國立臺灣大學生物資源暨農學院農業化學系 碩士論文
Graduate Institute of Agricultural Chemistry College of Bioresources and Agriculture
National Taiwan University Master Thesis
三泰芬及邁克尼干擾青鱂魚肝中細胞色素 P450 活性 及全反式視黃酸相關基因表現量之研究
Triadimefon and Myclubutanil Interfere the Activity of Cytochrome P450s and mRNA Expression of All-trans Retinoic
Acid-Related Genes in the Liver of Medaka (Oryzias latipes)
林均鴻 Chun-Hung Lin
指導教授:陳佩貞 博士 Advisor: Pei-Jen Chen, Ph.D.
中華民國 100 年 8 月
August, 2011
II
誌謝
這篇論文的完成,首先要感謝指導教授陳佩貞老師,在實驗室草創初期就願 意收我這個完全沒有經驗的學生,讓我能從品保品管、藥品與儀器的選擇等實驗 的基礎開始學習。感謝老師不論是研究還是課業上都不厭其煩的叮嚀指導,感謝 老師為了這份研究四處奔波商借儀器,感謝老師對我不成熟的為人與實驗技術的 包容與指正,讓我能完成這份研究。感謝系上的老師們在專題討論時提出的建議 與指正,讓這份研究能在各個方面進行修正。感謝實驗室的成員們,已經畢業的 蘇志翔學長、陳冠雯學姊、助理曾吉言學長、覃世偉同學和塗子毅、吳宛霖、廖 珮涵、朱思泓和江峻蔚在實驗期間給予協助。感謝鄭鑫源、陳奕邵、陳昱達、洪 偉峰從大學部開始六年來在各方面的指導與協助。感謝顏瑞泓老師、陳建德老師、
洪傳揚老師、林乃君老師、黃良得老師、森林系葉汀峰老師、科技共同中心湯凱 鈞先生提供各式儀器,讓這份研究能夠完成。感謝口試委員顏瑞泓老師、陳建德 老師、生工系廖秀娟老師以及成大環工系周佩欣老師撥允為這篇論文給予建議與 指教。最後要感謝我的家人,對我這段時間不規律的作息的包容、照顧與支持,
讓我能無後顧之憂的完成這篇論文。
I
摘要
三泰芬 (Triadimefon) 及邁克尼 (Myclobutanil) 為農業上廣為使用的康唑類 殺真菌劑 (Conazoles)。研究顯示三泰芬 (具致腫瘤性) 及邁克尼 (不具致腫瘤性) 會干擾小鼠肝中重要代謝酵素細胞色素P450 (Cytochrome P450, CYP) 基因表現及 酵素活性,進而加劇維生素A 活性形式全反式視黃酸 (All-trans retinoic acid, atRA) 之代謝。但三泰芬及邁克尼對水生生物之影響的相關研究仍有限,因此本研究探 討三泰芬及邁克尼對青鱂魚 (Oryzias latipes, medaka) 肝中 CYPs 基因表現量、酵 素活性及atRA 相關基因表現之干擾作用。結果顯示,暴露三泰芬及邁克尼 (2 mg/L) 3 天後青鱂魚肝中 CYP3A38 及 CYP3A40 之 mRNA 表現有提升趨勢。暴露三泰芬 溶液 (2.0-3.5 μM) 7 天後會提升 CYP1A 和 CYP3A 活性,而暴露邁克尼溶液 (2.0-3.5 μM) 則僅提升 CYP3A 活性。此外,合成 atRA 之酵素 ALDH1A2 之 mRNA 表現量不受實驗處理影響,而代謝內生atRA 之酵素 CYP26B1 之 mRNA 表現量則 受三泰芬 (2.0-3.5 μM) 誘導提升,顯示三泰芬造成水生生物中 atRA 過量代謝的可 能性較邁克尼高。在atRA 相關受體中,RARγ1 受三泰芬 (2.0 μM) 誘導,顯示三 泰芬可能經由RARγ1 途徑誘導 CYP26B1 基因表現;而三泰芬及邁克尼 (2.0 μM) 暴露後RXRα1 及 RXRβ1 之 mRNA 表現量皆下降,顯示三泰芬及邁克尼皆可能會 影響細胞對atRA 之感知能力,進而影響 atRA 參與調控之生理機制,但無法以此 做為康唑類殺真菌劑是否具有致癌性之生物指標。綜合以上結果,暴露於三泰芬 和邁克尼會提高青鱂魚肝臟中特定CYPs 基因表現及活性,並干擾 atRA 相關核受
體之 mRNA 表現,可能會進而影響維生素 A 之正常代謝及其參與之生理調控機
制。
關鍵詞:三泰芬 (Triadimefon)、邁克尼 (Myclobutanil)、細胞色素 P450 (Cytochrome P450)、全反式視黃酸 (All-trans-retinoic acid)、青鱂魚 (Oryzias latipes, medaka)
II
Abstract
Triadimefon and myclobutanil are triazole-containing conazole fungicides widely used in agriculture. Previous studies show that some conazoles (e.g. triadimefon) at sublethal dose induced gene expression and enzymatic activity of cytochrome P450s (CYPs) in the liver of mouse, leading to a decrease in hepatic level of all-trans retinoic acid (atRA), an active form of vitamin A with anticancer properties. This study demonstrates the effects of carcinogenic triadimefon and non-carcinogenic myclobutanil on the activity of CYPs and mRNA expression of atRA-related genes in the liver of an aquatic organism, medaka (Oryzias latipes). Enzymatic activity analyses show triadimefon (2.0-3.5 μM) induced the activities of CYP1A and CYP3A, while myclobutanil (2.0-3.5 μM) only induced the activity of CYP3A. Quantitative real-time PCR analyses revealed that only triadimefon (2.0-5.0 μM) induced mRNA expression of endogenous atRA metabolism enzyme CYP26B1. Results from both enzymatic and genetic analyses above indicating that triadimefon may enhance hepatic atRA metabolism more severely than myclubutanil could do. Gene expression of atRA-related nuclear receptors retinoid X receptors (rxrα1 and rxrβ1) was depressed by both triadimefon and myclobutanil, indicating that both conazoles may interrupt the ability of cell to sense atRA and leading to disruption of atRA-regulated mechanisms. Overall, both triadimefon and myclobutanil interfere with the activity of cytochrome P450s and mRNA expression of all-trans retinoic acid-related genes in the liver of medaka.
Keywords: Triadimefon, Myclobutanil, Cytochrome P450, All-trans-retinoic acid, medaka (Oryzias latipes)
III
目錄
摘要... I Abstract ... II 目錄... III 圖目錄... V 表目錄... VI
1. 緒言... 1
1.1. 康唑類殺真菌劑... 1
1.1.1. 三泰芬... 3
1.1.2. 邁克尼... 5
1.2. 三泰芬及邁克尼於環境中的宿命... 7
1.3. 三泰芬及邁克尼之相關毒理研究... 8
1.4. 細胞色素P450 ... 10
1.5. 全反式視黃酸... 13
1.6. 模式生物的選擇... 15
1.7. 研究目的... 15
2. 材料與方法... 16
2.1. 試驗設計... 16
2.2. 實驗動物... 17
2.3. 藥品與試劑... 17
2.4. 儀器... 19
2.5. 暴露實驗設計與樣品採集... 19
2.5.1. mRNA 表現量分析預實驗 ... 19
2.5.2. mRNA 表現量分析實驗 ... 20
2.5.3. CYP 酵素活性分析實驗 ... 20
IV
2.6. 分析方法... 21
2.6.1. mRNA 表現量分析 ... 21
2.6.1.1. RNA 萃取 ... 21
2.6.1.2. TURBO 去 DNA 處理 ... 21
2.6.1.3. 反轉錄... 21
2.6.1.4. 引子設計與品質檢測... 22
2.6.1.5. 即時定量聚合酶連鎖反應... 23
2.6.2. CYP 酵素活性分析 ... 26
2.6.2.1. 微粒體萃取... 26
2.6.2.2. EROD 法 ... 26
2.6.2.3. DBF 試驗 ... 26
2.7. 統計分析... 27
3. 結果與討論... 28
3.1. 引子品質及酵素分析條件測試... 28
3.1.1. 引子品質測試... 28
3.1.2. 酵素分析條件測試... 28
3.2. mRNA 表現量分析預實驗 ... 35
3.3. 肝體指數... 37
3.4. CYP1A 基因表現及酵素活性 ... 39
3.5. CYP3A 基因表現及酵素活性 ... 43
3.6. atRA 恆定相關酵素之 mRNA 表現... 48
3.7. atRA 相關核受體之 mRNA 表現... 50
3.8. p53 之 mRNA 表現 ... 53
4. 結論... 55
參考文獻... 56
V
圖目錄
圖1-1 康唑類殺真菌劑作用機制 ... 2
圖1-2 全反式視黃酸結構 ... 13
圖1-3 青鱂魚 ... 15
圖2-1 研究架構 ... 16
圖3-1 引子解離曲線 ... 29
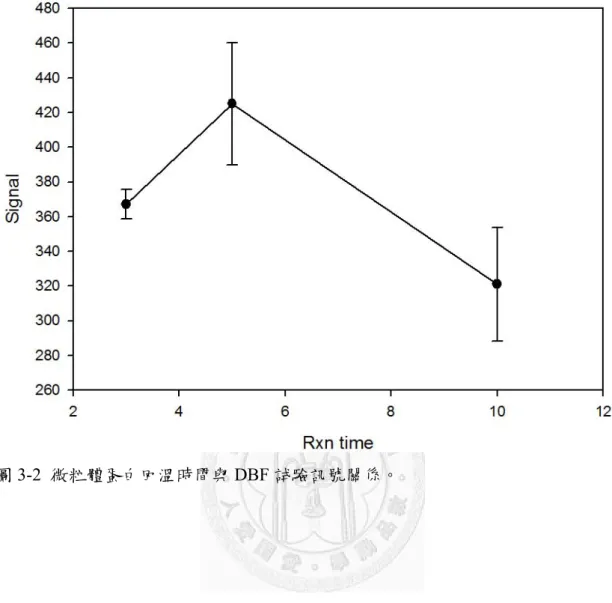
圖3-2 微粒體蛋白回溫時間與 DBF 試驗訊號關係 ... 31
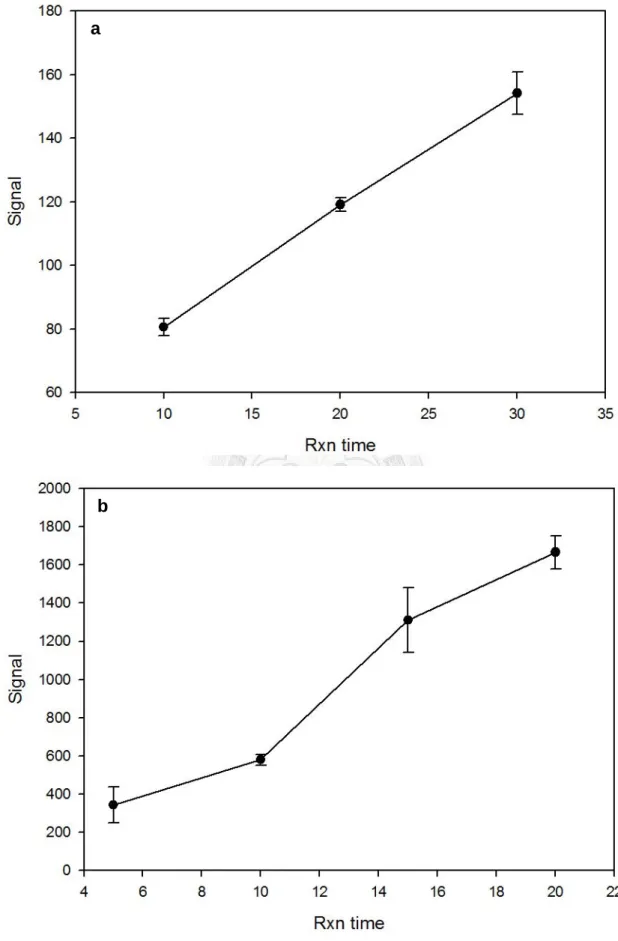
圖3-3 EROD 法與 DBF 試驗反應時間與訊號關係... 32
圖3-4 CYP1A 活性誘導試驗... 33
圖3-5 CYP1A 與 CYP3A 活性抑制實驗 ... 34
圖3-6 青鱂魚暴露於處理組及控制組 3 天後肝中 CYP1A, CYP3A38, CYP3A40 及PXR 之 mRNA 表現情形 ... 36
圖3-7 青鱂魚暴露於處理組及控制組 7 天後肝中 CYP1A 活性... 41
圖3-8 青鱂魚暴露於處理組及控制組 7 天及 14 天後肝中 CYP1A 及 AhR 之 mRNA 表現量... 42
圖3-9 青鱂魚暴露於處理組及控制組 7 天後肝中 CYP3A 活性... 45
圖3-10 青鱂魚暴露於處理組及控制組 7 天及 14 天後肝中 CYP3A38 及 CYP3A40 之 mRNA 表現量 ... 46
圖3-11 青鱂魚暴露於處理組及控制組 7 天及 14 天後肝中 PXR 之 mRNA 表現量 ... 47
圖3-12 青鱂魚暴露於處理組及控制組 7 天後肝中 ALDH1A2 及 CYP26B1 之 mRNA 表現量 ... 49
圖3-13 青鱂魚暴露於處理組及控制組 7 天後肝中 RARα1, RARγ1, RXRα1, RXRα2, RXRβ1 及 RXRβ2 之 mRNA 表現量 ... 52
圖3-14 青鱂魚暴露於處理組及控制組 7 天後肝中 p53 之 mRNA 表現量 ... 54
VI
表目錄
表1-1 三泰芬基本物化性質 ... 3
表1-2 三泰芬之公告使用範圍 ... 4
表1-3 邁克尼基本物化性質 ... 5
表1-4 邁克尼之公告使用範圍 ... 6
表2-1 水循環系統水質監測情形 ... 17
表2-2 引子序列 ... 24
表2-3 StepOne 儀器預設之即時定量聚合酶連鎖反應條件 ... 25
表2-4 EROD 法反應中各成分濃度 ... 27
表2-5 DBF 試驗反應中各成分濃度 ... 27
表3-1 青鱂魚暴露於處理組及控制組 7 天及 14 天後的肝體指數 (HSI) ... 38
1
1. 緒言
1.1. 康唑類 (conazole) 殺真菌劑
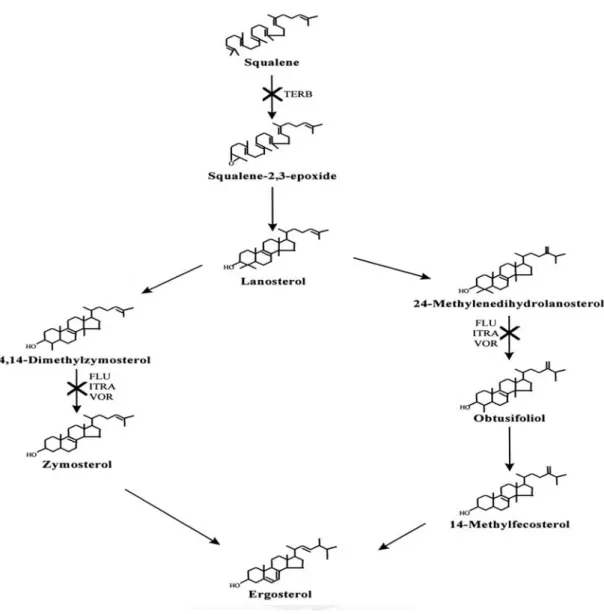
三泰芬 (triadimefon) 及邁克尼 (myclobutanil) 為康唑類殺真菌劑之一,用於 治療瓜果類、瓜菜類等作物的白粉病等病害。康唑類殺真菌劑為一類結構中含有 含 氮 戊 環 (azole) 的 殺 真 菌 劑 , 依 其 結 構 可 分 為 雜 環 中 含 兩 氮 的 咪 康 唑 類 (imidazole) 及含三氮的三唑類 (triazole)。康唑類藥劑的殺真菌作用機制主要為干 擾子囊菌及擔子菌中麥角固醇 (ergosterol) 的生合成。麥角固醇為真菌胞膜中重要 成分,可調控胞膜流動性及完整性。文獻指出,康唑類殺真菌劑會與真菌中細胞 色素P450 51 (cytochrome P450 51, CYP51) 的鐵蛋白 (heme protein) 結合,抑制其 對麥角固醇前驅物lanosterol 的 14α-去甲基 (14α-demethylation) 活性,其機制如圖 1-1 所示。此抑制作用會使麥角固醇合成量降低,進而造成真菌胞膜結構及功能受 損,最終導致真菌死亡 (Ghannoum and Rice, 1999)。部分咪康唑類殺真菌劑如克康 那唑 (ketoconazole) 和咪康唑 (miconazole) 等藥劑則具有更複雜之殺真菌機制,
如干擾真菌中脂質的生合成等生理機制等 (Sheehan et al., 1999)。
康唑類殺真菌劑的使用範圍相當廣泛,包括農業、醫療及個人保健用品中皆 有可觀之使用量 (Kahle et al., 2008)。在農業的使用中,康唑類殺真菌劑通常做為 系統性的殺真菌劑,依作物及防制病害種類不同而在幼苗期、花蕾萌出或發病初 期 時 使 用 。 其 他 用 途 包 括 蔬 果 與 穀 物 的 保 存 、 草 地 維 護 及 木 材 保 存 等 (Georgopapadakou and Walsh, 1996)。而在醫療及個人保健用品方面,康唑類殺真菌 劑則可做為系統性或局部性的殺菌劑,用於外用藥膏、塞劑、口服藥錠及洗髮精 等。除用於真菌治療外,部分康唑類化合物也有其他醫療用途,如克康那唑可阻 斷腎上腺類固醇的生合成,應用於治療前列腺癌中;而letrozole, anastrozole 等康 唑類殺真菌劑則可抑制芳香環轉化酶 (aromatase) 活性,應用於乳癌的治療中 (Zarn et al., 2003)。
2
圖1-1 康唑類殺真菌劑作用機制。(Ghannoum and Rice, 1999)
FLU, ITRA 和 VOR 分別為 fluconazole, itraconazole 及 voriconazole。此三種藥劑皆 為三唑類殺真菌劑。
3
1.1.1. 三泰芬
三泰芬為三唑類殺真菌劑之一,其結構及基本物化性質如表1-1 所示。三泰芬
目前於我國之主要用途為預防及治療農作物的真菌感染,表1-2 為我國農委會公告
之三泰芬使用範圍,包括瓜果類、葡萄及蘋果等作物的白粉病治療,乾豆類的銹 病治療等。以三泰芬為有效成分的農藥商品包括 Bayleton®, Amiral ®, Sadifon®及 Strike®等,而目前於我國登記使用的包括有洽益化學的好搭擋®、峰農的富貴通® 及世大農化的新世通®等產品。
表1-1 三泰芬基本物化性質。
中文名稱 三泰芬
英文名稱 Triadimefon
IUPAC 化學名稱 1-(4-chlorophenoxy)-3,3-dimethyl-1- (1H-1,2,4-triazol-1-yl)-2-butanone
CAS 編號 43121-43-3
結構式
分子式 C14H16ClN3O2
分子量 (g/mole) 293.75
水溶解度 (20℃) 64 mg/L
參考文獻:(EPA, 2006)
4
表1-2 三泰芬之公告使用範圍。
劑型
使用方法
作物名稱 使用範圍
每公頃每 次用藥量
稀釋倍數
25%
可濕性粉劑
3g/株 香蕉 葉斑病及葉黑星病
0.5 kg 2000 茶 茶餅病
5%
可濕性粉劑
2 kg 600 大豆 銹病
0.5 kg 2000 牛蒡 白粉病
0.5 kg 2000 瓜果類 白粉病
0.5-0.7 kg 2000 瓜菜類 白粉病
0.5 kg 2000 西瓜 白粉病
0.5-0.7 kg 2000 胡瓜 白粉病
2 kg 600 乾豆類 銹病
1000 梨 赤星病
1500 梨果類 白粉病
2000 葡萄 白粉病
1000 檬果 白粉病
1500 蘋果 白粉病
參考文獻:農委會公告農藥使用範圍
5
1.1.2. 邁克尼
邁克尼為一結構與三泰芬相似之三唑類殺真菌劑,其結構與基本物化性質如 表1-3 所示。邁克尼目前也僅用於農業上之真菌治療中,表 1-4 為我國公告之邁克 尼使用範圍,包括瓜菜類、葡萄及蘋果等作物的白粉病治療及蘋果的黑星病治療 等。以邁克尼為有效成分的農藥商品包括Nova®, Eagle®及Systhane®等,而目前於 我國登記使用的包括有萬德發的粉好效®、惠光的信心®及雅飛的勝生®等產品。
表1-3 邁克尼基本物化性質。
中文名稱 邁克尼
英文名稱 Myclobutanil
IUPAC 化學名稱 (RS)-2-(4-chlorophenyl)-2-
(1H-1,2,4-triazole-1-ylmethyl)hexanenitrile
CAS 編號 88671-89-0
結構式
分子式 C15H17ClN
分子量 (g/mole) 288.8
水溶解度 (20℃) 124 mg/L
參考文獻:(EPA, 2009)
6
表1-4 邁克尼之公告使用範圍。
劑型
使用方法
作物名稱 使用範圍
每公頃每次 用藥量
稀釋倍數
13.4%乳劑
0.25-0.35 L 4000 瓜菜類 白粉病
0.25-0.35 L 4000 胡瓜 白粉病
0.25 L 4000 香瓜 白粉病
40%
可濕性粉劑
6000 印度棗 白粉病
4000 柿 角斑病
0.1 kg 12000 柿 白粉病
3000 桃 縮葉病
8000 梅 白粉病
12000 葡萄 白粉病
12000 蘋果 黑星病
0.1 kg 12000 蘋果 白粉病
參考文獻:農委會公告農藥使用範圍
7
1.2. 三泰芬及邁克尼於環境中的宿命
雖然前人研究已發現目前廢水處理設備無法有效移除普克利 (propiconazole)、
氟康唑 (fluconazole) 等康唑類殺真菌劑 (Kahle et al., 2008),且已在環境水體中測 得ng/L 至 μg/L 濃度範圍的依普座 (epoxiconazole)、得克利 (tebuconazole) 等農業 用途之康唑類殺真菌劑 (Berenzen et al., 2005),但目前對於三泰芬及邁克尼在環境 中流佈的相關研究仍相當有限。
文獻指出,三泰芬在土壤中主要經由微生物作用途徑降解,於有氧及無氧的 砂質壤土中之半生期分別為5.6 及 23.1 天,屬於非持久性農藥 (EPA, 2006; Singh, 2005)。因三泰芬不易被水解,故在水中之主要降解途徑則為光降解與微生物降解,
而在無氧水體中半生期則可達 217 天 (EPA, 2006)。雖然三泰芬之土壤吸附係數 (adsorption coefficients, Koc) 為 387 L/kg,在土壤中移動性不高,但仍有可能因降 雨或灌溉行為而被淋洗出,並經地表逕流而進入表面水體中 (EPA, 2006)。
邁克尼在有氧土壤中之半生期大於70 天,為中度持久性 (moderately persistent) 農藥,在無氧土壤中之半生期則更長,為持久性農藥 (EPA, 2009)。因邁克尼微溶 於水,且不易被水解或光降解,故美國環境保護局 (Environmental Protection Agency, EPA) 推估淋洗、地表逕流及灌溉等作用或行為是邁克尼施用後的主要消散 (dissipation) 途徑 (EPA, 2009),因此邁克尼亦有進入環境水體中之風險。
8
1.3. 三泰芬及邁克尼之相關毒理研究
由於三泰芬及邁克尼皆為上市之商品,其對哺乳動物和環境水生生物的急毒 性、慢毒性等基礎毒理試驗皆已在小鼠、大鼠、虹鱒或藍腮魚等模式生物 (model organism) 中完成 (EPA, 2006; EPA, 2009)。三泰芬和邁克尼和其他康唑類殺真菌劑 相似,對哺乳動物的急毒性和慢毒性皆為輕微,對水生生物則具有中等毒性,而 在亞致死劑量下對大鼠皆具致畸性 (INCHEM, 1981; INCHEM, 1992)。而隨著科技 進步與人們對毒理效應的重視程度增加,關於三泰芬及邁克尼對生物正常生理機 制之影響的研究也陸續被進行與發表 (Allen et al., 2006; Chen et al., 2009; EPA, 2006; EPA, 2009; Sun et al., 2007)。
文獻指出,三泰芬和邁克尼會誘導小鼠肝中CYPs 酵素活性表現,而增加對維 生素A (retinol, vitamin A) 活性形式-全反式視黃酸 (all-trans retinoic acid, atRA) 的代謝,進而使小鼠體內atRA 濃度降低 (Chen et al., 2009)。此現象可能會導致維
生素 A 缺乏,並引發相關症狀,影響生物維持視覺、骨骼健康及免疫調控等生理
功能 (Morriss-Kay and Ward, 1999)。然而,有關三泰芬及邁克尼是否會影響環境生 物的瞭解仍十分有限,尤其在三泰芬及邁克尼經表面逕流等方式進入環境後,水 生生物為直接暴露於三泰芬及邁克尼的環境生物,故瞭解三泰芬及邁克尼對水生 生物之影響為評估其對環境衝擊中的重要議題。
另外,三泰芬與邁克尼影響atRA 代謝之研究最初起因於三泰芬致癌機制之探 討。文獻指出,三泰芬對小鼠具致肝腫瘤性,並對雄性大鼠具致甲狀腺腫瘤性,
因此三泰芬被美國環境保護局列為可能之致癌物 (C 類) (EPA, 1996)。而邁克尼在 相似劑量下於小鼠和大鼠中皆無致腫瘤反應,因此被美國環境保護局列為非致癌 物 (E 類) (EPA, 2000; INCHEM, 1992)。由於三泰芬及邁克尼在化學結構、使用範 圍及可能之流佈情形上有許多相似處,因此在許多三泰芬致癌機制的研究中皆以 邁克尼做為非致癌性的康唑類殺真菌劑代表 (Allen et al., 2006; Chen et al., 2009;
9
Hester et al., 2006; Ward et al., 2006)。
Ward 等學者以核酸微陣列 (microarray) 分析小鼠在餵食 1,800 ppm 三泰芬或 2,000 ppm 邁克尼 4 天後約 34,000 個基因之 mRNA 表現情形,發現共有 8 個機制 共259 個基因的 mRNA 表現量會受三泰芬干擾,而不受邁克尼影響 (Ward et al., 2006)。以此研究為基礎,許多學者針對三泰芬與邁克尼影響機制間的差異性深入 探討三泰芬可能之致癌機制,而維生素A 相關代謝機制便是其中之一 (Chen et al., 2009; Nesnow et al., 2009)。雖然在小鼠研究中,三泰芬與邁克尼對 atRA 代謝途徑 造成之影響相似,但此影響是否會有物種間的差異,也是評估其三泰芬及邁克尼 對環境生物影響中的重要議題之一。
此外,文獻指出三泰芬對小鼠及大鼠具有致腫瘤性,邁克尼則無此現象 (INCHEM, 1981; INCHEM, 1992),但尚無研究證實兩藥劑是否對水生生物具致腫 瘤性。前人研究指出,部分康唑類殺真菌劑會干擾小鼠體內腫瘤抑制基因 (tumor suppressor gene) p53 的基因及蛋白表現量 (Hester et al., 2006; Ho et al., 1998; Ward et al., 2006),而其是否可做為康唑類殺真菌劑對水生生物是否具致腫瘤性之生物指 標也將於本研究中探討。
10
1.4. 細胞色素 P450
細胞色素P450 是由許多單氧酶 (monooxygenase) 組成之酵素 superfamily,當 氨基酸序列相似度大於40%時分為同一 family (如 CYP1),大於 55%時則分為同一 subfamily (如 CYP1A)。目前在人類基因中已發現 18 個以上的 CYP family,合計超 過57 種 CYP 酵素 (Nebert and Russell, 2002)。不同 CYP 酵素對基質具有不同選擇 性,分別在生物體中參與不同生理機制,如人體中CYP11, CYP17 及 CYP19 參與 性賀爾蒙生合成、CYP24 代謝維生素 D 並調節相關的生理反應等,而體內內生之 atRA 濃度則由 CYP26 代謝調控 (Lewis, 2004; Nebert and Russell, 2002; Sakaki et al., 2005; Thatcher and Isoherranen, 2009)。
此外,CYPs 也是第一階段代謝 (phase I metabolism) 中重要的氧化酵素,可 藉由氧化作用代謝外來物質,產生親水性較高之代謝產物,或產生高反應性代謝 產 物 以 利 第 二 階 段 代 謝 (phase II metabolism),進而促進代謝產物排出體外 (Honkakoski and Negishi, 2000)。但在代謝過程中也可能會活化 (bioactivation) 外
來物質的毒性,或是影響藥物及外來物質在生物體內的相互作用,故瞭解CYP 之
作用機制為毒理研究中重要的議題。
人體中主要進行第一階段代謝的CYP 酵素包括了 CYP1, CYP2 及 CYP3,分 別 可 代 謝 不 同 的 外 來 物 質 (Lewis, 2003) 。 其 中 CYP1A 可 代 謝 多 環 芳 香 烴 (polycyclic aromatic hydrocarbons, PAHs)、異環芳香胺 (heterocyclic aromatic amines) 等物質 (Monostory et al., 1996; Smolowitz et al., 1992)。當這些物質進入人體後,會 與核受體 (nuclear receptor) aryl hydrocarbon receptor (AhR) 結合,調控 CYP1A 基 因及蛋白表現,進而氧化代謝外來物質 (Tompkins and Wallace, 2007)。然而在許多 PAH 的代謝中,由 CYP1A 氧化產生的高反應性代謝產物會和 DNA 作用,並進而 造成致癌性,此現象即為CYP1A 對 PAH 致癌性之生物活化 (Ma and Lu, 2007)。
在目前使用的人類用藥中,CYP2C 可代謝的種類僅次於 CYP3A,而 CYP2B
11
表現量雖較少,但也參與活化古柯鹼 (cocaine) 的肝毒性及黃麴毒素 (aflatoxins) 的致肝腫瘤性等機制 (Boelsterli et al., 1992; Guengerich, 2003; Robertson et al., 1983)。CYP2B 及 CYP2C 在人類及許多哺乳動物中主要由核受體 constitutive androstane receptor (CAR) 調控誘發,但目前在魚類中仍未發現 CAR, CYP2B 及 CYP2C 的存在 (Tompkins and Wallace, 2007)。
CYP3A 為人類肝臟中表現量最大的 CYP 酵素,可代謝的藥品種類也最多 (Williams et al., 2004)。文獻指出,CYP3A 主要由核受體 pregnane X receptor (PXR) 調控誘發,但因PXR 與 CAR 之功能有部分重疊,故也可經由 CAR 途徑誘發 CYP3A 表現 (Nannelli et al., 2008; Tompkins and Wallace, 2007)。由於 CYP3A 參與多種藥
物代謝,因此 CYP3A 活性會影響藥物及環境毒物在生物體中之相互作用,如
CYP3A 活性被抑制時可能會導致其他藥物或環境毒物累積而產生毒性,CYP3A 活 性被誘導時則可能會引起活化黃麴毒素B1的基因毒性及troglitazone (糖尿病用藥) 的肝毒性等現象 (Kassahun et al., 2001)。目前在水生生物中已發現多種 CYP3A 形 式 (isoform),如在青鱂魚中即有 CYP3A38 及 CYP3A40 兩種形式,雖然現今對於
水生生物中不同形式的CYP3A 酵素可代謝基質種類的瞭解相當有限,但已有研究
指出不同物質對不同形式CYP3A 的誘發效果不盡相同,因此不同形式 CYP3A 對 不同內生及外來物質可能具有不同之代謝能力 (Kashiwada et al., 2007; Yamauchi et al., 2008)。
文獻指出三泰芬及邁克尼會誘導小鼠及大鼠肝中AhR, CAR 及 PXR 等核受體 之mRNA 表達,並提升下游調控基因 CYP1A, CYP2B 及 CYP3A 之 mRNA 表達及 酵素活性 (Allen et al., 2006; Chen et al., 2009; Goetz and Dix, 2009; Goetz et al., 2006; Sun et al., 2007; Tully et al., 2006)。當 CYP 酵素活性受干擾時,除會影響對環 境汙染物及藥物的代謝及生物活化外,因CYP1A 及 CYP3A 具有代謝 atRA、性賀 爾蒙等內生物質的活性,當酵素活性被誘發時可能會加速體內重要物質的代謝速 度,進而影響正常生理機能的運作 (Chen et al., 2009; Williams et al., 1998)。因此,
12
瞭解外來物質如何影響CYP 的表現及活性在毒理研究中具有重大的意義。
雖然目前並無文獻探討三泰芬及邁克尼對魚類CYP 表現及活性的影響,但已 有許多研究證實撲克拉 (prochloraz)、克康那唑等康唑類殺真菌劑會干擾青鱂魚、
虹鱒等魚類中CYP 的表現情形 (Hasselberg et al., 2008; Zhang et al., 2008a),顯示
康唑類殺真菌劑具有影響水生生物CYP 表現的可能性,以及相關研究的急切與必
要性。
13
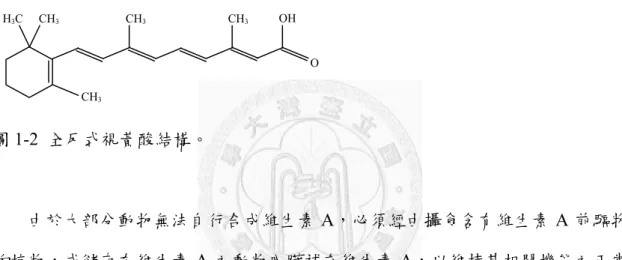
1.5. 全反式視黃酸
全反式視黃酸為具有生理活性的維生素 A 衍生物之一,其化學結構如圖 1-2 所示。廣義之維生素A 包含超過 4,000 種形式的維生素 A 衍生物及具維生素 A 活
性之物質。而在天然維生素 A 衍生物中,大部分的形式並不具有生理活性,如酯
質型態維生素A (retinyl esters) 即為動物儲存及運輸之維生素 A 形式 (Theodosiou et al., 2010)。
圖1-2 全反式視黃酸結構。
由於大部分動物無法自行合成維生素 A,必須經由攝食含有維生素 A 前驅物
的植物,或儲存有維生素 A 之動物內臟補充維生素 A,以維持其相關機能之正常
運作 (Blomhoff and Blomhoff, 2006)。Theodosiou 等學者回顧並發表了維生素 A 於
人體內之轉化、運作及代謝機制,動物在攝入維生素 A 前驅物或酯質型態維生素
A 後,經由一連串的代謝反應,最後由 aldehyde dehydrogenases 1a (ALDH1A) 酵 素生成具有生理活性的atRA 或 9-cis retinoic acid (9-cis RA),而 atRA 和 9-cis RA 在與核受體retinoic acid receptor (RAR) 和 retinoid X receptor (RXR) 結合後,可調 控相關基因表現 (Theodosiou et al., 2010),並藉由不同濃度的 atRA 調控胚胎發育、
器官分化等機制,或維持足量濃度以維持正常視力、皮膚健康等機制 (Morriss-Kay and Ward, 1999)。
當長期維生素A 攝取不足時,會造成維生素 A 缺乏 (vitamin A deficiency, VAD),
引起結膜乾燥、角膜軟化等症狀 (Sommer, 2008)。而在 atRA 濃度過高時,過量之
OH
O CH3
CH3
CH3
CH3
H3C
14
atRA 在和 RAR 結合後,會誘導 CYP26 基因及蛋白表現,進而由 CYP26 及其他第 二階段代謝酵素作用清除過量 atRA,以避免過量 atRA 造成發育畸形、神經損傷 等傷害 (Blomhoff and Blomhoff, 2006; Collins and Mao, 1999)。
1925 年,Wolbach 和 Howe 發現在 VAD 的患者中出現腫瘤組織 (Wolbach and Howe, 1925),自此開始維生素 A 與癌症之相關研究。1985 年,Kark 等學者分析 了174 人的血液中維生素 A 含量,發現其中的 85 名癌症患者中,血液中維生素 A 含量明顯較平均值低 (Kark et al., 1981)。而後在動物實驗和流行病學研究中也發現 了VAD 與腫瘤形成間具有良好關聯性 (Sun and Lotan, 2002)。目前對於維生素 A
與腫瘤形成間的詳細作用機制仍不完全瞭解,普遍認為維生素 A 具有抑制不正常
細胞增生、誘發不正常細胞凋亡及促進正常細胞增生及分化等功用 (Niles, 2000),
可抑制腫瘤細胞增生。相反地在維生素A 缺乏的情況下,腫瘤細胞則會加速增生。
因此,破壞生物體內維生素A 的恆定亦被認為是與致癌相關之機制之一。
另外,目前已在多種癌症前期細胞及組織中發現atRA 相關之核受體 RARs 及 RXRs 之表現量有降低情形 (Berard et al., 1996; Castillo et al., 1997; Houle et al., 1993; Lotan et al., 1995; Lotan et al., 2000; Picard et al., 1999; Qiu et al., 1999;
Widschwendter et al., 1997; Xu et al., 1994; Xu et al., 1997a; Xu et al., 1997b)。RARs 及RXRs 在人類中分別具有 α, β 及 γ 三種亞型 (subtype),每種亞型又有數種同功 酶 (如 RXRα1 及 RXRα2),不同 RARs 及 RXRs 的表現量比例依組織及發育時期 而不同,亦可能僅有單一RAR 或 RXR 表現 (Chambon, 1996)。雖然目前對於 atRA
相關核受體表現、維生素 A 缺乏及其相關致癌性的詳細關聯性及機制仍不完全明
瞭,但為探討在致癌性及非致癌性康唑類殺真菌劑暴露下對水生生物肝中atRA 相
關核受體表現是否會有所影響,以及 atRA 相關核受體之 mRNA 表現情形是否能 做為康唑類殺真菌劑具致癌性與否之生物指標,本研究也將分析青鱂魚暴露三泰 芬及邁克尼後,其肝中atRA 相關核受體之 mRNA 表現情形。
15
1.6. 模式生物的選擇
為探討三泰芬及邁克尼對水生生物肝中 CYP 酵素活性及 atRA 相關基因表現 量之影響,本研究選用青鱂魚 (Oryzias latipes, medaka) 做為模式生物。青鱂魚為 輻鰭魚綱 (Actinopterygii) 鶴鱵目 (Beloniformes) 異鱂亞目 (Adrianichthyoidei) 異鱂科 (Adrianichthyidae) 青鱂屬 (Oryzias) 之小型淡水魚種,原生於日本、台灣、
韓國等東南亞淡水域,成熟期約2-3 個月,成魚體長約 2-3 公分 (圖 1-3),不作食 用亦不作觀賞魚用。以魚類做為毒理研究之模式生物可提供快速、經濟的初步篩 選,並可做為化合物對環境生物影響之研究。
青鱂魚做為模式生物具有體積小、繁殖快、卵透明易觀察等優點,且為亞洲 地區原生之野生物種,而本研究主要利用其對多種環境物質具高敏感性之特性進 行研究。
圖1-3 青鱂魚。
1.7. 研究目的
本研究將探討青鱂魚暴露三泰芬及邁克尼後,對其肝中 CYP 活性及 atRA 恆
定相關基因表現量之影響,以評估三泰芬及邁克尼對水生生物中atRA 代謝情形之
影響,及比較三泰芬及邁克尼在青鱂魚及哺乳動物中造成之影響的差異。並將以 青鱂魚暴露三泰芬及邁克尼後,其肝中p53 和 atRA 相關核受體之 mRNA 表現量 改變情形,作為三泰芬及邁克尼是否對水生生物致腫瘤性之初步評估。
16
2. 材料與方法 2.1. 試驗設計
圖2-1 研究架構。
本研究將以即時定量聚合酶連鎖反應 (quantitative real-time polymerase chain reaction, q-RT-PCR) 及兩種酵素活性測定法分析青鱂魚暴露三泰芬及邁克尼後肝 中CYP 酵素活性與 atRA 相關基因表現量。q-RT-PCR 分析項目包括 CYP 與其相 關核受體、atRA 相關酵素及核受體、p53 之 mRNA 表現量。Ethoxyresorufin O-deethylase (EROD) 法及 Dibenzylfluorescein (DBF) 試驗則分別用於測定 CYP1A
及CYP3A 活性。因 CYP1A 及 CYP3A 皆為內質網 (endoplasmic reticulum) 上之膜 蛋白,故實驗時須先打破細胞,萃取出由內質網碎片形成的微粒體 (microsome) 部 分後,再以酵素活性測定法測定酵素活性。
本研究將以7 月齡青鱂魚雄魚進行暴露及分析,此時期之青鱂魚已達性成熟,
各項生理機制表現穩定,且萃取出之RNA 及微粒體蛋白足夠進行基因表現及酵素
活性分析。
17
2.2. 實驗動物
實驗用魚於實驗室附設魚房之水循環系統中飼養繁殖。每10-15 隻青鱂魚成魚 飼養於1 個 3 L 壓克力魚缸中,以微弱水流循環並以風管插入液面打氣維持溶氧量
充足。水循環系統每日更換1/3 過濾之除氯自來水以避免硝酸態氮累積。飼養環境
為每日14 小時光照,溫度為 26±1℃,每日餵食豐年蝦及乾飼料 otohime-β1 共 2-3 次,每週以市售水質檢測試劑監控系統中水質如表2-1。
表2-1 水循環系統水質監測情形。
監測項目 濃度
硝酸態氮 < 5 mg/L
亞硝酸態氮 < 0.5 mg/L
氨態氮 ~0.5 mg/L
總硬度 ~17.9 mg/L
碳酸鹽硬度 6-15 mg/L
溶氧 ~8 mg/L
氯 < 0.5 mg/L
2.3. 藥品與試劑
二甲基亞碸 (dimethyl sulfoxide, DMSO)、氯化鎂 (magnesium chloride, MgCl2)、 磷酸氫二鉀 (dipotassium hydrogen phosphate, K2HPO4)、磷酸二氫鉀 (potassium dihydrogen phosphate, KH2PO4)、Tris base、醋酸 (acetic acid)、甘油 (glycerol)、氫 氧 化 鈉 (sodium hydroxide, NaOH) 購 自 J.T. baker (U.S.A) 。 1- 溴 -3- 氯 丙 烷
18
(1-bromo-3-chloropropane)、異丙醇 (isopropanol)、乙醇 (ethanol)、乙烯二胺四醋 酸 (ethylenediaminetetraacetic acid, EDTA) 、 雙 丁 基 氫 氧 甲 苯 (butylated hydroxytoluene, BHT)、苯基甲基磺醯氟 (phenylmethylsulfonyl fluoride, PMSF)、乙 氧基間一苯二酚 (ethoxyresorufin, ER)、菸鹼醯胺腺嘌吟 (β-nicotinamide adenine dinucleotide phosphate sodium, NADP+)、葡萄糖-6-磷酸 (glucose-6-phosphate, G6P)、
葡萄糖-6-磷酸脫氫酶 (glucose-6-phosphate dehydrogenase, G6DPH)、β-奈黃酮 (β-naphthoflavone, β-NF) 購自 Sigma (U.S.A)。甲醇 (methanol)、甲醛 (formaldehyde) 購自Merck (U.S.A)。Dibenzylfluorescein (DBF) 購自 BD (U.S.A)。RNAlater®, TRI Reagent® Solution, Power SYBR® Green PCR Master Mix, Nuclease-Free Water 購自 Applied Biosystems (U.S.A)。
商業套組
TURBO DNA-free™ kit, High-Capacity cDNA Reverse Transcription Kit 購自 Applied Biosystems (U.S.A)。Pierce® BCA Protein Assay Kit 購自 Thermo Fisher Scientific (U.S.A)。
配置溶液
a.) DEPC 水:0.1% DEPC 配置於 milli Q 水中。
b.) TE 緩衝液:10 mM Tris-HCl, 1 mM EDTA 配置於 milli Q 水中。
c.) 均質用緩衝液:0.1 M Tris-acetate, 1 mM EDTA, 0.02 mM BHT, 0.1 mM PMSF 配 置於milli Q 水中,調整 pH 至 7.4。
d.) 保存用緩衝液:10 mM Tris-acetate, 1 mM EDTA, 20% glycerol 配置於 milli Q 水 中。
19
2.4. 儀器
離心機FORCE1418 (ForceMicro) 低溫離心機 (Eppendorf)
超高速低溫離心機Optima™ MAX-XP Benchtop Ultracentrifuge (Beckman Coulter) 均質器D-54518 Niersbach MICROMOT (PROXXON)
手動均質器Wheaton Tenbroeck Tissue Grinder (Fisher) 四位天平AC204 (METTLER TOLEDO)
五位天平 (denver)
分光光度計BioPhotometer (Eppendorf)
微量分光光度計Nanodrop ND-1000 (Thermo Scientific) 熱循環反應器T3000 Thermocycler (Biometra)
核酸即時定量PCR StepOne (Applied Biosystems) 恆溫培養箱LE09RD (裕德)
微盤讀儀Microplate reader BLX808 (Bio-Tek)
多功能微盤讀儀FlexStation3 microplate reader (MDS)
2.5. 暴露實驗設計與樣品採集
2.5.1. mRNA 表現量分析預實驗
為求得最佳反應之實驗條件,本研究先將青鱂魚暴露於亞致死劑量的三泰芬 及邁克尼3 天,分析其肝中 CYP1A, CYP3A38, CYP3A40 及 PXR 之 mRNA 表現情
形。實驗步驟為將 9 隻七月齡青鱂魚雄魚隨機平分為三組,於實驗前一天分別放
入3 個裝有 600 mL 除氯水之 1 L 燒杯中適應。實驗開始當天將燒杯中的除氯水 更換為暴露溶液以開始暴露實驗,處理組別包括溶劑控制組 (0.007% DMSO)、2 mg/L 三泰芬 (~6.8 μM) 及 2 mg/L 邁克尼 (~6.9 μM),暴露持續 3 天,每日餵食
20
otohime-β1 1 次,並於餵食 1 小時後更換暴露溶液。暴露條件為光週期為每日 14
小時光照,溫度為 26±1℃。適應及實驗期間皆以風管插入液面打氣以維持溶氧,
並於杯口套上尼龍網防止魚隻跳出。暴露結束後犧牲魚隻,取下肝臟保存於-80℃
冰箱中留待mRNA 表現量分析。
2.5.2. mRNA 表現量分析實驗
為求得更佳顯著的反應,mRNA 表現量分析實驗中將暴露時間拉長為 7 至 14 天,並為避免造成魚隻死亡而將暴露濃度降為2.0-5.0 μM。實驗步驟為將 80 隻七 月齡青鱂魚雄魚隨機平分為16 組,於實驗前一天分別放入 16 個裝有 600 mL 除氯
水之1 L 燒杯中適應。實驗開始當天將燒杯中的除氯水更換為暴露溶液以開始暴露
實驗,處理組別包括除氯水控制組、溶劑控制組 (0.001% DMSO)、不同濃度之三 泰芬 (2.0, 3.5, 5.0 μM) 及邁克尼 (2.0, 3.5, 5.0 μM) 共八組,每組 2 杯分別暴露 7
天及14 天。換水、餵食及其他暴露條件同預實驗。暴露完成時秤量魚隻重量、犧
牲魚隻取出肝臟、將肝臟浸於RNAlater®樣品保存液中秤量肝重後,依RNAlater® 原廠操作手冊,將樣品冷藏隔夜後置於-80℃冰箱中保存留待 mRNA 表現量分 析。
秤得之肝重與體重用於計算肝體指數 (hepatosomatic index, HSI),計算方式為 HSI = 肝重 / 體重。
2.5.3. CYP 酵素活性分析實驗
綜合mRNA 表現量分析預實驗及 mRNA 表現量分析實驗之結果,為避免高濃 度、長時間的暴露造成毒性,CYP 酵素活性分析實驗將最高暴露濃度降為 3.5 μM,
並僅進行7 天之暴露。實驗步驟為將 60 隻七月齡青鱂魚雄魚隨機平分為 12 組,
於實驗前一天分別放入12 個裝有 600 mL 除氯水之 1 L 燒杯中適應。實驗開始當 天將燒杯中的除氯水更換為暴露溶液以開始暴露實驗,處理組別包括除氯水控制
21
組、溶劑控制組 (0.001% DMSO)、不同濃度之三泰芬 (2.0, 3.5 μM) 及邁克尼 (2.0, 3.5 μM) 共六組,暴露持續 7 天,換水、餵食及其他暴露條件同預實驗。暴露完成 時秤量魚隻重量、犧牲魚隻取出肝臟、將肝臟浸於均質用緩衝液中秤量肝重後,
立即進行微粒體萃取。秤得之肝重與體重用於計算肝體指數,計算方法同上。
2.6. 分析方法
2.6.1. mRNA 表現量分析 2.6.1.1. RNA 萃取
將樣品自-80℃ 冰箱中取出解凍,並除去 RNAlater® 溶液後,使用 TRI Reagent® Solution 萃取青鱂魚肝臟中之 RNA (除因樣品較小而將操作體積減半外,
實驗步驟如原廠操作手冊)。萃取出之總 RNA 回溶於 50 μL DEPC 水,取出 3 μL
測定 RNA 濃度,其餘置於-20℃冰箱中保存。RNA 濃度測定以分光光度計
BioPhotometer 進行,以 TE 緩衝液為空白樣品校正後,將 3 μL RNA 樣品以 TE 緩 衝液稀釋25 倍,測量 260 nm 吸光值,並以 1 OD260 = 40 μg/mL 估算 RNA 濃度。
2.6.1.2. TURBO 去 DNA 處理
自總 RNA 樣品中取出 10 μg RNA,以 TURBO DNA-free™ kit 及 T3000 Thermocycler 移除 RNA 樣品中的 DNA (實驗步驟如原廠操作手冊)。RNA 濃度測 定以微量分光光度計Nanodrop 進行,以 Nuclease-Free Water 為空白樣品校正後,
取1.2 μL 去 DNA 之 RNA 樣品測量 260 nm 吸光值,並以 1 OD260 = 40 μg/mL 估算 RNA 濃度。將去 DNA 之 RNA 樣品置於-20℃冰箱中保存。
2.6.1.3. 反轉錄 (reverse transcription)
自去DNA 之 RNA 樣品中取出 10 μg RNA,以 High-Capacity cDNA Reverse
22
Transcription Kit 及 T3000 Thermocycler 進行反轉錄 (實驗步驟如原廠操作手冊)。
反應完成後之cDNA 樣品置於-20℃冰箱中保存。
2.6.1.4. 引子設計與品質檢測
引子序列設計以文獻中之序列為優先。查無相關文獻或文獻中之引子序列無 法 通 過 下 述 之 品 質 檢 測 時 , 使 用 文 獻 中 之 基 因 編 號 於 National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/) 中取得其完整序列,並使 用PrimerExpress 3.0 設計引子。
由於引子於即時定量聚合酶連鎖反應中需具有良好專一性,不產生自黏現象,
以及在實驗使用之 cDNA 濃度範圍中有穩定放大效果等性質,所有引子序列在分
析樣品前皆需經過品質檢測。引子品質檢測包括引子濃度測試及引子放大效率測 試兩部分,前者測試足量且不產生自黏現象之引子濃度,後者則測試引子於實驗 使用之cDNA 濃度範圍中之放大效果及不產生非專一性鍵結之 cDNA 濃度。
a.) 引子濃度測試
各基因分別配置100 nM 及 300 nM 引子濃度組合,進行 q-RT-PCR。確認引子 濃度300 nM 反應結果中解離曲線,如為單一波峰則表示此引子於 300 nM 反應時 不會產生自黏現象,即可以300 nM 進行反應。若具兩個以上之波峰,則確認引子 濃度100 nM 反應結果中解離曲線,如為單一波峰則以 100 nM 進行後續反應。若 仍具兩個以上之波峰,且其中之一的訊號值明顯隨引子濃度下降而減少,則再下 降引子濃度進行實驗。唯若兩波峰之訊號值皆隨引子濃度下降而明顯減少時,表 示此引子在反應所需之足量濃度下皆會產生自黏現象,需重新設計引子。
b.) 引子放大效率測試
將cDNA 樣品以 Nuclease-Free Water 做序列稀釋,進行 q-RT-PCR。各基因使
23
用之引子濃度皆為300 nM。作 log[cDNA]-CT圖之線性迴歸方程式,確認判定係 數 (R2) 大於 0.99,且斜率介於 -3.587 至 -3.103 間。若線性因高或低濃度點而偏 差,或CT值已低於 20 或高於 35,則於線性區間內重新稀釋數個濃度進行實驗,
以確認引子於實驗使用之cDNA 濃度範圍中之放大效果穩定,且所得之 CT值介於
ΔΔCT法可定量之範圍內。若反應結果中解離曲線具兩個以上之波峰,且其中之一
的訊號值於cDNA 濃度下降後消失,則表示高濃度反應中 cDNA 濃度過高,使引
子產生非專一性鍵結。此時需選用不於解離曲線中產生雙波峰之 cDNA 濃度進行
反應,唯若降低cDNA 濃度後 CT值高於35 時需重新設計引子。
2.6.1.5. 即時定量聚合酶連鎖反應
將cDNA 樣品稀釋 10 倍後,以 Power SYBR® Green PCR Master Mix 及 StepOne 進行即時定量聚合酶連鎖反應 (除因樣品較少而將操作體積由 50 μL 減少至 20 μL 外,實驗步驟如原廠操作手冊)。本實驗共測定 1 個參考基因 (rpl-7) 及 14 個目標 基因,每樣品每基因測定二重複。使用之引子序列如表 2-2 所示。PCR 反應條件 為StepOne 預設程序,如表 2-3 所示。實驗結果以 ΔΔCT法計算所有處理組相較於 DMSO 控制組之表現倍數。
24
表2-2 引子序列。
基因 引子序列 (F:前置、R:後置) 來源
rpl-7 F CGCCAGATCTTCAACGGTGTAT
(Zhang et al., 2008b) R AGGCTCAGCAATCCTCAGCAT
ahr F CCTTTCCCGAGGACGTCAT
自行設計 R GAGGTAGCTGACGCTAAGCCTTA
cyp1a F CGCAGAAAGTTGGCCTACAGT
(Hong et al., 2007) R TCTGCATTGCTGCCCTCTAG
pxr F GAGGAGCAAGAGCACGAATC
(Yamauchi et al., 2008) R ATGAAGCACAGAGGCTGGAG
cyp3a38 F AGGAAACAGAGATCCCCTTCGA
(Zhang et al., 2008b) R AGGCACCAGCTTCAGAAAGATG
cyp3a40 F AGATGGATGTCCAGGGTTTCC
(陳, 2011) R GGCGTGAGCTTCAGTTGGAT
aldh1a2 F GGTTTCCACCAGGAGTTGTCAA
(Zhang et al., 2008b) R GATAAGCTTGCCGACCTCAGTT
cyp26b1 F CTGCCTCGACATTCTGATCCTC
(Zhang et al., 2008b) R TTGTGCATCTGTGTGCGGTT
rarα1 F TGGACATTCTGATTCTGCGGAT
(Zhang et al., 2008b) R GGAGAAGGTCATGGTGTCCTGT
rarγ1 F CCTCGTGTCTACAAACCCTGCT
(Zhang et al., 2008b) R TGCGACGGAAGAAACCCTT
rxrα1 F CCGAGCAGGTTGGAATGAACT
(Zhang et al., 2008b) R CCCGTCTTTCACTGCTATGGAA
rxrα2 F TGGGGAGAACGAGGTTGAGT
(Zhang et al., 2008b) R ATCTTGTCCACGGGC ATCTC
rxrβ1 F TAGTGCCCATAGTGCTGGTGTT
(Zhang et al., 2008b) R CGAGTTGGACAAACCCTTAGCAT
25
rxrβ2 F CTGCCTTCGAGCTATTGTTCTG
(Zhang et al., 2008b) R TTAGAGAGCCCTTTAGCATCTGG
p53 F CGCCAGATCTTCAACGGTGTAT
(Zhang et al., 2008b) R AGGCTCAGCAATCCTCAGCAT
表2-3 StepOne 儀器預設之即時定量聚合酶連鎖反應條件。
步驟 溫度 (℃) 時間 (秒)
Holding 95 600
Cycling (40 cycles)
95 15 60 60
Melting Temperature
95 15 60 60 95 15
26
2.6.2. CYP 酵素活性分析 2.6.2.1. 微粒體萃取
微粒體萃取實驗方法為參考Tatarazako 等學者之研究,並將實驗步驟做部分修 正 (Tatarazako et al., 2002)。將 2 隻青鱂魚之肝臟合併為 1 樣品,在冰冷的均質用 緩衝液中均質,並在4℃中離心 (10,000 g,10 分鐘) 後,將上清液於 4℃中進行超 高速離心 (120,000 g,120 分鐘),再將沉澱物回溶於 200 μL 冰冷的保存用緩衝液 中,取出25 μL 以 Pierce® BCA Protein Assay Kit 試劑組測定微粒體蛋白濃度 (實驗 步驟如原廠操作手冊),其餘置於-80℃冰箱中保存。
2.6.2.2. EROD 法
EROD 法步驟為參考 Tatarazako 等學者之研究 (Tatarazako et al., 2002),並經 反應條件測試後修正部分反應條件。實驗步驟為取20 μg 微粒體蛋白 22℃下回溫 5 分鐘後,與基質ethoxyresorufin 和其他反應輔助因子於 22℃反應 20 分鐘,反應中
各成分濃度如表 2-4 所示。反應完成後,以甲醇停止反應,再以多功能微盤讀儀
FlexStation3 microplate reader 測定反應產物放射之螢光,測定條件為激發光 550 nm,
發散光585 nm。
2.6.2.3. DBF 試驗
DBF 試驗步驟為參考 Kashiwada 等學者之研究 (Kashiwada et al., 2007),並經 反應條件測試後修正部分反應條件。實驗步驟為取20 μg 微粒體蛋白於 25℃下回 溫5 分鐘後,與基質 dibenzylfluorescein 和其他反應輔助因子於 25℃反應 15 分鐘,
反應中各成分濃度如表2-5 所示。反應完成後,以 2N 氫氧化鈉停止反應,再以多 功能微盤讀儀FlexStation3 microplate reader 測定反應產物放射之螢光,測定條件為 激發光485 nm,發散光 538 nm。
27
表2-4 EROD 法反應中各成分濃度。
成分 濃度
Potassium phosphate buffer (pH 7.7) 50 mM
Magnesium chloride 5 mM
NADP+ 0.5 mM
Glucose-6-phosphate dehydrogenase 1 IU
Ethoxyresorufin 2 μM
Glucose-6-phosphate 10 mM
表2-5 DBF 試驗反應中各成分濃度。
成分 濃度
Potassium Phosphate Buffer (pH 7.4) 200 mM
Magnesium chloride 30 mM
NADP+ 2 mM
Glucose-6-phosphate dehydrogenase 0.5 IU
Dibenzylfluorescein 1 μM
Glucose-6-phosphate 25 mM
2.7. 統計分析
統 計 分 析 以 statistics analysis system (SAS) 9.1 軟體之單因子變方分析 (one-way analysis of variance, one-way ANOVA) 進行,並以 Dunnett’s t test 進行所 有處理組與DMSO 溶劑控制組之比較。各級差異顯著水準隻表示如下:*:p < 0.05;
**:p < 0.01;***:p < 0.001。所有圖表數據皆以平均值 ± 標準誤 (standard error) 表示。
28
3. 結果與討論
3.1. 引子品質及酵素分析條件測試
3.1.1. 引子品質測試
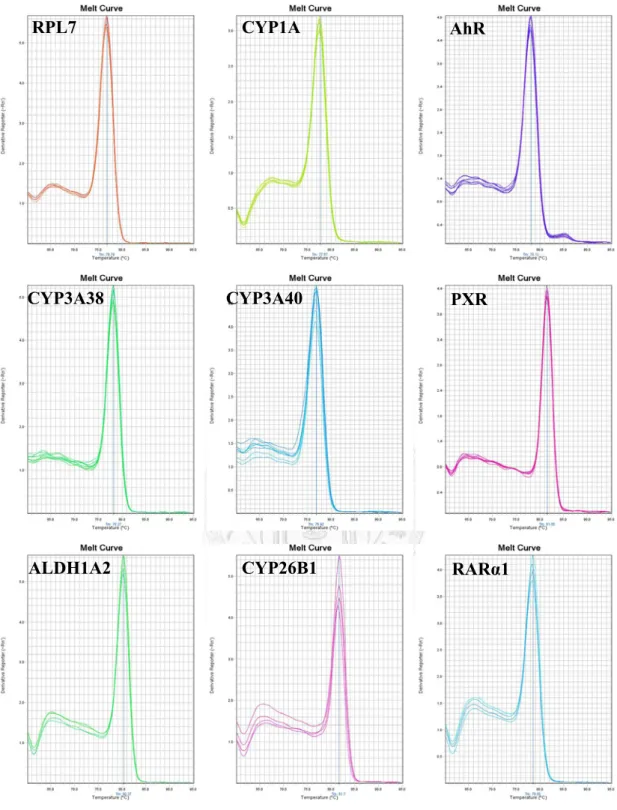
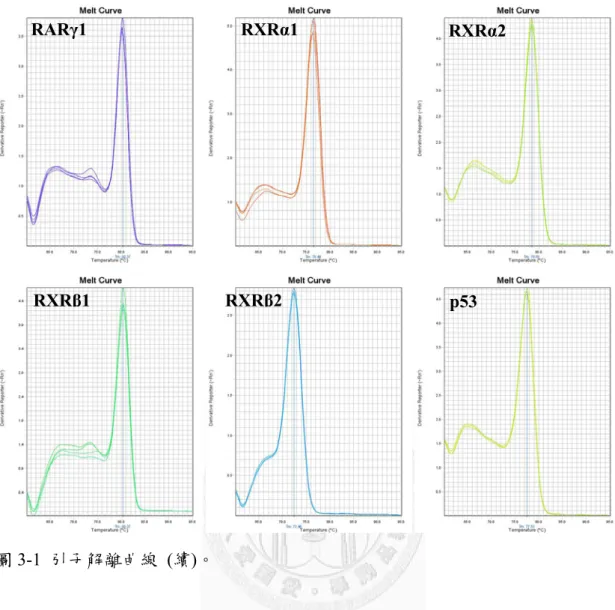
圖3-1 為本研究使用之引子之解離曲線,所有使用引子之 PCR 反應產物皆為 單一波峰,顯示反應中無非專一性結合或引子雙體等非預期之反應產物干擾反應 訊號。
3.1.2. 酵素分析條件測試
微粒體蛋白回溫時間以DBF 試驗測試,測試時間包括 3, 5 及前人研究使用之 10 分鐘。測試結果如圖 3-2 所示,在微粒體蛋白回溫 5 分鐘後進行的反應得到的
訊號值最高,而在回溫10 分鐘時其訊號值下降,故將接續之反應條件測試及正式
實驗中微粒體蛋白回溫修正為5 分鐘。
EROD 法反應時間測試條件包括 10, 20 及 30 分鐘,測試結果如圖 3-3a 所示,
測得之訊號值在30 分鐘內呈線性關係,與文獻之結果相符 (Tatarazako et al., 2002)。
故接續之反應條件測試及正式實驗中EROD 法之反應時間固定為文獻中使用之 20 分鐘 (Tatarazako et al., 2002)。而 DBF 試驗反應時間測試條件包括 5, 10, 15 及 20 分鐘,測試結果如圖3-3b 所示,反應 15 至 20 分鐘間測得之訊號改變量開始有下 降之現象,可能為反應進行效率降低、反應原料不足或訊號飽和等因素造成,故 接續之反應條件測試及正式實驗中DBF 試驗之反應時間固定為 15 分鐘。
EROD 法之正控制組以文獻中 CYP1A 誘導劑 β-naphthoflavone 進行 (Beijer et al., 2010),測試結果如圖 3-4 所示,青鱂魚暴露 1 mg/L β-naphthoflavone 達 48 小時 後,其肝中EROD 活性提升為 DMSO 控制組的 68.5 倍。EROD 法和 DBF 試驗之 酵素抑制實驗則以CYP1A, CYP3A 之活性抑制劑克康那唑進行 (Beijer et al., 2010;
Greenblatt et al., 2011),測試結果如圖 3-5 所示,微粒體蛋白加入 5 及 50 μL 克康 那唑後反應測應測得之訊號降低至與背景值相近。
29
圖3-1 引子解離曲線。
RPL7 CYP1A AhR
CYP3A38 CYP3A40 PXR
ALDH1A2 CYP26B1 RARα1
30
圖3-1 引子解離曲線 (續)。
RARγ1 RXRα1 RXRα2
RXRβ1 RXRβ2 p53
31
圖3-2 微粒體蛋白回溫時間與 DBF 試驗訊號關係。
32
圖3-3 (a) EROD 法與 (b) DBF 試驗反應時間與訊號關係。
a
b
33
圖3-4 CYP1A 活性誘導試驗。圖中 DMSO 為 0.01% DMSO 溶劑控制組,βNF 為 1 mg/L 之 CYP1A 誘導劑 β-naphthoflavone。暴露時間為 48 小時,其於暴露環境條 件同CYP 酵素活性分析實驗。βNF 處理組與 DMSO 控制組之比較以 ANOVA 進行。
各級差異顯著水準之表示如下: ***:p < 0.001。誤差線表示處理平均值的標準誤 (SE)。
34
圖3-5 (a) CYP1A 與 (b) CYP3A 活性抑制實驗。圖中 DMSO 和 Tri 3.5 為 CYP 酵 素活性分析實驗之樣品,Ket 為 CYP 活性抑制劑 ketoconazole,ERS 為能量再生系 統 (NADP+, glucose-6-phosphate dehydrogenase and glucose-6-phosphate)。
35
3.2. mRNA 表現量分析預實驗
前人文獻指出投以亞致死劑量的三泰芬及邁克尼會誘導小鼠肝中 CYP1A 及 CYP3A 之 mRNA 表現 (Allen et al., 2006; Chen et al., 2009; Goetz and Dix, 2009;
Goetz et al., 2006)。故本研究將青鱂魚暴露於 2 mg/L 三泰芬及邁克尼 3 天後採樣肝 臟分析CYP1A 及 CYP3A 之 mRNA 表現量。實驗結果如圖 3-6 所示,CYP1A 及
核受體 PXR 之 mRNA 表現於三泰芬及邁克尼處理組中皆無上升現象,而兩種
CYP3A 形式 CYP3A38 及 CYP3A40 之 mRNA 表現則於三泰芬及邁克尼處理組中
皆有上升趨勢。顯示三泰芬及邁克尼可能具有誘導CYP3A 基因表現及酵素活性之
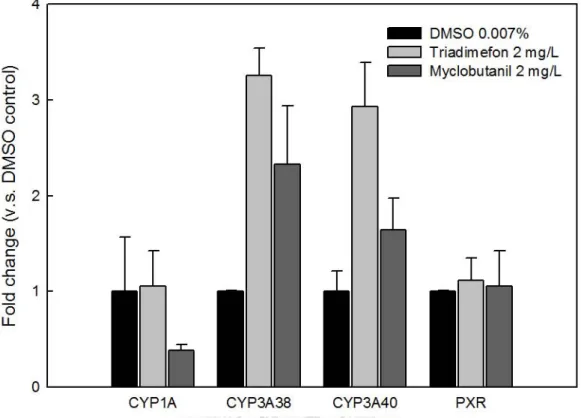
能力。但預實驗中因操作不慎而使DMSO 控制組之樣本數僅有 2 隻,無法進行統 計分析。而為得到更明顯之反應結果,mRNA 表現量分析之正式實驗中將暴露時 間延長為7 至 14 天,暴露濃度降低為 2.0-5.0 μM。
36
圖3-6 青鱂魚暴露於處理組及控制組 3 天後肝中 CYP1A, CYP3A38, CYP3A40 及 PXR 之 mRNA 表現情形。誤差線表示處理平均值的標準誤 (SE)。
37
3.3. 肝體指數
肝體指數為生物肝臟重與體重之比值。普遍而言,在條件較惡劣的環境下,
生物體內儲存的能量較少,而肝體指數較低,因此肝體指數一般用於評估環境是 否適合動物生存。而在毒理研究中,許多研究發現暴露毒物後,生物之肝體指數 相較於控制組有顯著提升。文獻指出,亞致死劑量之三泰芬及邁克尼可顯著提升
小鼠及大鼠之肝體指數,其機制可能為三泰芬及邁克尼誘導CYPs 蛋白大量表現,
進而造成肝腫大所致 (Allen et al., 2006; Chen et al., 2009; Sun et al., 2007)。故本研 究將青鱂魚暴露於2.0-5.0 μM 三泰芬及邁克尼,並於暴露第 7 天及第 14 天採樣肝 臟測量肝體指數,觀察肝體指數是否可做為三泰芬及邁克尼投藥成功之生物指標。
結果顯示,本實驗所有處理組之肝體指數與控制組間均無顯著差異 (表 3-1),顯示 肝體指數可能並非三泰芬及邁克尼於青鱂魚中投藥成功之良好生物指標。
目前在青鱂魚相關毒理研究中,肝體指數主要是做為暴露雌激素或類雌激素 之生物指標。文獻指出雌激素可誘導卵黃前質蛋白 (vitellogenin, VTG) 大量表現,
因而可使肝體指數顯著提升 (Ma et al., 2007; Ma et al., 2005; Zhang and Hu, 2008)。
而本實驗中肝體指數無顯著變化可能是本研究之暴露處理誘導之酵素表現量並未 如VTG 效應可造成顯著的肝體指數差異所致。
38
表3-1 青鱂魚暴露於處理組及控制組 7 天及 14 天後的肝體指數 (HSI)。
a.) 7 天
Conazole 處理組別 體重 (mg) 肝重 (mg) HSI (%) Controla 310.46 ± 14.88 4.68 ± 0.28 1.51 ± 0.06 DMSOa 0.001 % 288.94 ± 10.88 4.66 ± 0.35 1.61 ± 0.10 三泰芬 2 μMa 281.12 ±
07.16
4.64 ± 0.26 1.65 ± 0.08 3.5 μMb 279.31 ±09.67
4.52 ± 0.40 1.61 ± 0.12 5 μMc 312.92 ± 19.99 4.79 ± 0.70 1.51 ± 0.17 邁克尼 2 μMa 314.09 ±06.87
4.37 ± 0.31 1.38 ± 0.07 3.5 μMa 296.55 ±08.18
4.03 ± 0.29 1.37 ± 0.10 5 μMc 317.16 ± 29.50 4.74 ± 0.53 1.49 ± 0.05a n=13, b n=12, c n=5
b.) 14 天
Conazole 處理組別 體重 (mg) 肝重 (mg) HSI (%) Controla 303.00 ± 25.77 4.38 ± 0.52 1.43 ± 0.08 DMSOa 0.001 % 335.30 ± 15.33 4.72 ± 0.30 1.40 ± 0.05 三泰芬 2.0 μMb 306.55 ± 28.89 4.74 ± 0.56 1.54 ± 0.05 3.5 μMb 262.83 ± 32.75 5.09 ± 1.12 1.89 ± 0.29 5.0 μMb 314.55 ± 43.34 5.23 ± 1.04 1.62 ± 0.12 邁克尼 2.0 μMa 280.16 ± 30.28 3.88 ± 0.97 1.33 ± 0.24 3.5 μMa 280.88 ± 17.39 4.38 ± 0.52 1.54 ± 0.11 5.0 μMa 309.58 ± 22.54 5.93 ± 0.81 1.89 ± 0.15
a n=5, b n=4
39
3.4. CYP1A 基因表現及酵素活性
CYP1A 酵素活性為毒理研究中重要的生物指標之一,可代表實驗處理對
CYP1A 代謝基質能力之影響程度。前人研究指出,投以亞致死劑量的三泰芬及邁 克尼會使小鼠肝中CYP1A 活性顯著提升 (Allen et al., 2006; Goetz and Dix, 2009;
Goetz et al., 2006)。故本研究將青鱂魚暴露於 2.0-5.0 μM 三泰芬及邁克尼,並於暴 露第 7 天及第 14 天採樣肝臟分析 CYP1A 之 mRNA 表現量;另將青鱂魚暴露於 2.0-3.5 μM 三泰芬及邁克尼 7 天後,採樣肝臟以 EROD 法分析 CYP1A 酵素活性,
觀察三泰芬及邁克尼對青鱂魚肝中CYP1A 活性之影響。結果顯示,暴露 2.0 及 3.5 μM 三泰芬 7 天後,青鱂魚肝中 CYP1A 活性 (以 EROD 活性表示) 分別提升為約 2.8 及 3.9 倍,顯示此濃度下的三泰芬可誘導 CYP1A 酵素活性。相同暴露條件下,
邁克尼則不影響青鱂魚肝中CYP1A 活性 (圖 3-7)。然而,青鱂魚 2.0-5.0 μM 三泰 芬7 天後,肝中 CYP1A 之 mRNA 表現量於暴露下降約 36-63%,相同暴露條件下 邁克尼則使CYP1A 之 mRNA 表現量下降約 25-47%,但因組內個體差異較大而均 未達顯著水準 (圖 3-8a)。另外,前人研究指出 CYP1A 的基因及蛋白表現主要經由 AhR 調控 (Tompkins and Wallace, 2007),故本研究也進行青鱂魚肝中 AhR 之 mRNA 表現量分析 (實驗條件同 CYP1A 之 mRNA 表現量分析),結果顯示所有處 理組和控制組相比皆無顯著差異 (圖 3-8b),即無法判定 CYP1A 活性的提升是否為 經由AhR 途徑。
CYP1A 在許多脊椎動物中參與雌激素及多種環境汙染物之代謝。研究指出許 多境汙染物如PAH、戴奧辛 (dioxin) 等皆會誘導生物肝中 CYP1A 表現 (Monostory et al., 1996; Smolowitz et al., 1992)。然而 CYP1A 活性的提升一方面可能會過量代 謝內生雌激素,進而造成干擾性別分化、發育、生殖行為及能力等負面影響 (Williams et al., 1998);另一方面也會干擾多種環境汙染物及藥物在生物體內之代 謝情形。在CYP1A 代謝之外來物質中,目前以 PAH 之研究最為詳盡。CYP1A 可
40
藉由氧化PAH 以增加其水溶性,或產生高反應性的代謝產物以利第二階段代謝之
進行,如環氧化物 (epoxide) 即是高反應性之 CYP1A 代謝產物之一。但文獻指出 部分PAH 之環氧化物具致癌性,故 CYP1A 活性提升反而可能引起生物活化反應,
進而產生有毒或致癌之代謝產物 (Gabelova et al., 2004; Selkirk, 1977)。另外,文獻 指出人類及小鼠之 CYP1A 具有代謝 atRA 之活性 (Chen et al., 2000),顯示誘導 CYP1A 活性亦可能加速代謝體內 atRA,因而造成維生素 A 缺乏的可能性亦較高。
因此,提高青鱂魚肝中CYP1A 活性可能會干擾內生雌激素平衡、生物活化現象及
atRA 恆定,顯示三泰芬對水生生物造成的致癌性可能較邁克尼大。
在小鼠實驗中,亞致死劑量的三泰芬及邁克尼皆可誘導肝中 EROD 活性提升 (Allen et al., 2006; Goetz and Dix, 2009; Goetz et al., 2006);而本研究發現在青鱂魚 中僅有三泰芬會誘導肝中 EROD 活性,顯示康唑類殺真菌劑對 CYPs 活性之影響 具物種差異性,也顯示以環境生物評估農業用藥對生態影響的重要性。
本實驗中並未測得CYP1A 之 mRNA 表現量提升,可能是由於 CYP1A 的 mRNA 與蛋白表現有一時間差 (Leguen et al., 2010),因此本實驗設計僅測得 CYP1A 活性 提升,並未捕抓到最高的 mRNA 表達量;而暴露 14 天時 CYP1A 及 AhR 之 mRNA 表現量有下降趨勢,則可能是因為高劑量的暴露已造成某一程度的肝毒性,因而 抑制了CYP1A 及 AhR 之 mRNA 表現量,因此酵素活性試驗並未進行 14 天的暴 露處理。
41
圖3-7 青鱂魚暴露於處理組及控制組 7 天後肝中 CYP1A 活性 (以 EROD 活性表示)。
圖中Control 為除氯水控制組,DMSO 為 0.001% DMSO 溶劑控制組,Tri 為三泰芬,
Myc 為邁克尼,2.0 及 3.5 為濃度 (μM)。所有處理與 DMSO 控制組之比較以 Dunnett’s t test 進行。各級差異顯著水準之表示如下:*:p < 0.05;***:p < 0.001。
誤差線表示處理平均值的標準誤 (SE)。
42
圖3-8 青鱂魚暴露於處理組及控制組 7 天及 14 天後肝中 (a) CYP1A 及 (b) AhR 之 mRNA 表現量。圖中 Control 為除氯水控制組,DMSO 為 0.001% DMSO 溶劑控制 組,Tri 為三泰芬,Myc 為邁克尼,2.0-5.0 為濃度 (μM)。所有處理與 DMSO 控制 組之比較以Dunnett’s t test 進行。各級差異顯著水準之表示如下:*:p < 0.05。誤 差線表示處理平均值的標準誤 (SE)。
a
b
43
3.5. CYP3A 基因表現及酵素活性
CYP3A 為 CYPs 中表現量最大,可代謝基質最多的 subfamily。前人研究指出,
投以亞致死劑量的三泰芬及邁克尼會使小鼠肝中 CYP3A 活性顯著提升 (Allen et al., 2006; Chen et al., 2009; Sun et al., 2007)。故本研究將青鱂魚暴露於 2.0-5.0 μM 三 泰芬及邁克尼,並於暴露第 7 天及第 14 天採樣肝臟分析 CYP3A38 及 CYP3A40 之mRNA 表現量;另將青鱂魚暴露於 2.0-3.5 μM 三泰芬及邁克尼 7 天後,採樣肝 臟以DBF 為基質分析 CYP3A 酵素活性,觀察三泰芬及邁克尼對青鱂魚肝中 CYP3A 活性之影響。結果顯示,暴露2.0 及 3.5 μM 三泰芬 7 天後,青鱂魚肝中 CYP3A 活 性 (以 DBF 脫烷活性表示) 分別提升為約 3.0 及 2.8 倍,相同暴露條件下邁克尼則 使CYP3A 活性提升為約 3.2 及 4.3 倍,顯示此暴露條件下三泰芬及邁克尼皆可誘 導CYP3A 酵素活性 (圖 3-9)。基因表現部分,預實驗結果顯示青鱂魚暴露 6.8 μM 三泰芬及6.9 μM 邁克尼後 3 天後,肝中 CYP3A38 之 mRNA 表現量分別提升為約 3.3 及 2.3 倍,CYP3A40 之 mRNA 表現量則分別提升為約 2.9 及 1.6 倍 (圖 3-6)。
但14 天暴露實驗中除暴露 2.0 μM 三泰芬 7 天後青鱂魚肝中 CYP3A40 之 mRNA 表現量提升69% 達顯著差異外,其餘組別之 CYP3A38 及 CYP3A40 之 mRNA 表 現量於暴露 7 天後皆無顯著改變,暴露達 14 天時 mRNA 表現量更有下降之趨勢 (圖 3-10a, b)。另外,前人研究指出 CYP3A 的基因及蛋白表現主要經由 PXR 調控,
故本研究也進行青鱂魚肝中PXR 之 mRNA 表現量分析 (實驗條件同 CYP3A38 及 CYP3A40 之 mRNA 表現量分析),結果顯示,暴露 2.0-5.0 μM 三泰芬 7 天後,青 鱂魚肝中PXR 之 mRNA 表現量降低 64-74%,相同暴露條件下邁克尼則使 PXR 之 mRNA 表現量降低 33-50% (圖 3-11)。
文獻指出,CYP3A 代謝機制多為受基質調控,當雌激素或藥物等基質進入細 胞後,會與核受體PXR 結合,進而誘發 CYP3A 基因表現,再進而產生 CYP3A 蛋 白以代謝基質 (Tompkins and Wallace, 2007)。由本實驗中三泰芬及邁克尼可誘導
44
CYP3A 活性及 CYP3A38, CYP3A40 之 mRNA 表現之結果推測,青鱂魚中之 CYP3A38 及 CYP3A40 可能皆具有代謝三泰芬及邁克尼之能力,但是否以 PXR 途 徑誘發CYP3A 活性則仍不清楚。
然而CYP3A 除了參與多種藥物及環境物質的代謝與解毒外,也參與內生雌激
素或雄激素等固醇類荷爾蒙的代謝,因此提升CYP3A 活性亦可能干擾生物之性別
分化、發育、生殖行為及能力等機制。此外,在小鼠實驗中由三泰芬及邁克尼誘 發的CYP3A 酵素活性會提升對 atRA 的代謝能力 (Chen et al., 2009),可能造成體 內atRA 濃度下降,進而影響維生素 A 相關生理機制之正常運作。
45
圖3-9 青鱂魚暴露於處理組及控制組 7 天後肝中 CYP3A 活性 (以 DBF 脫烷活性 表示)。圖中 Control 為除氯水控制組,DMSO 為 0.001% DMSO 溶劑控制組,Tri 為三泰芬,Myc 為邁克尼,2.0 及 3.5 為濃度 (μM)。所有處理與 DMSO 控制組之 比較以Dunnett’s t test 進行。各級差異顯著水準之表示如下:*:p < 0.05;***:p <
0.001。誤差線表示處理平均值的標準誤 (SE)。
46
圖 3-10 青鱂魚暴露於處理組及控制組 7 天及 14 天後肝中 (a) CYP3A38 及 (b) CYP3A40 之 mRNA 表現量。圖中 Control 為除氯水控制組,DMSO 為 0.001% DMSO 溶劑控制組,Tri 為三泰芬,Myc 為邁克尼,2.0-5.0 為濃度 (μM)。所有處理與 DMSO 控制組之比較以Dunnett’s t test 進行。各級差異顯著水準之表示如下:*:p < 0.05。
誤差線表示處理平均值的標準誤 (SE)。
a
b
47
圖3-11 青鱂魚暴露於處理組及控制組 7 天及 14 天後肝中 PXR 之 mRNA 表現量。
圖中Control 為除氯水控制組,DMSO 為 0.001% DMSO 溶劑控制組,Tri 為三泰芬,
Myc 為邁克尼,2.0-5.0 為濃度 (μM)。所有處理與 DMSO 控制組之比較以 Dunnett’s t test 進行。各級差異顯著水準之表示如下:**:p < 0.01。誤差線表示處理平均值 的標準誤 (SE)。
48
3.6. atRA 恆定相關酵素之 mRNA 表現
本研究以q-RT-PCR 分析生物體內分別合成及代謝 atRA 的酵素 ALDH1A2 及 CYP26B1 之 mRNA 表現情形。結果顯示,暴露 2.0-5.0 μM 三泰芬 7 天後可誘導青 鱂魚肝中CYP26B1 之 mRNA 表現提升為約 3.0-4.2 倍,相同暴露條件下之邁克尼 則不造成影響 (圖 3-12b)。而暴露 2.0-5.0 μM 三泰芬及邁克尼 7 天皆不影響 ALDH1A2 之 mRNA 表現 (圖 3-12a)。
CYP26 為生物體內代謝 atRA 之酵素,正常情形下為受 atRA 誘導表現,藉以 維持體內atRA 濃度,避免過量 atRA 造成之傷害 (Thatcher and Isoherranen, 2009)。
以2.0-5.0 μM 三泰芬暴露 7 天後青鱂魚肝中 CYP26B1 之 mRNA 表現量提升,顯 示此濃度之三泰芬可能藉由誘導 CYP26 而提升對體內 atRA 之代謝,並有進而造 成維生素A 缺乏的可能。
ALDH1A 為生物體內合成 atRA 之酵素,可將視黃醛 (retinal) 代謝為具有活 性的 atRA,而文獻指出青鱂魚基因中缺少 ALDH1A1 及 ALDH1A3,因此僅有 ALDH1A2 一種合成 atRA 之酵素 (Canestro et al., 2009)。而暴露 2.0-5.0 μM 三泰芬 及邁克尼7 天皆不影響青鱂魚肝中 ALDH1A2 之 mRNA 表現,顯示此條件下三泰 芬及邁克尼對atRA 合成機制之影響可能較不明顯。
49
圖3-12 青鱂魚暴露於處理組及控制組 7 天後肝中 (a) ALDH1A2 及 (b) CYP26B1 之mRNA 表現量。圖中 Control 為除氯水控制組,DMSO 為 0.001% DMSO 溶劑控 制組,Tri 為三泰芬,Myc 為邁克尼,2.0-5.0 為濃度 (μM)。所有處理與 DMSO 控 制組之比較以Dunnett’s t test 進行。各級差異顯著水準之表示如下:**:p < 0.01;
***:p < 0.001。誤差線表示處理平均值的標準誤 (SE)。
a
b
50
3.7. atRA 相關核受體之 mRNA 表現
文獻指出,當 atRA 相關之核受體表現量受改變時,可能會干擾細胞對 atRA 之感知能力,而使 atRA 相關生理機制無法受到正確 atRA 濃度的調控 (Sun and Lotan, 2002)。且已在多種癌症前期細胞及組織中發現 atRA 相關之核受體表現量有 降低情形 (Berard et al., 1996; Castillo et al., 1997; Houle et al., 1993; Lotan et al., 1995; Lotan et al., 2000; Picard et al., 1999; Qiu et al., 1999; Widschwendter et al., 1997; Xu et al., 1994; Xu et al., 1997a; Xu et al., 1997b)。雖然目前對於 atRA 相關核
受體表現、維生素 A 缺乏及其相關致癌性的詳細關聯性及機制仍不明瞭,但為瞭
解在小鼠中具致肝腫瘤性的三泰芬與不具致肝腫瘤性的邁克尼對青鱂魚肝中atRA
相關核受體表現量之影響間是否有所差異,本研究以 q-RT-PCR 分析青鱂魚肝中 atRA 相關之核受體 RARs 及 RXRs 的 mRNA 於暴露 2.0-5.0 μM 三泰芬及邁克尼 7 天後之表現情形。結果顯示,青鱂魚肝中RXRα1 之 mRNA 表現量於暴露 2.0, 3.5, 5.0 μM 三泰芬 7 天後分別降低約 49, 39, 37%,而暴露 2.0, 5.0 μM 邁克尼後 7 天後 分別降低約49, 42% (圖 3-13c);RXRβ1 之 mRNA 表現量亦於暴露 2.0, 5.0 μM 三泰 芬後分別降低約70, 84%,暴露 2.0 μM 邁克尼後則降低約 53% (圖 3-13e);RARα1, RXRα2 及 RXRβ2 之 mRNA 表現量不受實驗處理影響 (圖 3-13a, d, f);而 RARγ1 之mRNA 表現量則於暴露 2.0 μM 三泰芬後提升約 1.5 倍 (圖 3-13b)。由以上結果
可發現,三泰芬及邁克尼皆可使部分atRA 相關核受體表現量降低,且兩者造成的
影響相當相似,顯示三泰芬及邁克尼皆可能影響atRA 相關生理機制的正常運作,
但無法判定是否干擾atRA 相關核受體表現量與康唑類殺真菌劑是否具有致肝腫瘤
性之關聯性。
當生物體內atRA 濃度過高時,atRA 會與 RAR 結合,RAR 再與 RXR 形成異 構雙體後,會誘發 CYP26B1 基因及酵素表現,代謝過量 atRA,藉以維持相關生 理機制之恆定 (Chambon, 1996)。RARα, RARγ 為同功酶,依物種、組織及發育時
51
期的不同,RARs 之表現量比例會有所不同,亦可能僅有單一 RAR 表現 (Chambon, 1996)。而實驗結果顯示三泰芬可誘導 RARγ1 及 CYP26B1 之 mRNA 表現量提升,
顯示三泰芬可能經由誘發RARγ1 途徑誘導 CYP26B1 表現。