行政院國家科學委員會專題研究計畫 成果報告
兩型斑馬魚肌肉調控因子,MRF4-I 及 MRF4-II,的分子結構、
在胚胎早期發育的角色、轉錄調控與啟動子分析
計畫類別: 個別型計畫 計畫編號: NSC92-2321-B-002-018- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學分子與細胞生物學研究所 計畫主持人: 陳曜鴻 計畫參與人員: 陳曜鴻, 蔡懷楨 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 9 月 15 日
行政院國家科學委員會補助專題研究計畫
■ 成 果 報 告 □期中進度報告兩型斑馬魚肌肉調控因子,MRF4-I 及 MRF4-II,的分子結構、在
胚胎早期發育的角色、轉錄調控與啟動子分析
計畫類別:■
個別型計畫 □
整合型計畫
計畫編號:NSC-92-2321-B-002-018
執行期間:92 年 8 月 1 日至 93 年 7 月 31 日
計畫主持人:陳曜鴻
協同主持人:蔡懷楨
成果報告類型(依經費核定清單規定繳交):■精簡報告 □完整報告 本成果報告包括以下應繳交之附件: □赴國外出差或研習心得報告一份 □赴大陸地區出差或研習心得報告一份 □出席國際學術會議心得報告及發表之論文各一份 □國際合作研究計畫國外研究報告書一份 處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列管計畫 及下列情形者外,得立即公開查詢 □涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:台灣大學分子與細胞生物研究所
中 華 民 國 九十三 年 九 月 十五 日
中文摘要 MRF4 的蛋白質結構與序列,與控制肌肉發育轉錄因子群高度相似,但其具 體的生物功能,尤其是在胚胎發育後期在分子生物層次面所扮演的角色,至今實 在了解極為有限。本報告複殖出斑馬魚的 mrf4 基因,並由染色體 mapping 的方 式得知其基因位於第四條染色體的長臂上結構與人類相類似。更進一步得知斑馬 魚有兩型 MRF4 cDNA (ZMRF4-1 與 ZMRF4-2),是由於另類剪接所導致。其中 ZMRF4-1 含有 206 個胺基酸,從 14-hpf 才開始被偵測到,而且逐漸變強,屬於 mid-somitogenesis 後才開始表現。ZMRF4-2 含有 239 個胺基酸,從 6-48-hpf 都 被偵測到,屬於 universal 型。兩型的 MRF4 都表現在體節中比較偏 hypaxia myotome 處。最後選用 morpholino knockdown 實驗,當內生性的 MRF4 被 knockdown 後,myosin heavy chain 的形成並未受阻,但是快肌與慢肌的連結產 生缺失。因此我們推論,MRF4 的功能是使肌肉纖維能產生正確的連結。 關鍵字:肌肉發育,轉錄因子,另類剪接,體節,斑馬魚
Abstract
The primary amino acids sequences of MRF4 are highly conserved with the muscle regulatory factors (MRFs), but the molecular mechanism of MRF4 involved in skeletal muscle cell specification and differentiation during late embryonic development of fish is still unclear to date. In this report, we cloned the zebrafish mrf4 gene and showed that is located at the long arm of chromosome 4 (LG4) by chromosome mapping. We also made the syntenic relationship of mrf4 gene between zebrafish and human, and showed that zebrafish LG4 fragment between EST markers fa05f06 and fk68a09 (including myf5) was related to human chromosome 12q13-12q21. We furthermore isolated two transcripts of mrf4 (ZMRF4-1 and ZMRF4-2) which were result from alternative splicing. ZMRF4-1 possesses 206 amino acids whereas ZMRF4-2 possesses 239 amino acids. The ZMRF4-1 transcripts were first detected at 14 h, and gradually increase until adult. In contrast, ZMRF4-2 transcripts were first detected from 6 h to adulthood. Both two types of mrf4 are expressed at the hypaxial region of somites. Finally, we check the MRF4-morphants phenotypes by using anti-myosin heavy chain antibody, and found that the myofibers perform incorrect connection. Based on these observations, we proposed that MRF4 is required for myofiber connection.
前言
脊椎動物的肌肉發育,是受到一群統稱為 muscle regulatory factors (MRF) 的 肌肉特異性轉錄因子群的調控。這群 MRF 主要有四個成員,包括:MyoD、 Myogenin 、Myf5 以及 MRF4,而且已經在人類(Braun et al., 1989)、小鼠(Davis et
al., 1987)、爪蟾(Hopwood et al., 1991)以及斑馬魚(Chen et al., 2000; 2001)中被複
殖出來。這群特異性的轉錄因子都具有 basic-helix-loop-helix (bHLH) 的 domain (也稱為 bHLH family)。藉著此 domain 與細胞內之其他的轉錄因子如 E12 或 E47 等形成異雙體 (heterodimer),再與基因上游啟動子區域之 E-box (CANNTG) 結 合,進而活化肌肉特異性基因的表現 (Chakraborty et al.,1992)。
然而,在小鼠的胚胎發育過程中,MRF4 的基因分兩個階段表現。其中第一 階段最早在授精後第九天出現,第十一天半消失,此時 MRF4 的生物功能可能 是執行 secondary MRFs 的功能(Summerbell et al., 2002)。第二階段 MRF4 的表現 是在第十六天再度出現然後一直持續到成體,此階段 MRF4 的生物功能一直不 清楚,直到去年才在滑爪蟾(Xenopus)的實驗中被證明可能是與肌肉的再生有關 (Ataian et al., 2003; Becker et al., 2003)。這些實驗證據指出,MRF4 可能具有相當 複雜的生物功能,必須利用不同模式動物才可將其生物功能研究透徹。在 MRF4 基因剃除鼠的實驗中,Braun 等人(1994)將 mrf4 基因的 exon 1 前端刪除,得到的
mrf4-/-同型合子小鼠一出生即死亡,具有粗而短的肋骨,背部肌肉缺失以及 myosin
heavy chain 降低;Patapoutian 等人(1995)將 MRF4 基因的 exon 1 後端刪除,得到
的 mrf4-/-同型合子小鼠也是一出生即死亡(少部分可存活),肋骨異常而且與胸骨
無法連結,肋間肌異常以及 myosin heavy chain 降低;Zhang 等人(1995)將整段
mrf4 基因進行取代,得到的 mrf4-/-同型合子小鼠全部都可存活至成體,雖然肋骨
異常但與胸骨正常連結,肌肉部分除了 myosin heavy chain 略為降低外大都正 常。這些結果顯示,在 MRF4 基因剃除鼠的實驗中,基因剔除區域很重要。由 於小鼠的 MRF4 基因下游約 8.5 kb 處,有 myf5 基因,上游約 5kb 處又有 Hox 基 因(Summerbell et al., 2000)。不同大小片段的基因剔除區,可能會影響 MRF4 基 因鄰近的基因表現,造成不同的表現型,因此導致 MRF4 的生物功能至今尚眾 說紛紜,並無具體結論。Ekker 等人(2001)利用化學修飾方法合成了基因特異性 的胚胎發育抑制劑(morpholino),會將特異性的目標基因 knockdown。因此利用 MRF4-morpholino 來將內生性的 MRF4 基因表現 knockdown,進而觀察胚胎的表 現型。如此一來則不會破壞染色體的結構進而影響鄰近基因的表現,二來也可以 比較有效率、符合經濟效應、符合科學原則的研究 MRF4 具體的生物功能。 相對於高等脊椎動物的研究,有關於魚類的 MRF4 的分子結構與生物功能, 更是一無所知。在本篇報告中,我們複殖出兩型的斑馬魚的 MRF4 transcripts, 為了能更深入探討 ZMRF4-1 與 ZMRF4-2 的生物功能,我們應用 morpholino 與 抗體染色等技術,在 in vivo 的狀況下去瞭解 MRF4 這種重要的肌肉核蛋白在胚 胎早期的調控以及蛋白質的表現。相信此研究結果,將可在學術上能進一步明白 兩型 MRF4 在魚類胚胎中所擔任的角色、因果及互動。
材料與方法 斑馬魚 AB strain 的飼育 在 28.5℃、光週期 14 小時的培養箱中,以 60x 20x 30 立方公分的玻璃水族 箱飼養,每缸平均 30 隻魚。每天傍晚餵食豐年蝦一次,日間不定時餵食人工乾 燥香魚飼料 1-2 次。 斑馬魚胚胎時期核糖核酸(RNA)之抽取 實驗用之斑馬魚,取從 3-96 hpf (hours post-fertilization)時期的胚胎,約 100 個胚胎裝成一管,收集後置於液態氮中儲存。抽取 RNA 時,每 100 個胚胎加入 約 0.5ml 之 TRI reagent (PROtech),混合並將胚胎均勻化;再加入 1/5 體積之
CHCl3 ,經離心去掉蛋白等雜質,最後以酒精沉澱法純化 RNA,並將最終濃度
調成 10μg/μl。
RT-PCR (Reverse Transcription-Polymerase Chain Reaction)
取約 10μg 的 RNA,加入 Oligo-dT 後先在 70℃下反應 10 分鐘,再依序加 入 DTT、dNTP、SuperScript (GibcoBRL)等藥品,置於 42℃下反應一小時,最後 加入 RNaseH 將 RNA 去除,這樣可得到 first-strand cDNA。由上述所得到的 first-strand cDNA 可用來當 PCR 的模板,加入 mrf4 之特異性引子後,於 94℃ 1 min、58℃ 1 min、72℃ 1 min 的條件下進行 30 個 cycles 後即可完成。
北方墨點法(Northern blotting)
取 21 與 24 hpf 之 RNA,用帶有 mrf4 基因的部份序列當探針,利用 Sambrook et al., (1989)方法確定 mrf4 cDNA 的大小。取 20-30μg 的 RNA,加入 20μl 的 RNA sample buffer 及 3μl 的 loading dye,於 65℃水浴 10 分鐘後,以含 17.6﹪formaldehyde 的 1﹪瓊脂膠體於 4℃,100volts 下電泳 3 小時,將膠體和 Immobilon-N transfer membrane 一起用 20×SSC 浸潤 30 分鐘。然後進行轉附作 用。之後將 membrane 風乾,用 UV 照射將 RNA 連結到 membrane 上,加入 prehybridization solution,65℃下反應 2-4 小時,然後再以 hybridization solution 於 65℃進行雜交反應,16-24 小時後,membrane 以含 2×SSC 及 0.5﹪SDS 之 solution,在室溫下清洗兩次,每次各 15 分鐘,再以 0.1﹪SSC 及 0.1﹪SDS,於 75℃清洗兩次,每次各三十分鐘。將 membrane 風乾後做自動放射顯影。
Riboprobe 的製備
本實驗採用 Roche Molecular Biochemicals 的 Riboprobe 合成套件構築,取 內含 mrf4 部分序列進行合成,其操作之詳細步驟參照廠商所建議之條件進行。 最後合成的 riboprobe 約有 250 ng,可用以進行北方墨點法及 whole mount in situ hybridization。
Whole mount in situ hybridization
將收集固定後的斑馬魚胚胎取出,先將其外膜去除後,再依序進行脫水、 覆水、prehybridization、hybridization、抗體呈色的反應。
100﹪的 methanol 內。 (2) 覆水:逐漸將胚胎中 methanol 的濃度降低,最後浸泡於 PBS 內。 (3) Prehybridization、hybridization:胚胎經由數次的清洗後,最後泡在 HYB 中 在 70℃下 prehybridization 約 4 小時,之後加入 mrf4 的 riboprobe 進行 hybridization。 (4) 抗體呈色:hybridization 完成後,需要不斷的清洗將未黏合的 RNA 探針移除, 最後加入 anti-DIG Ab,於 4℃下隔夜培養後再加入 NBT 與 BCIP 進行呈色。 定量 RT-PCR
(1) Primer 與螢光 probe 的設計:將複殖出的 mrf4 的 cDNA 序列輸入 ABI PRISM 7700 中的 Primer express software 中,設計出 Tm值約 60℃的 primer 及 Tm值 約 65℃的螢光 probe,所夾出之 replicon 約 100-200 bp。
(2) Primer 的測試:先用 RT-PCR 的方式測試所設計的 primer 是否只夾出 single band,以避免 non-specific 產物的干擾。若測試的結果可以,即可進行螢光 probe 的合成。
(3) 定量 RT-PCR:採用 Perkin Elimer 的套件進行之,最後採用 ABI PRISM 7700 上機分析。
Morpholino 之顯微注射法
先將 morpholino 依照廠商建議的方式,依序稀釋到適合的濃度,再進行顯 微注射實驗,步驟同 Chen and Tsai (2002)。
冷凍切片技術 室溫下以 4% PFA 固定胚胎 1 小時後,以 30%蔗糖滲透(過小不適合直接操 作之胚胎可先以瓊脂糖膠包埋),待胚胎由液面沉降至管底即可。以冷凍黏著劑 (O.C.T, SAKURA)包裹待切片之胚胎,置於已預冷(-15℃)之冷凍切片箱中,至其 完全凝固方可架於基座並栓緊,開始進行切片。於載玻片上註明樣本名稱及日 期,將切下之組織薄片直接沾黏至載玻片,冷凍黏著劑遇熱則瞬間將樣品薄片緊 緊黏於玻片上。本實驗切片厚度為 10 μm,切片樣品為受精後 48 hpf、受精後 7 天及成體等不同時期之斑馬魚。
結果與討論
兩型斑馬魚 mrf4 基因的複殖
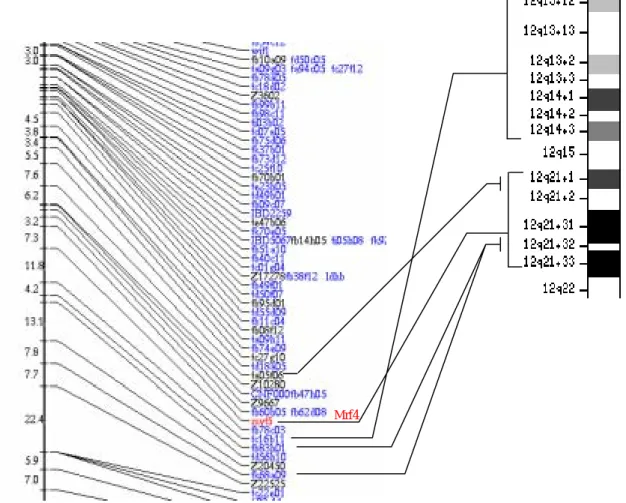
肌肉調控因子 mrf4 已在人類(Braun et al., 1990)、小鼠(Pin and Konieczny, 2002)、鳥類(Fujisawa-Sehara et al., 1992)與滑爪蟾(Jennings, 1992)中被複殖出,但是 魚類的 mrf4 基因至今尚一無所知。我們利用 RT-PCR 加上 5’-與 3’-RACE 以及北 方墨點法的技術分析,得知斑馬魚擁有兩型的 MRF4(ZMRF4-1 與 ZMRF4-2)。 其中,ZMRF4-1 含有 1122 bp,可轉譯出 206 個胺基酸;而 ZMRF4-2 含有 1071 bp, 可轉譯出 239 個胺基酸(Figs. 1 and 3B)。經由序列比對,發現 ZMRF4-2 與陸生脊 椎動物的保守性比 ZMRF4-1 高,因此我們推測,ZMRF4-1 可能是斑馬魚特有的 MRF4。 斑馬魚 mrf4 基因在染色體的位置上與人類具有相同的保守性 為了更進一步了解 mrf4 基因在演化上的保守性,我們使用了 radiation hybrid 的方法來訂出 mrf4 基因在染色體的位置。結果顯示,mrf4 基因位在第四條染色 體(LG4)的長臂上,介於 fb62d08 與 fb78c03 這兩個 EST 標記之中,而且距離 EST 標記 z9667 有 5.87 centiRay 之遙。由演化的保守性的觀點來看,斑馬魚 LG4 上 fa05f06 與 fk68a09 這兩個 EST 標記之中(包括 mrf4 基因)與人類第十二條染色體 12q13-12q21 的位置保守性相當高(Fig. 2)。
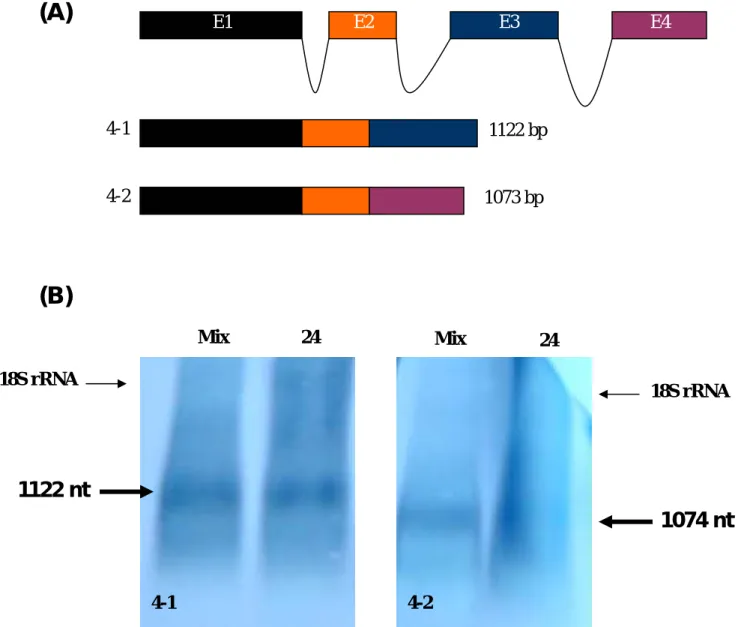
兩型斑馬魚 mrf4 是由於另類剪接所導致
將 mrf4 基因的序列輸入至 Ensembl (http://www.ensembl.org/)網站上進行序列 比對,結果比對到一個長約 1800 kb 斑馬魚的 contig 編號 ctg9418。接著將 ZMRF4-1 與 ZMRF4-2 的 cDNA 序列分別與 ctg9418 進行比對,結果發現斑馬魚 的 mrf4 基因是由 4 個 exons 與 3 個 introns 所組成,其中 ZMRF4-1 是由 exon1、 exon2 與 exon3 剪接而成;而 ZMRF4-2 是由 exon1、exon2 與 exon4 剪接而成(Fig. 3A)。至此,我們得知兩型斑馬魚 mrf4 是由於另類剪接所導致。與所有已發表之 物種的 MRF4 序列相比對,我們是第一個得知斑馬魚具有兩型另類剪接的 ZMRF4-1 與 ZMRF4-2,其他物種的 MRF4 並未有類似的報告。因此,研究斑馬 魚位什麼會具有兩型另類剪接的 MRF4,以及彼此間的生物功能,已成為刻不容 緩的話題。 兩型斑馬魚 mrf4 的時空分佈 我們首先選用定量定量 RT-PCR 的方法先來研究兩型另類剪接的 ZMRF4-1 與 ZMRF4-2 在胚胎早期表現的先後順序。定量 RT-PCR 的主要原理是及時偵測 PCR 產物的螢光強度,當螢光強度在 linear phase 時,每增加一個 cycle 數,產物 會增加兩倍。此時只要取 linear phase 的半數值時的 cycle 數(CT),則可利用 CT 值的大小來反應原始 temple 量的多寡。由實驗結果得知,ZMRF4-1 的 transcripts 從 16-hpf 才開始被偵測到,而且逐漸變強,屬於 mid-somitogenesis 後才開始表 現。反觀 ZMRF4-2 的 transcripts 從 6-48-hpf 都被偵測到,屬於 universal 型(Fig. 4)。
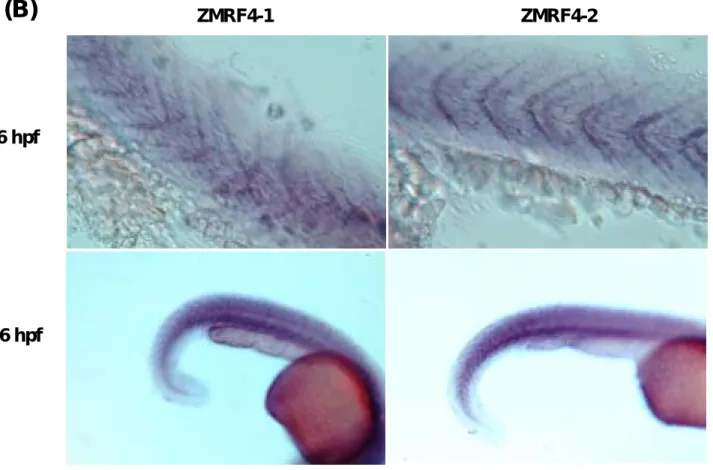
接著我們選用 whole mount in situ hybridization 的方法來研究 ZMRF4-1 與 ZMRF4-2 在胚胎早期的哪些位置表現?結果顯示 MRF4 在 14-、30-、54-及 74-hpf
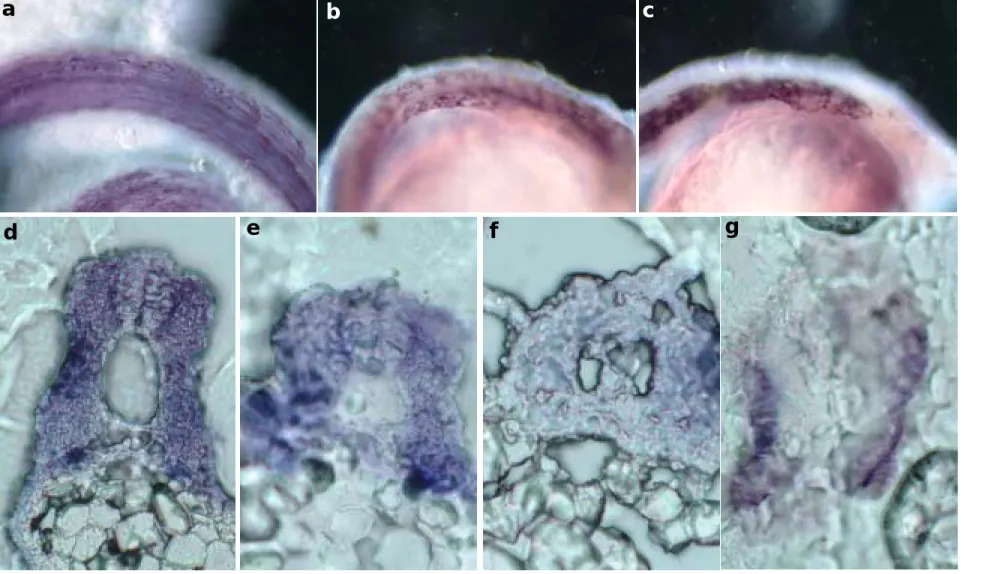
的 embryos 中表現,表現的位置雖然都在 somite 中,但是卻集中在 hypaxial 的位 置(Fig 5A),說明了 MRF4 與 hypaxial muscle 的形成有極大相關。Hypaxial muscle 的形成與 MyoD 有關,MyoD 是否透過調控 MRF4 的 signal 來調控 hypaxial muscle,需要更進一步的實驗證據證明。至於兩型 MRF4 分別的表現位置,由解 果可得知在 26-與 36-hpf 時,表現的位置也都集中在 hypaxial 的位置(Fig 5B)。 因此應該再多收集多一點發育時期的胚胎,才能更清楚的得知其所扮演的角色。 斑馬魚內生性 mrf4 被 knockdown 的表現型 由上一個實驗得知 MRF4 的表現位置,我們有興趣的是,MRF4 是與快肌或 慢肌的形成有關。於是我們選用可以同時標訂快慢基的單株抗體 F59 來進行免疫 染色。同時也選用野生種斑馬魚與 MRF4-MO 處理過的斑馬魚來進行實驗。結果 顯示,不論 slow or fast muscle 都偵測的到 signals,這表示 MRF4 是跟 slow and fast muscle 的形成都有關係。經過 MRF4-MO knockdown 的斑馬魚胚胎,發現其 slow and fast muscle 的形成都跟著消失,故推論 MRF4 是跟 slow and fast muscle 的形 成都有關係(Fig. 6)。
參考文獻
Ataian Y, Qwens J and Hinterberger T. 2003. MRF4 gene expression in Xenopus embryos and aneural myofibers. Dev. Dyn. 226: 551-554.
Becker C, Gaspera BD, Guyot M, Donsez E, Armand AS, Charbonnier F, Launay T and Chanoine C. 2003. Expression of MRF4 protein in adult and in regenerating muscles in Xenopus. Dev. Dyn. 227: 445-449.
Braun T, Buschhausen-Denker G, Bober E, Tannich E and Arnold HH. 1989. A novel human muscle factor related to but distinct from MyoD1 induces myogenic conversion in 10T1/2 fibroblasts. EMBO J. 8: 701-709.
Braun T, Bober E, Winter B, Rosenthal N and Arnold HH. 1990. Myf-6, a new member of the human gene family of myogenic determination factors: evidence for a gene cluster on chromosome 12. EMBO J. 9: 821-831.
Braun T, Bober E, Rudnicki MA, Jaenisch R and Arnold HH. 1994. MyoD expression marks the onset of skeletal myogenesis in Myf-5 mutant mice. Development 120: 3083-3092. Chakraborty T, Martin JF and Olson EN. 1992. Analysis of the oligomerization of myogenin and E2A products in vivo using a two-hybrid assay system. J. Biol. Chem. 267: 17498-17501.
Chen YH, Lee WC, Cheng CH and Tsai HJ. 2000. Muscle regulatory factor gene: zebrafish (Danio rerio) myogenin cDNA. Comp. Biochem. Physiol. 127B: 97-103.
Chen YH, Lee WC, Liu CF and Tsai HJ. 2001. Molecular structure, dynamic expression and promoter analysis of zebrafish (Danio rerio) myf-5 gene. Genesis 29: 22-35.
Chen YH and Tsai HJ. 2002. Treatment with myf5-morpholino results in somite patterning and brain formation defects in zebrafish. Differentiation 70: 447-456.
Davis RL, Weintraub H and Lassar AB. 1987. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell 51: 987-1000.
Ekker SC and Larson JD. 2001. Morphant technology in model developmental system. Genesis 30: 89-93.
Fujisawa-Sehara A, Nabeshima Y, Komiya T, Uetsuki T, Asakura A and Nabeshima Y. 1992. Differential trans-activation of muscle-specific regulatory elements including the mysosin light chain box by chicken MyoD, myogenin, and MRF4. J. Biol. Chem. 267: 10031-10038.
can activate muscle genes ectopically in early embryos. Development 111: 551-560. Jennings CG. 1992. Expression of the myogenic gene MRF4 during Xenopus development.
Dev. Biol. 151: 319-332.
Patapoutian A, Yoon JK, Miner JH, Wang S, Stark K and Wold B. 1995. Disruption of the mouse MRF4 gene identifies multiple waves of myogenesis in the myotome Development 121: 3347-3358.
Pin CL and Konieczny SF. 2002. A fast fiber enhancer exists in the muscle regulatory factor 4 gene promoter. Biochem. Biophys. Res. Commun. 299: 7-13.
Summerbell D, Ashby PR, Coutelle O, Cox D, Yee S and Rigby PW. 2000. The expression of Myf5 in the developing mouse embryo is controlled by discrete and dispersed enhancers specific for particular populations of skeletal muscle precursors. Development 127:3745-57.
Summerbell D, Halai C and Rigby PWJ. 2002. Expression of the myogenic regulatory factor Mrf4 precedes or is contemporaneous with that of Myf5 in the somitic bud. Mech. Dev. 117: 331-335.
Zhang W, Behringer RR and Olson EN. 1995. Inactivation of the myogenic bHLH gene MRF4 results in up-regulation of myogenin and rib anomalies. Genes Dev. 9: 1388-1399.
圖表 ZMRF4-1 MMDLFETNAYFFNDLRYLEGDHGTLDMPGVSPLYEGNGSPRSPGQDPVPSETGCESSGEEHVLAPPGLQ AHCEGQCLMWACKICKRKSAPTDRRKAATLRERRRLKKINEAFDALKKKTVPNPNQRLPKVEILRSAIN YIEKLQDLLHSLDEQEQSNDTDPYTYNLKENHVTPSEYHWKKTCQSWQENPDHSSSQMAGHREAPLLI ZMRF4-2 MMDLFETNAYFFNDLRYLEGDHGTLDMPGVSPLYEGNDSPLSPGQDPVPSETGCESSGEEHVLAPPGLQ AHCEGQCLMWACKICKRKSAPTDRRKAATLRERRRLKKINEAFDALKKKTVPNPNQRLPKVEILRSAIN YIEKLQDLLHSLDEQEQSNDTDPYTYNLKENHVTPSEYHWKKTCQSWQENPDHSSSQMAGHREGAVLES SESSSLRRLSSIVDSISTEEPKARCPSQISEK Fig. 1. 兩型斑馬魚 MRF4 基因的胺基酸次序
Zebrafish LG4 Human Chromosome 12
Fig. 2. 斑馬魚 mrf4 基因在染色體的位置上與人類染色體保守性之分析 Mrf4
4-1 4-2 1122 bp 1073 bp E1 E2 E3 E4 4-1 4-2 Mix 24 Mix 24 18S rRNA 18S rRNA
(A)
(B)
1122 nt
1074 nt
Fig. 3. 斑馬魚 mrf4 基因的 exon 與 intron 組成(A)以及所另類剪接之 ZMRF4-1 與 ZMRF4-2 的
0
10
20
30
40
50
60
70
No Template
C
on
tro
l
0.15
-
6-
12-
16
-
24-
36
-
48-hpf
Relative Expressio
n
數列1
數列2
Fig. 4. 斑馬魚另類剪接之 ZMRF4-1 與 ZMRF4-2 的定量 RT-PCR 分析14 h
30 h
54 h
74 h
ZMRF4-1 ZMRF4-2
26 hpf
36 hpf
Fig. 5. 斑馬魚另類剪接之 ZMRF4-1 與 ZMRF4-2 的 whole mount in situ hybridization 分析
Fig. 6. 野生種斑馬魚(a, b, d, e)與 MRF4 knockdown 斑馬魚(c, f, g)之快慢肌分佈情形分析