國立臺灣大學獸醫專業學院獸醫學研究所 碩士論文
Graduate Institute of Veterinary Medicine School of Veterinary Medicine
National Taiwan University Master Thesis
由 2014 至 2017 年從就診臺大動物醫院的犬貓所分離帶 有超廣譜性乙內醯胺酶大腸桿菌的盛行率及特性分析
Prevalence and characterization of the
extended-spectrum-β-lactamase (ESBL)-producing Escherichia coli isolated from the dogs and cats admitted to National Taiwan
University veterinary hospital from 2014 to 2017
黃伊萱 Yi-Hsuan Huang 指導教授:葉光勝 博士
Advisor: Kuang-Sheng Yeh, DVM, Ph. D 中華民國 108 年 6 月
June, 2019
誌謝
研究所是求學生涯過程中一個重要的階段,感謝指導教授 葉光勝博士在課 業及研究上各方面的教導,並以像爸爸般的角色關心著研究生們的生活,在學生 實驗遇到瓶頸時會適時的給予最大幫助,讓學生的實驗可以繼續順利地進行下去,
並給學生在實驗上有很大自由發揮的空間。也要謝謝教授包容學生的不細心,在 碩一透過擔任班代職務跟著教授學習如何細心地處理事務,讓原本不在乎小細節 的我也開始懂的做事的一些注意事項,並非交代一件事情完成就好而是連帶衍生 的問題也應該一併想清楚並完成,讓學生能夠不在像以前一樣莽撞做事。也非常 感謝在大學時期在 黃素華教授的帶領下,讓學生更加確定想要繼續就讀碩班的 想法,也是因為有素華教授的提攜使學生能夠進入臺大研究所繼續在學術領域上 深耕,學生在此對兩位教授獻上十二萬分的感謝。
感謝實驗室畢業的學長姐 (克銓學長、南綾學姊、正芳學姊等人)、同儕間 (幸 蓉、家容) 以及學妹 (佩歆、化臻) 的幫忙,感謝正芳學姊在我碩一時,帶領我 進入實驗室並熟悉環境,在課業上也會不吝嗇地分享筆記給我。感謝南綾學姊在 繁重工作之下還抽空幫我解決實驗上問題,讓我的實驗能夠順利完成。感謝克銓 學長把我的事情當成是他重要的事情般,一有問題向他請教他都能在最快的時間 為我解惑。也要感謝從大學到碩班一直以來陪伴我的實驗夥伴幸蓉,因為有妳才 讓我的碩班生涯不是當邊緣人的存在,一起在實驗室學習、接受新的挑戰,相互 交流討論一起努力一起成長,有妳的陪伴實驗室的生活變得美好且愉快。
感謝我的家人,爸爸、媽媽、妹妹們以及親友們在這求學路上,一路的支持、
鼓勵與陪伴,在遇到瓶頸的時候從旁默默的關心與付出,讓我知道到我不是一個 人在這孤單的道路上,在我低潮想放棄的時候給予我與許多關心並鼓勵我要永不 放棄,協助我走出低潮,讓我繼續朝著目標邁進。也要感謝我的兩位口試委員宣 詩玲和王裕智兩位教授,悉心指導學生的論文以及提供寶貴的意見,讓學生的論 文能夠順利完成且更加完整。最後感謝曾經在實驗上或者生活上給予幫助過的任 何人,讓我可以在這短短兩年碩班生涯中學習到很多,感謝有你們使得我碩班生 涯添加許多不同色彩在此敬致由衷的感謝。
黃伊萱 謹誌於 國立臺灣大學 獸醫學研究所 中華民國一百零八年六月
中文摘要
超廣譜性乙內醯胺酶(extended-spectrum-β- lactamases, ESBLs)為可以水解 β-內醯 胺類抗菌藥物的酵素,包括第三代頭孢子菌素,如獸醫常用的 ceftiofur。ESBL 的 產生主要是藉由質體傳遞,並且此類質體通常包含對其他種類的抗生素如胺基配 糖體和氟喹諾酮類具有抗藥性的基因,限制了可用於治療的有效抗微生物劑的選 擇。ESBLs 廣泛分佈於腸桿菌科的成員中,特別是在大腸桿菌中。在犬貓中最常 見的疾病為尿路感染,然而引起尿路感染 (urinary tract infections, UTIs) 的病原體 又以大腸桿菌居多,這時就需要抗生素的治療,進而可能會導致 ESBL-producing E.
coli 的產生。儘管帶有 ESBL 的微生物是全球性問題,但在臺灣,目前對於在獸醫 中產 ESBL 的微生物的報告有限。因此,本研究目的在於分析 2014 至 2017 年間,
就診臺大動物醫院的狗貓所分離帶有超廣譜性乙內醯胺酶大腸桿菌的盛行率及特 性。本研究收集了 285 個大腸桿菌的檢體,並經由臨床與實驗室標準協會所規範 的雙盤紙錠試驗去鑑定 ESBL 表現型,結果發現有 65 株大腸桿菌帶有 ESBL (22.8%)。所有的 65 株分離株都對 ampicillin 呈抗藥性,對 β- lactams 類的 ceftiofur、
四環素類的 doxycycline 以及喹諾酮類的 enrofloxacin、ciprofloxacin,則分別呈現 93.8%、73.8%、80%,以及 78.5% 的抗藥性。對於 β-lactams 類的 imipenem 與胺 基配糖體類的 gentamicin,則有相對較高的感受性。CTX-M-1 和 CTX-M-2 group 為最主要的 ESBL 基因型。屬於 CTX-M-1 group 的分離株共有 38 株 (58.5%),以 CTX-M-55 (24/65, 36.9%) 為主要的 ESBL 基因。屬於 CTX-M-2 group 的分離株共 12 株 (18.5%),其 ESBL 基因均為 CTX-M-124 (12/65, 18.5%)。多重基因座序列分 型法結果顯示 ST457、ST131、ST648 為最常見的無性繁殖系 (clone)。其中 ST131、
ST648 又是屬於全球流行高危險群的無性繁殖系。然而,ST131 O25b 是引起全球
人畜共通傳染並造成公共衛生問題的 clone。在本研究中發現有 8 株大腸桿菌屬於 ST131 O25b,又以 CTX-M-14 和 CTX-M-15 的衍生物為主要 ESBL 基因型。伴侶 動物如犬貓與人類會有親密的接觸,有可能成為人類產生 ESBL 細菌的潛在來源。
因此,從公共衛生和獸醫學角度來看,鑑定與分析 ESBL 細菌的抗藥性問題與流 行病學,是一個值得繼續深入探討的議題。[1]
關鍵詞:超廣譜性乙內醯胺酶、大腸桿菌、多重抗藥性、多重基因座序列分型法、
ST131/O25b
Abstract
Extended-spectrum-β- lactamases (ESBLs) is a group of enzymes that can hydrolyze β-latctams antimicrobial agents, including the third generation cephalosporins such as ceftiofur frequently used in veterinary medicine. Production of ESBL is primarily plasmid-mediated and such plasmids often comprise the genes that exhibit resistance to other classes of antimicrobials like aminoglycosides and fluoroquinolones, limiting the choice of effective antimicrobial agents available for treatment. ESBLs are widely distributed in the members of the family Enterobacteriaceae, particularly in Escherichia coli. In both dogs and cats, the most common pathogen was E. coli, which could cause urinary tract infections (UTIs) and should be treated by antimicrobial agents. Urinary tract infections (UTIs) is the most common disease in dogs and cats, and the most commonly seen pathogen is E. coli. Therapy of UTIs requires antimicrobials; however, usage of such antimicrobials may also result in producing of ESBL-producing E. coli.
Although presence of ESBL-producing microorganisms is a global problem, the information regarding the ESBL-producing E. coli is limited in veterinary medicine in Taiwan. Therefore, the objective of this study is to characterize the E. coli strains isolated in National Taiwan University Veterinary Hospital (NTUVH) from 2014 to 2017 in dogs and cats, analyzing the prevalence of ESBL-producing E. coli, the ESBL gene groups, and the sequence type of these ESBL producers. The double disc test specified by Clinical and Laboratory Standards Institute (CLSI) detected 65
ESBL-producing E. coli (22.8%) from 285 E. coli isolates recovered from dogs and cats in this study. All the 65 ESBL-producing E. coli were resistant to ampicillin. The
resistant rate to ceftiofur, doxycycline, enrofloxacin, and ciprofloxacin was 93.8%, 73.8%, 80%, and 78.5% respectively. The ESBL-producing E. coli were relatively more sensitive to imipenem and gentamicin. The CTX-M-1 and CTX-M-2 was the most
commonly found ESBL gene groups. A total of 38 isolates belonged to CTX-M-1 group;
the main ESBL gene was CTX-M-55 (24/65, 36.9%). Twelve isolates belonged to CTX-M-2 group and the ESBL gene were all CTX-M-124 (12/65, 18.5%). Multilocus sequence typing (MLST) showed that ST457, ST131 and ST648 were the most commonly found clones, and the ST131 and ST648 were the globally distributed high risk clones. The E. coli ST131/O25b is a zoonotic global clone of public health concern and our study found 8 E. coli ST131/O25b isolates whose major ESBL genes were CTX-M-14 and CTX-M-15 variants. Since companion animals like dogs and cats are in close contact with humans, From the perspective of public health and veterinary
medicine, characterization of the ESBL-producers and epidemiology is an issue warrants further investigation.
Keywords: Extended-spectrum β-lactamases, Escherichia coli, Multidrug-resistance, Multilocus sequence typing, ST131/O25b
目錄
中文摘要………...………i
英文摘要………iii
目錄………v
第一章 緒論………1
第二章 文獻回顧………3
第一節 大腸桿菌………3
2-1-1. 型態與背景介紹………3
2-1-2. 大腸桿菌型態與生化特徵………4
2-1-3. 大腸桿菌的血清型與毒素………4
第二節 大腸桿菌的抗藥性機制………6
2-2-1. 細胞膜通透性的改變………7
2-2-2. 藥物排出幫浦系統………7
2-2-3. 改變藥物的結合標的或流程………8
2-2-4. 產生酵素將藥物不活化………8
第三節 β-lactam 類抗生素……….9
2-3-1. 青黴素類………..10
2-3-2. 頭孢子菌素類………..11
2-3-3. 碳青黴烯類………..11
2-3-4. 單環內醯胺類………..11
第四節 超廣譜性乙內醯胺酶………..12
2-4-1. β-lactamaes 分類………..13
2-4-2. ESBLs 基因型………..15
2-4-3. ESBL 表現型的檢測………. 16
2-4-4. ESBL 基因型的檢測………..18
第五節 多重基因座序列分型法 (Multilocus sequence typing, MLST) …....18
第六節 ESBLs 流行病學相關研究………..19
2-6-1. ESBLs 在人醫的流行概況………..20
2-6-2. ESBLs 在獸醫的流行概況………..22
第七節 ST131/O25b 的流行概況……….………...23
第八節 研究目的………...25
第三章 材料與方法………...26
第一節 菌株來源………...26
第二節 ESBL 表現型鑑定……….…...26
第三節 萃取細菌 DNA (加熱煮沸法) ………....27
第四節 抗微生物藥物感受性試驗………...27
第五節 β-lactamaes 基因型測定………...28
第六節 MLST 序列定序分析………...29
第七節 PCR 檢測 ST131 O25b 之測試………...29
第四章 研究結果………...31
第一節 ESBL 菌株分離率與採樣來源………...31
第二節 ESBL 菌株之抗微生物藥物感受性試驗結果………...31
第三節 β-lactamaes 基因型檢測結果………...32
第四節 ESBL-producing E. coli 之 MLST 分析結果………...33
第五節 ESBL-producing E. coli 之 ST131 O25b 之檢測結果…………...34
第五章 討論………...35
5-1. ESBL 菌株分離率與採樣來源………...35
5-2. ESBL 菌株之抗微生物藥物感受性試驗結果……….…...35
5-3. β-lactamase 基因型測定結果………...37
5-4. ESBL-producing E. coli 之 MLST 序列定序分析………...37
5-5. ESBL-producing E. coli ST131 O25b 之檢測結果………...39
第六章 結論………... ... .41
第七章 參考文獻………...43
表次 Table 1. The primers used for ESBL-producing E. coli bla genes...….…... .56
Table 2. The primers used for multilocus sequencing typing …...57
Table 3. The primers used for ST131 O25b detection.………... ...58
Table 4. The specimen of the ESBL-producing E. coli isolated from the dogs and the cats in NTUVH. ………...58
Table 5. The prevalence of the ESBL-producing E. coli isolated from the dogs and the cats in NTUVH………...59
Table 6. Antibiotic susceptibility of 65 ESBL-producing E. coli……...60
Table 7. The bla genes of the 65 ESBL-producing E. coli………...61
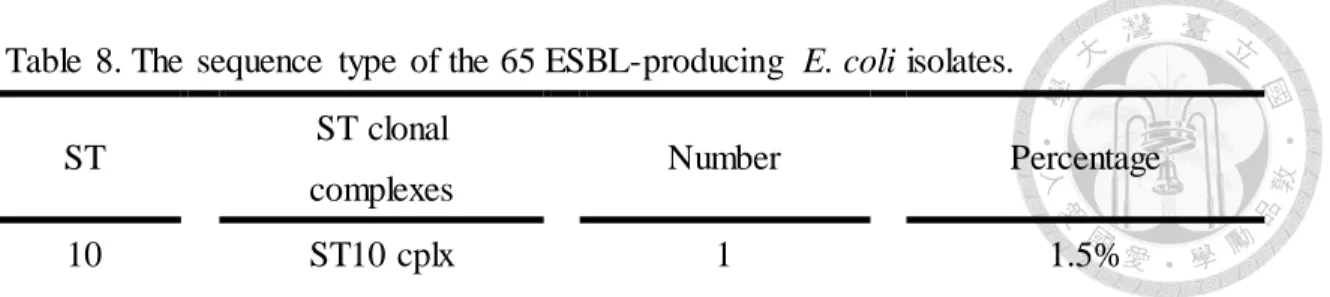
Table 8. The sequence type of the 65 ESBL-producing E. coli isolates... .. .62
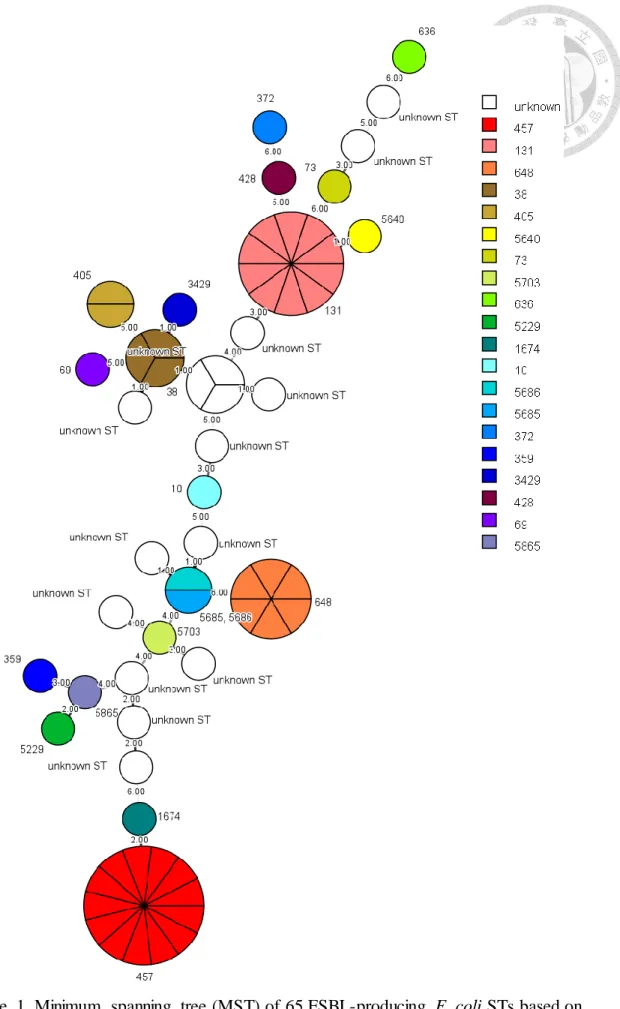
圖次 Figure 1. Minimum spanning tree (MST) of 65 ESBL-producing E. coli STs based on the degree of allele sharing. ………...63
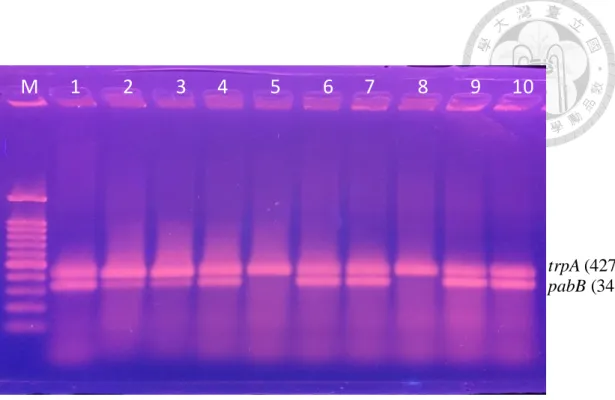
Figure 2. PCR detection for E. coli O25b-ST131 clone.…… ………...64
附錄 Appendix 1. MLST analysis of each ESBL-producing E. coli in 2014…....65
Appendix 2. MLST analysis of each ESBL-producing E. coli in 2015…....66
Appendix 3. MLST analysis of each ESBL-producing E. coli in 2016…....67 Appendix 4. MLST analysis of each ESBL-producing E. coli in 2017…....68
第一章 緒論
在 20 世紀 80 年代早期,第三代頭孢菌素開始引入臨床治療,被認為是抵抗 攜帶有β-lactamaes 抗藥細菌的重大突破,然而在 1983 年時,出現具有超廣譜性乙 內醯胺酶 (Eextened-spectrum β- lactamaes, ESBLs) 的腸內菌 (Enterobacteriaceae),
這些細菌不但可以對抗第三代頭孢菌素,且為多重抗藥性的菌株。雖然一開始僅 在歐美地區以及日本地區有零星分離報導,但現今以已遍佈全球,使得攜帶 ESBL 的細菌成為一個在公共衛生上非常重要的抗藥性議題 [2, 3]。然而具有 ESBL 的腸 內菌不只在人類發現,也可以在伴侶動物、經濟動物、野生動物、農作物和環境 中被分離出來 [4-9]。
大腸桿菌為腸道共生菌,若在腸道以外生長就有機會引起疾病,如在泌尿道中 引起尿路感染 (urinary tract infections, UTIs) [10]。然而犬貓中尿路感染為獸醫臨床 上常見的疾病,其病原菌又以大腸桿菌為主 [11],需要使用抗生素治療。因此若 是菌株對抗生素產生抗藥性,會導致治療上的失敗,也會增加醫療成本。且隨著 現今社會風氣的改變,犬貓不僅只是扮演寵物的角色,而更像是像自家孩子般,
飼主與犬貓居住一起且之間有著更親密的接觸,不管是飼主或是犬貓,若是帶有 ESBL-producing E. coli 就有可能造成抗藥性的傳遞,並且成為 ESBL-producing E.
coli 潛伏感染路徑的來源 [12, 13]。
在革蘭氏陰性病原體中,β-lactamase 的產生主要是導致細菌對於 β-lactam 類 抗生素產生抗藥性的原因。β-lactamase 主要是藉由水解 β-lactam 環,使 β-lactam 類的抗生素失去活性。而 ESBL 此種酵素則是由 β-lactamase 經由氨基酸的改變衍 生而來,擴增其所能作用的抗生素範圍,ESBL 能破壞所有青黴素類 (penicillins)、
第一代至第三代頭孢菌素 (cepahlosporins) 還有單環內醯胺類 (如:aztreonam) 的
抗生素,使其失去活性,增加臨床上治療的困難 [14]。而 ESBL 基因通常是位於 質體中,其中在插入序列 (insertion sequence, IS) 這類轉位子調節下,可使 ESBL 在各類細菌間相互傳播 [15]。由於攜帶 ESBL 的細菌為多重抗藥性菌株,導致人 醫和獸醫在使用抗生素治療時受到限制、導致增加治療失敗率以及發病率,也會 增加醫療費用 [16]。
已知細菌的抗藥性會隨著時間的推移而發生變化,因此,持續且長期的監控 對於研究細菌抗藥性的傳播是非常重要的 [11]。由於目前台灣對於帶有 ESBL 的 大腸桿菌在伴侶動物上的盛行率及特性分析尚未有全面性的研究報導,因此本研 究想先藉由研究臺大動物醫院中所收集到的犬貓檢體所分離出來的大腸桿菌,進 行 ESBL 的探討如 ESBL-producing E. coli 的盛行率、抗微生物藥物感受性試驗、
ESBL 基因型鑑定以及利用多重基因座序列分型法 (Multilocus sequence typing, MLST)分析各菌株之間的親緣性關係。了解在臺大動物醫院目前 ESBL-producing E.
coli 的流行概況,提供國內獸醫界一個參考。
第二章 文 獻回顧
第一節 大腸桿菌
2-1-1. 型態與背景介紹
大腸桿菌 (Escherichia coli) 在生物分類中屬於變形菌門 (Proteobacteria),γ- 變形菌綱 (Gammaproteobacteria),腸桿菌目 (Enterobacteriales),腸內菌科
(Enterobacteriaceae),埃希氏菌屬 (Escherichia)。為革蘭氏陰性菌 (Gram- negative) 兼性厭氧桿菌 (facultataive anaerobic bacillus),菌體大小約長 2-3 μm,寬 0.4-0.6 μm、
不產生芽孢,部分菌株會形成莢膜 (capsules) 或微莢膜 (microcapsules),具有線 毛 (pili) 及周鞭毛 (peritrichous flagella),具有運動性,觸媒反應呈陽性
(catalase-positive)、氧化酶試驗為陰性 (oxidase-negative),於 10-45℃均可以生長,
37℃為最適生長溫度 [1]。
大腸桿菌是由 Theodor Escherich 所發現,1884 年該學者研究腸道細菌對胃腸 道生理和病理的影響,且經由 Robert Koch 的學生 Wilhelm Frobenius 向 Theodor Escherich 介紹了細菌純培養和特性分析技術,使他致力於細菌學,並研究出一種 評估嬰兒糞便的新方法,在隔年 1885 年他公開演講論述在新生嬰兒的腸道所分離 出大腸桿菌的型態與特性,並命名為 Bacterium coli commune [17],直至 1945 年才 正式正名為 Escherichia coli [18]。
大部分的大腸桿菌為正常的腸道菌叢且無致病性,只有少數某些大腸桿菌會 引起下痢腹瀉等具有致病性,一般而言對宿主無害,且能合成人體所需及有益的 維生素 B12和 K2 [19, 20]。
2-1-2. 大腸桿菌型態與生化特性
大腸桿菌可以利用乳糖、葡萄糖等多數醣類,可以產酸及產氣。可以藉由培 養基或試管試驗檢測其生化特性,大腸桿菌生長在馬康基氏培養基 (MacConkey agar) 上可以清楚看見菌落呈粉紅色。在伊紅甲烯藍培養基 (Eosin methylene blue agar, EMB agar) 上可以看見菌落呈現帶有墨綠色金屬光澤。在呈色培養基 (CHROMT M agar ECC) 上可以觀察菌落呈現藍色。將大腸桿菌接種於三糖鐵培養 基 (Triple sugar iron agar, TSI agar) 時,底部 (butt) 和斜部 (slant) 均為黃色酸性 反應,且會產生氣體。也可以利用 IMViC test 進行鑑定結果如下:I:引垛 (indole) test 為會產生 indole,呈陽性反應,且不產生硫化氫。M:甲基紅 (methyl red) test 結果呈陽性。Vi:伏普 (Voges Proskauer) test 結果呈陰性。C:檸檬酸鹽利用 (citrate utilization) test 結果呈陰性,無法利用檸檬酸當做碳源 [1]。
2-1-3. 大腸桿菌的血清型與毒素
大腸桿菌可以依據 modified Kauffman scheme 將其抗原性分為三類:在細 胞壁上脂多醣 (LPS) 上的體抗原 (somatic antigen, O)、鞭毛上的鞭毛抗原 (flagella antigen, H) 、莢膜上的莢膜抗原(capsular antigen, K),目前已知的大腸桿 菌抗原至少有 170 種 O 抗原、50 種 H 抗原、100 種 K 抗原。血清型是由 O 抗原和 H 抗原特定組合所定義,某些血清型的大腸桿菌可能會引起相關之臨床症狀,但 原因並非是血清抗原所造成的毒性,例如 1983 年所發現的大腸桿菌 O157:H7 有三 種主要的致病毒力因子:志賀毒素 (Shiga toxins)、存在於細菌染色體上的 LEE 致 病基因群 (locus of enterocyte effacement pathogenicity island, LEE PAI) 以及質體 pO157 [21, 22]。
病原性大腸桿菌依據致病方式、臨床症狀、血清型別等特性,可以分為六種 亞群:
1. 腸毒性大腸桿菌(Enterotoxigenic E. coli, ETEC)
ETEC 會分泌兩種致病毒素,其分別為對熱不穩定腸毒素 (heat-labile enterotoxins, LT enterotoxins),包含 LT-I 和 LT-II,和對熱穩定腸毒素 (heat-stable enterotoxins, ST enterotoxins),包含 STa 和 STb。熱不穩定腸毒素與霍亂毒素有 80%的序列相似性,且兩者的作用位置也相同。會造成嚴重的腹瀉、腹痛、嘔 心以及脫水,成人和小孩均有可能會受到感染 [23]。
2. 附著性大腸桿菌 (enteroadherent E.coli, EAEC)
腸凝集性大腸桿菌 (Enteroaggregative E. coli, EAggEC) 為 EACE 的一種,
不分泌 LT 或是 ST,且不具有侵入性,但有吸附因子 (aggregative adherence fimbriae, AAF) 可附著於 HEp-2 細胞表面,產生凝集吸附現象 (aggregative adherence pattern)。EAEC 會附著在腸細胞之細胞膜上,損壞腸細胞並導致鄰近 微絨毛細胞喪失營養吸收之功能而引起腹瀉 [24]。
3. 腸侵入性大腸桿菌 (Enteroinvasive E. coli, EIEC)
EIEC 的致病機制為 EIEC 染色體和質體所攜帶的基因賦予細菌侵入結腸的 上皮細胞的能力。細菌進入結腸上皮細胞後,會在結腸上皮細胞內複製且擴散 到鄰近的細胞,引起發炎反應導致細胞被破壞,使糞便含血液、黏液和白血球,
此情況與腸出血性大腸桿菌不同,出血性大腸桿菌糞便也會帶血,但無白血球 [25]。
4. 腸致病性大腸桿菌 (Enteropathogenic E. coli, EPEC)
EPEC 具有附著因子 EPEC adherence factor (EAF)、bundle- forming pilus (BFP) 以及 intimate adherence (如 eae gene)。EPEC 會造成急性的腹瀉、發燒、
糞便帶有黏液等症狀,也是造成發展中國家嬰兒以及兒童腹瀉的主要原因,
EPEC 主要定植於近端小腸,黏附於上皮細胞形成菌落,這種黏附模式被稱為 局部黏附 (localized adherence, LA),無論是在人類或是動物受到感染時,在腸 道切片上可以看到黏附的細菌對腸道微絨毛細胞外型造成改變、細胞骨架受到 破壞,或又稱為 attaching and effacing (AE) lesions [26]。
5. 腸出血性大腸桿菌 (Enterohemorrhagic E. coli, EHEC)
又稱為佛羅細胞毒素大腸桿菌 (Verocytotoxin-producing E. coli, VTEC) 或是志賀毒素大腸桿菌 (Shiga toxin-producing E. coli, STEC)。EHEC 被認為黏 附在大腸以及遠端小腸段,其有最主要三個重要致病因子:Shiga-like toxin (Stx)、
attaching and effacing 基因 (eae gene),及含有溶血基因的質體 60-MDa pO157 (hemolysin plasmid),產生志賀毒素 Shiga-toxin 1 (Stx1) 和 Shiga-toxin 2 (Stx2) 作用於內皮細胞上,可導致溶血性尿毒症候群 (hemolytic uremic syndrome, HUS) 和出血性結腸炎 (hemorrhagic colitis, HC) 等臨床症狀 [27, 28]。
6. 擴黏性大腸桿菌(diffusely adherent E . coli, DAEC)
DAEC 與 EAggEC 不同的地方在於 HEp-2 細胞試驗中會產生廣泛性的吸附 特質,DAEC 表面具有線毛黏附因子 (fimbrial adhesion) F1845,利用
decay-accelerating factor (DAF, 是ㄧ種 cell-surface
glycosylphosphatidylinositol-anchored protein 用來保護細菌的細胞不被補體系統 破壞),DAEC 菌株誘導細胞病變,其特徵在於腸細胞如指狀突起,將其纏繞黏 附於細菌,通常是造成 1 歲以上幼兒的腹瀉感染 [29]。
第二節 大腸桿菌的抗藥性機制
大腸桿菌可以藉由不同的機制產生抗藥性,將其分為四類:
2-2-1. 細胞膜通透性的改變
通常抗生素要進入細菌體內時,須通過細胞外膜 (outer membrane, OM) 才能到 達所要標的的位置。革蘭氏陰性菌細胞外膜是由雙層脂質 (doubled-layer lipid) 和 孔蛋白 (porins) 組成,理論上疏水性抗生素如喹諾酮類 (quinolones) 和大環內酯 類 (macrolides) 是通過雙層脂質進入細菌體內,而親水性的抗生素如 β-內醯胺類 (β-lactams) 就藉由通過孔蛋白進入細菌體內。然而細胞外膜若是發生結構的改變 或者氨基酸的改變,就有可能影響細菌外膜對藥物的通透性進而產生抗藥性 [30]。
大腸桿菌有兩個外膜蛋白 (outer membrane protein, Omp)分別為:OmpF 和 OmpC,此兩者都是孔蛋白。他們作為穿過外膜的被動擴散孔,這兩個蛋白質的表 達受到滲透壓相互調控,在低滲透壓情況下 OmpF 會被優先製造出來,反之 OmpC 幾乎僅在高滲透壓被製造出來,OmpF 孔徑較大、通透性較高,而 OmpC 孔徑較 小、通透性較低,所以跨越外膜的被動擴散孔的大小對於大腸桿菌的調節有很重 要的關係,若是大腸桿菌減少 OmpF 並增加 OmpC 的表現,就有可能產生抗藥性,
若多種不同抗生素經由相同的 Omp 作為通道,這樣就有可能造成多重抗藥性 (multidrug resistance) 的出現 [31, 32]。
2-2-2. 藥物排出幫浦系統
排出幫浦 (Efflux pump) 的機制首次被發現是在 20 世紀 70 年代,在帶有抗四 環黴素 (tetracycline) 的抗藥性的大腸桿菌中所被發現 [30],細菌的藥物幫浦蛋白 通常分為五類,主要為 MFS 家族 (major facilitator super family)、ABC 家族 (ATP-binding cassette family)、RN D 家族 (resistance-nodulation-division family)、
SMR 家族 (small multidrug resistance family)、MATE 家族 (multidrug and toxic compound extrusion family)。除了 ABC 家族的幫浦蛋白是藉由水解 ATP 產生能量
差將藥物排出之外,其餘家族的幫浦蛋白需要有離子的催化例如氫離子和鈉離子,
才能產生能量差將藥物排出。Efflux 幫浦被認為是導致細菌具有多重抗藥性 (multi-drug resistance, MDR)的主要機制之一 [33]。
2-2-3. 改變藥物與標的物的結合或流程 (altered target or bypass)
細菌經由基因中的點突變或者因為抗藥性基因的轉移,改變抗生素的作用的 結合標的,使抗生素無法接合上去而失去作用,這改變對細菌的適應性影響最小,
例如合成的氟喹諾酮 (fluoroquinolone) 抗生素如環丙沙星 (ciprofloxacin) 目標為 拓撲異構酶 (topoisomerases),其在複製叉處鬆弛超螺旋 DNA 並在細菌 DNA 複製 期間對 daughter strain 進行脫離,導致原本環丙沙星抗生素可以作用的標的物現在 無法接合上去,而產生抗藥性 [34]。
質體介導的喹諾酮抗性 (plasmid- mediated quinolone resistance, PMDR),是一 個 target site 被保護很好的例子,一些蛋白質能夠保護旋轉酶 (gyrase) 免受喹諾酮 類 (quinolones) 藥物抑制,這些獨特的蛋白質進行編碼的等位基因,現在稱為 qnr,
並主要散佈於抗藥性質體 (resistant plasmids),大多伴隨著 ESBLs [30]。
2-2-4. 產生酵素將藥物不活化
細菌會分泌酵素,而這些酵素可以在抗生素上添加支鏈使抗生素無法進入菌 體中或導致無法與特定受器結合,也有可能破壞抗生素的部分鍵結,進而導致抗 生素的結構被破壞或失去活性,無法作用於菌體上。例如,細菌可以產生乙內醯 胺酶,而乙內醯胺酶可以與乙內醯胺酶環形成非共價鍵鍵結,再藉由醯基化 (acylation) 的作用將乙內醯胺環打斷,最後水解釋放出乙內醯胺酶及被去活化的乙 內醯胺抗生素結構,導致抗生素失效 [35]。由於此類抗藥性機制多存在於多重抗 藥性質體 (multidrug resistant plasmids) 上,因此容易在細菌之間傳遞,造成抗藥 性細菌的盛行。
抗藥性的產生除了上述 4 種抗藥性機制外,細菌還可以藉由其他方式獲得抗 藥性基因如 [33]:
1. Transformation:游離的 DNA 片段 (free DNA segment) 進入細菌菌體內與染色 體發生重組 (recombination),以獲得外源遺傳物質。
2. Transduction:噬菌體 (bacteriophages) 將帶有跳躍子 (transposable elements) 或 移動性遺傳因子 (mobile genetic elements) 的抗藥性基因以感染細菌的方式進 入菌體內。
3. Conjugation:細菌和細菌之間利用性線毛 (sex pili) 拉近彼此的距離,然後形 成接合管,傳遞帶有抗藥性的基因。
4. Mutation:細菌染色體因為突變產生抗藥性。
同種或者異種細菌之間可以經由質體或是抗藥性基因片段上的崁入序列 (insertion sequence, IS) 進行基因片段重組,使抗藥性基因在細菌間傳播。
第三節 β-lactam 類抗生素
β-lactam 類抗生素為最早被發現的抗生素種類,也是目前人醫以及獸醫在臨床 上用來對抗微生物主要用藥之一,所有β-lactam 類的抗生素皆是由乙內醯胺酶環 (β-lactam ring 或 cyclic amide) 作為最基本結構。由於環狀結構帶有兩個碳原子,
故稱為beta (β),此外在異環 (hetero ring) 結構部分分別帶有氮與羰基 (carbonyl moiety) [36]。
β-lactam 類的藥物作用機制主要是抑制細菌 peptidoglycan 的合成,使細菌無 法合成細胞壁,以達到殺菌的效果。因此β-lactam 類的抗生素抗菌效果與藥物對 細胞外膜的通透性和標的蛋白的親和性以及對抗細菌所分泌酵素的水解能力有關 [37]。β-lactam 類抗生素依照主結構的不同分為四大類:具有噻唑烷酮環
(thiazolidine ring) 的青黴素類 (penicillins)、具有 dihydrothiazine 的頭孢子菌素類 (cephalosporins)、碳青黴烯類 (carbapenems) 以及單環內醯胺類 (monobactams)。
2-3-1. 青黴素類 (penicillins)
青黴素類的藥物主要作用是阻礙細菌的細胞壁合成。盤尼西林結合蛋白 (penicillin binding proteins, PBPs)是 β- lactam 抗生素作用的目標,而 PBPs 的活性有 兩種 DD-transpeptidase 和 DD-carboxypeptidase,當 transpeptidase 被 β- lactam 作用 而失去活性時,將無法與胜肽醣類 (peptidoglycan) 結合至細胞壁上,導致細胞壁 無法合成。
該類藥物可以細分為六大類:第一和第二類的抗菌範圍相似,對於革蘭氏 陽性菌 、部分革蘭氏陰性菌和厭氧菌均有抗菌效果,第一類屬於原本就存在於自 然界的抗生素 (如:penicillin G, procaine penicillin 等)。第二類則是改良了第一代 青黴素對酸的不穩定之特性,發展成為口服藥物。第三類則是因為青黴素廣泛地 使用促使常見的金黃色葡萄球菌就產生青黴素酶 (penicillinase) 對既有 penicillin 產生抗藥性,因此第三類就是為了對抗金黃色葡萄球菌所產生青黴素酶
(penicillinase) 所研發,第三類也對鏈球菌具有抗菌活性,但對於其它具青黴素感 受性的菌種而言活性較低 (如:oxacillin、methicillin 等)。第四類屬於廣效型抗菌 範圍的青黴素,除了對原本有的抗菌能力外還增加了對抗革蘭氏陰性菌 (如:沙門 氏桿菌等),但目前細菌多為有抗藥性產生。第五類是為了對抗綠膿桿菌所研發的 青黴素 (如:carbenicillin、piperacillin 等),對鏈球菌以及厭氧菌等均有抗菌活性。
由於以上五類青黴素無法對抗乙內醯胺酶,會受到乙內醯胺酶影響而變得不穩定,
因此就有第六類青黴素產生,第六類是具有對乙內醯胺酶有抗性的青黴素 [38]。
2-3-2. 頭孢子菌素類 (cephalosporins)
頭孢子菌素類抗生素屬於弱酸類藥物,作用機制與青黴素類相似,依據抗菌範 圍分為四代,第一代頭孢子菌素主要用來革蘭氏陽性菌為主,常用的第一代頭孢 子素類包括 cephalothin、cefazolin、cephradine 及口服的 cephalexin 等,但對乙內 醯胺酶極為不穩定。第二代頭孢子菌素相對於第一代而言對乙內醯胺酶較穩定,
且比第一代多增加對抗革蘭氏陰性菌 (如:Bacteroides spp.等) 的活性。第三代則 是在多增加原第二代對抗革蘭氏陰性菌的效果,其中 ceftazidime 及 cefoperazone 可抗綠膿桿菌,但對革蘭氏陽性菌的活性卻比第一代差,與第二代相比差異最大 的是對乙內醯胺比較穩定且可穿透血腦障壁使得擴展適用範圍。第四代頭孢子菌 素類其抗菌範圍涵蓋了第一代抗革蘭氏陽性菌及第三代抗革蘭氏陰性菌的活性,
如:cefepime 及 cefpirome,該類藥效更強,而其對部分具有抗藥性的假單孢菌屬 細菌也有活性 [39]。
2-3-3. 碳青黴烯類 (carbapenems)
碳青黴烯類藥物的結構與青黴素類相似,但四氫噻唑 (thiazolidine) 中的硫被 甲基所取代,與肽酶 (peptidase) 作用進而抑制細菌細胞壁的合成。碳青黴烯類的 作用機制與青黴素類相同,但可以與更多的 PBPs 結合,因此藥物作用範圍比青黴 素類更廣泛。通常碳青黴烯類藥物被視為最後一道防線 (如:imipenem) 用於治療 嚴重的感染,對絕大部分的β-lactamase 具有活性 [40]。
2-3-4. 單環內醯胺類 (monobactams)
單環內醯胺類不會因對青黴素過敏而產生交叉過敏反應,因此對青黴素類過 敏的患者可以使用該類藥物治療。單環內醯胺類主要作用機制為抑制細菌細胞壁 的合成。常見的單環內醯胺類如:aztreonam,對革蘭氏陰性菌的 PBPs 親和性高適
合用來對抗革蘭氏陰性菌的抗生素,但不與革蘭氏陽性菌和厭氧菌的 PBPs 結合因 此不具有活性 [41]。
第四節 超廣譜性乙內醯胺酶 (extended-spectrum β-lactamase, ESBL)
β-lactamase 為細菌所分泌的一種酵素,可以與 β- lactam ring 形成非共價性鍵結,
再藉由醯基化 (acylation) 的作用將 β-lactam ring 破壞打開,最後水解 (hydrolysis) 釋出β-lactamase 及被去活化的 β-lactam 類抗生素結構。ESBL 主要是由革蘭氏陰 性菌所產生β-lactamase 的胺基酸序列經突變而來的。
β-lactamase 的分子結構大致相同,僅有一到四個胺基酸上的差異。而並非所 有的 TEM、SHV均為超廣譜性乙內醯胺酶 (extended-spectrum β- lactamase, ESBL),
如 TEM-1、TEM-2、TEM-13 以及 SHV-1 屬於原始的 β- lactamase,通常只對 penicillin、
第一代和第二代頭孢子菌素具有抗藥性,其抗藥性範圍較窄。若是β-lactamase 發 生胺基酸的突變 (mutation) 或者置換 (substitution),使原來的分子結構構型的改 變,變得更穩定,增加對第三代頭孢子菌素和單環內醯胺類等抗生素產生抗藥性,
提高了原始的β-lactamase 的抗藥性,將此類的 β-lactamase 稱為 ESBL。
ESBL 可以被 β- lactamase 的抑制劑所抑制,β- lactamase 的抑制劑是由 β- lactam 結構所衍生出來的藥物 (如:clavulanic acid、sulbactam 以及 tazobactam),具有 β-lactam ring 可以與 PBPs 結合或是與 β-lactamase 產生不可逆的反應。因此當 β-lactamase 抑制劑與 β-lactam 類抗生素一起使用時可以避免 β-lactam 類的抗生素 被β-lactamase 水解而失去活性,即使如此,細菌也產生出對應的生存之道,產生 過量的β-lactamase 或者改變外膜蛋白通透性等方式來抑制或者減少 β-lactamase 抑 制劑的活性而產生抗藥性。[42]。
2-4-1. β-lactamase 分類
目前對於β-lactamase 的分類主要可以分為兩大類:Bush Group 和 Ambler Class。
1. Bush Group
Bush 等人在 1989 年提出依據 β- lactamase 的功能性進行分類,但由於 TEM 和 SHV 衍生的 β- lactamase 數量增加,因此將其衍生物再加以分類,而分類的依據是 以 1989 年版本在新增子分類。分類方式是依據 β-lactamase 對抗不同 β- lactam 抗生 素的作用及受到 clavulanic acid 抑制程度可以分為四個 group 以及數個 subgroups [43]。
Group 1:以 cephalosporinase 且不被 clavulanic acid 抑制的為主,如 CMY、
ACT、DHA、FOX 和 MIR 等。在 Ambler 的分類中是屬於 Class C。
Group 2:以 cephalosporinase 或 penicillinases 且可被 clavulanic acid 抑制為主,
再依據對不同抗生素的水解能力再加以細分為 2a、2b、2be、2br、
2ber、2c、2d、2df、2e 和 2f 等 subgroups,其 TEM、SHV、OXA 和 CTX-M 衍生的 β-lactamase 屬於此類,TEM 和 SHV 所衍生的
β-lactamase 屬於 2b、2be、2br 以及 2ber,OXA 衍生的 β- lactamase 屬 於 2d、2de、2df,而 CTX-M 衍生的 β-lactamase 屬於 2be。在 Ambler 的分類中 Bush 2a、2b、2c、2e、2f 屬於 Class A,而 Bush 2d 屬於 Class D。
Group 3:含有鋅離子的酵素又稱金屬酵素 (metallo-enzyme),水解 penicillins、
cephalosporins 和 carbapenems,不被 clavulanic acid 抑制,但會被 EDTA、dipicolinic acid 或 1,10-o-phenanthroline 抑制。在 Ambler 的 分類中是屬於 Class B。
Group 4:不受 clavulanic acid 抑制的 penicillinases,目前 Ambler 對於該分類尚
未找到適合相對應的 class。
2. Ambler Class
Ambler 等人依據水解酶的氨基酸序列進行分類,將 β-lactamase 分為四個 classes:A、B、C、D。Classes A、C 和 D 是屬於 active-site serine-β-lactamases,
其β-lactamases 是通過 active site serine 形成 acyl enzyme 水解 β-lactam,而 class B 是屬於 metallo-β-lactamases,該類的 β- lactamases 是利用至少一個的 active-site 和 鋅離子催化反應促進β-lactam 水解,其中現在又以 class A 和 C 是為最常出現的 classes [44, 45]。
Class A:在 serine-β-lactamases 中佔了最大多數,具有水解 penicillin 的能力,
不論是在序列或是結構上皆與 PBPs 極為相
似,包含 TEM、SHV、PER/VEB/TLA-1 和 GES/IBC types
ceftazidimases、 SFO-1/BES-1 和 CTX-M types cefotaximases [46]。在 Class A 中最具代表性的 β-lactamase 為 TEM、SHV 和 CTX-M type。
Class B:屬於 metallo-β- lactamases (MBLs),需要二價離子的活化,通常以鋅離 子最為常見。IMP family、VIM family 等 carbapenemases 皆被歸類於 Class B β-lactamase 中。
Class C:屬於 cephalosporinases,多位於 Enterobacteriaceae 和一些其他生物的 染色體和質體上,第三代頭孢子菌素原本被使用來治療 β-lactamase- producing 的病原菌,但從 1980 年代開始,臨床上已經發現有可以對 抗第三代頭孢子菌素 (ceftazidime、cefotaxime、cefpodoxime 及 ceftriaxone) 和 monobactams (aztreonam) 的病原菌產生 [14]。
在 Class C 中其最具代表性的 β-lactamase 為 AmpC β-lactamase。
Class D:屬於 oxacillinase,主要是水解青黴素類的 oxacillin 和 cloxacillin,對
carbapenems 水解能力較差,在 Class D 中最具代表性的 β-lactamase 為 OXA-type β-lactamase。
2-4-2. ESBLs 基因型
ESBL 是在革蘭氏陰性菌中所發現,特別是在綠膿桿菌和腸內菌科,其中又以 Escherichia coli 和 Klebsiella pneumoniae 為最重要的兩種,在其他致病性細菌如 Salmonella、Shigella、Proteus、Enterobacter、Citrobacter 等細菌中也有發現 [47]。
ESBLs 的抗藥性基因大多位於質體上,藉由質體的複製與接合作用可以在不同細 菌間傳遞,也可能使原本不會產生β-lactamase 的腸道菌叢獲得抗藥性質體產生抗 藥性 [2]。
ESBLs 的相關基因型主要可以分為 TEM、SHV、CTX-M、OXA、PER、VEB、BES、
GES、TLA、SFO、IBC 等,其中又以 TEM、SHV、CTX-M 這三種為目前最常見 基因型 [2]。
1. TEM (Temoneria)
1965 年在希臘名為 Temoneria 的女性患者中,發現第一個存在於 E. coli 中藉由 質體傳播的β-lactamase,故就按照這名患者名字來命名為 TEM-1 [48],而後來 在 1985 年發現了 TEM-2,與 TEM-1 結構差異僅一個胺基酸不同,且對青黴素 類或者其衍生藥物具有水解的能力,TEM-1 和 TEM-2 在分類上為非 ESBLs [49]。
然而所有的 TEM type ESBLs 皆是 TEM-1 和 TEM-2 的衍生物,TEM-3 在 1984 年法國的 K. pneumoniae 分離株所發現,與 TEM-2 差了兩個胺基酸,但卻對 cefotaxime 產生抗藥性。目前約超過 200 個 TEM type 被發現,對於不同 TEM type 對於抗藥性程度也有所差異 [50]。
2. SHV (sulfhydryl variable)
SHV type 跟 TEM type 在結構上有許多相似之處,SHV-1 是在 1980 年代於 歐洲發現,對青黴素和第一代頭孢子菌素具有活性,目前的 SHV type ESBLs 皆為 SHV-1 衍生物,透過質體傳遞。其中大部份 SHV type ESBLs 胺基酸序列 位於第 238 位點會由原來的 glycine 取代為 serine,有的則是在第 240 位點從 glutamic acid 轉變為 lysine 或 arginine 時,促使對 ceftazidime 和 aztreonam 具有 水解能力。目前大約有 46 種的 SHV type 是屬於 ESBLs,有 150 種為非 ESBLs,
其中最常見非 ESBLs 的為 SHV-11 [51]。
3. CTX-M (Cefotaximase-Munich)
CTX-M type 被視為目前最重要的 ESBL β- lactamase 族群之一,屬於 Class A β-lactamase,過去研究指出帶有 ESBL 基因的細菌以 TEM type 和 SHV type 為 主,但自從 1989 年在德國慕尼黑 (Munich) 發現 CTX-M type 後,到 1995 年 就快速擴張至全球,ceftazidime 對 TEM 和 SHV 具有活性可以有效抑制攜帶 TEM、SHV 抗藥性基因的細菌,但對 CTX-M 不具活性,促使 CTX-M type 成 為近年來新興的 ESBL 基因原因之一,快速的分佈在世界各地。
在 CTX-M 的分類中,依據 Bonnet 在 2004 年用親緣分析 (phylogenetic analysis) 的方法將其分為五個 groups 分別為:CTX-M-1、CTX-M-2、CTX-M-8、
CTX-M-9 和 CTX-M-25,各 groups 之間約有大於 10%胺基酸差異,而各 groups 內每個 CTX-M type 彼此之間會以一個至數個不等的胺基酸差異 [52]。
2-4-3. ESBL 表現型的檢測
在 ESBL 表現型偵測中,不同實驗室使用不同的方法與參考的範圍可能 有所差異,在 ESBL 表現型偵測上目前比較常用的有雙紙錠協同測試法 (double disc synergy test, DDST)、最小抑菌濃度 (minimum inhibitory concentration,
MIC)、等電點聚焦 (isoelectric focusing, IEF) 和分子生物法 (例如:聚合酶連 鎖反應 polymerase chain reaction, PCR) 等等。再依據美國臨床與實驗室標準協 會 (Clinical and Laboratory Standards Institutes, CLSI) 準則,進行紙錠擴散 (disc diffusion) 方法分析,及 ESBL 表現型確認試驗 (phenotypic confirmatory test) [53]。
i 雙紙錠協同測試法 (double disc synergy test, DDST)
標準的 DDST 是以第三代頭孢子菌素紙錠與 β-lactamases 抑制劑 (如:
clavulanic acid) 進行測試,當第三代頭孢子菌素產生的抑制圈邊緣有明顯的往 抑制劑的方向延伸時,即為協同作用。但有些 ESBL 菌株同時帶有 AmpC
β-lactamases 不受 clavulanic acid 抑制且同時具有水解第三代頭孢子菌素的能力,
造成結果呈現偽陰性,使其表現型上造成誤判,故目前大多是使用改良的雙紙 錠協同試驗 (Modification of double-disk test, MDDST) 進行表現型分析,避免 偽陰性的產生。MDDST 除了使用第三代頭孢子菌素之外還要加上第四代頭孢 子菌素紙錠 (如:cefepime) 做檢測,當協同作用只作用於第四代頭孢子菌素時,
就表示該檢測菌株是 ESBL 和 AmpC β-lactamases 的 co-producer。
ii 等電點聚焦 (isoelectric focusing, IEF)
抽取待測菌株之 β-lactamases 進行電泳,電泳結束後再利用 nitrocefin 為受 質來呈色,並量測距離,再依據目前已知的β-lactamases 的 pI 值作為標準值,
畫出回歸曲線,將得到的結果與已知的β-lactamases 的 pI 值去做推論可能的 β-lactamases 種類。但因為蛋白質在進行電泳時,容易受到干擾造成位移或誤 差,因此 pI 值只能做為參考。
iii ESBL 表現型確認試驗 (phenotypic confirmatory test)
根據 CLSI 規範 [54],Klebsiella pneumoniae、Klebsiella oxytoca、
Escherichia coli、Proteus mirabilis 四種細菌可以用紙錠擴散擴散 (disc diffusion) 方法進行 ESBL 表現型確認試驗 (phenotypic confirmatory test)。此方法主要是 利用 ESBL 抑制劑 clavulanic acid 去抑制 β-lactamase 後,導致第三代頭孢子素 藥效增強,使抑菌圈增加的特性加以區別。
2-4-4. ESBL 基因型的檢測
在 ESBL 的基因型檢測中,較為常見的方法為聚合酶連鎖反應(polymerase chain reaction, PCR)、脈衝式電泳 (pulsed-field gel electrophoresis, PFGE) 與基 因定序 (DNA sequencing) 等方法。其中又以 PCR 為最常使用檢測 ESBL 基因 型別的方法 [55]。
依據 β-lactamase 基因序列 (如 blaT EM、blaSHV、blaCT X-M等) 設計相對應引 子 (primer),萃取待測菌株之核酸進行 PCR 片段增幅,檢測待測菌株是否帶有 ESBL 基因,再將所增幅之產物送定序,再將序列結果與 β-lactamase database (http://bldb.eu/BLDB.php?prot=A#CTX-M) [56]進行核酸序列比對,判定各基因 型別,除了 TEM-1、TEM-2、TEM-13、SHV-1 和 SHV-11 為非 ESBLs 之外,
其餘型別均為 ESBLs。
第五節 多重基因座序列分型法 (multilocus sequence typing, MLST)
MLST 是一種分子生物分析法,可以透過管家基因 (housekeeping gene) 核心 片段的 DNA 序列去分析比較該片段等位基因與各菌株之間的親緣性關係,而每個 細菌的管家基因不同,以 E. coli 為例,選取七個管家基因分別為:adk (adenylate kinase)、 fumC (fumarate hydratase)、gyrB (DNA gyrase)、icd
(isocitrate/isoprppylmalate)、mdh (malate dehydrogenase)、purA (adenylosuccinate
dehydrogenase)、recA (ATP/GTP binding motif),針對這七個管家基因片段進行 PCR 片段增幅與定序,至今已有 3,000 多的 Sepuence types (STs) 已建立於 MLST 資料 庫 (http://pubmlst.org/) 中。
早期定序還未盛行時,是使用 pulsed-field gel electrophoresis (PFGE) 分型法去 判別菌種間的親緣性關係,雖然這是最廣泛使用的細菌菌株分子分型工具,但該 方法存在一些缺點,例如:操作繁瑣屬於勞動密集型、難以標準化,且實驗室之 間的重現性有限 [16]。而 MLST 是一種相對較新的技術,其中針對多個管家基因 座進行測序以檢視親緣相關性並分析來自許多菌株的等位基因 (allele) 之間的序 列變異,可以提供更細微的序列上差異,使得更加容易去解釋菌株流行病學之間
的相關性,且不受限於各實驗室之間的相互比較,所以在有些情況下MLST 會比
PFGE 方法更具有鑑別力,不同的 STs 可以利用分子親緣關係畫出 minimum spanning tree (MST),用於長期監控與感染源追蹤上更加實用,不但能針對各國境 內各區菌株疫情的監控外,還可以應用於全球流行病學之研究 [57]。然而這些結 果並不能確切的表示人類和動物之間細菌的遺傳物質傳遞方向。
第六節 ESBLs 流行病學相關研究
ESBL 菌株一開始只在人類臨床醫療上觀察到,但近年來不論是動物還是環境 都陸續都有被分離出ESBL-producing E. coli,ESBLs 透過質體傳播的方式快速的
傳遞抗藥性基因,若是人類和動物腸道正常健康的腸道菌叢開始帶有ESBLs 抗藥
性基因而形成具有抗藥性的腸道菌,這樣在臨床上的治療勢必會增加治療上的困
難以及費用。依據不同國家所發表的文獻中可以得知,各基因型的ESBL 並不是
侷限在一個小地區,而是可以藉由宿主、交通、水源等環境因子將ESBLs 抗藥基
因散佈在世界各地 [58, 59]。
在 ESBLs 流行病學研究中顯示 1980 年代初期最常帶有 ESBL 的細菌是 K.
pneumoniae,而基因型又以 TEM 和 SHV 最常被發現,但在 1980-1990 年間,大多 數的ESBL-producing E. coli 和革蘭氏陰性菌中的 ESBL 都是 TEM group 或是 SHV group,在 1989 年 CTX-M 被發現後,盛行於 20 世紀 90 年代的 TEM 和 SHV type 逐漸已經被CTX-M type 所取代 [60, 61]
2-6-1. ESBLs 在人醫的流行概況
1. 北美地區
美國在 1988 年發現第一個 ESBL陽性分離株,為帶有 TEM-10 的 K. pneumoniae [62],之後又陸續發現 TEM-12、TEM-26 和 SHV-5。在美國的東南部社區醫院感 染ESBL-producing E. coli 數量從 2009 年的 17 例 (65%) 增加到 2014 年的 20 例 (77%),在女性患者中更為常見 54%,在泌尿道感染中分離出 61%為 ESBL-producing E. coli,且有不斷上升的趨勢 [63]。
加拿大主流的基因型與美國的不大相同,在加拿大中屬於基因型較為多樣性 可以分離出許多不同的 ESBL type,在 2000 年的調查中 K. pneumoniae 和 E. coli 的盛行率分別為13.7%和 6.3%,SHV 和 CTX-M 最為常見,其中 CTX-M-14 和一 些與CTX-M-14 較為相近的 ESBLs 曾在卡爾加里 (Calgary) 爆發 [14, 64]。
2. 南美洲
在拉丁美洲,產生 ESBL 的細菌在 K. pneumoniae 中佔 53%,在 E. coli 中佔 25%,智利也從原本盛行的 CTX-M-2 group 轉變為 CTX-M-1 group,這意味著傳播 機制的改變以及預防行動需要變化 [65]。
3. 歐洲
歐洲 ESBLs 的盛行率高過於北美洲,但比南美洲和亞洲低。在 1990 年代前大 多的 ESBLs 為 TEM 和 SHV 為主,其帶有病原體的細菌多和院內爆發有關,尤其
是在重症監護病房 (ICU),較少的社區感染的狀況,造成院內感染的菌株多為 K.
pneumoniae,但與現今結果發生變化,現在是以尿道感染的病患中所分離出來的 ESBL-producing E. coli 為最主要的分離株。
1983 年在德國檢測到第一個 ESBL,從重症監護病房住院患者中採樣不同腸桿 菌分離株中檢測到的,與 SHV-1 差一個氨基酸變化並將其命名為 SHV-2,1984 年 在法國的醫院在 K. pneumoniae 檢測到,先命名為 CTX-M-1,但後來經由鑑定之後 與 TEM-2 差了兩個氨基酸序列故將其正名為 TEM-3,德國在 1989 年在大腸桿菌 中發現第一個 CTX-M-1 group。
在保加利亞所分離到的 ESBLs 為歐洲比率最高高達 28%,塞浦路斯和羅馬尼 亞為 16%,葡萄牙為 12%。在 MYSTIC (Meropenem Yearly Susceptibility Test Information Collection) 監測計畫中,歐洲 ESBL 盛行率從 1997 年 2.1%到 2014 年 的 10.8%,可以看出 1997 年到 2014 年間產生 ESBL 的大腸桿菌有增加的趨勢 [66]。
4. 亞洲
在 2010 年-2013 年由 13 個位於亞太地區的國家或者城市,從腹腔感染所分離 到的革蘭氏陰性菌ESBL的盛行率,在中國 ESBL-E. coli盛行率為 66.6%位居第一,
其次為泰國ESBL-E. coli 盛行率為 49.8%,而臺灣在此研究中 ESBL-E. coli 盛行率 為12.8%,為亞太地區 ESBL-E. coli 盛行率倒數第三,其亞太地區 ESBL-E. coli 總 盛行率為38.2%,由全球性抗藥性監測計畫 SMART (Study for Monitoring
Antimicrobial Resistance Trends) 先前研究指出亞太地區相較於其他地區國家有特 別高的ESBL 分離率 [67]。
目前東南亞地區各國流行的基因型不一樣。中國在 2000 年後是以 CTX-M 的 E. coli ST131 為主,但在 2011-2012 年調查發現 ESBL-producing E. coli 在一般醫院
檢體分離率為42%,ST131 佔整體 14%且 CTX-M-1 (40.7%) 和 CTX-M-9 (48.7%) group 為最主要的 group [68]。
O25-ST131, CTX-M-15 在日本也有相關的報導,在日本近畿地區調查的 ESBL-producing E. coli 盛行率從 0.24%上升至 7.25%,以 CTX-M-9 group 為主要的 基因型 [69]。
5. 臺灣
經臺灣全國監測數據顯示,產 ESBL 的 大腸桿菌 的流行率 從 2002 - 2004 年 的4%增加到 2010 - 2012 年的 10.7% ,2007 年在 10 所主要教學醫院的重症監護病 房中 ,有 19.5%的 ESBL-producing E. coli 。而在台灣南部社區採樣兒童糞便中大 腸桿菌,其ESBL-producing E. coli 佔 8.3%且為 CTX-M-9 group 中的 CTX-M-14 (38.5%) 為最常見的基因型,O25b-ST131 是存在於兒童糞便中的最常見的產 ESBL 的大腸桿菌clone。在 2018 年的 SMART 中 ESBL-producing E. coli 佔 11.4%
[70-72]。
2-6-2. ESBLs 在獸醫的流行概況
自從 1988 年在日本由實驗犬隻糞便分離出帶有 ESBL 基因的大腸桿菌後,
ESBL 在伴侶動物和經濟動物中也開始受到重視 [73]。
1. 歐洲
葡萄牙在 2009 年的時候第一次報導在狗身上存在有 CTX-M-15, ST131 的 clone,Ewers 等人在 2010 年也去調查歐洲伴侶動物中 ESBL 的盛行率其結果發現 大約有 10% CTX-M-15, ST131 的 clone。其結果顯示在歐洲也是盛行 CTX-M-15, ST131 [12, 74]。
2. 美國
美國在 2010 年的時候首次在尿路感染相關的犬貓檢體中檢測出 ESBL 約佔 18%,而又以 CTX-M-1 group 中的 CTX-M-15 為最主要的基因型,其次為 SHV-12 和 CTX-M-14 [75]。
3. 中國
中國伴侶動物的檢體 ESBL-producing E. coli 檢測出來的比率高達 24.2%,其 中最常見的為 CTX-M-1 group 中的 CTX-M-15,其次為 CTX-M-123 和 CTX-M-1,
其主要流行的 ST type 為 ST131 和 ST10,也有檢測出 CTX-M-15, ST131 的 clone [76]。
4. 臺灣
臺灣在伴侶動物有關於 ESBL 盛行率的調查相較於其他國家是較為缺乏的,
於 2014 年回顧國立臺灣大學附設動物醫院在 2011 年至 2013 年間所採集之尿液樣 本進行 ESBL 的調查發現,其 ESBL-producing E. coli 的分離率約為 3.33% (2/60),
其 ESBL 基因型主要為 CTX-M (73.1%) 和 TEM (46.2%),多為 CTX-M-1 和 CTX-M-2 group,且菌株多為多重抗藥性菌株 (73.7%) [77]。
第七節 ST131 O25b 的流行概況
在人醫方面,目前導致產 ESBL 大腸桿菌的一個擴散關鍵因素是 clone 擴散,
例如高毒力 clone B2-O25:H4-ST131,其被認為是導致大流行病傳播的主要原因。
已經證實該 clone 已經出現和傳播於全球,但主要是感染人類造成尿路感染 (urinary tract infection, UTI) 和菌血症,而 CTX-M-15 在歐洲的醫院和社區環境中 均能發現其蹤跡,O25-ST131, CTX-M-15 這組 clone 相關的菌株已經出現在人類中 出現且容易被分離出來 [78]。
在獸醫方面,在 2009 年 Constança Pomba 等人發表一篇文獻,第一次指出 O25-ST131, CTX-M-15 在一隻 15 歲患有慢性膀胱炎的雌性犬中所分離出來 [74],
偶後就陸續有人發表在動物檢體中也有發現 O25-ST131, CTX-M-15 的蹤跡,但還 是無法證明該 clone 是由人傳遞給動物還是動物傳遞給人。
1. 歐洲:西班牙在 2004 年 ESBL-producing E.coli ST131 為 9%,到了 2006 年後 發現上升至 13%,也有其他研究在 2006-2008 年在西班牙的療養院病人
ESBL-producing E.coli ST131 為 22%,在法國 2001 年首次鑑定出 O25b-ST131,
並在 2006-2007 年 ESBL-producing E.coli ST131 為 25%。比利時在 2006-2007 年 ESBL-producing E.coli ST131 為 62%,在其他歐洲國家也是有發現該 clone 出現。
2. 美洲:加拿大在 2005-2007 年兩個尿路感染的分離株實驗中發現 ST131 的比率 為<3%和 1%,但在 2002-2004 年門診病患中發現 ST131 比率為 23%。在美國 芝加哥發現 ST131 的比率為 53%。
3. 亞洲:日本在 2002-2003 年發現 ST131 比率為 25%。在柬埔寨 2004-2005 年 ST131 clone 佔 ESBL-E. coli UTI 樣本中 27%。
4. 大洋洲:澳大利亞有兩項研究證實發現該國存在 ST131 其比率為 31%和 35%。
5. 非洲:非洲在 23 個樣本中有 43%且是表達 CTX-M-14 或 CTX-M-15 的 ST131 clone。在中非共和國中,產生 CTX-M-15 的大腸桿菌有 50%為 ST131。
由以上數據可以得知 O25b-ST131 是存在於世界各地的,而且發現 ST131 比例 不少 [79]。
第八節 研究目的
使用抗生素治療生病的伴侶動物是在獸醫臨床上常見的方法之一,目前台灣 對於 ESBL-producing E. coli 在伴侶動物犬貓的流行病學監控上尚未有全面性的研 究報導,因此本研究的目的在於分析 2014-2017 年間於臺大動物醫院中所收集到的 犬貓檢體中分離出來的大腸桿菌,進行 ESBL-producing E. coli 盛行率與基因型研 究,將所分離到的大腸桿菌菌株進行抗微生物藥物感受性試驗與多重基因座序列 分型法鑑定序列型別 (Sequence type, ST),以 BioNumerics Software version
(Applied Maths, Kortrijk, Belgium) 建構 minimum spanning tree (MST),了解各菌株 之間的親緣性關係,希望藉由本研究了解在臺大動物醫院目前 ESBL-producing E.
coli 的流行概況,提供國內獸醫界一個參考。
第三章 材 料與方法
第一節 菌株來源
本研究收集 2014 年至 2017 年間,從國立臺灣大學生物資源暨農學院附設動 物醫院看診犬貓檢體,凡藉由 Vitek 2 Compact (Biomérieux, Marcy-I’Etoile, France) 鑑定出的微生物菌種為大腸桿菌者,則加以收集,並使用商品化凍菌管
(MicrobankT M, Richmond Hill, Canada) 保存菌種凍存於-80 ˚C 冰箱,以作為後續實 驗所需。
第二節 ESBL 表現型鑑定
根據 CLSI 規範 [54],Klebsiella pneumoniae、Klebsiella oxytoca、Escherichia coli、
Proteus mirabilis 四種細菌可以用紙錠擴散擴散 (disc diffusion) 方法進行 ESBL 表 現型確認試驗 (phenotypic confirmatory test)。此方法主要是利用 ESBL 抑制劑 clavulanic acid 去抑制 β-lactamase 後,導致第三代頭孢子素藥效增強,使抑菌圈增 加的特性加以區別。本實驗所分離之 E. coli 會依據 CLSI 所規範進行 ESBL 表現 型鑑定分析。首先將待測菌株從-80 ˚C 冰箱中拿取出來,使用無菌接種針將凍菌保 存管中的玻璃珠取出一顆放至於 3ml 的 Mueller-Hinton broth (MHB) (BD Difco™, New York, USA) 中,以 37 ˚C 培養至 McFarland 值為 0.5 時,以無菌棉棒沾取菌 液於 Mueller-Hinton agar (MHA) (BD Difco™, New York, USA) 塗盤,再貼上四種 不同抗生素紙錠,分別為:Ceftazidime 30 μg (CAZ) (Mast group)、Ceftazidime 30 μg - Clavulanic acid 10 μg (CAZ/CA) (Mast group)、Cefotazime 30 μg (CTX) (Mast group)、Cefotazime 30 μg -Clavulanic acid 10 μg (CTX/CA) (Mast group),以 37 ˚C 培養 18-24 小時後判讀結果。
測量 CAZ/CA 與 CAZ 和 CTX/CA 與 CTX 的抑制圈差距,若 CAZ/CA-CAZ ≧ 5 mm 或 CTX/CA-CTX ≧ 5 mm 則判定為 ESBL 陽性菌株。
本實驗分別以 K. pneumoniae ATCC®700603 和 E. coli ATCC®25922 作為 ESBL 陽性與陰性對照組。
第三節 萃取細菌 DNA (加熱煮沸法)
以加熱煮沸法 (Boiling method) 進行 ESBL 陽性 E. coli DNA 萃取。先將保存 於凍菌管中的細菌以 37˚C 培養在 3 ml MHB 轉速 225 rpm 搖轉至隔天,取 1 ml 菌液,10,000 xg 離心 10 分鐘,倒掉上清液留下片狀沈澱物 (pellet),加入 0.5 ml 二 次去離子水 (distilled deionized water, ddH2O) 並震盪搖晃均勻,套上微量離心管防 爆蓋後放入沸水中加熱 10 分鐘,以 12,000 xg 離心 10 分鐘,取出含核酸之上清液,
保存於-20˚C 冰箱中,待後續 PCR 作為模板 (template) 使用。
第四節 抗微生物藥物敏感性試驗
根據 CLSI (2011 年) [80]動物源分離菌株抗微生物藥物敏感性試驗規範建議進 行紙錠瓊脂擴散試驗 (Disc agar diffusion method)。
將待測菌株從-80 ˚C 冰箱中拿取出來,使用無菌接種針將凍菌保存管中的玻璃 珠取出一顆放置 3ml 的 MHB 中以 37 ˚C 培養 McFarland 值為 0.5 時,以無菌棉棒 沾取菌液於 MHA 塗盤,再貼上藥物測試紙錠後以 37 ˚C 培養 18-24 小時後判讀結 果,依據抑制圈直徑大小對照 CLSI 所規範的標準去做判讀結果。
本實驗所使用之抗生素紙錠分別為 Amoxycillin/ clavulanic acid 20 μg/ 10 μg (Oxoid)、Ampicillin 10 μg (Oxoid)、 Imipenem 10 μg (Oxoid)、 Ceftiofur 30 μg (Oxoid)、 Doxycycline 30 μg (Oxoid)、 Enrfloxacin 5 μg (Oxoid)、 Ciprodloxacin 5 μg
(Oxoid)、 Gentamicin 10 μg (Oxoid)、 Sulfamethoxazole/ trimethoprim 23.75 μg/ 1.25 μg (Oxoid)。
第五節 β-lactamase 基因型測定
將萃取之細菌核酸針對 blaT EM、 blaSHV以及 blaCT X-M groups 包含:CTX-M-1、
CTX-M-2、 CTX-M-8、 CTX-M-9 和 CTX-M-25 基因群所設計之引子對進行 PCR 片段的擴增。其操作方法如下:
於 0.2 ml 微量管中加入 5 μl DNA template、25 μl 2X master Mix (
TIANGEN, Beijing, China) 以及 10 μM引子對各加入 1μl,在加入 18 μl ddH2O 使反 應總體積為50 μl,放入 PCR thermocycler (SensoQuest GmbH, Goettingen, Germany) 進行 PCR 反應,各引子序列見 Table 1。
PCR 反應條件:Pre-denaturation 95˚C、5 分鐘。 Denaturation 95˚C、30 秒。
Annealing 溫度依引子對而定 (Table1)、40 秒。 Extension 72˚C、1 分鐘,共 35 循環。 Final extension 72˚C、10 分鐘反應,最後產物保存於 4˚C。
PCR 反應結束後將 50 μl之 PCR 產物加入 0.7% (w/v) agarose gel,使用 Mupid-2 plus (含電源供應器) 以 100 伏特並於 0.5X TAE 緩衝液 (Omics Bio, Taipei, Taiwan) 中進行凝膠電泳25 分鐘,之後將膠體置入核酸安全染劑 (Omics Bio) 進行染色 25 分鐘,再將膠片置於紫外光透照相中 (Major Science, Saratoga, State of California) 確認DNA 片段擴增結果。
將預期產物片段切膠送至明欣生物科技,利用 ABI 3730xl 毛細管電泳 (capillary electrophoresis, CE) 自動定序儀和相對引子對進行定序,將序列結果與 β-lactamase database (http://bldb.eu/BLDB.php?prot=A#CTX-M) [56]進行核酸序列 比對,判定各基因型別。
第六節 MLST 序列定序分析
為了瞭解 ESBL 基因是透過自體複製傳播 (clonal spread) 或垂直傳播
(horizontal gene) 的方式傳遞,將具有 ESBL 特徵的 E. coli 菌株以 MLST 的方式進 行序列分析,依據Wirth 等人 [81] 將 E. coli 七段管家基因 (housekeeping gene):
adk (adenylate kinase)、 fumC (fumarate hydratase)、gyrB (DNA gyrase)、icd (isocitrate/isoprppylmalate)、mdh (malate dehydrogenase)、purA (adenylosuccinate dehydrogenase)、recA (ATP/GTP binding motif) 進行 PCR 片段擴增與定序,引子序 列見Table 2。其操作方法如下:
於 0.2 ml 微量管中加入 5 μl DNA template、25 μl 2X master Mix 以及 10 μM引 子對各加入1 μl,在加入 18 μl ddH2O 使反應總體積為 50 μl,放入 PCR thermocycler 進行 PCR 反應,各引子序列見 Table 2。
PCR 反應條件:Pre-denaturation 95˚C、5 分鐘。 Denaturation 95˚C、30 秒。
Annealing 溫度依引子對而定 (Table1)、40 秒。 Extension 72˚C、1 分鐘,共 35 循環。 Final extension 72˚C、10 分鐘反應,最後產物保存於 4˚C。
其各管家基因座定序結果與 MLST 資料庫 (https://pubmlst.org/) 做比對,最終 的Sequence types (STs) 以 BioNumerics Software version (Applied Maths, Kortrijk, Belgium) 建構 ESBL-producing E. coli minimum spanning tree 親緣樹狀圖。
第七節 PCR 檢測 ST131 O25b 之測試
依據 Clermont 等人 [82] 所提供之方法,將萃取之細菌核酸進行針對 ST131 O25b 所設計之引子對 (詳見 Table1) 包含:trpA 和 pabB 進行 PCR 片段擴增。操 作方法如下:
於 0.2 ml 微量管中加入 5 μl DNA template、25 μl 2X master Mix 以及 10 μ M引子對 (trpA 和 pabB) 各加入 1μl,在加入 16 μl ddH2O 使反應總體積為 50 μl,
放入 PCR thermocycler 進行 PCR 反應,各引子序列見 Table 3。
PCR 反應條件:Pre-denaturation 94˚C、4 分鐘。 Denaturation 94˚C、5 秒。
Annealing 65 ˚C、10 秒,共 30 循環。Final extension 72˚C、5 分鐘反應,最後產物 保存於4˚C。
第四章 研 究結果
第一節 ESBL 菌株分離率與採樣來源
本研究於 2014 年至 2017 年間共收集到 285 株經由 Vitek 2 Compact 所鑑定出 的微生物菌種為大腸桿菌之犬貓檢體,總計分離出 65 株 ESBL-producing E. coli,
其株採樣來源以尿液 (urine) 為主(44/65, 67.7%),其次為傷口 (5/65, 7.7%) 和膿 (pus) (4/65, 6.2%) 等等。(詳見 Table 4)
ESBL-producing E. coli 整體盛行率為 22.8% (65/285),若依據 2014-2017 年各 年份分離率如下:18.2% (22/121)、21.1% (15/71)、34.6% (18/52)、25.6% (10/38),
若是再細分的話,2014-2017 年犬隻 ESBL-producing E. coli 的總盛行率為 23.9%
(54/226),貓 ESBL-producing E. coli 的總盛行率為 18.6% (11/59)。在 2014 年犬、
貓盛行率分別為81.8% (18/22) 和 18.2% (4/22)。在 2015 年犬、貓盛行率分別為 93.3% (14/15)和 6.7% (1/15)。在 2016 年犬、貓盛行率分別為 83.3% (15/18) 和 16.7%
(3/18)。在 2017 年犬、貓盛行率分別為 70% (7/10) 和 30% (3/10)。(詳見 Table 5)
第二節 ESBL 菌株之抗微生物藥物感受性試驗結果
本研究使用臨床上常使用的 9 種抗微生物藥物進行實驗,使用 β-lactams 類:
amoxycillin/clavulanic acid、ampicillin、imipenem、ceftiofur,四環素類 (tetracyclines):
doxycycline,喹諾酮類(quinolones): enrofloxacin、ciprofloxacin,胺基配糖體類 (aminoglycosides): gentamicin,磺胺類 (sulfonamides):
sulfamethoxazole/trimethoprim。由實驗結果得知,分離出 65 株 ESBL-producing E.
coli 對於 β-lactams 類的抗生素中的 ampicillin 皆具抗藥性。而對 β-lactams 類的 ceftiofur (93.8%)、四環素類的 doxycycline (73.8%)以及喹諾酮類的 enrofloxacin
(80%) 、ciprofloxacin (78.5%),則有很高比率呈現抗藥性。然而,ESBL-producing E. coli 對於 β- lactams 類的 imipenem (96.9%) 與胺基配糖體類的 gentamicin (61.5%),
則有較高的感受性。在β-lactams 類的 amoxycillin/clavulanic acid,則呈現抗藥性 (38.5%) 與感受性 (38.5%) 比率為一樣的結果。在磺胺類中的
sulfamethoxazole/trimethoprim 的抗藥性 (50.8%) 微略高於感受性 (43.1%) 。 以犬貓做區分統計,在貓隻中,ESBL-producing E. coli 對
amoxycillin/clavulanic acid、ampicillin、ceftiofur、doxycycline、enrofloxacin、
ciprofloxacin 以及 sulfamethoxazole/trimethoprim 呈現抗藥性比率較高,而對
imipenem、gentamicin 則呈現較高比率的感受性。在犬隻中,ESBL-producing E. coli 對 ampicillin、ceftiofur、doxycycline、enrofloxacin、 ciprofloxacin 以及
sulfamethoxazole/trimethoprim 有較高的抗藥性比率,對 amoxycillin/clavulanic acid 抗藥性與敏感性比率各佔一半,而對 imipenem、gentamicin 則呈現較高比率的感 受性。 (詳見 Table6)
第三節 β-lactamase 基因型測定結果
本實驗所有 65 株 ESBL-producing E. coli 針對 TEM、SHV、CTX-M 這三種常 見 ESBL bla 基因型進行 PCR 檢測,將待測樣本送定序後,結果與軟體做比對,依 據定序結果判讀是屬於哪個 genotype。
由實驗結果得知,在 65 株分離株中,CTX-M-1 group 為最主要的 ESBL bla 基因型,其次分別為 CTX-M-9 group、TEM group、CTX-M-2 group、SHV group,
CTX-M-8 group 與 CTX-M-25group 則無發現。其 CTX-M-1 group 中的 CTX-M-55 和 CTX-M-9 group 中的 CTX-M-174、CTX-M-198 以及 TEM group 中的 TEM-215,
在 2014 年到 2017 年間經定序比對結果均有被發現。屬於 CTX-M-1 group 的分離