國立臺灣大學生命科學院生化科技學系 碩士論文
Department of Biochemical Science and Technology College of Life Science
National Taiwan University Master Thesis
利用電泳分析程式預測之阿拉伯芥中可能形成 R-loop 結構之基因
Analysis of the program-predicted R-loop forming genes in Arabidopsis by gel mobility shift assay
陳亞甄 Ya-Jhen Chen
指導教授:黃楓婷 博士
楊健志 博士 Advisor: Feng-Ting Huang, Ph.D.
Chien-Chih Yang, Ph.D.
中華民國 105 年 7 月
國立臺灣大學碩士學位論文
口試委員會審定書
中文題目 : 利用電泳分析程式預測之阿拉伯芥 中可能形成 R-loop 結構的基因
英文題目:Analysis of the program-predicted R-loop forming genes in Arabidopsis by gel
mobility shift assay
本論文係___陳亞甄_____君(學號 R02B22036)在國立
臺灣大學生化科技學系完成之碩士學位論文,於民國 105 年
7 月 21 日承下列考試委員審查通過及口試及格,特此證明
謝誌
在碩班的兩年半來,不長也不短,得到的收穫比我能付出的多很多,要感 謝的人也很多,很幸運能遇見這麼多願意幫我的人。
首先要感謝指導教授楊健志老師,楊老師總是很有耐心的教導我們,不管 是在實驗上或生活上,在我焦慮不安的時候總能出實驗方向,平常中午吃飯時 也願意跟我們分享大小事,也一直鼓勵我要有信心。再來謝謝另一位指導教授 黃楓婷老師,老師總能點出我實驗上的盲點,其實我每次看到老師都覺得有點 害羞而語無倫次,感謝老師的多方包容。還有這一年來跟我們一起 meeting 的 王老師,每次 meeting 完都讓我有種到了新世界的感覺,老師總能提醒我容易 被遺忘的小細節,非常謝謝王老師。還有我們口試委員們,廖憶純老師及陳佩 燁老師,謝謝你們提醒我在論文書寫上的缺漏,也謝謝你們在口試上提出的問 題,讓我能讓這本碩士論文變得更好。
再來要感謝實驗室的大小夥伴們,非常謝謝梁景興學長在實驗上給的指 導,雖然我有時候耳朵很硬,但最後總會證明學長是對的。還有跟我一起口試 的蓁華,我們也走到這一天了。實驗室的小學弟妹們,珆安、Helen、煥宇、法 理、Justin 和孫元恩,八月的我因為有你們的吵吵鬧鬧和說笑話的功力,感覺沒 有那麼疲憊。還有已經畢業的老爹學長,坐在你旁邊的日子真的非常有趣,感 謝你包容怪癖很多的我。以及我的同學們,旻壕、奕如、成浩、閔翔及哲倫,
過去的一年內我好懷念我們當初吵鬧的氣氛,希望我們未來都可以朝自己想去 的地方邁進。
最後要謝謝我的男朋友及家人,謝謝魏皓宇在我論文格式上的無限幫忙,
謝謝我爸媽支持我念完碩班,我總是不在家裡,還依靠著你們給我生活費,希 望未來可以換我來照顧你們。
目錄
目錄 ... 1
摘要 ... 3
Abstract ... 4
縮寫表 ... 5
第一章 緒論 ... 6
1.1 細胞中 DNA/RNA 雜合體的生成 ... 6
1.1.1 R-loop 生成的特徵 ... 7
1.2 R-loop 的存在與基因不穩定性( genomic instability) ... 8
1.3 調控 R-loop 的生理機制 ... 9
1.3.1 細胞中移除 R-loop 結構的方法 ... 10
1.3.2 防止 R-loop 形成的生理機制 ... 11
1.4 R-loop 的生理功能 ... 13
1.5 R-loop 與非編碼 RNA (non-codng RNA, ncRNA) ... 13
1.7 依 R-loop 的序列特徵建立計量模型 ... 14
1.8 建立 R-loop 基因搜尋程式 ... 15
1.9 本篇論文之研究起源與動機 ... 15
第二章 材料與方法 ... 17
2.1 植物材料及生長條件 ... 17
2.1.1 植物材料 ... 17
2.1.2 植物阿拉伯芥培養基 1/2 MS ... 17
2.1.3 種子表面消毒及低溫處理 ... 17
2.1.4 種子之無菌培養 ... 18
2.2 The gel mobility shift assay ... 18
2.2.1 以 Phusion DNA polymerase 進行聚合酶連鎖反應 (polymerase chain reaction, RCP) ... 18
2.2.2 DNA 瓊脂糖膠體電泳 (Agarose Gel Electrophoresis) ... 19
2.2.3 PCR 產物之分離與純化... 19
2.2.4 TA cloning ... 20
2.2.5 大腸桿菌培養基 ... 20
2.2.6 大腸桿菌熱衝擊轉形(Heat-shock Transformation) ... 21
2.2.7 菌落 PCR 檢定法 ... 21
2.2.8.1 利用實驗套組進行質體 DNA 小量製備 ... 21
2.2.8.2 以鹼裂解法進行質體 DNA 小量製備... 22
2.2.9 胞外轉錄 (in vitro transcription)方法一 ... 22
2.2.10 胞外轉錄方法二 ... 23
2.2.11 胞外轉錄方法三 ... 24
第三章 結果與討論 ... 25
3.1 利用 R-loop searching program 尋找阿拉伯芥中可能形成 R-loop 的基因 ... 25
3.1.1 挑選目標基因的條件 ... 26
3.2 以 Gel mobility sift assay 檢驗在 in vitro 條件下是否有 R-loop 結構存在... 27
3.3 13 個候選基因進行 GMSA... 29
第四章 未來展望 ... 37
參考文獻 ... 39
Figures ... 44
Table ... 75
Appendices ... 76
問答集 ... 95
摘要
R-loop 為 RNA 與互補 DNA 雜合後使另一股 DNA 分離而單獨存在所形成 的三股核酸結構。R-loop 的結構包含由轉錄後的 RNA 與模板 DNA (template strand DNA ) 重新配對,及單股的非模板 DNA (non-template strand DNA)。植物 中第一個 R-loop 有關之研究,是 COOLAIR 基因形成 R-loop 後,失去抑制 FLC 的作用,而共同調控植物開花。為了深入了解 R-loop 是否在植物扮演其他角色,
本實驗室謝閔翔與生物產業機電工程系陳倩瑜老師實驗室合作,使用程式語言
「C++」來設計出可以搜尋出可能形成 R-loop 的基因的程式,篩選出了在 53 個 在阿拉伯芥中可能形成 R-loop 的基因。謝 (2015)已研究 LBD18 在 in vitro 及 in vivo 形成 R-loop。本實驗挑選 13 個基因以胞外轉錄及三種 gel mobility shift assay 進行研究。13 個基因中,部分出現 in vitro R-loop 形成之特徵性色帶模糊。但同 一種基因中,三種 gel mobility shift assay 得到的實驗結果不盡相同,且 LBD18 在 三種方法中都沒有呈現色帶模糊。另外,本研究發現之前使用 urea 於染色體 DNA 抽取可能有缺點,因此使用改良的抽取方法,唯此方法未能再現 LBD18 在 native bisulfite sequencing assay 之結果。
Abstract
R-loop is a triple helix structure consisting of a DNA/RNA hybrid plus a single strand DNA. R-loops are formed when newly synthesized RNA threads back to anneal with the template strand DNA, leaving the non-template strand DNA remaining as a single strand. In Arabidopsis, the expression level of COOLAIR, the first gene discovered that can form R-loops in plants, coordinates flowering with FLC. To explore if R-loop formation is widespread in plants, Min-Hsiang Hsieh designed an R-loop searching program written using C++ together with Dr. Chien-Yu Chen (Department of Bio-industrial Mechatronics Engineering of National Taiwan University) .Fifty-three genes were predicted to be able to form R-loops by the program. Hsieh (2015) found that LOB domain-containing protein 18 (LBD18) form DNA/RNA hybrids in vitro and in vivo. In this study, 13 genes were tested by in vitro transcription and three kinds of gel mobility shift assay. Part of the 13 genes had the distinctive smear of DNA/RNA hybrid in vitro, but different protocol caused different consequences in some genes. In this study, LBD18 did not have smear pattern by three kinds of gel mobility shift assay.
Moreover, there was shortcoming of using urea to extract genomic DNA by Hsieh.
(2015). Modified genomic DNA extraction protocol did not repeat the native bisulfite sequencing result of LBD18.
縮寫表
AID activation-induced cytidine deaminase CSR class switch recombination
CRISPR clustered regularly interspaced short palindromic repeat
EDTA Ethylenediaminetetraacetic acid
EtBr ethidium bromide
G4 G quadruplex
LBD18 Lob domain-containing protein 18 PCR polymerase chain reaction REZ R-loop elongation zone
RIZ R-loop initiation zone RLFS R-loop forming sequence
TSS Transcription start site
TOP topoisomerase
qPCR Quantitative real time polymerase chain reaction
第一章 緒論
R-loop 為一種在 DNA 進行轉錄作用後,由兩股 DNA 及新轉錄生成之 RNA 所形成的三股螺旋結構。由於 R-loop 出現後,會有單股 DNA 形成,而增 高基因體不穩定性 (genome instability),因此生物體內發展了若干對抗 R-loop 的機制,如防止 R-loop 結構生成與移除以形成之 R-loop。R-loop 的研究目前多 在哺乳動物相關研究領域,本實驗室與生機系陳倩瑜教授研究室合作發展出 R- loop searching program (謝, 2015, 碩士論文),希望能在植物中找到可能形成 R- loop 的基因,以了解更多 R-loop 在植物中的生理功能。
1.1 細胞中 DNA/RNA 雜合體的生成
在轉錄作用進行時,轉錄泡 (transcription bubble) 中大約會有 8bps 的 DNA/RNA 雜合體片段 (Westover et al., 2004);而 DNA 複製作用中遲緩股 (lagging strand)上,則有引導 RNA 合成酶 (RNA primase) 所產生的約 11 個 bps 的 DNA/RNA 雜合體片段 (Reddy et al., 2011; Aguilera and Garcia-Muse, 2012),
這些雜合片段出現的時間短暫,在 RNA 離開後,DNA 又重新回到雙股配對。
R-loop 是一種由 DNA/RNA 雜合體形成的三股螺旋結構,在轉錄完成後,產 生的 RNA 與模板 DNA (template DNA) 重新配對形成雙股結構,而非模板 DNA (nontemplate DNA) 則形成單股結構。最早是發現在果蠅中被報導 (Thomas et al., 1976)。然而目前研究顯示,R-loop 的 DNA/RNA 雜合片段長度大約落在 110 到 1280 bps 之間 (Duquette et al., 2007)。
1.1.1 R-loop 生成的特徵
R-loop 是轉錄作用中所產生的結構。在一般的轉錄過程中,新合成的 RNA 會離開模板 DNA,使 DNA 又重新回到雙股螺旋的狀態。原先推測產生 R-loop 的原因是轉錄過程中 DNA/RNA 雜合體的延伸,但 2004 年在 RNA Polymerase II 的結晶結構發現,原先產生的 DNA/RNA 雜合體中的 DNA 與 RNA 會分別從 Pol II 上不同的通道離開,因此已推翻了這項說法 (Westover et al., 2004)。目前主流 的立論是 R-loop 的形成機制為轉錄出的 RNA 在離開 RNA polymerase 的通道 (exit channel) 後,與尚未重新黏合的雙股 DNA 中的模板 DNA 重新結合,稱 為”thread back” model。
2009 年的研究中指出,形成 R-loop 的序列有三個特徵: (1)鳥嘌呤 (guanine) 密度高,(2)負超螺旋 (negative supercoiling),(3)DNA 缺口 (nick)。R-loop 形成 的起點偏好由 G cluster,及非模板股啟動子序列的下游有具 DNA 缺口處開始,
而 DNA/RNA 雜合體的延伸及結構穩定性,藉由序列上高密度的鳥嘌呤和負超螺 旋所維持 (Roy and Lieber, 2009; Roy et al., 2010)。
研究顯示,DNA/RNA 雜合體的熱穩定性會較 DNA/DNA 雜合體高,而雜合 體的熱穩定度與 RNA 中嘌呤的比例有關,嘌呤越多則穩定度越高 (Shaw and Arya, 2008)。顯示熱穩定性高的證據為以∆G˚37為例,RNA/DNA (GG:CC) 雜合 體 的∆G˚37 為 -2.91 , 相 較 於 DNA/DNA (GG:CC) 雜 合 體 的 -2.75 更 為 穩 定 (Breslauer et al., 1986; Sugimoto et al., 1995)。對於 DNA/RNA 雜合體的穩定性比 DNA/DNA 雜合體高,目前提出了兩種解釋。第一種為 DNA/RNA 雜合體的構型 是介於雙股 RNA 所形成的 A form 與雙股 DNA 形成的 B form 之間 (Shaw and Arya, 2008)。第二種可能性為當 R-loop 形成時,非模板 DNA 為單股 DNA,其
上的序列特性易產生 G-四聯體 (G quadruplex , G4),進而穩定 R-loop 的結構 (Duquette et al., 2004)。因此在序列中若含有連續的鳥嘌呤將會拉大 RNA/DNA 雜合體與 DNA/DNA 雜合體之間的熱穩定性的差距,而促使 RNA 模板 DNA 黏 合,形成 R-loop 結構。亦即若轉錄出含有高度鳥嘌呤的 RNA,則可幫助 R-loop 的形成。但是若是轉錄轉錄出含有高度胞嘧啶 (cytocine) 的 RNA 的區域,卻不 會使得 R-loop 形成 (Ginno et al., 2012)。
1.2 R-loop 的存在與基因不穩定性( genomic instability)
在轉錄過程中出現的 R-loop 對 DNA 來說可能具有高危險性。一旦產生 R- loop,亦即含有單股 DNA 裸露在外,則單股 DNA 不穩定也則容易受到核酸水解 酶切除。目前對於 R-loop 產生時所造成的基因不穩定的機制還不清楚,現行有 兩種假說,第一種是 R-loop 在多個區域形成時,大量出現的單股 DNA 會容易受 到 DNA-damaging agent 破壞;第二種則是可以辨認單股 DNA 的蛋白質使 DNA 產生突變 (mutagenesis),最有可能參與此作用的蛋白質為 activation-induced cytidine deaminase (AID)。AID 是一種會促進免疫球蛋白 (Ig) 的重鏈 (heavy chain) 進 行 class switch recombination (CSR) 以 及 促 進 B 淋 巴 球 進 行 hypermutation,AID 會將單股 DNA 上的 dC 轉變為 dU (Muramatsu et al., 2000;
Revy et al., 2000)。當 R-loop 形成時,單股 DNA 被 AID 辨識,其上的 dC 突變,
而後續的複製則會讓 DNA 出現兩種序列,一種是正常的 DNA 序列,另一種為 產生 C-T 轉變的 DNA。除此之外,一旦 dU 產生,細胞可能會進行鹼基切除修 復 (base excision repair , BER)。鹼基切除修復會使得 dU 被去除而造成雙股螺旋 斷裂 (DNA double strand break , DSB), 或是在修補 DNA 缺口的過程中因為隨 機放入四種核苷酸而產生的 DNA 序列突變。以抗體基因為例,DNA DSB 產生 後,會被 recombination-mediated repair machinery 所辨認,而導致 CSR 或是 DNA translocation (Di Noia and Neuberger, 2002)。但 AID 只存在於 B 細胞以及 B 細
胞所衍生的 DT40 細胞內,在其他缺乏 AID 的細胞是如何產生 R-loop 形成後所 造成的基因不穩定則仍然未知。在一般情況下 dC 與 dU 之間也會有少部分的轉 換發生。
除了 DSB 外,另一種造成基因不穩定的可能性為 R-loop 干擾了 DNA 複製 的過程。在出芽型酵母菌 (budding yeast) 以及哺乳動物中發現,當 Pol II 停止 後,複製叉 (replication fork) 互相碰撞而造成 DNA break (Prado and Aguilera, 2005; Boubakri et al., 2010)。一旦 R-loop 在複製過程中形成後,單股 DNA 上的 傷害或是 DNA/RNA 雜合體本身會阻止 DNA 複製的進行,造成複製叉停止,而 複製叉的停止使得新合成的 DNA 具有 DSB,引起 recombination-mediated repair,
進而可能造成染色體重新組合 (chromosome rearrangements) 以及基因不穩定 (Aguilera and Garcia-Muse, 2012)。
2013 年發現了另一個可能是 R-loop 造成不穩定的原因,R-loop 形成後,會 促使 histone 3 Ser10 phosphorylation (H3S10P) 染色質壓縮 (compaction)的標記形 成,而造成了複製叉的停止,最終使得新合成的 DNA 具有 DSB (Castellano-Pozo et al., 2013)。
1.3 調控 R-loop 的生理機制
在轉錄過程中,R-loop 的形成會對於基因組的正確度產生破壞, R-loop 的 形成在演化上來說是具有保守性的機制,因此不同物種間也產生了相對應保護基 因組的方法 (Li and Manley, 2006; Aguilera and Garcia-Muse, 2012)。常見的方法 有:(1)利用 RNase H 降解 DNA/RNA 雜合體上的 RNA (Cerritelli and Crouch, 2009)。(2)利用 RNA/DNA helicase,例如酵母菌中的 Sen1 或是人類同源蛋白 Senataxin,可以解開 DNA/RNA 雜合體結構(Mischo et al., 2011; Skourti-Stathaki et
al., 2011),或是人類蛋白 DHX9 helicase 可以破壞 G4 結構 (Chakraborty and Grosse, 2011)。(3)利用 topoisomerase 鬆開 negative supercoiling 使得 R-loop 不易形成 (Drolet et al. 1994, 1995; Tuduri et al., 2009)。(4)防止 R-loop 形成的蛋白質。(5)抑 制幫助 R-loop 形成的蛋白質。前兩組可視為去除形成後的 R-loop,後三組則為 防止 R-loop 形成。
1.3.1 細胞中移除 R-loop 結構的方法
第一,RNase H 可以去除 DNA/RNA 雜合的中的 RNA,在大多數的物種裡,
有兩種 RNase H 存在,分別是 RNase H1 及 RNase H2 (Cerritelli and Crouch, 2009)。
RNase H1 出現在細胞核及粒線體中,對粒線體的複製很重要。過量表現細胞核 中的 RNase H1 可以有效去除 R-loop (Cerritelli et al., 2003),是目前在實驗上被廣 泛應用方法。RNase H2 的功能則為去除 DNA 複製過程中的 RNA 引子 (Cerritelli and Crouch, 2009)。
第二,酵母菌中的 Sen1 屬於 RNA/DNA 解旋酶 (helicase) 中的一員 (Kim et al., 1999),負責去除 R-loops 以及防止 R-loop 所造成的 DNA 傷害 (Mischo et al., 2011) 。Sen1 的突變株 sen1-1 發現有 R-loop 的累積,除此之外,Sen1 在 coding 以及 noncoding 基因也扮演終止因子 (termination factor) 的角色 (Ursic et al. 1997;
Steinmetz et al., 2006; Kawauchi et al., 2008),Sen1, Nrd1 及 Nab3 組成 NRD 複合 體,負責 sno/snRNA 的終止反應 (termination) (Ursic et al. 1997; Steinmetz et al., 2001; Chinchilla et al., 2012)。
人類的 Senataxin 是 Sen1 在的同源蛋白質,與轉錄作用中的終止反應有關 (Skourti-Stathaki et al., 2011; Padmanabhan et al., 2012; Becherel et al., 2013)。越來 越多的證據指向 Senataxin 是一種 DNA 修復酵素 (Becherel et al., 2013),以及用
來解開存在於 G-rich termination pause sites 的 R-loop。2013 年 Senataxin 被報導 在細胞分裂週期中的 S/G2 時期表現量上升,此時存在著 DNA 損害以及受傷害 的 DNA 複製 (Yüce and West 2013) 。此外,Senataxin 的表現量會隨著 R-loop 被 解開以及轉錄反應被終止後而明顯下降。2013 另一篇報導也指出,Senataxin 在 減數分裂中促進解開 R-loop 結構,以確保基因組的正確性 (Becherel et al., 2013)。
1.3.2 防止 R-loop 形成的生理機制
在 S. cerevisiae 中,由轉錄所產生的 R-loop 最初與四種核酸蛋白(nuclear proteins) (Hrp1, Tho2, Mft1, and Thp2)所組成的 THO 複合體一起被報導出來 (Aguilera, 2002) 。 在 酵 母 菌 中 , THO 突 變 後 , 會 產 生 跟 R-loop 相 關 的 hyperrecombination 性狀 (Piruat and Aguilera, 1998)。酵母菌中的扮演傳遞與處理 mRNA 的蛋白質 Npl3 也被發現可以預防 R-loop 所引起的基因不穩定,這也連結 了 RNA 的代謝與 R-loop 所引起的基因不穩定 (Santos-Pereira et al., 2013)。
DNA topoisomerase1 (TOP1) 具有抑制 R-loop 形成的功能,是演化上具有保 守性的因子 (El Hage et al., 2010)。抑制 R-loop 形成的可能原因為 TOP1 具有鬆 開 DNA 負超螺旋的能力,若缺乏 TOP1,則轉錄的過程中,負超螺旋的累積使 得 DNA 雙股螺旋打開,促進 RNA 與模板 DNA 重新配對而產生 R-loop (Drolet et al. 1995)。 2014 年發現,與 TPOI 同一家族的蛋白質, 拓樸異構酶 3B (TOP3B), 可以降低負超螺旋及 R-loop 的形成 (Yang et al., 2014)。TOP3B,與 methyl-arginine effector tudor domain-containing protein 3 (TDRD3) 產 生 交 互 作 用 可 辨 識 出 arginine methylation histone marks。目前提出的假說為 TDRD3-TOP3B 複合體會 被招喚至轉錄活躍的區域,TDRD3 辨識甲基化精胺酸組蛋白標誌區 (methyl- arginine histone marks) 使 Pol II 的 C-termanal domain (CTD)甲基化,而 TOP3B 解 開由 Pol II 所產生的負超螺旋,這與先前提到的 chromatin marks, H3S10P 擁有相
反的作用。
除了預防及解開 R-loop 的機制外,也存在著促進 R-loop 生成的機制。其中 一個是存在於 S. cerevisiae 中的 Rad51。真核生物的 Rad51 與細菌的 RecA 是同 源基因,在同源性重組扮演重要角色。2013 年研究發現缺乏 Rad51 的 S. cerevisiae 會降低 R-loop 的生成,以及後續由 R-loop 所帶來的基因不穩定 (Wahba at al.
2013)。
R-loop 依生成方式不同可分為 cis 及 trans 兩種,前者為與轉錄作用同時生 成,後者為由外部基因插入生成。研究指出,若在酵母菌中表現人類特定的 DNA 序 列 於 yeast artificial chromosome (YAC) 及 酵 母 菌 本 身 會 受 到 轉 錄 調 控 的 chromosome 中,轉錄開始後,這些被轉錄出的片段會入侵 YAC 中的同源基因,
造成遠離轉錄起始點的 trans R-loop 的生成,而 Rad51 可以幫助這些 trans r-loop 生成 (Wahba at al. 2013)。Trans 形式的 R-loop 相較於 cis R-loop 對於基因不穩定 性有更高的威脅,若轉錄作用發生在高度重複的的序列 (element),cis R-loop 只 會出現在特定的區域,然而 trans R-loop 可能會出現在許多位置,而製造出許多 基因不穩定度高的熱點 (hot spot)。
Trans R-loop 近年來也應用在基因修補上,稱為 CRISPR (clustered regularly interspaced short palindromic repeat)-Cas9 系統,CRISPR-Cas9 原先是微生物用來 對抗嗜菌體,以及其他外來基因片段的免疫系統。外來的 DNA 小片段插進宿主 DNA 的 CRISPR 基因位點 (loci),CRISPR 基因位點進行轉錄後, CRISPR RNA 進行 RNA 剪切產生成熟 (mature) 的 CRISPR RNA,其中有帶有 CRISPR 重複 序列的外來 RNA 序列會與 Cas9 酵素結合,而外來 RNA 序列辨認同外來基因位 置,產生了 trans R-loop,使外來基因靜默 (Terns and Terns, 2011)。
1.4 R-loop 的生理功能
R-loop 最早在 in vivo 中被提出之生理功能是在 B 細胞免疫蛋白重鏈上的類 型轉換重組 (class switch recombination, CSR)上。類型轉換重組基因位點序列長、
穩定、具有高度重複性以及富含 G 和 C,而類型轉換重組上的 R-loop 長度長且 安定,顯示出 R-loop 形成的難易度、穩定度和模板股上的基因序列顯著的關聯 性 (Yu et al., 2003)。
控制基因的表現需要定義基因的邊界: 5’端的啟動子及 3’端的終端子。R-loop 被報導常出現在基因的兩端 (Skourti-Stathaki et al., 2011; Ginno et al., 2012, 2013)。
在人類的基因組裡,R-loop 常出現在 5 端啟動子含有 CpG island 的下游,這些 區域通常是富含 G-C。而也有研究報導 R-loop 跟轉錄作用的中止反應有關,在 polyA 後形成的 R-loop 可以促使 Pol II 停止,達到轉錄終止的作用。當 R-loop 形 成後,轉錄停止,Senataxin 解開 R-loop 結後 Xrn2 促使 Pol II 離開基因,使得轉 錄終止。在這裡 R-loop 扮演雙重腳色,R-loop 的存在可以有效率的使轉錄終止,
但若在 Senataxin 突變株中,R-loop 大量累積,則會抑制這個過程(Skourti-Stathaki et al., 2011)。
1.5 R-loop 與非編碼 RNA (non-codng RNA, ncRNA)
在動物及植物中都有 R-loop 調控非編碼 RNA 的例子。普拉德-威利症候群 (Prader-Willi Syndrome, PWS) 以及天使症候群 (Angelman syndrome, AS) 是兩種 罕見的神經性遺傳疾病,發生原因為第 15 號染色體的基因缺失,主要是受到印 痕控制區域 (imprinting control region) 所調控,其中包含 Snrpn 基因的啟動子以 及複雜的非編碼 RNA,而非編碼 RNA 裡包括與 PWS 相關的 Snord116 基因所
產生的小片段 RAN 以及 Ube3a 基因的反義轉錄物 (antisense transcript)。已知 Ube3a 的缺乏會造成天使症候群,以及在神經專一的轉錄作用中,Ube3a 基因的 反義轉錄物會使父源的 Ube3a 靜默。
另外,2013 年也發現了在植物中 R-loop 調控非編碼 RNA 的例子。在阿拉 伯芥中,FLC 基因是抑制開花的主要因子,而 FLC 的反義轉錄物 COOLAIR 則 扮演著調控 FLC 基因表現量的角色。當植物資遭遇冷逆境後,COOLAIR 表現量 上升,進而抑制 FLC 表現,使得植物順利開花。COOLAIR 的啟動子區域會有 R-loop 形成,單股 DNA 結合蛋白, AtNDX, 結合其上之後,可以穩定該 R-loop 的結構,最終造成 COOLAIR 的轉錄受到抑制 (Sun et al., 2013)。
在 Ube3a 基因中,R-loop 的出現會造成染色質解凝,但在另一篇研究中發 現,R-loop 的生成會引起染色質凝聚 (condensation) (Castellano-Pozo et al., 2013),
兩個研究顯示 R-loop 的形成對於染色質的結構會造成完全相反的結果,表示 R-loop 與染色質的動態平衡可能有所關聯。
1.7 依 R-loop 的序列特徵建立計量模型
近幾年科學界中提出了一些關於 R-loop 中 DNA 的特徵,如可被轉錄的 DNA,DNA 序列鳥嘌呤含量高以及 DNA 的負超螺旋維持 R-loop 結構。因為 R- loop 中的 RNA 是由 RNA polymerase 在轉錄過程中所形成,所以唯有可以被轉 錄的 DNA 才能形成 R-loop。鳥嘌呤含量高則被視為 R-loop 形成的重要因子,
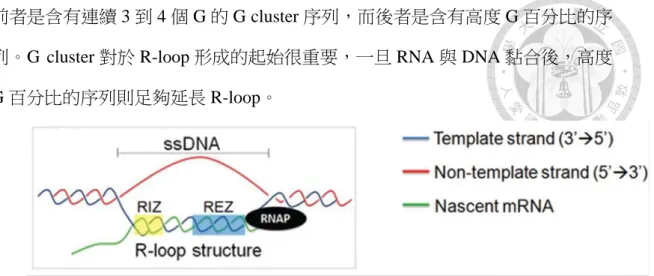
根據 in vitro 的實驗,科學家提出了 quantitative model of R-loop forming sequence (Roy and Lieber, 2009; Wongsurawat et al., 2012)。鳥嘌呤含量高的序列可分為兩種 特徵,R-loop initiation zone (RIZ) 以及 R-loop elongation zone (REZ) (Figure 1-1)。
前者是含有連續 3 到 4 個 G 的 G cluster 序列,而後者是含有高度 G 百分比的序 列。G cluster 對於 R-loop 形成的起始很重要,一旦 RNA 與 DNA 黏合後,高度 G 百分比的序列則足夠延長 R-loop。
Figure 1-1. The structure of the R-loop (Wongsurawat et al., 2012). R-loop initiation zone (RIZ) and R-loop elongation zone (REZ) are highlighted in yellow blue, respectively.
1.8 建立 R-loop 基因搜尋程式
本實驗室謝閔翔與台灣大學生物產業機電工程學系陳倩瑜老師實驗室合作 發展出 R-loop searching program,此程式是以 quantitative model of R-loop forming sequence (Roy and Lieber, 2009; Wongsurawat et al., 2012) 為基礎,利用 C++語言 所設計而來。此程式主要是尋找含有 R-loop forming sequence (RLFS) 的基因,
而 RLFS 由 RIZ 以及 REZ 所組成。本篇研究中,設定的參數值為 RIZ 為 50 鹼 基對,且序列中含有 4 個連續的 G,而 REZ 為 150 bp,序列中至少有 50% 的 G。尋找的範圍由基因的轉錄起始位置往後 1500bp 內,包含轉錄股與非轉錄股。
使用上述參數搜尋阿拉伯芥中可能含有 RLFS 的基因,得到 53 組結果 (Appendix 1)。
1.9 本篇論文之研究起源與動機
R-loop 在微生物及動物中已有許多重要科學研究,但在植物中,目前只知道 會調控 COOLAIR 的基因表現量。為了要探索在植物中更多 R-loop 的功能,首先
先找出可能會形成 R-loop 的序列,因此我們使用了生物資訊程式來搜尋序列特 徵相符合的基因,再搭配 gel mobility shift assay, 分析在 in vitro 情況下是否會形 成 R-loop 而造成泳動率改變。接著利用 native bisulfite sequencing 分析在 in vivo 是否有形成 R-loop,單股非模板 DNA 的存在。
利用以上方法,希望能找出更多可能會形成 R-loop 的基因以及未來進一步 找出 R-loop 對這些基因的作用機制,以了解更多 R-loop 在植物中的生理功能。
第二章 材料與方法
2.1 植物材料及生長條件
2.1.1 植物材料
本實驗使用的植物材料為 Arabidopsis thaliana Columbia ecotype (Col-0),其 種子購自 ABRC (Arabidopsis Biological Resource Center)。阿拉伯芥種子可於固態 培養基或培養土上種植,並置於溫控 22°C,照度 5000-7000 Lux 的植物生長箱中 培養,依照實驗條件所需可調整光照洲週期為長光照 (16 小時光照、8 小時黑暗) 或短光照 (12 小時光照、12 小時黑暗)。
2.1.2 植物阿拉伯芥培養基 1/2 MS
本實驗的固態培養基為 1/2 MS 培養基加上 1 % sucrose 及 0.8 %洋菜膠配製 而成,液態培養基則是不加入洋菜膠。以 1 L 固態培養基為例,取 2.2 g 1/2 MS salt 溶於約 800 mL 之 2 次水,再加入 10 g 蔗糖,調整 pH 值到 5.7~5.8,再補水 至 1 L,最後加入 8 g 洋菜膠。121°C高壓蒸氣滅菌 20 分鐘後,降溫至 60°C,於 無菌操作台分裝置培養皿中。
2.1.3 種子表面消毒及低溫處理
取適量的種子置於 1.5 mL 微量離心管,加入 1 mL 70%酒精,靜置一分鐘,
將酒精去除,加入 1 mL 無菌水清洗倒出,加入含有 15 %漂白水及 0.5 % Tween20 之消毒液 1 mL,上下搖晃 10 分鐘再倒出,最後以無菌水反覆沖洗種子,直到離
心管中水溶液不起泡為止。完成後以鋁箔紙包覆微量離心管避光,置於 4°C震盪 器緩慢旋轉 4 天,使種子同步發芽。
2.1.4 種子之無菌培養
於無菌操作台內將低溫處理後的種子以無菌水清洗數次,將種子懸浮在無菌 水中,使用無菌滴管吸取種子至 1/2 MS 固態培養基中,使種子均勻分布於培養 基中,再吸除多餘水分,於無菌操作台風乾 30 分鐘後,以石蠟膜封住培養皿後 放至植物生長箱中培養。
2.2 The gel mobility shift assay
將可能會形成 R-loop 的基因片段選殖入可雙向進行 in vitro transcription 的 T-A 載體建構質體,轉錄完後產物使用 RNase A 處理去除多餘單股 RNA,或再 同時加入 RNase H 進行處理,去除 DNA/RNA 雜合體上的 RNA,最後使用不含 ethidium bromide 之 1 %洋菜膠體進行電泳,染色後觀察其泳動率變化情況。
2.2.1 以 Phusion DNA polymerase 進 行 聚 合 酶 連 鎖 反 應 (polymerase chain reaction, RCP)
Phusion DNA polymerase (Thermal Scientific) 是一種高保真性之 DNA 聚合 酶,具有5’ 3’ DNA polymerase 及 3’ ->5’ exonuclease 活性,因此錯誤率大幅降 低。實驗條件如下:於 0.2 mL PCR 離心管中加入最終濃度為 1 倍的 Phusion GC buffer、0.2 mM dNTP、0.5 µM 專一性引子對、適量的模板 DNA 及 0.5 µL Phusion
制器內進行反應。反應條件為 98°C /30 秒,98°C /10 秒,55°C /20 秒,72°C/一分 鐘,以上四步驟重複 35 個循環,最後於 72°C作用 10 分鐘,產物可放置-20°C保 存。
2.2.2 DNA 瓊脂糖膠體電泳 (Agarose Gel Electrophoresis)
秤取適量瓊脂糖膠體 (依目標片段調整膠體濃度,一般使用 1 %),溶於 1 X TAE 緩衝液 (40 mM Tris (pH 7.6), 20 mM acetic acid 及 1 mM EDTA) 或 1 X TBE 緩衝液中(89 mM Tris (pH 7.6) , 89 mM boric acid, 及 2 mM EDTA),以微波爐加 熱至膠體完全溶解,冷卻後加入 ethidium bromide,混和均勻倒入鑄膠膜組中,
等待 30 分鐘使膠體凝固,選擇 50 V 或 100 V 進行電泳。
2.2.3 PCR 產物之分離與純化
為了提高 PCR 產物的品質並去除 PCR 反應中的多餘鹽類,本實驗利用 GeneJET PCR Purification Kit (Thermo Scientific) 純化 PCR 產物及瓊脂糖膠體中 的目標核酸片段,以利後續實驗進行。
GeneJET PCR Purification Kit: 將 RCR 產物加入 1 倍體積之 Binding Buffer,
混合液移至 GeneJET purification column 中 (最多 800 µL),離心 1 分鐘,去除過 濾液,加入 700 µL 之 Wash Buffer ,離心 1 分鐘,去除過濾液,再離心 1 分鐘。
將管柱取出移至新的 1.5 mL 微量離心管中,加入預熱之 50 µL 的 Elution Buffer,
離心 1 分鐘,所得之過濾液即為純化為之目標 DNA 片段。
GeneJET PCR Purification Kit: 將 RCR 產物進行瓊脂糖膠體電泳後,於 UV 燈箱上以刀片將含有目標片段之膠體接下,放至 1.5 mL 微量離心管中,加入 1
倍體積之 Binding Buffer,置於 55°C水浴直到膠體溶解 (約 10 分鐘),期間每隔 數分鐘取出上下搖晃,以增加溶解效率。將混合液移至 GeneJET purification column 中 (最多 800 µL),離心 1 分鐘,去除過濾液,加入 700 µL 之 Wash Buffer , 離心 1 分鐘,去除過濾液,再離心 1 分鐘。將管柱取出移至新的 1.5 mL 微量離 心管中,加入預熱之 50 µL 的 elution buffer,離心 1 分鐘,所得之過濾液即為純 化為之目標 DNA 片段。
2.2.4 TA cloning
利 用 Taq polymerase 將 純 化 好 的 目 標 DNA 3’ 端 多 帶 上 一 個 腺 嘌 呤 (Adenine),以 TA cloning 法使該 DNA 片段與 5’端多帶一個胸腺嘧啶 (thymine) 之 T vector 進行接合反應。先將 10 µL 的目標 DNA 片段與 2 mM dATP 及 Taq polymerase 在 72°C下反應 30 分鐘,再將 3’端接上腺嘌呤之產物與 pGM-T vector (GeneMark)、5 X Ligase Reaction Buffer、T4 DNA Ligase (Invitrogen) 混勻,補水 至 20 µL,在 16 °C下反應 16 小時,反應完之產物可直接進行大腸桿菌轉形。
2.2.5 大腸桿菌培養基
LB 成分包含 1 % tryptone、0.5 % yeast extract 及 1 % NaCl。配製 1 L LB medium 方法為秤取 25 g LB 粉末,加入二次水至體積 1 L,以高壓蒸氣滅菌法滅 菌,冷卻後即為液態培養基。若要配製固態培養基則是在滅菌前加入 15 g agar,
滅菌完後待冷卻至 55 °C,添加抗生素,於無菌操作台內分裝置培養皿中,可至 於 4 °C保存。
2.2.6 大腸桿菌熱衝擊轉形(Heat-shock Transformation)
取適量 DH5a 勝任細胞 (波仕特) 加入完成 TA cloning 之載體 DNA,劇烈震 盪 1 秒使其混合均勻,置於冰上 20 分鐘。接著以 42°C水浴作用 1 分鐘並置於冰
上 5 分鐘後,加入 1 mL 預冷之液態 LB,於 37°C 下培養一小時。之後以 6000 x g 離心一分鐘,去除多餘上清,留下 100 µL LB 菌體懸浮,在無菌操作台
內以無菌玻璃珠將菌液均勻塗抹於含有 100 µg/mL Ampcillin、0.5mM 的 IPTG 和 80µg/mL 的 X-gal 之 LB 固態培養基,於 37°C培養過夜。
2.2.7 菌落 PCR 檢定法
以微量吸管沾取菌落,加入最終濃度為 1x Taq 緩衝液、0.2mM dNTP、0.2µM 專一性引子對及 2.5U Taq DNA polymerase,混合均勻,並同時將菌液重新畫在新 的含有抗生素的固態培養基上。將離心管放入 PCR 溫度控制器中進行反應。PCR 反應條件為 95°C /五分鐘,98°C /20 秒,55°C /20 秒,72°C/1 分鐘,以上四步驟 重複 35 個循環,最後於 72°C作用 5 分鐘。反應完成後產物以瓊脂糖膠體電泳分 析,挑選帶有目標基因片段的菌落。
2.2.8.1 利用實驗套組進行質體 DNA 小量製備
使用 Plasmid DNA Mini Kit (Nautia) 進行質體 DNA 的小量製備。首先將帶 有目標載體之大腸桿菌培養於 3 mL 含有適當抗生素之 LB 液態培養基中,於 37°C 震盪培養過夜。隔天取 1.5 mL 菌液於微量離心管中,以 12000 x g 離心 1 分鐘,去除上清,重複以上步驟一次。加入 200 µL PS1 buffer,以微量吸管將菌 塊均勻懸浮,再加入 200 µL PS2 buffer,將離心管上下倒置 10 次使其均勻混和,
靜置 5 分鐘,再加入 300 µL PS3 buffer,快速且溫和地將離心管上下翻轉 10 次
使其均勻混和,以 12000 x g 離心 10 分鐘。取出上清至 spin column 中,以 12000 x g 離心 1 分鐘,將濾液去除並加入 400 µL wash buffer,12000 x g 離心 1 分鐘後 去除濾液,再離心 3 分鐘以去除殘餘之酒精。取出 column 至於室溫風乾 3 分鐘,
最後將 column 移至新的微量離心管中,加入預熱 55°C 之 30 µL elution buffer 於 濾膜上,靜置一分鐘後,以 12000 x g 離心 1 分鐘,收集之濾液即為質體 DNA。
2.2.8.2 以鹼裂解法進行質體 DNA 小量製備
首先將帶有目標載體之大腸桿菌培養於 3 mL 含有適當抗生素之 LB 液態培 養基中,於 37°C 震盪培養過夜。隔天取 1.5 mL 菌液於微量離心管中,以 12000 x g 離心 1 分鐘,去除上清,重複以上步驟一次。加入 100 µL MP I (25 mM Tris- HCl, pH 8.0; 10 mM EDTA, pH 8.0; 50 mM glucose),以微量吸管將菌塊均勻懸浮,
接著加入 200 µL MP II (0.2 N NaOH; 1% SDS; 使用前以 10 N NaOH, 10% SDS 稀釋混合配製),將離心管上下倒置 10 次使其均勻混和,置於冰上,再加入 150 µ L MP III,快速且溫和地將離心管上下翻轉 10 次使其均勻混和,置冰浴中 5 分 鐘,以 12000 x g 離心 5 分鐘。取出上清至新微量離心管中,加入 0.6 倍體積之 異丙醇,混合均勻,靜置室溫 2 分鐘。以 12000 x g 離心 10 分鐘,倒去上清液,
沉澱以 70 %酒精清洗兩次,每次各離心 1 分鐘。將上清液吸乾後,於 55°C 烘箱 中放置五分種使酒精完全揮發,再以 30 µL TE-8.0 回溶並於 55°C 加熱 5 分鐘,
產物即為質體 DNA。
2.2.9 胞外轉錄 (in vitro transcription)方法一
取 5 µg 之質體 DNA,加入 10 µL 10X RNA polymerase buffer (1X buffer: 40 mM Tris-HCl、6 mM MgCl2、1 mM DTT、2 mM spermidine)、0.5 mM NTP、2 µL (100 units) T7 RNA Polymerase (New England Biolabs) 或 3 µL (60 units) SP6 RNA
Polymerase (New England Biolabs),補 DEPC 水至 100 µL,再等分至於兩管 PCR 離心管,於 37°C 下反應 1 小時。反應完後的 100 µL 取兩份 45 µL 進行 RNase 處理,加入 4.5 µL RNase A (1 mg/mL) 或加 0.5U RNase H (Takara),於 37°C 下 反應一小時。反應完後,將約 50 µL 的產物至於新 1.5 mL 離心管中,加上 85 µL DEPC 水及 15 µL 3M Sodium Acetate (pH5.2),於抽風櫃中加入 150 µL Phenol,
震盪混勻後以 14000 x g 離心 10 分鐘,盡可能吸去下層溶液,再以 14000 x g 離 心 10 分鐘,吸取上層溶液至新的 1.5 mL 微量離心管,於抽風櫃中加入等體積之 Chloroform,震盪混勻後以 14000 x g 離心 10 分鐘,盡可能吸去下層溶液,再以 14000 x g 離心 10 分鐘,吸取上層溶液至新的 1.5 mL 微量離心管,進行異丙醇 沉澱。加入 1.1 倍體積之異丙醇,冰至-80°C 1 小時。完成後在 4°C 下以 14000 x g 離心 20 分鐘,去掉上清,加入 1 mL 70% 酒精清洗,在 4°C 下以 14000 x g 離 心 5 分鐘,倒去上清並盡可能將上清吸乾,於 37°C 烘箱中放置 10 分鐘使酒精完 全揮發,再用 20 µL DEPC 水回溶並於 37°C 加熱 10 分鐘。完成後將產物進行 1
%不添加 EtBr 之 TBE 瓊脂糖膠體電泳,電壓 50V,時間約 1.5 小時。電泳完成 後,進行 30 分鐘外染 EtBr,再至於清水中退去 EtBr,10 分鐘換一次水,再使用 照膠系統觀察結果。
2.2.10 胞外轉錄方法二
取 5 µg 之質體 DNA,加入 10 µL 10X RNA polymerase buffer、0.5 mM NTP、2 µ L (100 units) T7 RNA Polymerase (New England Biolabs) 或 3 µ L (60 units) SP6 RNA Polymerase (New England Biolabs),補 DEPC 水至 100 µ L,再等 分至於兩管 PCR 離心管,於 37°C 下反應 1 小時。反應完後的 100 µL 取兩份 20 µL 進行 RNase 處理,加入 2 µ L RNase A (1 mg/mL) 或加 0.5U RNase H (Takara),於 37°C 下反應一小時。反應完後,取 20 µL 進行 1 %不添加 EtBr 之 TBE 瓊脂糖膠體電泳,電壓 50V,時間約 1.5 小時。電泳完成後,進行 30 分鐘
外染 EtBr,再至於清水中退去 EtBr,10 分鐘換一次水,再使用照膠系統觀察結 果。
2.2.11 胞外轉錄方法三
取 5 µg 之質體 DNA,加入 10 µL 10X RNA polymerase buffer、0.5 mM NTP、
2 µ L (100 units) T7 RNA Polymerase (New England Biolabs) 或 3 µL (60 units) SP6 RNA Polymerase (New England Biolabs),補 DEPC 水至 100 µL,再等分至於 兩管 PCR 離心管,於 37°C 下反應 1 小時。將反應的 100 µL 取三份 30 µL 進行 EDTA 處理,終止 RNA polymerase 活性。加入 1/10 體積之 EDTA (100mM),於 70°C 下反應 20 分鐘。三份分別為不加入 RNase、1 µg RNase A 或加 1 µg RNase A 及 0.5U RNase H (Takara) ,後兩組先加入 1/10 體積之 MgCl2 (50mM)再行 RNase 處理,補 DEPC 水至 100 µL,於 37°C 下反應 1 小時。完成後將產物進行 1 %不添加 EtBr 之 TBE 瓊脂糖膠體電泳,電壓 50V,時間約 1.5 小時。電泳完成 後,進行 30 分鐘外染 EtBr,再至於清水中退去 EtBr,10 分鐘換一次水,再使用 照膠系統觀察結果。
第三章 結果與討論
先 前 本 研 究 室 謝 閔 翔 由 R-loop 搜 尋 程 式 所 找 出 的 53 個 候 選 基 因 中 (謝,2015,碩士論文) ,選出 LBD18 作為研究目標,藉由 gel mobility shift assay 及 native bisulfite sequencing assay 顯示 LBD18為可能形成 R-loop的基因。本研 究延續此主題,挑選更多 R-loop 搜尋所預測的候選基因進行三種實驗條件略有 不同的 gel mobility shift assay,比較不同實驗方法得到的結果,尋找最適合的 GMSA方法,也從中得知候選基因在胞外條件下,是否會形成DNA/RNA雜合體 結構。
3.1 利用 R-loop searching program 尋找阿拉伯芥中可能形 成 R-loop 的基因
本研究使用 R-loop 搜尋程式尋找在阿拉伯芥中可能會形成 R-loop 的基因,
希望能進一步了解 R-loop 結構在阿拉伯芥中是否有其他的生理功能。此程式撰 寫是根據 Lieber and Roy 的理論而來的在這個理論中 R-loop形成需要R-loop initiation zone (RIZ), linker 以及 R-loop elongation zone (REZ) 等三種序列特徵 (Wongsurawat et al., 2012)。而 R-loop 形成的第一步需要依靠 RIZ 中的 G cluster,所以G cluster的存在是非常重要的。再來是REZ中需要較高的 G 密度以 幫助延伸 R-loop的結構,但不必含有 G cluster。在進行搜尋時,需要給定的參 數包括: REZ的長度,REZ的G比例、RIZ+REZ的長度範圍 (由轉錄起始點 (transcription start site) 後開始計算), RIZ的長度,RIZ範圍內 G cluster 的數目 (連續三個G或四個G)。
本研究延續謝閔翔的碩士論文中利用R-loop 搜尋程式所尋找的基因,
其設定的參數是, REZ 長度為 150 鹼基對,其中至少包含50%的G,其座落的
範圍在轉錄起始點後 1500 鹼基對內,RIZ長度為 50 鹼基對,至少要包含一組 GGGG cluster。在以上參數設定下,R-loop 搜尋程式找出了 53個在阿拉伯芥中 可能會形成 R-loop 的基因 (Appendices 1)。本研究從以上的 53 個候選基因中,
挑選出 13 個基因進行選殖,接著以 gel mobility shift assay (GMSA) 和 native bisulfite sequencing 的方式研究以上基因在 in vitro 和 in vivo 的條件下,是否有 R-loop 結構存在。
3.1.1 挑選目標基因的條件
最初挑選基因的依據為優先選擇已被其他研究報導過的基因,如 AGP19/
At1g68725、DOT1/ At2g36120。為了進行大規模驗證,另外從 53 個候選基因中 挑選 20 個基因進行選殖,作為 GMSA 材料使用。原先的挑選依據為先將 53 個 基因依長度分群,1000 個鹼基對以內一群、1000~2000 個鹼基對一群及 2000 個 鹼基對以上一群,依各群占 53 個基因中的比例,挑選各群的個數。在進行 DNA 片段選殖時,考慮到選殖全長基因對於 R-loop 的形成可能有所影響,以 LBD18 為例,以全長 (2420 個鹼基對) 或包含 R-loop 形成序列的外顯子 (exon, 649 個 鹼基對) 進行 GMSA,得到不同的實驗結果 (謝, 2015, 碩士論文)。為了減少干 擾因子,本研究選擇選殖候選基因中包含最有機會形成 R-loop 的 600~700 個鹼 基片段,其中包含 RIZ+REZ。限制了轉殖序列的長度,也限制了引子對的位置,
候 選 基 因 中 成 功 由 引 子 對 設 計 網 站 設 計 出 引 子 對 個 數 為 19 個 (http://www.ncbi.nlm.nih.gov/tools/primer-blast/),這 19 個目標基因稱為 RL-1 ~ RL- 19,引子對分別命名為 RL-1-FP、RL-1-RF~RL-19-FP、RL-19-RF(Appendix )。使 用野生型阿拉伯芥基因組 DNA 作為模板 DNA,以 Phusion DNA polymerase 及引 子進行聚合酶連鎖反應對放大目標片段,再以瓊脂糖膠體電泳確定片段大小是否’
正確後,進行 PCR 產物之純化。若電泳結果出現若干片段,則選擇大小正確的
片段進行切膠純化。純化完後的產物與帶有 T7 及 SP6 的載體進行 TA cloning,
完成後,使用大腸桿菌進行熱衝擊轉形,再將大腸桿菌塗於含有 ampiclin、IPTG、
X-gal 的 LB 固態培養基,16 小時後進行藍白篩選。以藍白篩選挑出的菌落進行 colony PCR,確認挑選出的菌落內含轉型成功之大小正確的片段。最後進行質體 抽取,並將質體送定序,確認質體序列。19 個候選基因中成功選殖的共 11 個,
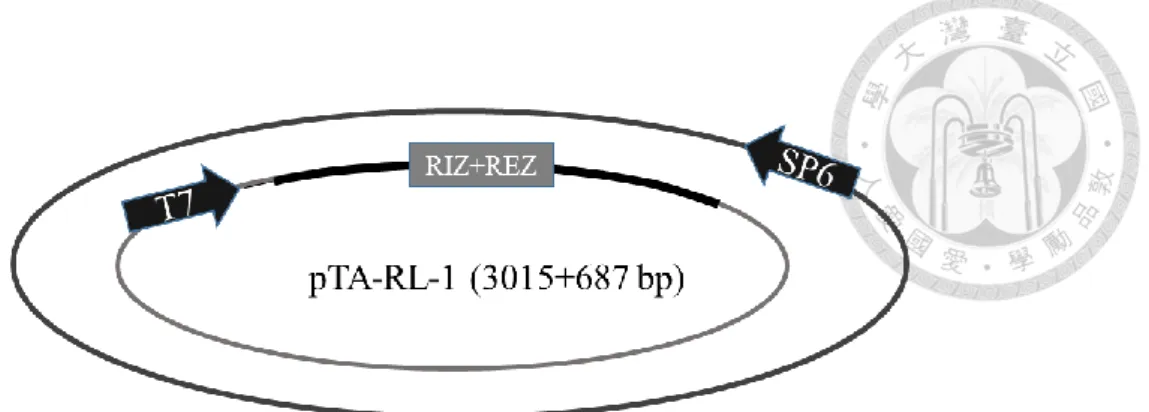
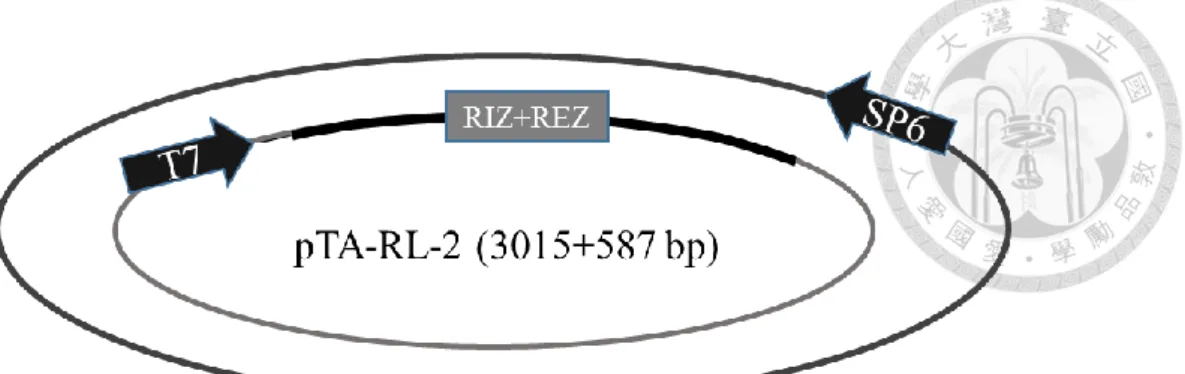
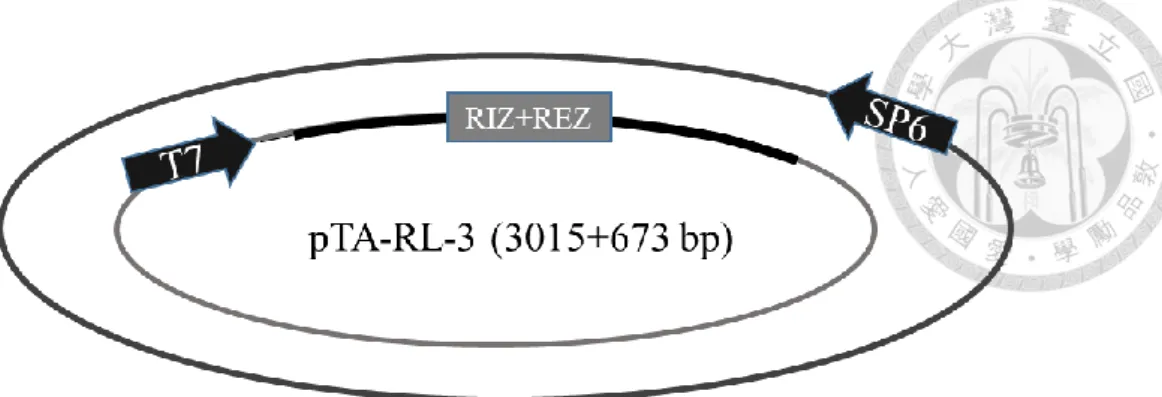
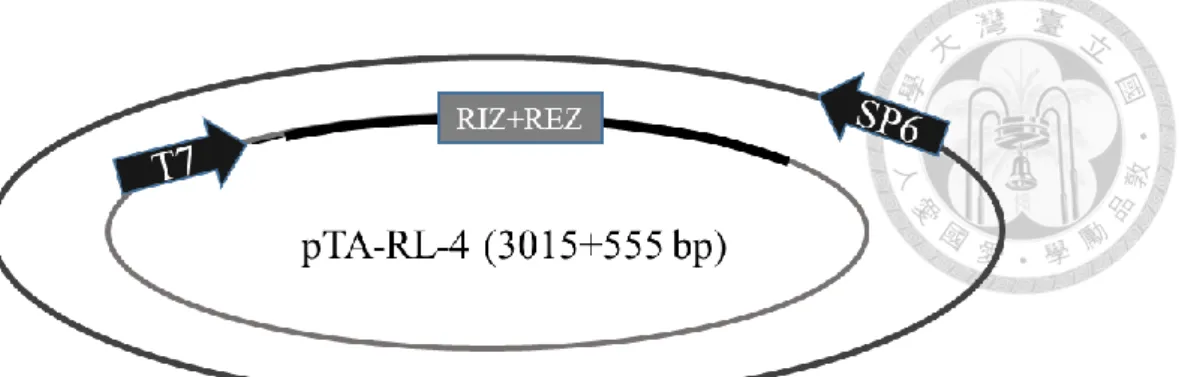
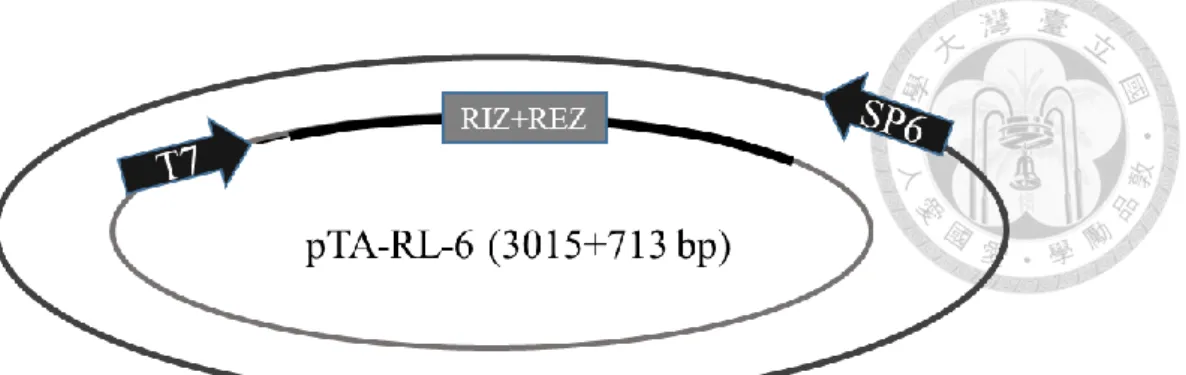
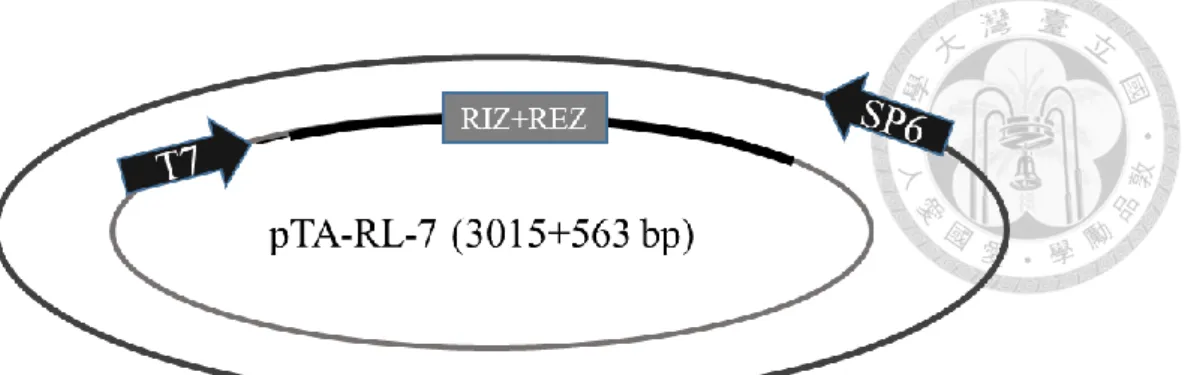
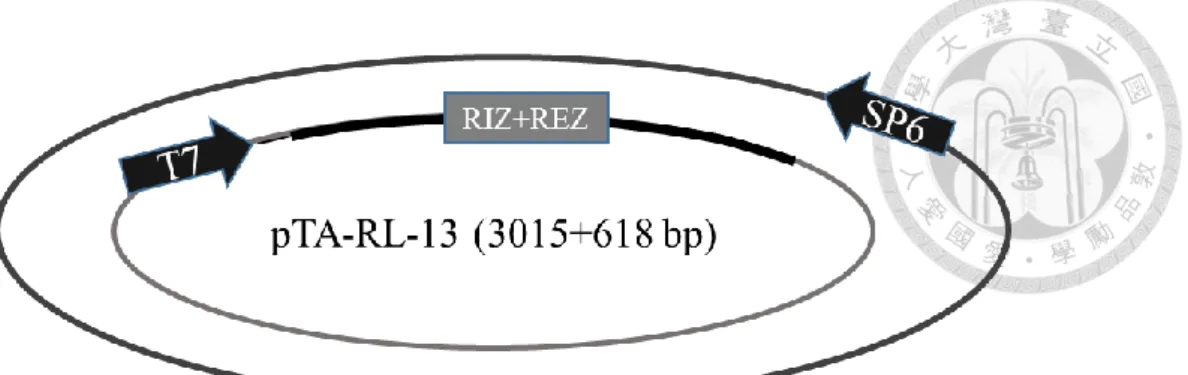
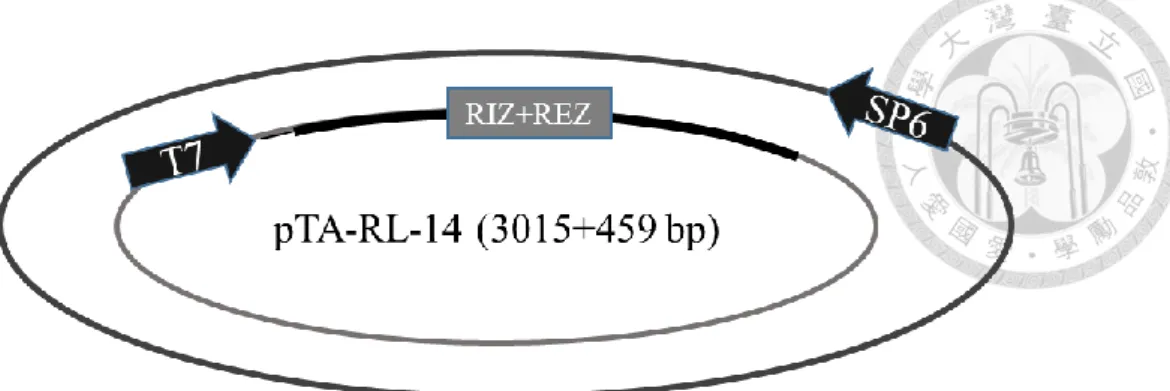
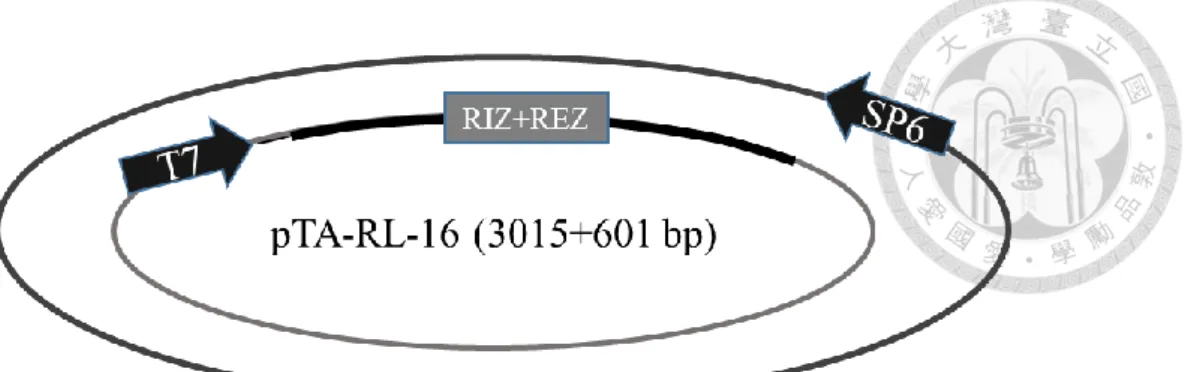
分別為 RL-1/At1g15830、RL-2/At1g15840、RL-3/At1g44191、RL-4/At1g62440、RL- 6/At1g77855 、 RL-7/At2g28490 、 RL-13/At4g33970 、 RL-14/At4g36230 、 RL- 16/At5g15780、RL-18/At5g53870、RL-19/At5g65390,其質體分別命名為 pTA-RL- 1 (figure 1c)、pTA-RL-2 (figure 2c、pTA-RL-3 (figure 3c)、pTA-RL-4 (figure 4c)、
pTA-RL-6 (figure 5c)、pTA-RL-7 (figure 6c)、pTA-RL-13 (figure 7c)、pTA-RL-14 (figure 8c)、pTA-RL-16 (figure 9c)、pTA-RL-18 (figure 10c)、pTA-RL-19 (figure 11c),
另外一開始挑選的 AGP19 及 DOT1 也選殖成功,其質體為 pTA-AGP19 (figure 12c)及 pTA-DOT1 (figure 13c)。上述 13 個質體中,RIZ+REZ 座落的位置皆於 T7 啟動子後。
3.2 以 Gel mobility sift assay 檢驗在 in vitro 條件下是否有 R-loop 結構存在
使用 GMSA 可以觀察到 DNA/RNA 雜合產生後泳動率的改變,而泳動率的 改變可能由兩個原因造成,(1) 含有 DNA/RNA 雜合體的質體,與沒有 DNA/RNA 雜合體的質體因構型不同而造成泳動率不同,(2)與 DNA 雜合的 RNA 片段長度 不一樣,使得泳動率有所差異。而泳動率不同的結果會反映在電泳圖上,讓電泳 圖有特徵性色帶模糊 (smear)的現象出現。RNase H 是一種可以分解 DNA/RNA 雜合體的酵素,若 GMSA 實驗結果上的色帶模糊在加入 RNase H 後消失了,則
可以推論色帶模糊是由 DNA/RNA 雜合體造成的。GMSA 包含質體進行胞外轉 錄( in vitro transcription)、RNase 處理及瓊脂糖膠體電泳分析。第一步為將選殖 成功的質體進行胞外轉錄,由於質體內含有 T7 及 SP6 兩種不同的啟動子,分別 加入 T7 及 SP6 polymerase 即可轉錄出兩股反義之 RNA。當 DNA 轉錄出含有高 度鳥嘌呤的 RNA 時,容易產生出 R-loop 結構,但轉錄出出含有高度胞嘧啶的 RNA 時,卻不會使得 R-loop 形成 (Ginno et al., 2012)。利用形成 R-loop 的序列 特徵,預期使用 T7 及 SP6 polymerase 進行胞外轉錄會得到不同結果。
質體進行完胞外轉錄後,為了去除多餘的 RNA 產物,後續進行 RNase A 處理,另外若電泳結果呈現出色帶模糊,為了分辨是否為 DNA/RNA 雜合體 所造成,需另外進行 RNase H 處理。完成胞外轉錄及 RNase 處理後,進行 phenol/chloroform 萃取和異丙醇沉澱,去除樣品中的蛋白質,最後進行 1 %不添 加 EtBr 之 TBE 瓊脂糖膠體電泳,電壓 50V,時間約 1.5 小時。電泳完成後,進 行 30 分鐘外染 EtBr,再至於清水中退去 EtBr。
本研究選擇 3 種條件略有不同的 GMSA 作為實驗方法 (材料與方法 2.2.9- 2.2.11)。方法一為完成胞外轉錄及 RNase 處理後,進行 phenol/chloroform 萃取 去除蛋白質和異丙醇沉澱,最後以電泳觀察結果。方法一可以確認在最後的電泳 結果上,不會有蛋白質殘留,可以除去因為有 polymerase 的存在、而使 DNA/RNA 雜合體產生的可能性或是 polymerase 與質體結合而產生色帶模糊的險象的可能 性,方法一是目前最為人採用的。
方法二為進行完胞外轉錄及 RNase 處理後,不做 phenol/chloroform 萃取,
以電泳觀察結果。方法二的好處是減少許多操作步驟,但樣品中會殘留沒辦法預 期是否會帶來干擾的 polymerase。
方法三為進行完胞外轉錄後,以 5 mM EDTA 螯合反應溶液內 6 mM 的鎂離 子,再以高溫 70°C 處理,使 polymerase 失去活性,也避免轉錄出的 RNA 在高 溫下被降解。後續因為 RNase H 需要鎂離子才有活性,故補上 5 mM 鎂離子後,
再進行 RNase 處理。反應完成後,以電泳觀察結果。方法三與方法二的差異是 方法三已經確定了 polymerase 在高溫處後失去活性,但是 polymerase 依然存在 於樣品中。另外,無法預期高溫處理會對 R-loop 結構造成的影響,有可能會使 DNA/RNA 雜合體結構被破壞。
值得一提的是,使用方法一進行 GMSA 時,是使用 1.1 倍體積的異丙醇沉 澱,而非 2.5 倍體積的酒精沉澱。若以 2.5 倍體積的酒精沉澱進行實驗,5 組實 驗組別幾乎沒有辦法同時回溶成功,原先認為是質體 DNA 的濃度太低,導致後 續進行完 phenol/chloroform 萃取回收的量不足以成功進行酒精沉澱。但將質體 DNA 的總量加倍,也沒有改善實驗結果。後改以加入 1.1 倍體積的異丙醇沉澱,
則可以成功得到完整的電泳結果。
3.3 13 個候選基因進行 GMSA
R-loop 的形成是由含有高密度 G 的 RNA 與模板 DNA 節結合,而非模板 DNA 形成單股 DNA。本實驗選殖的 13 個質體中,含有高密度 G 的 DNA 股皆 位於 T7 啟動子後,亦即加入 T7 polymerase 會轉錄出含有高密度 G 的 RNA,預 期加入 T7 polymerase 後,在電泳結果上可以看到色帶模糊出現。
三種 GMSA 的實驗方法除了在操作步驟上略有差異外,最後在電泳結果上 呈現的質體 DNA 濃度也有所不同。三種方法 GMSA 起始濃度理論上都相同,一 開始胞外轉錄反應溶液為 100 µL,但後續操作上的所取的體積有所差異。方法一 為 100µL 中取 45µL 進行後續實驗,而且因為有 phenol/chloroform 萃取去除蛋白 質的步驟,理論上會有 DNA 的損失。而方法二是 100 µL 中取 20 µL 進行後續操 作,只取 20 µL 的理由為最後進行電泳時,樣本槽可容納的最大體積約為 23 µL,
胞外轉錄後取 20 µL 加上後續添加的 RNase 及進行電泳時所需的追蹤染劑,體 積大約 23 µL。方法三則為 100 µL 中取 30 µL 進行後續操作,最終反應補 DEPC
水至 100 µL,取其中 20µL 進行電泳。而當初質體 DNA 的抽取方式為使用鹼裂 解 法 進 行 質 體 DNA 小 量 製 備 ( 第 二 章 材 料 與 方 法 2.2.8) , 沒 有 進 行 phenol/chloroform 萃取去除蛋白質,殘留的蛋白質可能干擾以 NanoDrop 測試質 體 DNA 濃度時的準確性。以方法三進行 GMSA 時,濃度參照 NanoDrop 的分析 結果。而後續以電泳分析再次判斷質體 DNA 的濃度,發現 NanoDrop 的分析結 果與實際上的濃度有所差異,故方法一、二與三的起始濃度理論上都為 5 µg 之 質體 DNA,但實際情況可能略有不同。
使用 pTA-RL-1 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以 觀 察到 色 帶模 糊 出現 (Figure 1A, 1B 和 1C) 。 三 種方 法都是 在 加 入 T7 polymerase 的組別可以觀察到色帶模糊出現,而這種現象在加入 RNase H 後消 失。比較 Figure 1A 和 1B 可以知道,進行 phenol/chloroform 萃取去除蛋白質對 實驗結果沒有很大的影響,而色帶模糊的型態為 supercoiled form 減少,在 supercoiled form 與 open circular form 之間出現色帶模糊,整體來說色帶模糊的位 置偏中下方 (Figure 1A, 1B)。但以方法三得到的實驗結果略有差異,色帶模糊的 型態為 supercoiled form 消失,supercoiled form 與 open circular form 之間出現非 常微弱的色帶模糊,幾乎看不見 (Figure 1C)。造成方法三沒有色帶模糊的原因可 能為高溫處理後,原先胞外轉錄後形成的 R-loop 結構被破壞,使得電泳圖上觀 察不到明顯的色帶模糊。
使用 pTA-RL-2 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以 觀 察到 色 帶模 糊 出現 (Figure 2A, 2B 和 2C) 。 三 種方 法都是 在 加 入 T7 polymerase 的組別可以觀察到色帶模糊出現,而這種現象在加入 RNase H 後消 失。三種方法得到的色帶模糊型態相同,為 supercoiled form 減少,在 supercoiled form 與 open circular form 之間出現色帶模糊,整體來說色帶模糊的位置偏中下
方。
使用 pTA-RL-3 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 3A, 3B 和 3C)。方法一與方法二得到的實驗結果 相似,色帶模糊的型態為為 supercoiled form 減少,在 supercoiled form 與 open circular form 之間出現色帶模糊,整體來說色帶模糊的位置偏中下方,而這種現 象在加入 RNase H 後消失 (Figure 3A, 3B) 。以方法三得到的實驗結果略有差異,
幾乎沒有色帶模糊出現 (Figure 3C),可能原因為經過高溫處理後,R-loop 結構 被破壞使得電泳圖上觀察不到色帶模糊。
使用 pTA-RL-4 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 4A, 4B 和 4C)。三種方法得到的實驗結果不盡相 同,使用方法一沒有色帶模糊出現 (Figure 4A)。而方法二與方法三的實驗結果相 似,雖然 supercoiled form 消失,但 supercoiled form 與 open circular form 中沒有 明顯色帶模糊出現, (Figure 4B 和 4C)。比較方法一與方法二、方法三所得到的 實驗結果,造成其中差異的原因可能為 phenol/chloroform 萃取步驟,方法二與方 法三的實驗結果中,supercoiled form 消失可能是由於 polymerase 的干擾所造成。
使用 pTA-RL-6 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 5A, 5B 和 5C)。方法一、二與三得到的實驗結果 都沒有觀察到明顯的色帶模糊,可以推論在在三種實驗條件下,pTA-RL-6 質體 不會形成 DNA/RNA 雜合體結構。
使用 pTA-RL-7 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 6A, 6B 和 6C)。由方法一及方法三得到的實驗結
果類似,色帶模糊的型態為 supercoiled form 減少,supercoiled form 與 open circular form 之間出現階梯狀色帶模糊,造成階梯狀的色帶模糊的可能原因為選殖片段 上有重複序列(Appendix 8),而這種現象在加入 RNase H 後消失 (Figure 6A 和 6C)。由方法二所得到的實驗結果為 supercoiled form 減少,色帶模糊不明顯,但 可以看到微弱的階梯狀色帶模糊,而這種現象在加入 RNase H 後消失 (Figure 6B)。
造成方法二與其他兩組實驗結果上的差異有可能為質體 DNA 的濃度偏低,即使 有色帶模糊出現也沒有辦法使用肉眼觀察到。
使用 pTA-RL-13 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 7A, 7B 和 7C)。由方法一及方法三得到的實驗結 果類似,色帶模糊的型態為 supercoiled form 減少,open circular form 增加,色帶 模糊出現的位置偏向 open circular form,而這種現象在加入 RNase H 後消失 (Figure 7A 和 7C)。此種色帶模糊型態最為接近 R-loop 形成時,supercoiled form 與 RNA 結合後,形成 open circular form 的情形。但由方法二的實驗結果觀察不 到色帶模糊出現(Figure 7B)。
使用 pTA-RL-14 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 8A, 8B 和 8C)。三種方法得到的實驗結果不盡相 同。方法一的色帶模糊型態為 supercoiled form 減少,open circular form 增加,
supercoiled form 與 open circular form 中有色帶模糊出現,而這種現象在加入 RNase H 後消失 (Figure 8A)。方法二的實驗結果為 supercoiled form 減少,但沒 有明顯色帶模糊出現,而這種現象在加入 RNase H 後消失 (Figure 8B)。方法三 為 supercoiled form 消失,supercoiled form 與 open circular form 中有些許階梯狀 色帶模糊,但選殖片段上沒有明顯重複序列 (Appendix 9 ),而這種現象在加入 RNase H 後消失 (Figure 8C)。
使用 pTA-RL-16 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 9A, 9B 和 9C)。使用方法一得到的結果為 supercoiled form 與 open circular form 中有些色帶模糊出現,這種現象在加入 RNase H 後消失 (Figure 9A)。使用方法二與方法三得到的實驗結果為沒有色帶 模糊出現 (Figure 9B 和 9C)。方法一觀察到的色帶模糊可能為過多的質體 DNA 所造成,而不是 DNA/RNA 雜合體出現所呈現的色帶模糊。
使用 pTA-RL-18 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 10A, 10B 和 10C)。方法一所得到的色帶模糊型 態為 supercoiled form 減少,open circular form 增加,兩者之間有些許階梯狀色帶 模糊但,選殖片段上有明顯重複序列 (Appendix 11 ),這種現象在加入 RNase H 後消失 (Figure 10A)。方法二與方法三得到的實驗結果為沒有色帶模糊出現 (Figure 10B 和 10C)。
使用 pTA-RL-19 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可 以觀察到色帶模糊出現 (Figure 11A, 11B 和 11C),三種方法得到的實驗結果皆沒 有觀察到色帶模糊出現,可以推論在在三種實驗條件下,pTA-RL-19 質體不會形 成 DNA/RNA 雜合體結構。
使用 pTA-AGP19 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別 可以觀察到色帶模糊出現 (Figure 12A, 12B 和 12C)。方法一與方法三得到的實 驗結果相同,沒有觀察到色帶模糊出現 (Figure 12A 和 12C)。而方法二的色帶模 糊型態為 supercoiled form 減少,靠近 supercoiled form 的位置有些許色帶模糊出 現,這種現象在加入 RNase H 後消失 (Figure 12B)。
使用 pTA-DOT1 質體進行三種 GMSA,預期在加入 T7 polymerase 的組別可以 觀察到色帶模糊出現 (Figure 13A, 13B 和 13C)。三種方法得到的實驗結果類似,
色帶模糊型態皆為 supercoiled form 減少至接近消失,open circular form 明顯增 加,兩者之間沒有明顯色帶模糊出現。造成此種現象的原因可能與 pTA-RL-13 質 體得到的類似實驗結果 (Figure 7) 的原因相同,此種色帶模糊型態最為接近 R-loop 形成時,supercoiled form 與 RNA 結合後,形成 open circular form 的情 形。但比較兩者的實驗結果還是略有差異,Figure 7A 與 7C 可以觀察到色帶模糊 出現,但 Figure 13 則否。
由 13 種質體 DNA 作為材料,比較三種實驗方法,可以得到相同實驗結果 的質體有 pTA-RL-2 (Figure 2)、pTA-RL-6 (Figure 5)、pTA-RL-19 (Figure 11)及 pTA- DOT1 (Figure 13),其中 pTA-RL-6、pTA-RL-19 沒有色帶模糊出現。
方法一、二結果相同,方法三不同的組別有 pTA-RL-1 (Figure 1)、pTA-RL-3 (Figure 3) 。 造 成 實 驗 結 果 不 同 的 可 能 原 因 為 方 法 三 經 過 高 溫 處 理 , 破 壞 DNA/RNA 雜合體結構。
方法二、三相同,方法一不同的組別有 pTA-RL-4 (Figure 4)、pTA-RL-16 (Figure 9)、pTA-RL-18 (Figure 10)。造成實驗結果不同的可能原因為方法一多操 作一步 phenol/chloroform 萃取,去除核酸以外的雜質,而方法二與方法三得到的 實驗結果有可能有雜質干擾。
方法一、三相同,方法二不同的組別為 pTA-RL-7 (Figure 6)、pTA-RL-13 (Figure 7)、pTA-AGP19 (Figure 12)。在 pTA-RL-7 中,造成實驗結果不同的可能 原因為質體 DNA 的濃度,pTA-RL-7 在方法二中的含量偏少。但目前沒有辦法解
釋 pTA-RL-13、pTA-AGP19 中,實驗結果的差異。
最後,三個方法得到不同的實驗結果的組別為 pTA-RL-14 (Figure 8),造成 實驗結果不同的可能原因有高溫處理破壞 DNA/RNA 雜合體結構與實驗結果有 可能有核酸以外的雜質干擾。三種 GMSA 實驗方法得到的實驗結果沒有一定的 規律性,高溫處理與雜質存在對於 GMSA 的實驗結果可能有一定的影響,日後 若要進行 GMSA,以方法一操作可能最為恰當。
而非由 R-loop 預測程式所找出的基因-PHY B,HHP1 及 ICE1,做為實驗的 負控制組,使用方法三作為實驗方法。結果顯示,都沒有色帶模糊的現象出現 (Figure 14)。
從本實驗所選擇的 13 個基因以 GMSA 觀察,最有可能形成 R-loop 結構的 為 RL-13 與 DOT1。為了瞭解由同一種參數條件下選出來的基因,為什麼在 GMSA 的得到不同的實驗結果,而進行 RIZ 和 REZ 序列分析。
首先從 REZ 區域的 G 富含程度來看,由於本研究給定的參數是 REZ 中至少 要有 50% 的 G,分析後發現可以看到 RIZ 區域的 G 富含程度除了 RL-7 是 89%
及 DOT1 是 54% 外,其他基因都是在 REZ 只有 50%的 G,這或許可以解釋 RL- 7 較明顯的色帶模糊 (Table 1)。
除此之外,RIZ 中的 G cluster 是形成 R-loop 的重要因子,所以我們也分析 了 RIZ 中 G cluster 的型態,由結果可以發現, RL-7、RL-14、RL-16,及 DOT1 在 G cluster 的分析上與其他基因有所不同。以上基因除了程式已找到的 GGGG cluster 之外,在 RL-7,RL-14 及 DOT1 中分別有一個 (RL-7)、一個 (RL-14)、兩 個 (DOT1)的 GGG cluster,而 RL-14 及 RL-16 各出現一個 GGGGG cluster。而其
他基因則只有一個 GGGG cluster (Table 1)。比較特別的是,謝閔翔先前選擇的研 究目標基因 LBD18,由序列分析發現,在 RIZ 的區域裡,除了原先設定的 GGGG cluster 外,沒有任何額外的 G cluster,本研究也對 LBD18 進行三種 GMSA。使 用含有 LBD18 全長的質體或包含 RIZ+REZ 的 LBD18 exon1 質體進行 GMSA,
加入 T7 polymerase 或 SP6 polymerase 皆沒有色帶模糊出現 (Figure15 和 16)。
以目前的實驗結果與序列分析尚未能得到一定的關聯性,或許在選殖序列的 RIZ 中沒有額外的 G cluster 對於 GMSA 有很大的影響,如 LBD18 在 GMSA 的 實驗結果觀察不到色帶模糊,但這需要更多的實驗結果來支持這項說法。
第四章 未來展望
本研究由 R-loop 搜尋程式所預測的 53 個基因中,挑選 13 個基因做為研究 目標,目前尚未發現 GMSA 實驗結果與候選基因之間的關聯性。為了進一步驗 證 R-loop 搜尋程式是否能更準確預測可能形成的 R-loop 的基因,可以從 53 個 基因中挑選更多基因進行研究,以增加樣本數。另外也從非預測基因中,選擇更 多基因進行研究,觀察是否有不具有原先設定的序列特徵而能形成 R-loop 的基 因,如 COOLAIR (Sun et al., 2013)。以 COOLAIR 進行 GMSA 結果顯示,沒有色 帶模糊出現 (Appendix ),暗示著會形成 R-loop 的基因不一定有序列特徵性。
除了 GMSA 外,還可以利用 native bisulfite sequencing assay 尋找細胞內的 R-loop 。當細胞內形成 R-loop 時,非模板股會形成單股 DNA 裸露在外,而此 時以 bisulfite 處理後,會使 DNA 上的 dC 轉換成 dU,接著進行進行聚合酶連鎖 反應,得到的產物則會出現 dC 到 dT 轉換的片段 (簡稱 C-T 轉換)。我們可以藉 由這些轉換片段知道由程式預測出的基因是否如預期一樣有 R-loop 的形成。
Native bisulfite sequencing assay 的第一步是抽取阿拉伯芥的基因組 DNA (genomic DNA) ,謝閔翔 (2015)的實驗流程是將植株均質化後,加入 urea- containing extraction buffer (7 M urea, 0.3 M NaCl, 50 mM Tris-HCl, pH 8.0, 20 mM EDTA, 及 1% sarcosine),接著使用 Phenol:Chloroform:Isoamyl Alcohol(PCI)去除 蛋白質跟脂質,異丙醇沉澱後即可得到基因組 DNA。使用 urea extraction buffer 是藉由 urea 讓蛋白質後,以及破壞 DNA 與水分子之間的氫鍵,再藉由 PCI 萃取 蛋白質,進而純化出基因組 DNA。
但有另一種更適合的抽取阿拉伯芥的基因組 DNA 的實驗方法: 使用 Honda
buffer (0.44 M Sucrose, 1.25 % Ficoll, 2.5 % Dextran T40, 20 mM Hepes KOH pH7.4, 10 mM MgCl2, 0.5 % Triton X-100, 5 mM DTT 及 Protease Inhibitors) 分離細胞 核,並以 miracloth 去除雜質,再使用 SDS 打破細胞核,以及利用 proteinase K 分 解蛋白質。最後使用 phenol/chloroform 萃取去除雜質,進行酒精沉澱即可得到基 因組 DNA。
本研究曾沿 續謝閔翔選擇的 LBD18 為研究目標,進行 native bisulfite sequencing assay,希望可以利用更適合植物的實驗方法,找出 LBD18 在細胞內 會形成 R-loop 的證據,但未能成功。原先認為是 native bisulfite sequencing assay 實驗上的條件不適當,但以含有 LBD18 序列的質體進行 GMSA 後觀察不到色帶 模糊出現,有可能是 LBD18 本來就為不會形成 R-loop 的基因,未來可以挑選 RL- 13 或 DOT1 作為研究目標進行 native bisulfite sequencing assay。
另外,未來也希望可以進行 DNA/RNA immunoprecipitation (免疫沉澱),
DNA/RNA immunoprecipitation 的實驗原理為利用可以偵測 DNA/RNA 雜合結構 的抗體, S 9.6, 結合 DNA/RNA 雜合體,搭配針對目標基因設計的引子對,進一 步使用即時定量聚合酶鏈鎖 (real time qPCR),觀察實驗結果推測目標基因是否 會形成 R-loop。另外,免疫沉澱完所得片段,可利用次世代定序 (Next Generation Sequencing , NGS),尋找出阿拉伯芥中可能會形成 R-loop 的基因。
參考文獻
Aguilera, A. (2002). The connection between transcription and genomic instability.
The EMBO Journal 21, 195-201.
Aguilera, A., and Garcia-Muse, T. (2012). R loops: from transcription byproducts to threats to genome stability. Molecular cell 46, 115-124.
Becherel, O.J., Yeo, A.J., Stellati, A., Heng, E.Y.H., Luff, J., Suraweera, A.M., Woods, R., Fleming, J., Carrie, D., McKinney, K., Xu, X., Deng, C., and Lavin, M.F. (2013). Senataxin plays an essential role with dna damage response proteins in meiotic recombination and gene silencing. PLoS Genet 9, e1003435.
Boubakri, H., de Septenville, A.L., Viguera, E., and Michel, B. (2010). The helicases DinG, Rep and UvrD cooperate to promote replication across transcription units in vivo. The EMBO Journal 29, 145-157.
Breslauer, K.J., Frank, R., Blöcker, H., and Marky, L.A. (1986). Predicting DNA duplex stability from the base sequence. Proceedings of the National Academy of Sciences of the United States of America 83, 3746-3750.
Castel, S.E., and Martienssen, R.A. (2013). RNA interference in the nucleus: roles for small RNAs in transcription, epigenetics and beyond. Nature reviews.
Genetics 14, 100-112.
Castellano-Pozo, M., Santos-Pereira, José M., Rondón, Ana G., Barroso, S., Andújar, E., Pérez-Alegre, M., García-Muse, T., and Aguilera, A. (2013).
R loops are linked to histone H3 S10 phosphorylation and chromatin condensation. Molecular cell 52, 583-590.
Cerritelli, S.M., and Crouch, R.J. (2009). Ribonuclease H: the enzymes in eukaryotes.
The FEBS journal 276, 1494-1505.
Cerritelli, S.M., Frolova, E.G., Feng, C., Grinberg, A., Love, P.E., and Crouch, R.J.
(2003). Failure to Produce Mitochondrial DNA Results in Embryonic Lethality in Rnaseh1 Null Mice. Molecular cell 11, 807-815.
Chakraborty, P., and Grosse, F. (2011). Human DHX9 helicase preferentially unwinds RNA-containing displacement loops (R-loops) and G-quadruplexes.
DNA Repair 10, 654-665.
Chinchilla, K., Rodriguez-Molina, J.B., Ursic, D., Finkel, J.S., Ansari, A.Z., and Culbertson, M.R. (2012). Interactions of Sen1, Nrd1, and Nab3 with multiple phosphorylated forms of the Rpb1 C-terminal domain in Saccharomyces cerevisiae. Eukaryotic cell 11, 417-429.
Crick, F.H. (1958). On protein synthesis. Symp Soc Exp Biol 12, 138-163.
Di Noia, J., and Neuberger, M.S. (2002). Altering the pathway of immunoglobulin