國立臺灣大學生命科學院微生物與生化學研究所 碩士論文
Institute of Microbiology and Biochemistry College of Life Science
National Taiwan University Master Thesis
以農桿菌媒介轉形法作為鮑魚菇異源表達之工具
Agrobacterium tumefaciens mediated transformation as a tool for heterologous expression in Pleurotus ostreatus
蔡媛禎 Yuan-Chen Tsai
指導教授:黃慶璨 博士 Advisor: Ching-Tsan Huang, Ph.D.
中華民國 98 年 7 月
July, 2009
I
謝誌
盼著盼著總算有寫謝誌的一天,首先最要感謝的是我的指導教授 黃慶璨老 師,總是不厭其煩地教誨我們做事與學習上該有的態度與思考方式,要我們將嚴 謹時時放在心中,也一直耐心鼓勵、包容我們;研究生涯期間同時承蒙 許瑞祥老 師指導,在菇蕈專業知識與實驗上給予許多寶貴意見,並且引領我們思考不同的 方向與觀念,在此對兩位老師致上最深的謝意與敬意。口試期間承蒙生化組 楊健 志老師、中正大學 陳浩仁老師以及中研院 常怡雍老師,在論文內容的細心審閱 與口試時的觀念引導,給予我論文寫作與科學方法修正上很大的幫助,也讓本論 文更臻完善,特此感謝!特別感謝中山大學 劉仲康老師在我大學期間給予我的指 導,以及研究所入學期間給予我的幫助,讓我得以進入微生所微生物組深造。
兩年在 106 的研究生活,時常受到 106 夥伴的照顧。感謝菇類轉殖大師父俊 毅在我最一開始進行研究時的協助與鼓勵,由於你先前的努力,才使得我有許多 材料與方向得以進行本研究。感謝琬婷與王董在實驗上給予的許多建議與教導,
你們總時詳細回答我實驗上的問題,也是使實驗室得以順利運作的推手,是實驗 室的精神榜樣。謝謝我的師父韻涵,是因為妳先前的研究結果使得我的題目能夠 誕生,在我初入實驗室時妳的親切讓我記憶猶新,妳的耐心指導,也讓我逐漸對 實驗上手。感謝學長昌廷時常協助我解決電腦上的疑難雜症,也提供寶貴的實驗 經驗給我。感謝學姐林錚,常常能夠從妳那裏學到不同領域的知識,也時常受到 妳的照顧與關心。感謝學長姐明玥、秀欣、家禎、千椀、境芳、敏慈、孟男,你 們總會熱心的分享實驗心得,謝謝你們給予我的指導與精神鼓勵。一起走過兩年 歲月的佳珮和佩璇,是我生活與實驗上的最佳夥伴,常常跟妳們討論、一同解決、
分享實驗上遇到的問題與瓶頸,彼此的鼓勵與打氣,是我在實驗遇到困境時,得 以堅持下去的動力之一。感謝升碩二的映慈、明毅,從軍樂的信豪,升碩一的翊 倫、瑞芸,因為你們的加入讓106 更熱鬧也更完整。謝謝所有 106 夥伴為實驗室 的付出,也祝福你們未來的路途順利朝自己的理想前進。
感謝大學同學芳寶、歐姐、映妃、珮寧、孝慈、旭祺、恆毅,高中同學乃嘉、
維新、安妮、小玉,國中同學尹岑和小學同學阿瑄,因為你們也在不同的地方、
不同的領域奮鬥著,難得的聯絡、聚會與分享,都是讓我努力學習、成長的動力。
特別感謝初到台北在聖本篤認識的郁萍、馥瑋、亭靜、盈涵等等,還有修女以及 許多姐姐妹妹們,是妳們給了我彷彿親人的溫暖讓我逐漸習慣在台北的生活。感 謝室友伯爵,每天回到宿舍跟妳聊天常是一天難得的愉快時光,謝謝妳陪我度過 碩二無數個煩惱與鬱悶的日子。希望你們所有人都能走出自己的一片天。也感謝 拉蒙卡哈,在我碩一的瓶頸期,是靠著你對年輕學者的一番鼓勵,使我不放棄。
最要感謝的,是我的家人。感謝我的父親 蔡精文先生和母親 簡靜蘭小姐含 辛茹苦拉拔我二十四個寒暑,總是掛心我的生活與健康,在台北的兩年日子他們 時常體貼的寄上許多物資與營養補給品給我,分享他們看到的資訊給我,也總是 給我許多可靠寶貴的意見;感謝大姊涵禎與二姊淳禎在我前面披荊斬棘、擋風擋 雨,讓我沒有經濟上的顧慮而能求學至今。無盡的感激難以訴諸筆墨,謝謝家人 無私地給予我精神與物質上的一切,你們永遠是我的驕傲與榜樣。
最後,僅以此論文獻給所有曾經關心、幫助過我的親友們,也祝福你們。
2009.8.8 媛禎 謹致
II
中文摘要
鮑魚菇在食用菇中的產量僅次於洋菇,其栽培容易且可利用多種農業廢棄材 做為基質,受到各國食用菇農業的青睞;建立穩定的鮑魚菇轉形方法可做為發展 鮑魚菇作為異源表達系統之工具,進而應用至食用疫苗的開發。
農桿腫瘤菌被譽為「自然的基因工程師」,農桿腫瘤菌媒介之轉形法相較其他 轉形方法有操作便利、耗費較低等優點。轉入的異源基因多以單一嵌入位嵌入宿 主染色體內,因此能穩定表現。在本研究中,我們成功建立了鮑魚菇以農桿菌媒 介之轉形系統,將質體帶有不同啟動子啟動報導基因綠色螢光蛋白質(Green f l u o r e s c e n t p r o t e i n , G F P ) 與 篩 選 標 記 潮 黴 素 磷 酸 轉 移 酶 ( H y g r o m y c i n phosphotransferase, hph)之農桿菌和鮑魚菇共培養之測試結果,鮑魚菇以蕈褶組織 做為轉形材料與農桿菌株(AGL-1、LBA4404)共培養能夠得到較多轉形株,約 80~90%的轉形效率,而均質菌絲液為轉形材料得到較少轉形株,以菌絲塊則無法 得到轉形株。螢光顯微鏡結果顯示,花椰菜鑲嵌病毒35S 啟動子(CaMV 35S)與洋 菇的甘油醛-3-磷酸脫氫酶 (glyceraldehyde-3-phosphate dehydrogenase, GPD) 啟動 子啟動之綠色螢光蛋白質傾向呈現點狀螢光,類似聚集體(aggresome)的情形;當 鮑魚菇 GPD 啟動子啟動 C 端融合內質網停留訊息胜肽的 GFP 時,亦有點狀綠色 螢光分佈菌絲內。而鮑魚菇、金針菇 GPD 啟動子啟動之 GFP 則在菌絲內呈現分 散的螢光。推測融合訊息胜肽可能有助於鮑魚菇內GFP 的累積而導致 GFP 聚集。
以三明治法酵素連結免疫吸附分析轉形株內可溶性 GFP,每克總可溶性蛋白質含
量最高為43.83 ng。此外,異源基因在轉形株內於有絲分裂與減數分裂後皆能穩定 表現。
關鍵字 : 鮑魚菇、農桿菌媒介轉形、甘油醛-3-磷酸脫氫酶啟動子、綠色螢光蛋白 質
III
Abstract
Pleurotus ostreatus is the second largest edible mushrooms next to Agaricus bisporus. P. ostreatus is popular worldwide due to ease of and use of agricultural wastes as substrates. Establishing a stable transformation system of P. ostreatus may provide a tool for its heterologous expression and even oral vaccine production in the future.
Agrobacterium tumefaciens has been known as “natural engineer”. There are many advantages of Agrobacterium tumefaciens mediated transformation (ATMT) over other transformation methods such as convenience and less expenssive. The transformed heterologous gene can be integrated into host genome by mostly single insertion, and consequently more stable.
In this study, we successfully developed the A.tumefaciens mediated transformation of Pleurotus ostreatus. Agrobacterium harboring plasmid with different promoters driving green fluorescent protein (GFP) and hygromycin phosphotransferase gene (hph) was co-cultivated with Pleurotus ostreatus. P. ostreatus gill tissues used as explants co-cultivating with A.tumefaciens AGL-1 and LBA4404 could get more transformants, with transformation efficiency around 80 ~ 90%. However, only a few of transformants were obtained when homogenized mycelia was used as starting material, and no transfomant was found using mycelia modified pellets (MMP). Fluorescent microscopic observation revealed that GFP can be driven by the califlower mosaic virus (CaMV) 35S promoter and Agaricus bisporus glyceraldehydes-3-phosphate dehydrogenase (GPD) promoter, in which GFP tended to appear as aggresome-like fluorescent spot. When GFP gene with C terminal endoplasmic reticulum retention signal was driven by P. ostreatus homologous GPD promoters, the GFP also formed green fluorescent spots. In the other hand, the Flammulina velutipes GPD and P.
ostreatus homologous GPD promoters without retention signal showed scattered green fluorescence. These results suggested that fused signal peptide led to GFP accumulation in P. ostreatu as aggregates. The highest amount of soluble GFP extracted from P.
ostreatus detected by ELISA is 43.83 ng per gram total soluble protein. Besides, heterologous gene could be stably expressed after mitosis and meiosis.
Keywords : Pleurotus ostreatus, Agrobacterium tumefaciens mediated transformation (ATMT), glyceraldehydes-3-phosphate dehydrogenase (GPD) promoter, green
fluorescent protein (GFP)
IV
目錄
謝誌 ... I 中文摘要 ... II Abstract ... III 目錄 ... IV 圖目錄 ... VI 表目錄 ... VIII
第一章 前言 ... 1
一、基因工程 ... 1
1. 基礎生物研究 ... 1
2. 分子育種 ... 1
3. 生產異源蛋白質 ... 1
二、異源表達系統 ... 2
1. 原核表達系統 ... 2
2. 真核表達系統 ... 3
三、鮑魚菇概述 ... 6
1. 形態特徵、分類地位、生活史 ... 6
2. 栽培優勢 ... 8
3. 藥理活性 ... 8
4. 其他應用價值 ... 8
四、食用菇類轉形系統 ... 10
五、農桿菌媒介轉形 ... 13
1. 農桿腫瘤菌 ... 13
2. 農桿腫瘤菌的感染過程 ... 15
3. 農桿腫瘤菌在基因轉殖的應用 ... 20
六、分生策略 ... 22
1.啟動子 ... 22
2.報導基因 ... 22
3.內含子與訊息胜肽 ... 23
4.篩選標記 ... 23
七、研究動機與目的 ... 24
第二章 材料與方法 ... 27
一、實驗材料 ... 27
1. 菌株及培養條件 ... 27
V
2. 質體 ... 28
3. 引子 ... 29
二、實驗方法 ... 36
1. 重組質體建構 ... 36
2. 農桿菌轉形 ... 42
3. 農桿菌媒介之鮑魚菇轉形 ... 43
4. 鮑魚菇轉形株篩選與分析 ... 46
第三章 結果 ... 49
一、質體建構 ... 49
1.重組質體確認 ... 49
二、農桿菌媒介轉形 ... 56
1. 重組質體轉入農桿菌株 ... 56
2. 農桿菌媒介之鮑魚菇轉形 ... 56
三、綠色螢光蛋白質於鮑魚菇之表現 ... 63
1. 螢光顯微鏡觀察 ... 63
2. 西方式雜合分析 ... 67
3. 三明治法酵素連結免疫吸附分析結果 ... 68
四、鮑魚菇轉形株異源基因穩定度 ... 70
1. 無性世代穩定性 ... 70
2. 有性世代穩定性 ... 70
第四章 討論 ... 73
一、鮑魚菇以農桿菌媒介之轉形系統建立 ... 73
1. 宿主材料之選擇 ... 73
2. 農桿菌株 ... 75
3. 誘導物濃度 ... 75
4. 共培養時間 ... 75
二、發展鮑魚菇作為異源表達系統 ... 77
1. 啟動子探討 ... 77
2. 訊息胜肽效應 ... 80
第五章、結論 ... 81
第六章、未來工作 ... 82
第七章、參考文獻 ... 83
VI
圖目錄
圖一 鮑魚菇之形態 ... 7
圖二 擔子菌灰蓋鬼傘之生活史與子實體發展過程 ... 7
圖三 VirA受環境因子以及AS誘導活化後之訊息傳導示意圖 ... 16
圖四 誘導農桿腫瘤菌 vir 基因表現的物質結構式 ... 16
圖五 農桿菌腫瘤引發質體與T-DNA移動及嵌入宿主染色體示意圖 ... 19
圖六 農桿腫瘤菌感染植物造成之T-DNA轉形過程示意圖 ... 19
圖七 本論文研究架構圖 ... 26
圖八 實驗室無菌栽培之鮑魚菇子實體。 ... 30
圖九 質體 pCAMBIA0380 與pCAMBIA 1302 之限制酶切位與特定基因圖譜 ... 31
圖九(續) 質體 pCAMBIA0380-PiH與pPiH-PiEGFP/SP之限制酶切位與特定基因圖 譜 ... 32
圖九(續) 質體 pCAMBIA0390-AH-AiEGFP 與 pCAMBIA0390-AH-FiEGFP 之限 制酶切位與特定基因圖譜 ... 33
圖十 pCAMBIA0380-PiH重組質體建構示意圖 ... 39
圖十(續) pCAMBIA0380-PiH-PiEGFP重組質體建構示意圖 ... 40
圖十(續) pCAMBIA0380-PiH-PiEGFP-SP重組質體建構示意圖 ... 41
圖十一 宿主轉形材料與農桿菌共培養操作示意圖 ... 45
圖十二 pCAMBIA0380-PiH Colony PCR產物之洋菜膠體電泳結果 ... 50
圖十三 pCAMBIA0380-PiH 限制酶截切確認結果 ... 51
圖十四 鮑魚菇GPD啟動子序列 ... 52
圖十四(續) 潮黴素磷酸轉移酶核苷酸序列 ... 53
圖十五pPiH-PiEGFP colony PCR與pPiH-PiEGFP-SP colony PCR 結果 ... 54
圖十六 pPiH-PiEGFP 與pPiH -PiEGFP-SP限制酶切結果 ... 54
圖十七 EGFP與EGFP-SP核苷酸序列 ... 55
圖十八 農桿菌colony PCR產物之洋菜膠體電泳結果 ... 58
圖十九 鮑魚菇MMP轉形株生長情形 ... 60
圖二十 以鮑魚菇雙核菌絲液作為轉形材料於選擇性培養基上初次生長情形 ... 61
VII
圖二十一 轉形株MLF-2 於選擇性培養基上生長情形良好 ... 61
圖二十二 以鮑魚菇蕈褶組織作為轉形材料於選擇性培養基上初次生長情形 ... 62
圖二十三 鮑魚菇蕈褶組織轉形株繼代生長情形 ... 62
圖二十四 鮑魚菇辨識CaMV 35S啟動子所表現之mGFP ... 64
圖二十五 鮑魚菇辨識同源性GPD啟動子所表現之EGFP ... 64
圖二十六 鮑魚菇辨識同源性GPD啟動子所表現之EGFP-SP ... 65
圖二十七 鮑魚菇辨識洋菇GPD啟動子所表現之EGFP ... 65
圖二十八 鮑魚菇辨識金針菇GPD啟動子所表現之EGFP ... 66
圖二十九 螢光顯微鏡觀察野生型鮑魚菇菌絲 ... 66
圖三十 鮑魚菇轉形株粗萃取蛋白質以西方式雜合分析定性結果 ... 67
圖三十一 轉形株菌絲生長速度不均之情形 ... 68
圖三十二 鮑魚菇轉形株蛋白質粗萃液之三明治法酵素連結免疫吸附分析結果 .. 69
圖三十三 鮑魚菇轉形株抗藥性基因穩定度測試 ... 71
圖三十四 螢光顯微觀察鮑魚菇轉形株液態培養產生之節生孢子萌發菌絲 ... 71
圖三十五 螢光解剖顯微鏡觀察鮑魚菇蕈褶組織 ... 72
圖三十六 螢光顯微鏡觀察鮑魚菇轉形株與野生型擔孢子 ... 72
VIII
表目錄
表一 不同表達系統生產人類醫藥用重組蛋白質之比較 ... 5
表二 鮑魚菇的藥理成分與功用 ... 9
表三 鮑魚菇轉形系統之發展 ... 11
表四 成功以農桿菌轉形之食用菇類 ... 12
表五 農桿腫瘤菌與其腫瘤引發病症於植物之部分研究 ... 14
表六 本研究使用之核酸引子序列 ... 34
表七 本研究所使用之菌株 ... 35
表八 限制酶反應之酵素比例與反應條件 ... 36
表九 聚合酶連鎖反應之條件 ... 38
表十 共培養使用之誘導培養基 (induction medium, IM)組成成分 ... 44
表十一 農桿菌株對應質體與鮑魚菇轉形株之簡稱 ... 59
表十二 鮑魚菇以農桿菌媒介轉形時所使用之材料比較 ... 74
表十三 已發表能辨識CaMV 35S啟動子之物種 ... 79
表十四 鮑魚菇以不同啟動子啟動GFP之表現情形 ... 79
1
第一章 前言
一、基因工程
基因工程(genetic engineering)為在生物體外進行 DNA 選殖與建構,完成基因 重組後,利用轉形系統將重組基因送入目標細胞中,其目的主要可概分為三類 1. 基礎生物研究
範圍涵蓋遺傳學、生理學、形態學等,包括針對生物體內特定基因剔除或 突變及回補功能分析;融合報導基因之應用如研究生物體之細胞生物學、目標 蛋白質於生物體內的位置分佈、生物分子之交互作用、功能探討以及尋找功能 性基因、啟動子強度分析等。
2. 分子育種
傳統育種方式為人工篩選具觀賞或食用價值之優良農畜品系進行雜交,其 子代經不斷繁衍篩選後,以獲得改良的品種,育種過程費力耗時。分子育種 (molecular breeding)以基因工程之方式導入特定性狀,改進外觀、產量或風味口 感、加快生長速度、延長保存時間等,藉此提高其經濟價值。
3. 生產異源蛋白質
從自然界中取得有用的蛋白質必須經過純化的步驟,並且可能潛藏汙染或 感染的風險,拜基因工程所賜,人類得以利用生物體做為平台大量生產異源蛋 白質,這種生物性平台亦稱為異源表達系統。以醫藥研發為例,如利用大腸桿 菌生產人類胰島素(97);以農業生物技術為例,如將農作物轉入蘇力菌基因、
抗殺草劑基因、胡蘿蔔素合成路徑之酵素(7)等,提高糧食產量或價值;而工業 及環境應用如利用微生物生產替代能源、提供生物質材料或進行污染物處理 等。
2
藉異源表達系統生產目標蛋白質,使得生產成本降低並能夠提供穩定且較 為安全的來源。其中,生產重組醫藥蛋白質之表達系統開發仍為目前生物技術 中最受重視的應用之一,因此相關表達系統的選擇與轉形技術的改進成為提高 經濟效益的關鍵。本研究開發食用菇類中的鮑魚菇(Pleurotus ostreatus)做為異源 表達系統,改進其轉形系統並探討其表現異源蛋白質之可行性。
二、異源表達系統
自基因工程發展以來,異源表達系統亦隨之快速演進,從原核系統至真核 系統如真菌、動物、植物、以至近年來新興之食用菇類表達系統(104)。各表達 系統簡介如下,其特性與發展之優缺點評估如表一。
1. 原核表達系統
原核表達系統具產量高、生長週期短、成本較低之優點。大腸桿菌
(Escherichia coli)為此系統之中發展最早的,然而過去研究發現,在大腸桿菌體 內過量表現異源蛋白質時,常會造成蛋白質的不正常摺疊導致其被細胞內的蛋 白酶分解或是會分泌至內涵體(inclusion body)而不具生物活性且不利純化,雖 然有研究提出以摺疊調節因子如伴護子(chaperone)或摺疊酶(foldases)來幫助異 源蛋白質正確摺疊(88),或是藉由培養大腸桿菌的溫度降低至 30oC 能夠使蛋白 質不被送至內涵體(81),但大腸桿菌產生之蛋白質以胞內形式存在,其破菌純 化步驟常伴隨大腸桿菌內毒素(endotoxin)的釋出,使其不利生產醫用蛋白質。
枯草桿菌(Bacillus subtilis)能夠將蛋白質產物送至胞外,省去純化步驟,其 外泌蛋白質每公升可達公克等級的產量(52),此外,枯草桿菌屬革蘭氏陽性菌,
因此沒有內毒素的問題,是原核系統中較適於作為醫用蛋白生產之系統。儘管 如此,原核系統因缺乏蛋白質轉譯後修飾作用使得其應用有限。
3
2. 真核表達系統
2.1 酵母菌
釀酒酵母(Saccharomyces cerevisiae)為單細胞真核生物,具類似原核系統之 快速生長與操作容易特性,是真核表達系統中最早開發者。但其具有表達之蛋 白質會有過度醣基化現象、轉形株質體不穩定性、部分蛋白質會停留在胞質週 緣區(periplasm)、工業化量產不易(異源蛋白質最大產量約 1~5%總蛋白量)等缺 點(62),促成其他酵母菌表達系統的發展:如嗜甲醇酵母菌 Pichia pastoris、
Hansenula polymorpha 等,這些菌株可利用甲醇作為主要碳源,具有能受甲醇 誘導的強力啟動子,能利用來大量生產異源蛋白質。此類酵母菌具有可大量生 產胞內與胞外蛋白質,及高等真核生物之蛋白質修飾作用如醣基化、雙硫鍵形 成與蛋白酶切作用(proteolytic processing)等優點。
其中,P. pastoris 因為具商業化之套組而受到研究人員廣泛使用;相對而 言,H. polymorpha 因為可忍受較高溫度(30~42oC;P. pastoris:30oC)、有較快且 高產量的發酵能力且能以非甲醇(甘油受限策略)誘導而較受產業界的喜愛 (62)。1996 年 Gellissen 及 Lepetic 團隊以 H. polymorpha 生產 B 肝疫苗(31, 55),
在一年後通過臨床試驗並於1997 年上市(43)。
2.2 絲狀真菌
以絲狀真菌為表達系統之特色在於其可將大量蛋白質分泌至胞外,主要使 用的菌種為麴菌屬如 Aspergillus oryzae, Aspergillus niger 以及木黴菌屬
Trichoderma reesei,其中麴菌屬異源蛋白質產量可達每公升 100 毫克(75)。近 來也有以新的絲狀真菌表達系統如 Chrysosporium lucknowense 生產工業蛋白 質,其特色在於轉形效率高、發酵時間短、黏滯度低、可在中性pH 值環境下 生產蛋白質等而倍受矚目(65)。
增進絲狀真菌同源蛋白質產量的方法通常為轉入多套基因數、使用強力的 同源性啟動子、使外來基因插入染色體的特定基因序列等,至於異源蛋白質之 生產則將該段外來基因融合能夠有效分泌的內源性蛋白質(65),或是利用蛋白 水解酶(protease)突變株來提高生產量(75)。也有研究指出 N 端融合胜肽能穩定
4
外來基因的mRNA,具有促進異源蛋白質分泌、保護異源蛋白質不被分解的作 用(37)。
2.3 分子農場
分子農場(molecular farming)是利用活細胞或生物體大量生產重組蛋白的 概念,通常意指以農作物或牲畜為表達系統。分子農場生產之目標蛋白質可直 接使用,省去純化之成本,以植物與菇類(mushrooms)而言,相較動物表達系 統之優勢在於食用上的安全性,沒有動物性病源的疑慮且成本較低、可大規模 栽種與生產。以下就兩者做簡介。
2.3.1 植物
最初以植物作為醫藥蛋白質表達系統的幾個重要研究均以菸草(tobacco) 為研究對象,包括1986 年以農桿菌轉殖獲得的人類生長激素為第一個以植物 生產的醫藥蛋白質;1989 年首次以植物表現了抗體 IgG1 (phosphonate ester)顯 示了植物能夠組裝數個次單元的複雜醣蛋白,1992 年則表現 B 肝病毒表面抗 原,在動物試驗中能引發免疫反應,進一步證實植物表現異源蛋白質在結構上 的正確性。近期以植物生產之蛋白質更延伸至工業酵素、研究用蛋白質等(59),
然而植物生長週期較長,且轉基因植物在田間種植時花粉的傳播對生態環境的 影響仍受到諸多質疑,因此生長週期短、室內栽培之菇類相對而言更具潛力。
2.3.2 菇類
菇類(mushrooms)係指真菌界有性世代產生肉眼可見之子實體,主要涵蓋 擔子菌門(basidiomycetes)以及部分子囊菌門(ascomycetes)。以菇類做為表達系 統,除可免去基因汙染之疑慮外,亦有生長週期短、附加營養價值高以及具 有多種藥用活性物質與免疫調節等優點。
據估計,在八萬種已知的真菌中,約一萬種為菇類,其中可食用並已商 業化生產的約四十種,約五到六種以工業化大量生產之菇類如洋菇、鮑魚菇、
香菇、金針菇等。
5
表一 不同表達系統生產人類醫藥用重組蛋白質之比較(59)
Table 1. Comparison of production systems for recombinant human pharmaceutical proteins.
System Overall cost
Production timescale
Scale-up capacity
Product
quality Glycosylation Contamination risks
Storage cost
Bacteria Low Short High Low None Endotoxins Moderate
Yeast Medium Medium High Medium Incorrect Low risk Moderate
Mammalian cell culture
High Long Very low Very high Correct Viruses, prions
and oncogenic DNA Expensive
Transgenic animals High Very long Low Very high Correct Viruses, prions
and oncogenic DNA Expensive
Plant cell cultures
Medium Medium Medium High Minor differences Low risk Moderate
Transgenic plants Very low Long Very high High Minor differences Low risk Inexpensive
6
三、鮑魚菇概述
1. 形態特徵、分類地位、生活史
鮑魚菇(Pleurotus ostreatus)形狀似蠔,又稱蠔菇(oyster mushroom),中國大 陸亦稱之平菇,生長時為叢生,其蕈柄(stalk)側生於蕈傘(cap),如圖一所示,
其蕈傘顏色依光照程度有些微不同,主要為灰白色、淡褐色至褐色。蕈褶(gill) 面為乳白色或淡褐色,孢子印為白色。
鮑魚菇在分類地位上屬於真菌界(Fungi),雙核亞界(Dikarya),擔子菌門 (Basidiomycota) , 傘 菌 綱 (Agaricomycete) , 傘 菌 目 (Agaricales) , 側 耳 科 (Pleurotaceae)中的側耳屬(Pleurotus)。
擔子菌的有性孢子著生於蕈褶上的擔子(basidia),因而稱之擔子菌,擔子 菌之生活史可分為有性世代與無性世代,有性世代自擔孢子萌發成單核 (monokaryon)菌絲後,不同性別之菌絲進行交配(mating),在接觸面的細胞壁水 解,細胞核移動而形成雙核(dikaryon)菌絲;雙核菌絲在菌絲分隔(septa)處具扣 子體(clamp cell),為菌絲在有絲分裂(mitosis)過程中細胞核移動之用。雙核菌絲 生長過程中受環境因子(水、光照、溫度)變化刺激生成原基體(primordia,又稱 菇蕾),並進一步分化成子實體(fruiting body)。子實體主要部分為蕈傘、蕈柄,
或部分擔子菌具蕈杯(volva),蕈傘內部雙核菌絲末端特化出雙核擔子,經核融 合(karyogamy)與減數分裂(meiosis)後產生單核之擔孢子(basidiospore);擔孢子 掉落後萌發成長之單核菌絲產生節生孢子(oidia or arthrospores)則為其無性世 代(15)。圖二以灰蓋鬼傘(Coprinus cinereus)為例說明擔子菌生活史與子實體發 展過程。
7
圖一 鮑魚菇之形態
Figure 1. Morphology of Pleurotus ostreatus 圖片來源:http://www.natur-lexikon.com/
圖二 擔子菌灰蓋鬼傘之生活史與子實體發展過程
Figure 2.The hallmark feature of the basidiomycete fungal life cycle is illustrated here by C. cinereus. The lower diagram illustrates the stages in the development of the fruiting body. Figure originated from Casselton, L., and M. Zolan., 2002 (15).
8
2. 栽培優勢
側耳屬菇類為白腐真菌(white rot fungi),可利用多種基質(如稻草、麥桿等 農業廢棄物)栽培,以木質纖維基質栽培則可省去堆肥、覆蓋土等步驟(21)。相 關研究指出蠔菇可利用香菇栽培使用過的基質,同為側耳屬的杏鮑菇對於使用 過的基質利用及出菇效率甚至更勝於未使用過的基質,顯示側耳屬菇類在栽培 上可大幅降低生產成本、減低木屑使用率。目前在台灣的商業化生產仍以木屑 太空包栽培為主,已有研究指出鮑魚菇的栽培基質可以蔗渣取代三分之二的木 屑量,同時能提升出菇產率。
除基質轉換率高以外,鮑魚菇菌絲生長最適溫度約25~30 oC,其出菇溫度 較高(14~20oC)且出菇時間短(約 2~3 週),使其在栽培上更具競爭力。
3. 藥理活性
據研究,鮑魚菇具許多藥理功效與活性成分,包含從菌絲體萃取之蛋白聚 醣,具抗腫瘤與免疫調節活性(80)、從菇體內萃取之新型凝集素(lectin),在動 物實驗證實具抗肝癌與抗腫瘤(94)功效,並且也有其他成分具抗氧化活性 (47),其他鮑魚菇已發表的藥理成分與功效請參見表二。
4. 其他應用價值
鮑魚菇為白腐真菌,具降解木質素與木質纖維的能力,含有多種具工業 應用價值之酵素如peroxidases、laccases、cellulases、hemicellulases以及
xylanases.,可應用於過氧化氫之生產、紙漿與造紙業及畜牧飼料業上;更有研 究指出鮑魚菇能夠代謝毒化物質,具生物復育潛力(21)。
9
表二 鮑魚菇的藥理成分與功用
Table 2. Medicinal effects and substances of Pleurotus ostreatu
Medicinal effects Substances References
Antiviral Ubiquitin-like protein (94)
Immunomodulating Glucan (71, 80)
Antitumor Glycopeptides
Mushroom grown on corncobs Mushroom
Lectin β-D-Glucan (pleuran)
(56) (103) (9)
Anticholesterolic Mushroom (67)
Antioxidant β-D-Glucan (pleuran) (8)
10
四、食用菇類轉形系統
食用菇類的轉形方法多以細胞壁水解酵素處理成原生質體(protoplast)的細胞 形式後,再以電穿孔(electroporation)、PEG (polyethylene glycol)法或是搭配限制酶 的 REMI (Restriction Enzyme Mediated Integration)法使外來基因能穩定嵌入染色 體。另有少數以基因槍轟擊法(particle bombardment)進行轉形之研究。
以鮑魚菇而言,早期為製作原生質體的方式進行電穿孔、PEG(72, 73)或 REMI 轉形(45)。2001 年 Irie 等人利用 PEG 法外加單股 DNA 有效提高原生質體方法為主 的轉形效率 50 倍(45),然而原生質體製做不易且細胞再生率低仍造成轉形效率降 低。2002 年 Sunagawa 與 Magae 以製作成菌絲塊(Mycelial disks)的方式搭配基因槍 粒子轟擊法(86),雖避免了製備原生質體,然而轉形率並無顯著改善。前述研究主 要多以同源或異源抗藥性基因為轉殖目標,2006 年 Li 等人的研究以 GFP 做為報 導基因進行PEG 法轉形,但綠色螢光在鮑魚菇轉形後 30 個小時消失(56)。2008 年 由 Kuo 建立鮑魚菇等食用菇類以酵素緩和處理的發芽擔孢子或菌絲片段之電穿孔 轉形系統使得轉形效率顯著提升,同時成功表現了真核的水母來源綠色螢光蛋白 基因(104)。但電穿孔轉形法對細胞傷害大且外來基因嵌入宿主染色體數目不一,
宿主生理受影響較多可能導致其異源基因較不穩定。此外,多基因嵌入對於以分 生策略提升異源基因表現量的比較也不易判定。鮑魚菇轉形系統之發展列於表三。
自 1998 年 Groot 等人利用農桿腫瘤菌媒介轉形法(Agrobacterium tmefaciens mediated trransformation)將抗藥性基因(hygromycin phosphotransferase)成功轉入洋 菇內表現後(24),農桿腫瘤菌為食用菇類的轉形帶來新的契機,表四列出成功以農 桿菌轉形之食用菇類。
農桿菌媒介轉形法轉入之異源基因為隨機嵌入宿主染色體且多是單一套數,
因此能穩定表現外來基因,在眾多轉形方法中其操作較為便利且可選擇材料範圍 大、成本低廉,甚至可將150 kb 大的片段轉形至植物宿主染色體且穩定遺傳至子 代(39),是深具潛力的轉形系統。除了應用於植物轉形外,也已經成功應用於真菌 類如酵母菌(11)、麴菌(36)、鐮孢菌(63)、木黴菌(101)及青黴菌屬(99)等。其中以子 囊菌最多,至少有五十種已成功轉形;接合菌、卵菌則占少數。而擔子菌至少十 二種(61),顯示農桿菌可轉形宿主範疇多樣,並不侷限於植物而有更多發展可能。
11
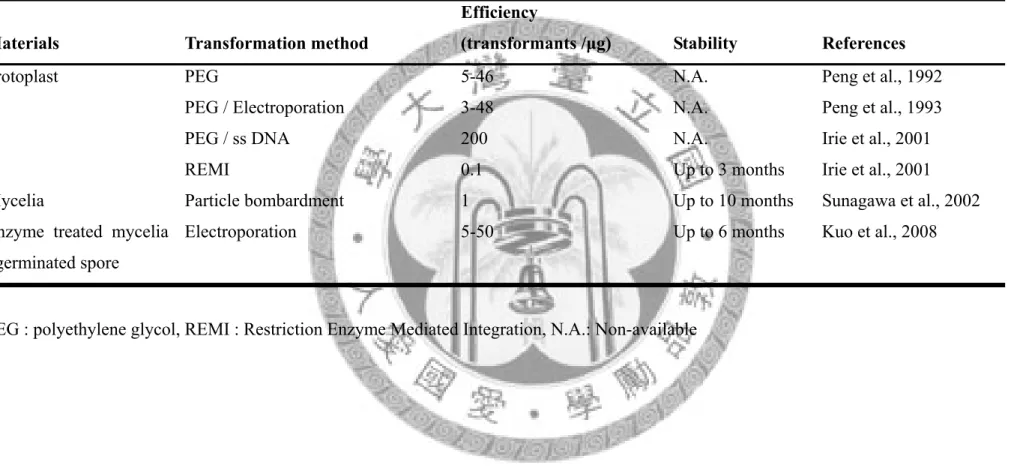
表三 鮑魚菇轉形系統之發展
Table 3. Progressing of transformation system in Pleurotus ostreatus
PEG : polyethylene glycol, REMI : Restriction Enzyme Mediated Integration, N.A.: Non-available Materials Transformation method
Efficiency
(transformants /μg) Stability References
Protoplast PEG 5-46 N.A. Peng et al., 1992
PEG / Electroporation 3-48 N.A. Peng et al., 1993 PEG / ss DNA 200 N.A. Irie et al., 2001 REMI 0.1 Up to 3 months Irie et al., 2001 Mycelia Particle bombardment 1 Up to 10 months Sunagawa et al., 2002 Enzyme treated mycelia
/ germinated spore
Electroporation 5-50 Up to 6 months Kuo et al., 2008
12
表四 成功以農桿菌轉形之食用菇類
Table 4. Edible mushrooms successfully transformed by Agrobacterium
hph Escherichia coli hygromycin resistence gene, afp budworm antifreeze protein gene, EGFP enhanced green fluorescent protein, GUS β-glucuronidase gene
Species Agrobacterium strain used Target gene Year References Agaricus bisporus (洋菇) AGL-1, LBA1100, EHA105 hph 1998, 2000, 2001 (17, 24)
Suillus bovines (乳牛肝菌) AGL-1, LBA1100 hph, EGFP 2002 (69)
Hypholoma sublateritium (栗蕈) AGL-1 hph 2004 (35)
Tricholoma matsutake (松茸) AGL-1, EHA105 hph 2006 (64)
Volvariella volvacea (草菇) EHA105, LBA4404 afp 2008 (95)
Antrodia cinnamomea (牛樟芝) EHA105 hph 2008 (16)
Flammulina velutipes (金針菇) AGL-1, LBA4404 hph, EGFP, GUS 2008 (105)
13
五、農桿菌媒介轉形
1. 農桿腫瘤菌
農桿菌(Agrobacterium spp.)廣泛存在於土壤中,主要由腐生菌組成,然而 部分農桿菌因帶有表現毒性蛋白質(virulence protein)的質體,為植物致病菌,
可以感染雙子葉植物及少數單子葉植物導致其生長變異,例如農桿根群菌
(Agrobacterium rhizogenes)使植物生成毛狀根(hairy root);農桿腫瘤菌
(Agrobacterium tumefaciens)造成植物冠癭腫瘤病徵(crown gall disease)等,
皆為典型例子 (29)。
農桿腫瘤菌的致病過程主要由腫瘤引發質體(Tumor inducing plasmid, Ti plasmid)所調控,該質體為一環狀巨型雙股DNA分子,全長約200 kb,質體可概 分為三部分:毒性蛋白質基因、由兩段約 25 bp 重複的片段Right border與Left border夾帶的T-DNA (Transferred DNA),以及與 opine 代謝相關的酵素; 原生 型T-DNA上帶有催化植物合成生長激素如auxin、cytokinin與opine的基因,在毒性 蛋白質的輔助下送入宿主細胞並嵌入宿主染色體中而表現,致使植物生長激素失 調而生成冠癭腫瘤,該腫瘤產生opine,而農桿腫瘤菌得以利用宿主生產的opine 作為營養源(32, 60)。1980年科學家首次將原生型T-DNA置換為異源基因,完成農 桿腫瘤菌媒介的重組DNA轉形植株,此後,農桿腫瘤菌以「自然的基因工程師」
普遍應用於基因轉殖研究,並且有更多與農桿腫瘤菌感染機制相關的研究仍在進 行中;有關首次探討農桿腫瘤菌所引發的冠癭腫瘤疾病以及其後將近一個半世紀 間,農桿腫瘤菌相關的重要研究整理於表五(29)。
14
表五 農桿腫瘤菌與其腫瘤引發病症於植物之部分研究
Table 5. Selected discoveries and insights in Agrobacterium biology(29)
Year Discovery References
1853 首次發表植物冠癭腫瘤的研究報告 Fabre, Dunal
1897 鑑定出Agrobacterium vitis 為造成葡萄冠癭瘤的病因 Cavara
1907 鑑定出 Agrobacterium tumefaciens 為造成 Paris Daisy 冠癭瘤的病因 Smith and Townsend 1947 植物腫瘤組織在激素缺乏的環境下能不斷增生,因而提出這些植物腫瘤細胞已被農桿菌轉形 Braun
1971 Agrobacterium tumefaciens 在 37oC 下培養不具致病力;致病力能在致病株與非致病株間轉移 Kerr 1974 Agrobacterium tumefaciens 的致病力與腫瘤引發質體(Ti plasmid)的存在有關 Zaenen 1977 Ti plasmid 上的 T-DNA 嵌入冠癭腫瘤細胞的染色體中,該 T-DNA 與腫瘤形成有關 Chilton 1980 被農桿菌轉形的細胞合成opine 提供良好環境予農桿菌生長 Guyon 1980 首度利用 Agrobacterium tumefaciens 轉形重組質體至植物 Francois
1984 T-DNA 上的致癌基因調控植物生長激素 auxin 和 cytokinin 大量產生 Klee et al ; Lichtenstein 1985 virA/virG 二元調節系統在農桿菌轉形中扮演訊息接收與傳導的角色 Stachel and Zambryski 1986 ~ 更多與T-DNA轉移過程相關的毒性蛋白被發現與探討;鑑定參與農桿菌轉形的宿主基因;
擴大農桿菌轉形宿主的範疇;針對Agrobacterium tumefaciens (C58)進行染色體DNA解碼
15
2. 農桿腫瘤菌的感染過程
植物受傷組織最易受到農桿腫瘤菌的感染,感染過程與機制大致如圖六,分 述如下:
2.1 附著宿主細胞
植物受傷部位會釋放胺基酸、有機酸以及糖類(sugars),這些物質吸引農桿腫 瘤菌附著到植物細胞上。農桿菌首先藉由合成乙醯多醣附著,隨後伸出纖毛固定 在植物細胞上(29)。
2.2 毒性蛋白質活化
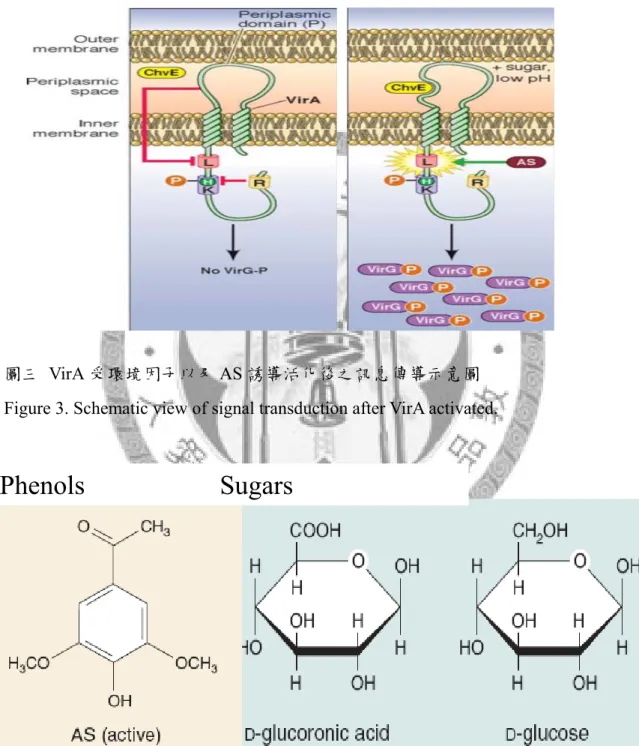
農桿腫瘤菌附著植物細胞的同時,腫瘤引發質體上的毒性蛋白質操作組亦開 始啟動表現,該基因群為T-DNA的轉移所必須,受到持續表現的VirA/VirG二元調 節系統調控,該系統主要藉由激酶 (Vir A組成之一) 對於下游調控蛋白(VirG)磷 酸化與否來控制往下的訊息傳遞 (98)。
植物受傷組織爲進行傷口修復,該處細胞苯基丙酸路徑(phenylpropanoid pathway)頻繁而生成大量酚類化合物,胞外環境呈酸性(pH 5.0-5.5),單糖累積以 合成細胞壁等(6),這些因子共同以直接或間接方式誘導穿膜受器激酶VirA自動磷 酸化(autophosphorylation)其組胺酸(Histidine)而活化,活化型VirA進一步將磷酸根 轉移至VirG的天門冬胺酸(Aspartate)上,磷酸化的VirG得以專一性結合毒性蛋白 基因上游調節位(vir boxes),調控毒性蛋白質基因表現(10)。VirA活化後引發之構 形變化與訊息傳導示意圖如圖三。
植物受傷位置細胞釋放毒性蛋白質誘導物如酚類化合物
3,5-Dimethoxyacetophenone(亦為acetosyringone [AS],其結構如圖四,重要結構特 徵包括芳香環上的氫氧基[aromatic hydroxyl]以及甲氧基[methoxyl group] )。最早 自植物根部及葉片原生質體檢測出,該成分含量高並且具高度誘導毒性蛋白質表 現的活性,其活性大小與甲氧基之多寡有關:帶有兩個甲氧基之結構活性最高,
單一甲氧基次之,不帶甲氧基者活性最小。無論是人工合成或是自植物苯基丙酸 路徑合成的酚類衍生物皆具誘導活性。另外,一些研究指出醛糖亦可作為誘導
16
物,形成如圖四的六環糖為活性關鍵,其中酸性糖具最高的誘導活性;而酸性環 境可能與VirA和ChvE (農桿腫瘤菌染色體上一段與致病力有關之基因編碼的膜蛋 白,與糖類結合蛋白有高度同源性)間的交互作用有關,亦影響訊息的傳遞,圖三 (60)。
圖三 VirA 受環境因子以及 AS 誘導活化後之訊息傳導示意圖 Figure 3. Schematic view of signal transduction after VirA activated.
圖四 誘導農桿腫瘤菌 vir 基因表現的物質結構式。酚類化合物 AS:
acetosyringone; 六環糖類:葡萄糖醛酸、葡萄糖。
Figure 4. Structural formula of substances capable of inducing vir gene expression.
Phenols Sugars
17
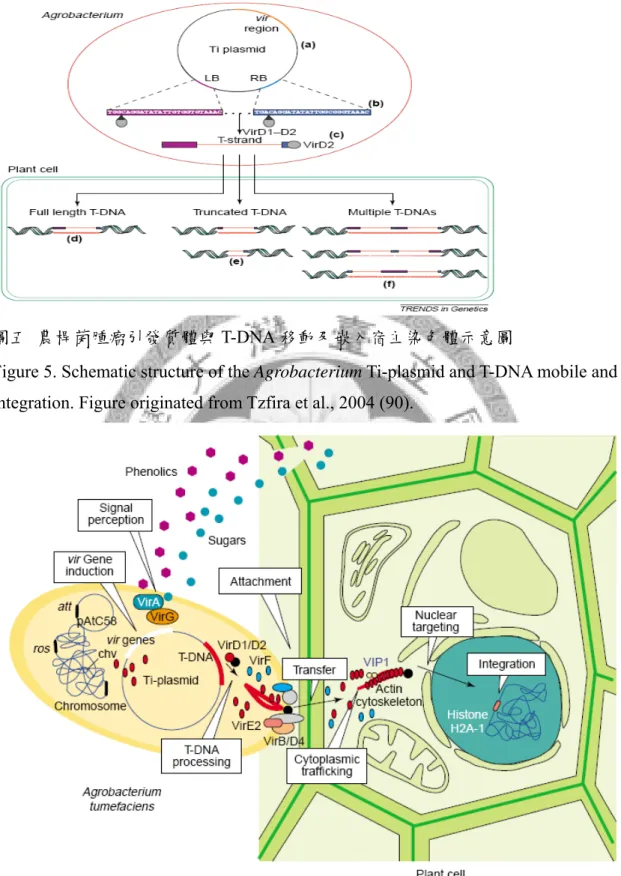
2.3 T-DNA形成
毒性蛋白質表現後,具單股核酸內切酶活性的VirD1與VirD2於T-DNA邊緣作 用,切開T-DNA底股(bottom strand)近RB與LB位置。新的一股DNA自RB位置開始 以頂股(top strand)為模板合成至LB為止,取代原本底股DNA,而裸露的底股DNA 即為可傳送的單股T- DNA (84)。毒性蛋白質VirD2結合T-DNA 5’端而與T-DNA共同 經由第四型分泌系統送入植物細胞。
2.4 傳送T-complex(Transport complex)與毒性蛋白質到宿主體內
第四型分泌系統(Type 4 Secretion System, T4SS)在細菌體內參與蛋白質與遺傳 物質傳送的過程,農桿菌利用此分泌系統運送物質到宿主細胞內。該系統為 VirB/D4複合體,主要由至少11個VirB與1個VirD4所組成,其功能包括提供運送的 能量、形成穿膜孔道以及構成接合線毛(又稱 T-pilus),這些組成結構協同作用下 幫助基質(VirD2-T-strand, VirE2, VirE3, VirF以及VirD5等)穿過農桿菌與宿主細胞間 的屏障。在農桿菌體內,VirD4為輔助蛋白質(Coupling protein),與T-complex上的 蛋白質(如VirD2)交互作用後,藉由與T4SS其他的組成蛋白質(如VirB10)結合,將 T-complex送至農桿菌細胞膜上的第四型傳送系統附近,進一步利用ATP水解產生 的能量將基質送到細胞外(14);此外,VirD4也能直接與其他毒性蛋白質C端結合(如 VirE2),幫助其送至T4SS附近(3)。研究顯示,VirE2在in vitro的情況下會形成可傳 送單股DNA的通道,其他研究亦提出VirE2與T-complex分別先後送入宿主體內,前 者在T-DNA進入宿主細胞的同時迅速與之結合並摺疊T-DNA,摺疊時釋出能量提 供T-DNA進入宿主細胞之所需(27, 28, 38)。此外,VirE2的結合也防止T-DNA受宿 主體內核酸酶降解。VirE2在農桿菌中為一多功能的蛋白質,藉由感染時傳送到宿 主細胞內,具有保護 T-DNA及與其他毒性蛋白質或宿主蛋白質發生交互作用等功 用 (96)。
2.5 T-DNA 進入宿主細胞核
T-DNA進入宿主細胞核的機制亦受毒性蛋白質媒介,包括VirD2與VirE2。VirE2 與VirD2被認為直接與T-DNA結合而形成T-complex (102),其進入宿主細胞核與宿 主自身的蛋白質有關。VirE2與VirD2由不同的機制進入細胞核內,VirD2由一種在
18
動物、酵母菌與植物細胞間具保守性的機制調控進入核內,而VirE2進入細胞核內 卻是由植物專一性的機制所調控(19)。此外,一已知可調節VirD2進入到核內的植 物蛋白質,阿拉伯芥的karyopherin α protein,無法辨認VirE2 (4),更顯示兩者是藉 由不同的方式進入宿主細胞核。後者在阿拉伯芥的研究主要藉由與宿主的蛋白質 VirE2 - interacting protein 1 (VIP1)結合而進入宿主體內(91)。宿主核外蛋白質進入 核內的目的在於啟動宿主的防禦基因活化表現,農桿菌進而演化出利用宿主此免 疫啟動的機制,以毒性蛋白質(如VirE2)與宿主調節防禦基因表現的蛋白質發生交 互作用,藉由宿主蛋白質的核轉移將T-DNA送入宿主細胞核內(26)。
2.6 T-DNA嵌入宿主染色體
在農桿菌媒介的植物轉形系統中,T-DNA插入宿主染色體位置多為隨機插 入,然而部分研究指出其插入位置序列與T-DNA的3’端有微同源性
(microhomologies)。在酵母菌的研究中,T-DNA可以非同源性重組(non-homologous recombination, NHR)或同源性重組(homologous recombination, HR)的方式插入酵母 菌染色體中,而植物方面的研究則發現依轉形組織的不同, T-DNA會以不同方式 嵌入宿主染色體(90),顯示T-DNA邊緣序列、宿主DNA序列以及農桿菌感染時宿主 的生理狀態和環境條件皆會影響T-DNA插入位置。在植物阿拉伯芥的研究中發 現,T-DNA傾向嵌入基因間隔區(intergenic regions)、啟動子、5’以及3’端非轉譯區 域(2, 79),但這些結果可能為轉形株經過人為抗藥性篩選所致,在未經篩選的轉形 株分析發現,T-DNA的插入確為隨機(34)。T-DNA多以單一拷貝數插入染色體單一 位置,或是以多個拷貝數重疊插入單一位置,如圖五所示,其T-DNA border可能完 整保留或部分保留,通常left border插入宿主染色體的完整度較高。
雖然有許多假說提出,但造成宿主染色體缺口的機制以及T-DNA接上宿主染 色體DNA的方式目前仍無定論,僅推測在T-DNA接合上宿主染色體的過程中,
VirD2與某些宿主蛋白質可能以接合酶的角色參與其中,此外,部分宿主蛋白質如 DNA修補酵素在T-DNA嵌入染色體過程中亦不可或缺(90)。
19
圖五 農桿菌腫瘤引發質體與 T-DNA 移動及嵌入宿主染色體示意圖
Figure 5. Schematic structure of the Agrobacterium Ti-plasmid and T-DNA mobile and integration. Figure originated from Tzfira et al., 2004 (90).
圖六 農桿腫瘤菌感染植物造成之 T-DNA 轉形過程示意圖
Figure 6. The model of T-DNA transformation process in plant cell during Agrobacterium tumefaciens infection. Figure originated from Gelvin, 2003 (33).
20
3. 農桿腫瘤菌在基因轉殖的應用
自1980年科學家將細菌的跳躍子基因(Transposon, Tn7)置入農桿菌腫瘤引發質 體而轉入植物細胞後(42),農桿菌即廣泛應用於植物基因工程上。以農桿菌進行轉 形時,轉入的異源基因套數較少,大部份的情況為異源基因以單一套數嵌入宿主 染色體,此情況相較於其他轉形方法,對宿主生理的影響較小因此能較為穩定的 表現異源基因(25),而未知基因標定也較為容易。儘管如此,農桿腫瘤菌在自然界 的宿主為雙子葉植物,因此仍有部分植物不易以農桿菌進行轉形,然而在實驗室 操作下,農桿菌可轉形的宿主範疇已擴大至真菌(24, 36)甚至是人類細胞(51)。
1995年Bundock等人的研究發現農桿腫瘤菌可以感染酵母菌而使T-DNA轉移 嵌入酵母菌染色體內,雖然其轉形機制似乎不同於農桿菌對植物細胞的轉形(11),
但該研究開啟了以農桿腫瘤菌應用於真菌轉形系統之先例。隨後的研究亦發現部 分絲狀真菌、菇類皆能夠以農桿腫瘤菌成功轉形且能穩定表現異源基因(24),農桿 腫瘤菌進而發展為部分真菌轉形或致突變的工具。然而,農桿腫瘤菌媒介之轉形 在應用於特定真菌物種前仍需經過感染條件測試與改進,其考量因素包括:
1. 轉形材料的選擇
真菌可選擇的材料如菌絲、菌塊、蕈褶組織、孢子或發芽孢子、原生質體 等,不同種類的真菌各有其最易感染的細胞形態,可以參考Caroline等人將已 知能被農桿菌成功轉形之真菌所使用的起始材料做的整理(61)。
2. 農桿腫瘤菌菌株與質體
過量表現特定毒性蛋白質的菌株對於片段較長之異源基因轉形效率可能 較 佳(39) , 而 不 同 菌 株 亦 有 其 最 易 感 染 的 物 種 對 象 , 例 如 在 板 栗 疫 菌 (Cryphonectria parasitica)的研究發現,農桿菌菌株A281與AGL-1的轉形效率較 LBA4404高(70),此外,草菇轉形之研究也提出EHA105的轉形效率較LBA4404 為佳(95)。
1983年Hoekema等人以及Framond發現毒性蛋蛋白質基因與T-DNA位於同 一農桿菌株之不同的質體上仍保有轉形效果後,便發展出雙偶型載體系統。雙
21
偶型載體系統之T-DNA位於雙偶型載體上,而傳送T-DNA相關之毒性蛋白質基 因則位於農桿菌體內的輔助質體(helper plasmid)上(54)。雙偶型系統發展25年 來,也有許多符合不同需求的設計與相關商品化質體,不同菌株搭配不同質體 的使用也會影響轉形效率(13),Lee及Gelvin(54)對較常使用之雙偶型載體特色 與常用農桿菌去致病性(disarmed)菌株做了較完整的介紹。
3. 共培養條件
包括誘導物在農桿菌前處理時的添加濃度以及共培養時的添加濃度,皆會 影響轉形效率(53);真菌與農桿腫瘤菌菌數的比例,真菌擔孢子與農桿菌數比 例或調整農桿菌數目皆會影響轉形效率(1, 16, 35);pH 值,已有研究指出pH值 介於5.0 ~ 5.3之間有最佳的轉形效率(87, 89);溫度,研究指出農桿菌感染酵母 菌時其 T-DNA 轉形效率隨共培養溫度下降(28oC降至22oC)而上升(30),亦有研 究指出真菌最佳感染溫度為22oC~25oC (22, 78);光照,針對農桿菌之研究發現 在光照情況下農桿菌泳動能力下降,對於植物根毛細胞的附著力降低,致病力 減弱(66)
22
六、分生策略
1.啟動子
本研究中使用之主要啟動子為鮑魚菇同源與異源如洋菇、金針菇之甘油醛-3- 磷酸脫氫酶(glyceraldehydes-3-phosphate dehydrogenase, GPD )上游啟動子。甘油醛 -3-磷酸脫氫酶除了在細胞內參與糖解(glycosis)與醣升(gluconeogenesis)作用外,也 與膜運送與胞吞作用、核內tRNA 傳送到細胞質、轉譯調節、核外 DNA 複製和修 補有關(5, 83),為持續表現的基因,因此使用其啟動子可望提高下游基因表現量。
另外使用花椰菜鑲嵌病毒(Califlower mosaic virus, CaMV)來源之啟動子測試 其是否能被鮑魚菇辨識及啟動子效果如何。花椰菜鑲嵌病毒常感染十字花科植 物,特別是甘藍菜、花椰菜、油菜、芥菜等,為了能在宿主體內複製自己的DNA,
病毒演化出壓過宿主調節系統的強力啟動子(35S 與 19S 啟動子),將其置於自身基 因之前,誘騙宿主持續表現其基因。目前在植物轉形中,最常使用的是 35S 啟動 子,幾乎應用於所有基因改造作物上,特別是基改玉米。CaMV 35S 啟動子重複序 列(duplicated)甚至能增加植物體內之基因表現達數百倍以上,對於下游較遠端之基 因亦有少許啟動作用(49)。
2.報導基因
本研究中以綠色螢光蛋白質 (Green fluorescent protein, GFP)為報導基因評估 異源蛋白質表現量的多寡。綠色螢光蛋白質有238 個胺基酸,其分子量約 27kDa,
於1962 年由日本學者 Osamu Shimomura 等人自水母 Aequorea victoria 體內純化並 研究其特性,1992 年 Douglas Prasher 等人選殖出綠色螢光蛋白質基因促進了此後 螢光蛋白質的相關研究與做為報導基因之應用,然而野生型綠色螢光蛋白質激發 波峰非單一(395 nm 與 475 nm)且在 37oC 下摺疊不易,促使研究人員開發了綠色螢 光蛋白質突變型,如S65T 突變型,使激發波峰發生紅移至單一波峰 488nm。綠色 螢光蛋白質由於不需基質、穩定而耐熱(可達 65oC),適於做為偵測基因表現量的指 標。
23
3.內含子與訊息胜肽
研究發現裂摺菌(Schizophyllum commune)表現潮黴素磷酸轉移酶以及洋菇 (Agaricus bisporus)、灰蓋鬼傘(Coprinus cinereus)表現綠色螢光蛋白質時,必須有 內含子(intron)存在才能表現,這可能是因為內含子能維持 mRNA 之穩定而可以在 細胞內累積足夠的量(12, 58);另外,在裂摺菌等真核表達的研究發現提高異源基 因之G+C 含量可增進表達原核基因的效率(82)。
另有研究提出蛋白質 C 端帶有訊息胜肽時,能提高異源蛋白質表現量,其目 的為使目標蛋白質停留在特定位置而累積,這些訊息胜肽如SEKDEL、KDEL、
HEEL 或 HDEL。
4.篩選標記
在植物與菇類表達系統中,最常使用的異源篩選標記為潮黴素磷酸轉移酶 (hygromycin B phosphotransferase),以潮黴素(hygromycin B)篩選轉形株。潮黴素為 胺基環醣醇類抗生素,具廣效活性,能影響真核與原核細胞生長,其方式為干擾 tRNA 辨識、與核糖體結合影響其移位(translocation),進而抑制蛋白質合成。多數 胺基環醣醇類抗生素被酵素作用而失去活性,包括乙醯化(acetylation) 三磷酸腺甘 酸化(adenylation)以及磷酸化(phosphorylation)作用。潮黴素磷酸轉移酶存在於對潮 黴素具抗性之細菌或產生潮黴素的菌株 Streptomyces hygroscopicus(76),成功轉入 並表現該酵素之轉形株即能在含有潮黴素之選擇性培養基生長。
鑒於異源抗藥性基因對於生態的影響,許多研究也以營養缺陷株、同源性篩 選 標 記 進 行 轉 形 株 的 篩 選 , 如 鮑 魚 菇 已 發 表 之 琥 珀 酸 脫 氫 酶(succinate dehydrogenase)其鐵硫蛋白(iron sulfur protein)經點突變後具有 carboxin 抗性,相關 研究中也認為同源性篩選標記的使用能夠提高轉形效率(44, 46)。而在草菇轉形的 研究中,使用夜蛾(budworm)的抗凍基因(Antifreeze protein, afp)進行抗凍轉形株篩 選,亦為免去細菌抗藥性基因汙染的策略提供不同的選擇(95)。
24
七、研究動機與目的
本實驗室先前已建立鮑魚菇等食用菇類之電穿孔轉形系統(104),但電穿孔轉 形法對細胞傷害大且外來基因嵌入宿主染色體數目不一,可能導致其異源基因較 不穩定。農桿菌媒介轉形法轉入之異源基因為隨機嵌入宿主染色體且大多是單一
套數,因此能穩定表現外來基因,其操作便利、成本低廉,近年由本實驗室 Hsu
成功建立金針菇之農桿菌媒介轉形法(105),然而不同物種有其最適轉形的細胞形 態與條件,若能發展作為鮑魚菇的轉形工具可望提高鮑魚菇的研究與應用層面。
本論文欲探討鮑魚菇利用農桿菌媒介轉形法之可行性並建立最適化條件,利 用實驗室先前選殖的鮑魚菇同源性 GPD 啟動子,配合其他菇類(洋菇、金針菇)來 源的 GPD 啟動子以及花椰菜鑲嵌病毒來源之啟動子,表現綠色螢光蛋白質基因以 比較不同來源啟動子啟動基因表現的情形與差異性;另比較報導基因融合內質網 停留訊息胜肽的蛋白質表現情形。期以農桿菌媒介轉形法能使異源基因於鮑魚菇 穩定表現進而在未來應用於醫藥蛋白質的生產。
欲達上述目的所需達成之目標包括:
1.表現質體建構
1.1 以 pCAMBIA0380 為骨架,置入鮑魚菇同源性 GPD 啟動子啟動之篩選標記 hph 1.2 將項目 1.完成之重組質體置入鮑魚菇同源性 GPD 啟動子啟動之報導基因 EGFP 1.3 將項目 1.完成之重組質體置入鮑魚菇同源性 GPD 啟動子啟動之 C 端帶有訊息
胜肽(HDEL)的報導基因 EGFP
2.鮑魚菇以農桿菌媒介轉形測試
2.1 將前述表現質體、本實驗室已建構帶有洋菇啟動子啟動之 GFP、金針菇啟動子 啟動之 GFP 質體以及商品化質體 pCAMBIA1302 藉電穿孔方式送入農桿菌株 AGL-1 與 LBA4404
2.2 製備鮑魚菇菌絲塊、菌絲液與蕈褶組織塊做為轉形材料
2.3 將帶有表現質體之農桿菌株與不同的鮑魚菇材料進行共培養,另測試共培養天 數、誘導物 AS 濃度。
25
3.轉形株篩選與分析
3.1 將生長於選擇性培養基上之菌絲以螢光顯微鏡觀察 GFP 表現情形 3.2 以西方式雜合分析進行 GFP 定性
3.3 以三明治法酵素連結免疫吸附分析對轉形株之 GFP 進行定量
4.轉形株穩定度探討
4.1 異源基因無性世代穩定性
轉形株於選擇性培養基上繼代四次後,於不含抗生素之培養基繼代四次,進 一步轉至選擇性培養基,觀察是否仍具抗性。轉形株於液態培養基培養時產 生之無性節生孢子以螢光顯微鏡觀察 GFP 表現情形
4.2 異源基因有性世代穩定性
將轉形株進行出菇後,以螢光解剖顯微鏡觀察蕈褶組織 GFP 表現情形,收集 擔孢子以螢光顯微鏡觀察 GFP 基因是否遺傳至子代並表現,另將擔孢子塗佈 於選擇性培養基上觀察生長情形。
26
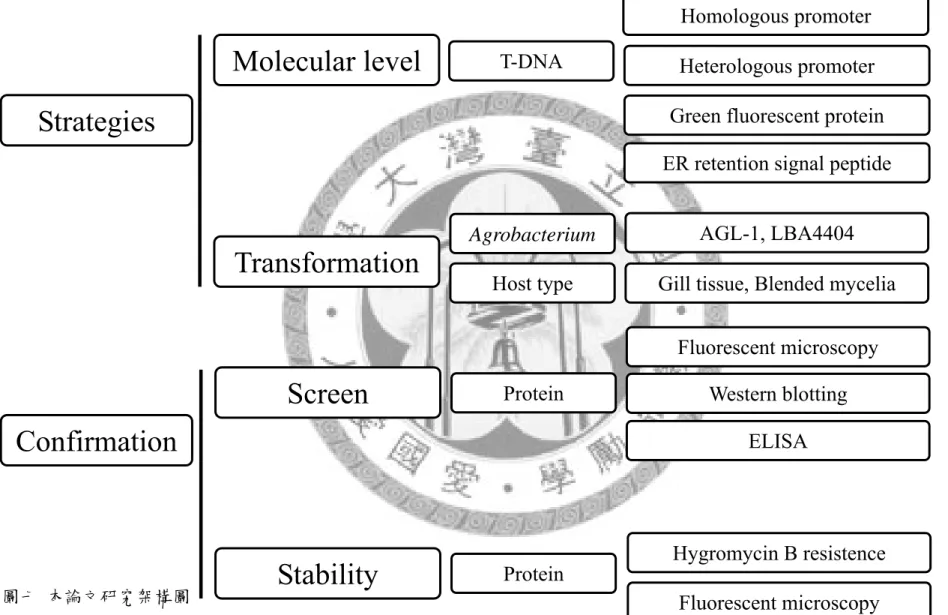
圖七 本論文研究架構圖
Figure 7. Schematic framework of this study
Strategies
Confirmation
Stability Molecular level
Transformation
Screen
T-DNA
Agrobacterium Host type
Protein
Protein
Homologous promoter Heterologous promoter
ER retention signal peptide Green fluorescent protein
AGL-1, LBA4404 Gill tissue, Blended mycelia
ELISA Western blotting Fluorescent microscopy
Hygromycin B resistence Fluorescent microscopy
27
第二章 材料與方法
一、實驗材料
1. 菌株及培養條件
1.1 真菌
鮑魚菇(Pleurotus ostreatus)雙核菌株與單核菌株為本實驗室保存。使用之培養 基為1.5% PDA (potato dextrose agar)平板培養基,於 25 oC 避光培養,三周後確認 菌絲形態無誤後,將平板培養基上菌絲生長邊緣之菌塊接種至1.5% PDA 斜面培 養基上保存,約每三個月接到新鮮PDA 斜面培養基。
將木屑培養基(木屑:米糠 = 3:1,含水量約 70 %)裝填於耐高溫之塑膠袋內壓 實,置於玻璃燒杯中滅菌降溫後,接種鮑魚菇雙核菌絲,待菌絲長滿後(約三周),
以滅菌之接種棒劃開塑膠袋及菌絲,添加水分並佐以15oC 低溫、光週期(9hr 光照 /15hr 黑暗)等出菇刺激後約兩到三周可得無菌子實體,如圖八。
1.2 細菌
大腸桿菌(Escherichia coli ) DH5α 菌株為質體保存使用之宿主細胞,大腸桿菌 Top 10 菌株則為分生操作使用之細胞,37 oC 下培養於含 1.5% Agar 之固態
Luria-Broth (LB, Acumedia Manufacturers, Michigan)平板培養基或液態 LB 培養基 中,液態培養震盪條件為250 rpm。另以含 25 % Glycerol 之 LB 培養基長久保存於 -80oC。
農桿腫瘤菌(Agrobacterium tumefaciens)菌株為 AGL-1 與 LBA4404,亦培養於 LB 培養基,培養溫度為 28 ~ 30oC,液態震盪條件為 250 rpm,長久保存條件同大 腸桿菌。
28
2. 質體
2.1 pCAMBIA0380
pCAMBIA0380 (CAMBIA, Australia)全長約 6.8 kb,為雙偶型載體,由長度各 25 bp 之 Left border 與 Right border 夾帶 T-DNA 共 582 bp,內含 pUC8 之多重選殖 位(Multiple cloning site),Nopaline synthase (nos) poly A signal。骨架上含胺基糖類 磷酸轉移酶(Aminoglycoside phosphotransferase)為卡那黴素 (Kanamycin)抗性基 因,做為細菌篩選之標記。
2.2 pCAMBIA1302
pCAMBIA1302 (CAMBIA, Australia)全長約 10.5 kb,為雙偶型載體,由長度各 25 bp 之 Left border 與 Right border 夾帶 T-DNA 共 4.3 kb,內含兩段重複的 CaMV 35S 啟動子序列啟動之潮黴素磷酸轉移酶,做為真菌抗藥性篩選之用;一段 CaMV 35S 啟動子啟動之 MGFP,其 3’端帶有 6 個 histidine 以及 nos poly A signal。
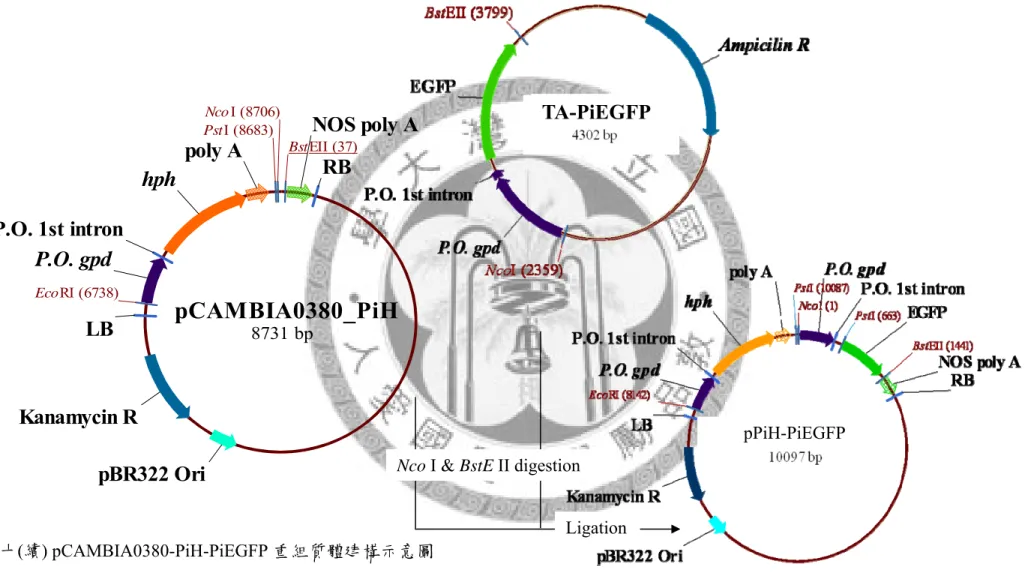
2.3 pCAMBIA0380-PiH
pCAMBIA0380-PiH 全長約 8.7 kb,以 pCAMBIA0380 作為骨架,其 T-DNA 之 MCS 近 LB 端置入鮑魚菇同源性 GPD 啟動子帶有第一個內含子啟動之 hph 抗藥性 基因。
2.4 pPiH-PiEGFP
pPiH-PiEGFP 全長約 10.1 kb,為 pCAMBIA0380-PiH 近 RB 端置入鮑魚菇同源 性 GPD 啟動子帶有第一個內含子啟動之 EGFP 報導基因。
2.5 pPiH-PiEGFP-SP
pPiH-PiEGFP-SP 全長約 10.1 kb,為 pCAMBIA0380-PiH 近 RB 端置入鮑魚菇同 源性 GPD 啟動子帶有第一個內含子啟動之 EGFP 報導基因,其 3’端帶有內質網停 留訊息胜肽HDEL。
29
2.6 pAiH-AiEGFP
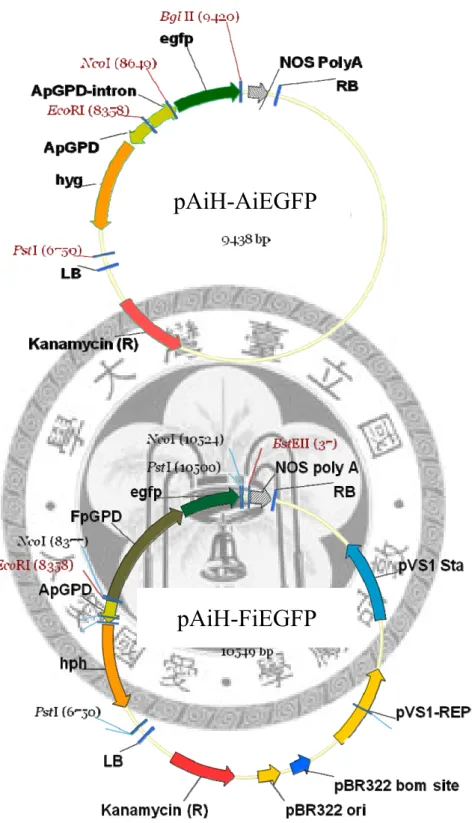
pAiH-AiEGFP 全長約 9.4 kb,以 pCAMBIA0390 作為骨架,其 T-DNA 之 MCS 近LB 端為洋菇 GPD 啟動子帶有第一個內含子啟動 hph 抗藥性基因,近 RB 端則 為洋菇 GPD 啟動子帶有第一個內含子啟動之 EGFP 報導基因
2.7 pAiH-FiEGFP
pAiH-FiEGFP 全長約 10.5 kb,以 pCAMBIA0390 作為骨架,其 T-DNA 之 MCS 近LB 端為洋菇 GPD 啟動子帶有第一個內含子啟動 hph 抗藥性基因,近 RB 端則 為金針菇 GPD 啟動子帶有第一個內含子啟動之 EGFP 報導基因。
上述質體之基因圖譜如圖九所示
3. 引子
本研究中所使之引子為委託生技公司合成 (禾鑫生技,台北),序列整理於表六;
使用之菌株整理如表七。
30
圖八 實驗室無菌栽培之鮑魚菇子實體。
上圖:玻璃燒杯內子實體生長情形;下圖:子實體傘面(左)與蕈褶面(右) Figure 8. Sterile fruiting bodies of P. ostreatus cultivated in laboratory.
31
pCAMBIA1302
10549 bp
MGFP Nos poly A RB
pBR322 Ori Kanamycin R
LB hph CaMV 35S(2)
CaMV 35S
CaMV 3' UTR
pCAMBIA0380
6812 bp
NOS poly A RB
Kanamycin R
pBR322 Ori
LB
MCS (pUC8)
BamHI (6748)
Eco RI (6738)
HindIII (6768) Nco I (6787) Pst I (6764)
Sma I (6745) XmaI (6743)
Bst EII (37)
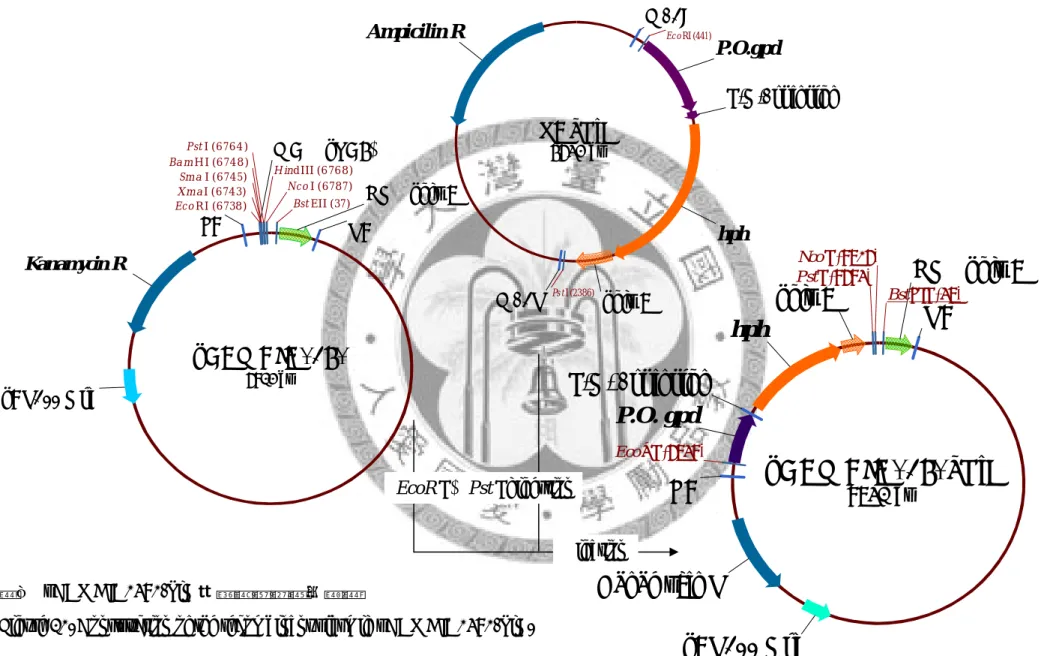
圖九 質體 pCAMBIA0380 與 pCAMBIA 1302 之限制酶切位與特定基因圖譜 Figure 9. Restriction sites and specific gene map of plasmid pCAMBIA0380 and pCAMBIA 1302.
32
pCAMBIA0380_PiH
8731 bp
NOS poly A RB
Kanamycin R
pBR322 Ori LB
P.O. gpd P.O. 1st intron
hph
poly A
Nco I (8706) PstI (8683)
EcoRI (6738)
BstEII (37)
圖九(續) 質體 pCAMBIA0380-PiH 與 pPiH-PiEGFP/SP 之限制酶切位與特定基 因圖譜
Figure 9.(Continued) Restriction sites and specific gene map of plasmid pCAMBIA0380-PiH and pPiH-PiEGFP (w/o signal peptide).
pPiH-PiEGFP-SP
pPiH-PiEGFP
33
圖九(續) 質體 pCAMBIA0390-AH-AiEGFP 與 pCAMBIA0390-AH-FiEGFP 之限 制酶與基因圖譜。本圖源自徐, 2008。
Figure 8. (Continued) Restriction sites and specific gene map of plasmid
pCAMBIA0390-AiH-AiEGFP and pCAMBIA0390-AiH-FiEGFP. Originated from Hsu, 2008)
pAiH-AiEGFP
pAiH-FiEGFP
34
表六 本研究使用之核酸引子序列 Table 6. Primers used in this study
Primer Sequence (5’Æ 3’)a Source
PO GPD i-F CCAGTTAGTGGAAATCAATGGCAGCCGT This lab
PO GPD i-R CCAAAGGTTGGGACGTTGACCTGAATACG This lab
Hph-f ACTAGTATGAAAAAGCCTGAACTCACC This lab
Hph-r CTGCAGACAACTTAATAACACATTGCG This lab
L GGCCATTACGGCCGGCTGGTGGCAGGATATATTGTGGTG This lab R GGCCGAGGCGGCCAACGCTCTTTTCTCTTAGGTTTACCC This lab
EGFP-F AGGTCAAAGGATCCATGGTGAGCAAGG This lab
EGFP-R GGTCACCTTACTTGTACAGCTCGTCCAT This lab
a引子內之限制酶切位以底線粗體標示,SpeI:ACTAGT;Pst I:CTGCAG;BamH I:GGATCC; BstE II:GGTCACC
35
表七 本研究所使用之菌株 Table 7. Strains used in this study
Strain Characteristics Source
P. ostreatus Dikaryon This lab
P. ostreatus (A1B2) Monokaryon This lab
P. ostreatus (A2B1) Monokaryon This lab
E. coli Top10 For molecular manipulation Invitrogen (Carlsbad, CA)
E. coli DH5α For plasmid conservation Invitrogen (Carlsbad, CA)
A.tumefaciens AGL-1 Disarmed strain, A.tumefaciens C58 as chromosome background with rifampicin and carbenicillin resistence. Harboring Ti plasmid pTiBo542ᇞT-DNA without marker gene.
A.tumefaciens LBA4404 Disarmed strain, A.tumefaciens Ach5 as chromosome background with rifampicin resistence. Harboring pAL4404 with spectinomycin and streptomycin resistence.
36
二、實驗方法
1. 重組質體建構
1.1 鮑魚菇啟動子序列確認
將實驗室保存之 TA-PiH 質體上鮑魚菇 GPD 啟動子及第一個內含子序列進行核 酸定序 (源資,台北),序列如圖十四。
1.2 質體抽取與限制酶截切
將帶有目標質體之大腸桿菌皆種於3 ml LB 培養(TA 為骨架者添加
ampicilin 最終濃度為 100 μg/ml,pCAMBIA 為骨架者則添加 kanamycin 最終濃度 為50μg/ml ),37oC 下 250 rpm 震盪培養 14~16 小時,離心收取菌體後,以 Plasmid Extraction Kit (Viogene,台北) 抽取細菌質體。
抽出之質體 TA-PiH、pCAMBIA0380 分別以酵素 Pst I 與 EcoR I 截切,
TA-PiEGFP、TA-PiEGFP-SP、pCAMBIA0380-PiH 分別以 BstE II 與 Nco I 截切,反 應組成與條件見表八。
表八 限制酶反應之酵素比例與反應條件
Table 8. The restriction enzyme ratio and reaction conditon
Restriction enzymes Percentage (v/v) Reaction time (hour)b
EcoR I 2 1
Pst I 2 1.5
BstE II 2 1
Nco Ia 2 0.6
a Digest first when pairing with BstE II. b Temperature used refer to New England Biolabs products information. All in buffer 3 system . The DNA volume is 25 ~ 30μl and total volume is 50μl.
37
1.3 洋菜膠體電泳與 DNA 回收
將抽取之質體或截切後之產物與 6x 追蹤染體(0.25% bromophenol blue, 0.25%
xylene cyanol, 0.1 M EDTA, 50% glycerol)混合後,以 1%洋菜膠體進行電泳,於 0.5x TAE buffer (40 mM Tris-acetate, 1mM EDTA, pH8.0)以電壓 140 伏特進行電泳 20 分 鐘。經ethidium bromide 染色 10 ~ 15 分鐘後,在 UV box 上照相、觀察結果,以 QIAquick Gel Extraction Kit (Qiagene, Valencia,CA)純化目標片段。
1.4 質體接合與轉形
將洋菜膠體純化出的目標片段再經洋菜膠體電泳後,確認其產量與大小,估算 莫耳數比vector:insert = 3:1 或 4:1,各加入所需體積與 1μl T4 接合酶、10x 緩 衝液A 、10 x 緩衝液 B (益生,台北),加入無菌水至最終體積為 10 ~ 20 μl。於 4
oC 反應隔夜後轉形至大腸桿菌 Top 10 或 DH5α 菌株,於含有 50 μg/ml kanamycin 之LB plate 培養 16 ~ 18 小時。各質體建構流程如圖十所示。
1.5 大腸桿菌轉形株篩選
以牙籤挑取生長於 Kanamycin 選擇性培養基上的菌落,加入專一性引子與試劑 進行菌落聚合酶連鎖反應(Colony PCR),PCR 反應條件見表九。反應完成後以洋菜 膠體電泳分析,確認目標產物大小無誤後,抽取質體進行限制酶截切確認並將正 確質體進行核酸定序 (源資,台北)。
38
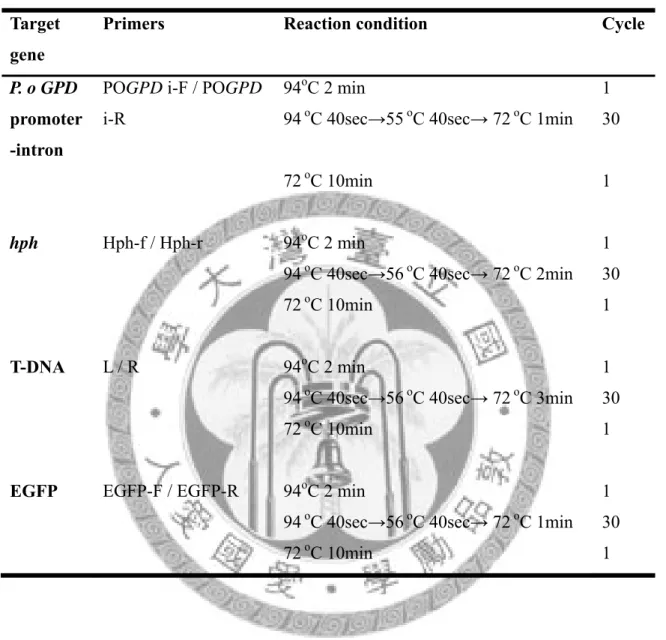
表九 聚合酶連鎖反應之條件
Table 9. Polymerase chain reaction condition used.
Target gene
Primers Reaction condition Cycle
P. o GPD promoter -intron
POGPD i-F / POGPD i-R
94oC 2 min
94 oC 40sec→55 oC 40sec→ 72 oC 1min 1 30
72oC 10min 1
hph Hph-f / Hph-r 94oC 2 min
94 oC 40sec→56 oC 40sec→ 72 oC 2min 1 30
72oC 10min 1
T-DNA L / R 94oC 2 min
94 oC 40sec→56 oC 40sec→ 72 oC 3min 1 30
72oC 10min 1
EGFP EGFP-F / EGFP-R 94oC 2 min
94 oC 40sec→56 oC 40sec→ 72 oC 1min 1 30
72oC 10min 1