國 立 交 通 大 學
生物科技學院
生物科技研究所
碩士論文
使用重組蛋白發展對 HPV、IHHNV 和 TSV
殼蛋白有專一性之單株抗體
Using Recombinant Protein to Develop Monoclonal
Antibody Specific to Coat Proteins of
Hepatopancreatic Parvovirus (HPV)、
Infectious Hypodermal and Hematopoietic Necrosis
Virus (IHHNV) and Taura Syndrome Virus (TSV)
研究生:陳 文 鴻
指導教授:吳東昆 博士
使用重組蛋白製造對 HPV、IHHNV 和 TSV 殼蛋白
有專一性之單株抗體
Using Recombinant Protein to Develop Monoclonal Antibody
Specific to Coat Proteins of Hepatopancreatic Parvovirus (HPV)、
Infectious Hypodermal and Hematopoietic Necrosis Virus (IHHNV)
and Taura Syndrome Virus (TSV)
研 究 生:陳文鴻 Student:Wen-Hung Chen 指導教授:吳東昆 博士 Advisor:Dr. Tung-Kung Wu
國 立 交 通 大 學
生 物 科 技 研 究 所
碩 士 論 文
A ThesisSubmitted to Department of Biological Science and Technology College of Science
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Biological Science and Technology June 2006
Hsinchu, Taiwan, Republic of China
使用重組蛋白製造對 HPV、IHHNV 和 TSV 殼蛋白有專一性之單株抗體
學生:陳文鴻 指導教授:吳東昆博士 國立交通大學生物科技學系碩士班
摘 要
根 據 聯 合 國 糧 食 及 農 業 組 織 (FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS,FAO)的數據指出,蝦養殖 產業為一個能夠提供高營養價值的食物來源,也能為發展中國家提供經濟 收入的一重要之產業。但近年來,因為病毒問題,使其遭受巨大之損失。 因此若是能有效且儘早偵測到病毒,可以針對所面臨之狀況,提早作出對 應,以降低損失。
本研究從 NCBI database 資料中,分別在肝胰小病毒 (hepatopancreatic parvovirus, HPV)、傳染性皮下及造血組織壞死病毒 (infectious hypodermal and hematopoietic necrosis virus, IHHNV) 和桃拉病毒 (taura syndrome virus, TSV) 等三種常見之蝦病毒中,各選出一段殼蛋白基因,利用重組蛋白的 方式,在大腸桿菌中大量表現。並利用親和性管柱純化,將純化所得之蛋 白質當作抗原注射至老鼠體內誘導,以期發展出針對這些蛋白質具有專一 性的單或多株抗體。 目 前 實 驗 已 可 以 成 功 地 利 用 大 腸 桿 菌 , 大 量 表 現 HPV 、 TSV 及 IHHNV 的重組蛋白,並利用重組蛋白上所帶的親和性標記進行純化,再 以這個純化的重組蛋白,在 BALB/c 老鼠體內引發免疫反應,經由 ELISA 及西方點墨實驗證實,可以免疫出對重組蛋白具有反應的單株抗體。之後 結合融合瘤技術發展單株抗體,在 HPV 方面,總共得到 2 株細胞株。在 西方點墨實驗中,均可以辦別到 HPV 重組蛋白。在蝦體的檢驗中,使用 3-24 這株單株抗體進行西方點墨實驗,其初步結果與病毒檢測試劑 (PCR
diagnosis) 所得結果相符。在 IHHNV 目前篩選到 3-62 這株細胞株,可辯認 到 IHHNV 重組蛋白。經由初步結果顯示,証實所發展的單株抗體,具有 發展成檢驗試劑的可能性。
Using Recombinant Protein to Develop Monoclonal Antibody Specific to Coat Proteins of Hepatopancreatic Parvovirus (HPV)、Infectious Hypodermal and
Hematopoietic Necrosis virus (IHHNV) and Taura Syndrome Virus (TSV) Student: Wen-Hung Chen Advisor: Dr. Tung-Kung Wu
Institute of Biological Science and Technology National Chiao Tung University
Abstract
As the FAO (FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS) reported, the aquaculture of shrimp is a very important industry. It can support high nutritional food and provide the income for the
developing nation. Because of the viral disease, this industry suffers huge loss
recently. Therefore, if the detection of virus can be effective as soon as possible, we may prevent the diseases from the shrimp.
In this study, we refer to the NCBI database to design primers which are used for amplification of the gene fragment of hepatopancreatic parvovirus (HPV)、 infectious hypodermal and hematopoietic necrosis virus (IHHNV) and taura syndrome virus (TSV). These gene-encoding structural coat proteins were cloned into expression vector and transformed into E. coli. The objective was to produce recombinant coat protein with a 6-histidine tag. After induction, the recombinant proteins were produced, purified by Nickel column and used for immunization of BABL/c mice for polyclonal antibody production. The mouse antiserum showed specific immunoreactivity to the recombinant protein as verified by ELISA and Western blot. In HPV, the western blot data indicated that two monoclonal antibodies against the HPV recombinant protein were constructed. The dectection of shrimps with monoclonal antibody 3-24 strain exhibited parallel result as compared with that of PCR diagnosis. In IHHNV, 3-62 strain showed immunoreactivity against the IHHNV recombinant protein or coat protein purified from the IHHNV infected shrimp.
謝 誌
首先,感謝我的指導教授,吳東昆 老師,在學生學涯遭受難關時,願 意幫助學生,這些日子以來,在生活上、實驗上給予甚多的指導與建議, 讓學生得以有機會完成這個學位。另外,很感謝 淡江化學的鄭建中 老 師、系上 袁俊傑 老師,百忙之中給予學生,在論文與實驗方面的建議與 指正,讓學生的論文更加完整。 感謝 林苕吟 老師,於前段求學日子中,在生活上的體諒與幫助,僅 獻上學生的祝福。 另外,感謝 毛仁淡 院長、邱顯泰 老師、賴美玲 師母及系辦 郭淑卿、 郭珍佑、呂聖鈴 小姐,在那段失意的日子裡,所給予的幫助與鼓勵。 完成我的碩士了,這段日子以來,感謝大家的幫忙與鼓勵,終於克服 了。實驗室的程翔、裕國學長、媛婷學姐、小芭、希希、阿美、晉豪、大 鳥、令宗、宏明、晉源、小妹、文萱、大景、皓宇以及志豪、昶文、秉 博、惠敏、勝致學長、文亮學長,還有智媛、惠娟、婉如等,很感謝大 家,這些日子以來的陪伴與幫助。 最後,謝謝兩個弟弟,俊豪、春元,在家裡經濟狀況不好的情況下, 能夠體諒我這個已居立業之年的哥哥,繼續就學。父親、母親,很感謝有 您們的支持,讓我可以一路求學過來,親愛的家人們,謝謝你們的支持, 謝謝。 文鴻 06 年 6 月 21 日于交大
目 錄
中文摘要……… i 英文摘要……… iii 謝誌……… iv 目錄……… v 圖目錄………viii 表目錄……… x 附錄……… xi 頁次 第一章 前言………1 1.1 水產養殖及養蝦產業概論………1 1.2 蝦病對養殖產業的影響………4 1.2.1 蝦病之簡介………4 1.2.2 主要蝦病毒之介紹………61.2.2.1 White spot syndrome virus (WSSV) ………6
1.2.2.2 Hepatopancreatic parvovirus (HPV) ………7
1.2.2.3 Infectious hypodermal and hematopoietic necrosis virus (IHHNV) ……….8
1.2.2.4 Taura syndrome virus (TSV) ………...8
1.2.2.5 其它病毒………..9 1.3 蝦病毒之檢測……….10 1.4 對抗蝦病毒之策略……….11 1.5 本研究之目的……….12 第二章 材料與方法……….15 2.1 抗源的製備……….15 2.1.1 病毒殼蛋白 (coat protein) 基因的選取……….15
頁次 2.1.2 引子的設計……….15 2.1.3 表現載體……….16 2.1.4 表現菌株……….18 2.1.5 病毒基因體的抽取與確認……….18 2.1.6 蛋白質重組……….20 2.1.7 表現與純化……….24 2.2 單株抗體之製作……….26 2.2.1 材料………...29 2.2.2 免疫反應………...30 2.2.3 骨髓癌細胞………...30 2.2.4 細胞融合………...31 2.2.5 細胞增殖與取樣篩檢………...32 2.2.6 抗體之生產………...32 2.2.7 腹水抗體製備………...33 2.3 抗體的測定……….33 2.3.1 材料………...33 2.3.2 ELISA………...35 2.3.3 SDS-PAGE………...35 2.3.4 Western blot……….36 2.4 儀器設備………37 第三章 結果與討論………37 3.1 病毒基因片段取得………37 3.2 重組蛋白的表現與純化………40 3.3 抗體的製作………43 3.3.1 單株抗體力價的測試………..43
頁次 3.3.2 單株抗體對抗原反應篩選………44 3.3.3 單株抗體特異性之測試………47 3.3.4 重組蛋白 Alignment 結果 (使用 Clustal W)………50 第四章 結論與展望………...51 第五節 參考文獻………...53
圖 目 錄
頁次
圖 1-1:水產品佔動物性蛋白攝取之比例………..1
圖 1-2:蝦養殖產量………..2
圖 1-3:蝦養殖產值………..3
圖 1-4 WSSV virions (a)及 WSSV nucleocapsid(b)之電顯圖……….7
圖 1-5 HPV virions 電顯圖………8 圖 1-6 TSV virions 電顯圖……….9 圖 1-7 YHV virions 電顯圖………..10 圖 2-1 細胞內核酸的合成路徑………31 圖 3-1-1 IHHNV PCR 產物………..37 圖 3-1-2 HPV 的確認………38 圖 3-1-3 HPV PCR 產物………...38 圖 3-1-4 TSV RT-PCR 產物……….39 圖 3-1-5 TSV Nest-PCR 產物………..39 圖 3-2-1 IHHNV 重組蛋白的表現與純化………..40
圖 3-2-2 IHHNV 重組蛋白與 anti-his taq 抗體西方點墨實驗………..40
圖 3-2-3 HPV 重組蛋白的表現與純化………..41 圖 3-2-4 HPV 重組蛋白與 anti-his taq 抗體西方點墨實驗………..41 圖 3-2-5 TSV 重組蛋白的表現與純化………..42 圖 3-2-6 TSV 重組蛋白在桿狀病毒系統中的表現………..42 圖 3-3-1 實驗接種小鼠血清力價測試,HPV 重組蛋白………..43 圖 3-3-2 實驗接種小鼠血清力價測試,IHHNV 重組蛋白……….43 圖 3-3-3 融合成功細胞產生的抗體對 HPV 重組蛋白抗原反應篩選…….45 圖 3-3-4 HPV 重組蛋白與抗體西方點墨結果………..45
頁次 圖 3-3-5 HPV 3-24 抗體與蝦檢體之西方點墨實驗結果………46 圖 3-3-6 融合成功細胞產生的抗體對 IHHNV 重組蛋白抗原反應篩選…..46 圖 3-3-7 IHHNV 3-62 抗體與蝦體之西方點墨實驗結果………...47 圖 3-3-8 HPV 3-24 抗體與重組蛋白 ELISA 實驗結果………..48 圖 3-3-9 HPV 3-7 抗體與重組蛋白 ELISA 實驗結果………48 圖 3-3-10 IHHNV 3-62 抗體與重組蛋白 ELISA 實驗結果………...49 圖 3-3-11 HPV 3-24 抗體與重組蛋白西方點墨實驗結果……….49 圖 3-3-12 HPV 3-7 抗體與重組蛋白西方點墨實驗結果………..49 圖 3-3-13 IHHVN 3-62 抗體與重組蛋西方點墨實驗結果………...50
表 目 錄
頁次 表 1.1 蝦養殖產量……….2 表 1.2 蝦養殖產值……….3 表 3.1 HPV 重組蛋白 polyclonal antibodies………...45附 錄
頁次 附錄一、實驗設計流程圖……….………..14 附錄二、 單株抗體製作流程圖………..28 附錄三、 IHHNV 選取之基因片段序列,及其蛋白質序列………61 附錄四、 HPV 選取之基因片段序列,及其蛋白質序列……….62 附錄五、 TSV 選取之基因片段序列,及其蛋白質序列……….63 附錄六、桿狀病毒表現系統………..64 附錄七、immuno-strips………..65 附錄八、抗原與免疫金膠抗體結合示意圖……….65 附錄九、immuno-PCR………..66 附錄十、抗體親和性管柱純化病毒示意圖………66第一章 前言
1.1 水產養殖及養蝦產業概論
根 據 聯 合 國 糧 食 及 農 業 組 織 (FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS,FAO)的報告( The state of world Fisheries and Aquaculture, 2004)指出,2002 年全球捕撈和水產養殖生 產工業總共提供了 1.01 億噸的食用水產品。其中至少提供了 20%的動物性 蛋白來源(圖 1-1)。但隨著長久對自然界水資源的開發,漁業的捕獲量已呈 停頓。因此水產養殖的角色在水產品供應上,日益重要。
圖 1-1 水產品佔動物性蛋白攝取之比例 資料來源: FAO Fisheries Circular No.886, Rev.2,
Review of the state of world aquaculture
而在水產養殖中,養蝦產業又具有較高的經濟價值,根據 FAO 的資 料顯示,從 1990 至 2003 年,年產量及產值明顯呈正向成長 (圖 2、圖
1-3)。2003 年時,年產量為一百八十萬噸 (表 1.1),產值為近十兆美元 (參考 表 1.2)。總而言之,此一產業能提供食物來源,以及增加經濟收入。 圖 1-2 蝦養殖產量 資料來源: FAO 表 1.1 蝦養殖產量 年 1990 1991 1992 1993 1994 1995 1996 年產值(噸) 679976 838229 897042 835203 881959 928238 917315 年 1997 1998 1999 2000 2001 2002 2003 年產值(噸) 932832 999370 1068297 1164408 1348275 1405367 1804932 資料來源: FAO
圖 1-3 蝦養殖產值 資料來源: FAO 表 1.2 蝦養殖產值 年 1990 1991 1992 1993 1994 1995 1996 年產值 (US$1000) 4232126932 5147831774 5564748338 5243870896 5808843324 6062515828 6101260906 年 1997 1998 1999 2000 2001 2002 2003 年產值 (US$1000) 6088002339 6029950994 6601383965 7467516197 8194251165 7804330709 9322858775 資料來源: FAO
1.2 蝦病對養殖產業的影響 一般魚類養殖往往因疾病造成嚴重之損失,蝦之養殖亦不例外,蝦之 存活率通常只有 50%,可見由疾病所造成之損失有多大。一般說來,蝦病 病因主要有兩類: (1)非傳染性病因 : 營養不平衡,如缺乏膽固醇或維他命 C、水溫太高或太 低、鹽度驟變、中毒、藥毒等物理化學之傷害。 (2)傳染性病因,如細菌、黴菌、病毒、寄生蟲等等。 目前全球養蝦產業所面臨到的問題,其中又以傳染性病因中之病毒感染所 造成之損失最大。主要是因其具有傳染性,且又有極高之致死率。 1.2.1 蝦病之簡介 蝦病主要有病毒、細菌、黴菌性疾病、原生動物感染、外共生性微生 物引起之疾病、寄生蟲病。而其中病毒,對於在亞洲、美洲等地,快速發 展中的養蝦產業,有很高的致死率,造成產能的大量損失。[1] 一.病毒性疾病 (Viral Disease) 對養殖產業來說,病毒性疾病是最大的病因,目前至少有二十二種病 毒被報導出來,[2] 以下六種病毒為目前影響較嚴重且常被研究者,(1)白 斑 病 毒 (White spot syndrome virus, WSSV) , (2) 肝 胰 小 病 毒 (Hepatopancreatic parvovirus, HPV) , (3) 桃拉病毒 (Taura syndrome virus, TSV) , (4) 傳 染 性 皮 下 及 造 血 組 織 壞 死 病 毒 (infectious hypodermal and hematopoietic necrosis virus, IHHNV) , (5) 草 蝦 桿 狀 病 毒 (MonodonBaculovirus, MBV) 及(6)黃頭病毒 (Yellow head virus, YHV)。 二、細菌性疾病 (Bacterial Diseases)
許多種類細菌可引起養殖蝦的疾病,尤其在蝦苗、幼蝦及中蝦期為 主,通常受感染後可引起以下三類型之疾病感染 :
(1)軀殼、附肢及鰓等體表之潰爛 (又稱爛殼病及細菌性壞死病) ,通常由產 生溶解幾丁質酵素 (Chitinase) 細菌感染而形成外殼之局部穿孔,然後再侵 入體內,引起體內局部炎症反應,如感染未能被有效遏止則可演變成全身 性敗血症死。(2)體內局部感染症,細菌可在內臟、鰓、肌肉等造成局部性 感染病灶而形成膿瘍。(3)全身性敗血症,病原菌侵入體內淋巴血液及各臟 器組織。一般感染細茵的症狀有 : 蝦體變色、鰓變色、壞死、爛鰓、爛尾 等,例如細菌性黑鰓病 (Bacterial black gill disease) 。絕大多數蝦病細菌均 屬二次感染性細菌 (Secondray pathogens) ,通常必須先有各種緊迫誘因 (Predisposing factors) ,如其他病原 (病毒或其他細菌) 之感染、寄生蟲寄 生、外共生性微生物附生、外傷、營養缺失、化學或物理性緊迫作用等之 影響下,才能使這些原來在水中或蝦體外附著之細菌有機可乘而引發細菌 感染造成病害。迄今自病蝦所分離到之細菌大約可分為兩大類,即一種為 自病蝦體內或局部病灶所分離到之病原細菌:蝦病原細菌,通常本屬細菌 可引起病變,另一種為附生在蝦體表面之共生細菌:蝦外部共生細菌,一 般不引起病變。 三、黴菌性疾病 (Fungal Diseases) 黴菌性蝦病可分為兩種 : (1)全身性黴菌感染症 (2)局部性黴菌感染症 四、原生動物感染 (Protozoan Infections) 寄 生 性 原 蟲 有 兩 種 (1) 微 孢 子 蟲 類 (Microsporidia) (2) 簇 蟲 類 (Gregarines)。
五、外共生性微生物引起之疾病 (Diseases caused by Epicommensals) 蝦體表外殼、附肢及鰓常被外共生菌、原蟲或藻類附著增生,此類微 生物之附著並不侵入體內亦不造成病變,但大量增生附著蝦體之結果可引 起蝦呼吸、運動、攝食、脫殼之障礙而往往導致蝦呼吸困難窒息而死亡。
六、寄生蟲疾病 (Parasitic Diseases)
包括吸蟲 (Flukes, Trematodes),條蟲 (Tampeworms, Cestodes) 及線蟲 或圓蟲 (Roundworms, Nematodes) 。一般吸蟲類寄生在蝦頭胸部各器官、 腹側肌肉或外骨骼下休眠 ; 條蟲則在消化腺組織中 ; 線蟲則在頭胸部各器 官及消化道內外寄生。養殖蝦類之寄生蟲病發生率小亦鮮有大量死亡情形 發 生,主要原因是一般養殖蝦池內,極少寄生蟲傳染所需之中間寄主棲 息,因此寄生蟲之生活史無法完成。 1.2.2 主要蝦病毒之介紹 病毒因具有傳染性,以及高的致死性,所以較受大家之注意。以下將 就目前在全球引發較大影響之病毒,作一簡單之介紹。
1.2.2.1 White spot syndrome virus (WSSV)
白斑病毒, White spot syndrome virus,簡稱 WSSV,即白點病,會在病
蝦蝦殼上產生白點。是目前對蝦子影響甚鉅的病原體之一,並且對全球之 養蝦產業造成嚴重的傷害。雖然此一病毒可以感染幾種甲殼類,例如蝦 子、螃蟹和龍蝦,[3, 4, 5, 6, 7] 但其對對蝦類 (penaeid shrimp) 特別具有高 的毒性,在感染三到七天內,死亡率高達 90~100%。[3, 8] WSSV 病毒顆 粒呈桿狀,包覆膜蛋白,大小約 275×120 nm。為雙股 DNA 病毒,基因體 大 小 為 290 kb , [8] 被 外 殼 蛋 白 capsid protein 所 包 覆 , 核 蛋 白 衣 nucleocapsid 大小約 300×70 nm。[9] 其有一顯眼之特徵,就是該病毒體有 著 如 尾 巴 的 一 端 ( 圖 1-4) 。 [11, 12] 這 個 病 毒 被 歸 類 在 尼 瑪 病 毒 科 (Nimaviridae) 這個家族 (“nima”在拉丁文中為“線”的意思) 。[13] 目前已經定出三個完整的 WSSV 序列,共有 532 個開放閱讀區 (open reading frames, ORFs) 。包含非結構蛋白 (醣核苷酸還原酶 ribonucleotide reductase, 蛋白激脢 protein kinase, the chimeric thymidine kinase/thymidylate
kinase, and DNA 聚合酶 DNA polymerase [14, 15, 16, 17]) 或是結構蛋白 (VP28, VP26, VP24, VP19, and VP15) 等等。[18~23]
圖 1-4 WSSV virions (a)及 WSSV nucleocapsid(b)之電顯圖 資料來源: Virology. 266:227-236, 2000.
1.2.2.2 Hepatopancreatic parvovirus (HPV)
肝胰小病毒, Hepatopancreatic parvovirus,簡稱 HPV。在 1984 年,第
一次在 Penaeus mergiensis 和 Penaeus indicus 這兩種蝦子中發現。[24] 這種 病毒為單股 DNA 病毒,基因體長度約 6321 bp。[25] 廣泛地分佈在養殖及 野生蝦子中,其中包括 Penaeus monodon 和 Penaeus chinensis。[26] 死亡率 約在 50~100%之間,病程約為 4-8 週,通常主要病變在肝胰臟,使該器官 萎縮壞死。嚴重感染時,會造成生長減緩,進而造成產量下降。其外形為 多面病毒體 (icosahedral) ,沒有膜蛋白包覆,直徑約 22 nm (圖 1-5) ,基因 體 大 小 約 6 kb , 有 三 個 大 的 ORFs, 基 於 這 些 特 徵 , 將 其 歸 類 在 Parvoviridae 家族 。[27, 28, 29]
1.2.2.3 Infectious hypodermal and hematopoietic necrosis virus (IHHNV)
傳 染 性 皮 下 及 造 血 組 織 壞 死 病 毒 , Infectious hypodermal and hematopoietic necrosis virus,簡稱 IHHNV。1981 年在 Hawaii 發現。[30] 致 死率達 90%,病毒感染蝦體時,通常破壞其皮下組織,前腸、後腸皮下組 織以及造血組織上皮及肌肉。此病毒為小、多面體,沒有膜蛋白包覆,單 股 DNA,基因體大小約 4.1 kb。[31, 32] 包含三個大的 ORFs。[33] 依外形 特微,被歸類為小 DNA 病毒科 Parvoviridae 家族。[31]
圖 1-5 HPV virions 電顯圖,Bar 代表 100nm. 資料來源: Journal of General Virology. 76:813-817, 1995.
1.2.2.4 Taura syndrome virus (TSV)
桃拉病毒 , Taura syndrome virus,簡稱 TSV。 1992 年時,在靠近
Taura 河 (厄瓜多, Ecuador) 出口的養蝦場中所養之 Penaeus vannamei 發現, 因此命名為 Taura,死亡率 50-90%。此病毒為多面體,大小約 31 至 32 nm (圖 1-6) ,單股 RNA,基因體大小約 9 kb,依外形被歸類為 Parvoviridae 家族。[34]受感染蝦群在急 性 期 症 狀 為 倦 怠 、 軟 殼 、 腸 內 無 內 容 物 ,
身 體 紅 色 素 增 加 而 變 淡 紅 , 尤 其 尾 扇 邊 緣 呈 紅 色 , 故 又 稱 紅 尾 病 , 常 可 見 脫 殼 期 之 死 亡 蝦 。 而 在 慢 性 期 症 狀 為 在 急 性 期 耐 過 後 出 現 , 病 蝦 可 能 仍 有 食 慾 , 但 體 表 甲 殼 有 多 發 性 黑 色 斑 點 。
圖 1-6 TSV virions 電顯圖,左邊的 Bar 代表 300nm.
資料來源: Journal of General Virology. 78:313-319, 1997.
1.2.2.5 其它病毒
其它對養蝦產業造成較大影響的還有黃頭病毒, Yellow head virus
(YHV)、草蝦桿狀病毒 monodon baculovirus (MBV)等。
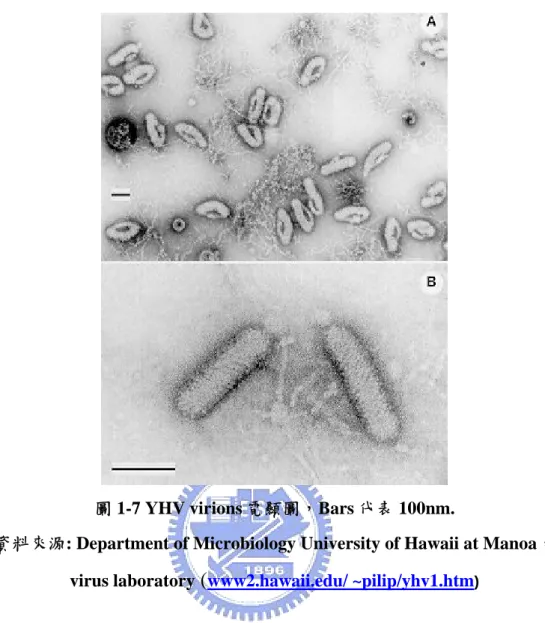
YHV, 黃頭症。1990 年時,在泰國發現。[35] 患病時會中斷進食,在 頭胸部 (cephalothoraxes) 和鰓 (gill) 會變成黃色。[36] 病徵出現 2-3 天就會 開始死亡。此病毒外表包覆膜蛋白,呈桿狀,大小約 40 × 170 nm (圖 1-7) ,有突起的外表,內部有螺旋狀的核蛋白。[37] 單股 RNA。[38] MBV, 草 蝦桿狀病毒。 1981 年時,第一次在台灣養殖的 Penaeus monodon 中發現。[39] 對草蝦來說,是個重要的病原體,患病時會造成高 致死率或生長遲緩。[40]
圖 1-7 YHV virions 電顯圖,Bars 代表 100nm.
資料來源: Department of Microbiology University of Hawaii at Manoa,
virus laboratory (www2.hawaii.edu/ ~pilip/yhv1.htm)
1.3 蝦病毒之檢測 因為蝦病毒對於養蝦產業影響之大,若是能有效且儘早偵測到病毒, 可以針對所面臨之狀況,提早作出對應,以降低損失。所以蝦病毒之檢測 發展就相形重要了。一個良好的檢測方法,必需考慮到快速、便宜、靈敏 及專一性。 目前所使用的方法主要有(1)組織病理學 (histopathology) 。[41, 42] 經 由 病 體 組 織 上 的 特 徵 , 判 斷 感 染 原 及 患 病 狀 況 。 (2) 免 疫 學 方 法 (immunological method) 。[43, 44, 45] 使用抗體對病原體的蛋白質作專一性 的辦認。 (3)聚合酶鏈鎖反應方法 (polymerase chain reaction, PCR)。[46, 47, 48] 設計引子,放大病原體某個核苷酸序列。 (4) DNA 探針方法 (in situ
hybridization methods using DNA probe)。[49, 50] 使用 DNA 探針辦認病原 體上特殊的核苷酸序列。除此之外,尚有一些其它改良的方法,例如 Dot-blot nitrocellulose enzyme immunoassays (DB-NC-EIA) 、 [51] miniarray system、[52] PCR-ELLISA。[53] 但其中,由於病理學檢測需長期的訓練, 且較費時。而使用聚合酶鏈鎖反應及 DNA 探針方法,操作人員需經過一 定的訓練,且需要聚合反應的機器。因此,免疫學方法,較有潛力發展成 兼具方便、專一性、便宜、靈敏之檢測工具。讓第一線的養殖漁民,可方 便地利用這項工具,快速 、正確地檢測,讓其損失降至最低。 1.4 對抗蝦病毒之策略 欲降低蝦病毒對養蝦產業所造成之衝擊,除了良好檢測工具的開發 外,對抗蝦病毒方法的研究也極具重要性。目前主要的作法有: 免疫能力 的增加、疫苗的開發、病原體的隔離及環境的控制,[54] 另外還有無特定 病原 SPF (Specific Pathogen-Free) 、對特有病原體有忍受性之種蝦的開發 SPR(Specific Pathogen Resistant)、及對病原的中和 (neutralization) 。以下對 這一些方法作簡略介紹:
(1) 免 疫 能 力 的 增 加 , 在 飼 料 中 加 入 一 些 免 疫 促 進 劑 ( 例 如 ß-1,3 glucan、lipopolysaccharide, LPS),藉以提升蝦子的免疫能力。[55, 56]
(2)口服疫苗的應用: 雖然蝦子不具有 adaptive immune response,但在 最近的研究中,發現一現象,quasi-immune response,可以對抗 WSSV。 [57] 因此可以利用失活的病原體,或是病原體的重組蛋白,來誘發蝦子對 病原體的抵抗能力。[58, 59] (3)病原體的隔離與環境的控制: 病原體可隨著宿者 [活體 (蝦子本身或 是昆蟲等) 或是冷凍食品] 四處散播,所以對於運送的方法就要有所注意。 而養殖所用的水,因為裡面也可能含有病原體,所以在使用前需要經過處 理。
(4)抗病種蝦及無特定病源種蝦的篩選: 可分析比對較高存活率與較低 者,其各基因之表現量。最終可以經由這些基因的表現量,來判斷該品系 是否能對抗病體。[60, 61, 62, 63]
(5)病原體的中和反應: 利用抗體技術尋找病原體感染蝦子時所需要的 結合位置,藉由抗體來中和病原體與蝦子的結合位,達到保護蝦子的目 的。例如黃頭病毒 (Yellow head virus, YHV) ,其表面蛋白 gp116 就是 YHV 與蝦子感染時的表面結合位,利用抗體中和這個蛋白,可降低 YHV 對蝦子的感染。[64] 1.5 本研究之目的 目前所發展之檢測方法,主要是以聚合酶鏈鎖反應方法、DNA 探針 和免疫學方法為主。但前兩者在操作上,需要有經訓練之人員、及昂貴的 機器。而使用免疫學方法則可發展成容易操作且不需機器,即可對檢體作 檢測。在免疫學方面,主要是取病原體之表面蛋白,或是以全病原體當作 抗原,用以在小鼠上引發免疫反應,再利用細胞瘤融合技術,作成單株抗 體。利用這些抗體,可辦認病原體之某些蛋白的性質,可發展成檢測試 劑、或是作為發展抗病藥物之研究基礎。 現今全球的蝦養殖產業,正因病毒的影響,造成產能嚴重損失。為了 解決病毒問題,我們的目標是發展出方便、快速、便宜之檢測工具,作為 評估現場狀況,讓養殖漁夫可在第一時間,作出最為符合經濟效益的判 斷。也可作為無特定病原蝦 (SPF) 的篩選工具。另外,也可透過抗體與病 毒表面蛋白間的結合關係,探討病毒感染宿主的機制。最終,對此發展可 有效對抗病毒之藥物。 因 為 病 原 體 之 純 化 不 易 , 所 以 利 用 基 因 重 組 技 術 , 表 現 蝦 病 毒 HPV、IHHNV、TSV 之蛋白,當作抗原。再利用融合瘤技術(hybridoma
technique),生產針對這些蛋白具有專一性之抗體。再利用這些抗體開發出 一快速、便宜、簡單、靈敏之檢測工具。 作為抗原的病毒蛋白,主要以殼蛋白為主。(1)TSV 方面,目前已有 五十一筆核苷酸資料,我們從中選取 AY826054,為一殼蛋白。(2)IHHNV 方 面 , 目 前 已 有 十 七 筆 核 苷 酸 資 料 , 從 NC002190 取 一 段 Gene ID: 1457864 之基因片段,為殼蛋白。(3)HPV 方面,有十一筆核苷酸資 料,從 NC007218 選取一段 GENE ID: 3562190 之基因片段。
附錄一、實驗設計流程圖
NCBI database search sequence of HPV、IHHNV、TSV protein gene I 給予小鼠抗原 PCR amplified Gene fragment
Sub clone into pET28a vector BL21(DE3) expressed Purification of protein via Ni column 細胞融合 ELISA 篩選 Antigen: protein 單株抗體 抗體細胞 小鼠腹腔注射 收集腹水 免疫金膠 抗體配對 檢體檢測 抗原製備
第二章 材料與方法
2.1 抗原的製備
2.1.1 病毒殼蛋白 (coat protein) 基因的選取
由 NCBI database,分別選取 IHHNV (附錄三)、HPV (附錄四)、TSV
(附錄五) 三種病毒,所欲表現之基因片段。
2.1.2 引子的設計 IHHNV
切位
Forward primer 5-ggAATTC-CATATg-ATgTgCgCCgATTCAAC-3' NdeI Reverse primer 5-CCg-CTCgAg-TTAgTTAgTATgCATAA-3' XhoI
HPV
切位
Forward primer 5-ggAATTC-CATATg-ATggCgAgCAAAggTgA-3' NdeI Reverse primer 5-CCg-CTCgAg -TTACCTggCgAAACATT-3' XhoI
TSV
切位
Forward primer 1 5-CCCCAAAACACgTgTgAggAgAAAgTCCC-3’
Forward primer 2 5-AAgTAgACAgCCgCgCT-3’
Forward primer 3 5-ggAATTC-CATATg-ATggCTAACCCAgTTgAAAT-3’ NdeI Reverse primer 5-Cg-ggATCC-TTAgACAATCCCCTgTTTAg-3’ BamHI
2.1.3 表現載體: pET28a, Novagen
pBacPAK8-MTEGFP, 國 衛 院 , 用 於 桿 狀 病 毒 表 現 系 統 (baculovirus
2.1.4 表現菌株: BL21(DE3), Novagen
可表現出帶有 His-tag 的融合蛋白 (fusion protein) 。
2.1.5 病毒基因體的抽取及確認
使用病毒檢測試劑組 (Pathotech, Magellan Biotechnology Co., Ltd.) 抽
取,由病蝦體上取下約 25-50 mg 組織,放到 1.5 ml 微量離心管,加入 500 µl Tissue Lysis buffer,用玻棒攪碎。使用桌上離心機離心 15 分鐘,轉速 10,000 rpm。取上清液 200 µl,加入 200 µl viral DNA binding buffer 及 50 µl Protease K (20 mg/ml),放到一新的微量離心管中,並反覆倒置混合。72℃ 靜置 10 分鐘。再加入 100 µl isopropanol,並再反覆倒置混合。將混合液置 入含有樹脂的 Filter tube 中,離心 1 分鐘,轉速 10,000 rpm。倒去過濾 液,再加入 500 µl Wash bufferⅠ,到 Filter tube 中,離心 1 分鐘,轉速 10,000 rpm。倒去過濾液,再加入 450 µl Wash bufferⅡ到 Filter tube 中,離 心 1 分鐘,轉速 10,000 rpm,倒去過濾液,再離心 30 秒,轉速 13,000 rpm 。 最 後 將 Filter tube 置 於 新 的 微 量 離 心 管 中 , 加 入 50 µl Elution buffer,並靜置 5 分鐘,離心 1 分鐘,轉速 10,000 rpm,即可收到病毒 DNA,存放於-20℃備用。
使 用 試 劑 組 中 所 附 之 六 種 病 毒 (WSSV 、 HPV 、 IHHNV 、 TSV 、 YHV、MBV)引子, 確認含有那些病毒。
PCR, 聚合鏈反應條件
WSSV、 HPV、 IHHNV、 MBV (DNA virus)
引子 2 μl
病毒 DNA 2 μl
滅菌二次水 16 μl
20 μl
94℃ 2 分鐘 94℃ 30 秒 55℃ 1 分鐘 72℃ 1 分鐘 35 cycles 72℃ 5 分鐘 4℃ ∞
YHV、TSV (RNA viral) RT-PCR (反轉錄聚合鏈反應) 引子 2 μl 病毒 RNA 2 μl 滅菌二次水 16 μl 20 μl *試劑組中所附紅色小管,內含反轉錄酶, dNTP (粉狀) 60℃ 5 分鐘 42℃ 30 分鐘 94℃ 30 秒 57℃ 30 秒 72℃ 1 分鐘 35 cycles 72℃ 5 分鐘 4℃ ∞ Nest-PCR (巢式聚合鏈反應) 引子 2 μl RT-PCR 產物 10 倍稀釋 2 μl 滅菌二次水 16 μl 20 μl *試劑組中所附紅色小管,內含反轉錄酶 (粉狀)
94℃ 2 分鐘 94℃ 30 秒 56℃ 30 秒 72℃ 30 秒 35 cycles 72℃ 7 分鐘 4℃ ∞ 2.1.6 蛋白質重組 2.1.6.1 材料 限制酵素 NdeI、XhoI、BamHI、SacI、NotI (購自 NEB)
100bp、1kb DNA marker, dNTP、10X PCR buffer、Taq DNA polymerase (5U/ul) 、Loading buffer 和 Plasmid Prep Kit (購自禾鑫公司)
SYBR Green I (10X)(購自 Life) ,使用 DMSO 將 10000X 稀釋成 10X 備用 T4 DNA ligase (購自進階公司)
Agrose gel (USB, 購自季勗公司)
Gel Band Purification Kit (購自 amersham 公司)
50X TAE buffer
Tris base 242 g, Glacial Acetic Acid 57.1 ml, 0.5 M EDTA, pH8.0 溶於 1 L 二 次水中,使用時稀釋成 1X
LB 培養液
10 g Bacto-Tryptone、5 g yeast extract 以及 5 g NaCl 溶在 1 L 二次水中,滅 菌備用
LB 培養基 每升 LB 培養液加入 15 g Agar 經過高溫滅菌後等冷卻至 70 ℃以下再倒盤 (~15 ml/ 每盤) 備用 Kanamycine 配製 : 取 250 mg kanamycine 溶於 10 ml 二次水中,使用 0.45μm filter 過濾,分裝備用 LB (kan)培養液 (基),kanamycine 最終濃度為 25 μg/ ml 2.1.6.2 基因片段放大 2.1.6.2.1 IHHNV、HPV 引子 (Forward,10 μM) 2 μl 引子 (Reversed,10 μM) 2 μl viral DNA 2 μl dNTP (2 mM) 2 μl 10X PCR buffer 2 μl
Taq polymerase (5U/μl) 0.5 μl
滅菌二次水 9.5 μl 20 μl 94℃ 2 分鐘 94℃ 30 秒 54℃ 30 秒 72℃ 1 分鐘 40 cycles 72℃ 5 分鐘 4℃ ∞
2.1.6.2.2 TSV RT-PCR 引子 (Forward 1or2,10 μM) 2 μl 引子 (Reversed,10 μM) 2 μl viral DNA 2 μl 滅菌二次水 14 μl 20 μl *使用病毒檢測試劑組中之紅色小管,內含反轉錄酶, dNTP 60℃ 5 分鐘 42℃ 30 分鐘 94℃ 5 分鐘 94℃ 30 秒 54℃ 45 秒 72℃ 1 分鐘 40 cycles 72℃ 5 分鐘 4℃ ∞ NEST-PCR 引子 (Forward3,10 µM) 2 μl 引子 (Reversed,10 µM) 2 μl RT-PCR 產物 10 倍稀釋 2 μl dNTP(2 mM) 2 μl 10X PCR buffer 2μl Taq polymerase(5U/µl) 0.5 μl 滅菌二次水 9.5 μl 20 μl 94℃ 2 分鐘 94℃ 30 秒 56℃ 30 秒 72℃ 30 秒 40 cycles 72℃ 7 分鐘 4℃ ∞
2.1.6.3 表現質體的建構
將載體 (Vector, pET28a) 和欲殖入的 DNA 序列 (Insert) ,IHHNV 和
HPV 利用 NdeⅠ、XhoⅠ兩個限制酵素;TSV 利用 NdeⅠ、BamHI 兩個限 制酵素 (pBacPAK8-MTEGFP-TSV,則是利用 SacI、NotI 將 TSV 基因片段 由 pCR-TOPO vector 上切下,接入 pBacPAK8-MTEGFP vector 中) ,於 37 ℃之恒溫水槽作用 (Digestion) 16 小時。再將 DNA 取出,加入 1 μl Loading buffer 及 1 μl SYBR GreenⅠ,利用 1%洋菜膠 (Agrose gel),電壓 100 伏特進行電泳分析,把正確大小的 DNA 及載體片段切下,使用 Gel Band Purification Kit (Amersham),將膠體中之 DNA 純化出來。純化出來之 基因片段及載體,以 6 : 1 (體積) 混合後,抽乾,用 10μl 二次水回溶,再 加入接合酵素 (T4 DNA ligase, progema),在 4℃下作用約 16 小時。再進行 轉形作用,將建構好之表現質體,送入 BL21(DE3)菌株中。
2.1.6.4 轉形作用
取 5 μl 接合產物,加到 100 μl BL21(DE3) 的勝任細胞 (competent
cell)中,在冰上靜置 30 分鐘。Heat shock,置於 42℃熱水中 1 分鐘,馬上 置於冰中 2 分鐘。加入 900 μl LB 培養液,於 37℃振盪培養 (200 rpm)1 小 時。取 200 μl 塗於 LB/kna 培養基上,在 37℃隔夜培養。挑單一菌落至 3 ml LB/kna 培養液中,37℃,振盪培養 (200 rpm) 16 小時,利用 Plasmid Prep Kit (GeneMark),將建構好之質體抽出,並用原先作接合之限制酵 素,來確認插入之 DNA 片段是否有接入載體之中。
2.1.7 表現與純化
IHHNV、HPV、TSV 三個基因均接在 pET28a,利用 IPTG (isopropyl-beta-D-thiogalactopyranoside) 誘發載體,使其表現所接入之基因片段。而 純化方面,均利用載體上所共同表現出來之 his-taq,使用 Nickel column 作 純化。
2.1.7.1 材料
IPTG (sigma, 購自季勗公司) Nickel resin (購自 Novagen)
PBS buffer
37g Na2HPO4, NaH2PO4 0.35g, NaCl 8.77g 溶於 1 L一次水中
Lysis buffer
NaCl 0.2 M, Tris-HCl 20 mM, Imidazole 50 mM, pH 7.9
Guanidine-Lysis buffer
6 M Guanidine Hydrochloride, NaCl 0.2 M, Tris-HCl 20 mM, Imidazole 50 mM, pH 7.9
Wash buffer
NaCl 0.2 M, Tris-HCl 20 mM, Imidazole 80 mM, pH 7.9
Elution buffer
透析液Ⅰ
0.25 M Guanidine Hydrochloride, NaCl 0.2 M, Tris-HCl 20 mM, Imidazole 50 mM, pH 7.9
透析液Ⅱ
NaCl 0.2 M, Tris-HCl 20 mM, Imidazole 50 mM, pH 7.9
2.1.7.2 表現
取菌養於 5 ml LB (kna) 培養液中,37℃振盪培養 (200 rpm) 16 小時
後。以 1:100 體積比,轉養至 300 ml LB (kna) 培養液中,37℃振盪培養 (200 rpm) 2~3 小 時 , 直 到 OD600=0.4~0.6 後 。 加 入 IPTG (final concentration 1mM) 誘導。IHHNV 在 25℃振盪培養 (200 rpm) 10 小時、 TSV 在 37℃振盪培養 (200 rpm) 10 小時、HPV 在 37℃振盪培養 (200 rpm) 6 小時後。以 4℃,轉速 9000g,離心 15 分鐘,倒掉上清液,菌體保存在-20℃中備用。 2.1.7.3 純化 2.1.7.3.1 IHHNV、TSV 取 100 ml 培養液所離心下來之菌體,加入 15 ml Lysis buffer 回溶菌 體。使用超音波震碎方式破細胞 (震幅 30%, 打 1 秒停 2 秒,共打 10 分 鐘)。以 4℃,轉速 10,000 g,離心 15 分鐘,取上清液進行純化工作。 將約 2 ml Nickel resin 裝填到管柱 (直徑 1.5 公分)中,取 20 ml Lysis buffer 平衡管柱。之後將上個步驟中,含有蛋白質的上清液,慢慢加到管 柱之中,待流完時,再用 15 ml Lysis buffer 沖洗管柱一次。取 12 ml Wash
buffer,沖洗管柱一次。再取 12 ml Elution buffer 沖洗管柱,每 1.5 ml 收一 管。 2.1.7.3.2 HPV 取 100 ml 培養液所離心下來之菌體,加入 5 ml PBS buffer 回溶菌 體,使用超音波震碎方式破細胞 (震幅 30%, 打 1 秒停 2 秒,共打 10 分 鐘)。以 4℃,轉速 6000 g,離心 15 分鐘。倒去上清液,加入 5 ml PBS buffer 回溶沈澱物,再使用超音波震碎,條件同上。離心 15 分鐘,4℃, 轉速 6000 g,到去上清液,加入 5 ml Guanidine-Lysis buffer,使用超音波 震碎回溶沈澱物,靜置冰上 1 小時。以 4℃,轉速 16,000 g,離心 30 分 鐘。取上清液,使用針頭,利用重力,慢慢加入 15 ml Lysis buffer,並將 之裝入透析膜 (MW 25,000)中,在 250 ml 透析液Ⅰ中,4℃透析 4~6 小時 後。再將之換到 1 升透析液Ⅱ中,4℃透析約 16 小時後。再以 4℃,轉速 16000g,離心 30 分鐘,取上清液作純化。
將約 2 ml Nickel resin 裝填到管柱 (直徑 1.5 公分)中,取 20 ml Lysis buffer 平衡管柱。將上個步驟中,含有蛋白質的上清液,慢慢加到管柱之 中,待流完時,再用 15 ml Lysis buffer 沖洗管柱一次。取 12 ml Wash buffer,沖洗管柱一次。再取 12 ml Elution buffer 沖洗管柱,每 1.5 ml 收一 管。
2.2 單株抗體之製作
抗體是由 B 細胞所分泌的蛋白質,目前廣為應用的抗體分為兩種,多 株抗體(polyclonal antibody)和單株抗體(monoclonal antibody)。多株抗體是 將抗原打入老鼠體內,經過數次免疫,採集血液,收血清,此為多株抗 體。操作過程雖然簡單,但其專一性低,對於相似的抗原結構都會有所反 應,稱為交叉反應(cross reaction)。1973 年,Milstein 等人,提出骨髓瘤細
胞(myeloma cell)之融合實驗,製造出單株抗體。因為血清中的每一種抗 體,是由單一種 B 細胞所製造,所以將 B 細胞由脾臟中取出,與骨髓瘤細 胞融合,成為可持續分泌抗體,又可在培養基中永久生長的細胞株,即為 融合瘤細胞株(hybridoma cell)。 其過程為,給予小鼠外來抗原,並同時以免疫佐劑加強小鼠免疫反 應,使脾臟內的 B 細胞產生特定抗體,將其與癌化細胞株融合形成可體外 培養且生產特定抗體之細胞株(附錄二)。
附錄二、 單株抗體製作流程圖 第 0 天-將佐劑 ( complete adjuvant) 和抗原等體積混和注射入老鼠體內 第 10 天-第二次注射 第 20 天-第三次注射,解凍 FO 細胞,且大量繁殖 第 27 天-第四次注射 第 28 天- FO 細胞 1:2 繼代培養 第 30 天-採血測 titer,取小鼠脾臟與 FO 細胞做細胞融合 第 37 天-加 50 µL 之 HY-HAT 培養液 細胞數佔孔洞約 50 - 90 %,培養液轉黃取 50 µL,做 ELISA 篩選 陽性細胞群落單株化 以 ELISA 篩選陽性單株細胞群落 大量繁殖陽性單株細胞,收集培養液,做後續實驗用
2.2.1 材料
Name Component Trademark
Dulbecco’s Modified Eagle Medium (DMEM)
Contains 4500 mg / L D-glucose, L-glutamine, and 110 mg / L sodium pyruvate but no sodium bicarbonate.
GIBCO BRL®
12800-017
Fetal Bovine Serum (FBS) < 10 EU / mL endotoxin, < 10 mg / dL hemoglobin, 40 nm flitered. HYCLONE® Defined Grade Penicillin / Streptomycin (P/S)
Contains 10,000 units of penicillin (base) and 10,000 µg of
streptomycin (base) / mL utilizing penicillin G (sodium salt) and streptomycin sulfate in 0.85% saline.
GIBCO BRL®
15140-122
H-Y Medium (HY)
Contains L-glutamine, bovine insulin, oxaloacetate, and sodium pyruvate but no sodium
bicarbonate. SIGMA H-9014 HYBRI-MAX ® HAT Media Supplement (50 X) (HAT)
Reconstitute contents of vial with 10 mL sterile cell culture medium. When reconstituted to 10 mL,
each vial contains 5×10-3 M
hypoxanthine, 2×10-5 M aminopterin, 8×10-4 M thymidine. SIGMA H-0262 HYBRI-MAX® PEG / DMSO Solution (PEG / DMSO)
Contains 50% (w/v) PEG (Av. Mol. Wt. 1450) and 10% DMSO (v/v) in DPBS without calcium.
SIGMA P-7306
碳酸氫鈉 (NaHCO3) (購自sigma)
DEAE resin (購自 amersham)
PB buffer
1 M Na2HPO4 68.4 ml, 1 M NaH2PO4 31.6 ml補一次水至 5 L, 調pH至 7.2
2.2.2 免疫反應
以純化所得之病毒蛋白為抗原。利用 4-8 週的 BALB/c 小鼠為免疫動
物,以抗原(100 µg / mouse) 加等體積之佐劑 (Freund’s complete adjuvant)
(Sigma) 混合成乳劑,於 0 天做小白鼠腹腔注射(IP)。之後再以抗原(100 µg
/ mouse) 加等體積之佐劑 (Freund’s incomplete adjuvant) (Sigma) 混合成乳 劑,於 10、 20 及 27 天再進行小鼠腹腔注射 (IP),誘發免疫反應,使 B 細 胞成熟增生並產生抗體。第 30 天時要採血測價數,確認小白鼠已被引發 免疫反應,確認後再進行細胞融合。 2.2.3 骨髓癌細胞 (myeloma cell) 所用之骨髓瘤細胞株極易增生,且與脾臟細胞融合後,可迅速發育且 分泌高濃度免疫球蛋白。目前最常使用為HGPRT (hypoxanthine guanine phosphoribosyl transferase) 缺乏性骨髓瘤細胞,如P3、P3 / 653、NS1、 SP2 、 及 FO 等 細 胞 , 當 細 胞 培 養 於 HAT (hypoxanthine aminopterin thymidine) 培養液中,其中aminopterin可阻止骨髓瘤細胞核酸之正常代謝 途徑 (de novo synthesis) 之進行,使細胞僅能以 HGPRT或TK酵素,依救急 代謝途徑 (salvage pathway) 利用hypoxanthine及thymidine合成核酸,若骨髓 瘤細胞缺乏HGPRT,將無法合成核酸而死亡,而融合細胞因為自脾臟細胞 取得HGPRT之基因而得以存活增生 (參考圖 2-1)。實驗室採用FO細胞株,
採購於食品工業研究所,使用DMEM培養液,添加 0.018 M碳酸氫納、10
% FBS及 1 % penicillin / streptomycin,培養於 37 ℃,5 % CO2。
圖 2-1 細胞內核酸的合成路徑 核酸的正常代謝途徑若被 aminopterin 阻礙,可由 救急途徑 (salvage pathway) 取用 thymidine (T) 及 hypoxanthine (H) 來合成 DNA。 但 FO cell 缺乏 TK 及 HGPRT 兩種酵素,因此在 aminopterin (A) 存在下 FO cell 無法生長;正常細胞 (如 B 細胞) 則可經救急途徑繼續生長。HAT 培養基含 aminopterin, thymidine 及 hypoxanthine,FO cell 在 HAT 中無法生長,除非經由 細胞融合導入 TK 或 HGPRT 兩酵素的基因 (可由正常脾臟細胞得來)。 2.2.4 細胞融合 (Fusion) 小鼠以乙醚將其迷昏後,採眼窩血,無菌操作解剖取出脾臟,以 DMEM於培養皿中清洗兩次。將脾臟至於培養皿中以 5 mL針筒末端平坦 處將脾臟壓碎,用 21G針頭抽吸使細胞分散,經由無菌紗布過濾至 50 mL 離心管,離心 5 分鐘,轉速 260 g,去除上清,再加入 10 mL DMEM沖
洗,重複兩次。同時取FO細胞,以DMEM沖洗細胞兩次,離心 5 分鐘,轉 速 260 g,回溶於 10 mL DMEM中。取部分細胞做細胞計數,用trypan blue 液計算活細胞率,以脾細胞:FO細胞=4:1 之比例將細胞混合,離心 5 分鐘,轉速 260 g,去上清液 。離心管輕擊桌面震散細胞,於 1 分鐘內緩 慢滴入 1 mL已加溫到 37 ℃之PEG / DMSO,同時輕輕搖晃細胞使其均勻 混合,於 37 ℃水浴 1 分鐘後取出,在 2 分鐘內緩慢滴入 10 mL已加溫到 37 ℃之DMEM,離心管輕擊桌面邊敲邊加,離心 5 分鐘,轉速 180 g,去 上清液,再以 20 mL DMEM沖洗細胞兩次。,以HY培養液 (已添加HAT、 0.018 M 碳酸氫納、20 % FBS及 1 % P / S) 懸浮細胞,並於 96 孔細胞培養
盤 (96 well culture plate)培養細胞,在 37 ℃,5 % CO2之條件進行融合後細
胞培養。
2.2.5 細胞增殖與取樣篩檢
融合後培養之細胞定期觀察其生長狀況,融合後第 7 天,需再補充 HY-HAT 培養液。14 天後檢查培養液之顏色,若有融合成功有融合瘤細胞 (hybridoma cell) 產生,則培養液顏色變黃,取其培養液做 ELISA 篩檢。太 早做篩檢易漏失陽性細胞,因其分泌出的抗體濃度尚低,但若太遲篩檢, 融合細胞會因過度增生而死亡。因此最好在細胞未長滿前篩檢一次,其陰 性者在培養液變黃時再篩檢一次。培養液經 ELISA 篩檢出含有抗體時,每 一次 ELISA 需作一個負向對照組實驗,將該細胞繼續培養。 2.2.6 抗體之生產 單株抗體之生產是使用組織培養法,組織培養法是以 175 cm2塑膠培 養盤 (T75) 大量培養,直到細胞過飽和開始死亡時,離心除去細胞,收集 上清液,保存在 -20 ℃冰箱中備用。
2.2.7 腹水抗體製備 (Ascetic fluid) 6~11 週大的BALB/C小鼠為實驗動物,第 0 天小鼠腹腔注射給予 0.5 ml Pristane (Sigma)。第 7 天收集單株抗體細胞以PBS清洗兩次後,小鼠腹 腔注射單株抗體細胞,3x106 cell/mouse。第 21 天觀察小鼠,腹部腫脹且行 動不便者,以 18 號針頭配 5ml針筒腹腔收集腹水,可收 2~3 次。將收集的 腹水離心去除組織碎片,最上層白色懸浮物為脂質先去除,收上清液。加 入 50﹪的硫酸銨 (ammonia sulfate),分三次加入,每次間隔 30 分鐘,最後 在 4℃中攪拌反應 2 小時,離心 30 分鐘,4℃,轉速 1620 g,倒掉上清 液,沈澱物以適量PB buffer回溶,再用PB buffer透析 16 小時。離心 30 分 鐘,4℃,轉速 1620 g,上清液用 0.45µm filter過濾備用。 使用 DEAE 管柱 (管徑 2.5 公分)進行純化,裝填 20 ml DEAE 樹脂, 加入 80 ml PB buffer 平衡管柱。將硫酸銨沈澱所得之過濾液倒入,將流過 管柱之液收起,大部份抗體因為不與 DEAE 樹脂結合,而分佈在這個部 份。再分別用含有 40mM、60m NaCl 之 PB buffer,各 40 ml 沖洗管柱,每 1.5 ml 收集一管。最後再用含有 1M NaCl 之 PB buffer 50 ml 沖洗管柱。 2.3 抗體的測定 2.3.1 材料
PBS: 1.37 g Na2HPO4, NaH2PO4 0.35 g, NaCl 8.77 g 溶於 1 L一次水中
goat anti-mouse conjugated HRP antibodies (KPL) TMB (KPL)
Blocking buffer
ABTS (2’-azinobis-3-ethylbenzthiazoline-6-sulfonic acid)
取 22 mg ABTS 溶於 100 ml 的 50 mM sodium citrate (pH 4.0)中,過 0.22
μm的filter中,置於 4℃備用。每次反應取 21 ml加入 36 μl 30% H2O2. DAB (3,3-Diaminobenzidine) 20 mM 3,3-Diaminobenzidine及 0.15% H2O2溶於PBS中 SDS running buffer 3.03 g Tris, 144 g glycine, 10 g SDS 溶於 1 L 一次水中 5× SDS loading buffer 二次水 4 ml, 0.5 M Tris-HCl (pH6.8) 1.7 ml, Glycerol 5 ml, 20% (w/v) SDS 0.5ml, 2-mercaptoetyhanol 2 ml, 0.05% (w/v) bromophenol blue 4~8 mg.
Coomassie Blue staining solution
400 ml methanol, 100 ml acetic acid, 0.1 g Coomassie Brilliant Blue R250 溶在 1 L 一次水中
退染溶液 I
400 ml methanol, 100 ml acetic acid, 補一次水至 1 L
退染溶液 II
50 ml methanol, 70 ml acetic acid, 補一次水至 1 L
西方點墨轉漬緩衝液 transfer buffer
2.3.2 ELISA(enzyme-linked immunosorbent assay)
先以老鼠原倍血清 (IgG) 當作正向控制 (positive control),blocking buffer作負向控制 (negative control)。實驗進行時,以純化所得之病毒蛋白 為抗原,於 96 孔盤中每個孔洞加入 100 µL,室溫下靜置 1.5 小時,以洗滌 液 (PBS) 洗 去 抗 原 溶 液 , 重 複 洗 三 次 後 , 加 入 blocking buffer 400 µL /well,室溫下靜置一個小時後,以洗滌液洗去blocking buffer,重複洗三
次,再加入以洗滌液稀釋的待測抗體即細胞培養液或腹水 100 µL /well,
室溫下靜置一個小時後,以洗滌液洗去待測抗體,重複洗三次,加入以洗 滌液稀釋的二級抗體(如goat anti-mouse conjugated HRP antibodies 100 µL /well),室溫下靜置一個小時後,以洗滌液洗去二級抗體,重複洗三次,再 以PBS洗一次,最後加入呈色劑(TMB或是ABTS) 50 µL /well,室溫下避光
反應 20 分鐘,使用 2 M H2SO4 , 50 µL /well終止反應,以ELISA reader
(Fusion Universal Microplate Analyzer, Packard)波長 450 nm (ABTS用 405 nm)讀取數值,即可知待測抗體是否能辨識抗原。
2.3.3 SDS-PAGE
在製膠器具中,加入約 3-4 mL resolving gel 於兩片玻璃間的溝槽中, 再以 70 % 酒精覆蓋其表面,靜置約 25-30 分鐘後待其凝結完全。去除酒 精,注入適當體積的 stacking gel,comb 立即緊貼玻璃面置入 stacking gel,靜置 25-30 分鐘。待其凝結後移去 comb。將凝結完成的膠片,架設 於電泳槽中,倒入 SDS running buffer,將樣品蛋白質與 5× SDS loading buffer 混合後,95℃熱浴 5 分鐘,注入孔中,以 100 伏特進行電泳,至電 泳結束。膠片浸泡於 Coomassie Blue staining solution 中 30-60 分鐘,換至 退染溶液 I 中約 1 小時,去除多餘背景染料之後,再換浸泡於退染溶液 II 以去除多餘背景染料。
12% SDS-PAGE, Separating gel ddH2O 1.6 ml 30 % acrylamide mix 2 ml 1.5 M Tris (pH8.8) 1.3 ml 10 % SDS 0.05 ml 10 % ammonium persulfate 0.05 ml TEMED 0.002 ml total volume 5 ml
5% SDS-PAGE, Stacking gel
ddH2O 0.68 ml 30 % acrylamide mix 0.17 ml 1.0 M Tris (pH6.8) 0.13 ml 10 % SDS 0.01 ml 10 % ammonium persulfate 0.01 ml TEMED 0.002 ml total volume 1 ml 2.3.4 Western blot (西方點墨) 硝化纖維膜與紙張先浸泡於轉漬緩衝液 transfer buffer 中,將完成的 SDS-PAGE 膠片置於硝化纖維膜上,上下再夾以三張紙張,置入溼式電傳 器中 (Mini Trans-Blot Cell, Bio-Red),90V,60 分鐘。完成後置入 blocking buffer 中,室溫下搖晃一個小時後,以 PBS 洗去 blocking buffer,重複洗三 次,再置入以 PBS 稀釋的一級抗體(待測抗體),室溫下搖晃一個小時後, 以 PBS 洗去一級抗體,重複洗三次,置入以 PBS 稀釋的二級抗體 (如 goat anti-mouse conjugated HRP antibodies),室溫下搖晃一個小時後,以洗滌液 洗去二級抗體,重複洗三次,後加入 DAB 呈色劑呈色。
2.4 儀器與設備
UVS400, Thermo Savant SPD SpeedVac
BECKMAN COULTER Allegra 21R Centrifuge ORBITOR SHAKER
Centrifuge 5415R, eppendorf
Orbital shaking incubator Model S300R, FIRSTEK SCIENTIFIC 550 sonic dismembrator, FIRSTEK SCIENTIFIC
Mini Trans-Blot Cell, Bio-Red
Fusion Universal Microplate Analyzer, Packard
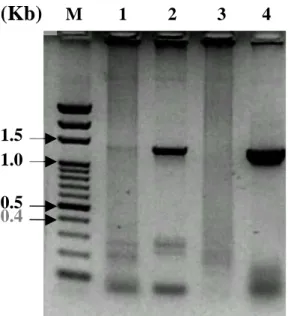
第三章 結果與討論 3.1 病毒基因片段取得 所得到之病毒基因片段均進行定序實驗,且證實所得之 DNA 片段序列 無誤。 3.1.1 IHHNV 使用商業用檢測試劑組所抽得之基因體當作模板,再用所設計之引子 進行 PCR 反應,即可得到一長度為 990bp 之基因產物 (圖 3-1-1)。 M 1
(Kb) Lane M: DNA marker (100bp, fermentas)
Lane 1: IHHNV PCR 產物,長度 990bp
1.0 0.8 0.8
3.1.2 HPV 使用檢測試劑得知,有四隻檢體體內具 HPV (檢體編號#1、2、 4、 5) ,因此將這四隻檢體所抽得之基因體,使用所設計之引子,進行 PCR 反應,其中,有三組可以得到預期之基因片段長度 (檢體編號#1、2、5) , 但其中一組所得之量甚少 (圖 3-1-3,Lane 1) 。此外,另有一組,在檢測後 得知有 HPV 存在,但在使用實驗室所設計之引子進行 PCR 反應後,卻無 法得到預期之基因片段 (檢體編號#4) ,因為其它兩組檢體 (檢體編號#2、 5) ,提供可放大出基因片段之模板,因此選定編號#5 之檢體,進行後續之 實驗。 圖 3-1-2 HPV 的確認 0.4 0.5 M 1 2 3 4 5 6 (Kb)
Lane M: DNA marker (100bp, fermentas) Lane 1~5: 五隻不同蝦子檢體,編號 #1~5,Lne 6: positive control,若檢體中 具有 HPV, 即可放大出一相同大小的片 段,該片段長 441bp。結果看來,五隻 檢體中,有四隻具有 HPV
圖 3-1-3 HPV PCR 產物
(Kb) M 1 2 3 4
Lane M: DNA marker (100bp, fermentas) Lane 1~4: 四個不來源,檢體編號為#1、 2、4、5(參考圖 3-2)的 HPV, 進行 PCR, 得到基因片段 1039bp,其中 Lane 4 所得 量大,以此檢體(編號#5)為來源進行後續 實驗。 0.4 0.5 1.5 1.0
3.1.3 TSV
TSV 屬於 RNA 病毒,因此需經由反轉錄之過程,以取得 cDNA,再 由其為模板,放大出所選取之基因片段。亦因如此,需在所選取之基因片 段前後,設計新的引子 (TSV Forward primer 1 and Forward primer 2) ,利用 反轉錄酵素,以放大出長度大於目標基因片段之 DNA 片段。再利用這個 片段,進行 Nest-PCR,以增加放大量。在圖 3-1-4 中,Lane 1 為使用 TSV Forward primer 1 和 TSV Reversed primer 所放大,其長度為 947bp。而在 Lane 2 則是利用 TSV Forward primer 2 和 TSV Reversed primer 所大出, 其長 度為 903bp。再利用這兩段 RT-PCR 產物當作模板,進行 Nest-PCR,不論 使用何者當模板,均可得到一片段長度為 876bp 的 DNA 片段 (圖 3-1-5) 。 圖 3-1-4 TSV RT-PCR 產物 圖 3-1-5 TSV Nest-PCR 產物 M 1 2 (Kb) 1.0 0.8 M 1 2 (Kb)
Lane M: DNA marker (100bp, fermentas)
Lane 1:使用 TSV foreard 1 及 TSV reversed primer 所作之 PCR 結果,可放大出一長 947bp 的片段
Lane 2: 使用 TSV foreard 2 及 TSV reversed primer 所作之 PCR 結果,可放大出一長 903bp 的片段
0.2 0.3 1.5 1.0
Lane M: DNA marker (100bp, fermentas) Lane 1:使用 947bp RT-PCR 產物當模板 Lane 2:使用 903bp RT-PCR 產物當模板
結果指出,不論用何種模板,均能放大出一長度 876bp 的 基因片段
3.2 重組蛋白的表現與純化 3.2.1 IHHNV 此一重組蛋白,在 37℃表現時,雖然有大量表現,但會形成 inclusion body。因此改在 25℃表現,雖然表現量降低,但較易於純化 (圖 3-2-1) 。 此外,此一重組蛋白易被分解,純化出來後,置於 4℃中保存,約七天再 拿出來進行蛋白質電泳時,原本單一蛋白質帶會變成兩條。純化出來的蛋 白質,亦有以 anti-his tag 抗體進行西方墨點實驗時,可以為其所辦認 (圖 3-2-2),所以初步判定,已能純化出此一重組蛋白,蛋白質分子量大小, 由表現載體與目標基因切接後,估計為 37 Kda。 97 66 45 30 20.1 14.4 (kDa) M 1 2 3
Lane M: Protein marker (amersham) Lane 1: lysate of BL21(DE3)(pET28a) Lane 2: lysate of BL21(DE3)
(pET28a-IHHNV)
Lane 3: 純化的 IHHNV 重組蛋白,分子量 估計為 37kDa
圖 3-2-1 IHHNV 重組蛋白的表現與純化
圖 3-2-2 IHHNV 重組蛋白與 anti-his taq 抗體西方點墨實驗
M 1 (kDa) 105 75 50 35 30
Lane M: Protein marker (amersham) Lane 1: IHHNV 重組蛋白
分子量估計為 37kDa
3.2.2 HPV HPV 重組蛋白,在 37℃表現時,會形成 inclusion body。改為 25℃表 現時,表現量降低甚多,與對照組相比,無法看出差異,僅能使用 anti-his tag 進行西方墨點實驗時,才能看出此重組蛋白分佈在上清液中。在 25℃ 表現雖然不會現在 inclusion body,但表現量太低。因此仍在 37℃表現, 使用 Guanidine HCl 將其失活 (denature) 之後,再利用透析方式使其再重疊 (refolding)。在再重疊的過程中,Guanidine HCl 濃度的改變必需要很緩 慢,若改變太快,容易產生沈澱。透析完後,仍有部份沈澱,離心完後, 此時上清液中,HPV 重組蛋白在全體蛋白質中已佔有很高的比例 (圖 3) 。使用 anti-his tag 抗體進行西方墨點實驗時,可以為其所辦認 (圖 3-2-4) ,所以初步判定,已能純化出此一重組蛋白,蛋白質分子量大小,由表 現載體與目標基因切接後,估計為 50 Kda。 圖 3-2-3 HPV 重組蛋白的表現與純化 圖 3-2-4 HPV 重組蛋白與 anti-his taq 抗體西方點墨實驗 105 75 50 35 (kDa) M 1 97 (kDa) M 1 2 3
Lane M: Protein marker (amersham) Lane 1: lysate of BL21(DE3)(pET28a) Lane 2: lysate of BL21(DE3)
(pET28a-HPV) Lane 3: 純化的 HPV 重組蛋白,分子量 估計為 50kDa 45 66 30 20.1 14.4
Lane M: Protein marker (fermentas) Lane 1: HPV 重組蛋白
3.2.3 TSV TSV 重組蛋白,在 37℃表現時,與對照組相比,無法看出差異,僅能 使用 anti-his tag 在西方墨點實驗中,證實該重組蛋白可能有表現。但表現 量很低,圖 3-2-4 中所純化之重組蛋白,經濃縮約六倍體積後才有如此結 果,蛋白質分子量大小,由表現載體與目標基因切接後,估計為 31 Kda。 也嚐試將此基因片段接入 PBACPAK8-MTEGFP 載體中,使用 baculovirus expression system 作表現 (附錄六),但初步實驗結果看來,與對照組相 比,似乎也是沒有表現 (圖 3-2-5) 。 97 66 45 30 20.1 14.4 (kDa) M 1 2 3
Lane M: Protein marker (amersham) Lane 1: lysate of BL21(DE3)(pET28a) Lane 2: lysate of BL21(DE3)
(pET28a-TSV) Lane 3: 純化的 TSV 重組蛋白,分子量估 計為 31kDa 圖 3-2-5 TSV 重組蛋白的表現與純化 圖 3-2-6 TSV 重組蛋白在桿狀病毒系統中的表現 103 77 50 34.3 28.8 20.7 M 1 2 (kDa)
Lane M: Protein marker (Bio-rad) Lane 1: lysate of Hi 5 cell as control Lane 2: lysate of Hi 5 cell (TSV) 分子量估計為 31kDa
3.3 抗體的製作 3.3.1 單株抗體力價的測試 當小鼠給予抗原後產生免疫反應,可測其血液中抗體力價(titer)高低, 決定是否適合進行融合實驗,通常給予抗原三次後,血液中抗體力價可達 高峰,因此在第 30 天給予抗原後,第 34 天眼窩採血,血液離心取血清連 續稀釋作 ELISA 實驗,值趨平緩者為其價數,由結果得知四次給予抗原小 鼠的血中力價,使用 HPV 重組蛋白者,可達 1 : 20000 (圖 3-3-1);使用 IHHNV 重組蛋白者,可達 1 : 50000 (圖 3-3-2)。 3.3.1.1 HPV 重組蛋白單株抗體力價之測式 0 0.5 1 1.5 2 2.5 0 10000 20000 30000 40000 50000 60000 Dilution factor A b so rb an ce o f O D 450 nm 圖 3-3-1 實驗接種小鼠血清力價之測試,HPV 重組蛋白 3.3.1.2 IHHNV 重組蛋白單株抗體力價之測試 0 0.5 1 1.5 2 2.5 3 3.5 0 10000 20000 30000 40000 50000 60000 Dilution factor Ab so rb an ce o f OD 450 nm 圖 3-3-2 實驗接種小鼠血清力價之測試,IHHNV 重組蛋白
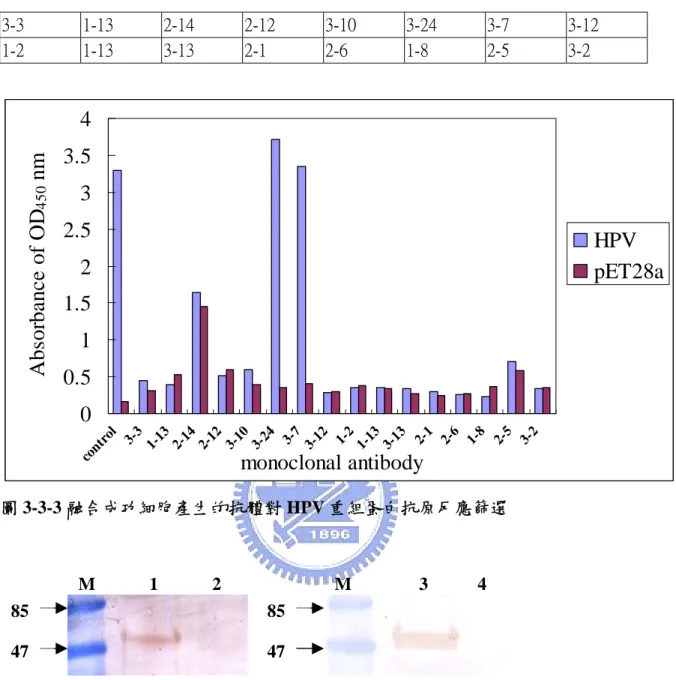
3.3.2 單株抗體對抗原反應篩選 3.3.2.1 HPV 在 HPV 方面,因為表現量很高,所以抗原的製備,是將蛋白質粗抽液 跑完 SDS-PAGE 後,經染色,將預期位置的帶切下,使用液態氮讓其結 凍,再將其磨碎。如此反覆數次,直至膠體顆粒大小可流過 23G 針頭,以 便於打到老鼠中。但免疫三次後,測血清價數,未達可作抗體之數,所以 再以粗抽蛋白免疫一次。因為粗抽蛋白中,尚有其它雜蛋白,所以會有認 其它蛋白的抗體。因此在篩選時,會使用 E. coli BL21(DE3)(pET28a)表現 後的粗抽蛋白當作負向選取 (negative selection),因為這兩組所表現之蛋白 質,理論上只差重組蛋白,所以可以利用這樣的方式,保留只會認重組蛋 白質的細胞株。所得到的十六株抗體 (表 3.1) ,由 ELISA 數據看來 (圖 3-3-3) ,均會認到其它雜蛋白,但其中,3-24、3-7 這兩株價數高且負向選取 的值也較低,代表這兩株細胞所生產之抗體,較不會認到雜蛋白,有較高 的專一性。雖然這兩株細胞,其負向選取的值與負向控制比起來,相對 高,但在西方點墨實驗中 (圖 3-3-4) ,卻僅認到 HPV 重組蛋白,這可能跟 DAB 呈色靈敏度較差,導致所認到的雜蛋白無法呈現出來。這樣的推論在
ECL (enhanced chemiluminescent) 呈色結果可得到證實 (data 未呈現) ,所得 到之單株抗體均會認到重組蛋白以外的雜蛋白。出現這樣的結果,與未作 單株化有關,僅靠負向篩選,又在不使用純的蛋白當抗原時,無法得到僅 認重組蛋白之單株抗體。在蝦體檢測方面,初步選了 13 隻未含 HPV 的蝦 子,取其組織粗抽液當抗原,與 3-24 單株抗體進行西方墨點實驗 (圖 3-3-5) ,初步結果指出,此株僅認到 HPV 重組蛋白。雖然此一結果並不能證 明該抗體可與天然的 HPV 殼蛋白作辨識,但也可證明,不會非特異性地 辦認到蝦體中其它的蛋白。
表 3.1 HPV 重組蛋白 monoclonal antibodies 3-3 1-13 2-14 2-12 3-10 3-24 3-7 3-12 1-2 1-13 3-13 2-1 2-6 1-8 2-5 3-2
0
0.5
1
1.5
2
2.5
3
3.5
4
cont rol 3-3 1-13 2-14 2-12 3-10 3-24 3-7 3-12 1-2 1-13 3-13 2-1 2-6 1-8 2-5 3-2monoclonal antibody
Ab
so
rb
an
c
e
o
f OD
450nm
HPV
pET28a
圖 3-3-3 融合成功細胞產生的抗體對 HPV 重組蛋白抗原反應篩選 M 1 2 85 47 M 3 4 圖 3-3-4 HPV 重組蛋白與抗體西方點墨結果。Lane 1、3, HPV 重組蛋白;Lane 2、4: BL21(DE3)(pET28a)表現後之粗抽蛋白,當作負向選取 (negative selection) ,左圖用 3-7 單株抗體,右圖用 3-24 單株抗體。85 47
1 2 3 4 5 6 M 7 8 9 10 11 12 13 14 15 36 26 20 118 85 47 圖 3-3-5 HPV 3-24 抗體與蝦檢體之西方點墨實驗結果。Lane 1~13, 蝦子檢體粗抽液; Lane 14, HPV 重組蛋白; Lane 15, BL21(DE3)(pET28a)表現後粗抽蛋白,當作負向控制 (negative control)。 3.3.2.2 IHHNV 在 IHHVN 方面,目前已得到一株細胞株,3-62。在 ELISA 實驗中 (圖 3-3-6),其價數與老鼠原倍血清同高。而在西方點墨實驗中也證實,此 一單株抗體辨認到 IHHNV 重組蛋白,在病原蝦粗抽液中也可以辨認到 IHHNV 天然之殼蛋白 (圖 3-3-7)。 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 Ab so rb an ce o f OD 450 n m positive control (mouse serium) negative contorol IHHNV recombinant protein BL21(DE3) crude protein (negative selection) 圖 3-3-6 融合成功細胞產生的抗體對 IHHNV 重組蛋白抗原反應篩選
1 2 M 3 4 5 6 7 47
36
圖 3-3-7 IHHNV 3-62 抗體與蝦檢體之西方點墨實驗結果。Lane1, BL21(DE3)(pET28a) 表現後粗抽蛋白,當作負向控制 (negative control) ; Lane2, IHHNV 重組蛋白; Lane3~7: IHHNV 病蝦檢體粗抽液。 3.3.3 單株抗體特異性之測試 抗體與抗原之辨認途徑有二個,一個是蛋白質序列、一個是蛋白質結 構。為避免所發展之抗體,會非特異性地辨認到其它蝦病毒,因此將所得 到之兩株 HPV 抗體與一株 IHHNV 抗體,交义與 TSV、HPV、IHHNV 三 種重組蛋白質進行 ELISA (圖 3-3-8~10) 及西方點墨實驗 (圖 3-3-11~13)實 驗。在 (圖 3-3-8)中,使用 HPV 3-24 這株 HPV 抗體,只會專一性辨認到 HPV 重組蛋白。在 (圖 3-3-9)中,使用 HPV 3-7 這株 HPV 抗體,只會專一 性辨認到 HPV 重組蛋白,雖然對 IHHNV 重組蛋白有一點反應,這可能與 單株化不佳有關。在 (圖 3-3-10)中,使用 IHHNV 3-62 這株 IHHNV 抗體, 只會專一性辨認到 IHHNV 重組蛋白,雖對 HPV 重組蛋白也是有一點反 應,這可能也是與單株化不佳有關。而在西方點墨實驗中,也是有類似的 結果。因此初步看來,所得到這三株抗體,在不同的重組蛋白中,不會出 現非特異性的辨認。
HPV 3-24 antibody 0 0.51 1.5 2 2.5 3 3.5 4 4.5 Posi tive Nega tive Nega tive sele ction TSV HPV IHH NV Ab so rb an ce o f OD 450 n m 圖 3-3-8 HPV 3-24 抗體與重組蛋白 ELISA 實驗結果。Positive, 使用老鼠原倍血清; negative, 使用 5%脫脂奶粉;TSV, TSV 重組蛋白;HPV, HPV 重組蛋白;IHHNV, IHHNV 重組蛋白。 HPV 3-7 antibody 0 0.5 1 1.5 2 2.5 3 3.5 Posi tive Nega tive Nega tive sele ction TSV HPV IHH NV A b so rb anc e of O D 450 n m 圖 3-3-9 HPV 3-7 抗體與重組蛋白 ELISA 實驗結果。Positive, 使用老鼠原倍血清; negative, 使用 5%脫脂奶粉;TSV, TSV 重組蛋白;HPV, HPV 重組蛋白;IHHNV, IHHNV 重組蛋白。
IHHNV 3-62 antibody 0 0.51 1.52 2.53 3.54 4.55 Posi tive Nega tive Nega tive sele ction TSV HPV IHH NV A b so rb anc e of O D 450 n m
圖 3-3-10 IHHNV 3-62 抗體與重組蛋白 ELISA 實驗結果。Positive, 使用老鼠原倍血 清;negative, 使用 5%脫脂奶粉;TSV, TSV 重組蛋白;HPV, HPV 重組蛋白;IHHNV, IHHNV 重組蛋白。 66 45 30 M 1 2 3 (kDa) 圖 3-3-11 HPV 3-24 抗體與重組蛋白西方點墨實驗結果。Lane 1, IHHNV 重組蛋白; Lane 2, HPV 重組蛋白;Lane 3, TSV 重組蛋白。
圖 3-3-12 HPV 3-7 抗體與重組蛋白西方點墨實驗結果。Lane 1, IHHNV 重組蛋白;Lane 2, HPV 重組蛋白;Lane 3, TSV 重組蛋白。 66 45 30 M 1 2 3 (kDa)