 μM/μl) 1 Primer 2 (10μM/μl) 1 10X Pfu Buffer 5 dNTP (10mM) 4 DDW 37 Pfu polymerase 1
Total reaction volume 50
表2-5 Quick Change Site-Directed Mutagenesis 聚合酶連鎖反應組 成。 部分 循環次數 溫度 時間 1 1 95℃ 2 分 95℃ 30 秒 Primer Tm – 5~8℃ 30 秒 2 18 72℃ 12 分 3 1 72℃ 10 分 4 1 4℃ ∞
表2-6 Quick Change Site-Directed Mutagenesis Kit 使用之聚合酶 連鎖反應放大程式。
2-2-4 脫輔基(apo-from)蛋白質的表現
取stock 菌養於 25 mL LB/kna 培養液中,於 37 ℃振盪培養(200 rpm)16 小時。以 1:40 稀釋轉養至 1 L 的新鮮 LB 培養液中,於 37 ℃
振盪培養(200 rpm) 3~4 小時,直到 OD600 = 0.5~1。加入 IPTG (final concentration 0.4mM),繼續在 37 ℃振盪培養(200 rpm)5 小時。於 4 ℃ 下,轉速 8000 rpm,離心 30 分鐘,倒掉上清液,取菌液沉澱物。最 後加入20 mL 溶菌緩衝液(Lysis buffer)回溶菌體。
2-2-5 包涵體(inclusion body)粗萃液之製備
用溶菌緩衝液(Lysis buffer)把菌液沉澱物回溶(每升的菌液用 20 mL 緩衝液回溶),置於冰上 1 小時之後,進行打破細胞的步驟 (Sonication)。將溶液置於冰上,設定超音波震碎儀能量至 30 %,以 震盪 2 秒,休息 1 秒方式,共震盪 30 秒,重複 6 個循環。將溶液於 4 ℃下,轉速 12,000 rpm,離心 30 分鐘,。倒去上清液,將沉澱物加 入20 mL 溶菌緩衝液(Lysis buffer)回溶後,置於冰上 30 分鐘,繼續於 4 ℃下,轉速 8,000 rpm 離心 20 分鐘,回溶再離心的動作重複 4 次。2-2-6 包涵體(inclusion body)的變性與再摺疊
最後一次離心的沉澱物以20 mL 的 disruption buffer 回溶,置於冰上 1 小時。之後進行打破包涵體的步驟 (Sonication),將溶液置於
冰上,設定超音波震碎儀能量至 30 %,以震盪 2 秒,休息 1 秒的方
式,共震盪 30 秒,重複 3 個循環。於樣品中緩慢加入 20 mL 的
solubilization buffer 稀釋 Gdm-HCl 的濃度。對 solubilization buffer 1L 透析5-6 小時,再對 equilibration buffer 2L 透析 3-4 小時 2 次。將溶
液於4℃下離心 30 分鐘,轉速 12000 rpm。取上清液。
2-2-7 脫輔基(apo-form)蛋白質的純化
利用離子交換管柱層析(DEAE)純化此蛋白,樣品以 BCA assay Kit 估計其蛋白質的總量,決定所要填充離子交換樹脂之體積。以 5 倍體積的 equilibration buffer 平衡管柱中的樹脂,慢慢注入樣品,收 集流出物(Flow-Through),利用試管每 5 mL 收集一管。
2-2-8 蛋白質分子量及純度分析
SDS-PAGE 是一種最常使用來分析蛋白質純度與定量的方法, 尤 其 是 在 進 行 蛋 白 質 純 化 時 , 於 膠 體(Polyacrylamide gel)中加入sodium dodecyl sulfate (SDS)用來使蛋白質變性(denature),SDS 會與 蛋白質結合使摺疊構形蛋白質拉成直鏈狀之一級結構,此複合體均帶
負電,所以在膠體中移動速率僅與蛋白質之分子量有關。以 20%
separating gel 及 4% stacking gel 的 SDS-PAGE(表 2-4)分析純化後之蛋 白質樣品。 組成物 20%的體積 ddH2O 2 ml 30% acrylamide mix 6.6 ml 1.5 M Tris (pH8.8) 2.5 ml 20% SDS 50 μl 10% ammonium persulfate 200 μl TEMED 10 μl 組成物 4%的體積 ddH2O 3 ml 30% acrylamide mix 0.67 ml 1 M Tris (pH6.8) 1.25 ml 20% SDS 50 μl 10% ammonium persulfate 50 μl TEMED 10 μl
組成份。 將經由離子交換管柱層析所得之每管樣品,取適量樣品與樣品緩 衝液(5X Sample Buffer)以 1:4 比例混合,於 95℃水浴加熱 10 分鐘 後,置入樣品槽內,以 90 伏特電壓進行電泳分析,當染劑(dye)跑至 下層膠片時,將電壓調至 120 伏特,直至染劑跑到膠片底下或還未完 全跑掉時即可關掉,取下膠片置於膠片染色液(Stain Buffer)中,染色 (含 Coomassie brilliant Blue R-250)1 小時後,用脫色溶液 I (Destain Buffer I)退染 30 分鐘,再用脫色溶液 II (Destain Buffer II)退染至隔天。
2-2-9 蛋白質輔基重組實驗
將經由離子交換管柱層析所得之蛋白質以BCA assay Kit 偵測蛋
白質濃度,以0.5 M 氫氧化鈉溶液將蛋白質溶液的 pH 值調至 12。取
莫耳濃度為蛋白質濃度2 倍的紫質溶於 250 μL 的 pyridine,加入 250 μL 的 equilibration buffer 並以超音波震盪 10 分鐘,將紫質溶液與 pH 12 的蛋白質溶液混合,於 4 ℃下,磁石攪拌(stir)10 分鐘,再以 0.5 M 鹽酸調整pH 值至 6.8,於 4 ℃下,磁石攪拌 6 小時。對 1 L 100 mM
磷酸鉀緩衝液pH 6.8 透析 2-3 小時,共 3 次。樣品以 0.22 μm 的濾膜 (filter)。利用 desalting column 純化樣品,收集流出物(Flow-Through), 利用eppendrof 以每 1 ml 收集一管。
2-2-10 酵素最適化條件之研究
2-2-10-1 最適反應環境之酸鹼值測定
配製不同pH值的受質反應溶液,將 1 mM ABTS及 7 mM H2O2溶
於不同pH值且適合於其酸鹼值範圍的各種鹽類溶液中,酸性範圍(pH 4-6)為檸檬酸(Citric acid)溶液,中性範圍(pH 6-8)為磷酸鈉(Sodium Phosphate) 溶 液 , 鹼 性 範 圍 (pH 8-10) 為 三 羥 甲 基 氨 基 甲 烷 - 鹽 酸 (Tris-HCl)溶液,緩衝液鹽濃度為 50 mM。取 1 ml不同pH值的受質反
應溶液與固定量的酵素溶液5 μl混合測定不同PH值的酵素活性。以不
同pH值對酵素活性作圖,可得酵素進行反應之最適作用pH值。
將固定量的酵素溶液分別於 4,25,37,42,50,60,70 與 80 ℃下作用30 分鐘後,置於室溫 20 分鐘。各取 5 μL的酵素溶液與 1 mL 的 1 mM ABTS及 7 mM H2O2,於 50 mM 磷酸鈉緩衝液pH 7.0 中混 合進行反應,測定酵素活性。以溫度對酵素活性作圖,可得到酵素的 耐熱性。
2-2-11 酵素動力學參數測定
配製不同ABTS濃度的受質液(0.05-5 mM)溶於含 7 mM H2O2的 50 mM 磷酸鈉緩衝液pH 7.0 中。取 1 mL不同ABTS濃度的受質液與 5 μL 適當量稀釋的酵素溶液混合進行反應,於室溫偵測 730 nm 吸光 值每隔5 秒測一次共偵測 2 分鐘,重複 3 次以上,取其平均值。取一 固定時間的吸收值除以ABTS游離基產物在 730nm的莫耳消光係數(ε = 1.4 × 104 M-1 cm-1)[8],最後換算酵素催化受質的反應速率單位為 (mM/min),以受質濃度對酵素反應速率作圖可得受質飽和曲線圖。 將受質濃度與反應速率均換算成倒數值,以Lineweaver-Burk plot雙倒 數作圖,加上趨勢線及其直線公式後可求得該直線的X截距及Y截 距 , 依 此 可 求 得 酵 素 催 化ABTS 之 apparent Km(K’m) 及 apparentVmax(V’max)值(K’m = -1/X截距;V’max = 1/Y截距)。 經由上述方式求得ABTS之K’m後,將ABTS濃度固定於大於K’m 之濃度,改變H2O2濃度,測定H2O2受質之K’m 值及V’max 值。配製 不同H2O2濃度的受質液(0.1-15 mM)溶於含 1 mM ABTS的 50 mM 磷 酸鈉緩衝液pH 7.0 中。取 1 mL不同H2O2濃度的受質液與 5 μL 適當量 稀釋的酵素溶液混合進行反應,於室溫偵測供氫體ABTS氧化產物 730 nm 吸光值,以H2O2濃度對酵素反應速率作圖可得H2O2受質飽和 曲線圖,並以Lineweaver-Burk plot雙倒數法求得酵素催化ABTS氧化 之H2O2受質K’m及V’max值。
第三章 結果與討論
3-1 目標基因質體之建構
可轉譯出完整的抹香鯨肌紅蛋白(sperm whale myoglobin)的 mb
基因,在利用6 條單股 DNA 引子以 PCR 方式黏合、延長並放大後,
可以得到一條全長 479 個鹼基對的雙股 DNA。經過 2% 洋菜凝膠
(Agarose gel) 電泳後可在約 500 bp 處看到 mb 的 DNA 質帶。
經過多次的截切與接合反應後,我們成功的將 mb 基因接入 pET28a(+)這個表現載體中,並經由限制酵素截切( Mapping )確認後 進行 DNA 定序的反應,可以確認我們成功的建構了 pET28-mb 這個 質體。
3-2 篩選目標
利用定點突變的技術來對蛋白質特定胺基酸殘基做改變以觀察 蛋白質的結構及活性變化,已是目前相當普遍的方法。本實驗室對這 項技術亦相當純熟。而先前所述肌紅蛋白中 His64 位置在突變成天門冬胺酸(Asp)後可能會增加對過氧化氫的極性與親合性來改善突變型
肌紅蛋白的氧化活性,而第 68 號 Val 的位置對加速受質到達活性部
位有很大的重要性。因此我們先以含有野生型抹香鯨肌紅蛋白(sperm whale myoglobin) mb 基因的質體 pET28-mb 為模板(template),利用定
點突變的技術得到改變2 個胺基酸殘基位置的 pET28-H64D/V68L mb 質體,即文獻中過氧化酶活性最好的雙重突變肌紅蛋白。並以此為模 板,挑選另一個可能會影響突變型肌紅蛋白過氧化酶活性的胺基酸殘 基。 經由野生型抹香鯨肌紅蛋白結晶結構來比對每個胺基酸殘基與 血基質鐵原子的最近距離後(表 3-1)。我們推測距離 6.76 Å 的第 107 號 Ile 位置的殘基對影響受質進出反應的活性部位也可能具有重要 的決定因素。因此我們嘗試飽和突變第 107 號 Ile 位置的胺基酸,將 它換成其它19 種的胺基酸,再來比較過氧化酶活性。 位置 殘基 最近原子 距離 位置 殘基 最近原子 距離 位置 殘基 最近原子 距離
1 Val CG1 -25.02 52 Glu OE2 20.47 103 Tyr OH -12.87
2 Leu CD1 -21.05 53 Ala CB 23.68 104 Leu CD2 -6.54
3 Ser OG -28.16 54 Glu OE1 21.17 105 Glu OE2 -17.46
4 Glu OE2 -27.26 55 Met CE 16.06 106 Phe CD2 12.84
5 Gly CA -28.67 56 Lys NZ 21.25 107 Ile CD1 6.76
7 Trp CH2 -18.96 58 Ser OG 15.55 109 Glu OE2 17.08
8 Gln NE2 29.89 59 Glu OE2 18.92 110 Ala CB 12.8
9 Leu CD2 27.28 60 Asp OD2 12.73 111 Ile CD1 10.53
10 Val CG1 18.79 61 Leu CD1 10.79 112 Ile CD1 17.33
11 Leu CD2 19.64 62 Lys NZ 19.57 113 His NE2 20.89
12 His ND1 27.01 63 Lys NZ 15.55 114 Val CG2 16.19
13 Val CG2 20.39 64 His CD2 8.48 115 Leu CD2 17.25
14 Trp CH2 13.11 65 Gly CA 9.56 116 His NE2 25.89
15 Ala CB 22.86 66 Val CG1 13.08 117 Ser OG 21.27
16 Lys NZ 27.49 67 Thr CG2 7.15 118 Arg NH2 19.52
17 Val CG1 15.96 68 Val CG2 4.99 119 His NE2 20.7
18 Glu OE2 16.25 69 Leu CD2 10.32 120 Pro CD 26.38
19 Ala CB 22.75 70 Thr OG1 12.44 121 Gly CA 29.08
20 Asp OD1 19.63 71 Ala CB 7.89 122 Asp OD2 27.24
21 Val CG1 13.9 72 Leu CD1 10.4 123 Phe CZ 20.28
22 Ala CB 16.56 73 Gly CA 13.49 124 Gly CA 27.54
23 Gly CA 17.5 74 Ala CB 13.52 125 Ala CB 28.7
24 His ND1 17.93 75 Ile CD1 -10.1 126 Asp OD1 28.5
25 Gly CA 12.16 76 Leu CD2 -15.02 127 Ala CB 24.54
26 Gln OE1 17.37 77 Lys NZ 18.69 128 Gln OE1 23.08
27 Asp OD1 17.94 78 Lys NZ -20.08 129 Gly CA 23.7
28 Ile CG2 10.73 79 Lys NZ -25.64 130 Ala CB 23.2
29 Leu CD2 7.64 80 Gly CA -19.4 131 Met CE 17.6
30 Ile CG1 15.2 81 His ND1 -22.87 132 Asn OD1 18.66
31 Arg NH1 18.98 82 His ND1 -15.76 133 Lys NZ -26.19
32 Leu CD2 8.35 83 Glu OE1` -20 134 Ala CB -17.52
33 Phe CZ 9.33 84 Ala CB -17.44 135 Leu CD1 14.03
34 Lys NZ 21.19 85 Glu OE2 -16.73 136 Glu OE2 -19.09
35 Ser OG 17.21 86 Leu CD1 -12.72 137 Leu CD1 -18.88
36 His NE2 15.61 87 Lys NZ -17.63 138 Phe CZ -8.17
37 Pro CG 17.72 88 Pro CB -11.24 139 Arg NH2 -14.07
38 Glu OE1 15.62 89 Leu CD1 -6.2 140 Lys NZ -21.4
39 Thr CG2 10.72 90 Ala CB -10.76 141 Asp OD2 -17.03
40 Leu CD1 12.3 91 Gln NE2 -13.71 142 Ile CD1 -9.6
41 Glu OE1 16.17 92 Ser OG -6.79 143 Ala CB -15.44
42 Lys NZ -11.36 93 His NE2 -2.18 144 Ala CB -18.04
44 Asp OD2 13.14 95 Thr OG1 -12.5 146 Tyr OH -8.47
45 Arg NH1 10.88 96 Lys NZ -10.07 147 Lys NZ -23.24
46 Phe CE2 8.94 97 His NE2 -5.69 148 Glu OE1 -19.11
47 Lys NZ 13.86 98 Lys NZ -14.52 149 Leu CD1 -14.36
48 His ND1 16.68 99 Ile CD1 -6.47 150 Gly CA -18.92
49 Leu CD1 14.93 100 Pro CD -10.07 151 Tyr OH -15.42
50 Lys NZ 24.72 101 Ile CD1 -11.69 152 Gln OE1 -16.45
51 Thr OG1 22.52 102 Lys NZ -18.05 153 Gly CA -16.09
表 3-1 野生型抹香鯨肌紅蛋白中各胺基酸殘基與血基質鐵原子 最近的原子及相距的距離。(註 1:最近的原子第一個為原子種類,之 後代表為殘基中的第幾號原子;註2:距離單位為 Å,負號代表為位 於血基質平面的近端組胺酸那一側)。
3-2-1 細胞粗萃液的活性篩選
以含有雙重突變H64D/V68L mb 基因的質體 pET28-H64D/V68L mb 模板(template),利用飽合突變的技術,我們得到將第 107 號 Ile 胺基酸殘基改變成其它 19 種胺基酸的三重突變株。經由 DNA 定序 確 認 後 , 我 們 將 包 含 雙 重 突 變 在 內 的 20 種 胺 基 酸 pET28-H64D/V68L/I107X mb 轉形(transform)至可以表現出完整肌紅 蛋白的 E.coli BL21 (DE3)中表現。為了達到可以快速的篩選飽和突變實驗中哪一株三重突變株具 有較好的過氧化酶活性,因此我們採用小量的培養包含雙重突變株在 內的20 種胺基酸的突變株於 3 mL LB 培養液中,並在 15 mL 試管中 於37℃ 震盪培養 12~16 小時。收集 1.5 mL 菌液,以 13200 rpm 離 心 1 分鐘,到掉上清液,加入 300μL 的溶菌緩衝液以回溶沉澱物。 利用溫和的破菌方法來打破細胞,隨即將樣品丟入液態氮中急速冷凍 約 1 分鐘,再迅速丟入 42℃水浴中約 2 分鐘,如此重複 3 次。最後 以 12,000 rpm 離心 1 分鐘,取上清液即為用來做活性比較的細胞粗 萃液。 經過蛋白質定量後,取含相同蛋白質量的細胞粗萃液與 90 μL 含1 mM ABTS、5 mM H2O2的50 mM 磷酸鈉緩衝液pH 7.0 在 96-well 的微量分析盤(microwell plate)中反應,並偵測ABTS氧化產物 405 nm 的吸光值以比較彼此間的過氧化酶活性大小,結果如圖3-1 所示。 結果顯示將第 107 號 Ile 胺基酸突變成 Met、Leu、Phe、Val、 Ala 這 5 種胺基酸的三重突變株的過氧化酶活性較優於文獻中的 H64D/V68L 的雙重突變株。因此我們將個別純化這幾種胺基酸的肌 紅蛋白再與H64D/V68L 的雙重突變進行過氧化酶活性的比較。
0 0.5 1 1.5 2 A V L M P F W G S T Y C Q D E H K R N I H64D/V68L/I107X Mb A bs . 405 nm 圖 3-1 含相同蛋白質量的三重突變肌紅蛋白H64D/V68L/I107X Mb細胞粗萃液 10 μL與 90 μL 的 1 mM ABTS、5 mM H2O2 反應後偵 測405 nm的吸光值。
3-3 脫輔基(apo-form)蛋白質表現及純化
由於實驗的需求,未來我們希望的是將突變後的脫輔基肌紅蛋白 與不同金屬的紫質及經過各種官能基修飾過後的紫質做輔基重組,再 研究其各種物理及化學特性。因此,我們參考文獻[16]將實驗採用的 肌紅蛋白表現成包涵體(inclusion body)的方式,再藉由包涵體的變性 與再摺疊獲得脫輔基形式的肌紅蛋白。如此可以擺脫以往利用 MEK 方法將紫質從肌紅蛋白中分離的繁複步驟,從包涵體純化脫輔基肌紅蛋白也可以去除許多雜蛋白,省去利用第二根管柱層析純化肌紅蛋白 的時間。 處理包涵體時由於蛋白質的過度堆疊,因此密度相當的大,所以 在破菌後只需利用低轉速就可將包涵體離心下來。接下來以 lysis buffer 不斷的回溶去除非包涵體的雜蛋白,經過幾次處理即可得到大 部分都是脫輔基肌紅蛋白的包涵體。 本 實 驗 利 用 Gdm-HCl 使 包 涵 體 變 性 , 再 利 用 透 析 去 除 Gdm-HCl 使脫輔基肌紅蛋白慢慢摺疊。透析時為了避免已變性的蛋 白又聚集在一起,因此會利用梯度透析的方法來避免此情況。最後只 需要利用DEAE 離子交換管柱層析收集沖提液,再將每管利用 BCA 蛋白質濃度測量分析後,收集代表含有蛋白質的吸收峰部分。 經 過 純 化 的 脫 輔 基 肌 紅 蛋 白(apo-Mb) 可 以 由 以 下 的 20% SDS-PAGE(圖 3-2 )偵測得知蛋白質的純度,並且利用 UV-Vis 光譜儀 掃描全波長並無發現紫質的410 nm 處的吸收峰,因此可以確認純化 的肌紅蛋白為脫輔基的形式。
M 1 2 3 (kDa) 14.4 30 20.1 97 65 45 圖 3-2 20% 的 SDS-PAGE 分析 Lane M:低分子量的蛋白質
marker。Lane 1:未加入 IPTG 誘導前。Lane 2:IPTG 誘導後經超音
波震盪離心之沉澱回溶。Lane 3:經純化後得到約 17kDa 的肌紅蛋白。
3-4 蛋白質輔基重組
在極性的環境(極酸或鹼)下肌紅蛋白會變性而去摺疊,紫質亦會 由 原 本 在 中 性 pH 值 時 的 高 度 聚 集 (aggregate) 現 象 而 形 成 單 體 (monomer)或雙體(dimer)的形式。利用此特性我們先將脫輔基肌紅蛋 白溶液與紫質溶液的 pH 值均調成 12,混合攪拌後再調回中性的 pH 值,以幫助脫輔基肌紅蛋與紫質進行蛋白質與輔基的重組(cofactorreconstitution)。 一些有機溶劑也會加入以幫助溶解紫質及穩定蛋白質,吡啶 (pyridine) 與 緩 衝 溶 液 等 比 例 的 混 合 可 幫 助 紫 質 更 不 容 易 聚 集 (aggregate)。另外一種有機溶劑二甲基亞石砜(Dimethyl Sulfoxide, DMSO)也被加在蛋白質溶液中以穩定蛋白質,防止用來溶解紫質的 吡啶(pyridine)加入後會造成蛋白質不穩定。 實驗中會加入約高於蛋白質莫耳濃度 2 倍量的紫質以確保每分 子的蛋白質均可與紫質結合,經過調整pH 值及透析去除有機溶劑後 多餘未被嵌入肌紅蛋白的紫質會高度聚集,利用孔徑極小的0.22 μm 濾膜(filter)及去鹽管柱(desalting column)可以將這些多餘未被嵌入肌 紅蛋白的紫質去除。 鐵(Fe3+)和鋅(Zn2+)兩種市面上可買到的不同金屬紫質在經過蛋 白質與輔基的重組實驗後已被成功的嵌入野生型及H64D/V68L雙重 突變型的肌紅蛋白中。經由UV-Vis光譜儀掃描全波長後,由下圖(圖 3-3)可以比較彼此間的不同。
圖3-3 UV/Vis 全光譜掃描圖 含鋅的紫質(Zinc Protoporohyrin IX, ZnPP)及含鐵的紫質(Iron(II) Protoporphyrin IX)即血基質(Heme)分別
與野生型及 H64D/V68L 雙重突變型的脫輔基肌紅蛋白中進行蛋白質 與輔基的重組後與對照的ZnPP 溶於 pH6.8 的 100 mM 磷酸鉀緩衝液 中。 含鐵的紫質即為自然界中常見肌紅蛋白中含的血基質(Heme),文 獻中指出MbH64D/V68L雙重突變肌紅蛋白會有過氧化酶(peroxidase)的活 性[10],因此為了測試蛋白質與輔基的重組實驗是否可行,我們將經 過蛋白質與輔基的重組實驗後血基質(Heme)已成功的被嵌入野生型 及突變型的肌紅蛋白與1 mM ABTS及 2 mM H2O2反應。結果雙重突 變肌紅蛋白MbH64D/V68L如文獻所指具有很高的反應活性,與野生型肌


![圖 1-5 在 chloroperoxidase 中 Glu-183 對形成 Compound I 的可能 功能。 另外,從結構的角度來看肌紅蛋白的 Val 68 位置對受質到達活性 部位有很大的重要性,將其置換成各種不同的胺基酸殘基會大大的改 變其活性區域的形狀與體積[10]。因此以 H64D 為模板,將 Val 68 突 變成各種不同胺基酸的雙重突變已被建構出來以測量其過氧化酶 (peroxidase)與過氧合酶(peroxygenase)酵素活性,在這些雙重突變中, H64D/V68L 突變在形成](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243603.171429/29.892.136.753.163.338/其置換成各種不同胺基酸殘基會大大變其活性區域形狀各種與過氧合.webp)
![圖 1-8 Mb(Fe III OH 2 )-Ru 2+ -BXV 4+ 在經過激發Ru(bpy) 3 後的能階與 選擇性路徑[15]。](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243603.171429/34.892.180.805.362.821/圖1MbFe2Ru2+BXV4+在經過激發Rubpy3後的能階與選擇性路徑1.webp)
![圖 1-10 圖示將 1 和 2 與脫輔基肌紅蛋白質包覆後而形成的肌紅 蛋白質三聚體[15]。](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243603.171429/37.892.169.795.108.691/圖1圖示將與脫輔基肌紅蛋白質包覆後而形成的肌紅蛋白質三聚體1.webp)