國立臺東大學生命科學系 碩士論文
Department of Life Science National Taitung University
Master Thesis (Draft)
指導教授:林志輝 博士 Advisor: Chih-Hui Lin, Ph.D.
臺東地區溫泉嗜熱/鹽菌酵素活性篩選與 Anoxybacillus flavithermus HY-TTH-D23 熱穩定半乳糖苷酶特性分析
Enzyme activity screening of thermophilic/halophilic bacteria in Taitung hot spring and characterization of Anoxybacillus flavithermus HY-TTH-D23 thermostable α-galactosidase
研究生:施政嘉
Graduate student: Cheng-Chia Shih
中華民國 104 年 7 月
July 2015
摘要
嗜熱性微生物為了適應高溫環境常會發展出獨特的生理特性,其中熱穩定性 的酵素系統是細胞生存所必須的。目前已有許多嗜熱菌生產的耐熱酵素應用在工 業製程上,包含食品、澱粉加工和藥物開發等領域。嗜熱性微生物偏好生存在溫 熱的環境,故目前許多相關的研究皆是以溫泉為主要對象。臺東地區溫泉自然資 源豐富,溫泉露頭數量眾多,且大多開發程度不高,是理想的嗜熱/鹽菌來源。本 研究針對臺東地區的五個樣區進行微生物調查,樣區包含綠島朝日溫泉、金崙溫 泉、紅葉溫泉、彩霞溫泉與綠島柚子湖潮間帶。本研究共鑑定出 110 株分離菌 株,其中有 12 株為可能的新種/屬。結果顯示高溫菌分離株 (T 組) 多為 Thermus 屬 與 Geobacillus 屬 , 而 耐 熱 菌 (TTH 組 ) 分 離 株 則 是 主 要 為 Anoxybacillus 屬,嗜鹽高溫菌組 (HT 組) 菌株則幾乎都是 Bacillus 屬,嗜鹽常 溫菌組 (HM 組) 菌株則無明顯的主要菌種。菌株酵素活性篩選的結果顯示高溫 菌組 (T 組) 具有較多種類的酵素活性,嗜鹽常溫菌組 (HM 組) 則是蛋白酶的 平均活性較強。綜觀所有菌株,Anoxybacillus flavithermus HY-TTH-D23 在 leucine arylamidase、α-chymotrypsin、α-galactosidase、α-glucosidase 與 β-glucosidase 的 活性皆為本研究中受測菌株最高者,且生長溫度可達 75oC,因此被選為酵素選 殖的對象。本研究成功從 A. flavithermus HY-TTH-D23 選殖出 α-galactosidase (AFGAL) 的基因並於 E. coli BL21 (DE3) 中進行異源表現。菌體經由超音波破 菌與 55oC 熱處理後,使用 HisTrap HP column 能有效純化重組 AFGAL。純化 後 rAFGAL 比活性為 15.2 U/mg,純化倍率和產率分別為 36.6 倍與 75.7 %。
rAFGAL 的分子量為 112 kDa,最適溫度為 55oC,經過 2 hr 後活性會提升至初 始的 164%,活性 pH 值範圍為 5-10,最適 pH 為 8-9。rAFGAL 為目前文獻 中第一個從 Anoxybacillus spp. 中選殖並異源表現生產的 α-galactosidase。與文 獻中異源表現的 Bacillaceae 科熱穩定 α-galactosidase 相較,具有最廣泛的 pH 活性範圍和最高的最適 pH,並且純化產率最高。本研究結果除了希望能對 Anoxybacillus spp. 的 α-galactosidase 基本特性有所描述與貢獻外,未來還能將 其應用在食品、製糖與飼料等高溫工業製程。
關鍵字:嗜熱菌、溫泉、Anoxybacillus flavithermus、α-galactosidase
Abstract
Thermophiles have evolved distinct characteristic such as thermozyme system to adapt high temperature environment. There are many thermozymes from thermophiles have applied to industrial processes such as food, starch processing and drug development. The major habitat of thermophiles are geothermal environment. There are several undisturbed hot-springs which are ideal thermophile and halophile study sites in Taitung. In the study, five study sites including JhaoRih, JinLun, HongYe, ChiXia and YouZaiHu were investigated. One hundred and ten of bacteria strains were isolated and identified in our study. The results of 16S rRNA gene sequence analysis showed that at least 12 new species of bacteria were isolated. Thermus sp. and Geobacillus sp. were the main species in T group. Anoxybacillus sp. was the main species in TTH group. Bacillus sp. was the main species in HM group. None of species of bacteria was dominate in HT group. The result of API ZYM® analysis showed that the thermophiles have more diverse enzyme activities than halophiles. Enzyme activity of lipase, phosphatase and glycosidase were common in thermophilic isolates, while protease was the most common enzyme in halophilic isolates. Among the bacteria isolates, the thermophilic Anoxybacillus flavithermus HY-TTH-D23 showed multiple promising enzyme activities includes leucine arylamidase, α-chymotrypsi, α- galactosidase, α-glucosidase and β-glucosidase. Besides, the highest growth temperature of A. flavithermus HY-TTH-D23 could up to 75oC.Thus the A. flavithermus HY-TTH-D23 was selected as our target for enzyme cloning. The gene encoding α- galactosidase of A. flavithermus HY-TTH-D23 (AFGAL) were successfully cloned and expressed in E. coli BL21 (DE3). After sonication and heat treatment, the rAFGAL was effectively purified from E. coli cells with a 36.6-fold purification and a final yield of 75.7% through HisTrap HP column. The specific activity against pNPG was 15.2 U/mg.
The rAFGAL had a molecular weight of 112 kDa, and remain active at pH 5-10 with optimum pH of 8-9. The rAFGAL was the first Anoxybacillus α-galactosidase that cloned and heterologous expressed in E. coli. In comparison with thermostable α- galactosidase cloned from Bacillaceae, rAFGAL had the most borad pH range, highest optimalpH and best purification yield. The results of this study could contribute to the knowledge of thermostable Anoxybacillus α-galactosidase and its application for industrial processes in foods, sugar and fodder industry.
縮寫表
縮寫 全名 中文
BSA Bovine serum albumin 牛血清白蛋白
BLAST Basic local alignment search tool 基礎本域比對搜尋工具
DNA Deoxynucleic acid 去氧核醣核酸
DTT Dithiothreitol 二硫蘚糖醇
EDTA Ethylenediaminetetraacetic acid 乙二胺四醋酸
HM Halomesophile 嗜鹽常溫菌組
HT Halothermophile 嗜鹽高溫菌組
IPTG Isopropylthio-β-galactoside 異丙基-β-D-硫代半乳糖苷
LB Luria broth 盧亞氏培養基
NCBI National center for biotechnology
information 國家生物技術資訊中心
NB Nutrient broth 營養肉湯培養基
PCR Polymerase chain reaction 聚合酶鏈鎖反應
PMSF Phenylmethanesulfonylfluoride 苯甲硫醯氟 pETAFGAL pET GFP vector include
α-galactosidase of A. flavithermus
帶有 AFGAL 基因的 pET GFP 載體
pETAFST pET GFP vector include subtilisin-type protease of A. flavithermus
帶有 AFST 基因的 pET GFP 載體
pNP p-nitrophenol 對硝基苯酚
pNPG p-Nitrophenyl-α-D-galactopyranoside 4-硝基苯-α-D-吡喃半乳糖苷 rAFGAL Recombinant α-galactosidase of
A. flavithermus HY-TTH-D23
重組 A. flavithermus HY- TTH-D23 α-半乳糖苷酶 rAFST Recombinant subtilisin-type protease
of A. flavithermus HY-TTH-D23
重組 A. flavithermus HY- TTH-D23 蛋白酶
SDS Sodium dodecyl sulfate 十二烷基硫酸鈉
SDS-PAGE Hodium dodecyl sulfate
polyacrylamide gel electrophoresis
十二烷基硫酸鈉聚丙烯酰胺 凝膠電泳
T Thermophile 高溫菌組
TTH Temperature tolerant-high 耐熱菌組
目錄
摘要 ... I Abstract ... II 縮寫表 ... III 目錄 ... IV 圖目錄 ... VI 表目錄 ... VIII
一、前言 ... 1
(一) 文獻回顧 ... 1
1. 嗜極端性生物 (extremophile) ... 1
2. 嗜熱性生物 (thermophile) ... 4
3. 臺灣的地熱環境與嗜熱菌的研究 ... 14
4. Anoxybacillus spp. 的發展與應用 ... 17
5. α-半乳糖苷酶 (α-galactosidase) 的種類與應用 ... 31
(二)、研究動機與實驗架構 ... 35
1. 研究動機與目的 ... 35
2. 實驗架構 ... 35
二、材料與方法 ... 37
(一) 菌株、藥品與試劑 ... 37
(二) 儀器與設備 ... 38
(三) 實驗方法 ... 40
1. 臺東地區溫泉菌株之篩選 ... 40
2. A. flavithermus HY-TTH-D23 酵素基因選殖 ... 55
3. α-galactosidase 異源表現與特性分析 ... 62
三、結果與討論 ... 66
(一) 菌株分離與鑑定 ... 66
2. 菌種初步鑑定 ... 73
(二) 酵素活性篩選 ... 80
1. API ZYM 酵素活性篩選 ... 80
2. API 50 CHB 碳源利用分析... 84
3. Anoxybacillus sp. HY-TTH-D23 的鑑定與生化特性分析 ... 92
(三) A. flavithermus HY-TTH-D23 熱穩定酵素的基因選殖與異源表現 ... 95
1. 熱穩定酵素的基因選殖 ... 95
2. 大腸桿菌表現載體的構築 ... 95
3. 熱穩定酵素的異源表現 ... 102
(四) rAFGAL 酵素特性分析... 113
1. rAFGAL 的表現與純化... 113
2. 溫度對酵素穩定度與活性的影響 ... 117
3. pH 值對酵素活性的影響 ... 119
4. 金屬離子對酵素活性的影響 ... 119
5. 其它化學試劑對酵素活性的影響 ... 119
6. 界面活性劑對酵素活性的影響 ... 123
7. rAFGAL 三級結構的預測... 123
四、結論 ... 126
五、參考文獻 ... 127
圖目錄
圖一、嗜冷性、嗜中溫性、嗜熱性與嗜超熱性微生物生存溫度關係圖。 ... 5
圖二、高溫原核生物最佳生長溫度 (Topt) 與最佳生長酸鹼度 (pHopt) 分佈圖。 7 圖三、臺灣溫泉地熱區與地質分佈圖。 ... 15
圖四、Bacillaceae 科的親緣關係樹。... 20
圖五、根據串聯 361 條同源基因的序列所建立的 Bacillaceae 親源性關係圖。 ... 22
圖六、以親源性關係樹比較 Anoxybacillus、Geobacillus 與 Bacillus 選殖出的酵 素。 ... 24
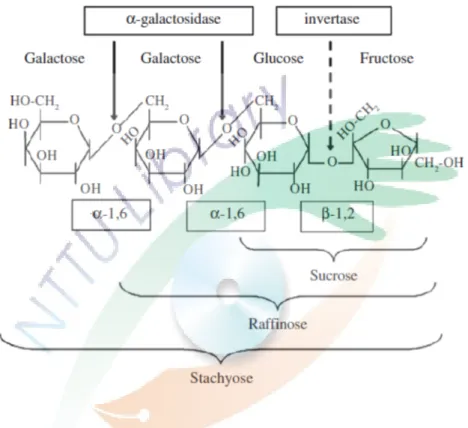
圖七、棉子糖和水蘇糖的 α-半乳糖苷寡構形與酵素催化水解的示意圖。 ... 32
圖八、本論文之研究架構。 ... 36
圖九、綠島朝日溫泉採樣點。 ... 41
圖十、金崙溫泉採樣點。 ... 43
圖十一、紅葉溫泉採樣點。 ... 46
圖十二、彩霞溫泉採樣點。 ... 48
圖十三、綠島柚子湖潮間帶採樣路線與地點。 ... 50
圖十四、pET GFP LIC 的選殖載體 (2GFP-T)。 ... 60
圖十五、各樣點中微生物有顯著生長的最高溫度/鹽度 ... 72
圖十六、本研究中微生物分離株數量統整。 ... 81
圖十七、本研究分離株種類及數量分布。 ... 82
圖十八、A. flavithermus HY-TTH-D23 的 API ZYM 試劑條分析結果。 ... 83
圖十九、ZYM 分析結果統整。... 90
圖二十、Anoxybacillus sp. D23 的細胞外觀。 ... 93
圖二十一、A. flavithermus HY-TTH-D23 酵素基因 PCR 選殖。 ... 96
圖二十二、A. flavithermus HY-TTH-D23 的 subtilisin-type proteinase 的親緣關係 樹。 ... 97
圖二十三、A. flavithermus HY-TTH-D23 的 alpha-galactosidase 的親緣關係樹。 ... 98
圖二十四、pET GFP LIC cloning vector 的 SspI 限制酶切。 ... 99
圖二十五、限制酶 SspI 的反應時間最佳化。... 100
圖二十六、A. flavithermus HY-TTH-D23 subtilisin-type proteinase 與 alpha- galactosidase 的 LIC cloning PCR。 ... 101
圖二十七、載體 pETAFST。 ... 103
圖二十八、載體 pETAFGAL。... 104
圖二十九、表現載體構築篩選。 ... 105 圖三十、E. coli BL21 (DE3) pETAFGAL 與 E. coli BL21 (DE3) pETAFST 菌體的
圖三十一、E. coli BL21 (DE3) pETAFST 與 E. coli BL21 (DE3) pETAFGAL 在 1
mM IPTG 誘導下的生長曲線。 ... 107
圖三十二、E. coli BL21 (DE3) pETAFST 與 E. coli BL21 (DE3) pETAFGAL 菌體 的綠色螢光曲線。 ... 108
圖三十三、E. coli BL21 (DE3) pETAFST 與 E. coli BL21 (DE3) pETAFGAL 在 IPTG 誘導後第 5 與第 7 小時的菌體外觀。 ... 109
圖三十四、E. coli BL21 (DE3) pETAFST 誘導過程的總蛋白電泳分析。 ... 110
圖三十五、E. coli BL21 (DE3) pETAFGAL 誘導過程的總蛋白電泳分析。 ... 111
圖三十六、重組 rAFST 與 rAFGAL 的 native-PAGE 螢光影像。 ... 112
圖三十七、rAFGAL 純化過程的 SDS-PAGE 分析。 ... 114
圖三十八、存放不同時間的 rAFGAL SDS-PAGE 分析結果。 ... 115
圖三十九、溫度對 rAFGAL 與咖啡豆的 α-galactosidase 的穩定性及活性的影 響。 ... 118
圖四十、pH 值對 rAFGAL 酵素活性的影響。 ... 120
圖四十一、金屬離子對 rAFGAL 酵素活性的影響。 ... 121
圖四十二、其它試劑對 rAFGAL 酵素活性的影響。 ... 122
圖四十三、介面活性劑對 rAFGAL 酵素活性的影響。 ... 124
圖四十四、以 Swiss-model 所預測 rAFGAL 的三級結構。 ... 125
表目錄
表一、已知對各種極端環因子耐受度最高的極端生物 ... 2
表二、生活於複數極端因子環境的嗜複極端因子性微生物 ... 3
表三、嗜熱菌全細胞的生物技術應用 ... 9
表四、嗜熱菌產生的高分子於已知或潛在的生物技術應用 ... 10
表五、 Bacillaceae 科的菌屬 ... 19
表六、文獻中 Anoxybacillus 物種的基本特性 ... 21
表七、由不同 Anoxybacillus 物種所生產的酵素生化特性 ... 23
表八、Anoxybacillus 的 glycosyl hydrolases (GHs) 與其相似物種的列表 ... 28
表九、綠島朝日溫泉 (JR) 採樣點特性 ... 42
表十、金崙溫泉 (JL) 採樣點特性 ... 44
表十一、紅葉溫泉 (HY) 採樣點特性 ... 47
表十二、彩霞溫泉 (CX) 採樣點特性 ... 49
表十三、綠島柚子湖潮間帶 (YZH) 採樣點特性 ... 51
表十四、API 50 CH 試劑組之碳水化合物基質成份 ... 53
表十五、API ZYM 酵素活性測試試劑組可檢驗的酵素種類 ... 54
表十六、目標酵素基因選殖用引子對 ... 57
表十七、預估的目標酵素基因片段大小 ... 58
表十八、LIC cloning 引子對 ... 61
表十九、各樣品之高溫菌組 (T) 培養生長情形 ... 67
表二十、各溫泉耐熱菌組 (TTH) 循環溫度之液態培養 ... 68
表二十一、各溫泉嗜鹽高溫菌組 (HT) 於液態培養之耐鹽程度 ... 70
表二十二、各溫泉嗜鹽常溫菌組 (HM) 於液態培養之耐鹽程度 ... 71
表二十三、高溫菌組 (T) 菌株之 16S rRNA 基因序列比對結果 ... 74
表二十四、各溫泉耐熱菌組 (TTH) 菌株之 16S rRNA 基因序列比對結果 ... 75
表二十五、各溫泉嗜鹽高溫菌組 (HT) 菌株之 16S rRNA 基因序列比對結果 76 表二十六、各溫泉嗜鹽常溫菌組 (HM) 菌株之 16S rRNA 基因序列比對結果 ... 77
表二十七、本研究中經由初步鑑定後可能的新細菌物種 ... 78
表二十八、部分高溫菌 (T) 分離株的 API ZYM 酵素活性測試結果 ... 85
表二十九、部分耐熱菌 (TTH) 分離株的 API ZYM 酵素活性測試結果 ... 86
表三十、部分耐嗜鹽高溫菌 (HT) 分離株的 API ZYM 酵素活性測試結果 .... 87
表三十一、部分耐嗜鹽高溫菌 (HM) 分離株的 API ZYM 酵素活性測試結果 ... 88
表三十二、酵素活性最強的分離株 ... 89
表三十三、部分分離株的 API 50 CHB 測試結果 ... 91
表三十五、rAFGAL 純化表 ... 116
一、前言
(一) 文獻回顧
1. 嗜極端性生物 (extremophile)
嗜極端性生物是指生活在一般生物不能正常生存、繁衍環境中的生物 (Rothschild and Mancinelli, 2001)。自 1960 年代首次發現可於 60°C 以上生 存的生物後 (Brock and Freeze, 1969),許多生存於特殊環境的生物被逐漸分 離出來,嗜極端性生物的概念開始被建立 (MacElroy, 1974),生命生存的極 限條件也一再被更新 (表一)。嗜極端性生物通常是以其所生存的極端環境 因子為描述依據,如嗜熱性生物 (thermophile)、嗜冷性生物 (psychrophile)、
嗜 酸 性 生 物 (acidophile) 、 嗜 鹼 性 生 物 (alkaliphile) 、 嗜 壓 性 生 物 (barophile/piezophile)、嗜鹽性生物 (halophile) 以及由其他特殊環境分離出 的生物。然而這樣的區分方式並不完全,因許多極端環境往往存在複數的限 制因子,如表二所示。因此由這些環境分離出來的生物,往往對多個極端環 境 具 有 抗 性 , 對 此 Rothschild and Mancinelli (2001) 提 出 了
「polyextremophilic organism」的概念。Canganella and Wiegel (2011) 也對嗜 極端性生物的分類提出 true (obligate) 與 facultative extremophile 的概念。
前者為只能生存於特殊環境,後者可生存於一般環境,但對於特殊環境亦有 耐受性。近年來,嗜極端性生物的研究包括 (1) 嗜極端性生物的起源: 是屬 於較古老的族群,或是經過二次演化適應的結果;(2) 於工業製程的利用性 ; (3) 地球生命起源的研究 ; (4) 對特殊環境的適應性與抗性 (Canganella and Wiegel, 2011)。
表一、已知對各種極端環因子耐受度最高的極端生物 (Rothschild and Mancinelli, 2001) Table 1. “Record-holding” extremophiles and their environmental limits
Environmental factor Organism Habitat Phylogenetic
affiliation Tolerance to stress
Temperature
High Pyrolobus
fumarii
Hot undersea hydrothermal
vents
Crenarchaeota
Maximum 113°C, Optimum 106°C,
Minimum 90°C Low Polaromonas
vacuolata Sea-ice Bacteria
Minimum 0°C, Optimum 4°C, Maximum 12°C
pH
Low Picrophilus oshimae
Acidic hot
springs Euryarchaeota
Minimum pH -0.06, Optimum pH 0.7,
Maximum pH 4 (is also thermophilic)
High Natronobacteriu
m gregoryi Soda lakes Euryarchaeota
Maximum pH 12, Optimum pH 10, Minimum pH 8.5 (is also halophilic) Hydrostatic
pressure Strain MT41 Mariana Trench Bacteria
Maximum >100 MPa, Optimum 70 MPa, Minimum 50 Mpa
Salt concentration Halobacterium salinarum
Salt lakes, salted
hides, salted fish Euryarchaeota
Maximum NaCl saturation, Optimum 250 g /L salt, Minimum 150
g/L salt
Ultraviolet and ionizing radiation
Deinococcus radiodurans
Isolated from ground meat, radioactive waste,
nasal secretion of elephant; true habitats unknown
Bacteria
Resistant to 1.5 kGy gamma radiation and
to 1500 J/m2 of ultraviolet radiation
表二、生活於複數極端因子環境的嗜複極端因子性微生物 (Rothschild and Mancinelli, 2001) Table 2. Some examples of "polyextremophilic" microorganisms, adapted to life at a combination of
several environmental extremes Environmental
factors Organism Habitat Phylogenetic
affiliation Tolerance to stress
Low pH, high temperature
Picrophilus
oshimae Acidic hot springs Euryarchaeota
Minimum pH -0.06;
Maximum temperature 65°C
Sulfurisphaera Acid hot spring Crenarchaeota Maximum temperature 92°C; Minimum pH 1
Cyanidium
caldarium Acidic hot springs Eukarya
Maximum temperature 57°C; Minimum pH
0.2 High pH, high salt
concentration
Natronobacterium
gregoryi Soda lakes Archaea - Euryarchaeota
Maximum pH 12;
NaCl saturation High temperature,
high pH
Thermococcus alcaliphilus
Shallow marine hydrothermal
springs
Archaea - Euryarchaeota
Maximum temperature 90°C; Maximum pH
10.5
High temperature, high pressure
Thermococcus barophilus
Thermal vent, Mid-Atlantic
Ridge
Archaea - Euryarchaeota
Maximum temperature 100°C; Requires 15- 17.5 MPa pressure at
the highest temperatures
High temperature, high radiation
Deinococcus
geothermalis Thermal springs Bacteria
Maximum temperature 50°C. Resistant to at least 10 kGy gamma
radiation Low temperature,
high pressure Deep-sea bacteria Deep sea
environments Bacteria
Live at 2-4°C and pressures of 50-110
Mpa
2. 嗜熱性生物 (thermophile) (1) 嗜熱性生物的定義
自嗜熱性生物被發現後,一般認為能以最適生長溫度做為標準,生物依 照生存溫度可分為數個區段 (圖一),其中最適生存溫度高於 45-50°C 者即 屬於嗜熱性生物。Stetter (1996) 於深海中高溫高壓環境的熱泉噴口 (deep- sea hydrothermal vents 或 deep-sea black smoker),發現可於生長於 113 °C的 Pyrolobus fumarii。之後 Stetter (1996) 將嗜熱性生物依照最適生長溫度 (Topt) 重新界定為超嗜熱性生物 (hyperthermophiles, Topt > or = 80°C),極嗜熱性生 物 extreme thermophiles, Topt 70°C to 80°C),與中度嗜熱性生物 (moderate thermophiles, Topt 45°C to 70°C),此定義一直被延用至今 (Stetter, 1996 ; Mesbah and Wiegel, 2008 ; Wagner and Wiegel, 2008)。
(2) 嗜熱性生物的生存環境
嗜熱性生物 (主要為嗜熱性微生物) 群落的存在環境為各種高溫環境。
這些高溫環境大多與火山活動有關,包括溫泉、間歇泉、火山和深海中的熱 泉噴口,且已成為主要的嗜熱菌的主要分離源。此外,人類活動產生的高溫 環境,如堆肥、煤灰處理場 (coal refuse piles)、工業廢水處理系統、造紙或 食品工業、熱水器及其冷卻水等也發現有嗜熱菌的存在 (Oshima and Moriya, 2008)。甚至在地殼深層也發現有嗜熱菌 (Gold, 1992)。不過除了熱源環境外,
Marchant 等人也於愛爾蘭地區的低溫或常溫土壤中,分離出至少包含兩個 種共 53 株的 Geobacillus 屬嗜熱菌 (Marchant et al., 2002)。另外許多天然 地熱環境也存在多極端因子,因此許多嗜熱性微生物常常也是嗜複極端因子 性生物 (polyextremophilic organism)。
(3) 嗜熱性生物的種類與分布
細菌、真菌、藻類以及古生菌都可能是嗜熱性微生物,其中以原核生物
圖一、嗜冷性、嗜中溫性、嗜熱性與嗜超熱性微生物生存溫度關係圖。(Madigan and Marrs, 1997)
Figure 1. Relation of temperature and growth rates for a typical psychrophilic, mesophilic, thermophilic and hyperthermophilic microorganisms.
The respective optimal growth temperatures Topt are indicated on the graph.
界的細菌以及古生菌的種類最為龐大。目前發現以下細菌屬中,均為嗜熱菌 的有:Desulfotomaculum (Campbell & Postgate, 1965)、Thermus (Brock & Freeze, 1969)、Chloroflexus (Pierson & Castenholz, 1974)、Thermothrix (Caldwell et al., 1976)、Sulfobacillus (Golovacheva & Karavaiko, 1978)、Meiothermus (Nobre et al., 1996a )、Thermotoga (Robert et al., 1986)、Rubrobacter (Suzuki et al., 1988)、
Thermoleophilum (Zarilla & Perry, 1984)、Sphaerobacter (Demharter et al., 1989)、
Rhodothermus (Alfredsson et al., 1988)、Thermosipho (Huber et al., 1989)、
Hydrogenophilus (Hayashi et al., 1999)、Anoxybacillus (Pikuta et al., 2000)、
Geobacillus (Nazina et al., 2001)、Caldimonas (Takeda et al., 2002)、Roseiflexus (Hanada et al., 2002) 與 Silanimonas (Lee et al., 2005) 等。此外有些菌屬中包 含 嗜 高 溫 及 嗜 中 溫 的 物 種 , 例 如 Deinococcus (Brooks & Murray, 1981;
Ferreira et al., 1997)、Bacillus (Fritze et al., 1990)、Marinobacter (Gauthier et al., 1992;Shieh et al., 2003)、Pseudoxanthomonas (Chen et al., 2002a; Finkmann et al., 2000) 與 Thermomonas (Busse et al., 2002) 等屬,可見地球上嗜熱性微生 物種類豐富,且有許多不同菌屬或是菌種的新穎嗜熱菌不斷地被發現 (魏,
2006)。
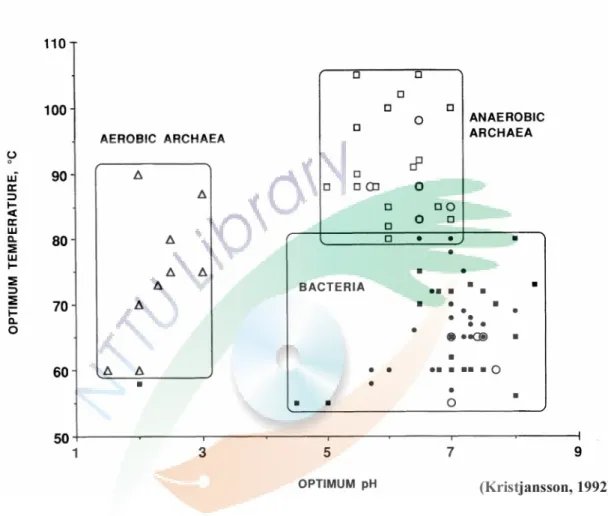
在高溫的環境中,溫度與酸鹼度是影響嗜熱菌分佈的最主要因子。好氧 產胞的桿菌群如 Bacillus spp.、Geobacillus spp.、Alicyclobacillus spp. 和 Sulfobacillus spp. 等可以存活於 pH 2-9 的高溫環境。好氧非產胞桿菌如 Thermus spp.、Meiothermus spp. 和 Rubrobacter spp. 等,則多分佈於中性或 鹼性的環境中 (Pask-Hughes & Williams, 1975;Brock & Boylen, 1973)。而厭 氧 性 嗜 熱 菌 如 Clostridium spp. 則 在 非 強 酸 性 的 環 境 中 廣 泛 地 分 佈 (Kristjansson and Stetter, 1992)。圖二為目前所知高溫原核生物其最佳生長溫 度與最佳生長酸鹼度分佈,好氧性的嗜熱古生菌多分佈於 pH 1-3,60-90 oC 的區間,厭氧性的嗜熱性古生菌則普遍存於 pH 5-7,80-110 oC 的區間,嗜 熱性細菌則存於 pH 4-9,55-80 oC 的環境中 (Kristjansson and Stetter, 1992)。
在環境中,地熱區的溫泉由於分佈於地表且為一個熱穩定性的環境,且其酸 鹼度與溫度呈現多樣化,故目前的許多高溫菌的研究皆以溫泉作為作為對象 (陳,2002)。
圖二、高溫原核生物最佳生長溫度 (Topt) 與最佳生長酸鹼度 (pHopt) 分佈圖。
(Kristjansson, 1992)
Figure 2. The distribution of optimal growth temperature and pH of the high-temp prokaryotes.
Hollow symbol : Archaea, and solid symbol : Bacteria. ○: methogens; △:
aerobic archaea; □: anaerobic archaea; ■: aerobic bacteria; ●: anaerobic bacteria
(4) 嗜熱性生物的生理特性
生存於高溫環境的嗜熱性細菌為了適應,常會發展出獨特的生理特性與 細胞結構以應付惡劣的生存環境,如耐熱的細胞膜、DNA 修補系統及具熱 穩定性的酵素系統等 (Stetter, 1998)。在細菌細胞的主要成份中,脂質、核酸 及蛋白質皆是對熱敏感的物質,因此存於嗜熱性細菌中的這些物質,都有其 獨特的特性以確保其熱穩定性,以 Thermotoga maritima 細胞膜的脂質為例,
主要為 n-fatty acid、diabolic acid 和 glycerol ether lipid (15, 16-dimethyl-30- glyceryloxytriacontanonicacid) 等脂肪酸,而在這些脂肪酸中,ether lipid 在 高溫下具不易被水解的特性,其存在可以增加膜的熱穩定性 (De Rosa et al., 1989)。除脂質的修飾有助於細胞膜的穩定外,嗜熱性細菌的 DNA 多為高 G+C 比例,可以增加 DNA 雙股螺旋的穩定性,加上其具有 reverse gyrase,
可以增加正向的超螺旋以幫助穩定高溫下的 DNA 分子結構 (Kikuchi and Asai, 1984;Bouthier de la Tour et al., 1990)。在蛋白質方面,目前並無法以胺 基酸的組成作為蛋白質是否具熱穩定性的依據,同源的嗜熱菌與中溫菌其蛋 白質的胺基酸組成近似,對於熱的差異性可能來自結構的不同所致,此外熱 休克蛋白質在嗜熱性細菌的耐熱特性上亦具相當的重要性 (Takai et al., 1998)。
(5) 嗜熱性生物的酵素與應用
許多嗜熱菌都是多嗜極性生物,表示他們可以適應其它極端的環境條件,
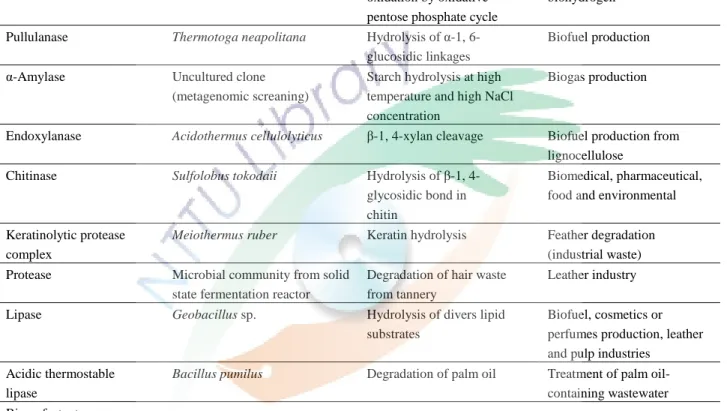
像是 pH、氧化還原電位、鹽度與有毒物質。嗜熱的微生物具有高代謝活性 可提生產率、表現耐熱的酵素與無須冷卻可降低生產成本等優點。嗜熱菌在 生物技術的應用可以分為以及高分子 (macromolecules) 的應用 (如表四與 表五)。
(a) 生物能源 (Bioenergy–biofuel)
由於石化燃料的減少與氣候的改變,尋找能源生產的新來源已經變成 世界各地感興趣的主題,生物燃料可以以固態、液態與氣態的形態存在,
液態生物燃料如生物乙醇、生物柴油、生物丁醇與生物煤油可以藉由發酵 澱粉與線狀纖維素等物質獲得,嗜熱菌可以產生熱穩定的酵素,有效的降
表三、嗜熱菌全細胞的生物技術應用 (Urbieta et. al., 2015) Table 3. Biotechnological applications of whole-cell thermophiles.
Whole cell application Thermophile Action
Biofuel production Caldicellulosiruptor bescii, Caldanaerobius polysaccharolyticus
Xylan degrading activity
Thermoanaerobacterium thermosaccharolyticum
Butanol production
Caldicellulosiruptor saccharolyticus Consolidated bioprocess for anaerobic hydrogen production
Methanoculleus thermophiles and other autochthonous thermophiles from coal mine
Methane production from coal mine substrates
Bioremediation Geobacillus sp.,
Anoxybacillus flavithermus, Thermus thermophiles, Thermococcus zilligii
Biosorption of toxic metals
Thermus scotoductus, Pyrobaculum. islandicum, Thermoanaerobacter sp.,
Thermoterrabacterium ferrireducens
Immobilisation of radionuclides
Aeribacillus sp., Geobacillus sp.
Biodegradation of recalcitrant aromatic
Anoxybacillus sp. Degradation of azo-dyes
Non characterised thermophiles Construction of biofilters for gas deodoration
Bioleaching Sulfobacillus sp., Ferroplasma sp., Acidianus infernus
Enhanced Cu extraction from chalcopyrite
Acidianus brierleyi, Acidianus manzaensis, Metallosphaera sedula, Sulfolobus metallicus
Metal solubilisation from nickel-copper sulphide
Food, animal feed, compost for agriculture, cultivation production
Clostridium sp., Caldicellulosiruptor sp.
Improved compost quality by cellulolytic activity
表四、嗜熱菌產生的高分子於已知或潛在的生物技術應用 (Urbieta et. al., 2015)
Table 4. Recent reports on macromolecules produced by thermophiles with known or potential biotechnological applications.
Macromolecule Thermophile/Source Activity Industry/Bioprocess
Thermozymes
Aldolase Sulfolobus solfataricus, S. acidocaldarius, Thermoproteus texas, Hypertermus butilicus
Stereoselectibe C-C bond formation
Pharmaceutical industry
G6PD, 6PGD, R5PI, others*
Geobacillus stearothermophilus, Moorella thermoacetica, Thermotoga maritima
Glucose oxidation Enzymatic fuel cell
Hydrogenase Pyrococcus furiosus Final stage of glucose oxidation by oxidative pentose phosphate cycle
Enhanced production of biohydrogen
Pullulanase Thermotoga neapolitana Hydrolysis of α-1, 6- glucosidic linkages
Biofuel production
α-Amylase Uncultured clone
(metagenomic screaning)
Starch hydrolysis at high temperature and high NaCl concentration
Biogas production
Endoxylanase Acidothermus cellulolyticus β-1, 4-xylan cleavage Biofuel production from lignocellulose
Chitinase Sulfolobus tokodaii Hydrolysis of β-1, 4-
glycosidic bond in chitin
Biomedical, pharmaceutical, food and environmental
Keratinolytic protease complex
Meiothermus ruber Keratin hydrolysis Feather degradation (industrial waste) Protease Microbial community from solid
state fermentation reactor
Degradation of hair waste from tannery
Leather industry
Lipase Geobacillus sp. Hydrolysis of divers lipid
substrates
Biofuel, cosmetics or perfumes production, leather and pulp industries
Acidic thermostable lipase
Bacillus pumilus Degradation of palm oil Treatment of palm oil- containing wastewater Biosurfactants
Thermostable lipopeptide- type
biosurfactant
Aneurinibacillus thermoaerophilus
Reduction of the surface tension of water
Agriculture, food, and pharmaceutical, petroleum and paper/pulp industries Glicolipid-type
biosurfactant
Alcaligenes faecalis Decrease of surface tension at air-water interface, enhance of emulsification
Oil industry
Cyclic lipopeptides Bacillus licheniformis, B. subtilis, B. subtilis
Oil mobilization, enhance in oil recovery
Oil industry
* G6PD (glucose-6-phosphate dehydrogenas), 6PGD (6-phosphogluconate dehydrogenase), R5PI (ribose-5-phosphate isomerase).
解線狀纖維素 (如木醣、纖維素與半纖維素) 產生液態的生物燃料,
Caldicellulosiruptor、Caldanaerobius 與 Clostridium spp. 產生的纖維素酶 與木聚醣酶展現有效的木質纖維素降解能力,具有生產多種液態生物燃料 的潛力 (Bhalla et al., 2013; Han et al., 2012; Su et al., 2013)。
(b) 生物採礦 (Biomining)
生物採礦包括不同的生物加工過程,主要的目的是提高從礦石中金屬 的回收率,生物浸取 (bioleaching) 和生物氧化 (biooxidation) 是兩種應用 在硫化礦物上的生物萃取過程,進行生物採礦的微生物須具備產生氧化和 酸性條件的能力,攻擊金屬硫化物使其釋放金屬至酸性溶液中變成可溶性 硫酸鹽,具備嗜酸性與可氧化鐵和硫等特性的原核生物適合參與生物浸取 和生物氧化的過程,從礦物溶液中釋放金屬,藉由微生物氧化硫產生的硫 酸保持低 pH 值的條件,以維持溶液中金屬離子的狀態 (Donati and Sand, 2007)。雖然生物採礦加工理論上可以應用在其他金屬,但主要商業應用 的發展是在銅的生物浸取和難處理的金礦石的生物氧化方面。大部分商業 生物浸取的應用是在 40-50°C 下運作,主要是因為研究較透徹的生物浸 取的微生物是中溫菌或中度嗜熱菌 (Donati and Sand, 2007),然而使用嗜 熱菌的優點是在高溫下操作可以提高整體反應速率,此外也可以除去冷卻 系統所需的能量。
(c) 生物復育 (Bioremediation)
生物復育被視為生態友善的污染處理技術,利用微生物 (當地或是人 工引進的) 低成本的清除、控制或轉換環境中有機或無機污染物,使其變 成良性的產物 (Tabak et al., 2005)。使用嗜極性微生物處理受金屬汙染的 地方已變成有潛力的選擇 (Sen et al., 2014),嗜熱菌經由生物復育可以固 化有毒的金屬離子,甚至是輻射汙染的熱水。
嗜熱酵素的優異的特性適用於溫度相關的特定產業,如紙漿、食品、
釀造和飼料加工產業。嗜熱菌對於嚴苛的條件具有高度的抗性,例如化學 變性劑、廣泛的 pH 值範圍與無水溶劑,嗜熱酵素包含纖維素酶、木聚醣 酶、果膠酶、幾丁質酶、澱粉酶、支鏈澱粉酶、蛋白酶、脂肪酶、葡萄糖 異構酶、醇脫氫酶與酯解酶 (Bergquist et al., 2014; Chai et al., 2012; Elleuche et al., 2014; Kahar et al., 2013),嗜熱酵素在工業應用扮演著重要角色。
嗜熱酵素最常用於澱粉與木質纖維素去生產生物燃料,轉化過程需要 高溫和低 pH 的條件,以及澱粉酶、葡糖澱粉酶與支鏈澱粉酶的催化反應,
為了生產生物燃料,木質纖維材料的複合結構必須先以化學和物理方法分 解,進而 得到纖 維素 、半纖維 素與木 質素 ,再由對 應的水 解酶 處理 (Elleuche et al., 2014),考慮到預處理的條件,嗜極性水解酵素會比中溫酵 素來得更適合。
(e) 生物界面活性劑 (Biosurfactant)
生物界面活性劑是由微生物產生的兩性分子化合物,有助於提高非水 溶性化合物的乳化和分散。生物界面活性劑可以是醣酯、酯多醣、酯蛋白、
磷酯、羥基化交聯的脂肪酸與完整的細胞。因其毒性較低、較高的生物降 解能力、更好的環境相容性與高泡沫的形成,可在極端的溫度、pH 值和 鹽度下有較高的活性,生物界面活性劑在食品、農業、醫藥、石油、造紙 /紙漿行業的使用有顯著的增加 (Sharafi et al., 2014)。如在其他領域,嗜熱 菌已經證明生產的生物界面活性劑具有較高的熱穩定性,並提高其他極端 物 理 化 學 參 數 如 pH 和 鹽 度 的 抗 性 , 迄 今 被 鑑 定 的 菌 屬 為 Aneurinibacillus、Geobacillus、Alcaligenes、Bacillus 和 Brevibacillus 屬 (Bharali et al., 2011; Joshi et al., 2008; Mnif et al., 2011; Sharafi et al., 2014)。
(6) 全球酵素市場概況
全 球 酵 素 市 場 可 分 為 工 業 酵 素 (industrial enzyme) 與 特 用 酵 素 (specialty enzyme) 兩大類,2006 年全球工業酵素市場為 22.52 億美元,其
中約 3/4 是用於食品、澱粉加工與清潔劑工業,2006 年市場已接近 50 億 美元,估計 2010 年時已超過 60 億美元 (許,2007)。
工業酵素屬於大宗酵素 (bulk commodity enzyme),每種酵素需求量大但 價格相對較低。隨著需求量成長和相關創新技術的進步,新的酵素與多功能 酵素將會帶動市場成長,預估此產業每年約以 4% 的速度成長。工業酵素 依產品種類可分為醣類水解酶、蛋白酶、脂肪酶和其他酵素,若依市場應用 面來分類,則分為食品及飼料、清潔劑、紡織、皮革、造紙和其他應用 (許,
2007)。
特用酵素是應用於藥物開發、抗生素開發、醫療診斷和食品安全檢驗等 領域,相對於工業酵素,為高單價的產品。由於量少但高單價的特性,特用 酵素市場規模預估會高於工業酵素,且市場成長速度較快。根據市場研究公 司 Global Industry Analysts 指出,全球 2006 年特用酵素市場為 27.4 億美 元,估計將以年複合成長率 7.88% 成長,至 2010 年達 37.7 億美元。特用 酵素依產品種類可分為特用醫療酵素 (specialty medical enzyme)、聚合酶 (polymerase)、核酸酶 (nuclease) 與其他酵素。若依市場應用面來分類,則分 為 製 藥 與 診 斷 (pharmaceutical & diagnostic) 、 生 技 研 究 (research/biotechnology) 以及其他應用市場 (other end use market) (許,2007)。
因為基因解碼與分子生物學蓬勃發展,使得基因與蛋白質工程領域突飛 猛進。這些技術的進步已經使酵素篩選、修飾與製造的規模放大,加惠於產 業。大量累積的數據也促使科技發展進入生物資訊階段,利用胺基酸序列與 X-光晶體解析獲得的蛋白質立體結構資訊,讓我們可以在電腦上進行分子模 擬,預測胺基酸改變對蛋白質結構的影響,有利於縮短酵素設計的時間與減 少資源耗費。隨著資料庫內涵的累積,未來生物資訊將協助研發人員針對特 定性質,例如專一性、親和力、反應溫度與酸鹼度等,進行有效率的酵素改 造,甚至創造新的催化功能,增加酵素運用的廣度與深度。產業界對於特用 酵素發展的看法相當樂觀,認為未來將有大量的特用酵素出現,甚至作為工 業應用,酵素市場的型態將逐漸由大宗性質轉變為專屬性質,未來工業酵素 與特用酵素的區別將越來越模糊 (許,2007)。
3. 臺灣的地熱環境與嗜熱菌的研究 (1) 臺灣的地熱環境
臺灣島地處環太平洋火山帶上,同時處於菲律賓板塊與歐亞板塊之聚合 與壓縮界線上,島上地殼變動與地熱特徵明顯,擁有各種不同的地熱極端環 境,特別是溫泉的密度相當高。截至民國九十二年之調查,有命名之溫泉即 達 128 座,分佈於三萬六千平方公里的面積內,除分佈於海邊之「金山」、
「南澳」、「綠島」與「龜山島」等溫泉外,亦有處於海拔高達 1,400 公尺以 上的「大分」與「樂樂」等高山溫泉,其它則多分佈於海拔高低不同之山谷 底部河床上或沖積平原上,如「金崙」、「紅葉」與「彩霞」等伴隨溪谷而生 的野溪式溫泉,另有少部份如「霧鹿」與「中埔」等見於斷崖和峭壁等地質 環境 (圖三) (Chung, 1995; 宋,2003)。
天然的地熱環境與地殼變動常具有相伴隨的關係,通常可依其形態區分 成兩大類,第一類為硫噴氣孔區 (solfatara),包含硫結晶、酸性土壤、酸性 溫泉及沸騰泥池等。而第二類為中性至鹼性的流動水溫泉及間歇泉 (Brock, 1971)。除了以形態區分外,地熱區通常可以以其熱源來區分,分成高溫地熱 區及低溫地熱區兩類。高溫地熱區指的是靠近活火山區,以地底之岩漿為熱 源的區域,此區除了溫度高外 (地底水溫可達 150-350 oC),亦常伴隨氣體與 蒸氣噴出,氣體主要為 N2、CO2、H2S 及 H2,另有些微量氣體如 CH4 和 CO 亦存於其間 (Arnorsson and Gunnlaugsson, 1985)。當大量的 H2S 達到地 表後會因高溫及氧氣而被氧化成硫酸,使得高溫區的環境變成 pH 2-3 的強 酸性環境,但因高溫故此區水份不多,而多以噴氣孔形態現,所以穩定度不 高,臺灣大屯火山區 (陽明山及北投) 的溫泉便是屬於此類。低溫地熱區指 不在火山活動區附近,由地底熔岩或死火山脈為熱源所形成的地熱區。地下 水流在加熱過程中會溶入礦物質與氣體 (如 SiO2 及 CO2),回到地表後其 酸鹼度會因地質的不同而產生變化,多為中性到鹼性。由於低溫地熱區具大 量的水源及較低的熱源變動性,相較於高溫地熱區則較具穩定性 (Brock, 1971),不過此種地熱區的範圍較小,常會因地殼變動所造成的地震或是天 災造成的土石流導致生成或消失。
溫泉根據成份、濃度、溫度與酸鹼度的不同,泉質也不盡相同,常見的
圖三、臺灣溫泉地熱區與地質分佈圖。(Chung, 1995)
Figure 3. Taiwan hot spring area and geological map.
There are 128 hot springs all around Taiwan. S : Sedimentary. M : metamorphic rock. V : volcano rock.
為硫磺泉、碳酸泉與硫酸鹽泉等,再加上臺灣的地層結構也不盡相同,包含 變質岩、火成岩與沉積岩等,使得臺灣的溫泉有很高的多樣性。臺灣全島的 溫泉主要的分布為北部的大屯火山群,以及中央山脈兩側。北起宜蘭,南到 屏東,另外綠島的溫泉更是全球難得一見的特殊泉質,與日本鹿兒島以及義 大利的西西里島並列世界三大海洋溫泉。在這些特殊且多樣性的溫泉當中,
必然蘊藏了許多臺灣特有的自然資源,無論是無機的礦物質或者是多樣的生 命形態,都是十分值得我們進一步去研究,而其中的嗜熱性微生物更是我們 能夠加以研究運用的寶貴資產,也是目前有關臺灣溫泉的研究領域上,最具 有潛力且研究最為廣泛的部分 (魏,2006)。
(2) 臺灣嗜熱性微生物研究
臺灣近年來對嗜熱性微生物的研究可以分成兩大方向。一個部份著重於 嗜熱性微生物基礎研究,像是嗜熱性微生物的分離與培養、生理及生態、鑑 定與分類等;另一部份則是著重於嗜熱性微生物所產生之熱穩定性酵素及其 產物的研究及應用。
(a) 臺灣嗜熱性微生物的分離與鑑定
謝等以海洋細菌為研究材料,於 1993 年針對綠島溫泉進行採集,先 後發表了兩種本土海洋性嗜熱菌:Altercoccus agarolyticus (Shieh et al., 1998) 以及 Marinobacter lutaoensis (Shieh et al., 2003)。陳等曾對全臺灣的 22 座溫泉進行取樣分析及嗜熱菌的分離,總共分離純化出 520 株嗜熱菌,
初 步 建 立 了臺 灣 嗜 熱性 細 菌 種 源庫 , 並 發表 兩 株 新 種:Meiothermus taiwanensis (Chen et al., 2002a) 及 Pseudoxanthomonas taiwanensis (Chen et al., 2002b),之後再從種源庫中篩選出抗輻射的新種嗜熱菌 Rubrobacter taiwanensis (Chen et al., 2004)。此外陳等曾發表新種嗜熱菌 Caldimonas taiwanensis (Chen et al., 2005)。另外陳等人則發表新種嗜熱菌 Tepidimonas taiwanensis (Chen et al., 2006)。
(b) 臺灣嗜熱性微生物所產生之熱穩定性酵素
過去研究曾自 Thermus taiwanensis WL-219 選殖出超氧歧化酶,在 80 oC 下加熱 20 min 仍保有 70% 活性 (陶,2002),而前人自新種嗜熱
菌 Me. taiwanensis 中,廖與李亦曾選殖出兩個 lon protease:MtaLonA1 (793 a.a) 和 MtaLonA2 (815 a.a),並透過表現與純化探討其結構與功能的 特性 (Liao et al., 2012; Li et al., 2013),之後再選殖出嗜熱的 galactokinase (MtGalK)。因其耐熱的特性有助於在高溫下催化 sugar-1-phosphates 的生 成,並可以結合 glucose-1-phosphate thymidylyltransferase (RmlA) 與 α- 1,4-galactosyltransferase (LgtC) 製備 globotriose analogs (Gb3) (Li et al., 2014);另外也有從 Deinoccocus sp. NTU-1079 選殖出 α,β-galactosidase,
並應用在寡糖的合成 (陳,2010);李等人將 Meiothermus taiwanensis 之 galactokinase 用於合成 Pk 抗原類似物 (李,2011);另外張等人從 Gb.
uzenensis 分離一耐熱和耐酸的脂解酶 (張,2008)。
4. Anoxybacillus spp. 的發展與應用
Bacillaceae 科的菌種所產生的細胞或酵素對生物加工和生物轉換來說是
很好的來源。屬於格蘭氏陽性菌,其中大部分是桿菌少部分是球菌,目前有超 過兩萬種的種、亞種、未描述的菌種與環境樣本被歸類在 Bacillaceae 科裡 (表 五)。Bacillaceae 科菌種間的親緣關係如圖四 (Goh et al., 2013),菌種間 16S rRNA 序列的相似度平均高於 80 %,Anoxybacillus 屬為其中獨立演化的物種,
16S rRNA 序 列 的 相 似 度 為 92–99 %。 與 Anoxybacillus 屬 相 近 的 物 種 為 Geobacillus、Saccharococcus 與 Aeribacillus,相似度在 90–95 % 之間,與 Caldalkalibacillus 與 Calditerricola 屬物種的相似度則是較低,只有 85–88 % (Goh et al., 2013)。
(1) Anoxybacillus spp. 的歷史
Anoxybacillus flavithermus (部份文獻是使用 A. flavothermus) 最初文獻 命名為 Bacillus flavothermus。從紐西蘭的溫泉中被發現,為會產生黃色色素 的兼性厭氧菌,生長溫度範圍為 30-70 °C (Heinen et al., 1982)。隨後有幾株 和 Bacillus flavithermus 16S rRNA 序列相似的細菌從黃石國家公園的八達通 溫泉 (Octopus Spring) 和澳大利亞的大自流盆地 (Australian Great Artesian Basin) 被發現 (Byers et al., 1998; Nold et al., 1996)。在 2000 年時,Pikuta 等學者從動物糞便中分離出厭氧菌株 K1T,並根據菌株 K1T 的 16S rRNA、
應該是 Bacillaceae 裡的一個新的屬。其命名是根據其厭氧的特性,anoxic or anoxia 指的是可溶性氧氣已經耗盡的情況。菌株 K1T 並因此被命名為 A.
pushchinensis。進一步的親源性分析的結果顯示,A. pushchinensis 和 B.
flavithermus 應為近親物種,並和 Bacillus spp. 分開,因此 B. flavithermus 被重新歸類為 A. flavithermus (flavothermus)。
自 從 Pikuta 學 者 在 2000 年 首 次 發 表 Anoxybacillus spp. 屬 後 , Anoxybacillus 屬的菌種陸續在世界各地被分離出來,至今約有 22 種及 2 亞種 (表六)。Anoxybacillus spp. 大部分是桿菌與少許的曲狀桿菌,且會形成 內胞子,其中有嗜酸性也有嗜鹼性的菌種,大部分的菌生活在中性 pH 的環 境。Anoxybacillus spp. 為中度嗜熱菌,最適生長溫度為 50-62 oC,比相近的 Geobacillus 的最適生長溫度 55-65 oC 略低,好氧菌或兼性厭氧菌皆有,值 得注意的是 Anoxybacillus spp. 的 genomes 平均大小為 Bacillaceae 科中最 小的 (圖五) (Goh et al., 2014)。
與 Geobacillus 以及 Bacillus 相比,Anoxybacillus 是相對較新的屬,因 為 其 耐 鹼 及 嗜 熱 的 性 質 , 具 有 產 業 應 用 上 的 潛 力 。 許 多 文 獻 指 出 , Anoxybacillus spp. 具有多種應用價值,像是分解澱粉和木質纖維素、環境廢 棄物的處理、酵素選殖與生產生質能源。
(2) Anoxybacillus spp. 的研究與應用
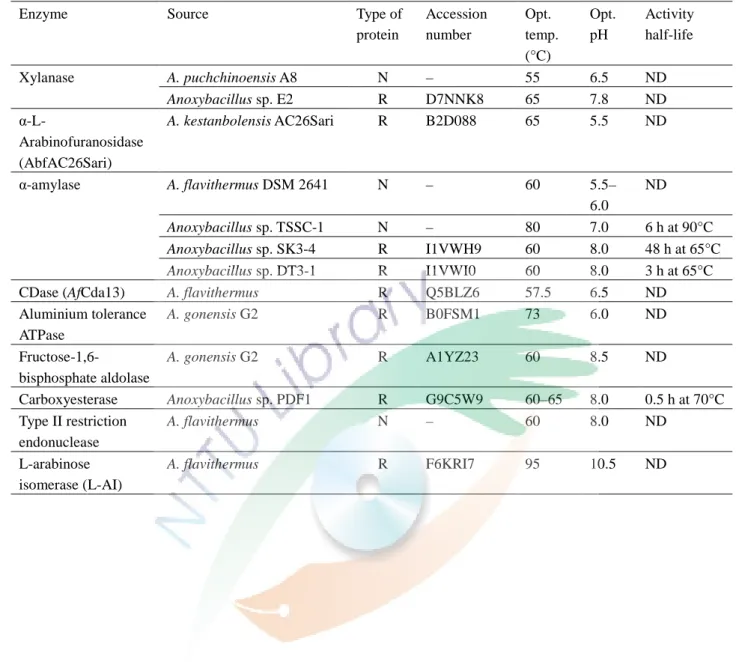
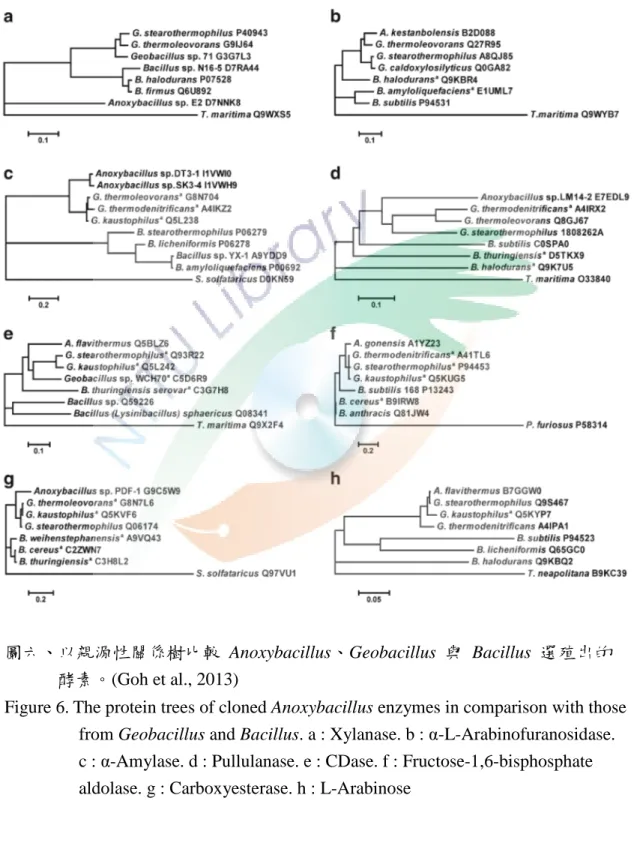
表七整理了 Anoxybacillus spp. 一些原態或選殖蛋白與其基本的生化特 性,一般來說這些酵素都具有熱穩定性與耐鹼性的能力,Goh 學者也利用 Anoxybacillus spp. 的選殖蛋白建立親源性關係樹 (圖六),由圖六可知大部 分選殖的蛋白序列皆和 Geobacillus spp. 相似且遠離 Bacillus spp. (Goh et al., 2013)。
(a) 木質纖維素水解酶
木質 纖維素 包括 纖維素、 半纖維 素和 木質素等 複雜的 結構 。由 Anoxybacillus spp. 所產生的纖維酵素相對來說較少,且較少與真菌像是 Trichoderma reesei 去 進 行 比 較 (Couturier et al., 2012) 。 文 獻 指 出 A.
flavithermus BC (Kambourova et al. 2007)、A. pushchinoensis A8 (Kacagan et al. 2008) 與 Anoxybacillus sp. E2 (Wang et al., 2010b) 這些從 Turkish 溫
表五、 Bacillaceae 科的菌屬 (Goh et al., 2013) Table 5. Genus in the Bacillaceae family
Genus Numbers Abbreviation used in Fig.
Aeribacillus 4 Aerib
Alkalibacillus 21 Alkal
Amphibacillus 48 Amphi
Amylobacillus– 1 –
Anaerobacillus 4 Anaer
Anoxybacillus 145 Anoxy
Aquisalibacillus 4 Aquis
Bacillus 16,545 Bacil
Caldalkalibacillus 5 Calda
Calditerricola 2 Caldi
Cerasibacillus 1 Ceras
Environmental 3 –
Filobacillus 11 Filob
Geobacillus 467 Geoba
Gracilibacillus 66 Graci
Halalkalibacillus 2 Halal
Halobacillus 516 Halob
Halolactibacillus 9 Halol
Lentibacillus 13 Lenti
Lysinibacillus 286 Lysin
Marinococcus 43 Marin
Microaerobacter 1 Micro
Natribacillus 1 Natri
Natronobacillus 1 Natro
Oceanobacillus 169 Ocean
Ornithinibacillus 39 Ornit
Paraliobacillus 10 Paral
Paucisalibacillus 4 Pauci
Piscibacillus 12 Pisci
Pontibacillus 55 Ponti
Psychrobacillus 4 –
Saccharococcus 11 Sacch
Salimicrobium 26 Salim
Salinibacillus 4 Salin
Salirhabdus 3 Salir
Salsuginibacillus 10 Salsu
Sediminibacillus 11 Sedim
Tenuibacillus 5 Tenui
Terribacillus 31 Terri
Thalassobacillus 45 Thala
Unclassified 425 –
Virgibacillus 256 Virgi
Vulcanibacillus 3 Vulca
– not incloud in figure 4
圖四、Bacillaceae 科的親緣關係樹。(Tamura et al. 2011) Figure 4. Cladogram of Bacillaceae family.
The 16S rRNA sequences of representatives were aligned using ClustalW and a neighbor-joining tree was build within the MEGA5 package. The transformedcladogram was redrawn using the FigTree program.
表六、文獻中 Anoxybacillus 物種的基本特性 (Goh et al., 2013) Table 6. General characteristics of Anoxybacillus species in previous reports
The type strains of Anoxybacillus species have been deposited in two or more public culture collections and their corresponding culture collection numbers are shown in the papers cited, respectively
A aerobe, F.a facultative aerobe, F.an facultative anaerobe, ND not determined
Species Source Place Growth
condition
Temp.
range
Opt.
temp pH range
Opt.
pH A. amylolyticus Geothermal soil Mount Rittmann, F.an 45–65 61 5.0–6.5 5.6 A. ayderensis Ayder hot spring Rize, Turkey F.an 30–70 50 6.0–11.0 7.5–8.5 A. bogrovensis Geothermal spring Sofia, Bulgaria F.an 40–69 65 6.0–10.0 8.0 A. caldiproteolyticus Sewage sludge samples Seletar, Singapore A 35–65 60 6.0–9.0 6.5
A. calidus Soil sample Denizi, Turkey F.an 35-70 55 6.5-9.0 8.0-8.5
A. contaminans Gelatin samples France F.an 50–60 50 4.0–10.0 7.0
A. eryuanensis Eryuan hot spring Yunnan, China F.an 35–75 55 7.0–11.0 8.0 A. flavithermus Unnamed hot spring Northern island of
New Zealand
F.a 30–72 60–65 5.5–9.0 7.0
A. flavithermus subsp.yunnanensis
Unnamed hot springs Yunnan, China F.a 30–66 60 5.5–10.0 7.0–7.5
A. gonensis Gonen hot springs Balkesir, Turkey F.an 40–70 55–60 6.0–10.0 7.5–8.0 A. Kamchatka Hot spring in
Geyser valley
Kamchatka, Russia
F.a 38–67 60 5.7–9.9 6.8–8.5
A. kamchatkensis subsp. asaccharedens
Talisdere hot spring Batman, Turkey A 35–65 55 5.5–9.5 7.5
A. kaynarcensis Kaynarca hot spring Izmir, Turkey A 35–70 60 6.0–10.0 7.0 A. kestanbolensis Kestanbol hot spring Canakkale,
Turkey
F.an 40–70 50–55 6.0–10.5 7.5–8.5
A. mongoliensis Tsenher hot spring Central Mongolia F.an 35–75 60 5.0–10.8 8.0 A. pushchinoensis Manure samples Moscow, Rusia F.a 37–66 62 8.0–10.5 9.5–9.7 A. rupiensis Unnamed hot spring Rupi basin,
Bulgaria
A 35–67 55 5.5–8.5 6.0–6.5
A. salavatliensis Well-pipeline sediment Aydin, Turkey F.an 37–69 60 5.5–9.5 8.0–9.0 A. tengchongensis Tengchong hot spring Yunnan, China F.an 30–75 50 7.0–11.0 8.5 A. tepidamans Geothermal heated soil Yellowstone
National Park, USA
F.an 39–67 55 6.0–9.0 7.0
A. thermarum Euganean hot Padova, Italy A 55–67 65 6.0–7.5 7.2
A. toebii Hay compost Korea A 45-70 60 6.0-9.0 7.5
A. vitaminiphilus Pung hot spring Xichang, china A 38-60 57-60 6.0-9.3 7-7.5 A. voinovskiensis Voinovskie hot spring Kamchatka,
Russia
F.a 30–64 54 7.0–8.0 ND
圖五、根據串聯 361 條同源基因的序列所建立的 Bacillaceae 親源性關係圖。
(Goh et al., 2014)
Figure 5. Phylogenetic tree of selected Bacillaceae based on concatenated sequences of 361 orthologs.
表七、由不同 Anoxybacillus 物種所生產的酵素生化特性 (Goh et al., 2013)
Table 7. Summary of basic biochemical properties of purified enzymes from various Anoxybacillus species
Enzyme Source Type of
protein
Accession number
Opt.
temp.
(°C)
Opt.
pH
Activity half-life
Xylanase A. puchchinoensis A8 N – 55 6.5 ND
Anoxybacillus sp. E2 R D7NNK8 65 7.8 ND
α-L-
Arabinofuranosidase (AbfAC26Sari)
A. kestanbolensis AC26Sari R B2D088 65 5.5 ND
α-amylase A. flavithermus DSM 2641 N – 60 5.5–
6.0
ND
Anoxybacillus sp. TSSC-1 N – 80 7.0 6 h at 90°C
Anoxybacillus sp. SK3-4 R I1VWH9 60 8.0 48 h at 65°C
Anoxybacillus sp. DT3-1 R I1VWI0 60 8.0 3 h at 65°C
CDase (AfCda13) A. flavithermus R Q5BLZ6 57.5 6.5 ND
Aluminium tolerance ATPase
A. gonensis G2 R B0FSM1 73 6.0 ND
Fructose-1,6-
bisphosphate aldolase
A. gonensis G2 R A1YZ23 60 8.5 ND
Carboxyesterase Anoxybacillus sp. PDF1 R G9C5W9 60–65 8.0 0.5 h at 70°C Type II restriction
endonuclease
A. flavithermus N – 60 8.0 ND
L-arabinose isomerase (L-AI)
A. flavithermus R F6KRI7 95 10.5 ND
圖六、以親源性關係樹比較 Anoxybacillus、Geobacillus 與 Bacillus 選殖出的 酵素。(Goh et al., 2013)
Figure 6.The protein trees of cloned Anoxybacillus enzymes in comparison with those from Geobacillus and Bacillus. a : Xylanase. b : α-L-Arabinofuranosidase.
c : α-Amylase. d : Pullulanase. e : CDase. f : Fructose-1,6-bisphosphate aldolase. g : Carboxyesterase. h : L-Arabinose
泉分離的株菌中皆有木質纖維素酶的活性 (İnan et al., 2011b)。上述菌株 所生產的木聚糖酶 (xylanase) 分子量大小為 38-92 kDa,且展現了不同的 生化特性。從菌株 I3、CT1Sari、BT2.1ACT2Sari 與 AC26 取得的木聚糖 酶 , 在 pH 為 9 的 情 況 下 有 最 大 的 活 性 (İnan et al., 2011b) 。 而 Anoxybacillus sp. A8 的 木 聚 糖 酶 最 適 pH 為 6.5 , 偏 好 酸 性 的 條 件 (Kacagan et al., 2008)。在其他的例子中,菌株 E2 產生的木聚糖酶可以耐 熱到到 60oC,展現了廣泛的 pH 耐受性 (pH 4.6-12.0),更重要的是沒有 纖維素酶的活性 (Wang et al., 2010b)。而且 A. flavithermus 木糖酶水解木 聚糖的能力是 Sulfolobus solfataricus β-xylosidase/α-arabinosidase 的三倍 以上,結合兩種嗜熱菌的酵素可以水解 63.6 % 的山毛櫸木聚醣,比 Bacillus thermoantarticus 在同條件的反應還好。
阿拉伯呋喃糖酶 (arabinofuranosidase) 的存在可以促進半纖維素的 α-1,2- 和 α-1,3-arabinofuranosidic 鍵的水解 (Saha, 2003)。在 2008 年時 Çanakçi 學 者 從 A. kestanbolensis AC26Sari 中 選 殖 出 α-L- arabinofuranosidase (AbfAC26Sari) 的基因並表現在 Escherichia coli BL21 中 (Çanakçi et al., 2008) 。酵素 AbfAC26Sari 與 Geobacillus 的 α-L- arabinofuranosidase 的胺基酸序列有 86-91 % 的相似度,且具有廣泛的多 醣水解能力,特別是對呋喃構型與 α-form 的糖苷鍵結。AbfAC26Sari 的 最適溫度為 65oC,最適 pH 為 5.5。相反的,從 A. flavithermus EHP2 分 離的纖維素酶 (cellulase) 對 avicel 微晶纖維素、羧甲基纖維素與纖維雙 糖有特別高的活性,其最佳的反應溫度為 75oC,最適 pH 範圍為 7.0-8.0 (Ibrahim and El-diwany, 2007)。Anoxybacillus sp. 527 的纖維素酶的最適反 應溫度為 70oC,最適 pH 為 6.0,額外添加 Ca2+ 和 NH4+ 離子可以提升 纖維素酶的活性。
Thitikorn 學者在培養 Anoxybacillus sp. JT-12 時發現有木聚糖酶 (xylanase) 、 木 糖 苷 酶 (β-xylosidase) 、 阿 拉 伯 呋 喃 糖 酶 (α-L- arabinofuranosidase) 與乙醯酯解酶 (acetyl esterase) 的活性,以含有樺木 木聚醣的培養基,在 55oC 且 pH 7.0 下培養 42 小時後,在菌液的上清 液發現有木三糖的產生,此研究證明 Anoxybacillus 可能是良好的木三糖 生產者 (Thitikorn et al., 2012)。