國立臺灣大學獸醫專業學院獸醫學研究所 碩士論文
Graduate Institute of Veterinary Medicine School of Veterinary Medicine
National Taiwan University Master Thesis
北臺灣白肉雞場污染沙門氏桿菌之風險因子分析暨 全臺家禽場飼料之沙門氏桿菌污染調查
The identification of risk factors for Salmonella contamination in broiler farms in northern Taiwan and the survey for Salmonella
contamination of poultry feeds in Taiwan
黃凰綺
Huang-Chi Huang
指導教授:蔡向榮 博士
Advisor:Hsiang-Jung Tsai, D.V.M., Ph.D.
中華民國九十八年六月
June, 2009
致謝
本論文係承蒙恩師 蔡向榮教授之指導,此論文在您對於學生研究方向的啟發、實驗工作 的全力支持並逐字細心地審閱下,始得以順利完成;感謝恩師長久以來充份給予學習的機會 與空間,並總是不棄學生駑鈍的耐心教導、慷慨借出書本,讓我從獸醫的門外漢到現在覺得 獸醫系就是自己的第二個家,謝謝您始終以寬厚的包容和支持,讓我有機會在這學習資源豐 富的校園內能藉著持續的努力,把夢想一歩步實現。
也感謝口試委員 連ㄧ洋博士與 周崇熙博士於百忙中審閱論文,愷切指導與斧正,使本 論文更趨完整。此論文在多方的協助下俾得以順利完成,特別感謝各縣市防治所與獸醫師的 協助,以及北區家禽保健中心志工 張啟德組長、周政宏先生、羅吉寰先生、郭進益先生以及 曾繁延先生等人,各位協助最辛苦的採樣工作並不厭其煩的指導許多寶貴的禽場知識與經驗 您的協助確實是推動此研究調查最有力的那雙手。
ㄧ路走來我曾受到許多溫暖的鼓勵與協助,感謝 李ㄧ凡老師對於我人生價值觀的指引與 開導、林逸莒醫師讓我對動物的單純熱愛轉為更近一歩付出的體認、王 穎教授開啟我對野生 動物生態保育的視野、以及臺灣動物科技研究所 楊平政老師與臺大醫學院 謝松蒼教授在學 術研究與未來方向的協助;以上各位都是學生的人生導師,您豎立了良好的典範,在我人生 不同階段中所帶來的啟發與指引,對我的人生影響甚鉅,故特在此獻上最誠摯的謝意。
同時也要感謝摯友皓媛、Lillian長久以來總是給予我最懇切的建議與支持,待我如親妹 妹的媺宜、靜宜學姊,還有燕萍、麗璇學姊在學業上的鼓勵與人生經驗的分享,可愛又貼心 的學妹彤安、瑋瑛、富琪、琳晶溫馨的陪伴與實驗上的協助,以及臺大人畜共通傳染病研究 中心優秀的夥伴們:育君、志嶽、健璁、金玉,謝謝你們豐富了我實驗室裡、培養皿外的生活。
最後感謝我最摯愛的家人,我的父母親 黃錦派先生與 江阿桃女士,謝謝您以長年的辛勞撫 育我成長,並賜與我們五姊妹最溫暖的港灣;亦感謝男友帥羊俊明多年來始終以體貼與包容,
陪伴我度過無數辛苦與挫折的時刻,我愛你們!
僅以本論文之成果與喜悅奉獻予各位,願本論文能為我國家禽場污染防治盡份心力。
黃凰綺 98 年六月於台大獸醫學研究所
摘要
沙門氏桿菌是重要的人畜共通傳染病原,長久以來家禽被認為是媒介人類感染的主 要來源之ㄧ,而白肉雞是消費量最大的肉雞品種,故其飼育過程之管控相當值得關注。為暸 解北臺灣白肉雞場污染沙門氏桿菌之風險因子以作為禽場飼養管理之參考,進行以問卷調查 及沙門氏桿菌檢測為基礎的病例對照研究,自 2008 年 4 月至 2009 年 5 月間共調查 41 個白肉 雞場,包括 14 場開放式與 27 場水簾式雞場,於各場擇一雞舍調查。經前預備試驗後首度建 立適用於北臺灣地區白肉雞場風險因子調查之研究方法,以連續 3 個時間點採樣並各採集雞 隻臟器與飼料進行檢測。結果顯示場陽性率為 56.1 % ,且雞場型態對於污染之影響未達顯著 差異 (p >0.05),經分析得知有 10 個風險因子雖未達統計學顯著水準,但其勝算比值顯示可 能與減少禽場污染有關,包括: (1)水簾式雞舍之型式為一層而非多層;(2)雞場入口具有需 以鑰匙開啟之門鎖;(3)當雞隻飲水非自來水但先經處理過再使用;(4)定期清掃雞場內之犬貓 糞便;(5)工作人員每天換洗工作服與工作鞋;(6)雞場具有圍牆與外界隔離;(7)進入雞舍前有 一獨立之室內空間可供更換工作服與清潔雙手;(8)雛雞運送至雞舍時之容器非重複使用之塑 膠籠;(9)雛雞來源為單一種雞場;(10)墊料使用前先移除發霉或結塊部份。另外,臺灣北部 之氣候因子可能值得特別注意,本研究結果顯示禽場污染於涼季顯著提高 (p <0.05)。
此外,有鑑於遭污染之家禽飼料可於禽場快速地傳播病原,故進行於家禽場進行之飼料 採樣調查,並比較不同採樣點與 6 種培養基搭配組合之分離效果;檢測結果顯示沙門氏桿菌 總分離率為 7.6 % (59/778)、場陽性率 21.4 % (33/154);檢測件共涵括 16 種商業品牌,陽性檢 體集中分離自 5 品牌。統計分析結果顯示: (1)不同飼料品牌之分離率呈顯著差異;(2)採樣 點中最易取得之飼料槽檢體分離結果顯示,白肉雞飼料之分離率顯著高於其他禽種;(3)不分 採樣點統計,種雞飼料之污染顯著高於其他飼料;(4)整體而言,禽場飼料分離沙門氏桿菌之 培養基分離效果以 RVS 搭配 XLD 最佳,RVS 搭配 CAS 居次 (p <0.05)。
關鍵詞: 沙門氏桿菌、白肉雞、風險因子、勝算比、飼料
Abstract
Salmonella is an important pathogen in terms of zoonoses, poultry has been thought to be one of the major medium for human salmonellosis. Broiler chickens are one of the most commonly consumed chicken breed in Taiwan, therefore monitor it’s rearing and feeding conditions are very important. In order to alleviate the risk at commercial broiler chicken farms, a questionnaire based retrospective case-control study was conducted to identify risk factors with Salmonella contamination during the broiler rearing period. Forty-one broiler farms in northern
Taiwan were studied from April 2008 to May 2009, included 14 open-sided and 27 wet pad cooling system housing type of chicken farms. All of the flock size was larger than 10,000 and only one house was studied in each farm. Through preliminary study, methods for the investigation of the broiler farms risk factor were established. Samples were taken at three consecutive time-points, visceral organs of chickens and feeds were also collected for further analysis. The results showed a flock positive rate of 56.1 % and the housing type of farms did not have a significant effect on Salmonella contamination (p >0.05). Through data analysis, odds ratios of ten risk factors
indicated that thry may associate with the decrease in poultry farm contaminations although not statistically significant. These risk factors includes: (1) pad cooling system house which are single layered instead of multiple; (2) the entrance of the farm is locked; (3) none-tap drinking water for animals are pre-treated; (4) dog/cat feces are regularly cleaned in the farm; (5) workers outfits are changed and cleaned on a daily basis; (6) the farm is separated from the outside by a wall; (7) a
single indoor room available for changing clothes and cleaning hands before entering the chicken house; (8) The cages used for transporting day-old chick are not reusable plastic cages; (9) a single source for day-old chick; (10) molds or coagulated litters are removed before use. Moreover, the weather factor of northern Taiwan is also worth mentioning, since contamination of Salmonella in broiler farms is significantly higher in colder seasons (p <0.05) .
Since contaminated poultry feeds may aid in the process of Salmonella spreading in poultry farms, an investigation of poultry feeds were also performed. Different sampling sites and isolation treatment methods were compared. The result shows a total Salmonella isolation rate to be 7.6 % (59/778), flock positive rate was 21.4 % (33/154). Feed samples included 16 different brands, Salmonella were isolated from five of the brands. The data analysis showed (1) isolation rate of different brands are significantly different; (2) isolation rate of feed troughts from broiler house showed a significantly higher isolation rate than from other types of poultry; (3) not taking sampling site into account, breeder chicken feeds have a higher contamination rate than other types of feeds; (4) Overall, the best culturing medium for Salmonella isolation from chicken feeds is to use RVS plus XLD. (p <0.05)。
Key word: Salmonella spp., salmonellae, broiler, risk factor, odds ratio, feed, feedstuff
目錄
中文摘要 ……… I
英文摘要 ……… II
目錄 ……… IV
表次 ……… VI
圖次 ……… VIII
第一章 前言 1
第二章 文獻回顧
第一節 沙門氏桿菌之研究背景
2-1.1 沙門氏桿菌症與宿主特異性……….. 2
2-1.2 沙門氏桿菌之毒力因子與致病特性……….. 2
2-1.3 臺灣地區沙門氏桿菌之流行病學調查……….. 7
第二節 家禽污染沙門氏桿菌調查 2-2.1 沙門氏桿菌於白肉雞產業之影響與概況…….……. 10
2-2.2 家禽沙門氏桿菌流行型別之改變與趨勢………….. 10
2-2.3 各國家禽污染沙門氏桿菌之風險因子調查概況..… 12
2-2.4 其他值得注意之風險因子……….. 17
第三節 家禽飼料污染沙門氏桿菌之調查與防治方法 2-3.1 各國的流行病學調查結果………... 18
2-3.2 飼料工業的防治方法………... 21
2-3.2.1 飼料製程之危害管制點………... 21
2-3.2.2 其他應用於飼料業之防治措施………... 21
第三章 材料與方法 第一節 北臺灣白肉雞場污染沙門氏桿菌調查 3-1.1 試驗設計………... 24
3-1.2 調查與試驗方法………... 25
3-1.2.1 雞場設施與飼育條件之問卷調查………... 25
3-1.2.2 採樣時間與樣本種類………... 26
3-1.3 沙門氏桿菌分離與鑑定方法………... 27
3-1.4 統計分析方法………. 28
第二節 臺灣地區家禽場飼料污染沙門氏桿菌調查 3-2.1 試驗設計………. 29
3-2.1.1 採樣策略………. 29
3-2.1.2 飼料檢體之採樣方法………. 30
3-2.2 沙門氏桿菌之分離與鑑定………. 31
3-2.3 統計分析方法………. 31 第四章 結果
第一節 北臺灣地區白肉雞場污染沙門氏桿菌風險因子調查
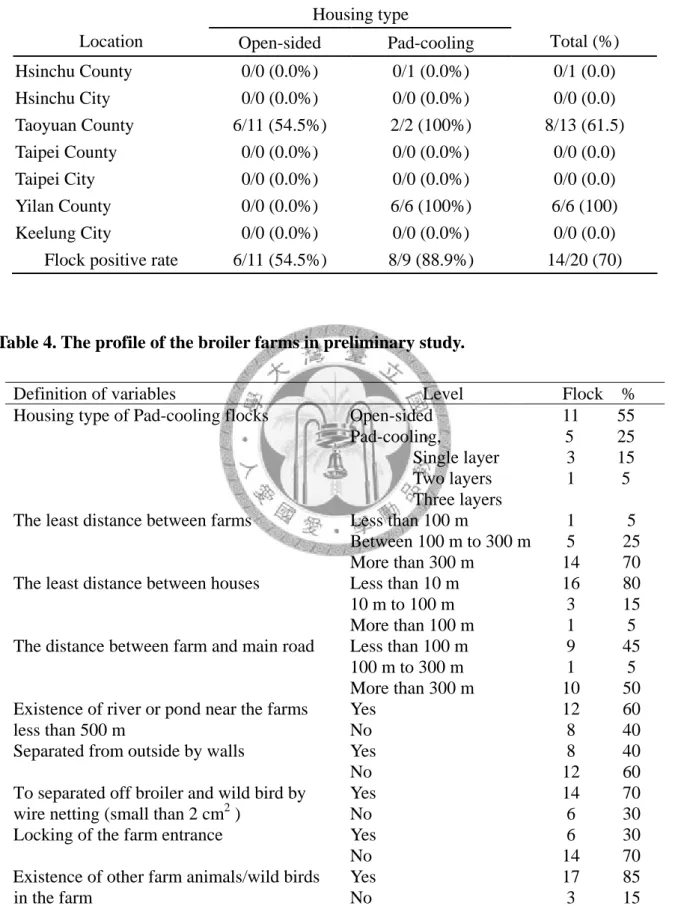
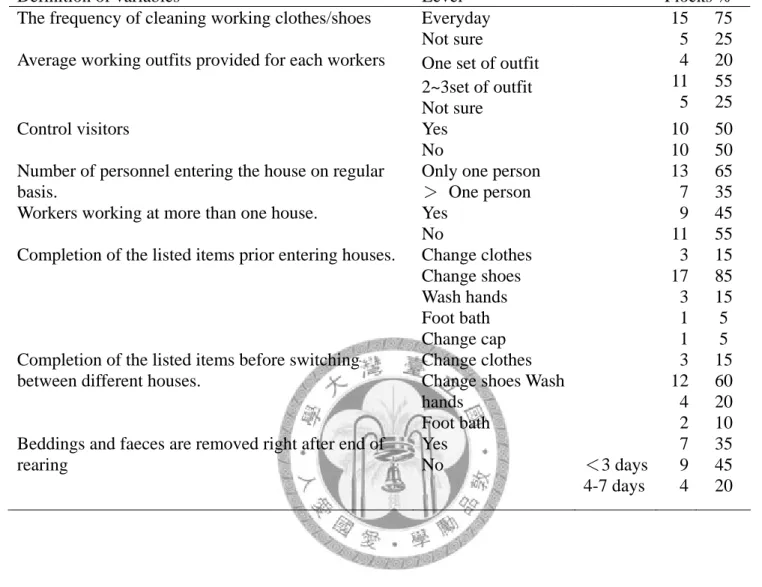
4-1.1 前預備試驗調查結果………... 32
4-1.2 北臺灣地區調查結果整合………... 34
4-1.3 北臺灣白肉雞場污染沙門氏桿菌之風險因子....…... 36
4-1.4 沙門氏桿菌污染於雞場之分佈情形.……….. 36
4-1.5 沙門氏桿菌分離株之血清型鑑定.……….. 37
第二節 臺灣地區家禽飼料污染沙門氏桿菌調查 4-2.1 前預備試驗……….……….. 38
4-2.2 調查結果整合………... 40
4-2.3 不同檢體種類與培養基分離效果之比較…………... 41
4-2.4 沙門氏桿菌分離株之血清型鑑定………... 41
第五章 討論 第一節 北臺灣地區白肉雞場污染沙門氏桿菌風險因子調查...……… 43
第二節 臺灣地區家禽飼料污染沙門氏桿菌調查…...……… 47
圖表……… 50
參考文獻……… 67
附錄一、家禽場之飼養管理與感染沙門氏桿菌之風險因子問卷……… 79
附錄二、白肉雞場污染沙門氏桿菌之風險因子分析結果……… 86
表次
Part I. The identified of risk factors for Salmonella contamination in broiler farms in northern Taiwan.
Table 1. The distribution of broiler farms with chicken flock size larger than 10,000 in northern Taiwan. Data was surveyed by Council of Agriculture, Executive Yuan, Taiwan
during 2008, 3rd season………. 49
Table 2. The distribution of flock sizes and type of housing in preliminary study……….. 49
Table 3. The distribution of salmonellae isolates from different location and housing type in preliminary study……….. 50
Table 4. The profile of the broiler farms in preliminary study………. 50
Table 5. The sanitation of equipment in preliminary study……….. 51
Table 6. The general controls on broiler farms in preliminary study………... 52
Table 7. The feeding plans in broiler farm in preliminary study……….. 53
Table 8. The salmonellae isolation rates of different sample type at various sampling time in preliminary study……….. 54
Table 9. The distribution of flock size and type of housing in the study………. 54
Table 10. The profile of the broiler farms in this study……… 55
Table 11. The hygiene apparatus and control in broiler farms………. 56
Table 12. The general controls on broiler farms………... 57
Table 13. The feeding plans in broiler farms……… 58
Table 14. Factors reducing risk for salmonellae contamination………... 59
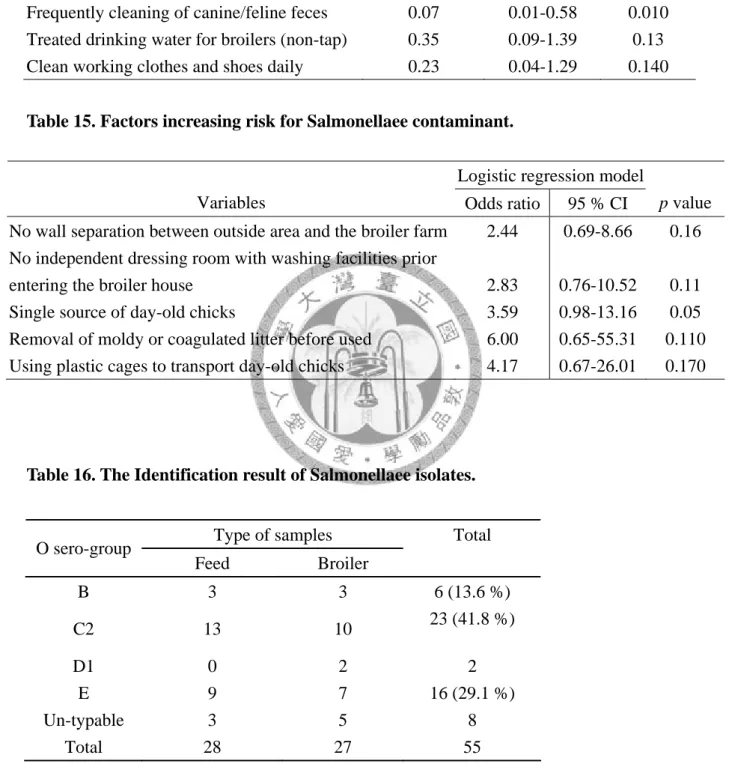
Table 15. Factors increasing risk for salmonellae contaminant………... 59
Table 16. The Identification result of salmonellae isolates……….. 59
Part II. The survey for Salmonella contamination of poultry feeds in Taiwan.
Table 17. Positive rates (%) of salmonellae from different locations and feed types and the total positive isolate rate in preliminary study.
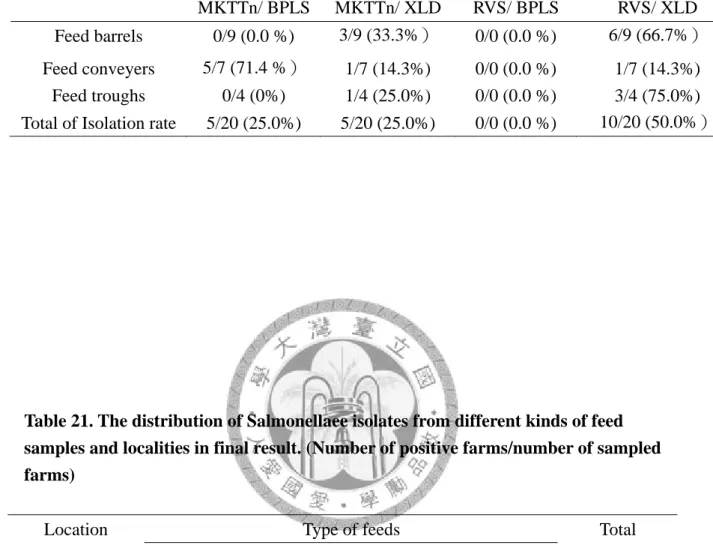
60 Table 18. Distribution analysis of sampling feed type and location in preliminary study. 60 Table 19. Salmonellae isolation rate of different feed brands in preliminary study. 61 Table 20. The effectiveness of isolation treatment in preliminary study. 62 Table 21. The distribution of Salmonellae isolates from different kinds of feed samples and
localities in final result.
62 Table 22. The frequencies of salmonellae strains isolated from different sampling sites. 63 Table 23. Identification of serotype of salmonellae strains from poultry feeds. 63
圖次
Fig 1. The experiment design flow chart of “The identification of risk factors for salmonellae contamination in broiler farms in northern Taiwan”
64 Fig 2. A flow sheet of the experiment design of the study of “the survey for
salmonellae contamination of poultry feeds in Taiwan”
65
第一章 緒言
沙門氏桿菌 (Salmonellae spp.) 為革蘭氏陰性桿菌,可依據其體抗原 (somatic antigen)、鞭毛抗原 (flagella antigen)以及莢膜抗原 (capsule antigen) 進行血清型鑑 定,目前已發現兩千五百多種血清型。沙門氏桿菌是重要的人畜共通傳染病原,除了 在世界各地人類與動物間造成疾病及經濟損失外,近年來也因其抗藥性問題越趨嚴重 而備受關注。許多研究證實在經濟動物藉由食物鏈傳播病原的途徑中,以食入受污染 的家禽類製品最常見的感染途徑。家禽是沙門氏桿菌的自然宿主,除了 S. Gallinarum 與 S. Pullorum 等少部分的血清型會造成臨床徵狀外,因感染其他種血清型時常呈現 不顯性感染,故易於禽場間被忽略,因此過去在改善禽類感染沙門氏桿菌的措施中以 屠宰場的管控等措施是最早被採納的防治重點。
研究證實沙門氏桿菌可藉由垂直傳播 (vertical transmission) 感染子代,並經常於 飼育或屠宰時藉由水平方式互相污染傳播,預防措施應全面性地串聯蛋雞、種雞,以 及肉雞之養殖層面,並藉由良好的屠宰衛生、食品衛生處理,使達成食品安全之最終 目的。飼育期間的污染防治在過去經常仰賴抗生素的使用,然而使用抗生素所衍生的 多重抗藥性菌株問題對家禽業之危害甚大;進行區域性家禽場之風險因子調查可獲得 該區域最適切之飼養管理重點以提升預防污染之效益,可減少以參考標準來決定防疫 重點時在衡量支出成本的考量下難以兩全之缺憾。
此外,近年來動物飼料的安全性在國際間逐漸獲得重視,即使部分報告指出飼料 成品污染沙門氏桿菌之現象並不嚴重,但飼料成份的污染卻是常見的,且要藉由飼料 製程完全滅除病原確實是目前尚無法克服的問題。在波蘭與英國的調查已發現飼料中 污染 S. Typhimurium DT104 等具多重抗藥性的菌株的現象 (Davies and Wray 1997;
Wasyl, Sandvang et al. 2006),遭高抗藥性菌株汙染的飼料一旦進入飼養環境,以直接 攝食與間接污染環境途徑快速散播,將對於禽場的疾病治療造成窒礙,也會對於公共 衛生將形成極大的威脅,故建立適用於飼料採樣與檢測方法以了解飼料之實際污染情 形或控制方法之效果,實為目前首要的工作。
第二章 文獻回顧
第一節 沙門氏桿菌之研究背景
2-1.1 沙門氏桿菌症與宿主特異性
無論溫血或冷血動物皆可作為沙門氏桿菌之自然宿主,動物經常呈現不顯性感染 (subclinical infection),而人類感染沙門氏桿菌症之途徑經常是透過吃下受污染的禽畜 肉品所造成;食入受污染的家禽食品被認為是人類感染最重要的途徑,根據荷蘭自 1990 年至 2006 間大規模監測調查即證實有超過 50 % 臨床病患之分離株屬於家禽源 的感染(Valkenburgh et al., 2007)。
學者曾提出會影響家禽對沙門氏桿菌感受性之因子至少包括以下八種:(1) 禽鳥 年齡;(2) 禽鳥是否受到環境、運輸或疾病所造成的緊迫;(3) 宿主基因遺傳背景;(4) 感染之血清型別與感染劑量;(5) 菌體與其他腸內菌叢的生長競爭的情況;(6) 菌體 經過胃低 pH 值環境後的存活能力;(7) 菌體是否存在於適合生長之環境;(8) 飼料的 添加物,如抗菌劑與抗球蟲藥等 (Foley, Lynne et al. 2007)。
部分血清型因僅於特定動物種別造成疾病或具有高分離率,因此被認為具有宿主 特異性之特質;像是在人類引起傷寒熱的 S. Typhi、引起副傷寒熱 ( paratyphoid fever ) 的 S. Paratyphi A、S. Schottmuelleri;分別在家禽引起雛白痢 ( pullorum disease ) 與家 禽傷寒 ( fowl typhoid ) 的S. Pullorum、S. Gallinarum ; 經常在牛隻分離到的 S.
Dublin、在猪分離到的 S. Choleraesuis 與 S. Typhisuis等 (Quinn P.J., et al., 1994)。
2-1.2 沙門氏桿菌之毒力因子與致病特性
致病機制之探討為近年來沙門氏桿菌研究的趨勢,毒力因子之研究被認為可能是 開發更適用之控制預防工具相當重要的基石。目前在沙門氏桿菌毒力因子的研究中尤
其以毒性質體(virulence plasmid)、毒力島(pathogenicity islands)與線毛(fimbriae or pili)之討論較為廣泛,以下簡介這三種毒力因子:
(1) 毒性質體(Virulence plasmid)
學者推測毒性質體與全身性感染時沙門氏桿菌大量增殖有關,目前發現具有毒性 質體之血清型包括S. Typhimurium、S. Enteritidis、S. Gallinarum、S. Pullorum、S.
Dublin、S. Choleraesuis與S. Abortusovis,但直得注意的是這些血清型的菌株並非都一 定具有毒性質體 (van Asten and van Dijk, 2005)。
(2) 毒力島(Pathogenicity Islands;pais)
是具有較大分子量的染色體DNA片段,亦稱疾病島;此DNA片段經常位於細菌 染色體的tRNA基因點內或附近,或位於與噬菌體整合有關的位點。毒力島的DNA片 度所含的G+C composition往往明顯的高於或低於宿主細胞,且其基因產物多屬於分 泌性蛋白與細胞表面蛋白,像是溶血素、血紅素結合因子等 (Amavisit, Lightfoot et al.
2003)。
(3) 線毛(fimbriae;pili)
線毛存在於大多數的革蘭氏陰性菌於及少數的革蘭氏陽性菌,大小介於7.5-10 nm x 0.5-6 nm之間。線毛的主體蛋白稱為fimbrins,而依功能性可以將線毛分為體毛 ( short attachment pili )與性毛 ( long conjugation pili ; F pili );體毛在細菌外圍呈現細 短毛狀,數量明顯多於性毛,負責協助細菌黏附於宿主的上皮細胞 ; 而性毛則具有 形狀較長的珠鍊狀外觀,負責進行菌體間的接合生殖。沙門氏桿菌的線毛大小為2-8 nm x 0.5-10 nm,利用線毛分子具有免疫原之特質製備疫苗之可行性受到關注 ( Jones and Bartlett, 2001 )。
不同血清型間毒力因子之研究
不同的毒力因子在不同血清型間所扮演的角色可能具有差異,以下針對近年來在 家禽與人類間特別受到重視或新興起的血清型:S. Typhimurium、S. Enteritidis、S.
Heidelberg、S. Montevideo、S. Schwarzengrund簡介其毒力因子之研究:
(a) Salmonella Typhimurium (ST)
S. Typhimurium是目前最被重視的血清型之ㄧ,不僅為最常造成人類沙門氏桿菌 症的血清型,在家禽也經常是分離率最高的血清型 (CDC, 2006a)。在美國FoodNet一 項長達四年的回顧性調查發現因感染沙門氏桿菌症死亡的病患中近半數都是感染此 血清型,故學者推測 S. Typhimurium不僅具較其他種血清型更優勢的生長能力外,於 致病性亦有特殊之處 (Kennedy, Villar et al. 2004)。目前已發現 S. Typhimurium含有多 重毒力因子,包括毒性質體與毒力島,這些毒力因子被認為與 S. Typhimurium 能夠 有效的侵犯並在腸道增殖有關 (Foley and Lynne 2007)。
約有88 %的 S. Typhimurium具有一個大小約60 MDa的質體,且當菌體為粗糙型 (roughly form) 時此質體大小為90 kb。此質體被賦予許多名稱,像是pSLT、MP10、
pRQ28、pSTV、cryptic plasmid以及毒性質體(virulence plasmid),其中以毒性質體 為最廣泛使用的名稱。多數學者推測毒性質體與全身性感染時細菌大量增殖有關,而 此機制也被認為與spv基因有關; spv基因可轉譯出約8 kb長度的毒性質體,並且使S.
Typhimurium可於腸道以外的臟器,尤其肝、脾進行增殖。(Ahmer, Tran et al. 1999)。
毒力島(pathogenicity islands)是S. Typhimurium另一個備受關注的毒力因子。目 前在S. Typhimurium所發現的毒力島至少有五種,包含SPI-1、SPI-2、SPI-3、SPI-4與 SPI-5。SPI-1為大小約40 kb的chromosomal locus,為沙門氏桿菌侵略宿主上皮細胞所 必須,且可誘導巨噬細胞的細胞凋零 ( apoptosis ) (Collazo and Galan 1997) ; SPI-2 鄰近 tRNAval gene,跟沙門氏桿菌於巨噬細胞中增殖、造成全身性感染有關 (Shea, Hensel et al. 1996) ; SPI-3包含十個基因,是沙門氏桿菌在巨噬細胞內存活與適應低鎂 離子環境所必須;SPI-4包含18個基因,被懷疑是沙門氏桿菌存活於巨噬細胞所必須
(Blanc-Potard, Solomon et al. 1999);SPI-5包含了六個基因,其中四個基因被認為與造 成小牛的腸炎有關 (Wood, Jones et al. 1998)。
目前毒力島的相關研究以SPI-1與SPI-2較多,學者也發現了這兩種毒力島間可作 用之宿主生理環境於不同沙門氏桿菌生長期間有極大差異,例如SPI-1易於低氧、高 滲透壓環境下且沙門氏桿菌處於指數生長期 (log growth phase)之早期時,呈現活躍 表現的狀態;SPI-2則較為活躍在缺乏磷酸、低鎂離子或低鈣離子的環境且沙門氏桿 菌處於指數生長期(logarithmic growth phase)之後期至平原期間(stationary growth phase)。
對應以上環境條件,研究亦發現當同時帶有SPI-1與SPI-2的沙門氏桿菌處於宿主 的不同生理條件下時基因表現量呈現差異,例如在低氧與高滲透壓的腸道時,SPI-1 的基因表現量遠大於SPI-2;而在鎂離子與磷酸量低的宿主細胞內,SPI-2的基因表現 量則遠大於SPI-1 (Hansen-Wester and Hensel 2001) 。
(b) Salmonella Enteritidis (SE)
在歐洲,S. Enteritidis是最常造成人類沙門氏桿菌症的血清型,於美國則僅次於S.
Typhimurium位居第二 (CDC.2006a; (de Jong and Ekdahl 2006)。在英國,S. Enteritidis 盛行率的消長有明顯的改變,在1981年造成10,000件病例,佔總沙門氏桿菌症的10
%;而到了1997年時則造成33,000件病例,佔總沙門氏桿菌症的70 % (Cogan and Humphrey 2003)。
S. Enteritidis因具有多種不同的毒力因子而影響其致病性甚巨,較特別的是利用多 種不同的線毛(fimbriae)來攻擊宿主細胞。目前在沙門氏桿菌所發現的線毛至少有 12種以上,而在S. Enteritidis已發現的線毛種類至少包括SEF14、SEF17、SEF18、
SEF21,以及由 lpf 基因所調控的long polar fimbriae與由pef 基因調控之plasmid encoded fimbriae等。以SEF14為例,線毛的命名方法為SEF代表Salmonellae Enteritidis Fimbriae的縮寫,而其後的數字則代表fimbrin monomer的長度,14代表14000 Mr (Muller, Collinson et al. 1991)。SEF14型的線毛只存在於沙門氏桿菌的體抗原D1血清
群,對於黏附在宿主的組織細胞與生殖道極為重要 (Turcotte and Woodward 1993)。
(c) Salmonella Heidelberg
S. Heidelberg是近年美國人類臨床分離率第四位的血清型,在猪隻與家禽的分離 率皆為前三名(CDC, 2006)。在過去此血清型幾乎都是從雞胸肉與碎火雞肉分離到 (Zhao, McDermott et al. 2006)。
目前美國每年平均會有84,000名感染 S. Heidelberg的病例發生,其中約有7 %的 病患因此死亡,其致死率僅次於 S. Typhimurium (Kennedy, Villar et al. 2004)。學者認 為該血清型傳染到人類時毒性有增強的情形,且人類感染時經常是以集體感染的方式 發生,像美國從1973年到2001年間發生的沙門氏桿菌症疫情共2,260件,其中感染該血 清型的案件共184件(8%),且污染源通常是家禽製品與蛋品 (Chittick, Sulka et al.
2006)。
長年來的監控調查發現在1990年代中期 S. Enteritidis盛行率減少的同時 S.
Heidelberg卻有明顯增加的現象,故學者推論這兩個血清型間具有相似的致病機制,
並且都具有感染家禽的生殖系統並侵入雞蛋的侵略性 (Gast, Guard-Bouldin et al.
2005)。
(d) Salmonella Montevideo
S. Montevideo屬於體抗原C1血清群,在過去經常是分離自冷盤食品、牛肉與家 禽,在食品製程中經常藉由交叉污染散播,有一些疫情的爆發則被發現於芝麻、巧克 力與蕃茄等 (Hedberg, Angulo et al. 1999) ; 此外,S. Montevideo 也被發現具有可侵入 蛋黃的能力,因此對於蛋品污染的可能性亦應有所防範 (Murase, Fujimoto et al.
2006)。此血清型也在幾次突發的疫情發現是藉由接觸禽鳥動物感染,尤其在復活節 時與幼雞幼鴨的接觸;為了預防類似的疫情一再發生,美國部分州郡甚至立法限制購 買鳥禽數量並公開勸導民眾不要飼養這類寵物 (2007) 。
(e) Salmonella Schwarzengrund
美國自1996至1999年間FoodNet調查顯示該血清型被認為是繼 S. Typhi、S.
Dublin、S. Paratyphi A 與 S. Choleraesuis之後,名列第五大具侵犯性的血清型 (Vugia, Samuel et al. 2004)。該血清型在動物主要分離自禽類與猪隻 (Limawongpranee, Hayashidani et al. 1999; Vugia, Samuel et al. 2004; Chen, Wang et al. 2006),而在家禽的 分離率一般約5至10 % 之間 (Poppe, Irwin et al. 1991; Poppe, Irwin et al. 1991);而無論 在經濟動物、食品甚至是病患臨床分離株都有嚴重的多重抗藥性問題 (Zhao,
McDermott et al. 2006; Aarestrup, Hendriksen et al. 2007)。於臺灣病患之沙門氏桿菌分 離株中佔了7.5 % (60/798),且對於nalidixic acid與ciprofloxacin等fluoroquinolone類藥 物具高抗藥性趨勢 (Lauderdale, Aarestrup et al. 2006)。
2-1.3 臺灣地區沙門氏桿菌之流行病學調查
(一) 經濟動物
食入受污染的禽畜製品是人類感染沙門氏桿菌最常見的途徑,基於血清型別間具 有宿主特異性 (host specific)、飼育與食品製造過程中可能發生連續性的交叉污染等因 素,分離株之血清型常呈現複雜且相異的分佈情形,像是在白肉雞最常見的血清型是 S. Albany、S. Enteritidis、S. Schwarzengrund,自蛋雞分離的血清型則以 S. Enteritidis 為最多 (蔡,2007),鴨隻以 S. Potsdam、S. Dusseldorf、S. Indiana 最常見 (Tsai and Hsiang 2005);仿土雞於屠宰場是以 S. Schwarzengrund 為主,而在傳統市場分離到的血清型 則以 S. Albany 為主 (郭,2006)。
前人研究顯示健康猪隻分離株最常見的血清型是 S. London、S. Panama、S.
Ttyphimurium (Hummel, Su et al. 1978),而猪隻屠體最常見的血清型是 S. Derby、S.
Anatum、S. Typhimurium、S. Schwarzengrund (Chen, Wang et al. 2006),而自猪場糞便、
堆肥場半成品 (未完成發酵) 及成品 (完成發酵) 堆肥、超市和傳統市場的猪肝分離 最常見的血清型則是 S. Derby、S. Agona 和 S. Albany (李,2006)。
於屠宰場之調查結果顯示具有宿主特異性之血清型其實不常自屠宰場分離,例 如 2000 至 2003 年間於臺灣各地區猪、雞、牛屠宰場的調查即發現 S. Choleraesuis 實 際在猪隻屠體分離株中僅佔少數,而可藉蛋傳播之 S. Enteritidis 則未自雞隻屠體分離 到,而於牛隻可能為潛在帶原或造成流產、仔牛腦膜炎、骨髓炎 (osteomyelitis) 之 S.
Dublin 亦未分離到,且在牛隻屠宰場連續四年的監控中也沒有分離到任何沙門氏桿 菌;整體而言於屠宰場最常分離到之血清型依序為 S. Derby、S. Anatum、S.
Schwarzengrund、S. Typhimurium,其中 S. Derby 及 S. Anatum 這兩種血清型只有在 猪隻屠體被分離到 (王,2004);以上結果說明操作與環境之交叉汙染源實為屠宰線重 要的危害管制點。
(二) 伴侶動物
而隨著社會結構老年化且少子化的趨勢與動物福利觀念的進步,人們對於伴侶動 物的需求與互動皆有增加的趨勢。犬貓動物與爬蟲類是沙門氏桿菌常見的帶原者,易 藉由與畜主共用生活環境或親密接觸而傳播病原並污染居家環境,實為極易受忽略的 污染源,因此美國疾病管制局已明確的指出並教育宣導伴侶動物為人類感染沙門氏桿 菌等人畜共通傳染病原的主要來源之ㄧ。近年來在臺灣地區針對家犬與流浪犬的沙門 氏桿菌分離調查發現在不同地區具有相異的血清型別分佈;北部最常見之血清型為 S.
Duesseldorf、S. Enteritidis、S. Derby (Tsai, Huang et al. 2007)、中部以 S. Newport、S.
Enteritidis 及 S. Senftenberg 為主 (廖,2007),而南部則以 S. Albany 及 S. Newport 居 多 (陳,2007)。
爬蟲類寵物分離株中有 43.9 %屬於人類常見感染之血清群(A、B、C、D、E), 並以血清群 B 最多;以動物種類區分則分離率以蛇類最高、蜥蜴類次之,龜鱉類檢出
率則顯著較低 (陳,2005)。
(三) 人類臨床分離株調查
早期於 1978-1987 年間的調查發現自臨床病患分離到的血清型多達 43 種,其中 最常見的血清型是 S. Typhimurium (43.7%),其次依序為 S. Muenchen、 S. Panama、
S. Krefeld、S. Bovismorbificans、S. Derby、S. Aanatum、S. Braenderup 等 (Peng 1992);
而近年來由國家衛生研究院所執行的全國微生物抗藥性監測計畫 (Taiwan
Surveillance of Antimicrobial Resistance; TSAR) 分別 於 1998 年、2000 年、2002 年間 從北中南東四個地區的醫學中心及區域醫院收集住院病人及門急診病人所分離出之 菌株,則發現最常分離到的血清型依序為 S. Typhimurium、S. Enteritidis、S. Stanley、
S. Schwarzengrund、S. Newport、S. Albany,其中 S. Typhimurium 與 S. Enteritidis 約占 所有分離株 42 % (Lauderdale, Aarestrup et al. 2006),略低於 2004 至 2006 年間於南部 地區的調查中的 55.8 % (陳,2007)。另外也發現近年來病患分離株對 fluoroquinolone (FQ)藥物具有高抗藥性的趨勢,尤其以 S. Choleraesuis 與 S. Schwarzengrund 這兩種血 清型最為常見 (Lauderdale, Aarestrup et al. 2006)。
第二節 白肉雞污染沙門氏桿菌調查
2-2.1 沙門氏桿菌於白肉雞產業之影響與概況
白肉雞由於具有生長快速、高換肉率且飼養期短的優點,是最普遍飼養的肉用雞 種,在世界各地皆有相當可觀的消費量,故其沙門氏桿菌污染問題相當值得重視。一 般而言白肉雞產業的生產流程經由以不同階段分層進行,首先自白肉雞種雞場配種產 蛋後,將受精卵送至孵化場 (hatchery),待孵育出小雞後於當天將一日齡雛雞分送至 各白肉雞場飼育,三到五週齡為雞隻快速生長,飼育期一般約38至42天即達可販售之 成熟度,再加上出雞後需有約兩週時間進行清潔消毒與淨空,故平均每批雞的生產包 括各批次間的清潔淨空期共約八週為一循環。
家禽飼養生產過程中各管控點就像一條條同源的支流般彼此牽動,一旦其中發生 污染,極易快速的水平傳播至整個雞場,再進一步污染增加屠宰線的污染機會,因此 家禽污染沙門氏桿菌的問題正如同每年難以估計的家禽產品消費量般,持續的存在於 各國的家禽產業。
白肉雞場之沙門氏桿菌盛行率在不同的國家具有相當的差異,例如泰國曾報告有 100 % (Sasipreeyajan et al., 1996)、加拿大 76.9% (Chambers et al., 1998)、法國 68.9 % (Rose et al., 1999)、丹麥 25.0 % (Chadfield et al., 2001)、巴西 21.7 % (Tavechio et al.,2002)、芬蘭 0.7 % (Anon., 1998);不同國家間盛行率有如此大的差異性可能跟不同 的衛生條件與設備、飼養管理方式等,以及進行調查時的採檢與檢測方法之差異有關。
2-2.2 家禽沙門氏桿菌流行型別之改變與趨勢
雖然沙門氏桿菌有高達二千多種血清型,但是相對而言在臨床病例中絕大多數的 血清型極少被分離到,僅少部分優勢的常見於流行病學調查中。國際間經常藉由長期 觀察血清型分佈趨勢來瞭解並評估相關控制預防措施的有效性,某些血清型可感染的
宿主範圍非常廣泛或具有重疊的流行性,故過去也經常以動物污染之血清型分佈作為 發生少見於人類之血清型疫情且尚未進行親源性分析時初步之來源推論 (Foley and Lynne 2007) 。
以美國為例,1990 年代初期 Salmonellae Pullorum 為美國家禽場最常見的血清 型,並因此引起的疾病造成極大的經濟損失,然而在美國政府自 1935 年開始施行 National Poultry Improvement Plan (NPIP) 制度後,1960 年代後 S. Pullorum 之盛行率 已明顯下降,顯示其污染獲得相的控制;然而同時 S. Enteritidis 盛行率則有明顯上升 的趨勢 (Shivaprasad, 2003; USDA, 1997)。此現象促使學者思考這兩個血清型別間交 替盛行之原由,因為在 1960 年前 S. Enteritidis 是很少自雞隻分離的。
由於這兩個血清型皆屬於體抗原 D1 血清群,故細胞壁具有相似的脂多醣構造,
學者認為基於類似構造的生態學基礎而導致兩血清型於家禽間生長競爭 (Gupta, Maiden et al. 1996; Velge, Cloeckaert et al. 2005),因此當 S. Pullorum 在家禽被大量抑制 後 S. Enteritidis 便成為優勢血清型,因而取代 S. Pullorum 而有高盛行率的現象 (Rabsch, Hargis et al. 2000),成為當時歐美地區最常見的血清型 (Velge et al., 2005)
而到了 1990 年代中期,美國的 S. Enteritidis 盛行率出現減少的趨勢,尤其從 1995 至 2004 年這十年間其盛行率減少了 50%;學者認為這可能與美國自 1989 年與 1994 年分別在 NPIP 中開始針對該血清型於蛋品與肉品的預防管制措施得到相當成效有關;
而在英國,該血清型的減少則被認為跟蛋雞施打 S. Enteritidis 疫苗有關 (Baumler, Hargis et al. 2000; Cogan and Humphrey 2003)。
當美國 S. Enteritidis 的污染趨緩後,S. Heidelberg 出現新興流行趨勢,且 S.
Heidelberg 與 S. Enteritidis 一樣可於家禽生殖系統增殖,並具有侵入蛋黃的能力 (Gast, Guard-Bouldin et al. 2004),而在之後的流行病學調查也顯示這兩個血清型之消長情形 亦呈現擷抗情形 (Foley, Lynne et al. 2007)。
雖然目前已知的血清型高達2500種以上,但是不同的血清型在不同動物間的盛行 率與致病性有很大的差異,血清型別流行性的起伏消長的資訊與其中演化的因果關係 尚更進一步的研究。
於白肉雞所流行的血清型於不同的國家不盡相同。荷蘭 2001 至 2006 年間最常見 的血清型是 S. Java (31~51.7 %)與 S. Infantis(13~16.4 %)(Valkenburgh et al., 2007);
在韓國白肉雞最常分離的血清型是 S. blockley、S. Seftenberg (Cheong et al., 2007),而 近年來 S. Enteritis 與 S. Typhimurium 之白肉雞分離株被確認與沙門氏桿菌症病患之分 離株有相似的脈衝式電泳型別表現 ( PFGE profile ),因而認為該國白肉雞為人類感染 這兩個血清型最主要的污染源 (Kim et al., 2007)。
而在參考各國文獻後發現近年來 S. Infantis 逐漸受到重視,此血清型是荷蘭第二 常於白肉雞分離之血清型 (Valkenburgh et al., 2007),日本自 1990 年後分離率快速增 加,所造成之人類沙門氏桿菌症於 2004 至 2005 年間僅次於 SE (Asai et al., 2007)。而 在匈牙利也發現,自從開始使用 ST 與 SE 的疫苗後,S. Infantis 不僅成為家禽最常見 的血清型,同時也成為第三種最常感染人類的血清型,在近年更衍生了多重抗藥性菌 株的問題 (Nogrady et al., 2007)。
2-2.3 各國白肉雞場污染沙門氏桿菌之風險因子調查概況
家禽場的污染防禦策略需建立在飼養管理與衛生條件的基礎上,然而不同國家、
地區,甚至在各個雞場間的生產條件都有相當程度的差異,因此針對各別區域進行白 肉雞場沙門氏桿菌污染之風險因子調查是必須的。下面將分別以丹麥與塞內加爾進行 之白肉雞場沙門氏桿菌污染風險因子調查為例,介紹風險因子之研究方法與調查結 果,並進一步介紹其他重要但常被忽略的風險因子,以及在白肉雞較具重要性的沙門 氏桿菌血清型。
(一) 丹麥白肉雞場之風險因子調查
自 1990 年代起,丹麥感染沙門氏桿菌症的患者有明顯增加的趨勢,且被認為與 家禽及家禽製品有極大關連性。丹麥的家禽製程是非常明顯的分層作業,每個層級皆
以非常嚴格的控管以避免沙門氏桿菌水平傳播污染的發生,例如丹麥家禽議會
(Danish Poultry Council)自 1989 年開始推動一個鼓勵雞農減少沙門氏桿菌在肉雞污 染的措施,並分別對於種雞、孵化場、肉雞以及屠宰場與飼料廠進行沙門氏桿菌的監 測。而在白肉雞的生產方面,雖然在該國境內有兩間種雞場,但大多直接自英國進口 一日齡小雞來飼養。普遍來說平均每個雞場約飼養三萬隻雞,每個雞舍每年約育成六 至七批雞,境內共有十一個白肉雞屠宰場,平均屠宰日齡為 37 天 (Angen et al., 1996)。
調查方法為自 1992 年二月至 1993 年十二月間共採樣 5921 場,各場相關飼育資 料採集自丹麥家禽議會。以盛行率 20 %以上,有 95 %的機會可採集到一個陽性檢體 的條件下,於每個雞場逢機採集 16 個三週齡的白肉雞盲腸扁桃結(caecal tonsils)檢 體,每兩個檢體混合在一起,故每場共有八個檢體進行沙門氏桿菌檢測。值得注意的 是風險因子調查之採樣方法因應學者的研究結果而有了調整與改變,像是在 1994 年 後改以採取每場 60 個糞便檢體,且每 12 個混合成一份檢測單位的方式來進行沙門氏 桿菌分離,學者認為這樣的採樣較簡單且可提高檢測的敏感度;而後又在 1997 年改以 採取雞農在雞場內走動後的鞋套 60 份,同樣的將每 12 雙鞋套混合成一個檢體的方式 進行檢測,根據前人的研究報告顯示此採檢方法的敏感度與採糞便相當,但是更為方 便 (Skov et al., 1999; Angen et al., 1996)。
調查結果發現在丹麥肉雞場,有四個風險因子對於感染沙門氏桿菌的影響性皆達 統計學意義 ( p < 0.05)。這四個風險因子分別是飼料廠、前批雞隻是否感染沙門氏 桿菌、季節性、每個雞場內的雞舍數量,下面詳加解釋各因子之統計結果:
(1) 飼料廠的規模大小對於飼料之加熱處理是否確實與其所造成的交叉感染機會 有相當的影響,生產量越大的飼料廠越容易發生有上述的問題,因此有較高 的污染風險。
(2) 前批雞隻若感染沙門氏桿菌,將容易污染飼育環境而造成下批雞隻的感染,
此部分同時與清潔消毒步驟與淨空是否確實,有顯著的交互作用。
(3) 季節性在丹麥是相當重要的一個風險因子,尤其在高濕氣並寒冷的秋天(九 至十一月),且該因子與飼料廠及雞場內的雞舍數也具有交互作用。濕氣與低
溫將影響雞舍在清潔、消毒與乾燥時的效果,而在飼料廠則是對於飼料加熱 的溫度與溼度,以及飼料儲存時的品質有直接的影響。而在雞舍數量較多的 雞場,也可能因此增加清潔消毒工作之困難性與交叉感染的可能性,增加了 污染的機會。
(4) 雞場內雞舍的數目,尤其是雞舍數大於三個的雞場,將會增加其雞舍間的互 相污染的機率。這是由於較大的雞舍數量也可能會導致工作人員的工作量較 大,而在進行清潔與消毒時容易發生工作時間被縮短或效果打折的情形。而 在飼養上,多雞舍數也較難做到統進統出的飼育方式,而增加了污染沙門氏 桿菌的機會 (Angen et al., 1996)。
(二) SE8 與 ST66 於丹麥白肉雞場之風險因子調查
有別於以上所進行的一般性風險因子調查,下面介紹一個當發生特定型別之菌株 爆發群體感染疫情時,為釐清並快速尋求現場有效減少互相傳播污染之飼養條件與方 法時,所進行的風險因子分析調查 (Gradel & Rattenborg, 2003)。
此案例發生於 1997 年,在丹麥發現有 42 個白肉雞場的 78 個雞舍感染了 S.
Enteritidis phage type 8 (SE8)或 S. Typhimurium phage type 66 (ST66),而 78 個雞 舍中,有 38 個雞舍感染 SE8、34 個感染 ST66,屬於單一感染,另外 6 個雞舍為 SE8 合併 ST66 之多重感染。
由於這兩種型別之沙門氏桿菌株以往並不曾於丹麥的白肉雞分離到過,且在該國 境內的白肉雞場從未有如此高的沙門氏桿菌感染情形,為了快速釐清可能的污染源並 找到最好的現場控制病原散播方法,故針對這些感染場進行風險因子調查。在這個案 例中,以病例對照研究(case control study,追溯性研究 retrospective field study)進行 調查,除了參考丹麥家禽議會(Danish Poultry Council)所建立的基本雞場資料外,
也另外設計問卷請雞場人員填寫,以期能更詳盡的瞭解這些雞舍的管理作業方式,並 於這些感染的雞場在雞隻三週齡時,採取鞋套檢體來進行檢測。
經過風險因子的調查與分析後,首先發現這些爆發疫情的雞場之雛雞皆來自同一
個孵卵場,而風險分析的調查認為下面的風險因子在這些感染場中,對於病原的傳播 扮演了高度的重要性:
(1) 進入雞舍前使用清潔劑與水清潔雙手後才進入雞舍,將可有效減少沙門氏桿菌間 接污染的情形。
(2) 處理死掉的雞隻時,以加蓋的容器覆載並自雞舍移除,可減少病原滋生傳播。
(3) 在雞舍四周圍鋪上細碎石頭(gravel),可減少病媒傳播。
(4) 經常性的確認室內的捕鼠餌置放處(rodent-bait depots)是否抓到老鼠。
(三) 塞內加爾(Senegal)白肉雞場之風險因子調查
塞內加爾位於西非,並如同一般的非洲國家,境內有相當多的愛滋病患者。由於 愛滋病患者免疫力缺失將導致在感染沙門氏桿菌時容易發生嚴重的菌血症致死,故感 染沙門氏桿菌在該區域將導致遠較於一般已開發國家嚴重的後果。
本研究由法國學者赴當地進行,於 2000 年 1 月至 2001 年 12 月間共調查 70 個白 肉雞場。在進行調查前會先拜訪雞農,解釋此調查之進行方法與目的,在得到雞農自 願性的允諾配合後使得以開始進行調查,並先擬定問卷調查內容,請 15 位雞農試寫 並提供意見,最後完成兩份共包含 80 個問題,預計可於 30 分鐘內完成之問卷。問卷 設計包含以下七大主題:(1) 雞場與雞舍間之差異性;(2) 一日齡雛雞之相關背景資 料;(3) 病雞與死雞的處理方式、管理措施;(4) 老鼠與畜養之寵物的管理控制;(5) 飼料與飲水的來源與管理;(6) 雞場工作人員與訪客之管理;(7) 清潔與消毒之程序。
此調查進行期間共拜訪每個調查場四次,時間點分別為:(1) 先選定一場次的雞舍 後,在該場次之前一批雞已養成且即將送至屠宰場之前(previous flock);(2) 前批雞已 出雞,且雞舍完成清潔消毒程序後;(3) 新一批雞入雛當天(day-old chicks);(4) 新一 批雞於飼養之尾聲,約第 32 至 40 日齡時(end of rearing period)。於第二次與第四次拜 訪時進行問卷調查,並於第一、三、四次拜訪時進行採樣。採樣方法乃是採取每個檢測 場內新鮮的排遺,將五個檢體混合成一份,共採集 60 份檢體。此檢測數目乃是根據當盛 行率大於 5 % 時,在 95 % 可信度水準下至少能檢出一個陽性檢體所需的檢體數。
調查結果發現大多數的經營者將雞場當成副業來經營,且這些雞場之雞舍型態皆 為開放式的(open-side house)無風扇等通風設備,最常使用的墊料是木屑,使用自 歐洲或巴西進口包括玉米、花生餅 (peanut cake)等飼料成份後於該國境內製造的飼 料。而在醫療行為上,普遍常規性地施打新城病(Newcastle disease)與傳染性華氏 囊病 ( Gumboro disease )疫苗,並治療球蟲。
沙門氏桿菌的分離結果為場陽性率 28.6%,最常分離到的血清型為 S. Hadar 與 S. Brancaster。而這兩個在白肉雞場最常見的血清型在該國後續的調查中也被確認了 白肉雞分離株與人類沙門氏症病患分離株具有相似的脈衝式電泳分型結果 (Cardinale et al., 2005)。而在風險因子分析的部分,共發現了四種會顯著增加感染沙門氏桿菌風 險的因子,分別是:
(1) 當前一批雞隻感染沙門氏桿菌時,下一批雞感染的機率顯著提高。此部分與前批 雞隻屠宰前,進下批雞之前的環境清潔與消毒工作具有相當的交互作用
(interaction)。
(2) 當一日齡雛雞感染沙門氏桿菌時,會增加該場感染的風險;當雞舍內有感染雞隻的 存在時容易污染環境,藉由水平傳播快速散播病原。
(3) 雞場有頻繁的訪客時將增加感染的風險,除了訪客可能攜帶外來的沙門氏桿菌入 場外,其衛生管理往往也較難以控制。
(4) 若病雞未移除至雞場外,易於雞場內形成傳染窩散佈病原,增加污染風險。
(5) 本調查認為於一日齡雛雞使用抗生素以及在雞場中以消毒劑進行清潔,將可減少 賽內加爾之雞場內沙門氏桿菌污染風險(Cardinale, et al., 2004)。
2-2.4 其他值得注意之風險因子
由於風險因子的調查需要對於該產業之製程有相當的瞭解,方得以在各個細微的 過程步驟中詳列舉可能影響的因子,再藉由病原的檢測結果配合統計分析的方式來確 認該因子對於所調查的對象或群體的感染是否具有顯著的影響,因此在參考相當的文 獻後,在此特地列舉兩個容易被忽略但卻可能重要的因子:輸送一日齡雛雞之運輸木 箱及野鳥。
輸送一日齡雛雞之木箱由於成本考量而經常重複使用,故易於不同批雛雞間造成 交叉污染。研究也發現即使是非頻繁的重複使用輸送木箱仍有藉此途徑傳播沙門氏桿 菌與彎曲桿菌之可能性,因此當基於成本考量而必須重複使用木箱時,建議在使用前 以消毒劑徹底清潔 (Slader et al., 2002)。
前人調查雞場附近的野鳥帶有病原菌的情形,結果發現沙門氏桿菌在排遺檢測出 之陽性率於不同雞場間約 0~33 %之間 (Craven et al., 2000),因此雞場附近的野鳥被認 為有污染環境、造成雞群感染的可能性。因此雞舍應避免與野鳥接觸,可以適當孔徑 之鐵網隔離野鳥與家禽,避免暴露於交互傳染的風險中。
第三節 家禽飼料污染沙門氏桿菌之調查與防治方法
2-3.1 各國的流行病學調查
(一) 瑞典
在瑞典,無論是動物或飼料中一旦檢出沙門氏桿菌,皆必須送往國家檢測單位進 行血清型鑑定與抗生素感受性試驗,且 S. Typhimurium與S. Enteritidis皆必須進一步進 行噬菌體分型 ( phage type)。
並且自1991年起對於飼料場進行以HACCP為基礎的風險因子管控作業,其重要 管控點主要在於環境以及製程中所產生的灰塵與碎屑,採樣方法為每週於各飼料廠最 少採取五個樣品進行檢驗,若同一地點單位在多次採樣皆分離到相同血清型及噬菌體 型別,則僅採計第一次分離的結果。
瑞典農業部在1993至1997年間飼料廠對於四大種類的檢體,包括不同的飼料成份
(包含植物性、動物性及未分類)、飼料成品、在飼料廠收集到的灰塵及碎屑及難以 分類的飼料廠檢體來進行採檢,共分離到957株沙門氏桿菌。檢測結果發現動物性原 料共分離到28株沙門氏桿菌,植物性原料則有194株,顯示該國植物性原料污染情形 較動物源嚴重。最常見的血清型是 S. Senftenberg、S. Mbandaka、S. Agona、S. Anatum 與 S. Cubana,而最常受污染的植物性原料種類則為大豆粉、玉米及油菜籽。此外,
在飼料廠收集到的灰塵及碎屑共分離到464株沙門氏桿菌(48.3 %, 464/957)為四類檢 體中分離率最高的檢體種類 (Boqvist, Hansson et al. 2003),顯示飼料廠應重視廠內環 境粉塵之清除是否確實,並盡量減少製程中所發生的粉塵量。
另外值得注意的是在進行調查的這段期間最常從動物性原料分離到的血清型S.
Livingstone (9.8 %) 不僅是首度於飼料分離到,且此血清型於1994年間經常可從該國 蛋雞(59.3 %)分離到,藉由PFGE ( pulse- field gel electrophoresis ) 分型結果,有學 者懷疑是藉由飼料污染雞隻的實例 (Eriksson, Lofstrom et al. 2005)。
分離結果並顯示一些較常從同時期瑞典境內動物分離到的血清型,像是 S.
Typhimurium、S. Enteritidis 與 S. Dublin 等則並不常從飼料中被分離到,因此學者認
為瑞典境內動物所發生的沙門氏桿菌污染除了 S. Livingstone 這個血清型外,似乎與 飼料源較無相關性。
(二) 荷蘭
在 1990 年七月至 1991 年四月間對於三種飼料成品:火雞飼料 (包括第一期、第 二期及第三期料)、蛋雞料及鴨飼料,以及四種飼料原料包括魚粉、肉骨粉、樹薯粉 及玉米進行檢測,並分別比較粉狀及粒狀飼料之沙門氏桿菌汙染率。
結果發現三種飼料共分離出 20 種血清型,總分離率約為 10 % (34/360),本調查 中粉狀飼料之沙門氏桿菌陽性率為 21.4 % (31/145),粒狀飼料之陽性率為 1.4 % (3/215),此結果與前人研究認為飼料製程中打粒的加熱步驟可以減少細菌污染的論點 相符 (Veldman, Vahl et al. 1995)。而三種飼料種類中以蛋雞陽性率最高(21 % ; 32/156),這個結果被認為可能跟此研究中蛋雞飼料多屬粉狀料有關。
四種飼料原料共分離出 16 種血清型,總分離率為 17% ,其中以魚粉分離率最高 31 % (40/130),玉米粉 27 %次之(4/15)。這個結果再次呼應沙門氏桿菌的汙染並非完 全以動物性成份為主,事實上植物性成分的汙染亦是不可輕忽。
而本調查中,飼料原料與成品共分離到 28 種血清型,然而原料與成品兩者間的 血清型分佈並沒有明顯的關聯性,顯示在飼料製程中可能有污染發生 (Veldman et al., 1995)。
(三) 美國
此研究同時對於三個每日飼料總產量為100,000至400,000公噸的飼料廠進行調 查,在春季(四月份)與夏季(八月份)間各採樣兩次,並分別於同一週內的週一、
週三及週五進行採樣。採樣地點分別為原料收件集處(ingredient receiving)、混合槽
(mixer)、打粒廠 ( pellet mill )、冷卻槽(cooler)及出貨區 ( load out ),每個檢體為 同批飼料內各採四至五份混合而成,並在採集前測取飼料打粒的溫度。
結果共採得886份檢體,檢體種類包含68件原料、189件粉塵及629件飼料成品,
飼料成品中有178件未經蒸汽加熱滅菌。此外,除了檢測沙門氏桿菌外並同時進行生 菌數測定。
調查結果發現粉狀飼料的陽性率為8.79 % (16/178),粒狀飼料為4.21 % (19/451),
這個現象再次驗證打粒加熱的過程可以降低飼料污染的菌量,而此調查中粉狀飼料陽 性率低於其他國家的調查結果。飼料原料中以玉米、大豆粉、大豆皮渣及小麥等植物 性原料在生菌數調查中陽性比例偏高,且也皆有沙門氏桿菌污染,也顯示植物性原料 受細菌污染的情形是常在的。
在不同採樣地點的調查結果發現在混合槽檢體受污染的情形顯著的比其他採樣 地點嚴重(P < 0.05),而季節差異性調查則發現春季的生菌數含量顯著高於夏季(P <
0.05),但是沙門氏桿菌的污染量則無季節性差異。
此外,不同的工作日(週一、週三、週五)對於飼料污染程度也表現出顯著的差 異,週五污染度最高,其次為週一,最低的是週三; 此現象推測應與週五的飼料製量 為了滿足週末飼料廠休工時農場的需求而必須有較高的生產量,導致在加熱打粒過程 中均勻受熱的效果較平日差所致 (Jones and Richardson 2004) 。
(四) 日本
於1998年一至十二月間在日本東部採集六間飼料廠的蛋雞飼料成品,採集點於 載運飼料的卡車從飼料廠到農場貯藏室的途中,共採得檢體4418件。檢測結果發現沙 門氏桿菌陽性率為3.3 % (143/4418),共分離到至少32種血清型,以 S. Enteritidis、S.
Livingstone、S. Bareilly及S. Derby是較常分離到的血清型, 其中分離率最高的S.
Enteritidis(8.1 %, 18/146)特別受到注意,因為這個血清型很少從飼料分離到,且一 般認為家禽場內的SE感染是以囓齒類傳播途徑為主,因這個調查結果而被認為飼料 是日本蛋雞感染SE重要的污染源。
與其他國家的調查結果相比,此研究認為日本飼料之沙門氏桿菌陽性率是偏低 的,學者認為應針對飼料研發更好的檢測方法,並加強飼料的檢測調查使之常規化的 進行 (Shirota, Katoh et al. 2000)。
2-3.2 飼料工業的防治方法
2-3-2.1 飼料製程之危害管制點
危害分析與重點管制計畫 ( Hazard Analysis and Critical Control Points, HACCP ) 是一個預防體系,目的在於以科學方法來控制、預防危害因子,並建立實行措施之標 準值。在飼料的預防策略中,包含了飼料原料的監控、飼料在混合加工等生產線之控 管,以及運送、儲存過程的管理等 (Billy and Wachsmuth 1997)。
以瑞典為例,其控制計畫包含了基本控制(basic control)、安全控制(safety control)、特別控制(special control)及相關數據的收集(data gathering)等,並於每 年秋季根據前一年間的監控結果檢討並進一步擬定或變更控制方法。
安全控制以飼料的營養成份的分析為主,像是礦物質、氨基酸及維生素等; 特別 控制以對人體、動物及環境可能具危害性的物質,包含了沙門氏桿菌與戴奧辛; 特別 控制以飼料製程中的空調系統為重點,以加裝過濾網的空調系統來減少室內外流動的 氣體相互污染的情形; 相關數據收集的部分包含相關買賣商的資料及產品的成份、添 加物等資料。在監測的部分,除了瑞典農業部(Swedish Board of Agriculture)會進行 例行性監測外也接受飼料公司的檢獲沙門氏桿菌陽性之通告案件,菌株統一經農業部 接收登記後由國家獸醫研究所(National Veterinary Institute)進行確認、血清型分型及 抗生素感受性試驗,當有S. Typhimurium、S. Enteritidis菌株時由傳染病控制研究所
(Swedish Institute for Infectious Disease Control)進行噬菌體分型。
2-3.2.2 其他應用於飼料業之防治措施
目前已發展之防治方法大致分為在飼料中加入添加劑或將飼料成品經特別處理兩個方 向; 添加劑包括化學添加劑與天然添加劑兩大類,而特別處理則包括加熱法與輻射線 照射等。
(一) 化學添加劑
常見添加的化學物質包括蟻酸 (formic)、氯化氫 (hydrochloric)、硝石 (nitric)、磷 (phosphoric)、丙酸 (propionic acid)、硫酸 (sulphuric acid)、異丙基醇 (isopropyl alcohol)、甲酸鹽 (formate) 、丙酸鹽 (propionic salt)與三鈉磷酸鹽 (trisodium
phosphate)。目前已知使用含氯水的效果並不好,前人研究指出若要有效消除污染紫花 苜蓿種子的S. Stanley,必須使用至少濃度2 mg/ml的氯水消毒 (Jaquette et al., 1996)。
上述的化學物質部份有使用及儲存的不便利性以及對環境與工作者健康的影響,
並且在飼料混合的過程中無法被確認完全均勻分佈等因素,故此類添加劑目前有被利 用自然改變家禽腸內菌叢優勢之緩衝有機酸( buffered organic acid )及益生菌所取代的 趨勢 (Maciorowski et al., 2004)。
(二) 其他飼料添加劑
為了改善使用化學性添加劑的缺點,天然添加劑的研發是目前的趨勢,此類添加 劑之材料的選擇十分多樣化,下面僅以幾種較常見的種類作一簡述。
益生菌添加劑利用競爭性排除 ( competitive exclusion ) 原理,建立理想的腸內環 境來達到自然具有對抗外來微生物狀態,此類型的添加劑的實驗效果普遍不錯,但是 在進行商業化生產時經常遇到困難 (Gaze et al., 2003 )。此外,還有一些醣類添加劑,
尤其是甘露醣、甘露聚醣 (mammaoligosaccharide)等,都被證實具有不錯的抑菌效果,
但是飼料業普遍因成本過高而難以接受 (David, R., 2002)。
(三) 加熱處理法
熱處理通常是在打粒 ( pelleting ) 過程進行,係指將粉狀的飼料混合料以高溫製 成粒狀,此溫度一般在70至90 ℃。有學者認為打粒的溫度至少要在83 ℃以上的溫度才 可消除沙門氏桿菌,進一步也有學者認為飼料中沙門氏桿菌的滅菌溫度和時間有很大 的變異性,這可能跟沙門氏桿菌對熱的抵抗力以及飼料的水份含量有關。
曾有研究以水份含量分別為5 %與15 %的飼料加入等量 S. Enteritidis後同時以82.5
℃加熱處理2.2秒,結果發現水份含量為5 %的飼料其含菌量僅下降1.5 log,而水份含量 為15 %的飼料則下降了4.5 log (Himathongham et al., 1996);此外也有研究證實像是丙 酸 (propionic acid)等化學添加劑搭配加熱處理可以增加滅菌效果 (Matlho et al., 1997)。
飼料加熱的目的是為了打粒、消除細菌,已知超過90 ℃容易變成泥濘狀飼料 ( slushy feed ),且過高的溫度會破壞飼料的營養,然而S. Typhimurium在含水量低的飼 料中經過100 ℃ 加熱處理60鐘後仍舊存活 (Kirby and Davies, 1990),說明了期望在飼 料製程中單憑加熱法來滅絕沙門氏桿菌並不容易。
(四) 輻射線照射法
一般使用的是 60Co gamma rays,乃是藉由改變細菌的DNA結構的控制方法。研究 發現輻射線處理可有效的減少肉品中的E. coli O157: H7,並有效減少穀粉、大豆粉、
肉骨粉及禽類半成品之病原菌; 研究認為飼料經大約15至35 kGy劑量的輻射線照射後 可確定為零沙門氏桿菌飼料,而經10至15 kGy劑量的輻射線則可使飼料中沙門氏桿菌 的含量減少到可檢測值以下 (Maciorowski et al., 2004)。然而這樣的方法並非萬無一 失,例如學者也發現輻射雖然可以有效消除魚粉的病原菌,但是因為無法消除屍氨
(cadaverine)中的proteolytic enzyme,反而有進一步產生scomboid poison等有毒物質 的風險(Urling et al., 1993)。
第三章 材料與方法
第一節 北臺灣白肉雞場污染沙門氏桿菌調查
3-1.1 試驗設計
自 2008 年 4 月至 2009 年 5 月間,於北臺灣地區包含宜蘭縣、基隆市、臺北縣市、
桃園縣、新竹縣市進行採樣,並根據農委會統計室 97 年度第二季與第四季之白肉雞 場規模調查,結果皆顯示北臺灣地區 95 % 以上的白肉雞場飼養規模皆落於 10,000 隻 以上,故擬定以此規模之雞場進行污染沙門氏桿菌之風險因子調查。
本調查之採樣分作兩階段進行,有鑑於國內外尚未建立雞場風險因子調查之採樣 統一標準,故於第一階段先進行前預備試驗來評估最佳採樣時間點與樣本種類後再進 行後續採樣調查。前預備試驗共進行 20 場次,於各雞場連續性於四個時間點採取三 種檢體類別進行沙門氏桿菌之培養分離。
第二階段根據在前預備試驗得到 70 % 之場陽性結果 (14/20),依據表一得知大 於 10,000 隻飼養規模之雞場數共 144 場次,以符合 10 % 精密度 (10 % precision)、
90 % 信賴區間 ( 90 % confidence interval ) 條件下,依據下列公式運算後得知在北臺 灣地區白肉雞場風險調查之雞場數量應達 41 場。
Npq n =
(N-1)B2 + pq
Z2 n= 需要抽樣的最小牧場樣本數
N=北臺灣地區白肉雞場數 (>=10000 隻之雞場飼養規模) p=某疾病的牧場盛行率 (在此為 70 %)
q=1-p (在此為 30 %)
B=估測誤差的界限 (在此設定為 10 %)
Z=標準常態分布係數 (90 % 可信度時,Z = 2.706)
為了簡化採樣方法以增加農民配合意願並符合調查成本考量,本調查得到前預備 試驗結果後經統計分析評估,選擇以最具分離效益之三個採樣時間點與兩種檢體類別 進行後續 21 場次之採樣。亦根據前預備試驗結果之北臺灣地區白肉雞場常見模式修 改問卷,試驗設計與流程圖請見圖一。
並因考量臺灣地處亞熱帶,全年為高溫高濕度的氣候條件下易使雞隻發生熱緊迫 而增加誘發疾病機會,故於四季溫差較大的北部地區採樣分析增加氣候條件之考量,
分別探討溫度與降雨量因子。根據氣象局統計資料將 2008 年 11 至 2009 年 5 月歸類 為均溫低於攝氏 25 度的涼季,其餘月份歸類於熱季;雨季與乾季之區別乃統計調查 期間之降雨平均值後,將降雨量平均值之月份歸類為雨季,各地降雨量之平均值分別 為新竹 152.5 毫米、桃園 178.4 毫米、宜蘭 206.1 毫米。屬於雨季的月份別為新竹 2008 年 4 至 7 月、9 月與 2009 年 3 月;桃園 2008 年 5 至 7 月、9、11 月與 2009 年 2、5 月;宜蘭 2008 年 7 月、9 至 11 月。
3-1.2 調查與試驗方法
本研究以病例對照法(case-control study ; retrospective study)進行,並根據採樣 分離結果與各因子之統計分析判斷各因子是否為影響白肉雞場內沙門氏桿菌污染之 風險因子。
3-1.2.1 雞場設施與飼育條件之問卷調查
北臺灣地區飼養的家禽種類以白肉雞為主,白肉雞是最大量生產且消費量最大的 肉雞禽種,為了解白肉雞生產過程中不同的飼養條件與階段中易造成沙門氏桿菌污染
之風險因子,於參考文獻與白肉雞場飼養管理手冊後彙整出與白肉雞場生產飼育管理 過程之相關因子並設計成一份問卷,藉由問卷調查結果來瞭解在各個調查場內飼養過 程中暴露於各因子的情形,問卷內容包含以下主題 :
ㄧ、雞場之基本資料,包含地理條件、雞舍數量等基本環境設施條件。
二、雞舍之衛生設備與管理,包含用水之來源與處理、斃死雞之處理 方式、各種病媒之預防設施、滅除頻率等。
三、雞場之常規管理,包含工作人員與訪客管理等。
四、飼養計畫,包含雛雞來源、防疫紀錄、發生疫情之管理、飼料 來源與使用情形、雞舍清空與消毒程序等。
完整問卷進行實際訪查前先經由九名家禽相關從業人員協助審核內容之適切性 與問題之明確性,加以增修後完成一份包含基本資料與 78 項問題之調查問卷,皆為 封閉或半封閉式題型,並經研究人員於現場訪查 7 位雞農後確認每份問卷皆約可於 30 分鐘左右完成,並為避免猜測性應答所造成的誤差,統一於出雞前兩週之採樣時進行 問卷調查,以確認對於該場次管理與飼養情形可得到完整明確的應答。最後依前預備 試驗結果之北臺灣地區白肉雞場常見模式來修改問卷,遂從 78 題精減為 56 題,本調 查實際使用問卷請見附錄一。
3-1.2.2 採樣時間與樣本種類
根據前預備試驗結果以最佳的採樣時間點,包括入雛、出雞前兩週與下一批雞入 雛時;檢體類別包含雞隻臟器、飼料,雛雞之採樣乃選取入雛後於雞舍內選取 24 小 時內活動力或健康狀況異常之淘汰雞。
以連續三個時間點進行雞隻檢測可確認不同飼養階段與兩批次間的雞隻有無感 染,以追蹤各時間點的污染情形; 飼料之沙門氏桿菌污染可能造成雞場大規模的水平 污染,故其檢測可瞭解飼料是否為臺灣地區沙門氏桿菌污染雞場的風險因子。
3-1.3 沙門氏桿菌分離與鑑定方法
所有檢體於採檢後皆盡速保存於低溫冷藏環境,當天以低溫宅配運送至實驗室,
在進行試驗前皆冷藏保存。分離方法依檢體種類而有所差異,雞隻先以無菌操作法取 得肝、膽、腸、胃等臟器後依循ISO 6579 : 2002/Amd.1: 2007 (E)之程序進行沙門氏桿 菌培養分離;飼料檢體之分離方法請參照3-2.2。
細菌鑑定方法為於每一種培養基上各選取3-5個典型或可疑菌落接種於tryptic soy agar (TSA; Merck, USA)繼代至少一代後進行生化鑑定。生化鑑定可使用 triple sugar iron agar (TSI;Merck)、sulfide-indole- motility medium (SIM;Merck)、lysine
decarboxylase test、urease test (URE;Merck) 等鑑定培養基或以商業生化鑑定套組
(API-20E;BioMerieux sa, MarcyI’Etoile, France)輔助進行。
惟須注意部份沙門氏桿菌血清型之生化特性並不一致,例如TSIA可能為K/A或 K/AG ( H2S+ )、CIT多數血清型呈現陽性而少數為陰性等。在培養基上是否具有硫化 氫 (Hydrogen Sulfide;H2S ) 並非絕對的選擇標準,因此在XLD、HE上挑選可疑菌落 時尤應注意勿過份依賴此生化性狀 (蔡,2006);目前已知至少有10 % S. Paratyphi、
50 % S. Choleraesuis、與部分發生基因突變之S. Heidelberg、S. Typhimurium皆不產生 硫化氫。
而在雞隻分離株尤應注意 S. Pullorum、S. Gallinarum皆不具運動性,S. Gallinarum 不產氣;並由於飼料之原料成份包含了複雜的來源,像是羽毛粉、動物性成分如魚粉、
肉骨粉等,因此例如當出現lysine、CIT皆呈現陰性時應繼續鑑定是否為S.
Choleraesuis,勿因該血清型少見於家禽而忽略排除。
血清學鑑定則利用Salmonellae O、Vi、H等抗原之市售標準抗血清 (Difco, USA;
Denka Seiken Japan) 進行凝集試驗,所得結果依廠商之說明書進行判讀並依據 Kaufmann-White-Schema進行血清型分型。
3-1. 4 統計分析方法
使用統計軟體 SAS® system 9.1 進行數據分析,探討風險因子與禽場污染沙門氏桿菌 之相關性使用卡方分析 (chi-square test),但若樣本數小於 5 時則以費雪精確檢定 (Fisher’s exact test) 進行單變項統計分析。此外亦計算各因子之勝算比(Odds ratio) 與其 95 %信賴區間 (95 % confidence interval ),並選取 P<0.25 之單一因子進一步 以多變項邏輯式回歸分析 (multiple logistic regression analysis) 進行資料分析以調 整變項間之干擾。