行政院國家科學委員會專題研究計畫成果報告
蜀黍單籽粒後裔法及族群遺傳歧異度之研究 (二年計畫)
第一年進度報告
Studies on Single SeedDescent Method and Genetic Diversity of Population in Sorghum
計畫編號: NSC 87 - 2313 - B - 002 - 021
執行期限: 86 年 8 月 1 日至 87 年 7 月 31 日
計畫主持人: 陳 成 執行單位: 國立臺灣大學農藝系
摘要
本試驗利用 SSD 法於田間疏植增進族群至 F5 世代,並於溫室繁衍 F4 世代而得不同 SSD 法密植淘汰之 F5 世代。於田間評估各世代族群之農藝性狀,發現 F2, F3, F4, F5 各世 代族群植株表現頗為一致,而比較 F5 世代各族群則在開花日數、綠葉數、莖徑及百粒重等 農藝性狀上有顯著差異存在,可知 SSD 法密植將會造成族群植株有差異的現象,而 10×10 cm2 密植將可能為蜀黍 SSD 法最可行的條件。利用分子標誌則可偵測出參試族群間的差異 存在。各農藝性狀與 RAPD 分子標誌間之相關性,在各世代族群間並不一致,此可能是各 世代植株的族群結構不同,或栽培環境不同,或取樣誤差及樣品數不夠大所致。由各世代族 群農藝性狀之標準化變數的一般化樣品變方 (R) 及 RAPD 分子標誌變異之 Shannon's 指數 (H0) 可知,若在較正常的栽培環境下,RAPD 技術將可應用於偵測族群遺傳歧異度。緣由與目的
蜀黍俗稱高粱為常異交作物,其育種的方式以往大多採用自交作物的育種方法 (Doggett, 1982),直到基因型雄不稔性 ms2 (Stephens, 1937) 和細胞質雄不稔性 (Stephens and Holland, 1954) 相繼被發現之後,才有雜種優勢 (heterosis) 的利用及具有商業性雜交蜀黍品種的生 產。1960 年 Webster 在 Nebraska 首先利用細胞質雄不稔性進行蜀黍族群的逢機交配創建基 礎族群 (Nordquist et al, 1973),隨後的育種家依據不同的育種需要相繼建立許多基礎族群, 以供育種選拔之用,在這些蜀黍族群中,大部分採用基因雄不稔性 (ms3 及 ms7) 做為雄不 稔材料的來源 (謝, 1985; Atkins, 1982; Bhola Nath, 1982; Nordquist et al., 1973; Ross et al., 1977)。目前主要生產區域無不利用其雜種優勢來進行雜交組合,然優良雜交組合需藉助選拔 完美的自交系提供交配評估與選育工作,所以若要選拔出完美自交系則需要視育種族群遺傳 變異的大小而定。由分離族群選育自交系的育種法,一般以譜系育種法 (pedigree breeding method)、混合 族群育種法 (bulk population breeding method)、單籽粒後裔法 (single seed descend method, SSD) 等為較常用的方法。在育種程序中,SSD 法因每株皆取一粒種子進行世代增進,在高 世代之族群變異仍維持與初期世代族群相當,所以比譜系育種法及混合族群育種法有更大的 遺傳變異 (Khalifa and qualset, 1975; Haddad and Muehlbauer, 1981;Tee and Qualset, 1975);另外 SSD 法由於可在溫室或生長箱中密植栽培,以增加每年之繁殖世代數,加速基因型的同型結
合育成自交系,縮短育種年限大幅提升育種效率 (Goulden, 1939),並節省大量的花費及栽培 面積 (Grafius, 1965),因此已漸被應用於小粒穀類 (small grains) 作物如小麥、水稻 … 等及 矮生莖桿作物如大豆、花生、番茄 … 等自交作物之上 (Brim, 1966; Casali and Tigchelear, 1975; Jink and Pooni, 1984; Tee and Qualset, 1975)。然而對於高莖的常異交蜀黍而言,此法之應用則 尚未有研究發表,因此利用 SSD 法快速地育成蜀黍自交系,以提供種原收集及往後的利用, 是一個非常值得研究與應用的方法。 雖然 SSD 法具有上述的優點,但由於每代每株僅取一粒種子繁殖,除非每代植株之發 芽率、成活率皆近 100%,否則幾代之後,此種指數關係的遺失將使族群數目縮小,可能導 致高世代之遺傳變異偏狹,但此論點必須在每代之遺失皆為非逢機性之前提下方能成立。另 外在 SSD 法欲節省栽培面積、加速世代之條件下,而以高密度種植,在日光、養分供應不 正常且植株間又具高度競爭力下所產生的缺株淘汰,其繁衍至高世代後,則可能造成整個族 群遺傳變異的改變。Roy (1976) 以四個小麥品系模擬 SSD 法於密植下的競爭作用,調查當 代各品系在混植、純植下的表現,認為 SSD 法應避免極度競爭以致死亡,因為某些競爭力 差者可能有高產之表現。Martin (1978) 以三個大豆組合於兩種密度下經 SSD 法繁殖四代, 結果遺失 19% 者其花色、軟毛色的基因頻度均未改變,而蓄意密植造成 55% 遺失者,其 基因頻度已不符卡方測驗。Cisar (1982) 嚐試利用 SSD 法密植以使燕麥產生單粒種子,經兩 代密植後,遺失 34% 的族群其株高、成熟期、含油量、產量與幾無淘汰的族群無顯著差異。 由以上的研究結果,SSD 法植株遺失淘汰的逢機與否,與作物之種類、淘汰率高低及調查的 性狀有關。因此本試驗將以 SSD 法繁衍蜀黍族群,並在不同的栽培密度下,探討蜀黍利用 SSD 法時所造成族群植株間的競爭淘汰是否會影響族群的遺傳變異?其改變是否逢機?若 非逢機則其偏向為何?這是在蜀黍 SSD 法實際應用前必須先加以研究的,也是本計畫所欲 研究的主題之一。 對作物育種而言,若育種材料之遺傳變異過於狹隘,不但使作物對環境的穩定性降低, 更限制產量及品質的提昇。為了更有效地利用作物種源,提高雜種優勢率,評估育種族群的 遺傳歧異度是必要的工作。對於遺傳歧異度之測量及分析方法,最早僅由外表型態分類性狀 加以分類;隨後即有育種學家利用田間外表連續性數量性狀進行族群遺傳變異的統計分析 (Fountain and Hallauer, 1996; Ouendeba et al., 1995; Stucker and Haullauer, 1992; Bittinger et al.,
1981),但此法需耗費大量土地、時間及人力,且植株評估易受環境左右而影響其精確性。隨 著生化技術的發展,利用電泳進行同功異構酵素 (isozyme) 的分析,可在實驗室快速地偵測 遺傳材料間的歧異度,增進育種效率 (Kephart, 1990; Kiang et al., 1992; Mymudes and Les, 1993),然而此法可用的標幟 (marker) 少,且結果易隨植株發育時期而異為其缺點。近來利 用 RFLP (restriction fragment length polymorphism) 技術進行植株 DNA 分子序列變異的分 析,可準確且迅速地得知育種材料之遺傳歧異度 (Bark and Havey, 1995; Fjellstrom and Parfitt,
1994; Keim et al., 1994),但由於其分析流程太長且需耗費大量之時間及人力,因此將 RFLP
納入育種計劃中有其限制存在。最近更有學者利用 RAPD (random amplified polymorphic DNA) 的技術來偵測族群遺傳歧異度,改進 RFLP 耗時、費工的缺點,此新的技術已漸被廣 泛地應用 (Chalmers et al., 1992; Dawson et al., 1995; Huff et al., 1993; Shah et al., 1994)。
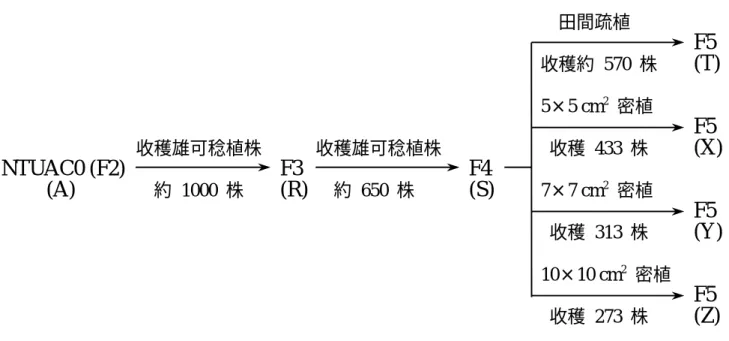
本研究室 (國立臺灣大學農藝系作物育種研究室) 以 9 個優良細胞質雄不稔品系為母 本及 11 個經過評估的優良 F1 雜種為花粉親,已成功地建立一套具廣泛遺傳變異、適合本 省栽植的基礎族群 (NTUAC0) (江等,1995)。利用 SSD 法於田間疏植增進族群至 F5 世代, 並於溫室繁衍 F4 世代而得不同 SSD 法密植淘汰之 F5 世代。為探討 RAPD 技術應用於偵 測蜀黍族群遺傳歧異度之可行性,本研究擬用上述各世代族群為材料,在實驗室進行族群
RAPD 的分析,求得各世代族群間遺傳歧異度之差異,並在田間進行族群變異的評估,以得 知 SSD 法密植淘汰對族群遺傳變異之影響,並探討 RAPD 技術應用於育種計畫之實用價 值。
材料與方法
1. 材料 NTUAC0 基 礎 族 群 利 用 SSD 法 於 田 間 疏 植 增 進 族 群 至 F5 世 代 , 即 由 NTUAC0 族群 (F2) 收穫雄可稔植株之自交種子,每株取一粒種子混合而得 F3 世 代,再種植 F3 世代並依上法得 F4 世代。F4 世代於田間疏植得 F5 世代,並同時 於溫室密植 F4 世代而得不同 SSD 法密植淘汰之 F5 世代。各世代族群之代碼及繁 衍流程圖如圖 1。 2. 方法 (1) 於田間評估各世代族群之農藝性狀,進行統計分析以得知各族群之差異。並以多變 數分析法求得農藝性狀之標準化變數的一般化樣品變方 R (Johnson and Wichern, 1992),以評估各世代族群之遺傳變異度。(2) 取各世代族群之植株進行 RAPD 之分析,利用有效逢機引子 (primer) 測試下在各 樣品間產生多型性之結果,存在多形性條帶者記錄為 1,不存在者記錄為 0,即所 有資料轉換成 1 與 0 的型式後輸入電腦,以 Shannon‘s 指數 (Shannon‘s index) 估算各世代族群之遺傳歧異度 (Saghai-Maroof et al., 1984),其算法如下所示。並與 田間之資料作比較,以探討 RAPD 技術應用於偵測族群遺傳歧異度之可行性。 H0 = -Σ piln pi pi:族群第 i 個 RAPD 多型片段之頻度 H0 值愈大表示族群的歧異度愈大
結果與討論
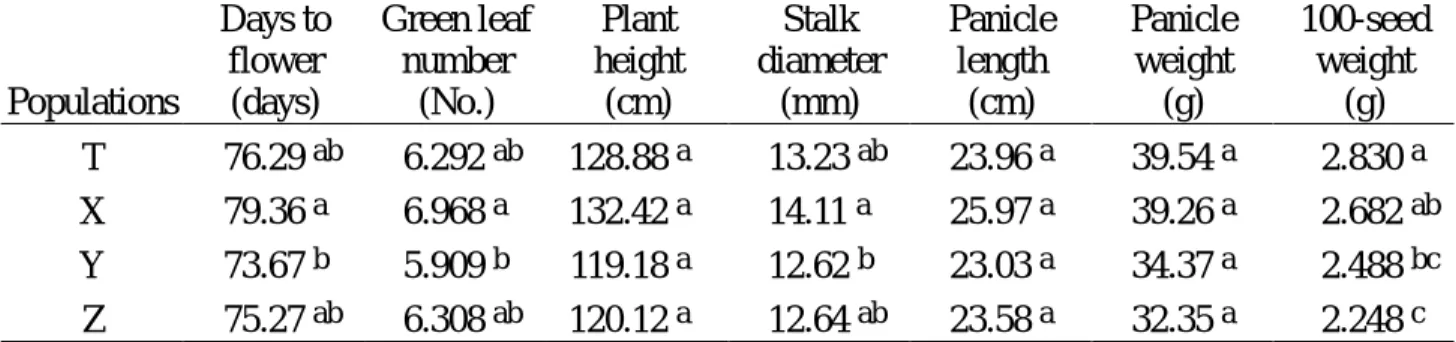
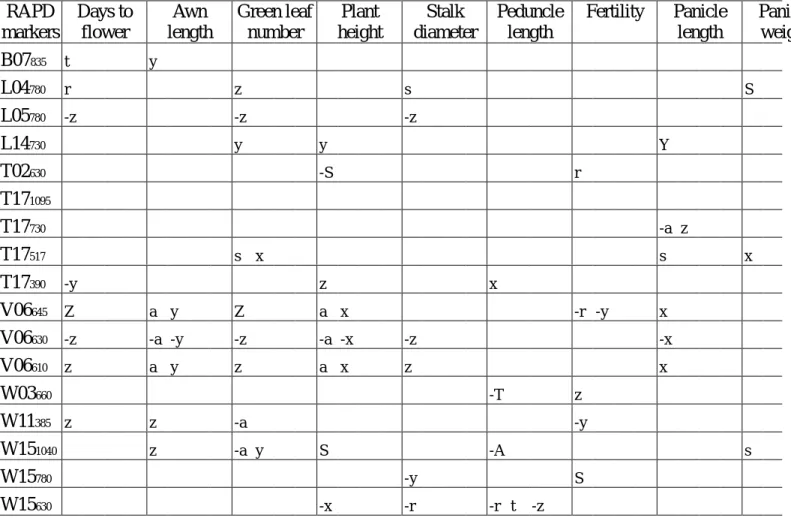
1997 年春作於臺北種植各世代族群之農藝性狀平均值差異顯著性測驗列於表 1。比較 F2, F3, F4, F5 各世代族群之平均值可知在全部所調查的農藝性狀皆無顯著差異存在,即在 此春作環境下,各世代族群植株表現頗為一致。而比較 F5 世代各族群之農藝性狀平均值 (表 2),可知各世代族群植株,在開花日數、綠葉數、莖徑及百粒重等農藝性狀上有顯著 差異存在,即在此春作環境下,以 5×5 cm2 密植族群植株所需開花日數較長、綠葉數較 多、莖徑較粗,而 7×7 cm2 密植族群植株則相反;但在百粒重方面,則以田間疏植 F5 世 代族群最重,10×10 cm2 密植族群植株最輕。可知 SSD 法密植將會造成族群植株有差異 的現象,尤其在 7×7 cm2 密植條件下更易造成與田間疏植間之差異表現,因此在此密植 條件下,所淘汰的植株可能為非逢機性,且為淘汰開花日數較長、綠葉數較多、莖徑較粗 及百粒重較重之植株。而仔細比較平均值可知,5×5 cm2 密植族群與 F5 世代疏植植株有 輕微差異,其植株淘汰方向恰與 7×7 cm2 密植族群相反;而 10×10 cm2 密植族群植株 之表現與田間疏植 F5 世代族群最類似,可見 10×10 cm2 密植將可能為蜀黍 SSD 法最可 行的條件。 1997 年春作於臺北種植各世代族群之 RAPD 分子標誌頻度差異顯著性測驗詳如表 3,其中在 T02630、T17517、T17390、V06645、V06630、V06610 及 W151040 等分子標誌上有差異性存在,可知雖然田間農藝性狀無法顯現出族群間的差異 (表 1) ,但利用分子標誌則 可偵測出其間的差異存在。而 1997 年春作於臺北種植 F5 世代各族群之 RAPD 分子標 誌頻度差異顯著性測驗詳如表 4,其中僅在 T17517、T17390 及 W151040 等分子標誌上有差 異性存在。 1997 年春作於臺北種植各世代族群各農藝性狀與 RAPD 分子標誌間相關性之顯著 性測驗詳如表5。至少有兩個族群顯現出農藝性狀與 RAPD 分子標誌間的相關性或有一個 族群達極顯著相關者有開花日數與 V06645,芒長與 V06645、V06630、V06610,綠葉數與 T17517、V06645、W151040,株高與 T02630、V06645、V06630、V06610、W151040,穗梗長與 W03660、 W151040、W15630,稔實性與 V06645、W15780,穗長與 L14730、T17730,穗重與 L04780,種子 色與 T171095、V06645,百粒重與 T17390、W11385。由上可知,相關性的表現在各世代族群間 並不一致,此可能是各世代植株的族群結構不同,即各族群之平均值、變異、歪度、峰度 等各不相同所致;不同栽培環境對植株之表現亦有相當大之影響,所以亦可能造成族群間 相關性不一致的現象;而取樣誤差及樣品數不夠大亦可能為不一致之因。 1997 年春作於臺北種植各世代族群農藝性狀之標準化變數的一般化樣品變方 (R) 及 RAPD 分子標誌變異之 Shannon's 指數 (H0) 詳如表 6。由於 1997 年受聖嬰現象的影響 在植株栽培期有綿連細雨,因此在此栽培環境下植株受多雨及日照不足影響而表現異常, 吾人利用目測人為將田間植株區分為正常與異常兩類,進行上述統計量之計算。利用田間 農藝性狀資料計算之變異 (R),在全部植株納入計算時以 5×5 cm2 密植族群之變異最大達 0.0766,而以 10×10 cm2 密植族群之變異最小;而僅正常植株納入計算則以 F3 世代族群 之變異最大達 0.1050,而以 F2 世代族群之變異最小。利用 RAPD 分子標誌變異之 Shannon's 指數,在全部植株納入計算時以 F3 世代族群之變異最大達 0.2671,而以 F2 世 代族群之變異最小;而僅正常植株納入計算則以 F5 世代族群之變異最大達 0.2687,而以 10×10 cm2 密植族群之變異最小。一般認為利用分子標誌所計算出之統計量應不受環境所 影響,但由上可知此二統計量在全部植株與正常植株的計算結果皆無一致的現象,此可能 是取樣誤差或樣本數不足所致。將兩統計量進行比較可發現,利用正常植株之田間資料與 RAPD 分子標誌所估算之族群遺傳歧異度大小之趨勢較全部植株一致,因此若在較正常的 栽培環境下,RAPD 技術將可應用於偵測族群遺傳歧異度。
計畫成果自評
1. 得知 10×10 cm2 為蜀黍 SSD 法密植之最適條件,故 SSD 法應可以利用於蜀黍育種 上。 2. 得知 RAPD 技術將可應用於偵測族群遺傳歧異度。參考文獻:
江明樹、林志東、王裕權、陳成。1995。早世代蜀黍逢機交配族群農藝性狀之探討。中 華農藝 5:261-276。
謝兆樞。1985。世界蜀黍之研究現況。科學農業 33(5-6):155-170。
Atkins, R. E. 1982. Registration of IAP3BR(M)C3 sorghum germplasm. Crop. Sci. 22:165-166. Bark, O. H., and M. J. Havey. 1995. Similarities and relationships among populations of the bulb
onion as estimated by nuclear RFLPs. Theor. Appl. Genet. 90:407-414.
Bhola Nath. 1982. Population breeding techniques in sorghum. In House, L. R., L. K.
Mughogho, and J. M. Peacock (eds) "Sorghum in the Eighties", pp. 421-434. ICRISAT , Patancheru PO, A. P., India.
Bittinger, T. S., R. P. Cantrell, J. D. Axtell, and W. E. Nyquist. 1981. Analysis of quantitative traits in pp9 random-mating sorghum population. Crop sci. 21:664-669.
Brim, C. A. 1966. A modified pedigree method of selection in soybeans. Crop Sci. 6:220.
Casali, W. D. V., and E. C. Tigchelear. 1975. Breeding progress in tomato with pedigree selection and single seed descent. J. Amer. Soc. Hort. Sci. 100(4):362-364.
Cisar, g., A. E. Howay, and C. M. Brown. 1982. Optimal population density and random vs. selective elimination of genotype under a modified single seed descent method with spring oats. Crop Sci. 22:576-579.
Chalmers, K. J., R. Waugh, J. I. Sprent, A. J. Simons, and W. Powell. 1992. Detection of genetic variation between and within populations of Gliricidia sepium and G. maculata using RAPD
markers.Heredity. 69:465-472.
Dawson, I. K., A. J. Simons, R. Waugh, and W. Powell. 1995. Diversity and genetic differentiation among subpopulations of Gliricidia sepium revealed by PCR-based assays. Heredity. 74:10-18.
Doggett, H. 1988. Sorghum. 2nd ed.Longman,Harlow. PP.70-79.
Doggett, H. A. 1982. A look back at the 70s. In : House, L. R., L. K. Mughogho, and J. M. Peacock (eds) “ Sorghum in the Eighties ”, ICRISAT, Patancheru Po, A. P., India. PP.15-24.
Fjellstrom, R. G., and D. E. Parfitt. 1994. Walnut(Juglans spp.) genetic diversity determined by
restriction fragment length polymorphisms.Genome. 37:690-700.
Fountain, M. O., and A. R. Hallauer. 1996. Genetic Variation within Maize Breeding Populations. Crop. Sci. 36:26-32.
Goulden, C. H. 1939. Problems in plant selection. Proceeding s of the Seventh International Genetical Congress, Edinburgh, Scotland. Cambridge University Press, Cambridge, PP.132-133. Grafius, J. E. 1965. Short curts in plant breeding. Crop Sci. 5:377.
Jink, J. L., and H. S. Pooni. 1984. Comparsion of inbred lines produced by single seed descent and pedigree inbreeding. Heredity. 53:299-308.
Johnson, R. A., D. W. Wichern. 1992. Applied Multivariate Statistical Analysis. Prentice-Hall, Inc.. Englewood Cliffs, New Jersey.
Haddad, N. I., and F. J. Muehlbauer. 1981. Comparsion of random bulk population and single-seed-descent methods for lentil breeding. Euphytica. 30:643-651.
Huff. D. R., R. Peakall, and P. E. Smouse. RAPD variation within and among natural populations of outcrossing buffalograss [ Buchloe dactyloides ( Nutt. ) Engelm.]. Theor Appl Genet. 86:
927-934.
Keim, P., W. D. Beavis, J. M. Schupp, B. M. Baltazar, L. Mansur, R. E. Freestone, M. Vahedian, and D. M. Webb. 1994. RFLP analysis of soybean breeding populations : I. Genetic structure difference due to inbreeding methods. Crop. Sci. 34:55-61.
Kephart, S. R. 1990. Starch gel electrophoresis of plant isozymes: A comparative analysis of techniques. Amer. J. Bot. 77(5):693-712.
Khalifa, M. A., and C. O. Qualset. 1975. Intergenotypic competition between tall and dwarf wheat. II in hybrid bulks. Crop Sci. 15:640-644.
Kiang, Y. T., Y. C. Ching, and N. Kaizuma. 1992. Genetic diversity in natural populations of wild soybean in iwate prefecture,Japan. Heredity. 83:325-329.
Martin, R. J., J. R. wilcox, and F. A. Laviolette. 1978. Variability in soybean progenies developed by single seed descent at two plant population. Crop Sci. 18:359-363.
Mymudes, M. S., and D. H. Les. 1993. Morphological and genetic variability in Plantago cordata
(Plantaginaceae), a threatened aquatic plant. Amer. J. Bot. 80(3):351-359.
Nordquist, P. T., O. J. Webster, C. O. Gardner, and W. M. Ross. 1973. Registration of three sorghum germplasm random-mating populations. Crop Sci. 13:132.
Ouendeba, B., G. Ejeta, W. W. Hanna, and A. K. Kumar. 1995. Diversity among African pearl millet landrace populations. Crop Sci. 35:919-924.
Ross, W. M., S. D. Kindler, H. L. Hackerott, T. L. Harvey, A. Sotomayor, O. J. Webster, and K. D. Kofoid. 1977. Registration of RP1R and RP2B sorghum germplasm. Crop Sci. 17: 983.
Roy, N. N. 1976. Inter-genotypic plant competition in wheat under single seed descent breeding. Euphytica 25:219-223.
Saghai-maroof, M. A., K. M. Soliman., R. Jorgenson, and R. W. Allard. 1984. Ribosomal-DNA spacer -length polymorphisms in barley : Mendelian inheritance, chromosomal location , and population dynamics. Proc. Natl. Acad. Sci. U. S. A. 81:8014-8018.
Shah, F. H., O. Rashid, A. J. Simona, and A. Dunsdon. 1994. The utility of RAPD markers for the determination of genetic variation in oil palm (Elaeis guineensis). Theor Appl Genet. 89:
713-718.
J. Am. Soc. Agron. 29:690-696.
Stephens, J. C., and R. F. Holland, 1954. Cytoplasmic male-sterility for hybrid sorghum seed production. Agron. J. 46:20-23.
Stucker, D. S., and A. R. Hallauer. 1992. Genetic variability as affected by selection in lowa stiff stalk synthetic Maize. Heredity. 83:410-418.
Tee, T. S., and C. O. Qualset. 1975. Bulk population in wheat breeding comparison of single seed descent random bulk methods. Euphytica. 24:393-405.
田間疏植
F5
收穫約 570 株(T)
5×5 cm2 密植F5
收穫雄可稔植株 收穫雄可稔植株 收穫 433 株(X)
NTUAC0 (F2)
F3
F4
(A)
約 1000 株(R)
約 650 株(S)
7×7 cm2 密植F5
收穫 313 株(Y)
10×10 cm2 密植F5
收穫 273 株(Z)
圖 1. SSD 法各世代族群之代碼及繁衍流程圖 表 1. 1997 年春作於臺北種植各世代族群之農藝性狀平均值差異顯著性測驗Table 1. Significant tests of agronomic character means for different generation populations planted in the spring of 1997 at Taipei.
Populations Days to flower (days) Green leaf number (No.) Plant height (cm) Stalk diameter (mm) Panicle length (cm) Panicle weight (g) 100-seed weight (g) A 73.88 a 5.875 a 112.75 a 13.33 a 23.25 a 33.43 a 2.635 a R 74.39 a 5.957 a 116.00 a 13.53 a 24.87 a 33.50 a 2.733 a S 74.70 a 6.424 a 115.00 a 14.38 a 23.36 a 39.17 a 2.620 a T 76.29 a 6.292 a 128.88 a 13.23 a 23.96 a 39.54 a 2.830 a a, b, c : Means with different letter are significantly different at 5% level
表 2. 1997 年春作於臺北種植 F5 世代各族群之農藝性狀平均值差異顯著性測驗
Table 2. Significant tests of agronomic character means for different F5 generation populations planted in the spring of 1997 at Taipei.
Populations Days to flower (days) Green leaf number (No.) Plant height (cm) Stalk diameter (mm) Panicle length (cm) Panicle weight (g) 100-seed weight (g) T 76.29 ab 6.292 ab 128.88 a 13.23 ab 23.96 a 39.54 a 2.830 a X 79.36 a 6.968 a 132.42 a 14.11 a 25.97 a 39.26 a 2.682 ab Y 73.67 b 5.909 b 119.18 a 12.62 b 23.03 a 34.37 a 2.488 bc Z 75.27 ab 6.308 ab 120.12 a 12.64 ab 23.58 a 32.35 a 2.248 c
a, b, c : Means with different letter are significantly different at 5% level
表 6. 1997 年春作於臺北種植各世代族群農藝性狀之標準化變數的一般化樣品變方 (R) 及 RAPD 分子標誌變異之 Shannon's 指數 (H0)
Table 6. Generalized sample variance of the standardized variables of agronomic characters and Shannon's indices of RAPD markers diversity for different generation populations planted in the spring of 1997 at Taipei.
R H0
Populations All plants Normal plants All plants Normal plants
A 0.0496 0.0167 0.2493 0.2522 R 0.0657 0.1050 0.2671 0.2522 S 0.0396 0.0375 0.2659 0.2646 T 0.0386 0.0725 0.2614 0.2687 X 0.0766 0.0873 0.2613 0.2638 Y 0.0697 0.0728 0.2635 0.2490 Z 0.0379 0.0408 0.2518 0.2371 表 3. 1997 年春作於臺北種植各世代族群之 RAPD 分子標誌頻度差異顯著性測驗
Table 3. Significant tests of RAPD marker frequency for different generation populations planted in the spring of 1997 at Taipei.
Popu-lation B07835 L04780 L05780 L14730 T02630 T171095 T17730 T17517 T17390 V06645 V06630 V06610 A 0.208a 0.822a 0.938a 0.333a 0.787a 0.729a 0.833a 0.500ab 0.333bc 0.182ab 0.795a 0.205b R 0.083a 0.854a 0.917a 0.188a 0.583b 0.660a 0.717a 0.500ab 0.565a 0.340a 0.596b 0.404a S 0.146a 0.787a 0.936a 0.271a 0.625ab 0.646a 0.688a 0.563a 0.521ab 0.146b 0.771ab 0.229ab T 0.125a 0.766a 0.830a 0.208a 0.625ab 0.583a 0.833a 0.333b 0.208c 0.146b 0.813a 0.188b
表 4. 1997 年春作於臺北種植 F5 世代各族群之 RAPD 分子標誌頻度差異顯著性測驗 Table 4. Significant tests of RAPD marker frequency for different F5 generation populations
planted in the spring of 1997 at Taipei. Popu-lation B07835 L04780 L05780 L14730 T02630 T171095 T17730 T17517 T17390 V06645 V06630 V06610 T 0.125a 0.766a 0.830a 0.208a 0.625a 0.583a 0.833a 0.333b 0.208ab0.146a 0.813a 0.188a X 0.130a 0.702a 0.875a 0.146a 0.688a 0.646a 0.750a 0.479ab0.208ab0.128a 0.872a 0.128a Y 0.125a 0.833a 0.938a 0.083a 0.667a 0.625a 0.681a 0.319b 0.149b 0.188a 0.708a 0.292a Z 0.083a 0.729a 0.917a 0.104a 0.750a 0.604a 0.813a 0.604a 0.354a 0.149a 0.787a 0.213a a, b, c : Means with different letter are significantly different at 5% level
表 5. 1997 年春作於臺北種植各世代族群各農藝性狀與 RAPD 分子標誌間相關性之顯著性 測驗#
Table 5. Significant tests of correlation between agronomic characters and RAPD marker for different generation populations planted in the spring of 1997 at Taipei.
RAPD markers Days to flower Awn length Green leaf number Plant height Stalk diameter Peduncle length Fertility Panicle length Panicle weight B07835 t y L04780 r z s S L05780 -z -z -z L14730 y y Y T02630 -S r r T171095 T17730 -a z T17517 s x s x T17390 -y z x V06645 Z a y Z a x -r -y x V06630 -z -a -y -z -a -x -z -x V06610 z a y z a x z x t W03660 -T z W11385 z z -a -y W151040 z -a y S -A s W15780 -y S W15630 -x -r -r t -z