National Chiao Tung University 國立交通大學
生物科技學系 碩士論文
Comparative characterization of the ferrous iron transport systems
FeoABC, SitABCD and EfeUOB in Klebsiella pneumoniae CG43
克雷白氏肺炎桿菌 CG43 中二價鐵螯合系統 FeoABC、SitABCD 和
EfeUOB 之特性探討與比較
Student: Li-Ching Kok
學生:郭俐君
Advisor: Hwei-Ling Peng, Ph.D
指導教授: 彭慧玲博士
July, 2014
克雷白氏肺炎桿菌 CG43 中二價鐵螯合系統 FeoABC、SitABCD 和 EfeUOB 之特性探討 與比較
Comparative characterization of the ferrous iron transport systems FeoABC, SitABCD and EfeUOB in Klebsiella pneumoniae CG43
研 究 生:郭俐君 Student:Li-Ching, Kok
指導教授:彭慧玲 Advisor:Hwei-Ling, Peng, Ph.D
國 立 交 通 大 學
生物科技學系
碩 士 論 文
A ThesisSubmitted to Department of Biological Science and Technology College of Biological Science and Technology
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Biological Science and Technology
July 2014
Hsinchu, Taiwan, Republic of China
中華民國一零三年七月
i
論文摘要
如同其他致病細菌,克雷白氏肺炎桿菌在宿主環境中需要克服缺鐵的考驗。克雷白 氏肺炎桿菌的二價鐵的螯合系統基因組 feoABC、sitABCD 和 efeUOB 在含鐵量豐富的培養 條件中,會受轉錄因子 Fur 抑制其表現。本研究以克雷白氏肺炎桿菌 CG43S3 為親本株分 別 建 構 了∆feoA 、 ∆feoC 、 ∆feoA∆fur 、 ∆feoB∆fur 、 ∆feoC∆fur 、 ∆sitCD 、 ∆sitCD∆fur 、 ∆efeUOB 、 ∆efeUOB∆fur 、 ∆feoB∆sitCD 、 ∆feoB∆efeUOB 、 ∆sitCD∆efeUOB 、 ∆efeUOB∆sitCD∆feoB 和∆efeUOB∆sitCD∆feoB∆fur 等基因缺損突變株,並比較分析這些基 因缺損後對生長及致病性相關的表現型之影響;同時,這三套螯鐵系統預測的啟動子片段 也被轉殖至啟動子報導質體 placZ15 中,再藉由 β-半乳糖苷酶(β-galactosidase)活性評估 其基因表現。結果顯示這三套螯鐵系統的啟動子活性皆可因 fur 基因缺損或缺鐵環境所誘 導;在缺鐵、微氧條件下,feo 的啟動子活性會被 FeoC 負向調控;而 sitABCD 活性最高。 SitABCD 系統能幫助細菌對抗氧化壓力;而 sit 及 feo 基因都剔除結果會使細菌的抗氧化能 力明顯下降;EfeUOB 表現可受酸性誘導,並被雙分子訊息系統的反應蛋白 CpxR 負向調 控,此結果顯示 EfeUOB 可能在有氧、缺鐵及弱酸的特定環境中扮演重要角色。莢膜多醣 體的生成與鐵的多寡有關,然而 FeoABC、SitABCD 或 EfeUOB 基因剔除並不影響克雷白 氏肺炎桿菌 CG43 的莢膜多醣體產生;而除了∆sitCD∆efeU 和∆feoC 之外,其他的基因缺損 株都會降低生物膜的生成,在添加了三價鐵螯合劑 Deferoxamine 後,這些缺損株的生物膜 減少更為顯著;有趣的是,受鐵濃度影響表現、而且是決定生物膜生成的主要因子第三型 線毛,其主要單位蛋白 MrkA 並沒有受到這些基因缺損的影響,這結果暗示這些螯鐵系統 並不是透過影響 MrkA 表現來影響生物膜的生成。
ii
Thesis Abstract
As the other pathogenic bacteria, Klebsiella pneumoniae often encounters the challenge of iron depletion surroundings in the host cells. The iron transport system encoding operon feoABC,
sitABCD, and efeUOB are present in the genome of K. pneumoniae CG43. Under iron rich cultured
condition such as LB medium, the expression of feoABC, efeUOB and sitABCD are repressed by the global regulator Fur (ferric uptake regulator). In the study, ∆feoA∆fur, ∆feoB∆fur, ∆feoC∆fur,
∆sitCD, ∆sitCD∆fur, ∆efeUOB, ∆efeUOB∆fur, ∆feoB∆sitCD, ∆feoB∆efeUOB, ∆sitCD∆efeUOB, ∆efeoUOB∆sitCD∆feoB and ∆efeoUOB∆sitCD∆feoB∆fur derived from K. pneumoniae CG43S3
have been generated. The mutation effects on the bacterial growth and virulence-related properties such as capsular polysaccharide (CPS) production, oxidative stress responses, type 3 fimbriae expression, and biofilm formation were analyzed and compared. The putative promoter of the three iron transport systems were isolated and individually cloned in front of the promoterless lacZ on the reporter plasmid pLacZ15 for the optimal expression and condition analysis using β-galactosidase activity. All three iron transport systems could be induced by the absence of Fur repressor and depleting iron from the culture medium. The feo promoter was negatively influenced under iron-depleted and microaerobic condition by FeoC, a predicted Fe-S sensor regulator. The microaerobically-induced sitABCD showed the highest promoter activity among these three systems. SitABCD plays important role in protecting bacteria from oxidative stress, with combined loss of sit and feo further decreased bacterial oxidative-resistance. EfeUOB is induced by weak acid and negatively regulated by the two-component regulator CpxR. This indicates its specific role in aerobic, iron-depleted and low pH environment. Although iron-availability affects CPS biosynthesis, FeoABC, SitABCD and EfeUOB transport systems do not involved in regulation of CPS biosynthesis. Except ∆sitCD∆efeUOB and ∆feoC, deletion of genes from these acquisition
iii
systems decreased biofilm formation of K. pneumoniae CG43S3, wherein iron chelators deferoxamine exerted more decreasing effects of biofilm formation. Interestingly, these deletion mutants did not decrease the expression of MrkA, the major pilin of type3 fimbriae which is iron dependent and the major determinant of biofilm formation. These results imply that these ferrous iron transport systems regulate biofilm formation independently of the expression of type 3 fimbriae.
iv
Acknowledgement (謝誌)
時間過得很快,還來不及將路上的風景看個盡興,就到達目的地。不知不覺,兩年 的光陰過去,兩年的碩士班生涯也終告一段落。要我為這兩年的生活做一個總結,那就是 計劃永遠趕不上變化:從一開始尋找指導教授、研究題目、說不盡的生活點滴、到畢業口 試的口試委員,結果永遠不是開始的那個 A 計劃,也因為如此,這兩年有如坐雲霄飛車的 般的生活非常充實。2012 年夏末,匆匆交接工作上的事項,隻身從屬於熱帶國家的馬來 西亞飛到有四季的台灣來深造,在身邊許多人的幫助下從不同的氣候、食物、文化以及習 慣中學習和適應,當中的感謝之情無法在短短的一文中傾訴。 慶幸有指導老師彭慧玲老師,讓我有緣加入彭家溫馨的大家庭。研究的過程總是充 滿著未知和重重關卡,在疲憊和躊躇的時候彭老師總是親和力十足地循循善誘,不辭勞苦 和學生們每週一 meeting,關心我們的實驗進度和生活近況;在徬徨的時候給予鼓勵,鬆 懈的時候適度的鞭策,還不忘為我們將來出社會後的生活未雨綢繆。老師,辛苦您了,謝 謝您! 感謝清華大學張晃猷老師在 Joint meeting 和口試時給我的建議和肯定,減輕我緊張 的心情,讓我可以順利口試畢業。感謝中興大學的鄧文玲老師在 Joint meeting 時的提議和 提問,提醒我忽略的盲點,使我對自己的研究主題有更深入的了解。感謝梁美智老師經常 請我們享用您用心製作的小點心和蛋糕,為我們加油打氣,和在口試中的指導。 感謝從一開始一步一步教導我這什麼都不懂的新手、溫柔體貼的靜柔學姐,使我的 實驗技術越來越熟練,在許多 try and error 中學習得更深入。學姐在我碰到問題總會放下 手上的事情來協助我,囑咐我在實驗上要注意的小細節;在冬天的時候體貼我怕冷的體質v 送我一條小毯子保暖,回想起來總有暖暖的感動。謝謝哲充學長在我碩班兩年來無數的幫 助,實驗上有什麼疑難雜症,學長都有不一樣的解決方案,揮走我頭上的烏雲,為實驗重 新找到光明。感謝同樣來自馬來西亞、外表內涵皆優的正妹燕曦,因為你的相助,我才有 緣和你們認識並加入彭家的家庭。雖然冠男總是‘嫉妒’我們倆同聲同氣,可是我們在很多 想法和興趣上總是不謀而合,和你的幾次長談讓我對你的認識更深一層,可惜之後我們都 各忙各的,希望還有機會和你一起 hang out!記得第一天加入彭家實驗室時,趕上了中秋 節的烤肉活動,坐我旁邊、一直為不小心將烤肉用的用具戳到我而道歉的蕙瑜,讓我見識 到原來還有人比我更迷糊、更粗神經。蕙瑜永遠有一股認真和拼命三郎的狠勁,有不到黃 河心不死的決心,生活上不拘小節,力氣大得驚人,只要她用力一扭,瓶瓶罐罐沒有打不 開的。我們兩個像機車衝鋒隊一樣,只要當下有興致就借機車往外衝,鬱悶的時候去散散 心、開心的時候去逛逛,非常開心有一個陪我瘋的好夥伴;在我搞不清一些生活細節時, 感謝你幫了我許多,你應該都不記得了,可我會一直記得的^_^ 希望你保持這積極樂觀的 態度,繼續用你賢惠的一面和笑聲感染身邊的人吧!同在那次的中秋烤肉活動中,熱情的 冠男手法嫻熟地為大家烤肉,親切的態度讓初來報到的我很容易融入新環境;實驗上有很 多小撇步讓我的實驗進行得比較順利,至今我還記得 WesternBlot 的黑膠概念 XD 非常懷 念那些空閒時我們聊得不亦樂乎的日子,希望你的補習中心越做越旺,成了彭董後要回來 關照一下我們哦~在休息室和酷酷的子祥做了鄰居一年多,對你一開始冷酷的印象到後來 的貓控熱血青年是怎麼一回事呢?你讓我見識到交大學生厲害之處:運動好、頭腦好、還 多才多藝,質優生就是如此吧!在碩班的第一年,和同為生科系的珍儀林一起修課,幾乎 每一餐都在可愛老闆的素食店用餐,還同寢了一學期,和活潑開朗的你一起時,我們會互 相鼓勵,給對方打氣,總覺得自己充滿正能量。在很多人對我都只有內向安靜的印象時,
vi 你卻觀察到我另一面,且不吝給我讚美,實在太讓我感動和開心,被了解的感覺太棒了! 修課最多、進度最快、腿最長的你發展得也最快,希望你在工作上繼續勇往直前,再創佳 績~和珍儀同姓又同月同日生日的小博士偉豐林,關於你的傳說已經很多了,簡短的介紹 就是彭家花園的小園丁兼外交官、大潤發忠實顧客、PTT 常駐鄉民、無論大小事都略懂的 豐哥,非常幸運能認識你。身為我們這屆碩班四位研究生中唯一的男生,在我們的話題圍 繞著女生話題時還能和我們聊的很開心,對大家的要求都不會拒絕的好好先生,從你身上 學習到很多知識和常識,感謝你聆聽我面對瓶頸時的訴苦,互相討論結果後都有茅塞頓開 的 feel,期待你接下來的發光發熱!碩班這兩年有你們三人的陪伴一起成長,是我畢生都 會回味無窮的日子,謝謝你們,別忘了保持聯絡喔! 謝謝即將成為彭家大支柱的博班學弟(?)家睿,總是默默地為實驗室完成很多繁 雜的事情,還不忘給休息室打掃,可從來不見你有急躁的時候,EQ 和 IQ 都一級棒,祝愿 你實驗能順順利利,加油!謝謝瑋芝和子芸的加入,讓實驗室充滿著一片新氣象。瑋芝的 背影總讓我錯覺是靜柔學姐,大姐姐性格的你讓人很放心把事情交給你處理;謝謝貼心和 細心的子芸,不時關心我有沒有吃好睡好,在我要搭公車時載我一程,還常請我吃小點心 可是子芸,你才應該要吃好睡好吧,你已經瘦得快被新竹的風吹走了……希望你們俩的 實驗越來越順利,越來越有信心!謝謝新加入的若迪、啟翔、銘軒、容毓、錦松、傑中等, 讓彭家的家族更加龐大,實驗室充滿著各種笑聲,希望你們保持這樂觀又積極的態度,迎 接並享受接下來的研究生涯。 感謝我在馬來西亞的朋友和前同事們,一佳、湘凌、慧韵、振業、振寧、Vernice、 Chin Fen、Mike So、Rizal、Yong、Kamariah、Sanjeeta、Juliana、KS 等。雖然減少了聯絡 卻從不忘關心我,在新聞上看到有關台灣的意外和天災時總會收到你們關心的信息;我自
vii 知我在忙碌的時候忘了和你們聯絡,可是我總會想起你們,讓我恢復元氣。最後感謝我最 親愛的家人,感謝無條件給予寬容和支持的爸爸媽媽,從來不求我有什麼大成就,只盼我 過得平安喜樂就好,也讓我無後顧之憂地追逐我的夢想!還要感謝我的弟妹們,在你們面 前我可以毫無保留地表達自己的喜怒哀樂,這些年有你們的陪伴太幸福了!感身邊的一切 人、事與物,不管好壞,都讓我的視野和心境變得更寬闊更豁達!感恩! 俐君 謹致於 交通大學生物科技學系 中華明國一零三年八月
viii
Table of Contents
論文摘要... i
Thesis Abstract... ii
Acknowledgement (謝誌) ... iv
Table of Contents ... viii
List of Tables ... x
List of Figures ... xi
List of Appendices ... xiii
Abbreviations ... xiv
1.0 Introduction ... 1
1.1. Iron is essential for bacterial growth and virulence ... 1
1.2. Regulation of iron homeostasis in bacteria ... 2
1.2.1 Fur ... 2
1.2.2 RyhB ... 3
1.3. Bacterial iron acquisition systems ... 4
1.3.1 The Feo transporter system... 5
1.3.2 The Sit transporter system ... 8
1.3.3 The Efe system ... 9
1.4. K. pneumoniae and the related studies ... 11
1.5 Specific aims ... 13
2.0 Materials and Methods ... 14
2. 1. Plasmid, primers, bacterial strains and growth conditions ... 14
2. 3. Bioinformatics analysis ... 15
2. 4. Construction of gene-deletion mutants ... 15
2.5. Construction of a pLacZ reporter system ... 16
2.7. Measurement of promoter activity through β-galactosidase activity assay ... 16
2.8. Sedimentation test ... 17
2.9. Biofilm formation assay ... 17
ix
2.11. Western blot analysis of the expression of type 3 fimbriae ... 18
3.0 Results ... 20
3.1. Generation of the specific gene deletion mutants ... 20
3.2. Effect of iron availability on the growth of the specific gene deletion strains ... 20
3.3. Analysis of feoABC, sitABCD and efeUOB promoter region ... 21
3.4. Effect of growth phase and oxygen availability on the expression of feo, sit and efe ... 22
3.5. Effect of fur deletion and iron-depletion on expression of feo, sit, and efe ... 23
3.6. Effect of feoC deletion and rstA deletion and on expression of feo, sit, and efe ... 23
3.7. Effect of manganese on expression of feo, sit and efe. ... 24
3.8. Effect of weak acid and cpxR deletion effect on the efe expression ... 24
3.9. Analysis of cumulative effect of acid and iron-depletion or iron-supplemented on growth ... 24
3.10. Analysis of ferrous iron transporter regulation in CPS biosynthesis ... 25
3.11. Analysis of deletion effects on oxidative stress response ... 26
3.12. Analysis of deletion effects on the biofilm formation under different culture conditions 27 3.13. Analysis of the deletion effects on the expression of type 3 fimbriae ... 27
4.0 Discussion ... 29
5.0 References ... 35
6.0 Tables ... 55
7.0 Figures... 60
x
List of Tables
Table 1. Bacterial strains used and constructed in this study ... 55 Table 2. Plasmid used and constructed in this study ... 57 Table 3. Primers used in this study ... 58
xi
List of Figures
Figure 1. Schematic representation of the feoA, feoB and feoC deletions and the verification of
deletions by PCR. ... 61
Figure 2. Schematic representation of the sitCD deletions and the verification of deletions by PCR. ... 62
Figure 3. Schematic representation of the efeUOB deletions and the verification of deletions by PCR. ... 63
Figure 4. Growth analysis of K. pneumoniae strains under iron-replete or iron-depleted media. 64 Figure 5. Growth analysis of K. pneumoniae ∆fur-derived strains... 65
Figure 6. Diagrammatic representation of K. pneumoniae CG43 feoABC, sitABCD and efeUOB promoter region and Pfeo::lacZ, Psit::lacZ and Pefe::lacZ fusion construction. ... 67
Figure 7. Analysis of growth phase and static-cultured effects on activity of Pfeo, Psit and Pefe in K. pneumoniae CG43S3 ∆lacZ (Z01) ... 68
Figure 8. Analysis of fur deletion effect and iron-depletion effect on Pfeo, Psit and Pefe activity .. 70
Figure 9. Analysis rstA, low pH and feoC deletion effect on Pfeo activity in different culture conditions. ... 72
Figure 10. Analysis of effect of manganese on activity of Pfeo, Psit and Pefe. ... 74
Figure 11. Analysis of weak acid and cpxAR effects on activity of Pefe ... 75
Figure 12. Effect of iron availability on growth of K. pneumoniae strains in acidic media. ... 76
Figure 13. Analysis of capsular polysaccharide (CPS) biosynthesis in of K. pneumoniae strains in different media. ... 78
Figure 14. Effect of deletion mutants and iron chelators on the oxidative stress response ... 80
xii
Figure 16. Biofilm formation analysis of deletion mutant strains. ... 82 Figure 17. Biofilm formation analysis of deletion strains under iron-depletion ... 83 Figure 18. Iron-depleted effect on the iron-dependent MrkA expression under shaking condition ... 84 Figure 19. MrkA expression of the ∆fur-derived deletion mutants under shaking condition ... 86 Figure 20. Iron- and oxygen-depleted effect on MrkA expression for the deletion mutants ... 87 Figure 21. Schematic representation (not to scale) of the proposed regulation of feoABC, sitABCD
and efeUOB in CG43S3. ... 88 Figure 22. Schematic representation of the proposed model of functional role and location of
xiii
List of Appendices
Appendix 1. Iron-acquisition genes in K. pneumoniae CG43 that expression were induced in ∆fur strain ... 91 Appendix 2. qRT-PCR analyses of the expression of iron-acquisition genes in K. pneumoniae CG43 ∆fur ∆ryhB and ∆fur strains ... 92 Appendix 3. The comparison of gene organization along with efeUOB operon between E. coli and
xiv
Abbreviations
BCIP 5-bromo-4-chloro-3-indolylphosphate CPS capsular polysaccharide
DIP 2‘2’-dipyridyl
DFX deferoxamine mesylate salt DTT dithiothreitol
EDTA ethylenediamine-tetraacetic acid
Fe iron
Fe(II) ferrous iron Fe(III) ferric iron
IPTG isopropyl-1-thio-β-D-galactopyranoside kb kilobase(s)
KCl potassium chloride kDa kilodalton(s) LB Luria-Bertani MgSO4 magnesium sulfate
Mn manganese
MnCl2 manganese chloride
Na2CO3 sodium carbonate
Na2HPO4 sodium phosphate dibasic
NaH2PO4 sodium phosphate monobasic
xv
ONPG ortho-nitrophenyl-β-D-galactopyranoside PAGE polyacrylamide gel electrophoresis PCR polymerase chain reaction
PVDF polyvinylidene difluoride
qRT-PCR quantitative reverse transcription polymerase chain reaction SDS sodium dodecyl sulfate
sRNA small RNA
1
1.0 Introduction
1.1. Iron is essential for bacterial growth and virulence
Iron is needed for many important cellular functions and processes, such as the transport and storage of oxygen and as catalyst of many enzymes. Enzymes that utilize iron are involved in many major biochemical processes such as photosynthesis, N2 fixation, respiration,
methanogenesis, trichloroacetic acid (TCA) cycle, gene regulation and DNA biosynthesis [1]. These enzymes include ribotide reductase, nitrogenase, peroxidase, catalase, cytochromes, and succinic dehydrogenase [2]. Iron biological function is depends on its incorporation with proteins, either as a mono- or binuclear or as a part of iron-sulfur clusters [3] or heme [1, 4].
In addition, iron is also an important factor in cellular events such as virulence, biofilm formation and quorum sensing [4-7]. Low iron in the environment can trigger the pathogens to induce virulence genes such as toxins or adhesins, although their activities are not linked to iron. For instance, Pseudomonas aeruginosa produces an extracellular toxin that inhibits protein synthesis in eukaryotic cells [8]. A number of studies have shown higher resistance to infection in animals in which level of iron in serum have been reduced by an deficient diet or iron-deficient mutants [2, 9-13]. The role of iron in many system indicates that virtually all living organisms require iron to survive and grow.
Iron is present in either an oxidized ferric [Fe(III)] or a reduced ferrous [Fe(II)] form. For most microorganism, iron is not easily available under aerobic conditions, due to the poor solubility of the oxidized Fe(III) (as low as 10-18 M at pH 7.0 [1]). For this reason, environmental microorganisms living in aerobic life-styles are often confronted with the shortages of bioavailability of iron. For pathogens, the iron-restriction problem is even more extreme since the
2
host limits iron availability by holding it within intracellular proteins. Most intracellular iron is found as hemoglobin, heme, ferritin, and hemosiderin. While the extracellular trace amount of iron is bound by the high affinity iron binding glycoproteins transferrin and lactoferrin [2, 14].
While reduced Fe(II) is more favored by bacteria in term of solubility, it can be hazardous to bacteria because Fe(II) participates in Fenton/Haber-Weiss reaction that lead to formation of reactive oxygen species (ROS) such as superoxide (O2-), hydrogen peroxide (H2O2) and the highly
destructive hydroxyl radical (•OH), which in turn causing harmful effects on fatty acids and other biological macromolecules [4, 15, 16].
1.2. Regulation of iron homeostasis in bacteria
1.2.1 Fur
To balance these dual aspects, bacteria must tightly regulate the cytoplasmic iron concentration, which is primarily mediated by the key regulator ferric uptake regulator (Fur) [1, 17, 18]. Under iron replete conditions, by using Fe(II) as a co-repressor, the Fe(II)-associated Fur functions as a transcriptional repressor of iron uptake-related genes by binding to its specific 19-bp consensus DNA sequences, called Fur box (GATAATGATwATCATTATC, w=A or T) [17], on the target promoters. The binding of Fur at the promoters impedes the binding of RNA polymerase thereby preventing transcription from these genes. We previously demonstrated several iron acquisition systems including iro, iuc, sit and feo were induced by fur deletion [19, 20].
On the other hand, the regulation by Fur is now known to be more complex, since Fe(II)-Fur can also activate genes in an indirect manner via the derepression of a small regulatory RNAs such as ryhB, nrrF and fsrA [4, 21-24] . In addition, regulation of Fur was revealed having
3
interesting link between redox stress management, acid tolerance management, adhesins factors, capsular polysaccharide (CPS) biosynthesis, some metabolic pathways with iron homeostasis [1, 17, 19, 20, 25, 26]. Finally, in some case, in the absence of iron, Fur functions as an activator on target gene promoter in apo-Fur form [4, 27].
1.2.2 RyhB
In E. coli, small RNAs (sRNA, non-coding RNA) is found involving in a variety of cellular functions such as a modulation of RNA polymerase activity (6S RNA) [28] protein tagging for degradation (SsrA and tmRNA) [29] and regulation of translation [30-33]. Some of these RNA use sequence specific RNA-RNA interaction to regulate mRNA synthesis. RyhB (also named as Sral) is a sRNA that initially discovered to be complementary to a portion of sdhCDAB operon encoding succinate dehydrogenase, affecting the cells growth in media containing succinate as a sole carbon source [34]. Later discovery explained this disability of growing in media lacks of specific carbon source is closely related to Fur.
The first step of sRNA RyhB mechanism of action initiated by pairing in an antisense manner with its mRNA targets. Then the pairing is further enhanced and stabled by the RNA chaperon protein Hfq [35, 36]. Following that, RyhB rapid degrades the sRNA with the mRNA target by RNase E, which is a part of complex called RNA degradasome [37, 38]. A double strand ribonuclease RNase III, is also involved in co-degradation of RyhB and its target mRNA [39]. Interestingly, the degradation leaves the promoter expression of the targeted genes intact while the mRNA is degraded [33]. This mechanism enables rapid shutdown of RyhB expression when condition returns to normal Recent discoveries suggested and revealed RyhB is involved in the positive regulation of Fur in certain genes [33, 40, 41]. Under iron-repletion, Fur exerts its
4
repression on ryhB transcription. However, in the absence of Fur or when iron becomes scarce in the cell, the repression of ryhB is relieved. RyhB targets only the non-essential genes for degradation [42, 43], enabling only essential proteins to have access to the free iron. The regulation is very critical during iron-deprivation, when Fur inactivation enables iron-acquisition genes and RyhB, which in turn reduces its usage to crucial proteins. Therefore, the interplay between Fur and RyhB balance the intracellular pool of available iron [44].
Previous study in K. pneumoniae CG43 suggested that RyhB plays a role downstream of Fur in the regulation of iron-acquisition systems and CPS biosynthesis. A qRT-PCR analysis implied that RyhB activates the expression of iucA, fepA, fepB, entC, fecA and fecE, but repress the expression of fhuA and sitA (Appendix 2). However, no apparent base pairing was found in the 5’-untranslated region of the iuc or fec operons a bioinformatics application (RNAhybrid) analysis showed no apparent base pairing was found, suggesting that the activation of iucA and fecA by RyhB is not a result of direct interaction. Yet, a possible pairing between RyhB with the adjacent sequence of translational start site of sitA was predicted [23].
1.3. Bacterial iron acquisition systems
Bacteria have evolved several iron uptake systems to both the ferric and ferrous form of elemental iron to meet their demands. In Escherichia coli, depending on the strain investigated, more than 10 iron-uptake routes known. In the presence of oxygen and at neutral pH environment, oxidized iron [Fe(III)] is more dominant, bacteria chelate iron by producing strong extracellular Fe(III) chelators, called siderophore. For Gram-negative bacteria, this transporter generally consisting of a Ton-ExbBD dependent outer-membrane receptor, an intermediate ferrisiderophore-binding protein presents in the periplasmic space and an inner-membrane ATP-ferrisiderophore-binding cassette
5
(ABC) permease. Once ferrisiderophore enters the cytoplasm, it is either degraded by an esterase, or undergo a reducing process leading to the release of ferrous iron and allowing the apo-siderophore to be recycled and reused [1, 25].
In contrast, at reducing or anaerobic conditions, the equilibrium of iron shift to the soluble Fe(II) iron form. The first bacterial ferrous iron (Feo) transport system was discovered in E. coli K-12 [45]. This type of iron transport system is quite different from the siderophore-dependent systems, and so, this transport system is also characterized as TonExbBD-independent transport system. Another type of bacterial iron transporter is the metal type ABC transport system that have specificity for iron but not necessarily require outer-membrane receptors or siderophores. Such systems include SfuABC, SitABCD, YfeABCD, FbpABC, FutABC and EfeUOB of Serratia
marcescens, Salmonella typhimurium, Yersinia pestis, Neisseria gonorrhoeae, Synechocystis PCC
6803 and E. coli [1, 46]. YfeABCD is part of the SitABCD family that regulate manganese and iron levels within the cell. The EfeUOB transport system is specific for ferrous iron transport, however, it only presents in certain pathogenic species [46-48].
1.3.1 The Feo transporter system
The Feo iron transport system is widespread among bacteria and thus appears to be a major route of ferrous iron acquisition [49, 50]. The feo operon in K. pneumoniae is constituted by three genes: feoA, feoB and feoC (also called as yhgG). In some Acidithiobacilli, it is preceded by a putative dedicated permease-encoding gene, feoP. FeoP is not contiguous with the feoABC cluster but its upstream predicted Fur box suggests its involvement in iron metabolism [51].
FeoA is a small Src-homology 3 (SH3)-like hydrophilic protein located in cytoplasm, predicted to act as a GTPase-activating protein (GAP) and/ or an Fe(II)-dependent repressor [49].
6
Recently, FeoA was shown to interact with FeoB in S. enterica [52] and is required for Fe(II) uptake in both S. enterica and Vibrio cholera in vivo [52, 53]. Further on, the 31P nuclear magnetic
resonance (NMR) study in E. coli proposed that FeoA may function by interacting with the highly conserved core region in the transmembrane domain of FeoB, instead of a GAP as suggested earlier [48]. An unpublished NMR structure of K. pneumoniae FeoA, which shares 90% sequence identity with E. coli FeoA protein, possess all secondary structure element same with the E. coli FeoA [48], suggesting they both may function similarly.
FeoB is thought to be the main Fe(II) transporter in the cytoplasmic membrane. It is a large protein containing a cytosolic N-terminal domain (NFeoB) that can divided into a Ras-like G domain and a helical S domain [54]. The G domain of NFeoB is thought to provide energy for the transport process or to regulate the transport by sensing the energy state of the cell and relays to the transmembrane [26, 48]. While the S domain may act as an open or close switch [55, 56]. The C-terminal region of FeoB is a helical transmembrane domain that likely form the porin functions as a Fe(II) permease [57, 58].
FeoC is a small, hydrophilic, winged-helix protein which is usually involved in DNA (sometime RNA) binding [49, 59]. In comparison with FeoA and FeoB, FeoC is not well conserved between specie and found only in γ-proteobacteria [49]. Multiple alignment of various bacteria, including K. pneumoniae, FeoC amino acid sequence shows that they possess four conserved cysteine residues with a consensus sequence Cx4CxxCx5-8C, that likely provides binding sites for
iron in the form of an iron-sulfur ([Fe-S]) cluster [49, 60]. Therefore, FeoC is suggested to act as a [Fe-S]-dependent transcriptional regulator, directly controlling feo expression. However, a study in Y. pestis suggested that FeoC did not regulate feo promoter [13] and no report has confirmed the DNA binding activity of FeoC.
7
Up until recently, two studies proposed that FeoC may function at post-translational level [53, 61]. In S. enterica, FeoC protein was showed contributing an increased FeoB protein level by protecting NFeoB from FtsH protease-mediated proteolysis [61]. Another evidence was discovered in Vibrio cheolerae, feoC encodes a protein that interacts with the cytoplasmic domain of FeoB, as determined using BACTH bacterial two-hybrid system [53]. Furthermore, structural study in K. pneumoniae showed FeoC protein binds to the NFeoB with high affinity and may coordinate the [4Fe-4S] cluster to regulate Fe(II) by modulating G protein activity [3, 55, 62].
Since the first description of the Feo system in E. coli [45], the importance of the Feo system has been confirmed by several bacterial systems. In both E. coli K12 and Salmonella, feoB mutant was reported to be deficient in uptake of ferrous iron and their colonization ability in mouse intestine [63, 64]. Besides, FeoB appears to be the main ferrous iron transporter and is required for
Helicobacter pylori colonization of mouse gastric mucosa under Fe(II)-restricted conditions. FeoB
is also required for the virulence of Porphyromonas gingivalis [55, 65]. Therefore, various studies have clearly established a role of the Feo system in bacterial colonization of the gut and in virulence. However, a deletion feoB mutant in K. pneumoniae CG43 was not attenuated in an 50% lethal dose (LD50) assay [19].
Anaerobiosis has been shown to influence persistence and virulence of enteric pathogen such as E. coli [66, 67], Salmonella spp. [68-70], V. cholera [71, 72] and Y. enterocolita [73]. The anaerobiosis response regulators Fnr and ArcA are the primary redox regulators responsible for the activation or repression of genes associated with the transition to anaerobiosis. Fnr has been shown to stimulate transcription of feoABC in E. coli and S. enterica under anaerobic condition [61, 74]. As well in Shigella spp., both Fnr and ArcA induced promoter activity of feo anaerobically [75].
8
RstA/RstB is a member of the OmpR subfamily of two-component regulatory system, consisting of the membrane sensor RstB and its response regulator RstA [76]. A DNA microaaray analysis in Salmonella revealed that under iron-replete conditions, expression of RstA protein from a plasmid lowered transcription levels of various genes involved in iron acquisition, including
fhuACDEF, sitA, entAB, exbD, cirA and tonB through Fur-Fe(II) protein. By contrast, the RstA
protein activated transcription of the feo operon by binding directly to the feoABC promoter [77].
1.3.2 The Sit transporter system
Many microorganisms have secondary Fe(II) transporters whose primary function is the uptake of other divalent metals such as Mn(II), Mg(II) and Zn, but that can also import Fe(II) in certain circumstances. This include the Nramp-like transporter, such as MntH [78] and metal ABC permease such as SitABCD [79].
The Salmonella iron transporter SitABCD was first identified in S. enterica Serovar Typhimurium as a homolog of the Y. pestis YfeABCD transporter [80]. This transporter is thought to be involved in iron uptake, although its specificity between iron and manganese is not clear. SitABCD resembles a typical ABC transporter with a periplasmic binding protein (SitA), an ATP-binding protein (SitB), and two inner membrane permease (SitC and SitD). The location of
Salmonella sit locus in the pathogenicity island SPI-1 might implicitly reveal its importance in
virulence. Inactivation of the genes encoding for Sit transporter resulted in decreased virulence in murine experimental infection model [81].
A variety studies show combining mutations in the Sit and Feo system or/ and other metal transporter MntH resulted in completely avirulent or more sensitive to oxidative stress. Such
9
observation is found in avian pathogenic (APEC) E. coli χ7122 [82], S. enterica Serovar Typhimurium [11, 12], S. flexneri 2a [83] and Y. pestis [84].
Recently, the association of sit with virulence was also observed in K. pneumoniae NTUH-K2044 [85]. When the bacterial cells was cultured in a nutrient- and iron-limited mammalian cell culture condition (DMEM), SitA protein was found significantly increased. Similar to the forementioned observation, deletion of sitA showed higher sensitivity to the stress of peroxide oxidation. Additionally, virulence of sitA deletion mutant, comparing to the wild-type strain, was attenuated in a mouse intraperitoneal infection model [85].
In S. enterica Serovar Typhimurium, transcription of sit is regulated by Fur and MntR. Both Fur and MntR are metallo-regulatory proteins that respond primarily to Fe(II) and manganese (Mn) respectively [86]. The presence of Mn and Fe in medium caused transcriptional repression of the sit promoter. In contrast, increased expression of sit was observed under high oxygen conditions and higher cell densities. However, sit did not appear to be controlled by either redox regulator Fnr and ArcA or the alternative sigma factor, RpoS [87]. Hence, the increased expression affected by oxygen level and growth phase might be caused by decreased availability of metals required for repression by the metallo-regulatory proteins [87].
1.3.3 The Efe system
Many bacteria possess a particular ferrous iron transport system with functional similarities to the Fet3p/Ftr1p system in yeast, fungi and algae [88-91]. E. coli O157:H7 encodes an elemental ferrous iron (EfeUOB) transport system that was recently identified as a highly specific ferrous iron transport system [46, 92, 93].
10
Only a few experimental studies on the function of EfeUOB proteins have been reported. So the large part of understanding about efeUOB comes from the sequence analysis. The EfeU protein is a homologue protein of yeast iron permease Ftr1p with seven transmembrane helices, two of these helices, contain an conserved REXXE motif [47], which was initially suggested to be involved in ferrous iron transport. However, later studies suggested that Fe(II) is taken up upon Fe(II) oxidation by an oxidoreductase mechanism that possibly involves the periplasmic EfeO and EfeB protein [46, 94].
EfeO possess an N-terminal cupredoxin-type domain (Cup domain) and two distinct iron binding sites, suggesting that this protein might be involved in ferrous iron oxidation [94]. It was further suggested that ferric iron could be subsequently be transferred to the EfeU permease, and that the oxidized state of the Cup domain could be regenerated by electron transfer to the third component EfeB. EfeB is a periplasmic homodimeric heme belonged to dye-decolorizing peroxidase (DyP) protein family, which is secreted by the twin-arginine translocation (Tat) pathway [95, 96]. Recent study showed that this protein can extract iron from heme without breaking the tetrapyrrole ring [97].
The latest proposed mechanism for EfeUOB transport system was suggested by Marcus et
al. [93] based on Gram-positive Bacillus subtilis bacterial model. Briefly, the binding protein EfeO
and the permease EfeU form a minimal complex for ferric iron uptake. The third component EfeB is a hemoprotein that oxidizes ferrous iron to ferric iron for uptake by EfeUO. In conclusion, the EfeUOB system contributes to high-affinity uptake of iron that is available in both oxidized and reduced state. Furthermore, in B. subtilis, the EfeB protein is able to promote growth under microaerobic conditions where ferrous iron is more abundant. More importantly, EfeB can provide
11
cell envelope stress protection by eliminating reactive oxygen species that accumulate in the presence of ferrous iron [93].
In E. coli, transcription of efeUOB operon is known to be induced under iron-depleted, low pH or in the presence of exogenous copper [46]. The interesting finding is, the expression of
efeUOB occurs under aerobic condition [46], wherein the amount of ferrous iron is relatively low.
The transcriptional factor Fur and the two component system CpxAR are involved in the iron- and pH-dependent expression, respectively. The OmpR-like response regulator CpxR appears to preferentially bind and repress the efeUOB promoter in its deactivated (de-phosphorylated) form. However, in respond to low pH, the sensor kinase CpxA phosphorylates CpxR and causes derepression of efeUOB.
1.4. K. pneumoniae and the related studies
K. pneumoniae is a Gram-negative, non-motile, facultative anaerobic,
heavily-encapsulated rod-shaped bacterium of the family Enterobacteriaceae. Its complex acidic polysaccharide capsule makes it distinct from other Enterobacteria, which is a major virulence factor avoiding phagocytosis [98]. It is an opportunistic pathogen, causing community acquired and nosocomial infections, including septicemia, pneumonia, urinary tract infection, meningitis and purulent abscess at various body sites particularly in immunocompromised patients [99]. Since 1980s, reported case of pyogenic liver abscess patients infected by K. pneumoniae started to be increasing [100, 101]. In Asian countries, especially Taiwan and Korea, K. pneumoniae is the predominant pathogen found in pyogenic liver abscess in diabetes patients [102-104]. Other than capsular antigen, lipopolysaccharide (LPS), adhesion, iron-acquisition system and serum resistance factors are also main virulence factors involved in K. pneumoniae infection [105].
12
K. pneumoniae CG43 used in this study is a clinical isolate belonged to K2 serotype with
a LD50 of 10 CFU for laboratory mice [106, 107], isolated from diabetes mellitus patients of Chang
Gung Memorial Hospital. Highly mucoid phenotype of K. pneumoniae CG43 has been correlated with the large virulence plasmid pLVPK [108]. The rmpA2 on pLVPK was shown to act as a transcriptional activator for the cps gene cluster expression, by binding to the putative promoters, Porf1-2 and Porf3-15 [109]. Interestingly, recent study revealed Fur represses the expression of rmpA2
in an Fe(II)-dependent manner [20], which implying the availability of iron affects CPS biosynthesis.
In addition to the CPS biosynthesis, expression of the major adhesion determinant type 3 fimbriae is also iron dependent. A previous study in K. pneomonniae CG43 showed that the major pilin of type 3 fimbriae MrkA and biofilm formation is depend on extracellular iron availability. MrkA expression and biofilm formation was reduced gradually by the increasing concentration of iron chelator 2,2’-dipyridyl (DIP) [26]. Most K. pneumoniae isolates expressed two types of fimbriae adhesins, type 1 and type 3 fimbriae [105]. Type 1 fimbria are virtually found in all
Enterobacteriaceae members, they mediate adhesion to mannose-containing structure on host
cells and their expression is phase variable, which is mediated by an invertible DNA element (fim switch) [110]. Type 3 fimbria are present in nearly all K. pneumoniae isolates and mediate adhesion to several cell types in vitro [111, 112]. However, the specific receptor for type 3 fimbriae has not been identified yet [113]. The adhesion filaments encoded by the mrkABCDF operon [114] in which MrkA and MrkF are the major and minor pilin protein, respectively [115, 116].
Iron affects biofilm formation in a variety of bacteria [117-120]. The role of type 3 fimbriae in biofilm of K. pneumoniae has been continually demonstrated [26, 113, 121-124]. Biofilm is recognized as surface-attached bacteria embedded in a self-produced matrix, composed mainly of
13
polysaccharide, but also containing proteins and nucleic acids [125]. Therefore, its strong coating provide protection to bacteria from hydrodynamic forces of urine flow, host defense and antibiotic [126]. The ability of bacteria to form biofilm on medical devices is believed to play a major role in development of nosocomial infections, including the K. pneumoniae caused catheter-associated urinary tract infections [126].
As analyzed by Lin et al. [20], homologues of ten genes encode for eight different iron-acquisition systems in K. pneumoniae NTUH-K2044 were identified in K. pneumoniae CG43 using Blast search program (http://blast.ncbi.nlm.nih.gov/Blast.cgi), as listed in Appendix 1. Increased expression of these genes in fur deletion background further revealed the negative regulation of Fur on them.
1.5 Specific aims
The main objective of this study is to characterize the individual role of the ferrous iron transport system FeoABC, SitABCD and EfeUOB in K. pneumoniae CG43. The deletion mutants of the presumed important ferrous iron acquisition genes feoB, sitCD, and efeUOB are generated. The phenotype of these bacterial mutants are determined by examining the gene deleting effects on the expression of iron-related virulence factors such as CPS, resistance to oxidative stress, biofilm, and type 3 fimbriae are analyzed. In addition, different combinations of the gene deletion of the three ferrous iron transporters are included to investigate their cumulative impact. Moreover, deletion feoA and feoC mutants are also generated for their regulation in feo system.
14
2.0 Materials and Methods
2. 1. Plasmid, primers, bacterial strains and growth conditions
The bacterial strains, plasmids and primers used and constructed in present study are describe in Table 1, Table 2 and Table 3 respectively. K. pneumoniae CG43 is clinical isolated strains from Chang Gung Memorial Hospital Linkou branch. E. coli and K. pneumoniae strains were generally cultured aerobically at 37 °C in Luria-Bertani (LB) broth or on M9 minimal medium supplemented with appropriate antibiotics. For static culture, 1: 20 dilution of overnight culture was added to fresh media and incubated for 20 hr. Otherwise indicated, the concentrations of antibiotics used include streptomycin (500 µg/ml), ampicillin (100 µg/ml), chloramphenicol (35 µg/ml), and kanamycin (25 µg/ml). The iron chelators added in media include deferoxamine mesylate salt (DFX, Sigma Aldrich), 2, 2’-dipyridy (DIP, Sigma Aldrich) and disodium salt dehydrate (EDTA, USB cooperation) with final concentration of 200 µM.
2. 2. DNA manipulation
Plasmids was purified by using High-Speed Mini kit (Geneaid). All DNA-modifying and –restriction enzymes was recommended by the manufacture (Fermentas). PCR amplifications were performed with Blend Taq DNA polymerase (TOYOBO) or Taq DNA polymerase (MDbio Inc), PCR products and DNA fragments were purified using the Gel/ PCR DNA Fragments Extraction it (Geneaid). The primers used in this study were synthesized by MDBio, Inc or Integrated DNA technology (IDT).
15
2. 3. Bioinformatics analysis
Homology search analysis and gene annotation were performed with the BLAST program provided by NCBI (http://www.ncbi.nlm.nih.gov) or vector NTI (Invitrogen Vector NTIM advance). Promoter prediction was carried out by SofBerry provider (http://www.softberry.com).
2. 4. Construction of gene-deletion mutants
The deletion strains constructed in this study are listed in Table 1. All the strains were made by allelic exchanged. Approximately 1000-bp sequences flanking both sides of the deleted region
(feoA, feoC, sitCD, and efeUOB) were PCR amplified with respective primer pairs (Table 3). The
amplified DNA fragments were then cloned into plasmid pKAS46 [127], a suicide vector containing rpsL, which allows positive selection with streptomycin for loss of the vector, to generate an in frame deletion plasmid. The resulting plasmid was then mobilized to K. pneumoniae CG43S3, K. pneumoniae CG43S3 ∆lacZ [128], K. pneumoniae CG43S33 fur [19] or K.
pneumoniae CG43S3-derived strains, through conjugation from E. coli S17-1 λpir. The
transconjugants, which carried constructed plasmid integrated in the chromosome via homologous recombination, were selected by ampicillin and kanamycin on minimal media (M9). Several of the colonies was grown in LB at 37 °C for 8 hours and then spread onto a LB plate containing streptomycin. The streptomycin-resistant and kanamycin-sensitive colonies were selected and verified by PCR and deletion of gene verified by PCR by designed primer sets described in Table 3.
16
2.5. Construction of a pLacZ reporter system
The putative promoter regions of feoABC (named as Pfeo1 and Pfeo2), sitABCD (named as Psit) and efeUOB (named as Pefe) were PCR amplified from CG43S3 by the designed primer pairs (Table 3) and subcloned into placZ15 [128] to fuse them with promoterless lacZ reporter system. The promoter-reporter plasmids placZfeo1, placZfeo2, placZsit, and placZefe, were individually mobilized into K. pneumoniae strains by conjugations from E. coli S17-1 λpir.
2.6. Measurement of bacterial growth in iron depletion or repletion conditions
Cultures of the parental strains CG43S3, along with deletion mutant strains were grown overnight in LB. 1:200 of diluted overnight cultures were inoculated into LB or LB with iron-depleted or iron-replete conditions. Iron-depletion condition was created by adding DFX or DIP to a final concentration of 200 µM; iron-repletion condition was created by adding ferric sulfate [(Fe)2(SO4)3] and ascorbic acid to a final concentration of 50 µM into LB broth. The cultures were
incubated at 37 °C agitatedly and the optical density was recorded every hour as the absorbance at 600 nm (OD600).
2.7. Measurement of promoter activity through β-galactosidase activity assay
β-galactosidase activity was determined according to the method of Miller [129]. In brief, overnight culture was diluted 1 : 20 in LB broth supplemented with appropriate antibiotic and incubated at 37 °C until it reached the lag phase (OD600 = 0.2), early (OD600 = 0.5-0.6) logarithmic,
late logarithmic (OD600 = 0.8-0.9) or stationary growth phase (OD600 >1.0). 100 µl of the bacterial
culture was added with 900 µl of Z buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1
17
vigorous shaking and incubated for 10 min at 30 °C. Subsequently, 200 µl of 4 mg/ml o-nitrophenyl-β-galactopyranoside (ONPG) was added and mixture was mixed thoroughly with vortex for 3 s, then incubated at 30 °C until yellow color was apparent. Finally, the reaction was stopped by adding 500 µl of stop solution (1 M Na2CO3) and the absorbance the supernatant was
measure at OD420.The activity was expressed as Miller units in which one unit of β-galactosidase
is defined as the hydrolysis of 1 nmol ONPG per min per mg or protein. Each sample was assayed in triplicate, and at least 3 independent experiments were conducted. The data shown were calculated from one representative experiment, and are presented as the means and standard deviations from triplicate samples.
2.8. Sedimentation test
The capsular polysaccharide biosynthesis was analyzed by sedimentation test. The bacterial strains were cultured overnight in LB broth at 37ºC and subjected to centrifugation at 4,000 x g for 5 min. Pictures of bacterial cells centrifuged by low speed sedimentation of were taken.
2.9. Biofilm formation assay
Overnight grown bacteria were diluted 1: 100 in LB broth supplemented with appropriate antibiotic and then inoculated 150 µl into each well of a 96-well microtiter dish (DPP), and propagated statically at 37°C for 24 hr. After removal of the bacteria, each well was washed by de-ionized water twice before added with150 μl of 1% (w/v) crystal violet and placing on an orbital shaker for 50 min at 60 rpm. After washing each well three times with de-ionized water, 200 µl
18
95% ethanol was added to solubilize the dye for 1 hr. The capability of biofilm formation was quantified by the absorbance at 595 nm (ELx800, BIO-TEK).
2.10. H2O2 sensitivity test
Sensitivity of deletion strains to H2O2 stress was determined by disk diffusion method on
LB plate and LB plate added with 0.2 mM of different iron chelators (DFX, DIP or EDTA). Overnight cultures were diluted 1:20 to a fresh LB grown agitatedly to an OD600 of 0.6-0.7. The
cultures were then supplemented with or without iron chelators for another 1 hr. Filter paper disks (6-mm diameter; Becton Dickinson) spotted with 5 µl hydrogen peroxide (30 %) were added onto the disks. The plates were then incubated at 37 °C for 16 hr. The diameters of inhibition zones were measured. The effect of iron chelators (deferoxamine, DIP and EDTA) was determined by its addition 1 hr prior to that H2O2.
2.11. Western blot analysis of the expression of type 3 fimbriae
Total cellular lysate from the bacteria were resolved by 12 % SDS-PAGE to determine the expression of type 3 fimbriae in CG43S3. The electrophoretic proteins were then transferred onto polyvinylidene difluoride (PVDF) membrane (Milipore, Billerica, MA, USA). After incubation with 5 % skimmed at 4 °C overnight, the membrane was washed three times with 1x Tris-buffered saline (TBS). Subsequently, the membrane was incubated at room temperature for 2 h with 25,000 fold diluted anti-MrkA serum. After washed three times with 1x TBS, a 5,000 fold diluted alkaline phosphate-conjugated immunoglobulin G (IgG) was added and the incubation continued for 1h. The bound antibodies were detected by chromogenic reagent BCIP (5-bromo-4-chloro-3-indolyl
19
phosphate), NBT (Nitro blue tetrazolium) and alkaline phosphatase butter (100mM NaCl, 5 mM MgCl2, 100 mM Tris-HCl pH 9.5).
20
3.0 Results
3.1. Generation of the specific gene deletion mutants
The coding sequence of feoA (290-bp), feoB (2297-bp), and feoC (266-bp) were individually deleted by allelic exchange method, and the deletions were confirmed by PCR analysis. Besides ∆feoA, ∆feoB, and ∆feoC, the double mutation mutants including ∆feoA∆fur,
∆feoB∆fur, and ∆feoC∆fur, were also obtained since Fur negatively regulates the iron uptake
systems. As shown in Fig. 1A, the specific primer pairs are delFeoA-check(+)/delFeoA-check(-) for the deletion of feoA, delFeoB-check(+)/delFeoB-check(-) for feoB deletion, and delFeoC-check(+)/delFeoC-check(-) for feoC deletion, respectively. The expected sizes of the PCR products are respectively 590-bp for feoA deletion (Fig.1B), 386-bp for feoB deletion (Fig.1C), and 870-bp forfeoC deletion (Fig. 1D).
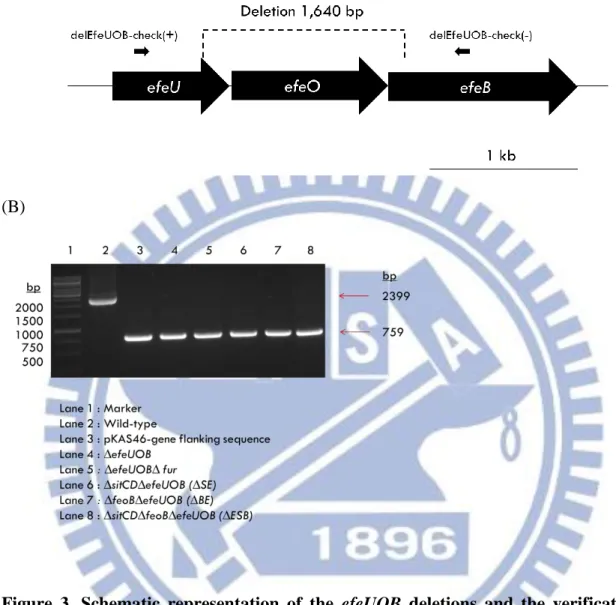
Fig. 2 and 3 respectively show the position of the designed primer pairs delSitCD-check(+)/delSitCD-check(-) and delEfeUOB-check (+)/delEfeUOB-check (-), and PCR analysis employed to confirm the sitCD and efeUOB deletion. A total of 1437-bp sitCD- and 1640-bp
efeUOB-containing DNA was deleted. The derived mutants are ∆sitCD∆fur, ∆sitCD∆fur, ∆sitCD∆feoB (∆SB), ∆efeUOB∆sitCD (∆SE), ∆efeUOB∆fur, ∆efeUOB∆feoB (∆BE), and ∆efeUOB∆sitCD∆feoB (∆ESB). As shown in Fig. 2(B) and Fig. 3(B), the amplicon of 546-bp for sitCD deletion and 759-bp for efeUOB deletion were obtained.
3.2. Effect of iron availability on the growth of the specific gene deletion strains
To investigate the effect of iron availability on these iron-acquisition systems, the growth of CG43S3 along with ∆feoA, ∆feoB, ∆feoC, ∆sitCD, ∆efeUOB, ∆sitCD∆feoB (∆SB),
21
in LB, iron-supplemented or iron-depleted medium was investigated. LB medium adding with 50 µM Fe2(SO4)3 and 50 µM ascorbic acid is the iron-supplemented condition, while iron-depleted
condition was created by adding 200 µM ferrous iron chelator DIP or 200 µM ferric iron chelator DFX. All the strains grown in LB medium reached OD600 of 1.2 in 6 h (Fig. 4A) and all appeared
to grow faster when supplemented with iron (Fig. 4B). As shown in Fig. 4(C) and 4(D), the bacteria grown in medium containing DIP or DFX grew slower compared to the growth in LB or LB loaded with iron. Interestingly, the deletion mutants grew slightly slower than the parental strain CG43S3 only in the LB medium added with DIP. Otherwise, they exhibited similar growth pattern.
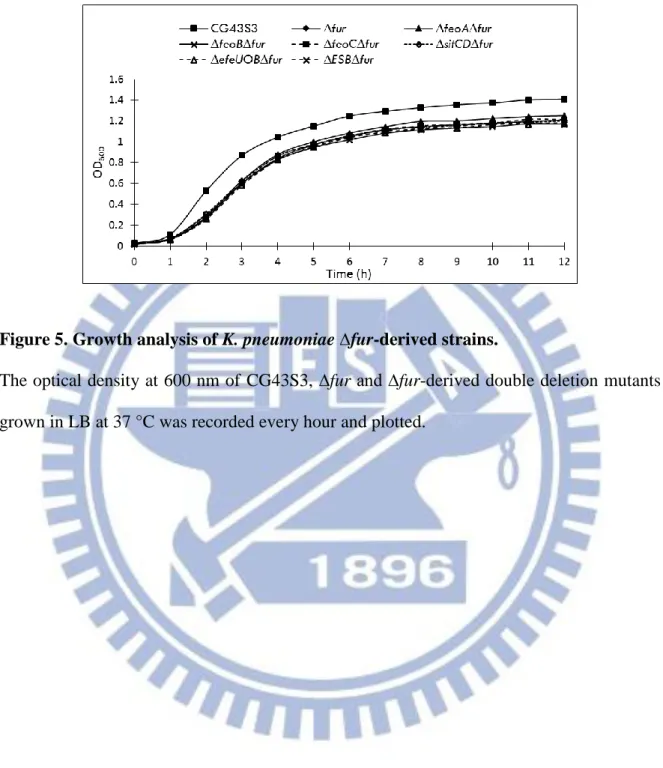
As shown in Fig. 5, the growth of double deletion strains ∆feoA∆fur, ∆feoB∆fur, ∆feoC∆fur,
∆sitCD∆fur, ∆efeUOB∆fur and ∆feoB∆sitCD∆efeUOB∆fur (∆ESB∆fur) had similar growth
pattern as ∆fur but grew slower than CG43S3 in LB medium. The ∆fur-derived strains reached OD600 of 0.25 while CG43S3 of 0.5 after 2 h growth.
3.3. Analysis of feoABC, sitABCD and efeUOB promoter region
As shown in Fig. 6(A), 6(B), and 6(C), a Fur box-like sequence could be identified in the putative promoter regions of feoABC, sitCD and efeUOB. Besides, other regulatory sequences predicted using Softberry are respectively RstA- and Fnr-box within Pfeo, ArcA- and MntR-box in Psit, and CpxR box in Pefe. The presence of Fnr-box in feoABC and ArcA-box in sitABCD promoter region suggests oxygen availability affects the expression of feoABC and sitABCD. However, whether Fnr regulates expression of feoABC remains unknown since the Fnr binding sequence overlaps with the Fur box in Pfeo. The manganese repressor MntR binding box (AAACATAGCN4GCTATGTTT) is located 81-bp upstream of the start codon of SitA. The
22
predicted CpxR box (GTAAAN4-8GTAAA) identified in efeUOB promoter region implies a
negative control by CpxR as reported in E. coli [46].
3.4. Effect of growth phase and oxygen availability on the expression of feo, sit and efe
To examine the promoter activity, the DNA fragments encompassing the putative promoter regions of feoABC, sitABCD and efeUOB were individually cloned in front of the promoterless
lacZ to generate pPfeo1-lacZ, pPfeo2-lacZ, pPsit-lacZ and pPefe-lacZ.
As shown in Fig. 7A, the activities of Pfeo, Psit and Pefe gradually increased following their growth. The activity of Pfeo, Psit and Pefe in stationary phase showed approximately 1.5-fold higher than in late log phase. Among these three Fe(II) uptake systems, sit exhibits the highest expression, following by efe and feo. It is interesting to note that Pfeo1 which contains two predicted 10 and -35 promoters had a much lower activity than Pfeo2 implying the presence of repressor binding element in the region only contained by Pfeo1.
Since Fnr and ArcA binding box are found respectively in promoter region of feo and sit, activity of Pfeo, Psit and Pefe was assessed under static condition (Fig. 7B). In compared to shaking condition, static condition did not apparently increase or decrease the activity of both Pfeo1 and Pfeo3. Interestingly, Psit::lacZ expression under static culture showed as high as 2000 Miller units in compared with 1000 Miller unit observed under shaking culture (stationary phase). In contrast, Pefe::lacZ expression was decreased under the microaerobic condition, showing merely 250 Miller unit of activity. These results supported the possibility that oxygen availability may affect the transcriptional level of sit and efe in adverse manner.
23
3.5. Effect of fur deletion and iron-depletion on expression of feo, sit, and efe
The effect of fur deletion on the expression of the feo, sit and efe was further verified by promoter activity assay. As shown in Fig. 8A, the activity of Pfeo1, Pfeo2, Psit and Pefe were apparently induced in Z01∆fur strain. In the meanwhile, Pfeo1, Pfeo2, Psit and Pefe activity were increased in response to addition of iron chelators (Fig. 8B). Interestingly, the inducible expression of feo was more effective by DFX than by DIP. On the other hand, both sit and efe promoters were more responsive to DIP than to DFX.
3.6. Effect of feoC deletion and rstA deletion and on expression of feo, sit, and efe
To investigate whether the two-component system response regulator RstA regulates feo expression, deletion effect of rstA on activity of Pfeo1 and Pfeo2 was examined. The result from left panel of Fig. 9(A) showed no apparent effect on Pfeo1 and Pfeo2 by rstA deletion. RstA has been shown to be involved in the acid stress response by regulating asr (acid shock RNA) expression in Salmonella [77], weak acid was treated to examine whether Pfeo is also acid-inducible. However, as shown in right panel of Fig. 9A, both Pfeo1 and Pfeo2 activity were not affected by acid.
Feo system is the first ferrous iron transport system identified in Enterobacteriae, and
FeoB is well known as the major ferrous iron transport system in many bacteria, but the role of FeoA and FeoC are not well studied thoroughly. FeoC was proposed to regulate Fe(II) uptake of FeoB by coordinating its iron-sulfur cluster [3, 55, 62]. In order to clarify the regulatory role of FeoC in feo expression, activities of Pfeo1 and Pfeo2 were studied in Z01∆feoC strain under different environmental conditions (shaking, static, iron-depleted and iron-supplemented culture). As shown in Fig. 9B (right panel), the promoter activity of Pfeo1 and Pfeo2 was induced by feoC deletion
24
when bacterial strains were cultured in LB supplemented with DFX under static condition. By contrast, the increased activity was not observed under shaking condition (left panel).
3.7. Effect of manganese on expression of feo, sit and efe.
MntR is a manganese repressor using Mn2+ as a cofactor to regulate Mn2+-related uptake systems. To investigate whether Psit is negatively regulated by MntR, the culture of CG43S3∆lacZ (Z01) was treated by 0.1mM MnCl2 and the activity of Pfeo, Psit and Pefe were examined. Interestingly, no suppressing effect was observed in Pfeo, Psit or Pefe, instead, their activities were induced by manganese (Fig.10).
3.8. Effect of weak acid and cpxR deletion effect on the efe expression
In E. coli O157:H7, expression of efeU was low-pH induced while repressed by CpxAR at high pH. The CpxR box (GTAAAnnnnnnnnGTAAA) found in promoter region of efeUOB suggests a possible role of CpxR in regulation of efeUOB in CG43. To investigate if efeUOB expression in CG43 works similar with O157:H7, Z01 and Z01∆cpxAR carrying Pefe-lacZ were grown in pH 7 or pH 5 LB media (Fig. 11). The expression of Pefe in Z01 at pH 5 was 4-fold higher than at pH 7. Deletion of cpxAR increased Pefe-lacZ expression under both pH 7 and pH 5 media, suggesting CpxR negatively regulates the expression of efeUOB under both acid and neutral pH conditions.
3.9. Analysis of cumulative effect of acid and iron-depletion or iron-supplemented on growth
From the result of promoter activity, acid and iron play a regulatory role on the expression of these specific iron-acquisition genes. Growth curve analysis of all the mutants cultured in pH 5
25
LB with additional Fe or Fe-depleted condition was performed. As shown in Fig. 12 (A) and (B), all growth curve of the mutant strains appeared similarly as the parental strain CG43S3 in pH 5 LB or with additional Fe. However, an apparent growth defect was observed when the mutants grown in pH 5 LB supplemented with DIP. As Fig. 12(C) shows the mutant strains displayed more acute slow-growing phenomenon of which the OD600 could only reach 0.8 at stationary phase. This
may be explained by no sufficient ferrous iron Fe(II) for the bacterial growth. No growth change was observed when Fe(III) iron chelator DFX was added to the cultures (Fig. 12D).
3.10. Analysis of ferrous iron transporter regulation in CPS biosynthesis
The ∆fur mutant strain exhibited profound CPS biosynthesis and hence the bacterial culture was precipitated much slower than CG43S3 [19]. The Fe(II)-associated Fur metalloprotein requires iron as cofactor to regulate expression of various genes. To investigate whether Feo, Sit or Efe system acts as the major source of Fe(II) for Fur, CPS production of the strains were analyzed by subjecting the overnight-grown bacteria to low-speed centrifugation. As shown in Fig. 13(A), the CPS formed by the mutant strains showed no apparent change compared to that of the parental strain CG43S3 and CG43S3∆fur, respectively. Fig. 13(B) shows that the mutant strains grown in LB with DFX displayed more viscous phenotype than those grown in LB media and hence could not be easily precipitated down. On the other hand, the addition of DIP resulted in poor growth of all the bacteria and the bacteria could be readily precipitated (Fig. 15C). Nevertheless, no apparent of change of the phenotype in comparing with that of the parental strain CG43S3 suggesting FeoABC, SitABCD or EfeUOB transport systems does not directly influence the CPS biosynthesis.
26
3.11. Analysis of deletion effects on oxidative stress response
Fe(II) could be hazardous to bacteria because of the Fenton reaction leading to ROS formation. As reported in many other bacteria, iron transport systems play a role in oxidative stress responses [12, 83, 85]. To investigate if any of the three iron transport systems is also involved, disk diffusion assay was employed with the disc immersed with 30% hydrogen peroxide (H2O2).
The effect of iron chelators (DFX, DIP and EDTA) on the oxidative stress response was also tested. By measuring the diameters of inhibition zones of the bacteria grown agitatedly in LB,
∆sitCD∆efeUOB (∆SE), ∆sitCD∆feoB (∆SB), and ∆efeUOB∆sitCD∆feoB (∆ESB) showed
increased sensitivity to H2O2 compared to CG43S3 (Fig. 14A). When comparing the H2O2
inhibition zone with that of CG43S3, only ∆feoC exhibited more susceptibility when the culture added with DIP (Fig. 14B) while ∆feoB, ∆sitCD and ∆efeUOB were more sensitive by the addition of DFX (Fig. 14C) than double deletion and triple deletion mutant strains ∆feoB∆efeUOB (∆BE), ∆sitCD∆feoB (∆SB), ∆sitCD∆efeUOB (∆SE) and ∆efeUOB∆sitCD∆feoB (∆ESB) (Fig. 14C). Fig. 14(D) showed the bacteria ∆sitCD, ∆sitCD∆feoB (∆SB) and ∆efeUOB∆sitCD∆feoB (∆ESB) which carrying deletion of sitCD displayed a more sensitive phenotype in the presence of the divalent-cation chelator EDTA.
Since FeoABC, SitABCD, and EfeUOB are repressed by Fur under LB medium, the derepressing effect of Fur on these system towards oxidative stress response was also tested. As Fig. 15 that ∆efeUOB∆sitCD∆feoB∆fur (∆ESB∆fur) is more sensitive while ∆feoB∆fur,
27
3.12. Analysis of deletion effects on the biofilm formation under different culture conditions
Iron plays an important role in bacterial biofilm formation. As shown in Fig. 16, all the deletion mutants except ∆sitCD∆efeUOB (∆SE) exhibited decreased biofilm forming activity compared to CG43S3. It is interesting to note that an apparent defect of biofilm formation was found by the deletion of fur.
Addition of DIP or DFX to the medium conferred different effects on the biofilm formation activity. As shown in Fig. 17, DIP negatively influenced the biofilm formation for CG43S3 as well as all the derived mutants. By contrast, addition of DFX allows identifying the deletion effect on the biofilm formation. Compared to CG43S3, only ∆feoC exhibited an increase of biofilm formation while the other mutants showed reduced biofilm formation in the presence of DFX. The
feoC deletion may induce the expression of feoAB thereby promote iron-uptake and increase
biofilm formation.
3.13. Analysis of the deletion effects on the expression of type 3 fimbriae
As described by Wu et. al., expression of the major pilin MrkA of type 3 fimbriae depends on the extracellular iron availability [26]. Consistent with their findings, Fig. 18(A) shows MrkA production is reduced by adding DIP or DFX to the culture, in which DIP almost abolished the MrkA expression. To investigate the role of ferrous iron uptake system on type 3 fimbriae expression, the deletion mutants along with wild-type strain were treated with different conditions and the MrkA production assessed by Western blot analysis. As shown in Fig. 18(B), ∆feoB and ∆efeUOB∆sitCD∆feoB (∆ESB) exhibited a slightly decreased production of MrkA compared with CG43S3. However, feoB deletion caused a slightly increased production of MrkA under iron
28
depletion condition (Fig. 18C). The iron depletion condition had no effect on the MrkA production of the other mutants.
We have shown that Fur positively regulates the expression of type 3 fimbriae. Whether FeoABC, SitABCD or EfeUOB is involved in this regulatory pathway is investigated. As shown in Fig. 19, ∆fur exhibited no MrkA production and further removal of either of feo, sit, or efe iron transport system could not rescue the deficiency caused by fur deletion. This implies that Fur-dependent expression is inFur-dependent to the iron-mediated expression of type 3 fimbriae.
MrkA expression was increased under microaerobic condition in CG43S3 and the derived mutant strains (Fig. 20A). In the presence of DFX, MrkA production was slightly decreased by the deletion of feoA, feoB, feoC, or efeUOB (Fig. 20B). Interestingly, ∆sitCD and ∆ESB revealed an increase of MrkA expression compared to CG43S3 (Fig. 20B).