行政院國家科學委員會專題研究計畫 期中進度報告
嗜甲基酵母菌中表現細菌血紅素以增進其生長及異源蛋白 質之表現(1/2)
計畫類別: 個別型計畫
計畫編號: NSC92-2214-E-011-017-
執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣科技大學化學工程系
計畫主持人: 李振綱
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 93 年 5 月 31 日
行政院國家科學委員會專題研究計畫 期中進度報告
研究類型:一般型研究計畫(個別型) 計畫編號:NSC 92-2214-E-011-017
執行期限:92 年 08 月 01 日 至 93 年 07 月 31 日 執行單位:國立台灣科技大學化學工程系
計畫主持人:李振綱 教授
報告類型:期中精簡報告 處理方式:本計畫可公開查詢
中華民國 九十三 年 五 月 三十一 日
嗜甲基酵母菌中表現細菌血紅素以增進其生長及 異源蛋白質之表現(1/2)
一、前言
在過去十五年中,嗜甲基酵母菌P. pastoris 已經成功地表現出各種異源蛋白 質,它因為具有下列幾個特點,所以近幾年來受到重視:(1)只需簡易的分子 基因操作即可建構完成表現系統;(2)具有表現胞內和胞外異源蛋白質的能力,
並且表現量高;(3)具有真核細胞轉譯後修飾的能力,例如醣基化、雙硫鍵和 蛋白質分解過程等;(4)無過度醣基化(hyperglycosylation)的現象,不會造 成免疫原性改變、蛋白質活性降低或是改變存在於血清中的半衰期等[Van Arsdell et al., 1987;Lemontt et al., 1985];(5)易於發酵培養達到很高的細胞濃 度;(6)所使用的表現載體是屬於插入性質體(integrative plasmid),藉由這 種質體將外來基因插入在其染色體上,可以提高外來基因的穩定性,使其在大規 模培養時不會因為質體的不穩定性而使蛋白質表現量降低[Romanos, 1995]。
當P. pastoris 用在大量生產表現異源蛋白質時,必須進行高密度的細胞培養 以達到高產率,而在此時氧氣的需求極高,因為P. pastoris 喜好呼吸生長而非嫌 氧的發酵生長。此外,代謝甲醇(用來誘導異源蛋白質表現)也需要氧氣參與氧 化反應,所以氧氣的供應對於異源蛋白質的表現是非常重要的參數[Villatte et al., 2001;Ohashi et al., 1999]。Vitreoscilla 是一株極度好氧性細菌,當它處於氧氣不 足的環境時,可以合成出血紅素(VHb)而存活並生長[Dikshit and Webster, 1988;
Khosla and Bailey, 1988]。此血紅素可能的作用是從環境中吸附氧氣,並幫助傳 送到呼吸膜上的最終氧化酵素,因而提升呼吸與 ATP 的形成。血紅素已在許多 原核細菌與真核細菌中表現,並具有提升細胞生長、蛋白質與代謝物生成的能力 [Bulow et al., 1999;Chen et al., 1994;Kallio and Bailey, 1996]。這種對氧氣有很 高的吸附與釋放能力之特性,使它成為一種非常適合在 P. pastoris 表現的蛋白 質,以解決在高密度發酵中甲醇誘導時期極度需要氧氣的問題。
二、研究目的
本年度計畫裡我們主要是以基因重組技術分別將原生種之血紅素及合成之 血紅素基因引入嗜甲基酵母菌P. pastoris 的染色體中,再探討原生種及基因合成
之血紅素VHb 基因在 P. pastoris 細胞質是否能正常表現,並且利用 CO difference spectrum 法來確認分析 VHb 的功能性。由於 VHb 能幫助菌體在為氧環境下的氧 氣的傳送,所以我們接著利用微生物發酵槽來加以確認在人工微氧環境下,VHb 對菌體生長的影響。另外,由於P. pastoris 中酒精氧化酵素對氧氣的親和力很差,
所以我們也利用基因技術將 VHb 基因透過過氧化氫體標的訊息 1(peroxisomal targeting signal 1;PTS1)表現送到過氧化氫體中,藉以提高過氧化氫體胞器的氧 溶度,而有利甲醇的代謝,進而觀察VHb 對菌體生長與蛋白質合成上的效應。
三、研究方法
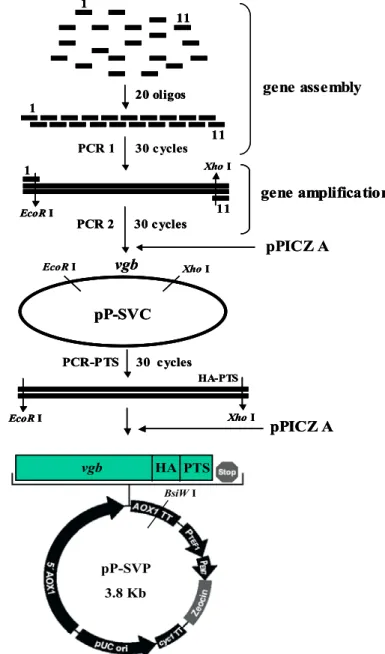
1. 人工合成 vgb 基因及表現質體之建構
首先全合成 vgb 基因的序列是利用 DNAWorks 這一套軟體[Hoover and Lubkowski, 2002],並根據 P. pastoris 的密碼使用率(codon usage)來設計出最佳 基因序列來表現蛋白。圖1.1 為設計出來 vgb 全合成的序列,合成方法[Stemmer et al., 1995]:從庫存(stock)濃度為 1 mg/ml 的寡核苷酸中取等量體積混合後,
用水稀釋到每一條寡核苷酸最終濃度是1 ng/µl,此寡核苷酸混合物用 PCR 溶液 再稀釋五倍使用。最後的組成是每一寡核苷酸0.2 ng/µl,20 mM Tris-HCl(pH 8.8),10 mM KCl,10 mM (NH4)2SO4,6 mM MgSO4,0.1 ﹪(v/v)Triton X-100,
0.1 mg/ml BSA,0.2 mM dNTP,2.5 U pfu polymerase。基因組合的 PCR 條件是 95 ℃ 5 分鐘(”hot start” PCR)→ 95 ℃ 1 分鐘→ 72 ℃ 1 分鐘,進行 30 個循 環,最後再進行72 ℃ 10 分鐘,其 DNA 電泳分析圖如圖 1.2。接著取 1 µl PCR 產物當作模板,使用最外側兩端之寡核苷酸作為引子,來進行基因放大,除了將 黏合溫度提高至58 ℃外,其餘條件與基因組合相同,最後可得到全長的基因,
如圖1.2 所示。將合成出的 vgb 片段,經純化與 EcoRI、XhoI 作用後,接入載體 pPICZA 中,基因序列確認無誤後,得到 VHb 表現於細胞質的質體 pP-SVC(如 圖1.3)。
為了建構 VHb 在過氧化氫體表現的質體,因此 VHb-HA-PTS1 的基因便可 以用5’ CCG GAA TTC ATG TTG GAT CAA CAG ACT ATC AAC A 3’與 5’CCG CTC GAG TTA CAA CTT AGA AGC GTA ATC TGG AAC ATC GTA TGG GTA TTC AAC AGC TTG AGC GTA CAA ATC AGC 3’引子以 pP-SVC 為模板經 PCR
放大後即可獲得。經萃取與純化後,用 EcoRI 與 XhoI 作用後,接入 pPICZA,
經過篩選與確認基因序列後,便命名為pP-SVP(如圖 1.3)。
Figure 1.1 Nucleotide sequence of the synthetic vgb gene according to preferred codons of P. pastoris.
1 CCGGAATTCATGTTGGATCAACAGACTATCAACATCATCAAGGCTACTGTTCCAGTTTTG 60 CAACCTAGTTGTCTGATAGTTGTAGTAGTTCCGATGACAAGGTCAAAAC 61 AAGGAACATGGTGTTACTATCACTACTACTTTCTACAAGAACTTGTTTGCTAAGCATCCA 120 TTCCTTGTACCACAATGATAGTGATGATGAAAGATGTTCTTGAACAAACGATTCGTAGGT 121 GAAGTTAGACCATTGTTTGATATGGGTAGACAAGAATCTTTGGAACAACCAAAGGCTTTG 180 CTTCAATCTGGTAACAAACTATACCCATCTGTTCTTAGAAACCTTGTTGGTTTCCGAAAC 181 GCTATGACTGTCTTGGCTGCTGCTCAGAACATTGAGAACTTGCCAGCTATCTTGCCAGCT 240 CGATACTGACAGAACCGACGACGAGTCTTGTAACTCTTGAACGGTCGATAGAACGGTCGA 241 GTTAAGAAGATTGCTGTTAAGCATTGTCAAGCTGGTGTTGCTGCTGCTCATTACCCAATT 300 CAATTCTTCTAACGACAATTCGTAACAGTTCGACCACAACGACGACGAGTAATGGGTTAA 301 GTTGGTCAAGAATTGTTGGGTGCTATCAAGGAAGTCTTGGGTGATGCTGCTACTGATGAT 360 CAACCAGTTCTTAACAACCCACGATAGTTCCTTCAGAACCCACTACGACGATGACTACTA 361 ATCTTGGATGCTTGGGGTAAGGCTTACGGTGTTATTGCTGATGTCTTCATTCAAGTTGAA 420 TAGAACCTACGAACCCCATTCCGAATGCCACAATAACGACTACAGAAGTAAGTTCAACTT 421 GCTGATTTGTACGCT 459 CGACTAAACATGCGAGTTCGACAACTTATTGAGCTCGCC
1 2
3
4
5
6 7
8
9 10
11
12 14 13
15
16 17
18 19
20
Figure 1.2 Agarose electrophoresis analysis of vgb gene synthesis.
Figure 1.3 Construction of the VHb expression vector pP-SVC and pP-SVP starting from assembly of synthetic VHb gene fragments.
0.5 kb
Assembly PCR
Amplification
0.5 kb 0.5 kb
Assembly PCR
Amplification
1
11
20 oligos 1
PCR 1 30 cycles 11
gene assembly
gene amplification
1
EcoR I 11
Xho I
Xho I EcoR I
pP-SVC vgb
PCR 2 30 cycles
pPICZ A
PCR-PTS 30 cycles HA-PTS
EcoR I Xho I
pPICZ A vgb HA PTS
pP-SVP 3.8 Kb
vgb HA PTS
pP-SVP 3.8 Kb
BsiW I 1
11 11
11 11
20 oligos 20 oligos 1
11 1
PCR 1 30 cycles 11
gene assembly
gene amplification gene amplification
1
EcoR I 11
Xho I 1
EcoR I 11 EcoR I
Xho I
Xho I EcoR I
pP-SVC
vgb Xho I
EcoR I
pP-SVC vgb
PCR 2 30 cycles PCR 2 30 cycles
pPICZ A
PCR-PTS 30 cycles HA-PTS
EcoR I Xho I
pPICZ A
PCR-PTS 30 cycles HA-PTS
EcoR I
EcoR I Xho IXho I
pPICZ A vgb HA PTS
pP-SVP 3.8 Kb
vgb HA PTS
pP-SVP 3.8 Kb
BsiW I
vgb HA PTS
pP-SVP 3.8 Kb
vgb HA PTS
pP-SVP 3.8 Kb
BsiW I
2. 血紅素活性分析
以CO difference spectrum 法分析,方法如下:加入少量的亞硫酸鈉(sodium hydrosulfite)還原劑於含有 VHb 的溶液中並均勻混合,再通入一氧化碳約 30 秒,
並利用可見光光譜分析儀從400 ~ 500 nm 進行偵測。而對照組則是不通入一氧化 碳,其餘部份相同。
3. 電穿孔轉殖法
電穿孔轉殖法的進行如下:首先製備P. pastoris 勝任細胞(competent cells),
在含有5 ml YPD(1 ﹪yeast extract, 2 ﹪peptone, 2 ﹪dextrose)培養液的 50 ml 錐形管中培養P. pastoris 過夜。取 0.1 ∼ 0.5 ml 菌液接種於含有 250 ml 新鮮 YPD 培養液之2 公升錐形瓶中,使 P. pastoris 培養至 OD600值為1.3 ∼ 1.5,約需 12 ∼ 18 個小時。將菌液於4 ℃1500 × g 離心 5 分鐘,再以 250 ml 冰冷無菌水回溶,而 後再於4 ℃1500 × g 離心 5 分鐘,以 125 ml 冰冷無菌水回溶。再於 4 ℃離心 1500 × g 5 分鐘,以 10 ml 冰冷 1 M 山梨糖醇(sorbitol)回溶;再於 4 ℃離心 1500 × g 5 分鐘,而後以 0.5 ml 冰冷 1 M 山梨糖醇回溶,使最後體積約為 0.75 ml;如此便成為 P. pastoris 勝任細胞。之後取 80 µl 的 P. pastoris 勝任細胞與 5 ∼ 20 µg 以限制酶切割成線狀的表現載體混勻後置入冰浴過的電穿孔轉殖專用管
(cuvette)中;於冰上放置 5 分鐘,準備進行電穿孔轉殖。電穿孔轉殖法所利用 的儀器為Bio-Rad 公司 Gene Pulser 機型,而選擇內建的 P. pastoris 電穿孔轉殖參 數進行電擊,馬上加入1 ml 冰冷 1 M 山梨糖醇,而後將內容物移至滅過菌之 1.5 ml 離心管中,於室溫下培養 2 小時,取 200 ∼ 600 µl 菌液塗在適當的抗生素 Zeocin 的 YPD 固態培養基上,於 30 ℃培養 2 ∼ 4 天直到菌落出現。因為轉型株有抗 Zeocin 基因在,故可以在含有 Zeocin 的 YPD 固態培養基上挑選,逐步提高 Zeocin 的濃度由0.5,1.0,1.5 至 2.0 mg/ml,最後從含 2.0 mg/ml Zeocin 的 YPD 固態培 養基上挑選出來當作VHb+菌株,以供VHb 共同表現實驗用。依照同樣的程序,
不含VHb 的質體 pPICZA 也轉殖入 GS115/ZA,而得到VHb-菌株。
4. 培養條件
在50 ml 離心管中裝入 15 ml 的 YPD 或 YPM 培養基,於 30℃轉速為 250 rpm 的培養箱中培養。若欲進行微氧培養,就將螺紋蓋轉緊,每24 小時鬆開蓋子通 氣 3 小時,若進行有氧培養,則將螺紋蓋蓋上但不轉緊即可。在 YPM 與 YPD 培養時,每 24 小時加入甲醇使其最終濃度為 0.5 ﹪(v/v)。細胞密度的測定是 利用分光光度計,測量波長為600 nm 之吸光值測得。在培養期間,固定時間取 樣並保存於-20℃以便進一步分析。
在微生物發酵槽的低溶氧實驗中,主要利用發酵槽,以通氣量0.416-vvm、
轉速500rpm、溫度 30℃及 pH 5.5 的微氧環境,並且每 24 小時加入甲醇使其最 終濃度為 0.5 ﹪(v/v),接續培養 Pichia pastoris pP-SVC 轉型菌株與 Pichia pastoris pPICZA 轉型菌株三天。在培養期間,細胞密度的測定是利用分光光度 計,測量波長為600 nm 之吸光值測得。
5. 萃取 P. pastoris 染色體 DNA
在10 ml 的 YPD 培養基中,培養條件為 30℃,轉速為 250 rpm,培養至 OD600
約到達5 後,以 1500×g 離心 10 分鐘,用 10 ml 的無菌水清洗一次,再經過一次 離心之後,細胞回溶在2 ml 的 SCED 緩衝溶液(1 M sorbitol, 10 mM sodium citrate, pH 7.5, 10 mM EDTA, 10 mM DTT),加入 0.1 mg 的 Zymolyase,在 37℃中反應 50 分鐘達到約 80﹪的 spheroplasting。此時加入 2 ml 的 1 ﹪SDS,緩慢的混合並 置於冰上5 分鐘,再加入 1.5 ml 5 M 的醋酸鉀(pH 8.9),緩緩的混合後立即以 10,000×g 在 4℃離心 10 分鐘。將上清液取出後,加入兩倍體積的酒精,在室溫 下靜置15 分鐘。以 10,000×g 在 4℃下離心 20 分鐘後,緩慢的用 0.7 ml 的 TE 緩 衝液(10 mM Tris-HCl, pH 7.4, 1 mM EDTA, pH 8.0)回溶沉澱物。用等體積的 phenol:chloroform(1:1 v/v)緩和的萃取,再經一次等體積的 chloroform:isoamyl alcohol(24:1 v/v)萃取 DNA 後,小心取出水層相,並加入 1/2 體積的 7.5 M 醋酸胺(pH 7.5)與兩倍體積的酒精,於-20℃靜置 1 小時。以 10,000×g 在 4℃下 離心20 分鐘後,用 1 ml 70 ﹪酒精清洗沉澱物,經風乾後,用 50 µl 的 TE 緩衝
液回溶,並定量之。
6. 初步分離過氧化氫體
在500 ml 的 YPD 中,於 30℃過夜培養,以 1500×g 離心 10 分鐘,再用無菌 水清洗細胞一次,回溶細胞在2 L 的 YPM 中,使 OD600從0.5 開始培養,讓細 胞生長約16 小時使之誘導並增生過氧化氫體,此時 OD600約為2,以 1500×g 離 心10 分鐘,以 30 ml TEM 緩衝液(100 mM Tris-HCl, 50 mM EDTA, 200 mM β-mercaptoethanol, pH 7.5)回溶細胞,於室溫靜置 20 分鐘,再以 1500×g 離心 10 分鐘後,用30 ml 的 KPi/sorbitol 緩衝液(10 mM potassium phosphate buffer, pH 7.4, 1.2 M sorbitol)清洗一次,並再次以 30 ml 的 KPi/sorbitol 緩衝液回溶細胞,加入 5 mg 的 Zymolyase,在 37℃中反應 45 分鐘。以 1500×g 離心 10 分鐘,收集 spheroplasts 並用 20 ml 冰 Dounce 緩衝液(5 mM morpholinoethanesulfonic acid
(MES), pH 6.0, 0.5 mM EDTA-Na, 1 mM KCl, 0.1﹪ethanol, 0.8 M sorbitol, 1 mM PMSF)回溶之,倒入 Dounce 均質器(homogenizer)後,以 10 次來回推送來 打破spheroplasts,在 4℃下以 1500×g 離心 10 分鐘,將沒打破的 spheroplasts 與 細胞殘骸離心下來,而上清液則以3,000×g 在 4℃下離心 10 分鐘,將粒腺體離心 下來,然後其上清液再一次離心(20,800×g, 4℃離心 30 分鐘),將過氧化氫體離 心下來,而上清液則為細胞質液[Monosov et al., 1996]。
四、實驗結果與討論
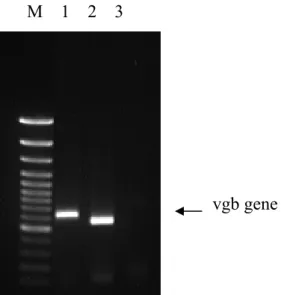
1. 人工合成與原生種 VHb 基因的建構與轉型株之確認
透過基因重組技術我們成功地建構出人工合成及原生種的 VHb 表現質體,
並且也成功地表現於嗜甲基酵母菌P. pastoris 中,並且透過染色體萃取之基因篩 選也確認 VHb 以融合入 P. pastoris 染色體中,如圖 1.4 所示,並且也經由 P.
pastoris 菌株的表現證實 VHb 可透過甲醇的誘導而表現。
M 1 2 3
Figure 1.4 Phenotype analysis of P. pastoris peroxisome VHb (lane 1),cytosome VHb(lane 2) ,and host strain based on VHb gene existence in their chromosome by PCR method.
2. VHb 對細胞生長的影響
在 VHb 表現於細胞質之菌株與對照組菌株經過微氧發酵槽培養三天之後,
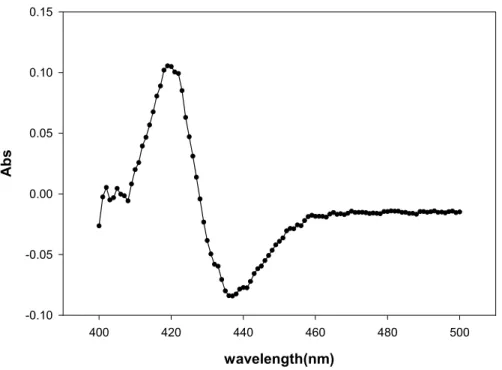
我們可以明顯得知在有VHb 表現的菌株培養後其細胞濃度較對照組來的高(如圖 1.5 所示),並且在相較於對照組菌株的本身顏色上,我們也可看出有 VHb 表現 的菌株其顏色會呈現粉紅顏色,而且透過VHb CO difference spectrum 活性的分 析也可證明VHb 在 P. pastoris 細胞質中可被表現出來(如圖 1.6 所示),並且在微 氧環境下可增進細胞的生長。
vgb gene
Time(hr)
0 6 12 18 24 30 36 42 48 54 60 66 72
OD600
0 10 20 30 40 50
VHb- strain VHb+strain
Figure 1.5 Growth curve of the VHb+({)and VHb-(z)cytosol strain cultivated in YPD.
wavelength(nm)
400 420 440 460 480 500
Abs
-0.10 -0.05 0.00 0.05 0.10 0.15
Figure 1.6 CO difference spectrum of VHb expressed in cytosol of P. pastoris
3. 西方免疫點漬分析
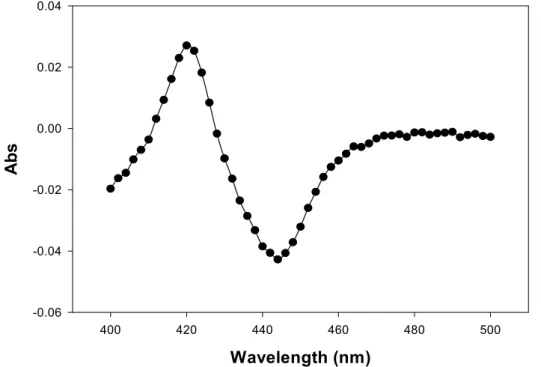
經過 CO difference spectrum 的分析後發現,無論是 VHb 表現在細胞質或過
氧化氫體中的 VHb+菌株,在 YPD 培養基下細胞確實有表現出血紅素 VHb,如 圖 1.6 與 1.7 所示,而且在過氧化氫體表現之 VHb 菌株經過胞器分離、西方墨 漬法初步分析後,我們可發現 VHb 確實有表現,並且可知道經由過氧化氫標的 訊息的引導,VHb-PTS1 確實有進入過氧化氫體中(如圖 1.8 所示)。
Figure 1.7 CO difference spectrum of VHb expressed in peroxisome of P. pastoris
Figure 1.8 Subcellular fractionation and Western blot analysis of VHb-PTS1 expressed in peroxisome of P. pastoris using anti-HA antibody as probe.
10 15 25 35
kDa
Peroxisome cytosol
VHb 10
15 25 35
kDa
Peroxisome cytosol
VHb Wavelength (nm)
400 420 440 460 480 500
Abs
-0.06 -0.04 -0.02 0.00 0.02 0.04
VHb-PTS1
在此部分的實驗結果當中,無論是人工合成或原生種的 VHb 均可表現於嗜 甲基酵母菌Pichia pastoris 中,並且其幫助溶氧的特性功能也經由 CO difference spectrum 分析加以證實了。而在微氧發酵槽培養下,我們也可看出 VHb 在微氧 環境下可幫助細胞的氧氣傳送,進而增加細胞生長,提供高密度發酵氧氣不足的 解決方向,除此之外,為了解決Pichia pastoris 表現的氧氣需求,發現在添加過 氧化氫標的訊息的VHb,一樣可以表現於 Pichia pastoris 中,並且經由透過簡易 的胞器分離及西方免疫分析發現,VHb 能經由標的訊息的牽引能正確的表現於 胞器中,對於接下來一連串的分析有很大的幫助。
五、參考文獻
1. Bulow L., Holmberg N., Lilius G., Bailey J.E. The metabolic effects of native and transgenic hemoglobins on plants. Trends Biotechnol 1999;17: 21-24.
2. Chen W., Hughes D.E., Bailey J.E. Intracellular expression of Vitreoscilla hemoglobin alters aerobic metabolism of Saccharomyces cerevisiae. Biotechnol Prog 1994;10: 308-313.
3. Dikshit K.L., Webster D.A. Cloning, characterization and expression of the bacterial hemoglobin gene from Vitreoscilla in Escherichia coli. Gene 1988;70:
377-386.
4. Khosla C., Bailey J.E. Heterologous expression of a bacterial haemoglobin improves the growth properties of recombinant Escherichia coli. Nature 1988;
331: 633-635
5. Kallio P. Bailey J.E. Intracellular expression of Vitreoscilla hemoglobin enhances total protein secretion and improves the production of α-amylase and neutral protease in Bacillus subtilis. Biotechnol Prog 1996;12: 31-39
6. Lemontt J.F., Wei C.M., Lawman M.J., Gray P.W. Expression of active human uterine tissue plasminogen activator in yeast. DNA 1985;4:419-428.
7. Ohashi R., Mochizuki E., Suzuki T. A mini-scale mass production and separation system for secretory heterologous proteins by perfusion culture of recombinant Pichia pastoris using a shaken ceramic membrane flask. J Biosci Bioeng 1999;87:
655-660
8. Romanos M.A. Advances in the use of Pichia pastoris for high-level gene expression. Curr Opin Biotechnol 1995;6:527-533
9. Van Arsdell J.N., Kwok S., Schweichart B.L., Ladner M.B., Gelfand D.H., Innis M.A. Cloning, characterization, and expression in Saccharomyces cerevisiae of endoglucanase I from Trichoderma reesei. Bio/Technology 1987;5:60-64.
10. Villatte F., Hussein A.S., Bachmann T.T., Schmid R.D. Expression level of heterologous proteins in Pichia pastoris is influenced by flask design. Appl Microbiol Biotechnol 2001;55:463-465
六、 計畫成果自評
本年度計畫成功地表現了異源蛋白 VHb 於 Pichia pastoris 中,並且也發現 VHb 在細胞質的表現初步能增加氧氣的傳送,幫助細胞在微氧環境下的生長,
並且也發現透過過氧化氫標的訊息的牽引,VHb 能有效的表現於過氧化氫體中,
對於解決Pichia pastoris 在高密度發酵中甲醇誘導時期極度需要氧氣的問題提供 了研究的方針,因此預計第二年的計畫將針對 VHb 表現在過氧化氫體中,對於 菌體生長及蛋白質合成的效應,以充分利用了解 VHb 的功能,除此之外,我們 也將探討VHb 表現於粒線體中,對於細胞本身的連鎖效應。