群聚變異梯度長度對排序結果的影響
許皓捷
1,2,3)李培芬
1,2)摘 要
排序(ordination)是探討生物群聚與環境之間關係常用的多變數統計方法。本研究以模擬數據及鳥 類群聚資料,探討主成分分析(principal components analysis, PCA)、對應分析(correspondence analysis, CA)、以及降趨對應分析(detrended correspondence analysis , DCA)三種常用的排序法在不同群聚變異梯 度長度下的排序表現。結果發現三種排序法適用的群聚梯度長度不同。DCA運算結果的軸長可作為群 聚梯度長度判斷的參考;梯度軸長在 2 SD以上,且CA因為物種沿環境梯度的非線性分布而產生明顯 拱形效應(arch effect)時,以 DCA的結果較佳;但若樣點在CA的空間未有明顯拱形排列時,則以使用 CA 較適當。軸長在 2 SD以下時,以使用 CA及 PCA較佳, DCA則可能因為降趨(detrending)及重新刻 劃(rescaling)的程序,而嚴重扭曲數據原本蘊含的生態意義。排序方法的選擇應考慮生物群聚之梯度長 度,以獲得合理的結果。
關鍵詞:對應分析、降趨對應分析、梯度長度、排序、主成分分析。
許皓捷、李培芬。 2003 。群聚變異梯度長度對排序結果的影響。台灣林業科學 18(3):201-11 。
Re sea rch paper
Influence of Gradient Length on Community Ordinations
Hau-Jie Shiu,
1,2,3)Pei-Fen Lee
1,2)【Summary】
Ordina tio n is th e c olle ctive t erm for mu ltiv ariable te chn ique s t hat elu cida te the variat ion in co mmun ities a nd dete ct relatio nsh ips bet ween co mmun ity gra die nts and en viro nme ntal fa ctors. We us ed artific ial an d a vian commu nit ies t o inve stiga te the p erformanc e of 3 o rd ina tio n tec h-nique s o f p rin cip al comp one nts an aly sis (PCA), c orresp ond enc e a nalysis (CA), an d d etrend ed co rre spon den ce ana lysis (DCA ) for d iffere nt grad ien t leng ths of commun itie s. Res ult s su gge st tha t ap plic ability of o rdin atio n me thod s va ries acco rdin g to gra dien t le ngth . If the len gths of the ordina tio n axes are gre at er th an a bou t 2 sta nd ard d ev ia tio ns (SD), th e res ults of t he CA may displa y t he a rch effect, an d on e s hould co nsider using t he DCA. Howeve r, if t he c onfiguratio ns of s it e po in ts of t he CA do n ot disp la y th is effe ct , on e sh ou ld us e th e CA . If th e ordina tion
1)國 立 台 灣 大 學 動 物 學 系 , 10 6 臺 北 市 羅 斯 福 路 四 段 一 號 D ep ar tm en t of Zo ology, N a tion al T aiwa n
Un ive rsity. 1 Roo sev elt Rd., Sec. 4, T a ipe i, T aiwan 10 6.
2)目 前 地 址 : 國 立 台 灣 大 學 生 態 學 與 演 化 生 物 學 研 究 所 Pr esent address: In stitut e o f Ecolo gy and Ev
o-lutio nary Biolo gy, Na tion al T aiwan Un iver sit y.
3)通訊作者 Co rre spo nding autho r, e- mail:jay@oik os.zo .nt u.e du. tw
le ngth s a re less tha n ab out 2 SD, the DCA may de stro y t he e cologic ally me anin gfu l in formation b y de tren ding a nd res ca ling , an d th erefore o ne s ho uld co ns id er u sing t he PCA o r CA . Th e leng th of the gradient stron gly in fluenc es the correc tness of the ordin ations ; inve stigat ors sh ould ca refully c hoo se a n a ppropriat e meth od acc ord ing to gra die nt len gth s.
Ke y wor ds : co rres po nd en ce a na lys is , de tren de d co rre sp on de nc e an alys is, g ra dien t le ng th ,
ordin ation, principal co mpo nen ts a nalysis.
Shiu HJ, Lee PF. 2003 . In flue nce o f gradie nt le ngth on c ommu nity ordinatio ns. Taiwan J For Sci
18(3):201-11.
緒 言
依據耐受法則(law of tolerance),生物在環 境梯度上有一特定分布範圍,並在其中的最適 環境有最大的豐富度(abundance)。由於不同物種 在環境梯度上耐受範圍的差異及豐富度的變 化,導致生物群聚(community)的結構(物種組成 及豐富度)沿環境梯度而改變。排序(ordination) 是依據物種組成及豐富度,將取樣點依序排列 的多變數技術之集合名詞(Austin 1985, ter Braak 1995)。排序的目標是使物種組成及豐富度類似 的取樣點彼此排列位置較為接近,而差異較大 的取樣點之間則分隔較遠。利用排序方法,可 以得到生物群聚結構的主要變異梯度軸;以之 與環境因子做相關分析,則可以找出與生物群 聚變異有關的主要環境因子。 群聚生態學家最常使用的排序方法為主成 分分析(principal components analysis, PCA)與其 衍生之重複分析(redundancy analysis, RDA),以 及對應分析(correspondence analy sis, CA; also named as reciprocal averaging, RA)及其衍生之降 趨對應分析(detrended correspondence analysis, DCA)與典型對應分析(canonical correspondence analysis , CCA) (Gauch 1982, t er Braak 1986, ter Braak and Prentice 1988, ter Braak 1995)。每一種 排序方法各有其適用場合與優缺點。 PCA 適用 於線性資料;在豐富度沿環境梯度呈現高斯分 布(Gaussian dist ribution)的生物群聚資料上, PCA常產生嚴重的馬蹄形效應(hors eshoe effect) (Gauch 1982, Legendre and Legendre 1998),即樣 點在前二軸的空間分布上產生馬蹄形狀排列, 並且有軸端反轉與壓縮的現象,而扭曲真實的群聚梯度。CA在高斯分布特性的生物資料上, 避免了 PCA 軸端反轉的問題,但是仍然存在軸 端壓縮與多餘第二軸的拱形效應(arch effect)現象 (Gauch 1982, ter Braak 1995, Legendre and Legendre 1998)。 DCA 則以非線性重新刻劃(nonlinear rescaling)及降趨(detrending)的方式修正 CA 的軸 端壓縮,以及多餘第二軸的缺點,而使生物群 聚的變異梯度能正確反映在排序軸上(Hill and Gauch 1980)。至於 RDA 及 CCA ,則分別是 PCA 及 CA 的直接梯度分析形式,即在分析過 程中以樣點的環境因子測量值對排序分數作修 正,以使環境因子對排序結果有最佳的解釋。 大致而言, PCA及其衍生法適用於線性反 應模式,當物種的豐富度隨潛在環境變數的數 值線性遞增或遞減時,即適合用 PC A 進行排 序。CA及其衍生方法則適合物種豐富度沿環境 梯度呈鐘形反應模式的資料。空間尺度影響物 種豐富度沿環境梯度所呈現出的反應模式。在 較小的空間尺度之下,其環境梯度亦較窄,物 種豐富度常呈線性反應模式;在較大的空間尺 度之下,物種豐富度多呈現鐘形反應模式。因 此,隨著空間尺度不同,所採用的排序方法亦 應有所不同。過去對於 PCA 在大尺度下之馬蹄 形效應已有許多討論(e.g., Gauch 1982, Pielou 1984),但是 CA 及其相關排序法在較小梯度下 的表現,則缺乏深入探討;同時,空間尺度影 響排序結果仍是概念性的討論,精確的定量研 究亟待發展。 綜觀國內的群聚生態學研究,早期很少以 排序為分析工具,直到Su (1987)將DCA在國內
推廣後,以排序技術探討生物群聚與環境之間 關係的研究才明顯增多。 Su 在該文中介紹了 PCA 、 CA 、 DCA 及其它排序方法,並說明了 PCA及CA的缺點,同時認為DCA修正了CA拱 形效應之缺陷,因此「不論梯度數目之多寡及 長度大小,目前以 DCA 之使用最為普遍」。其 後國內的研究者在生物群聚的排序分析上,大 量採用 DCA 。我們整理了 1988-2001 年在「中 華林學季刊」及「台灣林業科學」發表的文章, 以及國內各大學相關研究所有採用排序分析的 碩博士論文,共計 67篇文獻,發現其中 73%的 研究採用 DCA ,另有 16% 使用 CCA , 10% 使 用 PCA,而使用 CA的只有 1%,沒有任何研究 採用RDA。我們另外檢索了Biological Abstracts 資料庫的Ecology, Forest Ecology and Management,
Plant Ecology 三種期刊於 1988-2001 年刊登的生 物群聚排序分析之文獻計94篇,卻發現DCA的 使用僅有 27%,而有 40%使用CCA ,11%使用 PCA , 13% 使用 CA ,另有 9% 使用 RDA 。綜 合上述,國內的研究者明顯偏好使用 DCA;對 國內的群聚生態學研究而言,各種排序方法的 優缺點,以及其使用時機或適用條件,顯然有 重新討論的必要。 本 研 究 以 模 擬 群 聚 及 鳥 類 資 料 , 探 討 PCA 、 CA 、 DCA 在各種梯度長度下的排序表 現。我們同時討論這些排序方法在不同梯度長 度下的適用性,並歸納排序方法選擇之原則。
材 料 與 方 法
一、模擬群聚 假設物種 k 的豐富度沿環境梯度分布遵循 常態分布機率密度函數,則物種 k 在環境梯度 上任一取樣點 i 的豐富度估值 Eyki可以下式求得 (ter Braak 1995): _________________ Eyki= ck exp -0.5(xi-uk)2 tk2 其中ck是物種k的最大豐富度,出現在最適 環境值 uk的位置;xi是樣點 i 在環境梯度軸上的 分數(位置);uk是物種 k 最適的環境梯度位置之 分數;tk是物種 k 耐受力的介量,亦即標準差σ (Fig. 1)。 若群聚具有兩個變異梯度,且任一物種的 豐富度沿任一環境梯度之分布均遵循常態分布 機率密度函數,則物種k在二維環境梯度上的任 一取樣點 i 的豐富度可以下式求得: Eyki= ck exp -0.5[(xi1-uk1)2+(x i2-uk2) 2] tk2 其中 xi 1及 xi 2分別是樣點 i 在兩個環境梯度 軸上的分數;uk1及 uk2是物種 k 分別在兩個環境 梯度上的最適位置之分數。 高斯曲線兩端為趨近但不相交於 0 的漸近 線,因此取用uk ± 3 tk為模擬物種在環境梯度上 的耐受範圍,其豐富度合計約佔全部的99.74%。 為量化梯度長度,將此耐受範圍定義為一個T (= 6 tk),以 T 值度量梯度的長度。 我們將群聚梯度模型分為 ty pe I 和 ty pe II 兩種, t yp e I model 包含梯度的兩個軸端(Fig. 2a),亦即物種在環境梯度上的最適值 u 均位於 模擬的梯度長度範圍內。 Ty pe I model 存在於 海岸到山頂、島嶼南北兩端之間或草原到成熟 林等,因地理或環境限制的梯度中。 T y p e II model 則不包含梯度的兩個軸端(Fig. 2b),亦即 物種最適值 u 允許位於模擬的梯度長度範圍之 外。Type II model存在於一個長的環境梯度中, 任意截取一段不包含軸端的梯度段落,例如玉 山地區某一海拔 2,000-2 ,500 m 的成熟林( 在 2,000 m 以下及 2,500 m 以上仍為成熟林)。 Fig. 1. Gaussian respons e curve displaying a unimodal relation between the abundance value of a s pecies and an environmental variable. (u = optimum; t = tolerance; c = maximum) (Redrawn from ter Braak and Prentice 1988).一維群聚梯度模型設定2, 1.5, 1, 0.75, 0.625, 0.5, and 0.25 T 七種梯度長度。在模擬的環境梯 度上,每一物種的最大豐富度c相等;每一物種 在環境梯度上的最適值u成等距排列;每一物種 的耐受力 t 相等。 二維群聚梯度模型的第一軸長度設定為第 二軸的兩倍,以模擬包含一個主要變異梯度軸 及一個次要變異梯度軸的生物群聚。物種在兩 環境梯度軸上的最大豐富度 c 相等,耐受力 t 亦 相等;在任一環境梯度上,物種的最適值u成等 距排列;物種之間排列的順序,在兩個環境梯 度軸上分別以隨機方式產生。共建立四組資料 矩陣,包括(1) t y pe I model ,梯度長度 2 及 1 T;(2) ty pe II model ,梯度長度 2 及 1 T ;(3) type II model ,梯度長度 1 及 0.5 T;(4) type II model ,梯度長度 0.5 及 0.25 T 。 二、真實群聚 本研究以鳥類調查資料,進行排序方法適 用性之探討。由於海拔是影響台灣山區鳥類群 聚變異的主要環境因子(Lee 1995, Shiu 1995), 因此以海拔高度差作為選擇群聚梯度長度之主 要依據。我們選取數個不同梯度長度的鳥類群 聚,包括:(1)海拔 400-3,700 m 鳥類群聚,共 170 個鳥類取樣點,廣泛分布於台灣北中南山 區,棲地類型包括草地、灌叢、先鋒闊葉林、 成熟闊葉林、針闊葉混生林及針葉林;(2)大武 山鳥類群聚,位於北大武山海拔 1,600- 2,600 m ,包括闊葉林、針闊葉混生林及針葉林;(3) 玉山鳥類群聚,位於玉山沙里仙溪溪谷及楠梓 仙溪溪谷,海拔 1,800-2,500 m ,包括闊葉林、 針闊葉混生林及針葉林;(4)拉拉山鳥類群聚, 海拔 800-1,200 m,棲地類型包括天然闊葉林及 林下小徑木經擇伐處理之天然林;(5)丹大山鳥 類群聚,海拔 2,900-3,000 m ,二葉松林及冷杉 林。其中拉拉山鳥類群聚資料取自 Fang (1996) 之野外調查數據,其餘鳥類資料均來自 Sh iu (1995)及 Shiu 1998-2000 未發表之資料。 上述野外調查均於鳥類繁殖季以定點計數 法(point count)進行。排序以 PC-ORD ver. 4.20 (M cCune and M efford 1999)進行。為避免優勢 種過度主導排序結果,分析前,鳥類密度資料 先以平方根做數值轉換。不同排序方法之間的 結果以相關性檢測比較之,方法是先將每一排 序軸的最小值訂為0,前兩軸之中數值最大者訂 為 1 ,將所有樣點排序分數轉換為 0 與 1之間的 相對值,再以 Bray -Curtis measure (Krebs 1999) 計算各樣點在前二軸排序分數的相異度矩陣, 然後用 M ant el t est 進行矩陣間之相關性檢測 (Douglas and Endler 1982)。
結 果
一、模擬群聚 在模擬梯度的探討中,排序的正確性採用 樣點在排序軸的分數與其環境梯度值之相關係 數評估之;須注意的是,樣點彼此之間的關 係,係表現在排序軸上的相對位置與距離,相 關係數的正負值對排序正確性並沒有影響,因 此在本研究的結果及討論中均予以省略。在模 擬的維度上,相關係數愈高,表示排序愈正 Fig. 2. Community gradient models. (a) TypeI model, including both terminals of the gradient; (b) type II model, excluding the ter-minals of the gradient. The curves shown have equally s paced optima, equal tolerances, and equal maximums . A Gauss ian res ponse curve is defined as 1 T. Gradient length of an artificial community is measured by the number of Ts. Both communities have a length of 2 T.
確;相對的,在非模擬的次維度上,相關係數 則應該愈低愈好。另外, PCA 及 CA 的特徵值 (eigenv alue)及解釋變異量,以及以標準偏差 (standard deviation, SD)度量的 DCA 軸長,亦為 評估排序表現的指標之一。模擬維度與非模擬 的次維度之間的解釋變異量或軸長的差距愈 大,表示排序結果愈理想,亦即數據結構變異 大部分反映在正確的維度上;相對的,如果差 距很小,則顯示數學運算過程造成一定量的數 據結構變異反映在不正確的維度上。值得注意 的是, DCA在運算過程中的降趨步驟,使第一 軸以後的特徵值不能作為解釋該軸變異量的指 標。我們依據:(1)樣點在排序軸的分數與其環 境梯度值之相關係數在0.995以上;(2)沒有軸端 反轉現象;(3)正確梯度軸之特徵值或軸長至少 是多餘的次一軸的兩倍以上,整理了 PCA 、 CA 、 DCA 在不同梯度長度下的適用性,詳如 Table 1。 PCA 在較小的梯度下有較理想的排序結果
(Fig. 3)。在type I model較大梯度長度下,排序 結果會有軸端樣點反轉及壓縮的畸變現象。於 梯度長度 0.5 T (含)以上時,在第一軸靠近梯度 兩端樣點的排列順序與模擬的順序相反,同時 樣點間的排列位置相對於中間樣點亦較為緊 密;由特徵值及解釋變異量,亦可發現其產生 一個多餘的第二軸。 PCA 第一軸的樣點排序分 數與其在環境梯度軸上的分數之相關係數,大 致隨梯度長度減小而增加,軸端反轉及壓縮的 畸變現象也隨之減緩,顯示梯度愈小,排序結 果愈理想。 Ty pe II model 的 PCA 排序結果與 type I model 有極大差異。大致而言,不論梯度 長度為何,均無軸端反轉現象,但梯度較長 時,有明顯中間樣點壓縮及軸端樣點疏離的情 形,並且產生多餘的第二軸。但隨梯度減小, 排序結果愈趨於理想。 CA在較小梯度下,有較佳的排序表現(Fig. 3)。兩種梯度模型的結果類似,梯度愈長,樣點 之間的拱形狀排列與軸端壓縮現象愈明顯;同
Table 1. Applicability of ordination techniques with different gradient lengths of communities
Artificial Gradient leng th Ap plicab ility of ord ina tio n t ech niq ue co mmu nity T1) DCA a xes (SD) PCA CA DCA
On e d ime nsion Ty pe I mod el 2 11.3 - - Yes 1.5 8.2 - - Yes 1 5.2 - - Yes 0.75 3.6 - Yes Yes 0.625 2.8 - Yes -0.5 2.1 - Yes -0.25 0.6 Yes Yes -Ty pe II mod el 2 12.1 - - Yes 1.5 9.0 - - Yes 1 6.1 - - Yes 0.75 4.5 - Yes Yes 0.625 3.7 - Yes Yes 0.5 2.9 Yes Yes -0.25 1.4 Yes Yes -Two d imen sio ns
Ty pe I mod el 2, 1 12.3, 6.5 - - Yes Ty pe II mod el 2, 1 11.9, 6.4 - - Yes 1, 0.5 5.7, 2.9 - - Yes 0.5, 0.25 3.2, 1.5 Ye s Yes
Fig. 3. Behavior of the 3 ordination techniques, PCA, CA, and DCA, with different gradient lengths of 1-dimens ional artificial communities. The circles are data of the type I model, and the cros ses are data of the type II model. Note that the s econd axes are s uperfluous.
時,第二軸與第一軸的特徵值及解釋變異量的 差距亦愈小。整體而言,第一軸的樣點排序分 數與其在環境梯度值之相關係數大致隨梯度減 小而增加,而第二軸的解釋變異量則隨梯度減 小而減小,顯示在較小的梯度長度下,CA排序 結果較為理想。 DCA 在梯度較長時,有較佳的排序表現 (Fig. 3)。兩種梯度模型的結果類似,在較長梯 度下, DCA排序第一軸與環境梯度之相關係數 較高,且軸長明顯高於第二軸。較小梯度長度
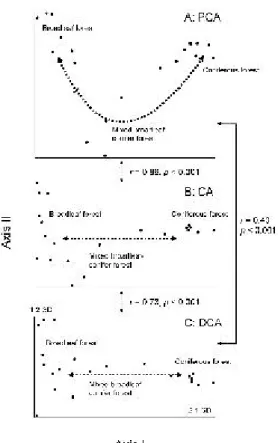
下,則會產生類似 CA 的拱形效應;梯度更小 時,第一軸與第二軸軸長的差異減小,且樣點 在第二軸的排序分數與環境值之相關係數增 加,顯示在較小的梯度下, DCA運算結果產生 了多餘的第二軸。DCA產生多餘第二軸的臨界 軸長,以 DCA 第一軸度量,約為 3 SD 。 二維梯度模擬結果與一維梯度類似,DCA 在較長的梯度有較佳的表現, CA 及 PCA 則在 較小的梯度下較為理想。在 2 與 1 T ,以及 1 與 0.5 T 的梯度長度下,DCA在前二軸的相關係數 均甚高,第三軸的相關係數則較低,且第二軸 的軸長約為第一軸的一半,顯示排序結果非常 理想。 PCA 及 CA 則只有在第一軸有較佳的表 現,次要變異梯度則表現在第三軸,CA並呈現 嚴重的拱形效應。但當梯度長度減小到 0.5 與 0.25 T 時,PCA及CA不論在相關係數或排序軸 的解釋變異量上,均有理想的表現;DCA 雖在 前二軸有理想的排序結果,但是第三軸與次要 環境梯度亦有高度相關,同時其軸長與第二軸 亦類似,顯示在較小梯度下, DCA產生了多餘 的第三軸。CA將次要梯度表現在正確第二軸的 梯度長度,以DCA第二軸軸長度量,約在2 SD 以下。 二、鳥類群聚資料 海拔400-3,700 m取樣點鳥類資料的排序分 析, DCA 第一軸軸長為 6.1 SD ,第二軸為 2.5 SD 。 PCA 及 CA表現出與模擬梯度類似的馬蹄 形及拱形效應;相對的, DCA則有較理想的排 序結果。 北大武山海拔 1,600-2,600 m 成熟林鳥類群 聚, DCA 前二軸軸長分別為 2.1 及 1.2 SD 。三 種排序方法對群聚變異趨勢的分析結果大致類 似(Fig. 4),惟 PCA 前二軸的樣點分布略呈倒鐘 形趨勢,但未有軸端反轉的現象,顯示這是一 個符合 type II model 的群聚梯度,但對 PCA 而 言,梯度稍長。 玉山海拔 1,800-2,500 m 成熟林鳥類群聚, DCA前二軸軸長亦分別為2.1及1.2 SD。樣點在 PCA及CA前兩軸的空間分布均呈現拱形狀(Fig. 5a, b);沿第二軸,海拔梯度兩側的闊葉林與針 葉林樣點在軸上端,而海拔梯度中段的針闊葉 混生林樣點在下端,樣點排列並沒有任何生態 意義,顯示這是由於排序方法造成的拱形現 象,而非數據結構本身造成。相對的, DCA的 樣點分布主要沿第一軸開展(Fig. 5c),其排序結 果較為合理。 DCA 前兩軸與 CA 前兩軸樣點分 布的相關係數僅 0.60 ,但 DCA 前兩軸與 CA 第 一軸及第三軸的空間分布相關係數則高達 0.95 (Fig. 6),顯示CA的鳥類群聚次要變異梯度表現 在第三軸,此結果與二維梯度模擬時,在較長 梯度下的結果類似。 拉拉山海拔800-1,200 m成熟林鳥類群聚, DCA前二軸軸長分別為1.14及1.06 SD。樣點在 前二軸的空間分布以 CA 與 DCA 較為類似(r = 0.92, p < 0.001),PCA與前兩者的相似性較低, Fig. 4. Ordination diagrams of Mt. Dawu bird data. (a) PCA shows an arch shape; and both (b) CA and (c) DCA s how relatively ideal configurations.
Fig. 5. Ordination diagrams of Mt. Jade bird data. Both (a) PCA and (b) CA s how arch shapes , and (c) DCA shows a relatively ideal configuration.
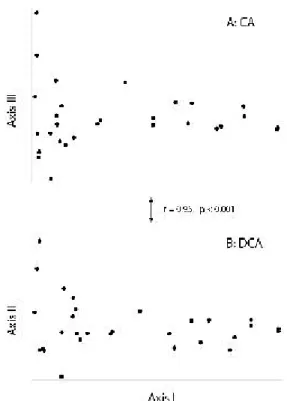
Fig. 6. Ordination diagrams of Mt. Jade bird data. (a) Configuration of s ite points in the first and third axes of CA and (b) in the first 2 axes of DCA. The diagrams are very similar. 但均無拱形狀等畸變的現象。丹大山海拔2,900-3,000 m 針葉林鳥類群聚, DCA 前二軸軸長分 別為1.0及0.6 SD。三種方法排序結果均無畸變 現象;樣點在前二軸的空間分布亦以 C A 與 DCA 較為類似(r = 0.84, p < 0.001)。 整體而言,群聚梯度愈長, PCA 及 CA 的 馬蹄形或拱形效應愈明顯,樣點在 DCA第一軸 上則相對有較理想的排列。 PCA 及 CA 變形的 臨界梯度長度並不一定,須視資料結構而定, 例如大武山鳥類群聚 DCA第一軸軸長2.1 SD, CA沒有拱形排列現象,但玉山鳥類群聚軸長亦 為 2.1 SD ,卻產生明顯拱形效應;無論如何, 就台灣山區鳥類群聚的排序分析而言,此一臨 界梯度大致在 2 SD 左右。當梯度小於 2 SD , 則樣點在三種排序的空間分布均沒有拱形或軸 端反轉現象,但 DCA 可能在高次維產生多餘 的軸。
討 論
群聚變異梯度長度對排序正確性有決定性 影響,依據梯度長度選擇適當的方法,才能得 到理想的排序結果。依本研究的結果, PCA 及 CA 適合在較短的梯度下使用, DCA 則適用於 長梯度。 PC A 可視為直線與平面的最小平方迴歸 (leas t-s quares regress ion)之延伸(Pielou 1984, Digby and Kempton 1987, ter Braak 1995, Lynn and McCulloch 2000)。因為 PCA 建立在物種豐 富度沿環境梯度呈線性反應的模式,故只有在 極短的空間尺度或群聚變異梯度下, PCA 才適用。值得注意的是,過去對於 PCA 在生物群聚 排序之應用,多只討論長梯度下的馬蹄形效應 (e.g., Gauch and Whittaker 1972, Gauch 1982, Pielou 1984);本研究將梯度模型分為包含環境梯度軸 端的 type I model 與不含軸端的 type II model , 結果發現馬蹄形效應只出現在type I model。在 比較長的梯度之下, ty pe II model 的軸端並不 會反轉,而是向兩側延展,形成鐘形分布,此 乃梯度兩端的分布中心位於梯度範圍外之物種 導引的結果;也就是說在任一連續環境梯度中 截取一段不包含梯度端點(如山頂、林緣)的範圍 進行分析時, PC A 可以容許較長的梯度。另 外,本研究在分析過程中,雖將type I model的 梯度模擬至 0.25 T ,但在真實狀況下,比較小 的梯度應多屬於type II model;而根據模擬梯度 的結果,任一梯度長度下,type II model的PCA 排序結果均較 ty pe I model 理想。亦即 PCA 在 生物群聚排序的適用性,應比過去受限於type I model 馬蹄形效應的印象來得廣。 在比較長的群聚變異梯度之下, CA 避免 了嚴重的馬蹄形效應,但仍有拱形效應的問 題。CA的拱形效應導因於數學運算過程中,豐 富度沿梯度的非線性分布模式(ter Braak 1985), 不過有些學者認為此一問題與 PCA 的馬蹄形效 應相比,並不特別嚴重,因為CA的軸端並不反 轉(e.g., Gauch 1982)。 CA 比較特別的問題是資 料結構的第二梯度可能延遲到較高的軸才顯現 出來,一如本研究在二維梯度模擬及玉山成熟 林鳥類群聚資料中所見。Gauch (1982)指出這是 因為當第二梯度的軸長小於第一梯度的一半 時,導致拱形效應的二次方畸變之特徵值大於 資料結構本身第二梯度的特徵值所致。儘管CA 可能有上述的問題,然而在較小的梯度下,CA 的排序效果是非常理想的(Lynn and McCulloch 2000)。 DCA由CA修改而來,它的目的在修正CA 的軸端壓縮以及多餘第二軸(Gauch 1982, t er Braak 1995)兩項主要缺點。 DCA 以降趨技術修 正拱形效應投影在第二軸上的值,而拱形效應 導致第一軸軸端壓縮的問題則以非線性重新刻 劃的方式修正(Hill and Gauch 1980)。經過這兩
種程序,在比較長的模擬梯度之下, DCA表現 的排序結果甚為理想。一些學者支持 DCA的方 法,例如 Peet et al. (1988)認為降趨及非線性重 新刻劃有助於生態意義的詮釋。但很多學者卻 不贊同 DCA 修正的方式。例如 Pielou (1984)即 主張 DCA 針對 CA 缺點的修正,有矯枉過正的 嫌疑,可能在不知不覺中破壞了資料訊息蘊含 的生態意義。 Wartenberg et al. (1987)認為拱形 是樣點之間存在的距離,而非數學的人工結 果,因此無須對此進行修正。Kenkel and Orlóci (1986)發現 DCA 在多重梯度下的表現並不好。 Minchin (1987)則認為DCA會因為降趨或非線性 重新刻劃,或兩者同時的影響,而造成與梯度 有關的資料訊息之減損。 Jackson and Somers (1991)亦發現 DCA 的樣點排列會受到非線性重 新刻劃時任意決定的段落數(number of segments) 的影響,而其結果在生態詮釋上也有很大的差 異。 Ter Braak (1995)認為軸端壓縮不是太嚴重 的問題,因此建議避免習慣性的使用非線性重 新刻劃。 Legendre and Legendre (1998)則主張除 了估計群聚梯度長度的特殊目的之外,根本應 該避免使用 DCA 。 本研究的結果則發現,在比較短的模擬梯 度下, DCA會在次一維上產生多餘的梯度。我 們認為在比較長的梯度下, DCA理想的排序結 果不容置疑,然而在較小的梯度下,因為CA的 拱形效應甚為輕微,並不需要進行降趨與非線 性重新刻劃的修正工作;在比較小的梯度下使 用DCA,反而會嚴重扭曲資料原有的生態訊息。 在DCA的非線性重新刻劃過程中,會將變 數予以標準化(standardiz ation),而以標準偏差 取 代 物 種 高 斯 曲 線 的 範 圍 ; 以 此 物 種 轉 換 (species turnover)的平均標準偏差作為軸的尺度 (Legendre and Legendre 1998)。一個高斯曲線的 起落約跨越4 SD (Fig. 1),而因為標準化的關係, 大部分物種具有此一耐受範圍。因此兩個樣點 間在梯度軸上的分布若差距 4 SD 以上,則可預 期這兩個樣點之間沒有共同種的存在(t er Braak 1995, Legendre and Legendre 1998);若物種轉換 一半則約跨越 1到1.4 SD (Legendre and Legendre 1998)。 Ter Braak (1995)建議以 SD 值作為排序
方法選擇的依據;若軸長小於 2 SD ,則大部分 的物種豐富度反應曲線屬於線性,可以考慮使 用PCA或由其衍生之RDA;若大於4 SD,則應 使用 CA, DCA ,或由 CA 衍生之排序法 CCA; 若排序軸長約 3 SD 時,則 PCA 與 DCA 的樣點 排序結果類似。然而需注意的是, ter Braak 的 建議大抵屬於理論推衍性質,而非實證結果。 由本研究的結果,我們則認為當軸長 2 SD 以上時,應優先使用 CA;尤其若 CA 的結果沒 有拱形時,應該避免使用 DCA 。而當 CA 有明 顯拱形排列時,為判斷此為資料結構本身,或 只是數學運算造成的拱形效應,應該檢視軸與 相關的環境因子之間是否合理或具有生態意 義,並測試及比較 CA 及 DCA 的排序結果;只 有在判斷CA的拱形是數學造成的人工結果下, 才使用 DCA 。另一方面,當軸長在 2 SD 以下 時, CA 及 PCA 較沒有拱形或馬蹄形效應的顧 慮,因此均可使用。 CA 與 P CA 各有其優缺 點, CA 同時對物種及樣點排序,而 PCA 則僅 能對樣點作排序;但是 PCA 在樣點與環境的雙 疊圖(biplot )上提供比 CA 更多的定量資訊(t er Braak and Prent ice 1988, ter Braak 1995),並且 PCA 類似線性迴歸,排序軸蘊含的訊息容易被 了解。至於 CCA 及RDA ,由於其分別為 CA及 PCA 的直接梯度分析形式,選用原則可遵循上 述之準據。 在國內,研究者在生物群聚梯度分析上, 明顯過於偏好使用DCA,這可能是對群聚變異 梯度長度在排序方法上的限制了解不夠所致。 很多研究者並未注意梯度長度與排序方法適用 性之間的關係;在使用 DCA的文獻中,有許多 分析結果,或第一軸軸長即已小於 2 SD ,或選 用多個軸,然其選用的高次維軸長過小(e .g., Huang 1994, Chung 1997),可能係多餘的梯度。 建議往後的研究者應該視群聚變異梯度長度與 資料結構,選擇適當的排序方法;切勿習慣性 的使用單一排序法,以避免不適用時產生錯誤 的結果。
結 論
每一排序方法均有其適用的梯度長度,排 序方法的選擇應該視群聚變異梯度長度與資料 結構而定。 PCA 及 CA 適合在較短的梯度長度 下使用, DCA 則適用於較長的梯度。 DCA 運 算結果的軸長可作為群聚梯度長度判斷的參 考。當梯度軸長在 2 SD 以上,且 CA 的樣點分 布因數學運算而有明顯拱形排列時,可以考慮 使用 DCA;但若 CA 沒有拱形效應產生時,則 以使用 CA 較適當。軸長在 2 SD 以下時, DCA 的運算結果可能在高次維產生多餘的梯度軸, 相對的,在此較短的梯度長度下, CA 及 PCA 則沒有拱形或馬蹄形效應的顧慮,因此以使用 CA 及 PCA 較佳。群聚變異梯度長度影響排序 的正確性,排序方法的選擇應考慮生物群聚之 梯度長度,切忌習慣性的使用單一排序法於所 有尺度的群聚生態學研究。謝 誌
本研究感謝方韻如小姐允用其野外資料, 陳一菁小姐在內容上提供許多寶貴意見。我們 同時感謝兩位審查委員之寶貴意見。引 用 文 獻
Austin MP. 1985. Continuum concept,
ordina-tion method s, and niche theory. A nn Rev Ecol Syst 16:39-61.
Chung CC. 19 97 . The v ariat io n of fo liag
e-height pro files in the succe ssion of Machi
lus-Ca sta no psi s n atu ra l fore st of no rth we ste rn
Taiwan [master’s dissertation]. Taipei, Taiwan: Nat ional Taiwa n Univ. 40 p. [in Chinese with English s ummary].
Digby PGN, Kempton RA. 1 987. Multivariate
analysis of ecological communities . New York: Chapman & Hall. 206 p.
Douglas ME, Endler JA. 1982 . Quan tit ative
matrix comparisons in ecological and evolution-ary inve stigations. J Theor Biol 99:777-95.
Fang YJ. 1996. The relationship between forest
the influence of the timber stand imp rovement program in mid-elevation area, northern Taiwan [maste r’s d is serta tion]. Taipe i, Ta iwan: Na-tional Taiwan Univ. 70 p. [in Chinese with En-glish su mmary].
Gauch HG, Whittaker RH. 1972. Compa rison
of ordin ation techniqu es. Ecology 53:868-75.
Gauch HG. 1982. Multivariate analysis in
com-mu nity ec olog y. Camb ridg e, UK: Cambrid ge Univ. Pres s. 298 p.
Hil l MO, Gauc h HG. 198 0. De trende d
corre-spo nden ce an alys is: a n imp rove d ord inat ion techniqu e. Vegetatio 42:47-58.
Huang CS. 1994. Studies on the forest
dynam-ics and population structure of the Kawak amii fir fo rest in the northern slope of Hsue h-Shan [maste r’s d is serta tion]. Taipe i, Ta iwan: Na-tional Taiwan Univ. 89 p. [in Chinese with En-glish su mmary].
Jackson DA, Somers KM. 1991. Putting things
in order: the ups and d owns of detren ded cor-respondenc e analysis. Am Nat 137:704-12.
Kenkel NC, Orlóci L. 198 6. App lying me tric
and nonmetric multidimensional scaling to eco-logical studies: some n ew results. Eco logy 67: 919-28.
Kr ebs CJ . 19 99 . Ec olog ic al met ho do lo gy .
Menlo Park, CA: Addison-Welsey Educational Publishers. 620 p.
Lee CK. 1995. A comparison of bird
communi-ties between conifer plantation and natural broa-dlea f forest [mas te r’s diss erta tion ]. Taipe i, Taiwan: Nat ional Taiwan Univ . 68 p. [in Chi-nese with English su mmary].
Le ge ndr e P, Le ge ndr e L. 19 98 . Nu me rica l
eco logy. 2nd English e d. A mst erda m, Nethe
r-lands: Elsevier Science BV. 853 p.
Lynn HS, McCulloch CE. 2000. Using principal
component analysis and correspondence analy-sis for estimatio n in late nt v ariab le mo dels . J Am Stat Asso c 95:561-72.
McCune B, Mefford MJ. 1999. PC-ORD.
multi-variate an alysis of ecological data, versio n 4. Gle nede n Be ach, OR: MjM Software Design. 237 p.
Minchin PR. 1987. An e valuation of the
rela-tiv e ro bust ness of t echn ique s for eco logical ordination. Vegetatio 69:89-107.
Peet PK, Knox RG, Cas e JS, Allen RB . 1988.
Pu tt in g th in gs in orde r: t he a dv an ta ge s of det rende d correspo ndenc e ana lysis . Am Nat 131:924-34.
Pielou EC. 1984. The interpretation of
ecologi-cal data : a primer on classification and ordina-tion. New York: Wiley. 263 p.
Shiu HJ. 19 95 . The relation sh ips b et we en
avian co mmunity struct ure and enviro nmental factors in mature forests of mid-elevation moun-ta in a rea s in Ta iwan [maste r’s d iss ert at ion ]. Taipei, Taiwan: National Taiwan Univ. 98 p. [in Chinese with English s ummary].
Su HJ. 1987. Studies on the multivariate
analy-sis in v egetation ecolog y III: detrende d corre-sp on de nce a na lys is an d re lat ed o rdina tion tec hnique s. Q J Chin For 20(3):45-68. [in Chi-nese with English su mmary].
ter Braak CJF. 1985. Correspond ence
analy-sis of incidence a nd abund ance da ta: pro per-ties in terms of a u nimoda l resp ons e mode l. Biometrics 41:859-73.
ter Braak CJF. 1986 . Ca nonical correspo
n-denc e analysis: a new eigenv ector technique for multivariate direct grad ient ana lysis. Ecol-ogy 67:1167-79.
ter Braak CJF. 1995. Ordination. In: Jongman
RHG, ter Braak CJF, van Tongeren OFR, editors. Da ta an alys is in co mmunity an d la nds ca pe ec olog y. Cambrid ge , UK: Cambrid ge Univ. Press. p 91-173.
ter Braak CJF, Prentice IC. 1988. A theory of
gradient an alysis. Adv Ecol Res 18:271-317.
Wartenbe rg D, Fe rs on S, Rohlf FJ. 1 98 7.
Putting thin gs in order: a critique of detrended corresponde nce analysis. Am Nat 129:434-48.