5-Fluorouracil之新抗藥性機制與r-胸腺核苷合成酶在乳癌表現之關聯性; Correlation between a novel resistance to 5-Fluorouracil and expressions of rTS in breast cancer
67
0
0
全文
(2) 目. 錄 頁次. 目錄. 2. 中文摘要. 3. 英文摘要. 4. 誌謝. 6. 圖目錄. 7. 第一章、前言. 8. 第二章、文獻探討. 13. 第三章、材料與方法. 26. 第四章、結果. 43. 第五章、討論. 48. 第六章、結論. 52. 參考文獻. 53. 附錄. 59. 碩士論文電子檔案上網授權書. 67. 2.
(3) 摘要 5-Fluorouracil(5-FU)是一種應用廣泛的抗腫瘤藥物,近年來 隨著抗藥性的產生,限制了其在臨床上的用途及發展。因此,本研究主 要探討 r-胸腺核? 合成酵素(Antisense Thymidylate Synthase; rTS) 在乳癌的表現與 5-FU 抗藥性間的關聯性。我們利用五株不同的乳癌細 胞株,分別為 MCF-1、MCF-7、BT-20、60055 及 T47D 來進行研究。首先, 利用西方墨點法及免疫細胞染色法來測定 rTS 的表現量,再用細胞毒性 分析法來測定 5-FU 對五株乳癌細胞株的毒殺作用,以不同的濃度:1.6 μM 至 50 mM,將五株細胞株放置 72 小時後、計算 5-FU 對五株細胞株 的 IC50 (50% growth inhibitory concentration),並製作劑量-反應曲 線圖。由實驗結果得之,五株乳癌細胞株除了 T47D 外,其餘的細胞株 都有 rTS 的表現,且有 rTS 表現的細胞株其 5-FU 之 IC50 的濃度比 T47D 明顯 高了 3 至 195 倍。換句話說,有 rTS 表現量的細胞株,其會明顯 降低 5-FU 的細胞毒殺作用,產生 5-FU 的抗藥性,然而其確立的機轉尚 未清楚。回溯有關 5-FU 抗藥性的研究,大多數是以 TS 與大腸癌細胞株 為主,本研究希望藉由乳癌細胞株來建立 rTS 與 5-FU 抗藥性的關聯性, 以改善 5-FU 在乳癌之輔助性療法的效果,並增加 5-FU 在臨床上的地 位。藉由 rTS 的表現量,可做為 5-FU 在臨床上的治療指標。. 3.
(4) Abstract 5-Fluorouracil (5-FU) is a commonly used therapeutic drug in breast cancer. In this study, we investigated the correlation between a novel resistance to 5 -FU and expressions of rTS in breast cancer. We used five breast cancer cell lines ( MCF-1, MCF-7, BT-20, 60055, T47D) in this study. First, we measured rTS expression levels by western blotting and immunocytochemistry . Next, we investigated sensitivity of the cells to 5-FU by cytotoxic assay. The cells were exposed to 5 - FU at concentrations from 1.6μM to 50 mM for 72 hr. Dose-response curves from these five breast cancer cell lines were plotted and IC50 was measured. Our results show that four breast cell lines overexpressed rTS (MCF-1, MCF-7, 60055, B-20 ), but T47D. The IC50 of 5-FU for T47D was 0.013 μM, which was 3 to 195 times lower than that of other four cell lines (MCF-1, MCF-7, 60055, BT-20), showing significantly more sensitive to 5-FU. Therefore, expression of rTS could result in reduced 5-FU cytotoxic effect and increase resistance to 5 -FU. In conclusion, Our data demonstrated a relationship between rTS expression and resistance to 5-FU in breast cancer. Although the mechanism of. 4.
(5) resistance is not yet clear, Our results provide a forcus for future studies to elucidate the mechanism by which drug resistance in breast cancer could be mediated by rTS expression.. 5.
(6) 誌謝 在這鳳凰花開的畢業時節,已睽違學校十年的我,竟然還有機會 重溫學生舊夢,是機緣也是福份。在這兩年的研究過程中,首先要感謝 本院藥劑部的大家長蔡輝彥主任,也是我求學過程中的恩師,不僅提供 我們藥師進修的管道,還幫我爭取留職停薪的機會,讓我在這兩年中可 以專心學習學校的課業及研究,並實現我跨進醫學研究的大門,真的非 常感謝蔡主任的提攜支持。再者,最是要感謝國立中興大學生醫所的周 寬基教授的指導。是自己運氣好,也是與周博士有緣,當我還在化療室 時,一次偶然的機緣,竟然能幸運的窺探分子生物的堂奧,由於周博士 深入淺出的提綱契領,讓我對基礎醫學產生了濃厚的興趣,不僅教導我 在實驗上的技術與方法,也提供我很多寶貴的建議與想法。最重要的是教 導我如何思考事情、解決問題及簡報技巧。由於這兩年的訓練,讓我受. 益一輩子,除了感謝,還是感謝! 此 外,我要特別感謝淑芬、智尤、俊豪、冠樺、加麗及永生的技術指 導,也要感謝華浩、佩瑜在課業上的幫忙。另外,還要感謝國立中興大學 6.
(7) 獸微所邱繡河教授及本校醫研所的陳玉芳老師的指導。 最後,要感謝我最親愛的家人,尤其是我先生成家的支持與鼓勵,讓 我在這兩年的研究生活中,無後顧之憂。在此將本論文獻給我的家人及所 有曾經指導及幫助過我的人。日後將秉承師長的教導,不負眾人的期望, 並為癌症病患盡一份藥師應盡的職責與服務。. 圖目錄 頁次 圖一. 5-FU 的代謝途徑. 59. 圖二. 5-FU 抑制 TS 的作用機轉. 59. 圖三. Leucovorin 和葉酸的代謝環. 60. 圖四. Leucovorin 的作用機轉. 60. 圖五. rTSβ 乳癌細胞株的表現. 61. 圖六. TS 在乳癌細胞株的表現. 61. 圖七. 不同 LV 的濃度在乳癌細胞株的生長抑制作用. 62. 圖八. 5-FU 在 60055 細胞株之 IC50 的反應曲線. 62. 圖九. 5-FU 在 BT-20 細胞株之 IC50 的反應曲線. 63. 圖十. 5-FU 在 MCF-1 細胞株之 IC50 的反應曲線. 63. 圖十一 5-FU 在 MCF-7 細胞株之 IC50 的反應曲線. 64. 圖十二 5-FU 在 T47D 細胞株之 IC50 的反應曲線. 64. 圖十三乳癌細胞株在 5-FU 濃度 5 mM 時之細胞存活率 7.
(8) 的比較. 65. 圖十四乳癌細胞株的細胞免疫染色圖. 66. 第一章、 前言 第一節、研究緣起 惡性腫瘤自民國七十一年起已躍居台灣十大死因的榜首,這二十幾 年來一直穩居首位。根據衛生署統計台灣地區十大女性癌症之發生率 中,乳癌僅次於子宮頸癌躍居第二位。由九十二年的資料顯示,乳癌排 名在台灣婦女因癌症死亡原因的第四位(1)。更值得注意的是,乳癌在 這十年來的死亡率正逐年增加中,且有年輕化的趨勢。 一般台灣婦女 乳癌的好發年齡為四十至五十歲之間,較歐美國家的好發年齡約提早十 歲,但任何成年婦女皆有機會罹患此症。因此,乳癌實為女性最重要的 癌症之一。就全球而言,乳癌在歐美佔婦女癌症的第一位,近年來亦有 上升的趨勢(2)。 乳癌在臨床上的治療方式,主要是根據乳癌的分期,一般仍以手術 治療為主,放射治療、化學治療及賀爾蒙治療為輔。許多病人經過根除 性乳房切除術後,並不代表已完全根治痊癒,因此須靠術後的輔助性治. 8.
(9) 療( Adjuvant therapy )來預防乳癌復發。目前乳癌的輔助性化學治療 是 以 5-Fluorouracil (5-FU) 為 主 的 合 併 療 法 ( combination therapy)(2)。5-FU 是一種應用廣泛的抗腫瘤藥物,其為 pyrimidine 的相似物,本身為前? 藥(prodrug)不具有抗癌活性,必須要經由代謝 轉 變 為 5-fluorouracil -2'- deoxyuridine -5'- monophosphate (FdUMP) 的活性物,才能與細胞內的 deoxyuridine monophosphate (dUMP)去競爭胸腺核? 合成? (Thymidylate synthase;TS),藉由抑制 胸腺核? 合成? (Thymidylate synthase)來阻止 DNA 的合成及修復。 因 此 , 5-FU 會 與 Thymidylate synthase(TS)及 5, 10-methylene tetrahydrofolate 形成一個穩定的三物複合體(ternary complex),來 阻斷 dUMP 轉變為 deoxythymidine monophosphate (dTMP)的機轉(如圖 一) (3,4) Thymidylate synthase (TS)在自然的酵素反應中是將 dUMP 甲基化 成為 dTMP 的催化酵素,一旦受到抑制,細胞內缺乏 dTMP,就會影響到 DNA 的修復與複製。雖然 dTMP 的製造有回收路線(salvage pathway)的 存在,但在大多數的哺乳動物細胞內,由 dUMP 轉化為 dTMP 的自然製造 路線仍然是最重要的。因此,有學者認為 Thymidylate synthase (TS) 可做為抗癌藥物的治療指標,因為它在 DNA 的合成反應上佔有非常重要 的地位 (5,6)。也因為如此,近年來在臨床上我們對於已接受輔助性化. 9.
(10) 學治療的乳癌患者採用以 5-FU 為主的合併療法,發現所使用的劑量日 益增加或在臨床反應上已不盡理想,這是否意謂著已有抗藥性產生?而 TS 與 5-FU 抗藥性之間是否有關聯?其產生的機轉為何?目前尚未確立 完全。 根據早期有關 TS 與 5-FU 抗藥性研究,大多數是以大腸癌為主。因 為 5-FU 早在西元 1957 年由 Heidelberger 等學者研發至今,已廣泛且 有效的治療大腸癌超過 40 年的歲月(7)。5-Fu 除了成功應用在治療大腸 癌外、其亦可治療乳癌、頭頸癌、胰臟癌、甚至胃癌等固態瘤,其抗癌 效果顯著。但近年來由於許多臨床試驗顯示,5-FU 隨著抗藥性的產生, 限制了其在臨床上的用途及發展。以治療轉移性大腸癌為例,5-FU 在化 學治療上劑量的使用,隨著時間的投予、有逐漸增加的趨勢,且在臨床 的反應率(Response rate)都不盡理想,僅達到約 30% (8,9)。於是探 討 5-FU 抗藥性的機轉就成為重要的課題。根據 Robet 等學者回顧有關 5-FU 抗藥性的探討,其提出相關的機轉包括 5-FU 的 Target enzyme、 如 : Thymidylate synthase (TS) 的 過 度 表 現 或 改 變 Thymidylate synthase 與 FdUMP 的親和力、以及降低 5-FU 活性代謝物的產生等因素 (8,10-17) 。 最 近 有 學 者 專 門 針 對 Thymidylate synthase (TS) 與 antisense Thymidylate synthase (rTS)和 5-FU 抗藥性的關聯性做研 究(18)。. 10.
(11) rTS 基因和 TS 基因同位於 chromosome 18 的位置上,含有 17 個 exons, 可經由 alternative splicing 作用轉錄成 rTSα及 rTSβ兩段產物,其 中 rTSα分子量為 41 kDa 而 rTSβ為 47 kDa。rTSαRNA 的 3’端恰巧 與 TS RNA 的 3’端互補。所以,廣義上, rTSα是 TS 基因的 antisense RNA (19,20,21)。早期,有些學者發現具有抗藥性的細胞株,其 TS 基 因表現下降,同時伴有 rTS 基因表現上升的現象 (22,23,24,25)。從 原核生物中發現的 naturally occurring antisense RNAs,本來就會抑 制基因表現。而 rTS 顯然的在各種不同的階段也扮演著調節 TS 基因的 功能,包括:轉錄減弱(transcriptional attenuation)、影響 RNA processing、造成 RNA 不穩定、以及抑制轉譯等(19,24,26)。在真核生 物中,雖然有漸漸多的 naturally occurring antisense RNAs 被發現, 但是在哺乳動物細胞中,這些 naturally antisense RNAs 調控相對 sense RNA 表現的功能還沒被建立(19,26),或者 rTS 與 TS 的相互消 長就是一個很好的例子。 根據 Dr. Dolnick 所做的研究顯示,對 5-FU 具有抗藥性的大腸癌 細胞株 H630-1 與大腸癌細胞株 H630 之 rTS 基因與 TS 基因表現量的比 較,發現 H630-1 細胞株的 TS 基因有 DNA amplification,因此 TS 的表 現量高過 H630 細胞的 6 至 10 倍。由於 rTS 基因也包括在整個 amplicon 中,所以在 H630-1 細胞中 rTS 的表現量也高過 H630 細胞的 40 至 70 倍. 11.
(12) (22,23) 。依據 Dr.Dolnick 的研究發現,顯示 rTSβ可負向調控 TS 基因 的表現,而 TS 基因表現在快速生長細胞中則扮演著相當重要的角色。 於是開始探討在一個快速生長的癌細胞中,TS 與 rTSβ基因的變化關 係,進而去研究 rTSβ基因的表現量,對大腸癌病人所使用的 5-FU 在臨 床的反應(19),其結論可能與 rTSβ的表現較能代表 5-FU 的抗藥性。. 第二節、研究目的 因此在本研究中,我們希望藉由乳癌細胞株來探討 rTSβ基因在乳癌 的表現與 5-FU 抗藥性之間的關聯,利用西方墨點法及免疫細胞染色法 來測定 rTSβ的表現量,再用細胞毒性分析法來測定 5-FU 對乳癌細胞株 的毒殺作用,進而證明 TS 與 rTSβ基因在乳癌細胞株的變化關係。. 12.
(13) 第二章、文獻探討 第一節、 乳癌的簡介 一、 何謂乳癌 乳癌是由乳房乳腺管細胞或是腺泡細胞經由不正常分裂、繁殖 所形成之惡性腫瘤。這些惡性腫瘤除了侵犯乳房之局部器官,更可能 轉移到遠處器官如骨骼、肺、肝、腦等,而破壞身體重要器官的功能, 造成身體健康之損害,甚至危害生命。. 二、 乳癌在台灣的現況 根據行政院衛生署的統計,乳癌的發生率與死亡率正逐年增加 中。由民國九十二年癌症登記報告顯示,乳癌的發生率為女性好發 癌症的第二位,每年約新增三千一百多位乳癌患者。死亡率自民國 八十五年來更超越子宮頸癌,躍居台灣地區女性主要癌症死亡排名 的第四位,在這十年來、乳癌的死亡率增加一倍以上且有年輕化的. 13.
(14) 趨勢(1)。 台灣乳癌好發年齡在四十至五十歲之間,較歐美國家的 好發年齡約提早十歲,但任何成年婦女皆有機會罹患此症。近年來 由於醫學進步,經由適當的治療,乳癌的十年存活率平均達百分之 六十,第一期乳癌的存活率 則高達百分之八十,零期乳癌甚至接近 百分之百(2) 。 三、乳癌發生的機轉 乳癌主要是由乳腺產生,其發生的機轉最主要可分成三階段,即 為啟動期(initiation phase)、促進期(promotion phase)和進展 期(progression phase)(27)。正常細胞經過環境中放射線或化學 物質的影響,若產生基因突變,就可能會發展成癌細胞,但還不會形 成腫瘤。癌細胞在雌性素的幫助下生長比正常細胞好,就會逐漸發展 成癌症。癌症的形成往往由數種致癌基因(oncogenes)的活化或數種 抑癌基因(tumor suppressor genes)的功能喪失所累積而成的 (28)。其中與乳癌相關的致癌基因包括 erb-B、c-myc 等,而抑癌基 因則有 p53、BRCA1 及 BRCA2 (27)。乳癌的形成過程通常需要經過「多 步驟性致癌機轉」(multistep carcinogenesis),一旦細胞的基因突 變時,便會啟動本身所具有的修補機制,使細胞週期停滯,並活化許 多修補性的酵素,試圖將受損的基因體修復,以回復至細胞原本的正 常生理狀態;然而,細胞受到的傷害超過修補機制所能修補的範圍. 14.
(15) 時,細胞便會啟動計劃性凋亡(programmed cell death , apoptosis) 的機制,使細胞進入死亡。此修補機制及細胞計劃性凋亡機制能有效 地維持個體的細胞數及品質,使個體能維持正常的生理功能。此外致 癌因子亦可能會導致細胞內修補機制產生缺失而失去修補基因體的 作用,使得受損的基因體無法修復,而使癌細胞數量增加(27,28)。 就乳癌而言,遺傳的影響大約是 10%,其他大多數為環境與雌性素的 影響(2)。. 四、乳癌的診斷 乳癌最重要的臨床表徵是可觸摸到的乳房腫塊。雖然大部份 的乳房腫塊均是良性,乳癌僅為一小部份,但摸到任何乳房腫塊均 仍需要分辨良性與惡性。除了自我檢查及醫師的理學檢查外,必要 時也須安排進一步檢查,包括乳房超音波、乳房X光攝影及細針抽 吸細胞學檢查。然而病理組織切片可以藉由手術切片或以粗針穿刺 取檢體做組織病理學診斷,這是乳癌診斷的黃金標準(2)。組織切片 除了確定病理診斷,另一個重要目的為檢測乳癌預後因子,如荷爾 蒙接受體或與乳癌相關的蛋白表現,以作為治療依據(29,30)。. 五、 乳癌的分期. 15.
(16) 乳癌分期的主要目的在確立治療方式的選擇,目前乳癌的分期 是根據 TNM 系統依照腫瘤大小( tumor size;T),腋下淋巴腺轉移 與否( node;N),遠處是否轉移(metastases;M)等來分期( 2) 。 乳 癌的零期,即原位癌、為最早期的乳癌,癌細胞仍在乳腺管基底層 內。第一期的乳癌為腫瘤小於兩公分以下的浸潤癌且腋下淋巴結無 癌轉移。第二期指腫瘤在兩公分至五公分之間的浸潤癌;或腫瘤小 於兩公分,但腋下淋巴結有癌轉移。到了第三期已成為局部廣泛性 的乳癌,腫瘤已大於五公分的浸潤癌且腋下淋巴結有任何癌轉移或 有胸壁皮膚的浸潤乳癌。至於第四期則為轉移性乳癌,因為已有遠 處器官轉移,最常見的是轉移至骨骼、肺、肝、腦等(2)。. 六、乳癌的治療 一般而言,仍以外科手術治療為主,放射線治療、化學治療及 荷爾蒙治療為輔。但針對第二期以後的乳癌,通常還是以合併化學 治療為主軸,然而在臨床上,癌細胞的抗藥性仍然是不可避免的障 礙(2)。. 第二節、 抗藥性機轉 16.
(17) 癌細胞隨著抗藥性產生,會降低抗癌藥在臨床上的效果,一般 的抗藥機轉可分為生理性(physiology)及遺傳性 (genetic) 兩 種。所謂的生理性抗藥多半是指藥物不能到達作用的腔室或血液供 給不足而造成藥物無法充分作用,甚至藥物會受癌細胞周期所調 控。至於遺傳性抗藥機轉、近年來隨著基因工程及分子生物學的突 飛猛進,也帶動一股研究風潮,目前的研究大多數是以多重抗藥性 基因(multi- drug resistance gene;MDR)為主,其主要的作用 會增加藥物的流出及降低藥物的吸收。其餘還包括增加或改變 target 基因產物、或增強 DNA 的修復以造成抗藥性產生( 31)。反 觀 5-FU 的抗藥機轉是不屬於上述的作用路徑(32)。. 一、5-FU 的抗藥機轉: 5-FU 是一種應用廣泛的抗腫瘤藥物,其為 pyrimidine 的相似 物,本身為前? 藥(prodrug)不具有抗癌活性,必須要經由代謝轉 變為 FdUMP 的活性物,才能與細胞內的 dUMP 去競爭胸腺核? 合成 ?. (TS),藉由 TS 來阻止 DNA 的合成及修復 (3,33)。然而當 5-FU. 的抗藥性產生時,容易造成合併化學療法的失敗,於是探討抗藥性 機轉就成為重要的課題。目前與 5-FU 抗藥性有關的因素包括:5-FU 的 target enzyme 的改變如 TS 的過度表現或改變與 FdUMP 的親和. 17.
(18) 力(7,8,34)、 或降低 5-FU 活性代謝物或增加不活性代謝物的產 生以及有關 5-FU 藥物動力學的抗藥性(7.8,32,35)。. 二、5-FU 的藥物動力學: 5-FU 可經由口服或靜脈注射給藥,其口服吸收率為 28 至 100%, 主要是決定於肝臟的 dihydropyrimidine dehydrogenase (DPD) 能 代謝多少比率的 DHFU 代謝物,除此之外、其主要是經由肝臟代謝 成 FdUMP 的活性物才具有抗癌作用。可分佈於腦脊髓液(CSF)、組 織外液(ECF)、肋膜滲出液及腹水中,最後可經由呼出 CO2 排出體 外,在血漿半衰期為 8 至 13 分鐘。一般的使用劑量是根據不同的 癌症所使用的合併處方而不同,最常見的毒性除了噁心、嘔吐、腹 瀉外、還會造成骨髓抑制、禿頭及皮膚病變如:hand-foot syndrome (36)。. 三、Leucovorin 增強 5-FU 的細胞毒殺作用: Leucovorin 是 glutamic acid 的衍生物,亦稱為 5-formyl -tetrahydrofolate 或亞葉酸,屬弱酸性、親水性高。其含有蝶啶 環(pteridine ring)的構造,因此在碳 6 的位置上能以 l 型或 d. 18.
(19) 型兩種立體形式等比例存在(如圖三)(37,38)。Leucovorin 在細胞 內會被轉變為 5,10- methylenete - trahydrofolate (mTHF)的還 原葉酸輔助因子,因此有能力即時補充細胞內不足的還原葉酸,可 作為? 氫葉酸還原酵素抑制劑(dihydrofolate reductase,DHFR) 如 methotrexate (MTX)抗癌藥的拮抗劑,目的是用來防止葉酸拮 抗劑所造成的毒性(37,38,39)。 5-FU 的活性代謝物 FdUMP 會與細胞內的 dUMP 競爭胸腺核? 合 成酵素(TS),與 mTHF 及 TS 形成一個穩定的共價鍵之三物複合體 (ternary complex)(如圖二)(3)。另外 mTHF 會加速複合體的結合 和分解,以及 dTMP 和? 氫葉酸(dihydrofolate)的形成,因而控制 調節 DNA 的聚合作用。? 氫葉酸也經? 氫葉酸還原酵素抑制劑(DHFR) 和麩氨酸梭甲基轉換酵素(serine hydroxymethyl transferase)再 生為 mTHF 輔助因子。所以在有充分的 mTHF 存在時,TS 的活性就會 被 FdUMP 抑制,而不能產生 dTMP。只有 FdUMP 而沒有 mTHF 存在時, 有些癌細胞就會有抗藥性產生,因為沒有足夠的 mTHF 來產生三物 複合體。因此在這種情況下,可藉由與 Leucovorin 合併使用或增 加 Leucovorin 的濃度來增強 5-FU 的細胞毒殺作用,並扭轉 5-FU 的抗藥性(如圖四) ( 3,37,38,39)。. 19.
(20) 第三節、Thymidylate synthase (TS)、Antisense Thymidylate synthase (rTS)與抗藥性的關係 一、Thymidylate synthase (TS) 的作用與抗藥性關係 TS 的分子量為 36 kDa,在自然的酵素反應中是將 dUMP 甲基化 成為 dTMP 的催化酵素,一旦受到抑制,細胞內缺乏 dTMP,就會影 響到 DNA 的修復與複製(33)。然而 5-FU 的作用就是去抑制 TS 而阻 止 DNA 的合成及修復(3,33),同時也可在細胞內轉化成 FUTP (fluorodeoxyuridine triphosphate ) 而 incorporate 到 RNA 中, 影響 RNA 的功能(19,20)。關於 TS 與 5-FU 的抗藥性研究,有學者 指出 5-FU 的抗藥性與 TS 的作用改變有關,包括 TS 的過度表現或 是減少葉酸的多麩氨酸鹽化(polyglutamylation)而造成 FdUMP 與 TS 的親和力下降,又加上缺乏 mTHF,而形成不穩定的二物複合體 (binary complex),因此 FdUMP 抑制 TS 的作用降低而可能導致 5-FU 的抗藥性產生(7,40)。 根據 Peters 等學者的研究顯示,在臨床上投予 5-FU 治療的病 人,經過 48 小時後其可抑制 50%的 TS,如果與 Leucovorin 合併使 用則可抑制高達 70%以上的 TS,且發現 5-FU 對 TS 過度表現的病人. 20.
(21) 其抑制效果較差,甚至沒有反應(33)。又有學者指出,TS 可做為 治療乳癌的預後指標,因為發現在乳癌細胞中,TS levels 與 S-phase fraction (SPF)即細胞的增生、DNA polymerase α及淋 巴的侵犯程度有明顯的關連性,因此他以 115 位乳癌病患進行實 驗,得知有 TS 高度表現的病人其存活率較低,且與乳癌惡化的程 度有關。如果使用 5-FU 的合併療法來治療乳癌,可能會影響 5-FU 的臨床效果(29,41-44)。最近更有學者以基因轉殖的技術將 TS 基 因轉殖到子宮頸細胞株內,以證實 TS 的高度表現的確會降低 5-FU 的敏感性。他們利用細胞毒性分析法來測定 5-FU 對子宮頸細胞株 的毒殺作用(42,45)。根據以上的結果也曾發表在 TS 高度表現的胃 癌細胞株(45, 46,47)、及大腸癌抗藥株上(45,48)。近年來有關 TS 與 5-FU 的抗藥性研究,已有學者專門針對 Thymidylate synthase (TS)與 antisense Thymidylate synthase (rTS)的關係著手,並奠 定今後有關 5-FU 抗藥性研究的新方向(19-25)。. 二、Thymidylate synthase (TS)與 antisense Thymidylate synthase (rTS)的關係 近年來 Dr. Dolnick 專注於探討 rTS 基因的功能及應 用,他指出 rTS 基因和 TS 基因同位於 chromosome 18 的位置. 21.
(22) 上,含有 17 個表現子(exons),可經由選擇剪接 (alternative splicing) 作用轉錄成 rTSα及 rTSβ兩段產物,其中 rTSα分 子量為 41 kDa、存在量很少,而主要的產物為 rTSβ其分子量 為 47 kDa (20,21)。rTSα和 rTSβ都含有一個可轉譯成 361 個氨基酸和 416 個氨基酸的開放讀序框(open reading frame ),這兩個蛋白質在 C 端 341 個氨基酸是完全相同的。 經過電腦比對,兩者同屬於 mandelate racemase 的 super -family,這類酵素可作用在許多不同的受質,主要功能是將 質子從 carboxylic acids 中的αcarbons 抽離(19,49)。 rTSα RNA 的 3’端恰巧與 TS RNA 的 3’端互補。所以, 廣義上, rTS 是 TS 基因的 antisense RNA。早期,他們發現 有一些抗藥性細胞株,其 TS 基因表現下降,同時伴有 rTS 基 因表現上升(19-21,24)。根據 Dr. Dolnick 的假說認為 rTSβ 的表現增加會降低 TS 的蛋白表現,在哺乳動物細胞中,這些 naturally antisense RNAs 調控相對 sense RNA 表現的功能 還沒被建立(19,26)。因此引發他們去研究 rTS 是否具有調節 TS 基因。他們首先證實,rTSα的存在會造成在 TS mRNA 特定 點的切割,而降低 TS mRNA 的表現量(19,22,26)。另外、為了 要證實 rTSβ的過度表現可能會調降 TS 基因並與 5-FU 的抗藥. 22.
(23) 性有關,於是利用了 H630 及 H630-1 這兩種細胞株進行實驗。 H630 是大腸癌細胞株,H630-1 則是由 H630 衍生出來、對 5-FU 具有抗藥性的細胞株。H630-1 細胞株的 TS 基因有 DNA amplify - cation,因此 TS 的表現量高過 H630 細胞的 6 至 10 倍。由 於 rTS 基因也包括在整個 amplicon 中,所以在 H630-1 細胞中 rTS 的表現量也高過 H630 細胞的 40 至 70 倍(22,23) 。將 H630 細胞以 固定濃度種在培養瓶中,之後每天分析細胞週期的變化,以及細胞 內 TS 與 rTSβ的表現量,會發現隨著細胞滿度(confluency)的增 加,處於 S 期的細胞比例漸減,TS 表現漸減,而 rTSβ的表現則越 來越多,這個現象符合了 TS 是一個 S 期的酵素。當他們以高濃度 細胞種在培養瓶中,24 小時後就去分析細胞週期的變化,及細胞內 TS 與 rTSβ的表現量,雖然處於 S 期的細胞比例很低,但 TS 與 rTSβ的基因表現卻沒有變(19)。 所以他們認為是細胞長期培養分泌了某些化學物質,造成細胞 內 TS 基因表現的變化。於是他們利用 trans-well,在細胞不 互相接觸的情況下,觀察 rTSβ有高表現量的 H630-1 細胞是否 分泌物質調控 H630 細胞內的 TS 表現量,結果顯示有 rTSβ過 度表現的細胞會分泌造成細胞內的 TS 基因表現下降的物質 (19,22,23,49,50)。. 23.
(24) 由於 H630 及 H630-1 這兩種細胞株在顯微鏡下的外型 morphology)相差許多,單就在培養液改變 methionine 的濃度 就可以改變 H630 的外型,讓 H630 看起來更像 H630-1,因此他 們覺得這個物質可能是 methionine 代謝物(19,50,51)。依據 Dr. Dolnick 的研究發現,顯示 rTSβ可負向調控 TS 基因的表 現,而 TS 基因表現在快速生長細胞中則扮演著相當重要的角 色。於是開始探討在一個快速生長的癌細胞中,TS 與 rTSβ基 因的變化關係。. 第四節、rTSβ在乳癌細胞的表現與 5-FU 抗藥性的關係 根據 Dr. Dolnick 先前所作的研究顯示,H630-1、對 5-FU 具有抗藥性的細胞株其 rTSβ的表現量高於 H630 細胞株達 40 至 70 倍之多,甚至在許多不同的抗藥細胞株如 K562B1A、 HCT-8/DF2 其 rTSβ的表現量與上述結果相似,都有過度表現 的現象(22,23,24)。於是推論,近年來關於 5-FU 的抗藥性探 討可能與 rTSβ的表現比較有關、至於與 TS 的關係、顯示 rTS β可負向調控 TS 基因的表現,而 TS 基因表現在快速生長細胞 中則扮演著相當重要的角色。截至目前為止,沒有文獻報導有 關 rTSβ在乳癌細胞的表現與 5-FU 抗藥性的關係。. 24.
(25) 第五節、研究動機 近年來、由於沒有文獻報導有關 rTSβ在乳癌細胞的表現 與 5-FU 抗藥性的關係,於是讓我們有動機去研究 rTSβ在乳癌細胞 的表現與 5-FU 抗藥性的關係,並以乳癌細胞株來進行實驗。根據 Dr. Dolnick 先前所作的研究結果,我們假設如果有 rTSβ表現的 乳癌細胞,可能會降低 5-FU 的細胞毒殺作用而造成 5-FU 的抗藥性 產生,並影響 5-FU 在臨床上的療效。. 25.
(26) 第三章、材料與方法 第一節 、實驗材料 一、 儀器、器材 1、. 無菌操作箱(laminar flow). 2、. 二氧化碳細胞培養箱(CO2 incubator). 3、. 細胞培養皿 (T25 及 T75 規格)(購自 FALCON). 4、. 細胞計數器(Haemocytometer)(購自 Boeco). 5、. 96 孔細胞培養盤(購自 FALCON). 6、. 離心機(centrifuge). 7、. 位相差顯微鏡(phase-contrast microscope). 8、. 酵素免疫分析儀 (ELISA reader) (購自 Anthos Labtec, Australia). 9、. 分 光 光 度 計 : Metertech , UV spectrophotometer. 26. / VIS-SP8001.
(27) 10、 分光光度計偵測管 11、 Trans-Blot SD Semi-Dry Transfer Cell ( 購 自 美 國 Bio-Rad 公司) 12、 Hybond-PVDF transfer membrance (Millipore ; lmmobilon-P transfer membrane) 13、 電泳裝置系統(購自美國 Bio-Rad 公司) 14、 電泳? 膠器(購自美國 Bio-Rad 公司) 15、 Pipetmen 16、 均質器 17、 微量離心管(購自季勗) 18、 冷凍管 19、 3M filter paper 20、 X-ray film (FUJI) 21、 微量天平. 二、 藥品試劑 1、. RPMI medium 1640 【內含 10% fetal calf serum(FCS)、 2 mM L-glutamine、100 units/ml Penicillin G、 100 µg/ml streptomycin sulfate】(購自 GIBCO). 27.
(28) 2、. Trypan blue (購自 SIGMA). 3、. Dimethyl Sulfoxide(DMSO)(購自 SIGMA). 4、. 5-Fluorouracil (購自 SIGMA;編碼:F6627). 5、. Leucovorin(購自 Wyeth). 6、. Trypsin – EDTA. 7、. Phosphate buffer saline (PBS)【由 NaCl, KCl, Na2HPO4, KH2PO4 組成】. 8、. Quick Cell Proliferation Assay Kit : WST-1 (購 自 Biovision). 9、. Bio-RAD Protein assay dye reagent concentrat, 450 ml. 10、 Protein Standard : bovine serum albumin (1 µg/ul) 11、 Enhance chemiuminescent (ECL)( 購自美國 AmershanPharmacia ) 12、 Ammonium persulfate(APS)(購自 SIGMA) 13、 Tetramethyl ethylenediamine (TEMED)(購自 SIGMA) 14、 Phenylmethylsulfonylfluoride (PMSF)(購自 SIGMA) 15、 一級抗體 :rTSβ type (由本研究室 周寬基博士自行研 發而成)及 TS106 16、 二級抗體:antimouse IgGb. 28.
(29) 17、 30% H2O2 18、 脫脂奶粉(blotto milk)(購自安佳品牌) 19、 goat serum 20、 AEC 試劑(購自 MERCK) 21、 Haematoxylin 試劑. 三、 人類乳癌細胞株的來源: 人類乳癌細胞株 5 株 (細胞株編號:MCF-1、MCF-7、60055、 BT-20、T47D)分別從 ATCC(American Type Culture Collection) 購買。其皆屬於乳腺癌細胞株(mammary gland. carcinoma cell. lines)。. 第二節、方法 : 一、細胞毒性分析 (Cytotoxic assay) (一) 人類乳癌細胞株的培養: 首先將存放人類乳癌細胞株(編號:MCF-1、MCF-7、60055、 BT-20、T47D)的冷凍管從液態氮中取出後,插上浮板,迅速放入 37℃水浴中使其快速解凍。取出冷凍管以擦手紙擦拭水滴後,以 70%酒精噴灑冷凍管外部後,移入無菌操作台。之後、將已解凍 的細胞株從冷凍管取出細胞加在含 5 ml 10%胎牛血清之培養基,. 29.
(30) 離心、 800 rpm、5 分鐘後,將上清液去除(儘量吸乾),再以 tip 來回抽吸方式打散細胞並加入 10 ml RPMI 的培養液於 T25 培養皿中,再將 5 株細胞株置於含 5% CO 2 的 37℃恆溫培養箱內 培養。每二天更換一次新鮮的培養液,每株細胞均間隔三至四天 就必須分盤培養。細胞分盤、首先將培養皿中舊的細胞培養液吸 出,加入 10 ml 1xPBS 的緩衝液清洗二至三次後,再加入 1 ml 的 trypsin-EDTA 溶液(0.005% trypsin, 0.002% EDTA)於培養 皿中,再放入恆溫培養箱 3 分鐘後,觀察細胞是否以脫離培養皿 (倘若未完全脫離,可用手輕輕拍打培養皿邊緣,以幫助細胞脫 落。) ,待細胞脫落後,加入 1 ml 的培養液稀釋中和 trypsin-EDTA 的作用,並將細胞吸出至含 5 ml 培養液的試管中,離心、 1500 rpm、5 分鐘後,將上清液去除再加入 2 ml 的培養液,利用 pipet 以抽吸的方式儘量將細胞打散。再將細胞懸浮液均分到新的培養 皿中,進行分盤或經過細胞計數後種入適當數目的細胞至 96 孔 細胞培養盤中預備進行細胞實驗。最後在於培養皿或培養盤中加 入 10%胎牛血清之培養液,即可放入恆溫培養箱培養。 一般在進行細胞實驗時,所使用的細胞株需將細胞解凍後,以繼 代培養三代以上方可進行實驗,因為細胞的生長及特性表現會趨 於? 定。. 30.
(31) 1.細胞數目計數法: 計算細胞數目是用血球計數盤(如右圖所示), 以 70% 酒精清洗擦乾,蓋上蓋玻片再取細胞 樣品 20µl 滴入下方三角形之區域,讓樣品依 虹吸現象吸入血球計數盤中。 一般血球計數盤有 2 個 chambers,每個 chamber 中細刻 9 個 1 mm2 大正方形,其中 4 個角落之正方形再細刻 16 個小格,深度均 為 0.1 mm。當 chamber 上方蓋上蓋玻片後,每 個大正方形之體積為 1 mm2 ×0.1 mm=1.0×10-4 ml。 使用時,計數每個大正方形內之細胞數目,壓線 的細胞一律計數,之後、乘以稀釋倍數,再乘以 104,即為每 ml 中的細胞數目。(如右圖所示) 一般在計數細胞時,應只計數樣品中的活細胞,依照 dye. 31.
(32) exclusion 的存活測試原理,利用藍色之 trypan blue 染料會 滲入死細胞中而呈色的特性,而活細胞因細胞膜完整,染料無 法滲入,故無法呈色,因此只計數白色的細胞數。 步驟:取 50μl 細胞懸浮液與 50μl trypan blue 等體積混合均 勻於 1.5ml 微量離心管中。取少許混合液(約 20μl)自血球計 數盤的 chamber 上方凹槽處加入,蓋上蓋玻片後,再放入倒立顯 微鏡下觀察,活細胞不染色,死細胞則為藍色。然後計數五個大 方格的細胞總數,再除以 5,乘以稀釋倍數,最後乘以 104 ,即 為每 ml 中細胞懸浮液的細胞數。若所含細胞密度過高(每一大 格超過 250 顆細胞),所算出的細胞數目,誤差可能較大,必須 先稀釋細胞後,再重新計數。稀釋時必須詳細計算稀釋倍數。若 樣品先經過稀釋才計數,則原樣品中的細胞數應再乘回稀釋倍數 (例如:染 trypan blue 時,樣品與 trypan blue 等體積混合, 故稀釋倍數至少乘以 2)。. 2.細胞毒性分析法: 本實驗採用較新的 tetrazolium salt 的 kit,此 kit 是由 WST-1 試劑(lyophilized)與 ECS(Electro Coupling Solution) 25 ml 混合而成。其測定原理是根據 MTT 的測定方法,基本上是. 32.
(33) 利用細胞本身的酵素對受質的作用,產生顏色的變化,再進一步 測定其吸光值。如果細胞受到細胞裂殖素的刺激,增生越多,則 活的細胞愈多,酵素的活性就會較高,可以測到較高的吸光值。 利用此一原理,也可以來測定細胞的增 生 (cell proliferation) 、 存 活 率 (percent of viable cells) 及 細 胞 毒 性 (Cytotoxicity)。但是此種測定方法通常對附著性細胞的準確度 較高,對懸浮性細胞則較不理想。針對 WST-1/ECS 試劑的優點 為:不需要沖洗,直接加入即可作用,也不需要經過溶解的步驟, 相較於 MTT 的測定方法,此 kit 所實驗出來的結果較快速且敏感 性佳。 步驟:將以計數過的細胞,分別各取 5000 顆至 20000 顆,放入 內含 100μl 之細胞培養液的 96 孔細胞培養盤中,再放進含 5% CO2、37℃的細胞培養箱內,放置 24 小時後,取出加入抗癌藥物 5-FU,其濃度分別為 1.6 μM 至 50 mM。另一組加入 5-FU 同時 又加入 leucovorin (LV) 1.25 mM,混合均勻後,再放入細胞培 養箱,經過 72 小時後,取出 96 孔細胞培養盤,各別加入 WST-1/ECS 試劑 10μl 在每一個 well 中,混合均勻後,分別放 置 1 小時、2 小時及 4 小時,之後取出振搖一分鐘,再利用酵素 免疫分析儀(ELISA reader)去測定第 1 小時、第 2 小時及第 4. 33.
(34) 小時的 OD 值(波長設定為 450 nm、630 nm)。每一種細胞株都 做三重覆,以求實驗結果更加精確。. *乳癌細胞株的生長曲線製作: 利用血球計數盤取出五種不同的細胞株,以 40000 顆的細胞為 主,用 1 比 2 的稀釋比例配製成以下細胞數,分別為 20000 顆、 10000 顆、5000 顆、2500 顆、1250 顆、625 顆及 313 顆,並種入 96 孔細胞培養盤中,再放入細胞培養箱,經 24 至 72 小時後,利 用倒立顯微鏡來觀察五種不同細胞株的生長情形並繪製生長曲 線。. 二,西方墨點法(Western blotting) 利用此法來檢測人類乳癌細胞內與抗藥性相關蛋白質的表現 (一)、細胞蛋白質萃取: 先將已長滿細胞的 T75 細胞培養盤從培養箱取出後,吸出培養 廢棄液,再以 10 ml 的 1 x PBS 緩衝液小心清洗 3 次後,加入少 量的 1 x PBS 緩衝液於 T75 細胞培養盤中,用刮杓將培養盤中的 細胞全部刮出,之後倒入離心管中離心,2000 rpm、10 分鐘後, 倒掉上清液,加入 150 μl 的 lysis buffer (1% Nonidet P-40. 34.
(35) +10% SDS+0.5% sodium deoxycholate ) 及 10 mg/ml PMSF 適 量體積(約總體積的百分之一),之後置於冰上超過 20 分鐘或用 研磨器研磨數分鐘,再吸取液體放入 1.5 ml 微量離心管中離心, 10000 rpm、4℃離心、5 分鐘後,取上清液,即含細胞總蛋白質。 萃取後的蛋白質需存放在 -30℃的冰箱。 (二)、蛋白質定量: 蛋白質的濃度是依據 Bradford 的方法測定。其分析原理是根 據 BCA 法。BCA 於鹼性條件下可還原雙價的銅離子(Cu2+)成為單 價的亞銅離子(Cu+),再由兩分子的 BCA 去螯合亞銅離子,而產 生藍紫色的產物。以胎牛血清蛋白(Bovine serum albumin;BSA) 為蛋白質標準品。利用 Bio-Rad Protein Assay Kit (美國 Bio-Rad 公司)套裝式劑,來進行蛋白質定量的檢測。. 1.蛋白質標準品檢量線製作: 濃度(mg/ml) 2 mg/ml BSA ddH2O (μl). Bio-RAD Proteinn Assay ddH2O assay dye reagent. (μ l). 0.2. 4. 16. 200 μl. 800 μl. 0.4. 8. 12. 200 μl. 800 μl. 0.6. 12. 8. 200 μl. 800 μl. 35.
(36) 0.8. 16. 4. 200 μl. 800 μl. 1.0. 20. 0. 200 μl. 800 μl. 【總體積為 1020 μl, 每個濃度配製二套 】. 將配製好的蛋白質標準品的樣品放入石英管中,利用分光光度計波 長 595 λ來測其吸光值。二套之間的吸光值其偏差不能超過 10% , 否則需重新配製。經過平均後的吸光值與濃度畫出標準品檢量線, 並求出趨勢線方程式及 R2 值。. 2.樣品蛋白質定量 : ∼依照下列方式製作蛋白質樣品: 稀釋倍數(X) 樣品蛋白質(μ l) ddH2O (μl). Protein. assay. reagent(μl) 5 X. 4. 16. 1000. 10 X. 2. 18. 1000. 20 X. 1. 19. 1000. 【總體積為 1020 μl, 每個濃度配製二套 】. 本實驗的蛋白質樣品製作,是將樣品稀釋成 5 倍,再將配製好的蛋 36.
(37) 白質樣品放入石英管中,利用分光光度計波長 595 λ來測其吸光 值。二套之間的吸光值其偏差不能超過 10% ,否則需重新配製。將 樣品吸光值的平均值代入趨勢線方程式,則可算出稀釋後樣品之蛋 白質濃度,最後再乘上稀釋倍數,則可求得實際蛋白質樣品濃度。. (三)、膠體電泳分析法(SDS-PAGE) 測定 rTS 及 TS 蛋白質 先配製 0.75 mm 厚度之 10% 下層膠及 4%的下層膠,其配製法如下。 * 10 % 下層膠 (10% Separating):兩片量 先加入 5 ml 的 ddH2O 在 50 ml 的離心管中、再加入 4 ml 的 30% acryamide/Bis(37.5:1) ,之後再依序加入 3 ml 的 1.5 M Tris-HCl (pH8.8)、各 120 μl 的 10﹪SDS 及 10﹪APS,最後再加入 6 μ l 的 TEMED (目的是促進下層膠凝集),等混合均勻後,即為 10 % 下 層膠。 * 4 % 上層膠(4% stacking): 先加入 1.8 ml 的 ddH2O 在 15 ml 的離心管中,再加入 0.36 ml 的 30% acryamide/Bis(37.5:1),之後再依序加入 o.75 ml 的 0.5 M Tris-HCl(pH 6.8)、各 30 μ l 的 10﹪SDS 及 10﹪APS,最後再加 入2μ l 的 TEMED (目的是促進下層膠凝集),等混合均勻後,即為. 37.
(38) 4 % 上層膠。 *. 6X Sample buffer 配製法如下: 先加入 1.5 ml 的 2M Tris-HCl(pH 6.8)在 15 ml 的離心管中, 再依序加入 0.93 gm 的 DTT、1.2 gm 的 SDS 及 Bromophenol blue 、再加入 6 ml 的 100% Glycerol,最後再加 ddH2O 至總體 積為 10 ml,並分裝在 1.5 ml 的微量離心管中,於 -20℃下保存。. * 10 X Running buffer(10 X Tris-glycine)配製法如下: 取 30 g 的 Tris-base 、再取 144 g 的 glycine 後,加入 100ml 的 10﹪SDS,最後再加 ddH2O 至總體積為 1000 ml,4℃ 保存,使 用前先稀釋成 1X。 步驟:將配製好的 10% 下層膠注入? 膠台後,緩緩加入 ddH2O 以 去除氣泡,並壓平下層膠之上緣,放置 30 分鐘等膠體凝固後,再 將 4 % 下層膠緩緩注入,並插入樣品槽梳子(comb)至未凝固的 膠體中,避免氣泡出現於樣品槽(wells)下緣。放置約 20 分鐘 等膠體凝固後、再拔出樣品槽梳子並以 ddH2O 小心沖洗樣品槽內, 避免雜質殘留。將? 好的樣品槽放入電泳槽中,加入 1X Running buffer 至高過樣品槽,用針筒以抽吸的方式去除膠體下緣殘留的 氣泡。之後、再準備 loading 預測定的蛋白質樣品。 首先、先 loading 5 μ l 的分子量標準品(rainbow marker),再. 38.
(39) 依序由左至右 loading 已配製好的樣品(此樣品與 6X Sample buffer 依照比例混合後,經 95℃加熱 5 分鐘後,於-20℃下冷卻 5 分鐘)至樣品槽中,以設定 75 伏特、跑膠 2 小時。. (四)、西方墨點法(Western blotting): Semi-dry 轉漬法: * Transfer buffer 配製法如下: 取 8.72 g 的 Tris-Hcl 、再取 4.4 g 的 glycine 後,加入 1.5 L 的 ddH2O,混合均勻後,將所配製的溶液調整其 PH 值至 8.4,再 加 5.6 ml 的 10﹪SDS,最後加入 300 ml 的 Methanol 即為 transfer buffer。 步驟:先將semi-dry轉漬夾打開,鋪上3張以Transfer buffer 浸潤過的3M濾紙,並將跑完的SDS-PAGE gel依序放在3M濾紙的上 層,再依序加入PVDF轉漬膜(須用100%甲醇潤濕20秒),最後再 鋪上3張的3M濾紙於最上層。以設定10伏特、通電1小時來進行蛋 白質轉漬的步驟。待轉漬完成後,取出PVDF轉 漬 膜 來 進 行 Blocking的步驟(將PVDF轉漬膜放入5%脫脂奶粉中、混合振搖30 分鐘) 。之後、取出轉漬膜於小盒中並以0.05% PBST 緩衝液清洗. 39.
(40) 3次、每次間隔10分鐘,再將轉漬膜以鑷子夾入含一級抗體( rTS β以1比6000的稀釋比例加在5% blotto blocking solution中; TS以1比200的稀釋比例加在5% blotto blocking solution中) 的密封盒,放入4℃的冰箱overnight。隔天取出密封盒,在室溫 下振搖1小時後,用0.05% PBST 緩衝液清洗3次、每次間隔10分 鐘,接下來再取出轉漬膜加入二級抗體(antimouse IgGb以1比 5000的稀釋比例加在5% blotto blocking solution中) ,在室溫 下振搖1小時後,同樣以0.05% PBST 緩衝液清洗3次、每次間隔 10 分 鐘 。 再 取 出 轉 漬 膜 加 入 冷 光 劑 ( enhanced chemico -luminescent;ECL)之冷光受質Solution 1、及 Solution 2 各 取 1 ml等量混和均勻後,將轉漬膜浸潤1 分鐘後,取出再放入 壓片夾中,至暗房壓片顯影(採用FUJI medical X-ray film )。 其曝光時間為30秒至 10分鐘 ,經過抗體辨識之蛋白質影像 (band),會呈現在 X-ray底片上。. (五)、染色呈色法: 利用 AP buffer(組成:100 mM Nacl+5 mM MgCl2+100 mM Tris-Hcl;pH 9.5)加上 66 μ l/10 ml 的 NBT(p-Nitro blue tetrazolium chloride)及 165 μl/10 ml 的 BCIP(5-Bromo-4-. 40.
(41) chloro-3-indoxyl phosphate )之染劑來呈色。先將轉漬後的 PVDF 轉漬膜放入 10 ml 的 AP buffer/66 μ l NBT/165 μl BCIP 之染劑中,直到轉漬膜上出現欲表現之蛋白質影像(band)時, 即可取出轉漬膜,並放在大量清水中,以終止反應。隨後將轉 漬膜取出並自然晾乾。 三、免疫細胞染色法(Immunocytochemistry;ICC): 首先將已長滿細胞的載玻片,放入以 50%甲醇與 50%丙酮等量之 混合液中,浸泡 15 分鐘,目的是讓細胞株固定在載玻片上。取出 後晾乾,再用 3%的過氧化氫(100 μ l 30% H 2O2 +100 μ l sodium azide + 800 μ l ddH 2O)處理 15 分鐘後,取出載玻片,利用 ddH2O 沖洗 3 次並晾乾,接下來則進行 blocking 的步驟(blocking solution 之配製為 5% goat serum + 1x PBS)。將載玻片加入 少量的 5% goat serum,放置 5 分鐘後,直接加入一級抗體(rTS β以 1 比 100 的稀釋比例加在 5% goat serum 中)於載玻片上作 用 2 小時。之後用 ddH2O 沖洗 3 次便可加入 biotinylated 的二級 抗體,於室溫下作用 15 分鐘,再用 ddH2O 沖洗 3 次後,即可加入 AEC 試劑呈色(此試劑組成為 500 μl AEC+500 μ l1M NaOAc +50 μl 3% H 2O2+4000 μ l ddH 2O),並在室溫下作用 20 分鐘,注意此 步驟需避光操作。之後再用 ddH2O 沖洗 3 次後,將已呈色的細胞. 41.
(42) 株浸泡在 haematoxylin 的試劑(將 haema- toxylin 與 ddH 2O 等 比例混合)中,作用 20 分鐘,其目的是染背景色。再以活水(自 來水)沖洗玻片約 10 分鐘,使背景呈藍紫色以方便觀察染色的結 果。最後將玻片自然晾乾後,再利用 Gelatin 封片保存。. 四、 統計方法 本實驗利用 PRISM4 的電腦軟體(購自 Graph Pad 公司)及線性 迴歸(linear regression)等方法來分析細胞的生長曲線,癌細 胞的存活率及細胞毒性作用(即癌細胞被抑制 50%時的濃度; IC50)。. 42.
(43) 第四章、結果 第一節、 西方墨點法:檢測 rTSβ及 TS 蛋白在人類乳癌 細胞株的表現量 一、 利用西方墨點法來檢測 rTSβ蛋白在五株乳癌細胞株的表現 量,將每株細胞株都 loading 30μl 的量,如圖五實驗結果顯 示,在 47 kDa 分子量的標記處以 H630-1 細胞株為 positive 對 照組,發現在這五株細胞株中僅有 T47D 這株細胞株幾乎不表 現 rTSβ蛋白或表現量很弱,其餘的細胞株都有 rTSβ蛋白的 表現且表現量以 60055 及 BT-20 這兩株細胞株為最多,其次依 序為 MCF-1 及 MCF-7 這兩株細胞株。. 二、 利用西方墨點法來檢測 TS 蛋白在五株乳癌細胞株的表現 量,將每株細胞株都 loading 40 μl 的量,如圖六實驗結果 顯示,在 36 kDa 分子量的標記處以 HL-60 細胞株為 positive. 43.
(44) 對照組,發現在這五株細胞株中皆有表現 TS 蛋白,而 T47D 細胞株的 TS 表現量與其餘四株細胞株比較起來並沒有明顯 的差異。這樣的結果恰巧與圖五的 rTSβ蛋白的表現剛好相 反。. 第二節 、細胞毒性分析 : 利用快速細胞增生分析套組 (WST-1 kit 來檢測 5-FU 對人類乳癌細胞株的 毒殺作用 一、利用 WST-1 分析法來檢測 5-FU 對人類乳癌細胞株的毒殺作 用,其中將五株乳癌細胞株以不同的 5-FU 濃度及 5-FU 加上 leucovorin (LV) 1.25 mM 的濃度培養 72 小時後,評估 5-FU 及 5-FU 加 LV 之藥物對五株乳癌細胞株的增生影響。首先我們先評 估以不同的 LV 濃度對五株乳癌細胞株的增生作用,如圖七發現 LV 對五株乳癌細胞株的增生作用會隨著濃度的增加而有明顯的 抑制效果。尤其在 10 mM 的濃度下,幾乎所有乳癌細胞株的生長 都被 LV 抑制了,只是抑制的程度不同。所以選擇適當的 LV 濃度 與 5-FU 合併作用是很重要的研究設計,LV 的濃度選擇必須對乳 癌細胞株的生長要完全沒有抑制作用,如此才能評估 5-FU 加 LV 之藥物後,是否能增強 5-FU 對乳癌細胞株的毒殺作用?因此由. 44.
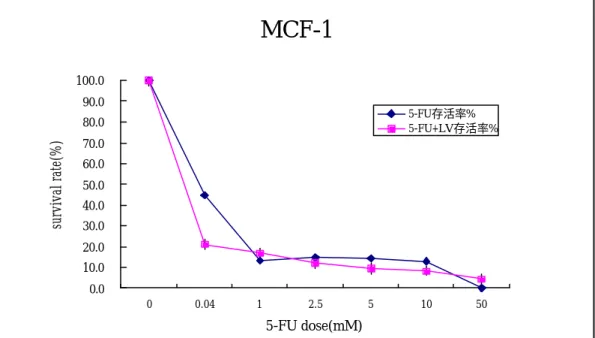
(45) 圖七的實驗結果而選擇 LV 的 1.25 mM 做為與 5-FU 合併作用的基 準濃度。. 二、 接著再進行評估 5-FU 及 5-FU 加 LV 之藥物對五株乳癌細胞 株的細胞毒殺作用,發現 5-FU 對乳癌細胞株如圖八、圖九顯示 5-FU 在 60055 及 BT-20 這兩株細胞株的 IC50 之反應曲線,其 IC50 濃度分別為 0.042 mM 及 2.54 mM,且加上 LV 之藥物後都有明 顯增強這兩株細胞的毒殺作用,使細胞的存活率明顯下降。又如 圖十、十一顯示 5-FU 在 MCF-1 及 MCF-7 這兩株細胞株的 IC50 之 反應曲線,其 IC50 濃度分別為 0.41 mM 及 0.36 mM,加上 LV 藥 物後其細胞毒殺作用增強的曲線就不如 60055 及 BT-20 這兩株細 胞株明顯。針對 T47D 細胞株的實驗結果如圖十二所示,5-FU 對 T47D 細胞株的增生會隨著濃度的增加而有明顯的抑制效果,且 毒殺作用也很敏感,其 IC50 的濃度為 0.013 mM ,加上 LV 藥物 後使其細胞毒殺作用增強為 6 倍,與其餘四株細胞株做比較,會 發現幾乎不表現 rTSβ蛋白的 T47D 細胞株其對 5-FU 的敏感度 佳,相對於有 rTSβ蛋白表現的細胞株其對 5-FU 的敏感度較差, 換句話說、即有較高的抗藥性。. 45.
(46) 三、 如表 1 的結果顯示,發現具有 rTSβ蛋白高度表現的細胞, 如 60055 及 BT-20 這兩株細胞株,及 rTSβ表現量中等的 MCF-1 及 MCF-7 這兩株細胞株,其 5-FU 之 IC50 的濃度與幾乎不表現 rTS β蛋白或表現量很弱的 T47D 細胞株做一比較,會發現有 rTSβ 蛋白表現量的細胞株其 5-FU 之 IC50 的濃度會比幾乎不表現 rTS β蛋白的 T47D 細株明顯多了 3 至 195 倍。換句話說、有 rTSβ 蛋白表現的細胞株,有較高的抗藥性產生,且會明顯降低 5-FU 的細胞毒殺作用。. 四、針對 5-FU 對人類乳癌細胞株的敏感性試驗,選定單一 5-FU 濃度:5 mM 以進行實驗。如圖十三的結果顯示,發現 BT-20 細 胞株在 5 mM 的 5-FU 濃度下,竟然還高達 42﹪的細胞存活率, 而 60055、MCF-1 及 MCF-7 這些細胞株也依序有 18﹪、14.1%及 11﹪的細胞存活率,與 BT-20 細胞株做比較也有 2 至 4 倍的差 異,但相較於 T47D 細胞株已沒有細胞存活了。因此、我們發現 T47D 細胞株對 5-FU 的敏感性的確比其餘四株都來的好。. 46.
(47) 第三節、免疫細胞染色法:檢測 rTSβ蛋白質在人類乳 癌細胞株的表現 利用免疫細胞染色法來檢測 rTSβ蛋白在人類乳癌細胞株的表 現。如圖十四的染色結果顯示,以 H630-1 細胞株為 positive 對 照組,發現圖 A 的 H630-1 細胞株經過 AEC 試劑及 haematoxylin 的試劑染色後,其細胞核呈現很明顯的紅色,即為 rTSβ在細胞 內的表現,其表現的強度是以含有 rTSβ細胞比例的數目為依據 來定義:一般超過總細胞數 50%以上,則定義為高度表現;10-50% 為中度表現;低於 10%以下為微弱表現甚至不表現(根據病理科的 guideline 判定)。從圖 B、C 可發現 60055 及 BT-20 這兩株細胞 株,其 rTSβ在細胞核的含量很多,屬於高度表現,而圖 D、E 的 MCF-1 及 MCF-7 這兩株細胞株,其 rTSβ多表現在細胞質中, 則屬於中度表現。至於圖 F 的 T47D 這株細胞株,我們發現它在 細胞核及細胞質中幾乎不表現 rTSβ。. 47.
(48) 第五章、 討論 本實驗利用西方墨點法來檢測 rTSβ及 TS 蛋白在人類乳癌 細胞株的表現量,結果發現在這五株細胞株中僅有 T47D 這株細胞 幾乎不表現 rTSβ蛋白,其餘的細胞株都有 rTSβ蛋白的表現且表 現量以 60055 及 BT-20 這兩株細胞株為最多。相反的、在 TS 蛋白 的表現細胞間的表現量沒有明顯差異。根據 Dr. Dolnick 先前所作 的研究顯示,H630-1、對 5-FU 具有抗藥性的細胞株其 rTSβ的表現 量高於 H630 細胞株達 40 至 70 倍之多,甚至在許多不同的抗藥細 胞株如 K562B1A、HCT-8/DF2 其 rTSβ的表現量與上述結果相似,都 有過度表現的現象,但 TS 的表現量並沒有明顯差異( 19,22,23 ) 。 於是推論,5-FU 的抗藥性可能與 rTSβ的表現比較有關、至於與 TS 的關係、顯示 rTSβ可負向調控 TS 基因的表現(19)。我們可由 T47D 細胞株觀察到此現象。因為 T47D 細胞株幾乎不表現 rTSβ,僅稍有 表現 TS 蛋白,而此細胞株對 5-FU 的敏感性佳。至於 60055 及 BT-20. 48.
(49) 這兩株細胞株皆屬於 rTSβ高表現量,但對 5-FU 的敏感性相較於 T47D 而言則比較差。我們進一步發現一些有趣的結果為:60055 與 BT-20 這兩株細胞株雖然同屬於 rTSβ的高度表現,但對 5-FU 的敏 感性彼此間也有些許的差異。例如 60055 細胞株與 BT-20 細胞株在 5 mM 的 5-FU 濃度下其存活率分別為 42.1%及 18.2% (如圖十三), 彼此的存活率相差了兩倍多,因此、我們推論,60055 細胞株對 5-FU 的敏感性較 BT-20 細胞株好。至於是什麼原因或機轉會造成這樣的 結果,目前尚未清楚,有待未來再做進一步的研究與探討。 再者,我們利用快速細胞增生分析法來檢測 5-FU 對人類乳 癌細胞株的毒殺作用,結果發現 5-FU 加上 Leucovorin 的合併療 法,的確比僅用 5-Fu 有明顯的增強效果,且增強約 2 至 4 倍(如表 一)。根據 Chow 等學者指出,Leucovorin 在細胞內會被轉變為 5,10- methylenetetrahydrofolate (mTHF)的還原葉酸輔助因子, 而 5-FU 的活性代謝物 FdUMP 會與細胞內的 dUMP 競爭 TS,與 mTHF 及 TS 形成一個穩定的共價鍵之三物複合體(ternary complex)(如 圖二) ( 3,37)。所以在有充分的 mTHF 存在時,TS 的活性就會被 FdUMP 抑制,而不能產生 dTMP。相反的,只有 FdUMP 而沒有 mTHF 存在時,有些癌細胞就會產生抗藥性,那是因為沒有足夠的 mTHF 來產生三物複合體。因此在這種情況下,可藉由 Leucovorin 合併. 49.
(50) 使用或增加 Leucovorin 的濃度來增強 5-FU 的細胞毒殺作用,並扭 轉 5-FU 的抗藥性(如圖四) (37)。但並非所有細胞使用 5-FU 與 Leucovorin 的合併療法會明顯增加 5-Fu 的毒殺作用。我們由實驗 結果得之,MCF-1 及 MCF-7 這兩株細胞株在 5-Fu 之 IC50 的濃度時, 加入 Leucovorin 之後,的確能增加 5-Fu 約兩倍的細胞毒殺作用(如 表一),但超過了 1 mM 的濃度時,Leucovorin 似乎無法明顯提升 5-Fu 的作用,可能是因為 MCF-1 及 MCF-7 這兩株細胞株在超過 5-Fu 之 1 mM 濃度下,細胞的存活率僅介於 10 至 20%之間,無法明顯提 升 5-Fu 的作用。至於 T47D 細胞株、其對 5-FU 的敏感性佳,加入 Leucovorin 之合併療法後更可明顯增加 5-Fu 約 6 倍的細胞毒殺作 用(如表一)。且發現有 rTSβ蛋白表現量的 4 株細胞株其 5-FU 之 IC50 的濃度會比幾乎不表現 rTSβ蛋白的 T47D 細株明顯多了 3 至 195 倍,於是我們推論,有 rTSβ蛋白表現的細胞株有較高的抗藥 性產生,且會明顯降低 5-FU 的細胞毒殺作用,但相對於 TS 蛋白的 表現量並沒有明顯差異。根據 Dolnick 先前的發現,顯示 rTSβ可 負向調控 TS 基因表現,而 TS 基因表現在快速生長細胞扮演相當重 要的角色,所以他們也開始探討在一個快速生長的癌細胞中,TS 和 rTSβ基因的變化關係。並分析各種臨床檢體一對對正常和癌細 胞 TS 和 rTSβ的比例,結果發現、在乳癌、大腸癌、腎癌、肺癌及. 50.
(51) 卵巢癌細胞中,rTSβ基因表現有意義的被調降了;這暗示 rTSβ基 因可能是一個抑癌基因(tumor suppressor gene) (19, 21-26)。 回顧有關 rTSβ在乳癌細胞的表現與 5-FU 抗藥性的研究,截至 目前為止,沒有文獻報導有關 rTSβ在乳癌細胞的表現與 5-FU 抗藥 性的關係。根據我們的假設,認為 5-FU 的抗藥性可能與 rTSβ的表 現比較有關與 TS 的表現量則沒有明顯的差異。因此,藉由本實驗 的結果,證實有 rTSβ表現的乳癌細胞,可能會降低 5-FU 的細胞 毒殺作用而造成 5-FU 的抗藥性產生,並影響 5-FU 在臨床上的療效。. 51.
(52) 第六章、 結論 根據本研究的結果證明了五株人類乳癌細胞株除了 T47D 外、其餘 的細胞株都有 rTSβ的表現,而且有 rTSβ表現量的細胞株其 5-FU 之 IC50 的濃度都比 T47D 明顯多了 3 至 195 倍。換句話說、有 rTSβ表現 量的細胞株,其會明顯降低 5-FU 的細胞毒殺作用而造成 5-FU 的抗藥 性產生。至於與 TS 表現量的關係,可由圖六的結果得之;在這五株人 類乳癌細胞株中,並沒有明顯的差異。 再者,本研究希望藉由人類乳癌細胞株來建立 rTS 與 5-FU 抗 藥性的關聯性,以改善 5-FU 在乳癌之輔助性療法的效果,並增加 5-FU 在臨床上的地位。並藉由 rTSβ的表現量,可做為 5-FU 在臨床上的治 療指標。同時也期望未來能利用此實驗模式應用在不同的癌症治療上, 以探討不同化療藥物的抗藥機轉,進而提升化療藥物在臨床上的應用。. 52.
(53) 參考資料 1. 行政院衛生署生命統計,2003。 2. 國家衛生研究院(NHRI)/台灣癌症臨床研究合作組織(TCOG)/乳癌 診斷與治療共識,2003。 3. Daniel B. Longley, D. Paul Harkin & Patrick G. Johnston. 5-Fluorouracil:Mechanisms of action and clinical strategies. Nature Reviews.2003;3:330-338. 4. Diasio, H. & Harris, B.E. clinical pharmacology of 5fluorouracil. Clin. Pharmacokinet. 1989;16:215-237. 5. William R. Montfort & Andrzej Weichesl. Thymidylate synthase:structure, inhibition, and strained conformations during catalysis. Pharmacol. Ther.1997;76: 29-43. 6. Carreras, C. W. & Santi. D. V.The catalytic mechanism and structure of thymidylate synthase. Annu. Rev. Biochem. 1995;64:721-762. 7. Peter GJ, Jansen G, Schilsky RL, Milano GA, Ratain MJ. Principles of antiplastic drug development and pharmacology, Marcel Dekker, New York, 1996;543-585. 8. Robert M. Mader, Markus Muller & Gunther G. Review article: Resistance to 5-Fluorouracil.1998;31:661-666. 9. Sobrero A.F. Aschele C. and Bertino J.R. Fluorouracil in colorectal cancer – a tale of two drug::implications for biochemical modulation. J. Clin. Oncol.1997;15:368-381. 53.
(54) 10. Pinedo HM, Peter GJ. 5-Fluorouracil : biochemistry and pharmacology. J. Clin. Oncol.1988;6:1653-1664. 11.Peter GJ. Jansen G. Resistance to antimetabolites. In Schilsky RL, Milano GA, Ratain MJ, eds. Principles of cancer drug pharmacology. Marcel Dekker, New york,1995,in press. 12.Spear CP, Gustavsson BG, Berne M, Frosing R, Bernstein L, Hayes AA. Mechanisms of innate resistance to thymidylate synthase inhibition after 5-fluorouracil. Cancer Res. 1988;48:5894-5900. 13.Priest DG, Ledford SE, Doig MT. Increased thymidylate synthase in 5-fluorodeoxyuridine resistant cultured hepatoma cells. Biochem Pharmac. 1980;29:1549-1553. 14.Yin MB, Zakrzewski SF, Hakala MT. Relationship of cellular folate cofactor pools to the activity of 5 -fluorouracil. Mol. Pharmac. 1983;23:190-197. 15.Berger SH, Hakala MT, Relationship of dUMP and FdUMP pools to inhibition of thymidylate synthase by 5 -fluorouracil. Mol. Pharmac.1984;25:303-309. 16.Berger SH, Barbour KW, Berger FG. A naturally occurring variation in thymidylate synthase structure is associated with a reduced response to 5-fluoro-2’-deoxyuridine in a human colon tumor cell line. Mol. Pharmac.1988;34:480-484. 17.Kinsella A.R. and Pickard M. Resistance to chemothera – peutic antimetabolites : a function of salvage pathway involvement and cellular response to DNA damage. Br. J. Cancer.1997;75:935-945. 18. Dolnick BJ,and Black A. Alternate splicing of the rTS Gene product and its overexpress in a 5 -fluorouracilresistant cell line. Cancer Res.1996;56:3207-3210. 19.Dolnick BJ, and Norman J.A novel function for the rTS gene. Cancer Biology & Therapy.2003;2:19-24. 20.Dolnick BJ, Cloning and characterization of a naturally 54.
(55) occurring antisense RNA to human thymidylate synthase mRNA. Nucleic Acids Res. 1993;21:1747-52. 21.Dolnick B, Su J. GenBank:GenBank;2000. 22.Dolnick BJ, Black AR, Alternate splicing of the rTS gene product and its overexpression in a 5-fluorouracilresistant cell line. Cancer Res. 1996;56:3207-10. 23.Black AR, Dolnick BJ, Expression of rTS correlates with altered growth regulation of thymidylate synthase. Cancer Res. 1996;56:700-5. 24.Dolnick BJ, Lu K, Yin MB, Rustrum YM, Recent advances in the study of rTS proteins. rTS expression during growth and in response to thymidylate synthase inhibitors in human tumor cells. Adv Enz Reg. 1997;37:95-109. 25.Chu J. Natural antisense (rTS α ) RNA mediated downregulation of thymidylate synthase gene expression [Ph.D.]. Buffalo, NY, SUNY at Buffalo;1999. 26.Chu J. Dolnick BJ, Natural antisense (rTSα) RNA induces site-specific cleavage of thymidylate synthase mRNA. Biochim Biophys Acta. 2002;1587:183-93. 27.Sugerman PB, Joseph BK, Savage NW. Review article:the role of oncogenes, tumor suppressor genes and growth factors in oral squamous cell carcinoma: a case of apoptosis versus proliferation. Oral Dis.1995;1:172-188. 28.Rebecca S. Carol B. Concepts in oncology therapeutics. 2000;2nd:2-18. 29.Nishimura R, Nagao K, Miyayama H, Matsuda M, Baba K, Matsuoka Y, Yamashita H, Fufuda M, Thymidylate synthase level as a therapeutic and prognostic predictor in breast cancer. Anticancer Res. 1999;19:5621-26. 30.Nishimura R, Nagao K, Miyayama H, Matsuda M, Baba K, Matsuoka Y, Yamashita H,Higuchi A, Suzuki S, Ishihara A,Prognostic significance and application to adjuvant therapy of flow cytometric DNA analysis in breast cancer. Jpn J Breast cancer. 55.
(56) 1994;9:287-292. 31.Ejendal KF, Hrycyna CA. Multidrug resistance and cancer: the role of the human ABC transporter ABCG2. Curr Protein Pept Sci. 2002 ;3 : 503-11. 32.Peter GJ, Backus HHJ, Freemantle S,Triest B, Smid K. Lunec J. Review article: Induction of thymidylate synthase as a 5-fluorouracil resistant mechanism. Biochimica et Biophysica Acta.2002;1587:194-205. 33.Peter GJ, Van Der Wilt CL, Van Triest B, Codacci-Pisanelli G, Johnston PG, Pinedo HM. Thymidylate synthase and Drug Resistance. Eur J Cancer.1995;31:1299-1305. 34.Wang FS, Aschele C, Sobrero A, Chang YM,Bertino JR. Decreased folylpolyglutamate synthase expression:a novel mechanism of fluorouracil resistance. Cancer Res. 1993; 53:3677-3680. 35.Peter GJ, Van Groeningen CJ. Clinical relevance of biochemical modulation of 5-fluorouracil.Ann Oncol. 1991; 2:469-480. 36.Dominic A, Linda R, Polly E, Marck C. Drug information handbook for oncology. 2000-2001;2nd:254-256. 37.Chow KC, Chow SL, Chi KH & Chen KY. The application of leucovorin in the chemotherapy of cancers. Therapeut. Radiol. Oncol. 1995;2:79-85. 38.Bleyer WA. New vista for leucovorin in cancer chemotherapy. Cancer 1989;63:995-1007. 39.Erlichman C, folate-based fluorouracil Chemotherapy. Mitrovski B. Comparative cytotoxicity of inhibitors of thymidylate synthase and +/- leucovorin in MGH-U1 cells. Cancer & Pharmacology. 1994;34:51-56.. 40.Wang FS, Aschele C, Sobrero A, Chang YM, Bertino JR. Cancer Res. 1993;53:3677-3680. 41.Komaki K, Kamamura Y, Ohmine Y, Sasa M, Tanaka K, Inoue H, Uyama T, Morimoto T, Monden Y. Difference in thymidylate 56.
(57) synthase activity in involved nodes compared with primary tumor in breast cancer patients. Breast Cancer Res Treat. 1995;35:157-162. 42.Pestalozzi BC, Peterson RD, Gelger A, Goldhirsch A, Guselerson BA, Trithia H, Lindtner J, Corles-Funes H, Simmoncini E, Byrne MJ, Golouh R, Rudenslam CM, Castiglione-Gerlsch M, Altegra CJ, Johnston PG. Prognostic importance of thymidylate synthase expression in early breast cancer. J Clin Oncol. 1997;15:1923-1931. 43.Mitamura T, Sakamoto S, Kudo H, Susuki S, Maemura M, Nakayama T. Clinicopathological stages and mammary DNA synthesizing enzyme activity in patients with mammary cancer. Medicine and Biology. 1996;133:61-64. 44.Larsson PA, Carlsson G, Gustavsson B, Spears CP. Thymidylate synthase in advanced gastrointestinal and breast cancers. Acta Oncologica. 1996;35:469-472. 45.Saga Y, Susuki M, Mizukami H, Kohno T, Takei Y, Fukushima M, & Ozawa K. Overexpression of thymidylate synthase mediates desensitization for 5-fluorouracil of tumor cells. Int. J. Cancer. 2003;106:324-326. 46.Suda Y, Kuwashima Y, Tanaka Y, Uchida K, Akazawa S. Immunohistochemical detection of thymidylate synthase in advanced gastric cancer:a prognostic indicator in patients undergoing gastrectomy followed by adjuvant chemotherapy with 5-fluoropyrimidines. Anticancer Res. 1999;19:805-10. 47.Habara K, Ajiki T, Kamigaki T, Nakamura T, Kuroda Y. High expression of thymidylate synthase leads to resistance to 5-fluorouracil in biliary tract carcinoma in vitro. Jpn J Cancer Res.2001;92:1127-32. 48.Wang W, Marsh S, Cassidy J, McLeod HL. Pharmacogenomic dissection of resistance to thymidylate synthase inhibitors. Cancer Res. 2001;61:5505-10. 49.Schaefer A, Hanzelka B, Parsek M, Greenberg E. Detection, purification, and structural elucidation of the acyl57.
(58) homoserine lactone inducer of vibrio fischeri luminescence and other related molecules. In Ziegler M, Baldwin T, editors. Bioluminescence and chemiluminescence. San Diego::Academic Press;2000:288-301. 50.Camara M, Daykin M, Chhabra SR. Detection, purification and synthesis of n-acylhomoserine lactone quorum sensing signal molecules. Meth Microbiol 1998;27:319-30. 51.Waston WT, Minogue TD, Val DL, von Bodman SB, Churchill ME. Structural basis and specificity of acyl-homoserine lactone signal production in bacterial quorum sensing. Mol Cell 2002;9:685-94.. 58.
(59) 附錄. 圖一、 為 5-FU 的代謝途徑 (Nature Reviews Cancer 3:330-338,2003). 59.
(60) 圖二、 為 5-FU 抑制 TS 的作用機轉。 (Nature Reviews Cancer 3:330-338,2003). 圖三、. Leucovorin 和葉酸的代謝環. 60.
(61) 圖四、. H630-1. T47-D. MCF-7. MCF-1. rTSβ 在乳癌細胞株的表現. 47kDa. H630-1 為 positive control. 圖五、 rTSβ 在乳癌細胞株的表現. 61. HL-60. T47-D. MCF-7. MCF-1. BT-20. H630-1 為 positive control. 60055. 47kDa. BT-20. 60055. 圖四、Leucovorin 的作用機轉.
(62) 36kDa. 36kDa. 圖六、 TS 在乳癌細胞株的表現 HL60 為 positive control LV effect 60055survival rate 100. MCF-1 survival rate. 80. MCF-7 survival rate T47-D survival rate. 70 60 50 40 30 20 10. 10. 2.. 5. 5. 0 1.2 5. survuval rate (%). BT-20 survival rate 90. LV conc. (mM). 圖七、不同 LV 的濃度在乳癌細胞株的生長抑制作用. 62.
(63) 60055 100.0 5-FU存活率%. 90.0. 5-FU+LV存活率%. 80.0. 60.0 50.0 40.0 30.0 20.0 10.0 0.0 0. 0.0016. 0.008. 0.04. 0.2. 1. 5. 10. 5-FU dose (mM). 圖八、5-FU 在 60055 細胞株之 IC50 的反應曲線. BT-20 100.0. survival rate (%). survival rate(%). 70.0. 90.0. 5-FU 存活率%. 80.0. 5-FU+LV 存活率%. 70.0 60.0 50.0 40.0 30.0 20.0 10.0 0.0. 0.008. 0.04. 1. 5. 5-FU dose (mM). 63. 10. 50.
(64) 圖 九、5-FU 在 BT-20 細胞株之 IC50 的反應曲線. MCF-1 100.0 90.0 5-FU存活率% 5-FU+LV存活率%. survival rate(%). 80.0 70.0 60.0 50.0 40.0 30.0 20.0 10.0 0.0 0. 0.04. 1. 2.5. 5. 10. 50. 5-FU dose(mM). 圖十、5-FU 在 MCF-1 細胞株之 IC50 的反應曲線. MCF-7. survival rate(%). 100.0 90.0. 5-FU存活率%. 80.0. 5-FU+LV存活率%. 70.0 60.0 50.0 40.0 30.0 20.0 10.0 0.0 0. 0.04. 1. 2.5. 5-FU dose (mM). 64. 5. 10. 50.
(65) 圖十一、5-FU 在 MCF-7 細胞株之 IC50 的反應曲線. T47D 100.0 90.0. 5FU存活率%. 80.0. 5-FU+LV存活率%. survival rate(%). 70.0 60.0 50.0 40.0 30.0 20.0 10.0 0.0 0. 0.0016. 0.008. 0.04. 0.2. 1. 5. 10. 5-FU dose (mM). 圖十二、5-FU 在 T47D 細胞株之 IC50 的反應曲線 表一、乳癌細胞株在不同的 5-FU 及 5-FU+LV 1.25mM 的濃度下之 IC50 的 比較. 乳癌細胞株 BC60055. 5-FU (mM). 5-FU (mM) + LV 1.25mMRatio(X). 0.042. 0.017. 2.47. BT-20. 2.54. 0.98. 2.59. MCF-1. 0.41. 0.21. 1.95. MCF-7. 0.36. 0.19. 1.89. T-47D. 0.013. 0.0021. 6.19. 65.
(66) 5-FU, 5mM , survival rate 100% 90%. 5-FU存活率. 80%. 5-FU+LV存活率. surivival rate. 70% 60% 50% 42.1%. 40% 30% 20.3%. 20%. 14.10% 9.40%. 10%. 11.20% 9.9%. 18.2% 10.1%. 0% 0%. 0% T47D. MCF-1. MCF-7. BT-20. 60055. cell lines. 圖十三、乳癌細胞株在 5-FU 濃度 5mM 時之細胞存活率的比較. 66.
(67) 圖十四、乳癌細胞株的細胞免疫染色圖;以 H630-1 為 positive 對照 組。其中 A 圖為 H630-1 細胞株,B 圖為 60055 細胞株,C 圖為 BT-20 細 胞株,D 圖為 MCF-1 細胞株,E 圖為 MCF-7 細胞株,F 圖為 T47D 細胞株 (以 上圖片放大 100 倍) 。. 67.
(68)
數據
Outline
相關文件
[r]
10 SF12259A 黃文豊 一項隨機分配、開放性、多中心臨床 試驗,對於先前未接受過治療的 CD20 表面抗原陽性之瀰漫性大型 B 細胞淋 巴瘤或 CD20 表面抗原陽性之濾泡性
SF11310 張基晟 比較 Ipilimumab 加上 Etoposide/Platinum 與 Etoposide/Platinum
S08176 王輝明 一個亞太區、非隨機、開放性、第二期臨床試驗,用以評 估讓 KRAS 基因野生型的轉移性大腸直腸癌病患使用單
評估以 S-649266 或最佳現有療法進行治療罹患抗 Carbapenem 革蘭氏陰性菌感染的患者的臨床結果 (包括詴驗用藥 S-649266
Rapiacta 因不經肝代謝,故透過 CYP 機轉與其他藥物發生 交互作用之可能性應該很低,就目前所知的排除途徑以及從 體外試驗可推知 Rapiacta 並不會誘導或抑制 CYP 450。 1)
考生只能攜帶或使用 5
• 是細胞不正常增生,且這些增生的細胞可
