國 立 交 通 大 學
生物科技研究所
碩士論文
利用重組鋅
-原紫質及鋅-乙炔苯酸紫質
之不同肌紅蛋白突變株進行其光能轉化
學能的研究
Light-Chemical Energy Conversion Study of
Reconstituted Zinc-Protoporphyrin/Zinc-
Carboxyphenylethynyl Porphyrin Myoglobin
Mutants
研究生:歐書涵
指導教授:吳東昆
博士
利用重組鋅
-原紫質及鋅-乙炔苯酸紫質之不同肌紅蛋白突變
株進行其光能轉化學能的研究
Light-Chemical Energy Conversion Study of Reconstituted
Zinc-Protoporphyrin/Zinc- Carboxyphenylethynyl
Porphyrin Myoglobin Mutants
研究生:歐書涵
Student: Shu-Han Ou
指導教授:吳東昆
博士 Advisor: Prof. Tung-Kung Wu Ph.D
國 立 交 通 大 學
生物科技研究所
碩士論文
A Manuscript of Dissertation
Submitted to Department of Biological Science and Technology
College of Biological Science and Technology
National Chiao Tung University
in partial Fulfillment of the Requirements for
the Degree of Master of Philosophy
in
Biological Science and Technology Hsinchu,
Taiwan, Republic of China
July, 2010
I
利用重組鋅
-原紫質及鋅-乙炔苯酸紫質之不同肌紅蛋白突變
株進行其光能轉化學能的研究
中文摘要
經過定點突變的技術,可以將野生型肌紅蛋白 His-64、Val-68、Ile-107 分別 突變成 Asp、Leu、Met。研究指出,經突變的肌紅蛋白具有下列兩種特性:(1) 過氧化酶活性,(2) 能夠穩定帶有自由基的陽離子 (FeIV = O Por+‧ ),進而延長電 荷分離的時間。肌紅蛋白的結構已經藉由高解析X-ray 解出,且基本特性也已分 析完成,因此肌紅蛋白很適合用來當作電子轉移以及化學活性分析的模型。為了 將 野 生 型 及 突 變 型 肌 紅 蛋 白(MbWT,MbH64D, MbV68L, MbI107M, MbH64D/V68L, MbH64D/V68L/I107M) 應用在以蛋白質為基礎的光-化學能量轉換的系統中及比較其 突變的影響,因此,我們將輔基團 鋅-原紫質 (ZnPP) 以及 鋅-乙炔苯酸紫質 (ZnPE1) 重組到肌紅蛋白內。藉由紫外光-可見光光譜及螢光光譜分析其生物物 理特性以確定輔基團成功地重組到蛋白質的活性中心,並藉此算出其能隙 (energy gap)。此外,我們也經由循環伏安法及微差脈衝伏安法求出重組金屬紫質 -肌紅蛋白複合體的 HOMO/LUMO 電位。在螢光生命期的比較中發現,將輔基 團重組到肌紅蛋白內的生命期比單獨輔基團溶解在四氫呋喃 (THF) 內還要長。 以蛋白質為基礎的光化學能量轉換的系統包含重組金屬紫質-肌紅蛋白複合體當 做感光酵素、三乙醇胺 (TEA) 當作電子提供者以及菸鹼胺腺呤雙核酸磷酸鹽 (NADP+) 當作電子接受者。比較不同輔基團以及突變點的重組金屬紫質-肌紅蛋 白複合體中,ZnPE1-MbV68L具有最好的NADP+ 還原效率 (94.7%) 以及連續照光 6 小時的反應速率 (0.209 mM/hr)。II
Light-Chemical Energy Conversion Study of Reconstituted
Zinc-Protoporphyrin/Zinc- Carboxyphenylethynyl
Porphyrin Myoglobin Mutants
Abstract
After replacing His 64 with Asp, Val 68 with Leu, and Ilu 107 with Met of myoglobin (Mb) via molecular engineering, two properties exist: (1) peroxidase activity and (2) the ferryl porphyrin cation radical (FeIV = O Por+‧
) stabilization for prolonging charge separation. Because the structure of myoglobin had been successfully solved by high-resolution x-ray crystallography and possess the characterization of availability, it is a suitable model to understand the electron-transfer (ET) reaction as well as the chemical reactivity. In order to apply these proteins into protein-based photo-chemical energy conversion system and compare the mutation effect, various Mb mutants, ranging from mono (MbH64D, MbV68L, MbI107M), double (MbH64D/V68L), and triple (MbH64D/V68L/I107M) mutations were reconstituted with prosthetic groups (ZnPP and ZnPE1) to generate metallo-porphyrin
Mb complex. Biophysical properties characterization of the metallo-porphyrin Mb complexes, using UV-Vis and fluorescence spectrometry, validates the successful reconstitution of the prosthetic group inside the active site pocket and energy gap of the reconstituted metallo-porphyrin Mbs. Cyclic voltammetry and differential pulse voltammetry techniques further confirmed the HOMO/LUMO of the reconstituted metallo-porphyrin Mbs. In the picosecond fluorescence decays also showed apo-Mb reconstituted with porphyrin prolonged the lifetime compared with porphyrin in THF. The protein-based photo-chemical energy conversion systems were constructed, using
III
mutated apo-Mb reconstituted with different metalloporphyrin-derivatives as photosensitizers, triethanolamine (TEA) as electron donor and nicotinamide adenine dinucleotide phosphate (NADP+) as an electron acceptor. Among different reconstituted metalloporphyrin-Mbs in photoirradiation systems, MbV68L reconstituted with ZnPE1 exhibited the best energy conversion efficiency (94.7 %) and reaction rate
within 6 hours photoirradiation (0.209 mM/hr), as comparable to that of the myoglobin reconstituted with ZnPP in photo-chemical energy conversion system.
IV
謝誌
(Acknowledgement)
經過了兩年的努力,終於如願以償拿到碩士學位。在這個過程中要感謝很多人 陪我ㄧ起度過。碩士班生涯中,最感謝的人莫過於指導教授- 吳東昆博士,在研 究方向以及觀念上的導正與指引,老師無不費煞苦心,殷殷指導,在此獻上我最 由衷的敬意。同時亦感謝 楊裕雄老師、李耀坤老師在百忙之中撥空參加學生的 口試以及審閱論文,並給予許多寶貴的意見與指正,使我的論文能夠更加完善。 此外,讓我能夠在兩年順利畢業,要特別感謝帶我的老大-晉豪學長。在他的 細心指導下,我不只學到了實驗的技術及應有的態度,在論文上也給予許多中肯 的建議。不管是在實驗上或是電腦方面有問題,老大總是會不厭其煩熱心地的給 予協助。小麵包學姊總是很有耐心教導我實驗,並且不吝於分享實驗「小撇步」; 不管什麼事情,她總是用認真嚴謹的態度去處理,我也期許自己能夠像她一樣。 感謝裕國學長在我研究感到困惑時,能夠給予建議,並且幫我建立自信心。感謝 程翔學長提供不同種類的早餐,讓我不再天天都吃宿舍樓下的摩斯和全家;當我 實驗遇到瓶頸的時候,學長總是會扮演「心靈導師」的腳色,幫我做好「心理輔 導」,讓我更有動力地繼續下去。媛婷學姊獨有的思考能力,帶給實驗室不凡的 效率與嚴謹,足以讓學弟妹以此為楷模。晉源學長對待事情樂觀的態度,讓我不 再畏懼眼前面臨的困難。小紅學姊總是懷抱著開朗的心情,讓整個實驗室充滿著 愉悅的氣氛。文鴻學長嘗試以幽默的態度教導我在實驗上的最基本原則,讓我明 瞭培養好奇心與求知慾是一切研究的骨架;感謝學長給予包容與耐心,指導著一 點一滴成長的我。感謝Allen學長像個家庭醫師一樣,可以提供免費的問診以及 醫學相關知識。感謝Mili學姊的好手藝,讓我品嘗到道地的印度料理也讓我的英 文會話有一點進步。感謝已畢業的學長姐,宏城學長、宏明學長、小妹學姊、天 昶學長在生活上以及實驗上的幫忙。感謝碩士班一起打拚的同學們,靜婷、小花、 奕齊,在這一段學習的過程中,有你們在旁邊加油打氣互相勉勵,與你們相處的 時光讓我受益良多。學弟妹世穎、欣芳、欣怡、怡臻、富生、孟兒、欣樺、婉婷、 幫忙分擔實驗室公共事務,有你們在的實驗室格外溫馨,雖然你們還在累積研究 上的經歷,但我特別感謝你們對實驗室的付出與照顧,我衷心期盼未來的你們能 成為學弟妹的典範。感謝專題生孟儒、俞靜、小彥、聖為、奕汝、瑛婷、家嘉, 有你們的加入,實驗室多了一份難能可貴的稚氣,調和大家在實驗之虞的緊繃氣 氛。感謝林敬堯老師實驗室成員幫忙合成ZnPE1化合物;感謝德茹學姊、翊暐撥 空幫忙量測樣品的半生期。 最後,我要感謝我的家人,爸爸、媽媽、小阿姨一家人,因為有你們的支持與 鼓勵,讓我可以無後顧之憂的專心於課業上,順利地走過碩士班兩年的日子。碩 士班生涯已經告一段落,往後的日子又是另一個開始,我會帶著老師們的教誨與 各位的祝福,勇敢面對挑戰。V
Table of Contents
中文摘要 ... I Abstract ... II 謝誌 (Acknowledgement) ... IV Table of Contents ... V Table of Figures... VIII Table of Tables ... XIII1. Introduction ... 1
1.1 THE ENERGY ISSUE ... 1
1.2 PHOTOSYNTHESIS ... 2
1.3 ARTIFICIAL PHOTOSYNTHESIS ... 6
1.4 PORPHYRIN ... 8
1.5 AGGREGATION FORMS OF PORPHYRIN ... 10
1.5.1 H-aggregate ... 11
1.5.2 J-aggregate ... 14
1.6 MYOGLOBIN ... 17
1.6.1 Structure of myoglobin ... 17
1.6.2 Molecular engineering of myoglobin ... 19
1.6.3 The application of myoglobin in electron transfer study ... 21
1.7 PHOTOTRIGGERED CHEMICAL REDUCTION OF RECONSTITUTED ZINC PORPHYRIN MYOGLOBIN ... 25
VI
2. Specific aims ... 29
2.1 RECONSTITUTION OF APO-MYOGLOBIN WITH METALLO-PORPHYRINS... 29
2.2 OPTIC PROPERTIES OF RECONSTITUTED METALLO-PORPHYRIN-MYOGLOBIN (UV-VIS, FLUORESCENCE, AND TCSPC) ... 30
2.3 ELECTROCHEMISTRY ANALYSIS (CYCLIC VOLTAMMETRY ANALYSIS (CV) AND DIFFERENTIAL PULSE VOLTAMMETRY (DPV))... 30
2.4 THE LIGHT-CHEMICAL ENERGY CONVERSION ANALYSIS PHOTOIRRADIATION ASSAY ... 31
3. Experimental materials and procedures ... 32
3.1 EXPERIMENTAL MATERIALS ... 32
3.2 EXPERIMENTAL APPARATUS ... 33
3.3 PURIFICATION OF MUTATED MYOGLOBIN AND EXTRACTION OF REMAINED HEME BY METHYL ETHYLKETONE METHOD ... 33
3.4 RECONSTITUTION OF APO-MYOGLOBIN WITH ZNPP AND ZNPE1 ... 34
3.4.1 Reconstitution of apo-myoglobin with ZnPP ... 34
3.4.2 Reconstitution of apo-myoglobin with ZnPE1 ... 35
3.5 CIRCULAR DICHROISM (CD) SPECTROSCOPOLARIMETRY ... 35
3.6 PICOSECOND FLUORESCENCE DECAYS ... 35
3.7 CYCLIC VOLTAMMETRY ANALYSIS ... 36
3.8 PHOTOIRRADIATION EXPERIMENT ... 36
4. Results and discussion ... 38
4.1 PURIFICATION OF APO-MYOGLOBIN MUTANTS ... 38
4.2 UV-VIS ANALYSIS RESULTS OF RECONSTITUTED METALLO-PORPHYRIN-MBS 39 4.3 CIRCULAR DICHROISM (CD) SPECTROPOLARIMETRY ... 43
VII
4.4 FLUORESCENCE ANALYSIS ... 45
4.5 PICOSECOND FLUORESCENCE DECAYS ANALYSIS ... 48
4.6 CYCLIC VOLTAMMETRY (CV) ANALYSIS ... 55
4.7 PHOTOIRRADIATION ASSAY ... 56
5. Conclusion and future perspectives ... 69
6. References ... 71
7. Appendix ... 75
7.1 CYCLIC VOLTAMMETRY (CV) ANALYSIS ... 75
VIII
Table of Figures
Figure 1-1 The photosynthetic apparatus of the purple bacteria. ... 3
Figure 1-2 The scheme for photosynthesis. ... 5
Figure 1-3 Diagram of an artificial photosynthesis system based on liposome membrane. ... 7
Figure 1-4 Illustration of the Phe-RC/ WO3-TiO2 PE and photo-induced ET. ... 8
Figure 1-5 The structure of chromophores are based on porphyrin. ... 10
Figure 1-6 The interaction between porphyrin and porphyrin macrocycle. ... 12
Figure 1-7 The structure of protoporphyrin IX. ... 12
Figure 1-8 The (A) UV-vis, (B) RLS, and (C) fluorescence emission of protoporphyrin IX. ... 13
Figure 1-9 The molecular model of the H-aggregate at pH 12. ... 13
Figure 1-10 The structure of tetraary-substituted porphyrins. ... 14
Figure 1-11 Absorption spectra of (A) TPPH2 (B) TPyPH2 in dichloromethane and (C) TMPyPH2 (D) TCPPH2 (E) TSPPH2 in water. ... 15
Figure 1-12 Fluorescence spectra of (A) TPPH2 (B) TPyPH2 in dichloromethane
and (C) TMPyPH2, solid line spectra (for excitation wavelengths of
414, 413 and 421 nm, respectively) and dashed line spectra (for excitation wavelengths of 433, 400, and 441 nm, respectively) relate to the absence and presence, respectively, of trifluoroacetic acid (10 vol.%). (D) TCPPH2 (the open circles (a) and solid circles (b)
correspond to fluorescence resulting from excitation at 439 and 467 nm, respectively, when trifluoroacetic acid is present (10 vol.%). (E)
IX
TSPPH2 (the open circles (a) and solid circles (b) correspond to
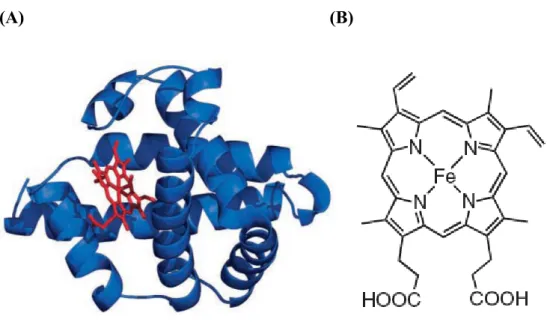
fluorescence resulting from excitation at 432 and 490 nm, respectively, when trifluoroacetic acid is present (10 vol.%). ... 16 Figure 1-13 Two possible linear J-aggregates. ... 17 Figure 1-14 (A) The structure of myoglobin. The backbone of myoglobin consists
of eight α-helices (blue) that wrap around a central pocket containing a heme group (red). (B) The structure of heme. ... 19 Figure 1-15 The proposed mechanism of compound I formation. ... 21 Figure 1-16 Schematic illustration of the reconstitution of (FeIIIOH2)-Ru2+-BXV4+
and (Zn)-Ru2+-BXV4+ with apo-myoglobin to afford the myoglobin-based triads. ... 23 Figure 1-17 The structures of triads. ... 23 Figure 1-18 (A) Energetics and selected pathways for Mb (FeIIIOH2)-Ru2+-BXV4+
in buffer solution after excitation to the Ru(bpy)3 excited state. (B)
Energy levels of the locally excited state of the Ru2+(bpy)3 moiety
and the hypothetical ion-pair states of Mb(Zn)-Ru2+-BXV4+ and the pathways available for their interconversion. ... 24 Figure 1-19 UV-visible spectral. 1 M TEA, 10 μM ZnPP-Mb, and 1 mM NADP+
in 10 mM phosphate butter (pH 9.0) under photoirradiation. ... 26 Figure 1-20 (A) A schematic presentation of the operating principles of the
dye-sensitized solar cell (DSSC). (B) Chemical structure of N-3 dye-cis-di-(thiocyanato)bis(2,2-bipyridyl)-4,4-dicarboxylate)
ruthenium-(II). (C) Schematic drawing showing the currently used embodiment of the DSSC. ... 28
X
Figure 4-1 The SDS-PAGE results of Mbs purification. ... 39 Figure 4-2 UV-Vis spectra results of metallo-porphyrins. ... 41 Figure 4-3 UV-Vis spectra results of (A) ZnPP in THF, 100 mM KPi buffer and
reconstituted with apo-Mb in 100 mM KPi buffer (B) ZnPE1 in THF,
100 mM KPi buffer and reconstituted with apo-Mb in 100 mM KPi buffer. ... 42 Figure 4-4 The CD spectrums of metallo-porphyrins (A) ZnPP (B) ZnPE1
reconstituted with MbWT, MbH64D, MbV68L, MbI107M, MbH64D/V68L, and MbH64D/V68L/I107M. ... 44 Figure 4-5 UV-Vis spectra and fluorescence emission of (A) ZnPP (B) ZnPE1
dissolved in THF, and the exciting wavelength is 435 nm. ... 45 Figure 4-6 UV-Vis spectra and fluorescence emission of (A) ZnPP-MbWT (B)
ZnPP-MbH64D (C) ZnPP-MbV68L (D) ZnPP-MbI107M (E) ZnPP-MbH64D/V68L (F) ZnPP-MbH64D/V68L/I107M. ... 46 Figure 4-7 UV-Vis spectra and fluorescence emission of (A) ZnPE1-MbWT (B)
ZnPE1-MbH64D (C) ZnPE1-MbV68L (D) ZnPE1-MbI107M (E)
ZnPE1-MbH64D/V68L (F) ZnPE1-MbH64D/V68L/I107M. ... 47
Figure 4-8 The correlation of protein concentration and fluorescence intensity in 100 mM KPi buffer. (A) ZnPP-MbWT (B) ZnPP-MbH64D (C) ZnPP-MbV68L (D) ZnPP-MbI107M (E) ZnPP-MbH64D/V68L (F) ZnPP-MbH64D/V68L/I107M. ... 49 Figure 4-9 The correlation of protein concentration and fluorescence intensity in
100 mM KPi buffer. (A) ZnPE1-MbWT (B) ZnPE1-MbH64D (C)
XI
ZnPE1-MbH64D/V68L/I107M. ... 50
Figure 4-10 Picosecond fluorescence transients of reconstituted ZnPP-Mb in 100 mM KPi buffer with λex = 435 nm at λem = 600 nm. ... 52
Figure 4-11 Picosecond fluorescence transients of reconstituted ZnPE1-Mb in 100
mM KPi buffer with λex = 435 nm at λem = 630 nm. ... 53
Figure 4-12 Picosecond fluorescence transients of reconstituted ZnPP and ZnPE1
in THF with λex = 435 nm at λem = 640 nm (ZnPP) and λem = 621 nm
(ZnPE1). ... 54
Figure 4-13 The energy diagram of TEA, ZnPP-Mb, ZnPE1-Mb, and NADP+. .. 56
Figure 4-14 UV-Vis spectral change for 10 μM ZnPP-Mb, 2.5 mM NADP+, and 1 M TEA in 100 mM KPi pH 9.0 under room temperature. ... 58 Figure 4-15 UV-Vis spectral change for 10 μM ZnPE1-Mb, 2.5 mM NADP+, and
1 M TEA in 100 mM KPi pH 9.0 under room temperature. ... 59 Figure 4-16 UV-Vis spectral change for 10 μM ZnPP-Mb, 2.5 mM NADP+, and 1
M TEA in 100 mM KPi pH 9.0 under room temperature and keep in dark for 5 days. ... 60 Figure 4-17 UV-Vis spectral change for 10 μM ZnPE1-Mb, 2.5 mM NADP+, and
1 M TEA in 100 mM KPi pH 9.0 under room temperature and keep in dark for 5 days. ... 61 Figure 4-18 The reduction rate of NADP+ in photoirradiation reaction within 6
hours. ... 63 Figure 4-19 Absorbance change at 340 nm in the solution containing 10 μM ZnPE1-Mb, 1 M TEA, and 2.5 mM NADP+ in 100 mM phosphate
XII
XIII
Table of Tables
Table 4-1 Fluorescence lifetimea of the excited state of ZnPP-Mb in 100 mM KPi
... 54
Table 4-2 Fluorescence lifetimea of the excited state of ZnPE1-Mb in 100 mM KPi ... 55
Table 4-3 Photoirradiation reaction of various ZnPP-Mb mutants on NADP+ reduction ... 64
Table 4-4 Photoirradiation reaction of various ZnPE1-Mb mutants on NADP+ reduction ... 64
Table 4-5 Reaction rate of metallo-porphyrins reconstituted with myoglobin within 6 hours ... 65
Table 4-6 The Soret band changed within 6 hours of ZnPP-Mb ... 65
Table 4-7 The Soret band changed within 6 hours of ZnPE1-Mb ... 66
Table 4-8 Conversion efficiency of ZnPP-Mb ... 68
1
1. Introduction
1.1 The energy issue
For past decades, the major source of energy most came from coal, oil, and natural gas. However, these fossil fuels resources are non-renewable, and they will be run out eventually. In addition, pollution and greenhouse gases were produced and contributed to global warming while using these energy sources. Therefore, finding an alternative energy source is a task of top priority.
Fortunately, sunlight provides organisms energy to maintain the daily metabolism. Plants capture solar energy to catalyze photosynthesis which is a process of photoinduced electron transfer to yield long-lived charge-separated state and facilitates chemical energy production. Besides, the sun energy is the most sufficient, available and easily renewable resource which does not require gaseous fuels to be combusted. For these reasons, the application of sunlight energy can not only decrease greenhouse effect but also be a suitable substituent energy source.
Furthermore, the supply of energy from the sun to the Earth is 3×1024 joules per year1, and more widely than the global population currently consumes. Namely, covering 0.1% of the Earth’s surface with solar cells with an efficiency of 10% would satisfy our present needs1. If we can covert solar energy into an efficient form we can use, it will decrease the requirement of non-renewable resources. Thus, solar energy is suitable and providing an alternative way in finding substituent resource, and we want to construct a system to mimic the photosynthetic process as an alternative energy source.
2
1.2 Photosynthesis
Sunlight energy storage system is found in photosynthetic organisms, including plants, algae and variety types of bacteria. All these organisms utilize sunlight to drive cellular and metabolism processes. After absorbing light by an antenna pigment, these organisms perform photosynthesis by a cascade-like system in the primary reaction site called photosynthetic reaction center (RC) which is an integral membrane pigment-protein that carries out light-driven electron transfer reactions2. Without the energy produced by photosynthesis, these organisms will not be able to continue the chemical reactions to fuel themselves. Hence, understanding the mechanisms of photosynthesis in organisms is a critical role in development of energy transfer system by sunlight energy. The photosynthetic systems are generally divided into bacteria and plants and the details are as following.
Before two billion years, photosynthetic bacteria containing bacteriochlorophyll and carotenoid have existed on earth3, thereby obtaining nutrients to maintain life by absorbing light energy. Among these various photosynthetic bacteria, purple bacteria are the most studied and best characterized. In 1985, the three-dimensional X-ray structure of photosynthetic reaction center (RC) in purple bacteria (Rhodopseudomonas viridis, Rps. V) has been demonstrated4. In most purple bacterial, the photosynthetic membranes consists two types of light-harvesting complexes, light-harvesting complex I (LH-I) and light-harvesting complex II (LH-II). The photosynthetic unit (PSU) involves photosynthetic reaction center (RC) and exists in photosynthetic bacteria or higher plants generally. The PSU is surrounded by LH-I which contacted with LH-II. When the light-harvesting complex absorbs light, excitation energy is transferred to the RC via LH-II→LH-I. The RC binds QB which
3
electron is across the membrane by the exothermic reaction. Therefore, an electron is turned back into RC by cytochrome c2 complex. On the other hand, when electron
crosses the membrane can produce large proton gradient which drive the synthesis of ADP→ATP by the ATPase (Fig.1-1)5, 6.
Figure 1-1 The photosynthetic apparatus of the purple bacteria. The RC (red) is surrounded by LH-I (green). Purple bacterial photosynthesis begins when a photon is absorbed by the light-harvesting apparatus, and excitation is transferred to the RC via LH-II (green) →LH-I. QB is reduced into QBH2 form
and an electron is across the membrane after QBH2 is oxidized by the bc1
complex. Then an electron is back to RC by cytochrome c2 complex (blue). The
synthesis of ATP occurs when electron crosses the membrane produced large proton gradient5.
In plants, photosynthesis begins when light is absorbed by antenna pigments.
Antenna pigments transfer light energy to one of two types photochemical reaction centers which are located on the thylakoid membrane of the chloroplast: P700, which is
4
transferred to electron acceptors, leaving the reaction center in an oxidized state7. In addition, photosynthesis is divided into two stage processes: the light-dependent process (light reactions) in the grana and the light-independent process (dark reactions) in the stroma of the chloroplasts. In light-dependent process, it produces NADPH and ATP when solar energy is captured, and these products are used in the dark reaction to convert CO2 into sugar. In light-dependent process, it contains two forms:
non-cyclic and cyclic reaction. In the non-cyclic reaction, the chlorophyll in the
light-harvesting antenna complex of photosystem II and other accessory pigments absorb light. After getting sufficient excitation energy from adjacent antenna pigments, an electron transfers to the primary electron-acceptor molecule (named pheophytin) via a process called photoinduced charge separation. Electrons pass through by an electron transport chain---Z-scheme8, which generates a chemismotic
potential across the membrane and connects photosystem I and photosystem II. When the photosystem I is excited by absorbing light, a second electron carrier accepts the electron, which pass down the lowering energy of electron acceptors. At the same time, energy releases and drives ATP synthase activated, thus ATP and NADPH synthesized (Fig. 1-2). On the other hand, the cyclic reaction takes place only at photosystme I, and excited electrons from photosystem I can use an electron transport chain to reduce oxidized P700. When the electron is transferred from the photosystem I,
the electron is passed down the electron acceptor molecules and returned back to photosystem I to generate ATP (Fig.1-2)9.
5
(A)
(B)
Figure 1-2 The scheme for photosynthesis. The photosynthetic apparatus in green plants is organized into two-photosystems, photosystem I and photosystem II (PS-I and PS-II, respectively). (A) Noncyclic photophosphorylation (B) cyclic photophosphorylation10.
6
1.3 Artificial photosynthesis
Photosynthesis in bacteria and plant demonstrates the viability of directly converting sunlight into chemical fuels, which involves storing the energy from the incident solar irradiation. Artificial photosynthesis is a system which mimics natural photosynthesis and applies the fundamental scientific principles of the natural process to the design of solar energy conversion systems11. These constructs use different materials, and researchers tune them to produce energy efficiently and in useful forms to humans. Many scientists are attracting extensive interest for the development of artificial photosynthesis. Artificial photosynthesis is also a strategy for producing sustainable fuels.
In 1998, Gali et al. reported liposome-based artificial photosynthesis system12,
13(Fig. 1-3). F
0F1-ATP synthase was incorporated into liposomal bilayers containing
the components of the proton-pumping photosystem: liposomes, F-type ATP synthase, carotene-porphyrin-naphthoquinone molecular triad (1, C-P-Q), lipophilic quinine (Qs). The proton pump was driven by irradiation visible light resulted in generating
the species C‧+—P—Q‧+. This process accompanied with proton translocation by a lipophilic quinine (Qs). Qs accepted an electron to form Qs‧﹣, then Qs‧﹣ was be
protonated. The semiquinone H Qs‧﹣ deliversed the proton and electron to the site of
oxidation potential (C‧+—P—Q) located near the inner membrane surface by either diffusion or self-exchange reaction among the Qs molecules. Oxidation of HQs. near
the internal aqueous interface results in proton ejection to the intraliposomal volume and the generation of protonmotive force (p.m.f.). On accumulation of sufficient p.m.f., proton flow through the coupling factor with the concomitant formation of ATP from adenosine diphosphate (ADP).
7
Figure 1-3 Diagram of an artificial photosynthesis system based on liposome membrane. Included in this diagram is the photocycle that pumps protons into the interior of the liposome and the F0F1-ATP synthas. C-P-Q :
carotene-porphyrin-naphthoquinone molecular triad;Qs:lipophilic quinine12.
In 2006, Kong et al. constructed a novel photoelectrode which contains pigment-exchanged photosynthetic reaction center trapped on the mesoporous WO3-TiO2 film (Fig.1-4). This WO3-TiO2 film provide high surface, promote the
photo-induced ET (electron transfer) of RC, and result in higher capability of separating photogenerated carriers than single TiO2 or WO3. In addition, improved
photo-induced ET (electron transfer) from RC mutant to the WO3-TiO2 film
(replacement of pigment Bphe with Phe) via experimental results of femtosecond (fs) pump-probe dynamics, fluorescence (FL) emission and near-infrared (NIR) photoelectric responses, and this strategy decreases the charge recombination of RC
8
itself and also provide an alternative system to probe the photo-induced multiple-pathway ET of photosensitive chromophores6, 14.
Figure 1-4 Illustration of the Phe-RC/ WO3-TiO2 PE and photo-induced ET. Cb:
conduction band; Vb: valence band; P:bacteriochlorophyll (Bchl) dimmer; Bphe: bacteriopheophytin; Phe; pheophytin; QA: the primary quinine; QB: the
secondary quinine6.
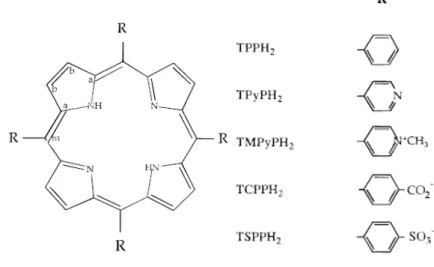
1.4 Porphyrin
Porphyrin is a heterocyclic macrocycles characterized by the presence of four pyrrole rings interconnected via methine bridges15, 16. Porphyrins may occur in nature and physiological function, and the most well-known are the pigment in red blood cells and photosynthesis. In photosynthesis of plants and bacteria, light energy collected by antenna pigments, such as chlorophyll (Fig.1-5A) which is a
9
chromophore, and is efficiently converted to chemical energy. The basic structure of chlorophyll is porphyrin and it plays a critical role in photosynthesis process. In our system, zinc-5-(4-carboxy-phenylethynyl)-10, 20-biphenylporphines (ZnPE1) and
Zinc (II) protoporphyrin (ZnPP) will be used as chromophores in the following analysis (Fig.1-5B, C). ZnPP contains a porphyrin-based structure as a light-harvesting center and two carboxylic acid groups. The structure of ZnPE1 is
comprised by three parts: a zinc biphenylporphyrin (ZnBPP) unit serves as light-harvesting center to provide photoinduced electrons, the phenyl group (PE) group is connected to the meso position of the porphyrin to transfer the electrons efficiently, and the carboxylic acid (CA) attached to the end of the spacer serves as an anchoring group to bond tightly onto the surface of TiO2 nanoparticles17.
10
(A) (B)
(C)
Figure 1-5 The structure of chromophores are based on porphyrin. (a) The structure of chlorophyll a (b) zinc-5-(4-carboxy-phenylethynyl)-10, 20-biphenylporphines (ZnPE1) (c) Zinc (Ⅱ) protoporphyrin (ZnPP).
1.5 Aggregation forms of porphyrin
Because the planar structure of porphyrin, it does not easily exist in a monomer form. The structure of porphyrin is macrocycle which is considered as a rigid hydrophobic box, and it has a particular packing character. The aggregation is a geometry which places the pyrrole ring of one porphyrin directly above the π-cavity at the center of the other; such an arrangement minimizes π-π repulsion and maximizes attraction between the σ-framework around the inner edge of the π-cavity of one
11
porphyrin with the an-electrons of the pyrrole ring immediately above. The role of the metal enhances porphyrin aggregation resulting from metalation places a large positive charge in the central cavity of the porphyrin which leads to a favorable interaction with the π-electrons of the pyrrole of the other porphyrin, and it dimerizes with a lateral shift. The geometric dimensions of aggregation box are 0.7×0.7×0.35 nm-3(Fig.1-6)18, 19. The aggregation forms of porphyrins contain two types : face-to-face arrangement to form H -aggregate and head-to-tail to form J-aggregate. The details of aggregation forms are as following.
1.5.1
H-aggregate
The aggregates in solution exhibit distinct changes in the absorption band as compared to the monomer forms. The H-aggregate causes hypsochromically shift (blue shifts); hypsochromically shift is a change of spectral band to a shorter wavelength (higher frequency)20, 21.
In 1991, Inamura et al. determined the aggregation of protoporphyrin IX at different pH values by using a combination of UV-Vis spectroscopy and size-exclusion gel chromatography22. Protoporphyrin IX is an iron-free form of hemin which is the most common natural porphyrin23(Fig. 1-7). At pH 1, the sharp band in UV-Vis spectroscopy is at 406 nm; the fluorescence emission showed two bands at 604 nm and 660 nm and the intensity is greater than others. At pH 4.8, a broadened split Soret band is observed with maxima at 352 nm and 450 nm; the fluorescence emission is almost quenched result in no signals. At pH 12, the Soret band is blue shift to 382 nm; the fluorescence emission is red-shifted respect to that of the species existing at acidic pH 4.8 (Fig.1-8). These results indicated that the aggregation behavior of protoporphyrin IX at pH 1 was a monomer form resulting from the protonation of both the propionate end groups and the two nitrogen atoms of the core;
12
the higher aggregate obtained at pH 4.8; the dimmer form of protoporphyrins at pH 12 exhibits the spectral features of H-dimer. According to molecular model for the H-dimer at pH 12, two porphyrin arranges on opposite site via face-to-face H aggregation, and one-fourth of a porphyrin ring overlaps (Fig.1-9)23.
Figure 1-6 The interaction between porphyrin and porphyrin macrocycle18.
13
Figure 1-8 The (A) UV-vis, (B) RLS, and (C) fluorescence emission of protoporphyrin IX under different pH conditions: pH=1 (solid line), pH=4.8 (dotted line), pH=12 (dashed line)23.
14
1.5.2
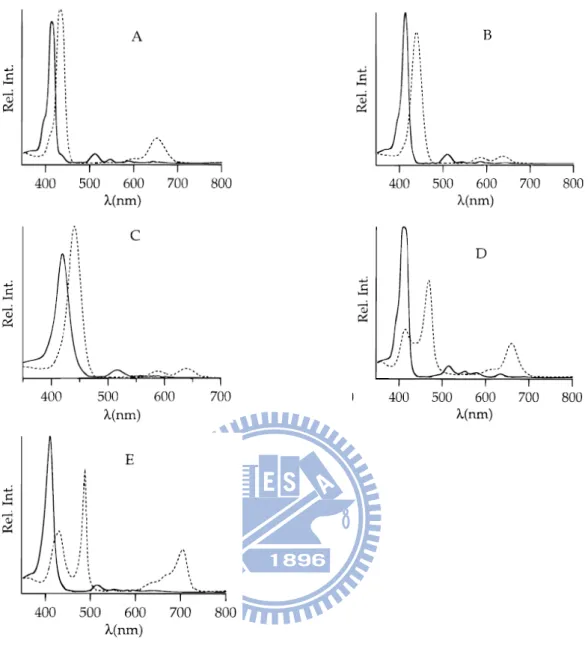
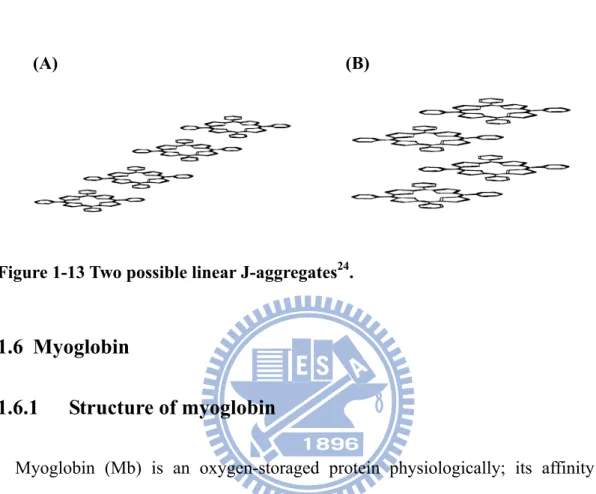
J-aggregate
In general, the J-aggregate causes bathrochromically (red-shifted wavelengths)20, 21. In 1996, Akins et al., studying the aggregation of tetraary-substituted porphyrins (Fig.1-10) in homogeneous aqueous acidic solution (trifluoroacetic acid)24. From these spectra (Fig.1-11), all spectral patterns shifted to the longer wavelengths (red-shifted) upon addition of trifluoroacetic acid (10 vol. %). In addition, TCPPH2
and TSPPH2 showed the second more red-shifted band in the absorption spectra
(Fig.1-11D, E). Fluorescence spectra are show in Fig.1-12. After addition of trifluoroacetic acid, the fluorescence emission patterns are different from absence of acid. Besides, using longer wavelengths to excite TCPPH2 and TSPPH2 showed the
emission wavelength more red-shifted. Upon addition of trifluoroacetic acid, the protonation of TCPPH2 and TSPPH2 (TCPPH4+ and TSPPH4+) in the two imino
nitrogen atoms of the pyrrolenine-like ring interact to the negative aryl group resulting in coplanar alignment. Such arrangement is the head-to–tail alignment define J-aggregate and leads to red-shifted (Fig.1-13).
15
Figure 1-11 Absorption spectra of (A) TPPH2 (B) TPyPH2 in dichloromethane and
(C) TMPyPH2 (D) TCPPH2 (E) TSPPH2 in water. Solid line and dashed line
spectra relate to the absence and presence, respectively, of trifluoroacetic acid (10 vol. %)24.
16
Figure 1-12 Fluorescence spectra of (A) TPPH2 (B) TPyPH2 in dichloromethane
and (C) TMPyPH2, solid line spectra (for excitation wavelengths of 414, 413 and
421 nm, respectively) and dashed line spectra (for excitation wavelengths of 433, 400, and 441 nm, respectively) relate to the absence and presence, respectively, of trifluoroacetic acid (10 vol. %). (D) TCPPH2 (the open circles (a) and solid circles
(b) correspond to fluorescence resulting from excitation at 439 and 467 nm, respectively, when trifluoroacetic acid is present (10 vol. %). (E) TSPPH2(the
17
open circles (a) and solid circles (b) correspond to fluorescence resulting from excitation at 432 and 490 nm, respectively, when trifluoroacetic acid is present (10 vol. %)24.
(A) (B)
Figure 1-13 Two possible linear J-aggregates24.
1.6 Myoglobin
1.6.1
Structure of myoglobin
Myoglobin (Mb) is an oxygen-storaged protein physiologically; its affinity of oxygen is between those of hemoglobin and cytochromes. In diving animals of the sea such as whales, seals and penguins, they contain extremely sufficient myoglobin in muscle tissue that can enhance the time of diving in the sea. Myoglobin is a single-chain globular protein which folded by eight α-helix containing 153 amino acids (Fig.1-14). The molecule weight is approximately 17 kDa, and it is also a member of hemoprotein family25, 26.
A functional myoglobin contains a prosthetic group named heme (protoporphyrin IX iron complex) via non-covalent interactions that actually combines with oxygen. At the center of the heme is a Fe (II) atom which has six coordination sites. Four coordination sites around Fe (II) atom are occupied by nitrogen atoms to from a planar porphyrin ring. The fifth coordination site is occupied by a nitrogen atom from
18
a histidine side chain on one of the amino acids in the protein. The last coordination site is available to bind an O2 molecule. For this reason, heme is the oxygen-carrying
part of the myoglobin molecules.
In 1958, John Kendrew and associates successfully solved the structure of myoglobin by high-resolution x-ray crystallography which shows the position of residues for myoglobin27. The hydrophobic amino acid residues are folded inner; the hydrophilic ones are positioned at outer surface. Accordingly, this character provides myoglobin solubilized in water28.
Over the past years, myoglobin has been investigated in structure and function by many scientists. Because of its less bulky effect, it is a suitable model to understand the electron-transfer (ET) reaction and chemical reactivity of ferryl myoglobin29. On the other hand, myoglobin has many advantages: (1) high-level expression of sperm whale myoglobin by E. coli system30, 31, (2) the cavity of apo-myoglobin can accommodate macrocycle, such as ZnPE1 and ZnPP, that prevents porphyrin
aggregation, (3) the interaction between myoglobin and heme is non-covalent that provides an easy way to replace the prosthetic group32. Owing to above properties, myoglobin is a suitable protein for protein engineering and to reconstitute with different porphyrins for further study.
Myoglobin is a protein with a rich and varied history, has recently become the object of renewed interest regarding its potential roles beyond those previously characterized.
19
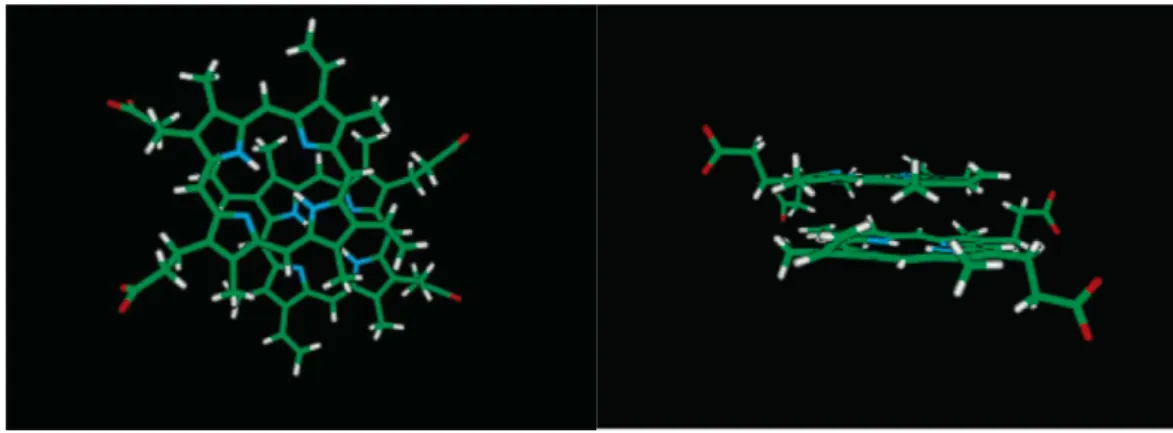
(A) (B)
Figure 1-14 (A) The structure of myoglobin. The backbone of myoglobin consists of eight α-helices (blue) that wrap around a central pocket containing a heme group (red). (B) The structure of heme25.
1.6.2
Molecular engineering of myoglobin
The four general functions of hemoproteins contain (1) electrons transporter (cytochrome b5) (2) oxygen carrier and storage (hemoglobin and myoglobin) (3) the
sensor of oxygen or carbon monoxide (FixL, CooA) (4) catalysis of redox reaction (horseradish peroxidase, cytochrome P450, catalase, NO synthease, NO reductase). Though these different functions, all of the proteins have iron protoporphyrin IX (heme) as a prosthetic group. It’s suggested that the amino acid residues surrounding the prosthetic group control the intrinsic functions of heme33.
The heme iron usually coordinates and reacts with H2O2, oxygen, and other ligands.
When peroxidase reacts with H2O2, a special species - the ferryl porphyrin cation
20
H2O2 binds to the iron of heme, and the distal histidine serves as a general base to
protonation. The protonated distal histidine then transfers the proton to the singly protonated oxygen atom of the bound HO2- as a general acid resulting in O-O bond
cleavage and release water molecule. The positive charged distal arginine can help the polarization of O-O bond, and the negative charged distal histidine stabilize the transition state of O-O cleavage. The catalysis functions of the distal histidine and arginine has been described as the “pull effect” (Fig.1-15)33, 34.
A common strategy for studying the relationship between structure and function is molecular engineering. In 1999, Ozaki et al attempted to replace the distal histidine on position-64 from sperm whale via site-directed mutagenesis that two properties have been proved via a series of myoglobin molecular engineering: (1) peroxidase activity and (2) the ferryl porphyrin cation radical (FeIV=O Por+‧) stabilization for prolonging charge separation35. Furthermore, the residue 68 is important in the access for the substrate to the heme pocket. In 2003, Ozaki et al., using the H64D/V68X mutated myoglobins to study the influence in peroxidase activity36, 37. In 2006, Lin et
al., constructed the triple mutation H64D/V68L/I107M to further improve the
peroxidase activity of myoglobin. All these mutated myoglobins not only can stabilize the compound I but also enhance charge-separation state. In order to further study the role of myoglobin on the property of prolonging charge separation, we use these mutated myoglobins to do the sequential study.
21
Figure 1-15 The proposed mechanism of compound I formation. The distal histidine first serves as a general base to protonation, than the protonated distal histidine cheavage O-O bond results in the release of a water molecule. The positively charged distal arginine also contributes to this process by polarization of the O-O bond33.
1.6.3
The application of myoglobin in electron transfer study
The fundamental energy conversion process of photosynthesis is to yield the long-lived charge-separated stages. The electron transfer steps in the photosynthetic bacteria are needed with efficient and fast moving from the initial bacteriochlorophylls molecule via bacteriophenophytins intermediate to the final quinones acceptors. Some of these small molecules are embedded in the protein matrix. The structural protein controlling the charge separation or the electron transfer is believed a major important aspect in photosynthesis process. Artificial photosynthetic reaction centers were first successfully constructed to satisfy many requirements in the nature.
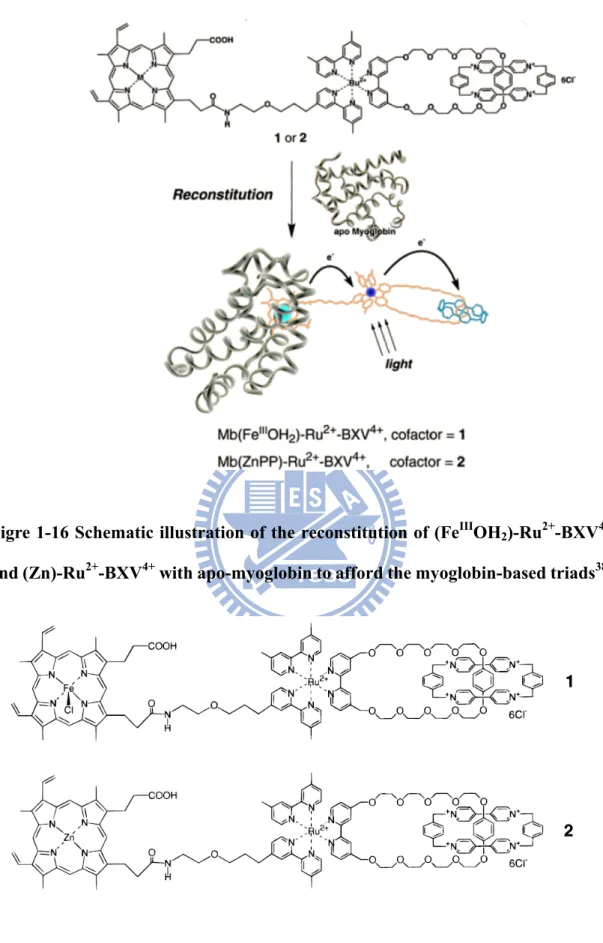
In 2000, Hu et al incorporated a synthetic dye into apo-myoglobin that constructed a semisynthetic chromophore-protein system (Fig.1-16)38. This system mimics the
22
function of photosynthetic organisms to convert light energy to chemical energy in the form of long-lived charge-separated states. The dyed-molecule consists two parts, (FeIIIOH2)-Ru2+-BXV4+ and (Zn)-Ru2+-BXV4+ which linked in a catenane type with
the sensitizer (Ru(bpy)3). Reconstitution of apo-myoglobin with dyed-molecule forms
Mb(FeIIIOH2)-Ru2+-BXV4+ and Mb(Zn)-Ru2+-BXV4+ (Fig.1-17). The electron transfer
processes in the Mb-triads complex has been investigated by steady-state UV-Vis absorption, fluorescence and transient absorption spectra (Fig.1-18). The result showed without Mb matrix, the BXV3+‧ radial absorption spectra rapidly decayed within 1 μs and the multistep electron transfer did not occur. In summary, fixation of the chromophore in a protein matrix not only regulates the vectorial, multistep electron transferring pathway but also stabilizes the charge separated state. The structural aspect might come from the stabilizing or extending the U-shape-like conformation of triad to more expanded form by its incorporation into apo-Mb. Therefore, protein plays a critical role in biological electron transfer.
23
Figre 1-16 Schematic illustration of the reconstitution of (FeIIIOH2)-Ru2+-BXV4+
and (Zn)-Ru2+-BXV4+ with apo-myoglobin to afford the myoglobin-based triads38.
Figure 1-17 The structures of triads. 1: (FeIIIOH2)-Ru2+-BXV4+; 2:
24
(A)
(B)
Figure 1-18 (A) Energetics and selected pathways for Mb (FeIIIOH2)-Ru2+-BXV4+
in buffer solution after excitation to the Ru(bpy)3 excited state. The energy of the
excited Ru(bpy)3 component was calculated from the absorption and emission
25
estimated from the corresponding redox potentials of models for the components of Mb(FeIIIOH2)-Ru2+-BXV4+. The energy levels are not corrected for any
Coulombic stabilization of the intermediates. (B) Energy levels of the locally excited state of the Ru2+(bpy)3 moiety and the hypothetical ion-pair states of
Mb(Zn)-Ru2+-BXV4+ and the pathways available for their interconversion. For comparison purpose, the excited states of ZnPP are not present38.
1.7 Phototriggered chemical reduction of reconstituted zinc
porphyrin myoglobin
In 1999, Nishiyama et al. constructed a simple model of photosystem I by forming NADPH39, 40. Phototriggered electron transfer occurred when zinc porphyrin (ZnPP) was reconstituted with apo-myoglobin to forming ZnPP-Mb as a photo-sensitized enzyme, and the triethanolamine (TEA) as an electron donor. Upon photoirradiation, the absorbance increase at 340 nm ascribed to the reduction of NADP+ to form NADPH (Fig. 1-19). On the other hand, the longer lifetime of triplet state for ZnPP-Mb could have more ability to transfer excited electrons to NADP+ to cause the photoinduced reduction. It’s implied that protein plays a critical role in these electron transfer system.
26
Figure 1-19 UV-visible spectral. 1 M TEA, 10 μM ZnPP-Mb, and 1 mM NADP+ in 10 mM phosphate butter (pH 9.0) under photoirradiation. (a) Before photoirradiation. (b) After 3.5 h of photoirradiation with visible light (λ > 390 nm) at an intensity of 2.8 mW. Following the photoirradiation, spectra were measured after (c) 1, (d) 2, (e) 3, (f) 6, and (g) 9 days in the dark under an atmosphere of N240.
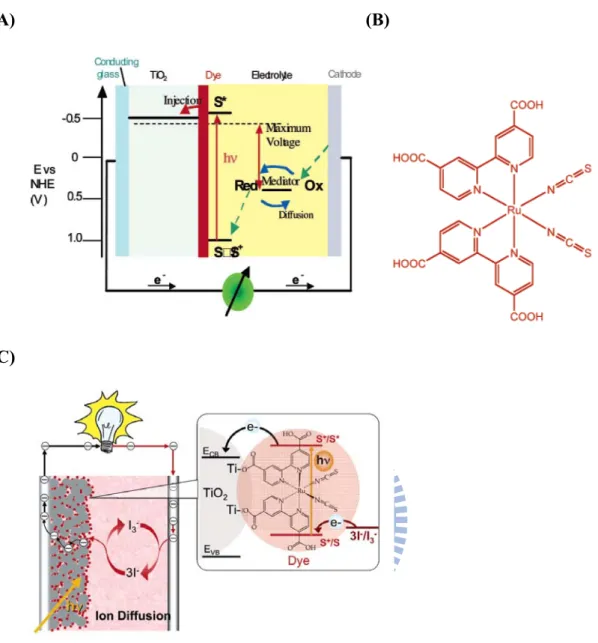
1.8 Dye-sensitized solar cell
Photovoltaic (PV) or solar cell is a device converting energy of sunlight into electric form by photovoltaic effect, which intriguing many scientists in this promising and renewable technique. Photovoltaic has been dominated by solid-state junction devices, which made from single crystal, polycrystal, and amorphous silicon. Among them, the dye-sensitized solar cell (DSSC) have promising potential and regard as a low-cost alternative to conventional p-n junction solar cells41. Typical DSSC consists of (1) two conductive transparent glasses (conducting glasses and
27
cathode), (2) photoactivated-dye molecule, (3) mesoscopic semiconductor oxide film (TiO2, ZnO, Nb2O5), and (4) a liquid electrolyte containing the I3-/I- redox couple.
Monolayer photoactivated-dye molecule is adsorbed on a mesoporous of network with wide band gap semiconductor TiO2. When the dye is excited by photoexcitation,
an electron is injected into the conductor band of the semiconductor oxide. To complete the circuit, the dye molecule regenerated an electron by the electrolyte I3-/I
-oxidation (electron donor) which recaptured an electron of migration through the external load (Fig.1-20)42.
The most common dye material is Ru-based dye (Fig.1-20 b, c). A compound named as N3 (cis-di-(thiocyanato) bis(2,2-bipyridyl)-4,4-dicarboxylate) ruthenium-(II)) or N-719 (bis(tetrabutylammonium) cis-di(thiocyanato) bis (2,2’-bypiridine-4,4’ carboxylic acid)ruthenium-(II)) dye were found to be an outstanding solar light absorber and charge-transfer sensitizer43-45. N-719 which was absorbed on mesoscopic TiO2 film exhibited the incident monochromatic
photo-to-current conversion efficiency (IPCE) 90 %46; on the other hand, it also showed 0.72 fill factor yielding 9.18 % conversion efficiency.
In addition, the dye molecule absorbed on TiO2 has to satisfy the following
demanding conditions: (1) it must be steadily onto the semiconductor oxide surface and inject electrons into the conduction band with a quantum yield of unity, (2) to regenerate rapidly via electron donation from the electrolyte, the redox potential should be greatly high, (3) it should be stable enough to sustain at least 108 redox turnovers under illumination43.
28
(A) (B)
(C)
Figure 1-20 (A) A schematic presentation of the operating principles of the dye-sensitized solar cell (DSSC). (B) Chemical structure of N-3 dye-cis-di-(thiocyanato)bis(2,2-bipyridyl)-4,4-dicarboxylate) ruthenium-(II). (C) Schematic drawing showing the currently used embodiment of the DSSC. It employs mesoscopic semiconductor oxide film TiO2 as light-harvesting units. The
sensitizer is N-719 dye. The redox system employed to regenerate the dye and transport the positive charges to the counter electrode is the iodide/triiodide couple dissolved in an organic electrolyte or in a room-temperature ionic liquid42.
29
2. Specific aims
Due to the structure of porphyrins are easily to self-aggregation resulting in broad band, band shift, fluorescence quenched, and decrease the lifetime of charge separation, we reconstituted metallo-porphyrins with apo-myoglobin to let them as a monomer form inside the protein and attempt to stabilize and prolong the lifetime of excited state.
On the other hand, in the light energy conversion mechanism, the photo-sensitized molecule should have longer charge-separation lifetime to prevent charge recombination. After molecular engineering, myoglobin has peroxidase activity and stabilization of the ferryl porphyrin cation radical (FeIV=O Por+‧) for prolonging the lifetime of charge separation. Previously, we have shown myoglobin plays a critical role in photo-induced electron transfer. Therefore, it gives us an aspect to investigate whether the mutated apo-myoglobin reconstituted with metallo-porphyrins can enhance the lifetime of charge separation.
2.1 Reconstitution of apo-myoglobin with metallo-porphyrins
First of all, we have to get apo-myoglobin via expressed myoglobin in inclusion bodies within E. coli system and ethyl methyl ketone method47. After getting apo-myoglobin, we reconstitute it with metallo-porphyrins by adjusting pH to induce apo-myoglobin unfolding and refolding. This procedure can prevent porphyrin from aggregation and prolong the lifetime of charge separation. This method is convenient to us for further experiments.
30
2.2 Optic properties of reconstituted metallo-porphyrin-myoglobin
(UV-Vis, fluorescence, and TCSPC)
When successfully reconstituting apo-Mb with metallo-porphyrin, we use UV-Vis spectra to check and compare the peak shift with only metallo-porphyrin. In order to make sure the secondary structure of metallo-porphyrin-Mb after reconstitution, circular dichroism (CD) spectroscopolarimetry can be used to insure the protein keeping in natural form. The fluorescence emission spectra which are excited at Soret band will be analyzed and getting preliminary information of photo-induce electron transfer process. We also try different concentration condition in fluorescence analysis to study the relationship between concentration and the intensity of fluorescence. In addition, we use fluorescence and UV-Vis spectra to calculate the energy gap. In the time-correlated single-photon-counting system (TCSPC) and fluorescence optically gated (up-conversion) system (FOG), we distinguish photo-induced charge-separated state and understand the details of the photo-induced electron transfer mechanism. In our proposal, these results are essential basic optic properties to the following experiments.
2.3 Electrochemistry analysis (cyclic voltammetry analysis (CV) and
differential pulse voltammetry (DPV))
Cyclic voltammetry (CV) is a method to study the redox and electron transfer process of substance. We can use the reduction potential, oxidation potential, fluorescence and UV-Vis analysis to calculate the HOMO, LUMO and energy gap. We can explain the rationality of light conversion mechanism by means of the energy levels diagram. Differential pulse voltammetry (DPV) is more sensitive than cyclic voltammetry, so we confirm the redox potential via this technique.
31
2.4 The light-chemical energy conversion analysis by
photoirradiation assay
We study the light-chemical energy conversion by photoirradiation assay which can detect light energy converted to chemical energy. Phototriggered electron transfer occurs when metallo-porphyrin is reconstituted with apo-myoglobin as a photosensitized enzyme, triethanolamine (TEA) as an electron donor, and NADP+ as an electron acceptor. We compared the reaction rate within 6 hours and photo-induced NADP+ conversion efficiency after 5 days of different reconstituted metallo-porphyrins-myoglobin mutants.
32
3. Experimental materials and procedures
3.1 Experimental materials
Acetic acid (Merck)
30% Acrylamide (GE Healthcare) APS (GE Healthcare)
2, 2-azino-bis (3-ethybenthiazoline 6-sulfonic acid) (ABTS) (Sigma) Amicon ultrafitration membrane (Millipore)
Coomassie® Brilliant blue R 250 (Merck)
DEAE Sephacel (Amersham Pharmacia Biotech)
Didodecyldimethylammonium bromide (DDAB) (Aldrich) Dimethyl sulfoxide (MP Biomedicals)
Ethyl methl ketone (Merck) Hydrogen chloride (Merck)
LMW-SDS Marker Kit (Amersham Pharmacia Biotech) Methanol (Merck)
Metallo-5-(4-carboxy-phenylethynyl)-10, 20-biphenylporphines (C.-Y. Lin Lab) Nicotinamide adenine dinucleotide phosphate (NADP+) (Fluka)
Potassium chloride (Merck) Potassium diphosphate (Merck) Potassium phosphate (Merck) Sodium acetate (Merck) Sodium bromide (Merck) Sodium chloride (AMRESCO)
33
Sodium hydroxide (Merck) TEMED (GE Healthcare) Tris base (USB)
Triethanolamine (TEA) (Merck)
3.2 Experimental apparatus
AllegraTM 21R Centrifuge (Beckman Coulter)
8453 UV-Visible Spectrophotometer (Agilent Technologies) Fluorescence Spectrophotometer (USB2000FLG, Ocean Optics) CHI electrochemical analyzer (CH Instruments)
Jasco J-815 spectropolarimeter
Photochemical reactor PR-2000 (Panchum)
Time-correlated single photon counting (Fluo Time 200, PicoQuant)
3.3 Purification of mutated myoglobin and extraction of remained
heme by methyl ethyl ketone method
The clone of myoglobin mutants have been constructed and stocked at -80 ℃ by previous lab members. 3 mL LB media contained the stock of myoglobin and 3 μM kanamycin was with a 37 ℃ overnight culture. Amplifying in 1 L LB culture medium was induced by IPTG, and the culture was grown at 37 ℃ until it reached an OD600 of
between 0.5~1 when IPTG was added to a final concentration of 0.4 mM. The bacteria culture was harvested by centrifugation, resuspended in 20 ml/L lysis buffer, and sonicated the cell 6 cycles at 30 % energy. After centrifuging and lysing cell, it formed inclusion body. The inclusion body was dissolved via 20 mL solubilization
34
buffer and sonication, then diluting guanidine hydrochloride by 100 mM Tris-base (pH 8.0). Dialysis was with 10 mM KPi solution overnight. The dialysis product was centrifuged and collected the supernatant. The DEAE column was equilibrated with 10 mM KPi buffer and the protein was recovered in the flow throughout. The examination of expression yield and protein profile purification was performed by 15 % SDS–polyacrylamide gel (SDS–PAGE).
3.4 Reconstitution of apo-myoglobin with ZnPP and ZnPE
13.4.1
Reconstitution of apo-myoglobin with ZnPP
The method of reconstitution apo-myoglobin with ZnPP was followed Hoffman protocol48. Briefly, the protein solution was adjusted pH value to pH 12 and reacted for 15 min, then added the same volume of 10 mM KPi and adjusts to pH 7.0 and reacted for 15 min. A 1.5-fold molar excess of the zinc-porphyrins was dissolved in a warm CH3OH/DMSO (3:1 v/v) solution. After being cooled to 4 °C, the porphyrin
solution was added dropwise to the apo-Mb solution with stirring at 4 °C, and the protein solution was kept in the dark at 4 °C for 2 h. A second 1.5-fold molar excess of porphyrin was then added in the same fashion, and the solution was kept at 4 °C for another 2 h. The solution was transferred to a dialysis bag against phosphate buffer (10 mM KPi buffer pH 6.8) overnight. The protein solution was filtered with 0.45 μm cellulose membrane before applied to a Sephadex G-25 column. The column was equilibrated with 100 mM KPi buffer pH6.8, and eluted with the same buffer at 4 °C. The protein was collected and stored at -20 °C for later experiments.
35
3.4.2
Reconstitution of apo-myoglobin with ZnPE
1Reconstitution of ZnPE1 into apo-Mb followed the modified method of Axup et al49.
The apoprotein solution was maintained at ice bath temperatures for the duration of the reconstitution process. A 1.5-fold molar excess of ZnPE1 in dimethyl sulfoxide
(DMSO) (3:1 v/v) solution was added dropwise to a 10 mM KPi/DMSO (4:1 v/v) buffer containing apo-mypglobin at pH 12. After incubation for 15 min, the solution was adjusted to pH 7.0 and allowed to stir for 6 hr at 4 °C. The solution was transferred to a dialysis bag against phosphate buffer (10 mM KPi buffer pH 6.8) overnight. The protein solution was filtered with 0.45 μm cellulose membrane before applied to a Sephadex G-25 column. The column was equilibrated with 100 mM KPi buffer pH 6.8, and eluted with the same buffer at 4 °C. The protein was collected and stored at -20 °C for later experiments.
3.5 Circular dichroism (CD) spectroscopolarimetry
Circular dichroism (CD) spectroscopolarimetry was recorded and analyzed by Jasco J-815 spectropolarimeter. All measurements were performed in thermostated quartz cuvette of 1 mm path length. CD spectroscopy is a form of light absorption spectroscopy that measures the difference in absorbance of right- and left-circularly polarized light by a substance. The wavelength of measurement is from 500 nm to 190 nm, and the scanning speed is 100 nm/min at room temperature.
3.6 Picosecond fluorescence decays
36
system (TCSPC, FluoTime 200, PicoQuant). The excitation source came from either a femtosecond laser system (Verdi-10 and Mira 900D, Coherent) at 435 nm (doubled from an 860 nm beam out, PicoQuant); the instrument response is ~30 ps using the fs laser system while it increases to ~80 ps using the ps laser system. The temperature of the fluorescence decay experiments was kept at 10°C.
3.7 Cyclic voltammetry analysis
In electrochemical measurement, experiments employed a three-electrode cell with Ag/AgCl reference electrode, a platinum wire auxiliary electrode and a glassy carbon electrode (GCE) as working electrode. Before each experiment, the glassy carbon electrode was pulished with alumina oxide powder and rinsed with distilled water as a pretreatment. 10 mM DDAB suspensions were dissolved in water and ultrasonicated it for 30 min. 10 mM DDAB containing 0.25 mM protein was dropped on the surface of glassy carbon electrode (GCE) to make films. Equal volume of protein and DDAB were spread on a GCE and dried overnight. Cyclic voltammetry at DDAB-protein film was performed in 50 mM acetate buffer pH 7. Prior to measurements, buffer solution was purged with nitrogen gas at least 15 min, and then stopped apply nitrogen gas during the experiment.
3.8 Photoirradiation experiment
The reduction efficiency of reconstitution of apo-myoglobin with ZnPP and ZnPE1
were recorded with photochemical reactor PR-2000 (Panchum). 10 μM ZnPP-Mb (or ZnPE1-Mb), 2.5 mM NADP+, and 1 M TEA in 100 mM KPi pH 9.0 was co-incubated
37
38
4. Results and discussion
4.1 Purification of apo-myoglobin mutants
To produce highly pure and large amount of myoglobin variants, we follow the procedure which produces inclusion bodies proposed by Ribeiro et al.. When the inclusion bodies were isolated, the miscellaneous proteins can be removed easily.
First, E. coli was induced by IPTG to produce inclusion bodies of our myoglobin variants. When the cells were lysed, many inclusion bodies were in the crude extracts. The inclusion bodies were isolated via centrifugation. They were expected to be in the pellet because of their high density, and the miscellaneous proteins in the supernatant were able to be efficiently removed. The collection of inclusion bodies was re-dissolved by guanidine hydrochloride (8 M Gdm-HCl). Upon removal of the Gdm-HCl by dialysis, myoglobins would slowly refold to avoid subsequent aggregation. Last, myoglobins were purified by DEAE column, and analyzed by 15% SDS-PAGE to confirm the purity of the extracted proteins.
This collected protein contained few heme in the cavity of myoglobin and was with absorbance at 410 nm. Therefore, we used methyl-ethyl ketone (MEK) substitution method47 to extract heme from hemeprotein. MEK is a little miscible with water, enable heme and protein to be rapid and completely separated to organic layer and liquid layer. Figure 4-1 showed a 15% SDS-PAGE of the wild-type apo-myoglobin and its mutants for the characterization of molecular weight and purity. The results showed that these myoglobin variants were pure and expressed in large quantities.
39
Figure 4-1 The SDS-PAGE results of Mbs purification. Lane 1: marker; lane 2: MbWT; lane 3: MbH64D; lane 4: MbV68L ;lane 5: MbI107M ; lane 6: MbH64D/V68L ; lane 7: MbH64D/V68L/I107M.
4.2 UV-Vis analysis results of reconstituted metallo-porphyrin-Mbs
In order to confirm reconstitution of apo-myoglobin with porphyrin success or not, UV-Visible spectra was used to check the peak shift. The absorbance spectra of reconstituted metallo-porphyrin-Mbs have two bands: Soret (B) band in higher energy region and Q band in lower energy region, traditionally identified as the S0→S2
(n→π*) and S0→S1 (π→π*) transitions, respectively. Reconstituting myoglobin
variants (MbWT, MbH64D, MbV68L, MbI107M, MbH64D/V68L, MbH64D/V68L/I107M) with ZnPP and ZnPE1 shows the absorbance of Soret band at 426 nm and 443 nm and Q band at
554 nm, 597 nm, 568 nm and 616 nm, respectively (Fig.4-2).
On the other hand, we also compared the absorption of ZnPP and ZnPE1 in organic
solvent (THF) and phosphate buffer (100 mM KPi pH6.8), and the UV-Vis spectra results were shown in Fig. 4-3. Metallo-porphyrins almost could not dissolve in
1 2 3 4 5 6 7 65 kDa 14.4 kDa 30 kDa 20.1 kDa 45 kDa 91 kDa
40
phosphate buffer result in aggregation, so the bands in UV-Vis spectra are broad and shift. Soret band and Q band of ZnPE1 dissolved in phosphate buffer were not obvious
by comparing with ZnPP in phosphate buffer. A conjecture was that the structure of ZnPP has two carboxylic groups (COOH) and of ZnPE1 has three phenyl groups and
only one carboxylic group (COOH). It results in ZnPP is more hydrophilic than ZnPE1 and slight dissolving in phosphate buffer.
In addition, the Soret band of ZnPP and ZnPE1 in KPi buffer was broader than
ZnPP-Mb and ZnPE1-Mb in KPi buffer, and it was shifted to longer wavelength
(red-shift). According to Periasamy et al.20, the aggregation forms contain two types: J-aggregates and H-aggregates. J-aggregate cause red-shifts and H-aggregate cause blue shifts compared to ZnPP-Mb and ZnPE1-Mb in KPi buffer. Namely, the
aggregation types of ZnPP and ZnPE1 were J-aggregates (head-to-tail arrangement).
It’s also observed that reconstitution of metallo-porphyrin-Mbs can increase the solubility in KPi buffer.
41
(A)
(B)
Figure 4-2 UV-Vis spectra results of metallo-porphyrins (A) ZnPP (B) ZnPE1
reconstituted with MbWT, MbH64D, MbV68L, MbI107M, MbH64D/V68L, and MbH64D/V68L/I107M in 100 mM KPi buffer.
42
(A)
(B)
Figure 4-3 UV-Vis spectra results of (A) ZnPP in THF, 100 mM KPi buffer and reconstituted with apo-Mb in 100 mM KPi buffer (B) ZnPE1 in THF, 100 mM KPi
43
4.3 Circular dichroism (CD) spectropolarimetry
Circular dichroism (CD) spectropolarimetry can determine the secondary structure of protein in the far-UV region (190-250 nm). In order to analyze the secondary structure of metallo-porphyrin-Mb after reconstitution, we use circular dichroism (CD) to insure the protein keeping in natural form. According to the reported CD spectrum of a protein, the conformation characterization of α-helix occurs in the spectral region is 200-250 nm. Figure 4-4 showed the CD spectrum of ZnPP-Mb and ZnPE1-Mb, and
44
(A)
(B)
Figure 4-4 The CD spectrums of metallo-porphyrins (A) ZnPP (B) ZnPE1
reconstituted with MbWT, MbH64D, MbV68L, MbI107M, MbH64D/V68L, and MbH64D/V68L/I107M.
45
4.4 Fluorescence analysis
The fluorescence emissions of reconstituted metalo-porphyrin-Mbs are shown in Fig. 4-5 to 4-7. The emission spectrum of ZnPP and ZnPE1 in THF showed the
maxima at 588 nm and 621 nm, respectively. The emission spectral is the mirror image of Q band. When reconstituted ZnPP-Mb and ZnPE1-Mb in KPi buffer, the
emission spectrum of them showed the maxima at 600 nm and 630 nm, respectively. Compared these results showed that increasing the polarity of environment leads to red shift of absorption peak in porphyrins.
(A) (B)
Figure 4-5 UV-Vis spectra and fluorescence emission of (A) ZnPP (B) ZnPE1
46 (A) (B) (C) (D) (E) (F)
Figure 4-6 UV-Vis spectra and fluorescence emission of (A) ZnPP-MbWT (B) ZnPP-MbH64D (C) ZnPP-MbV68L (D) ZnPP-MbI107M (E) ZnPP-MbH64D/V68L (F) ZnPP-MbH64D/V68L/I107M.
47 (A) (B) (C) (D) (E) (F)
Figure 4-7 UV-Vis spectra and fluorescence emission of (A) ZnPE1-MbWT (B)
ZnPE1-MbH64D (C) ZnPE1-MbV68L (D) ZnPE1-MbI107M (E) ZnPE1-MbH64D/V68L (F)
48
4.5 Picosecond fluorescence decays analysis
The fluorescence intensity of reconstituted metallo-porphyrin-Mb was related with protein concentration. High concentration of chromophores can increase fluorescence intensity; on the contrary, low concentration causes weak intensity. However, when the concentration of chromophores is too high, the intensity decreases and results from self-quenching. It is contributed that charge transfer and exciplex formation50, 51. The former is that when chromophores are too close, result in energy transfer and decrease fluorescence intensity; the latter is chromophores intermolecular collide to form complex and self-quenching. To prevent from high concentration effect, we test different concentration of reconstituted metallo-porphyrin-Mb in fluorescence analysis to find the suitable concentration to do picosecond fluorescence decays. Fig. 4-8 to 4-9 showed the fluorescence emission in different concentrations, and the optimum protein concentration for picosecond time-resolved fluorescence measurement are 10 μM in ZnPP-Mb and 5 μM in ZnPE1-Mb.
49
(A) (B)
(C) (D)
(E) (F)
Figure 4-8 The correlation of protein concentration and fluorescence intensity in 100 mM KPi buffer. (A) ZnPP-MbWT (B) ZnPP-MbH64D (C) ZnPP-MbV68L (D) ZnPP-MbI107M (E) ZnPP-MbH64D/V68L (F) ZnPP-MbH64D/V68L/I107M. Obtain fluorescence intensity at λem = 600 nm (ZnPP-Mb) with excitation λex = 435 nm.
50
(A) (B)
(C) (D)
(E) (F)
Figure 4-9 The correlation of protein concentration and fluorescence intensity in 100 mM KPi buffer. (A) ZnPE1-MbWT (B) ZnPE1-MbH64D (C) ZnPE1-MbV68L (D)
ZnPE1-MbI107M (E) ZnPE1-MbH64D/V68L (F) ZnPE1-MbH64D/V68L/I107M. Obtain