DOI:10.6152/jaa.2016.12.0005
從人骨和獸骨之骨膠原碳與氮穩定同位素組成看
圓山文化人的攝食特徵
李政益
*、陳瑪玲
、林立虹
***、
Peter Ditchfield
****、
王珮玲
*****、林秀嫚
******、
A. Mark Pollard
*******、羅清華
********、蔡錫圭
*********摘 要
人骨與獸骨同位素組成分析已被成功應用於解析史前人類的攝食特徵和生 業系統。本研究嘗試引此分析圓山遺址所出土的七具人骨和三具獸骨樣本之骨膠 原碳、氮同位素組成,進而探討圓山人的攝食體系。分析結果顯示圓山人的飲食 成分以 C3食物源,例如 C3植物和以 C3類食物為食的動物(鹿和豬),以及海 洋食物源(可能主要來自貝類)為主,至於 C4食物源的貢獻則較小。此一結果 與先前由生態遺留和出土器物等證據推論所得的攝食特徵與生業型態是吻合 的。綜合言之,本研究成果不僅呼應前人研究,即圓山人以狩獵採集,並行 C3 植物耕作為其生業活動;結合同位素分析結果和考古證據,則將此認識更加深化。 關鍵字:古攝食特徵、生業策略、碳與氮同位素、骨膠原、圓山文化* 國立臺灣大學地質科學研究所博士後研究員 國立臺灣大學人類學系教授 *** 國立臺灣大學地質科學研究所教授 **** 牛津大學考古暨美術史研究所穩定同位素研究室主任 ***** 國立臺灣大學海洋研究所副教授 ****** 國立臺灣史前文化博物館博士級助理研究員 ******* 牛津大學考古暨美術史研究所教授 ******** 國立臺灣大學地質科學研究所教授 ********* 國立臺灣大學醫學院教授
Paleodietary pattern of the Yuan-Shan Neolithic people
in Taiwan:
Evidence from carbon and nitrogen stable isotopic
analyses of human skeletons and faunal remains
Cheng-Yi Lee
*, Maa-Ling Chen
**, Li-Hung Lin
***, Peter
Ditchfield
****, Pei-Ling Wang
*****, Hsiu-Man Lin
******, A. Mark
Pollard
*******, Ching-Hua Lo
********, Hsi-Kuei Tsai
*********ABSTRACT
The application of isotopic analysis on human skeletons and faunal remains has successfully provided insights into the dietary patterns and subsistence systems of prehistoric populations. In this study, seven human skeletons and three faunal remains collected from the Yuan-Shan archaeological site were subjected to carbon and nitrogen isotopic analyses in order to investigate the dietary pattern of Yuan-Shan people. The isotopic results indicated that their diet may have consisted of mainly C3-based foods, such as C3 plants and the animals (e.g. deer and pig) that consumed C3 foods, and marine resources, potentially marine shellfish. The contribution of C4-based foods to the overall diet seemed to be minor. The isotope results conform to the previous paleodietary studies based on the evidences of ecofact and artifact remains of Yuan-Shan site. All available lines of evidence suggest
* Post-Docs Fellow, Department of Geosciences, National Taiwan University ** Professor, Department of Anthropology, National Taiwan University *** Professor, Department of Geosciences, National Taiwan University
**** Stable Isotope laboratory Manager, Research Laboratory for Archaeology and the History of Art, University of Oxford
***** Associate Professor, Institute of Oceanography, National Taiwan University
****** Assistant Research Fellow, Division of Research and Collection, National Museum of Prehistory
******* Edward Hall Professor of Archaeological Science, Research Laboratory for Archaeology and the History of Art, University of Oxford
******** Professor, Department of Geosciences, National Taiwan University ********* Professor, College of Medicine, National Taiwan University
that the Yuan-Shan people practiced hunting, gathering, and C3-plant cultivation as their subsistence activities.
Keywords: paleodietary pattern, subsistence strategies, carbon and nitrogen isotopes, bone collagen, Yuan-Shan Culture.
前言
關於史前人群的攝食特徵和生業體系等資訊,可藉由遺址中所出土的器物功能分 析和生態遺留種類來進行推測。可是這類研究往往僅能提供一個概略的定性描述。器 物和生態遺留也容易受到埋藏環境的影響,其中植物殘餘尤其不易被保存(Zhao 2011),因此可能無法提供完整且確實的攝食資訊。此外生態遺留只能顯示群體層次的 攝食與生業策略,而非個體層次。相較之下,人類與動物骨骸遺留的骨膠原(collagen) 碳、氮同位素組成分析可同時提供個體及群體層次的攝食習性資訊,不僅能用以評估 各種食物在一個社會中的相對重要性,從而推論出該人群的生業體系和經營策略;更 有助於比較個體間的攝食差異,觀察一人群中可能存在的社會結構或性別差異等文化 社會現象。 以骨膠原碳、氮同位素組成重建古食譜的研究起源於 70 年代末。包括玉米農業在美洲的引入時間和傳播路徑(參見van der Merwe and Vogel 1978),以及史前人類飲食
中的海洋食物和陸地食物的相對重要性(參見Tauber 1981)。隨著分析技術的進步,其
後有許多類似的研究持續且大量地被執行(參見 DeNiro and Epstein 1978, 1981; Lee-Thorp 2008; Pollard and Heron 2008; Schoeninger 2011; Schwarcz and Schoeninger 2012)。但此一分析技術卻尚未被廣泛應用於臺灣的考古骨骼遺留上。推究其中一個可 能的原因為溫暖潮濕地區的骨骼遺留,容易快速被成岩同位素置換作用(diagenetic isotopic exchange processes)1影響,導致骨膠原流失或其化學成分受到置換而失去其原 始的同位素訊號,因此較不適合進行同位素攝食研究(Krigbaum 2003)。目前已知的少 數研究結果似乎也支持此一觀點。例如米田穣等(2008)曾分析臺灣 19 個布農族人骨 以及八個史前遺址的 21 具人骨之骨膠原碳、氮同位素組成,結果顯示 21 具史前人骨 中,僅有七具個體保存了原始的生物同位素訊號。而這七具人骨分別出自六個不同的 遺址,因此各遺址的樣本數尚不足夠進行任何議題的深入討論。除上述研究外,陳光 祖(2004)於其回顧文章中提及 Tsang(1992)曾討論過澎湖鎖港遺址的一筆人骨碳、 氮同位素分析資料,但分析結果與考古證據有所牴觸,因此需要更多的研究來釐清。 另李匡悌(1995)亦曾於鵝鑾鼻第二遺址以及龜山遺址執行同樣的分析工作,認為該區 域的史前人群以海洋資源和 C4植物為食;但考古證據和植矽石分析結果卻似乎較強調 稻米(C3植物)的重要性(李作婷、鄭建文 2008)。 有鑑於此,本研究試圖針對台灣的人骨和獸骨遺留樣本進行系統性採樣,並嘗試
粹取骨膠原以分析其碳、氮同位素組成。根據可取得的材料,本研究選擇圓山遺址作 為初步研究對象,其研究旨趣包括: 一、評估圓山遺址的骨骼遺留是否適合進行同位素攝食研究。我們將以骨骸遺留 中的骨膠原產量和碳、氮元素莫耳比做為評估準則。 二、根據圓山遺址所出土的圓山文化遺留如器物和各樣生態遺留,學者描繪出的 圓山人之攝食體系極為複雜,食物資源也相當繁多。因此,期冀藉由分析圓山人骨膠 原碳、氮同位素組成,或可進一步獲得關於圓山人攝食體系與生業策略的明確資訊, 例如在這樣一個採用廣泛攝食體系的社會中,那些食物資源可能是更為重要的。 三、此類研究在臺灣地區仍屬起步階段,因此本研究將可建立並累積臺灣地區的 區域性生物同位素基礎值,以供臺灣地區或鄰近其它區域後續的同位素攝食研究比較 之基礎。
同位素攝食研究的原理與限制
骨頭組織由於會不斷更新,其更替(bone turnover)時間尺度長達 10 至 15 年,因 此人骨中的元素和同位素能記錄一個個體的長期平均攝食特徵 (Hedges et al. 2007)。 人骨中的元素及同位素組成已被證實與人類所吃的食物有關,因此可藉以重建史前人 群的攝食特徵和生業策略(Lubell et al. 1994; Polet and Katzenberg 2003; Richards et al. 2006)。骨頭可概略分為骨膠原和骨頭礦物兩個部分,其中骨膠原的主要組成元素包括 碳(C),氮(N),氧(O),氫(H)和硫(S),而又以碳和氮同位素組成最常被 用來探討個體的攝食特徵。 同位素組成(δ 值)的定義如下: δ (‰) = ((Rsample/Rstandard) – 1) 1000。 其中R 為某元素的重同位素除以輕同位素的莫耳比值,例如13C/12C 或15N/14N。碳同位 素的國際通用標準品為VPDB(Vienna Pee Dee Belemnite),而氮同位素的國際通用標 準品為空氣。骨膠原的碳、氮同位素組成能顯示所攝取的食物種類,亦即有機碳、氮的供應 源。普遍而言,有機碳的供應源可概略分為陸地源或海洋源;各自又可根據其初級生 產者的光合作用路徑區分成C3、C4和CAM(crassulacean acid metabolism)三大類型。
光合作用的進行可將大氣中的二氧化碳(δ13C = -7 ‰,Chisholm et al. 1982)經過一連串 的生化反應而轉換成為植物組織。在此過程中,植物可讓碳同位素發生分化(亦即植物 偏好使用12C 做為組織合成的原料)。其中 C3植物會產生較大的碳同位素分化量,因此 δ13C 值較低。以陸域環境為例,C 3植物的組織δ13C 值介於-35 – -22 ‰(平均為 -27 ‰, Farquhar et al. 1989) 之間。相比之下,C4植物組織的δ13C 值則介於-16 – -9 ‰之間(平 均為 -12.5 ‰,Farquhar et al. 1989)。由於 CAM 植物能依照環境的乾燥程度選擇使用 C3或C4光合作用路徑,因此CAM 植物的光合作用對碳同位素的分化程度會介於這兩者 之間,使得CAM 的 δ13C 值介於-27 – -12 ‰。大部分植物都屬於 C 3植物,包括各種溫 帶地區的草本植物、灌木、根莖類以及木本植物,重要的穀類作物如小麥、大麥和稻 米也屬於 C3植物。少部分熱帶地區的草本植物,包括小米、玉米、高粱和甘蔗則屬於 C4植物。CAM 植物多生長於沙漠地區,包括仙人掌,鳳梨和其它多肉植物(succulent plant)。 海洋環境中的碳源跟陸地不同,海洋裡的初級生產者以溶解於海水中的無機碳 (dissolved inorganic carbon,δ13C 為 0 ‰)做為碳源(Chisholm et al. 1982)。溶解的 無機碳比氣態二氧化碳更富集13C,因此生活於海洋環境中的生物 δ13C 值會比生活在陸 地環境中的C3植物δ13C 值高,而較為接近 C4植物的δ13C 值。以加拿大做為例子,其 陸地動物的平均δ13C 值為-25.7 ± 0.3 ‰,而海洋動物的平均 δ13C 值則為-17.8 ± 0.3 ‰ (ibid.)。綜合來說,骨膠原的碳同位素組成可用以辨別出該個體所攝取的食物屬於 C3 或C4植物來源,以及來自陸地或海洋(Schoeninger et al. 1983)。 有機氮的來源也可區分成陸地源或海洋源,使海洋植物的 δ15N 值通常比陸地植物 高。以大洋洲為例,其海洋藻類的δ15N 值約介於 4 – 8 ‰之間,而陸地植物 δ15N 值約為 2 – 7 ‰(Kinaston et al. 2013)。在陸地上不論是 C3還是C4植物,其δ15N 值與局部的 土壤環境較有關係,而與光合作用無關。除了環境中的氮源種類及其同位素組成外, 生物組織的δ15N 值與其所處的營養階層(trophic level)也有關。肉食者組織的 δ15N 值 一般比草食者高,草食者又比植物高(Minagawa and Wada 1984),這是因為生物偏好
排出較輕的氮(14N),造成體內15N 的富集所致。生物所處的食物階層不同而產生之同
位素組成的階層差異被稱為食階分化量(trophic level effect)。一個食階所造成的 δ15N 食階分化量介於3 – 5 ‰之間(Schoeninger and DeNiro 1984)。例如南非濱海地區的氮 同位素生態研究指出海洋貝類的δ15N 值約為 8 ‰,而以貝類為食的魚類或鳥類的 δ15N 值約為11 – 15 ‰,更高階的動物如海豹的 δ15N 值則高達 19 ‰(Sealy et al. 1987)。此
外海洋環境中的食物鏈階層數量比陸地多,因此海洋生物的 δ15N 值會高於陸地生物。 譬如Schoeninger and DeNiro(1984)曾收集 100 種海洋和陸地的現代魚類、鳥類和哺乳
動物的骨膠原碳、氮同位素數據,發現海洋動物的骨膠原δ15N 值平均比陸地動物高約 9 ‰。 除了指示有機碳、氮的供應源(食物)種類,骨膠原的碳、氮同位素組成還可用以 估算各類供應源的相對貢獻量(%)。理論上只需以消費者的骨膠原碳、氮同位素組成 扣除食階分化量後,即可獲得其整體飲食的同位素組成;再將之與各類食物的同位素 組成進行比較,就可求得各類食物在其整體飲食中的相對貢獻量(%)。簡單地說,n 種食物的相對貢獻量,可由n-1 種同位素解聯立方程式而求得,此被稱為線性混合模型
(Linear mixing model)。
以碳、氮兩種同位素為例,假定某人的整體飲食中包含三種食物(設為 x,y,z, 其各自的碳、氮同位素組成分別為δ13C x,δ15Nx;δ13Cy,δ15Ny;δ13Cz,δ15Nz)。又假定 其人的整體飲食碳、氮同位素組成分別為δ13C diet、δ15Ndiet。則這三種食物在其整體飲食 中的相對貢獻量(a,b,c)可由下列公式來計算: δ13C diet = a δ13Cx + b δ13Cy + c δ13Cz δ15N diet = a δ15Nx + b δ15Ny + c δ15Nz a + b + c =1 雖然骨膠原的同位素分析能協助估計各類食物在人類的整體飲食中的相對貢獻 量。但此類研究仍有一些限制和考量,包括:一、每個遺址或研究區域之各類食物的 同位素組成是否能被恰當地建立。這通常需仰賴動物考古學以及植物考古學的工作, 以協助瞭解人類的食譜組成;且需要各類生態遺留樣本以求得其同位素組成。由於不 同食物的同位素組成可能彼此重疊,單獨使用人骨膠原的同位素數據並無法直接獲得 其飲食中各類食物的相對貢獻量。換言之,人骨膠原同位素分析的結果,還必須與同 遺址或同地區的動物骨膠原和植物同位素數據加以比對、並與器物研究、植物考古 學、動物考古學、甚至是民族學的研究與證據加以檢視與辯證,方能正確地重建出一 個人群的攝食與生業體系。二、成岩同位素置換過程的考量。遺址中所出土之生態遺 留樣本的同位素組成可能會受到同位素置換而改變,要評估樣本的同位素組成是否受 到同位素置換過程的影響,通常可藉由光學技術如紅外線光譜和拉曼光譜(Beasley et al. 2014),或某些化學指標如碳含量(%C)、氮含量(%N)與碳氮元素莫耳比(molar C/N
ratio)協助判斷骨頭樣本的保存品質(DeNiro 1985)。三、是否能取得代表一遺址的食 階分化量。食階分化量為處於食物鏈某一位階的各類生物之同位素平均值,與其下一 階層(代表其整體飲食)的同位素平均值之間的差值。因此食階分化量可用以鑑別某生 物在其所處的食物鏈中的位階,並可據以推估該生物的整體飲食同位素組成。由於每 個遺址中所出土的生態遺留種類和其食物鏈基礎碳,氮源的同位素組成都不盡相同, 因此需要分析遺址中的肉食動物、草食動物和植物遺留同位素組成,進而推論出代表 該遺址的食階分化量。但若無法取得足夠的樣本,則通常會根據一經驗值作為食階分 化量的代表值。這類經驗值通常來自動物攝食實驗。實驗表示動物骨膠原的 δ13C 值通 常會比整體飲食的δ13C 值高約 3.4 ‰;而骨膠原 δ1%N 值則比整體飲食 δ1%N 值高約 3.0 ‰(Ambrose 1993)。因此假設某人類的骨膠原 δ13C 和 δ1%N 值分別為-20 ‰以及 9 ‰, 則可推估其整體飲食的δ13C 和 δ1%N 值應分別為-16.6 ‰以及 6 ‰。 雖然有這些限制存在,但仍有許多研究成功地應用骨膠原的碳,氮同位素組成獲 得人類飲食中各種食物資源的貢獻量,進而解析史前人群的攝食習性與生業策略,如 土地使用、畜牧和農業的經營方式、主要和次要的經濟行為等。而根據個體取食特徵 的差異也獲得如社會地位差異或性別差異等有具社會文化意涵的資訊。因此人骨膠原 碳、氮同位素分析在台灣考古學的研究中絕對是值得發展的一項重要技術。
研究對象
圓山遺址位於臺北盆地中基隆河河岸的一處小丘陵上,海拔高度約為 3 – 36 公尺 (圖1)。圓山遺址於 1897 年為伊能嘉矩與宮村榮一所發現,而於 1953 至 1954 年由國 立 臺 灣大 學和 臺 北文 獻委 員 會合 作進 行 第一 次正 式 發掘 (石 璋 如 1954;宋文薰 1954)。後續的發掘分別在 1986 到 1987、1991、1992、1997、和 1999 年。圓山遺址因 其重要性已於1988 年被指定為一級古蹟,並於 2006 年改定為國定遺址。 圓山遺址包括多層文化層,分別為先陶文化、大坌坑文化、訊塘埔文化、圓山文 化、植物園文化以及十三行文化。從這些文化層中曾出土相當豐富的考古遺物,包括 陶器、石器、玉器、大量獸骨、以及聞名的圓山貝塚(宋文薰 1954;連照美 1988;黃 士強 1989, 1991, 1992;黃士強、劉益昌 1999)。此外,從圓山貝塚中還曾出土五具人 骨(宋文薰 1954)。圓山文化至今已發現超過 50 個遺址,分布於臺北盆地以及臺灣北 部和西北濱海區(李娜莉 2003;郭素秋 2002;黃士強 1991)。過去關於圓山遺址之圖1 圓山遺址的位置
圓山文化的年代有許多討論(朱正宜 1990;宋文薰、張光直 1964;宋文薰等 1992; 黃士強、劉益昌 1999;劉益昌 2000)。本文作者亦嘗試由已發表的木頭和泥炭共十一 個碳十四定年結果以CALIB rev 5.0 軟體重新計算其校正年代(校正所使用的數據組來 自INTCAL04,Stuiver et al. 2005)。經過校正後的圓山文化年代介於 4200 – 2200 cal BP
(表1)。其中兩個泥炭的年代似乎較其餘九個偏早,可惜原報告中並無提及此二泥碳 定年的出土層位(黃士強 1992),因此無法確立其與文化層的關係。另劉益昌(2000) 曾提及圓山遺址所出土的貝殼碳十四定年皆有年代過早的問題,將貝殼年代經過減數 修正後並結合泥炭的碳十四年代,建議圓山文化的年代介於3200 – 2300 年前之間,後 期文獻皆多以此為圓山文化的年代依據。 根據圓山遺址圓山文化層所出土的器物和生態遺留,使我們得以初步了解圓山人 的生活方式(石璋如 1954;宋文薰 1954;李娜莉 2003;連照美 1988;黃士強 1989, 1991, 1992;黃士強、劉益昌 1999;Tan 1934)。首先,根據圓山貝塚中出土的貝殼種
類,指出貝塚形成當時的臺北盆地為一鹹淡水交會的溼地環境,而圓山人可能以貝類 為主要的食物來源之一。其中最常見的貝類為大蜆(Corbicula maxima Prime)。其它可 鑑定的貝類有田螺(Viviparus sp.)、網蜷(Melanoides sp.)、牡蠣(Ostrea sp.)、鐘螺 (Trochus sp.)和織紋螺(Nassarius sp.)(李娜莉 2003;連照美 1988)。其次,出土 器物中如帶倒刺的脫頭箭頭、投擲器(spears)、網墜,以及具有網紋和魚骨印紋的陶 片,都指出圓山人能從河流、湖泊或濱海濕地區取得各種水生資源(宋文薰 1954;黃 士強 1991, 1992)。部分出土的魚脊椎骨直徑可達 5 公分,推測圓山人對於大型魚類的 捕捉與食用。根據獸骨的鑑定,可知圓山人的陸地肉類來源可能包括梅花鹿(Cervus
nippon)、水鹿(Cervus unicolor)、山羌(Muntiacus reevesi)、豬(Sus sp.)、狗(Canis
sp.)、鼠、蛇與鳥等(宋文薰 1954;李娜莉 2003;黃士強 1991, 1992)。狗被認為是 人所飼養來作為嚮導或打獵幫手,並可在必要時提供肉類食物(宋文薰 1954)。此外, 鐮刀的出土顯示圓山人可能有耕種行為(宋文薰 1954),而碳化稻米粒的發現則暗示 稻米為可能的作物(黃士強、劉益昌 1999)。從這些器物和生態遺留,考古學者推論 圓山人的生業策略包括採貝、魚撈、狩獵以及耕作(宋文薰 1954)。基於以上學者的 研究成果,本研究試圖進一步對圓山人骨與獸骨進行碳、氮同位素分析,期冀能提供 新的資訊以對圓山人的食物資源與生業策略能有更細緻的討論與認識。
分析材料和實驗方法
樣本
本研究的分析樣本來自1953-54 年發掘出土的圓山人骨和獸骨樣本。初步先以肉眼 和放大鏡檢視並排除掉可能遭受囓齒動物或微生物破壞的骨頭樣本。由於本研究的目 標之一是要了解這批樣本是否具有保存良好的骨膠原,且是否適合用以進行同位素攝 食研究,因此初步採樣僅挑選出 20 個樣本。其中針對發掘報告中所記錄的五具人骨 (M1 – M5,宋文薰 1954)共採集 11 個樣本;而在樣本檢視過程中另外辨識出兩個個 體(M6、M7),因此從中再分別選取一個樣本。此外尚包括個體歸屬不明者(M?)共 計4 個樣本。總計,人骨樣本共 17 個,分別屬於七個或七個以上的個體(表 2)。其中 包含六個成年人(M1 – M5)和一個年輕個體(M7),但性別皆無法辨識。獸骨部分, 針對鹿、豬、魚三種動物各選取一樣本。骨膠原萃取
本研究所使用的骨膠原萃取流程係依循牛津大學考古暨美術史研究所 (RLAHA: Research Laboratory for Archaeology and the History of Art)的常規萃取法(參見 Pollard et al. 2011)。其大致步驟如下: 1. 選取約 0.5 – 1 公克的骨頭樣本,以碎鑽鑲嵌的鑽頭進行樣本表面的清理,其目 的為清除掉附著於樣本表面的可能汙染物質,包括土壤或黏膠。 2. 因海綿骨(cancellous bone)較易被填塞汙染物質,故移除海綿骨,僅留存皮質 骨(cortical bone)。 3. 將皮質骨以瑪瑙研缽仔細磨碎,秤重後移置於 15 毫升的玻璃試管中。 4. 加 入 10 毫 升 0.5 M 的 鹽 酸 , 置 放 於 4oC 的 冰 箱 中 , 使 樣 本 去 礦 化 (demineralization)。每隔兩天更換一次鹽酸,直至完成去礦化(亦即不再有氣泡的產 生)。 5. 移除鹽酸,並以 18mΩ-cm 純淨水淋洗樣本數次,直到水溶液呈現 pH 中性。 6. 加入 pH 3 的水(以濃鹽酸加入純淨水所調配),並以 75oC 持續加熱兩天,使骨 膠原溶解。 7. 經過濾後,把含骨膠原的水溶液置放於-80oC 的冰箱中結凍,再移置冷凍乾燥機 中使水分蒸發以獲取乾燥的骨膠原。秤重以計算骨膠原產量(%)。
骨膠原碳、氮同位素分析
在進行碳、氮同位素分析之前,本研究先以臺灣大學海洋研究所的元素分析儀(EA: Elemental Analyzer)分析每個骨膠原樣本的碳、氮含量,並計算其碳-氮元素莫耳比以判 斷骨膠原品質,並據以估計質譜儀分析所需的樣本量。 根據計算結果,將每個骨膠原樣本秤量約0.5 mg,包入錫箔小杯(tin capsule)內 後,置放於自動進樣器中。同位素分析工作分別在臺灣大學地質科學研究所(12 個樣 本)以及牛津大學(五個樣本)執行。兩處所使用的分析儀器皆為元素分析儀裝載連續 流質譜儀(EA-IRMS: EA continuous flow isotope ratio mass-spectrometer;臺灣大學: Thermo Finnigan MAT 253,牛津大學: Sercon Geo-20/20)。分析所得的碳、氮同位素組 成分別相對於各自的內部標準品進行校正(臺灣大學: 酪蛋白-casein,其 δ13C 和 δ15N 值 分別為-27.0 ‰和 5.9 ‰;牛津大學: 丙胺酸-alanine,其 δ13C 和 δ15N 值分別為-26.9 ‰和 -1.6 ‰)。此外牛津大學內部標準品也在臺灣大學的質譜儀中進行分析,以確保兩間實驗室的測量值可以進行比較。在樣本分析期間,標準品之δ13C 和 δ15N 的誤差值在臺灣 大學分別為0.18 ‰和 0.05 ‰(n = 34);而在牛津大學則皆為 0.02 ‰(n = 8)。
分析結果
圓山人、獸骨之骨膠原產量、碳含量、氮含量、碳氮元素莫耳比、以及碳、氮同 位素組成等分析結果詳列於表3 和表 4。而碳、氮同位素組成另繪於圖 2 和圖 3。骨膠原品質評估
圓山人、獸骨的骨膠原產量不高(1.68 – 11.74 %,表 3、表 4),平均值為 5.87 ± 3.13 %(1σ)。新鮮骨頭的骨膠原含量一般可達 20 – 25 %(Pollard et al. 2011)。此外, 這批樣本的骨膠原碳含量和氮含量也皆低於新鮮骨頭的預期值(C: 45 %;N: 16 %, Pollard et al. 2011)。這代表圓山骨頭樣本在其長期埋藏中,曾受到成岩同位素置換過程 影響,致使部分骨膠原流失。不過,對考古骨骸樣本而言,只要骨膠原產量> 1 %即可 被視為保存良好(DeNiro and Weiner 1988)。根據此原則,本研究所選取的 20 個人骨與獸骨樣本的骨膠原產量雖然不高,但都符合標準(表3、表 4)。 另當骨膠原的碳氮元素莫耳比介於2.9 至 3.6 之間,就可判定為保留著原始的同位 素訊號(DeNiro 1985)。根據此原則,有 14 個人骨樣本和 3 個獸骨樣本符合此標準。 因此這 17 個骨膠原樣本可被判定為保有原始碳、氮同位素組成,並適合用以討論圓山 人的攝食特徵。
人骨、獸骨樣本同位素組成
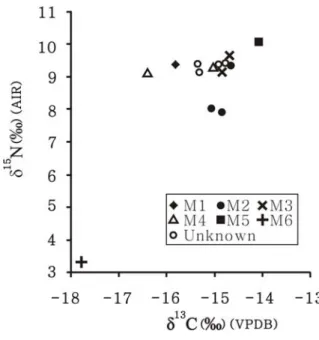
14 個通過骨膠原品質評估的人骨樣本,分屬六個或六個以上的個體(表 3)。其中 個體歸屬確立者有六個個體,共計10 個樣本。這六個個體的 δ13C 和 δ15N 值的範圍分別 為-14.1 – -17.8 ‰以及 3.3 – 10.1 ‰;而平均值分別為-15.5 ± 1.3 ‰(1σ)和 8.3 ± 2.5 ‰ (1σ)。此六個個體的同位素組成範圍和一個標準差都相當大,主要是因為其中有一個 個體(M6)的同位素組成偏離了其他個體。若把 M6 排除掉,則 M1 – M5 的 δ13C、δ15N 平均值分別為-15.0 ± 0.7 ‰和 9.3 ± 0.6 ‰,顯示出較集中的同位素組成(圖 2)。其中, M2 個體包括三個肋骨樣本,其 δ13C、δ15N 值的範圍分別為-15.1 – -14.6 ‰以及 7.9 – 9.4 ‰(δ13C 平均值為-14.9± 0.2 ‰;δ15N 平均值為 8.4 ± 0.8 ‰)。M5 相較於其他個體有較 高的δ13C、δ15N 值。圖2 圓山人骨膠原碳、氮同位素組成 三個獸骨樣本都通過骨膠原評估(表4)。其中魚的 δ13C、δ15N 值最高,分別為-12.3 ‰和 10.6 ‰。豬的 δ13C 值低於鹿(-17.1 ‰ vs -15.1 ‰);但 δ15N 值卻較高(7.2 ‰ vs 4.0 ‰)。
討論
台灣遺址的骨膠原品質評估
臺灣地區的骨骼遺留樣本是否適合進行骨膠原碳、氮同位素分析,以重建史前人 的攝食特徵? 除了圓山遺址外,本研究也曾針對烏山頭遺址、卑南遺址、鵝鑾鼻遺址 和番仔園遺址進行人骨膠原萃取的工作,根據這些目前能取得的骨骼樣本之初步分析 結果,臺灣各地的埋藏環境皆有其地域特徵,因此一個遺址的人骨和獸骨是否含有足 夠且品質佳的骨膠原以進行同位素攝食研究,須視其所在環境與形成過程而定。圓山 遺址的骨骼遺留雖然在骨膠原產量上並不高,大部分樣本的碳氮元素莫耳比也偏向可 接受範圍(2.9 – 3.6)的上限,但基本上仍符合標準,因此適合進行同位素古攝食研究。 但如烏山頭遺址和卑南遺址的人、獸骨樣本則完全萃取不出骨膠原,顯見其在埋藏環 境中已遭到地下水淋洗或其它分解作用致使骨膠原完全流失(李政益、林秀嫚 2015)。其他如鵝鑾鼻遺址人骨樣本的骨膠原產量偏低(平均僅約2.5 %,樣本數量= 9,李政益 等 未發表資料),但碳氮元素莫耳比仍落在可接受範圍;番仔園遺址 105 個人骨和獸 骨樣本的骨膠原產量平均為 7.4 %,碳氮元素莫耳比平均為 3.3(李政益等 未發表資 料),是骨膠原品質最好的一批樣本。綜合上述,可知臺灣各地的埋藏環境殊異,可能 導致不同遺址的考古骨骸骨膠原之保存狀況不一。一般而言,潮濕炎熱的環境較不利 於骨膠原的保存(Krigbaum 2003)。但若埋藏環境中有碳酸鹽類的存在,例如石灰岩層 或貝塚,則可讓骨骸遺留免於快速分解或溶解(Lee-Thorp and Sponheimer 2003)。綜觀 這幾個遺址的埋藏環境,圓山遺址、番仔園遺址皆伴隨貝塚的發現,而鵝鑾鼻遺址中 也存有石灰岩和珊瑚礁岩。因此很可能就是這些碳酸鹽類的存在,使其骨骸遺留保存 較其它遺址為佳。此外較年輕的骨骼樣本(如番仔園遺址),骨膠原的品質也相對較 好。這些結果建議在臺灣地區或者其它氣候狀況相似的區域進行人骨同位素分析研究 前,可先以遺址埋藏環境中是否伴隨碳酸鹽類以及遺址年代做為骨骸遺留保存狀況的 初步判斷依據,然後可採集小部分樣本進行骨膠原萃取和元素濃度分析以評估其骨膠 原品質,爾後才作進一步的同位素分析規劃。
圓山鹿、豬、魚的同位素組成與攝食特徵
根據圓山三個獸骨樣本的碳、氮同位素組成可評估這些動物的取食特徵。其中鹿 骨樣本(F01k-3)的 δ15N 值(4.0 ‰)符合草食動物的同位素特徵值。而根據其 δ13C 值 (-15.1 ‰),指出這隻鹿的食物為 C3和C4植物的混合,間接說明這兩類植物都存在於 其棲地的原生環境中。此推論與台北盆地孢粉分析所顯示的結果相一致(曾美惠、劉平 妹 1999)。 豬骨樣本(F01k-2)的 δ13C 值(-17.1 ‰)雖也指出其食物為 C 3類和C4類食物的混 合,δ15N 值(7.2 ‰)卻較鹿為高,因此顯示其食階位置較鹿高且呈現雜食特徵。此豬 骨樣本的骨膠原碳、氮同位素組成指出其食物選擇極為多樣,頗類似現代野豬的覓食 習性,亦即以環境中能取得的各種植物、昆蟲、小型爬蟲類或小型哺乳類為其食物來 源(趙榮台、方國運 1988)。李娜莉(2003)曾針對圓山遺址的豬骨形態進行比較分 析,也建議圓山的豬尚屬野豬一類。結合形態學與同位素分析結果,可推測這隻豬應 屬於野豬,曾在圓山遺址鄰近的溼地或濱海地帶四處覓食。 魚骨樣本(F02b-5)呈現較高的 δ13C 值(-12.3 ‰)和 δ15N 值(10.6 ‰),顯示出 海水魚類的同位素特徵值。圓山人的攝食特徵與生業體系
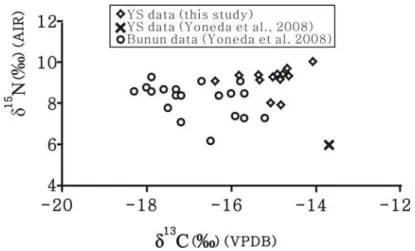
排除M6 後,M1 – M5 的骨膠原碳、氮同位素組成(δ13C = -15.0 ± 0.7 ‰、δ15N = 9.3 ± 0.6 ‰)指出其飲食是由 C3類食物和C4類/海洋類食物混合而成,且大致由肉類資源 作出貢獻。根據圓山遺址所出土的獸骨種類,指出圓山人的肉類資源可能包括陸地動 物如豬、鹿等,以及水生動物如魚、貝類(李娜莉 2003;宋文薰 1954;黃士強 1991, 1992)。是否這些動物都是圓山人的主要肉類來源?此議題可由人類和動物的骨膠原氮 同位素組成的比較來嘗試釐清。 動物攝食實驗的結果指出,若無其它食物階層的食物輸入,則單一食階所造成的 骨膠原δ15N 食階分化量會介於 1 – 6 ‰之間(Hedges and Reynard 2007;O’Connell et al. 2012)。比較圓山人骨膠原的 δ15N 平均值與圓山鹿和豬的骨膠原 δ15N 值,發現人相對 於豬和鹿的δ15N 差值分別為 2.2 ‰和 5.4 ‰,這說明豬和鹿的確屬於圓山人的肉類選擇 來源。但相比之下,海洋魚類的骨膠原δ15N 值卻高於圓山人的骨膠原 δ15N 平均值,指 出海水魚似乎並非圓山人的主要肉類資源,雖然其生活環境離海並不遠。 除了直接比較圓山人骨膠原和獸骨膠原的氮同位素食階分化量外,還可利用線性 混合模型以及圓山人和圓山豬、鹿、魚的骨膠原碳、氮同位素組成,進而評估這三種 動物類食物於圓山人飲食中的相對貢獻量。在進行評估之前,需先將人骨和獸骨的骨 膠原同位素組成進行必要的校正(圖3),包括:一、人類所吃的部位通常是動物的肌 肉,因此必須考量這三種動物的肌肉和骨膠原這兩種組織的碳、氮同位素組成差異。 實驗指出此差異量在碳同位素為2.4 ‰、在氮同位素為 0.8 ‰(Ambrose 2000; Tieszen and Fagre 1993)。換言之,將圓山豬、鹿、魚的骨膠原 δ13C、δ15N 值分別扣除 2.4 ‰和 0.8 ‰,即可獲得其肌肉的 δ13C、δ15N 值。二、由於人類飲食的同位素組成並不等同於 其骨膠原的同位素組成,因此需由人類骨膠原同位素組成扣除掉食階分化量後以求得 人類的飲食同位素組成。此食階分化量在碳同位素為3.4 ‰,在氮同位素為 3.0 ‰。 以經過上述校正所得之豬、鹿、魚之肌肉碳,氮同位素組成(豬δ13C = -19.5 ‰, δ15N = 6.4 ‰; 鹿 δ13C = -17.5 ‰,δ15N = 3.2 ‰; 魚 δ13C = -14.7 ‰,δ15N = 9.8 ‰),以 及圓山人飲食的碳、氮同位素組成(δ13C = -18.4 ‰,δ15N = 6.3 ‰),可計算出豬、鹿 和魚這三種動物在圓山人飲食中的相對貢獻量分別為66 %, 19 %和 15 %。此計算結果 表明若圓山人飲食中只包含這三種動物,則海水魚的重要性確實低於另外兩種陸地動 物,也建議圓山人的肉類來源可能主要是陸地動物。不過必須強調的是現階段所分析 的獸骨樣本數量過少,僅用單一樣本代表該種類食物的同位素組成當不具代表性;兼之圓山遺址出土動物遺留種類繁多,因此尚待日後取得更多種類及數量的獸骨樣本以 進行同位素分析,才能確實估算各種動物資源在圓山人飲食中的貢獻量。此外,若要 評估植物類食物的貢獻,也需要植物遺留作為分析材料。 圖 3 圓山人骨、獸骨之骨膠原碳、氮同位素組成。圓山人骨同位素組成(十 字)為平均值±一個標準差(M6 未被納入計算)。三種動物的肌肉碳、 氮同位素組成是由其各自的骨膠原碳、氮同位素組成(三角、星號、以 及方塊)分別扣除2.4 ‰和 0.8 ‰而獲得。人類飲食的同位素組成則以人 骨膠原同位素組成經食階分化量的校正而得(δ13C 值扣除 3.4 ‰; δ15N 值 扣除3 ‰)。 由於現階段仍缺乏足夠的動植物同位素數據以明確討論圓山人的攝食特性,因此 本研究暫以前人研究中發表的同位素數據做為比較基礎,進一步檢視現有分析結果的 資訊意涵。米田穣等(2008)分析 19 個布農族人骨膠原 δ13C、δ15N 值。其結果指出這 些個體的食物除了C3植物的貢獻外,還包括淡水魚類。與其分析結果相比較(圖4), 可發現圓山的人骨δ13C、δ15N 值皆較高(圓山人 δ13C 平均值為-15.0 ‰,δ15N 平均值為 9.3 ‰;布農族 δ13C 平均值為-16.8 ‰,δ15N 平均值為 8.2 ‰)。這代表圓山人的飲食中
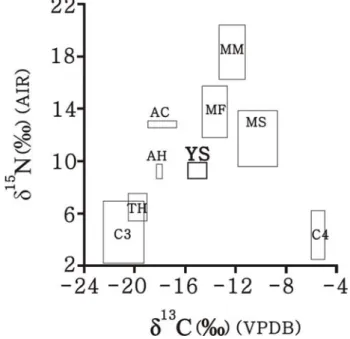
圖4 臺灣的人骨膠原碳、氮同位素組成。包括採自 Yoneda et al.(2008)的布 農族和圓山人骨膠原同位素數據,以及本研究的圓山人骨膠原同位素 值。 除了有較多肉類含量,也有較高的C4食物源或海洋食物源的輸入。 要釐清圓山人飲食中究竟是 C4食物源還是海洋食物源較為重要,可嘗試將圓山人 的骨膠原 δ13C、δ15N 值與東亞地區的各類動、植物之同位素基礎值做比較。由於此類 資料目前僅在日本被完整發表(Yoneda et al. 2004),因此本研究暫時僅以此批數據進 行比對(圖5)。圖 5 所示之各類動、植物的碳、氮同位素組成範圍,乃該研究收集前 人研究中所發表的同位素測量值進行適當的校正所得。其中 C3植物、C4植物、海洋貝 類和海洋魚類都是屬於現代樣本,因此其各自的δ13C 值需經過13C Suess 效應2的校正。 而陸地草食動物、淡水生物和海洋哺乳動物屬於古代樣本,因此不需要扣除 13C Suess 效應。不過這類樣本是以骨膠原做為同位素分析的材料,因此必須考量肌肉和骨膠原 兩組織間的碳、氮同位素組成差異。此差異量在碳同位素為 2.4 ‰、在氮同位素為 0.8
‰(Ambrose 2000, Tieszen and Fagre 1993)。最後還必須考量食階分化量,亦即消費者
的骨膠原與其飲食之間的碳、氮同位素組成差異。此食階分化量在碳同位素為 3.4‰,
在氮同位素為 3.0‰(Ambrose 1993)。將各類動、植物校正過的碳、氮同位素組成與
圓山人的骨膠原碳、氮同位素組成相比較 (圖 5),發現圓山人的食物可僅由陸地 C3
食物源 (包括 C3植物和吃C3類食物的草食或雜食動物)以及海洋食物源共同組成,而
圖5 圓山人骨膠原碳、氮同位素平均值±一個標準差(M6 未被納入計算), 以及日本的各類食物同位素組成(Yoneda et al. 2004)。圖上所示之各類 食物的碳、氮同位數值都已經過不同的校正考量。其中C3 植物(C3)、 C4 植物(C4)、海洋貝類(MS)和海洋魚類(MF)屬於現代樣本,因 此針對13C Suess 效應進行校正。陸地草食動物(TH)、淡水生物(包 括AH:水生草食動物,AC:水生肉食動物)和海洋哺乳動物(MM)屬 於古代樣本,雖不需要扣除 13C Suess 效應,但這類樣本是以骨膠原做 為同位素分析的材料,因此已經過組織(骨膠原 vs. 肌肉)同位素組成 差異量的校正。各類食物的同位素組成也都已經過食階分化量的校正 (詳細校正方式參見內文)。 上述推論亦可進一步由線性混合模型的計算來獲得證實。假設圓山人的飲食中僅 包含C3植物、C4植物和海洋資源這三類食物,並參考各類食物的碳、氮同位素組成(經 過13C Suess 效應校正後的日本現代 C3植物δ13C = -25.4 ‰,δ15N = 1.2 ‰;C4植物δ13C = -10.0 ‰,δ15N = 1.0 ‰;海洋魚貝類肌肉 δ13C = -16.3 ‰,δ15N = 9.4 ‰)以及經過食階 分化量校正的圓山人飲食同位素組成(δ13C = -18.4 ‰,δ15N = 6.3 ‰),則可推算出 C 4 植物在圓山人飲食中所佔的比例僅有8 %,而 C3植物和海洋類食物的貢獻量分別為29
%和 63 %。這代表若圓山人飲食中只包含這三類食物時,則海洋類食物的貢獻的確大 於C4植物。 結合現有的考古器物研究、生態遺留以及人骨同位素證據,可推測圓山人的飲食 主要包括C3食物源以及海洋食物源;至於C4食物源的貢獻則相對較小。其肉類來源則 以陸地動物為主。根據生態遺留的相對比例,圓山人飲食中的海洋類食物很可能主要 來自貝類,而海洋魚類的重要性則相對較低。雖然不同種類的生態遺留或許會因埋藏 作用而產生數量差異,因此依據生態遺留相對比例推測食物的重要程度有其可能的偏 差。不過在黃士強先生的多次發掘中,發現魚類數量雖不及貝類,仍出土許多細小魚 骨(李娜莉2003;黃士強 1989, 1991, 1992),顯示此地的埋藏環境對魚骨與貝殼遺留 所造成的保存差異應該不大。若此推論無誤,則圓山遺址的生態遺留佔比應仍可做為 這兩類資源的相對重要性之判斷依據。
個體攝食差異
米田穣等(2008)所分析的史前人骨樣本中,有一個取自圓山遺址(編號 B)。不 過,此一圓山人骨樣本的同位素組成落在本研究的五個圓山人骨同位素範圍之外(圖 4),呈現出偏低的 δ15N 值(6.0 ‰)和偏高的 δ13C 值(-13.7 ‰),代表其食物中有較多 C4植物類食物。而本研究中樣本編號M6 的個體 δ15N 值更低,δ13C 值也較小(表 3), 代表M6 的食物幾乎僅由 δ15N 值極低的 C 3類植物所組成,可能的候選者為豆類植物。 豆類植物屬於 C3類植物,但由於豆類植物與根瘤菌共生,因此能直接利用大氣的氮氣 作為其氮源,使豆類植物的δ15N 值通常接近 0 ‰。雖然同位素組成支持 M6 以豆類為主 食的可能性,不過圓山遺址迄今尚未有豆類植物遺留的出土,因此暫不以此做為定 論。 雖然我們能取得的樣本數量不足以進行統計運算以評估這兩個個體(本文中的 M6 以及米田穣等文中的 B)是否屬於異常值(outlier),但其攝食特徵與其他圓山人仍有 相當差異,推論造成此現象的幾個可能因素為: 一、此二個體屬於外來者。 新石器時代中期,臺灣北部地區除了圓山文化外,尚有芝山岩文化與晚期可能有 部分重疊的植物園文化(郭素秋 2002)。即便是圓山文化本身,其遺址就分布在不同 的地理位置,包括臺北盆地內部以及北部或西北部的濱海地區(李娜莉 2003;黃士強 1991)。不同的遺址之間可能因婚嫁或貿易而有人群的移動或遷移。張光直(1957)亦曾從圓山遺址出土的人骨與穿孔門齒推論圓山人有獵頭習俗,若此推論可能,則代表 人群間的衝突也可能迫使俘虜被埋葬於異地中。由這些考古文獻來看,這兩個取食特 徵較特殊的個體可能來自同時代的其它文化人群或者同文化的其它人群。不過鄰近遺 址目前尚無同時代的人骨碳、氮同位素組成可供比較,故無法確定他們是否真為外來 者。建議能針對其它相鄰遺址的人骨進行碳、氮同位素分析,提供人群之間的攝食體 系比較基礎,從而釐清這些取食特徵異常的個體的可能來源區。或者也可進一步針對 這些個體進行骨頭礦物的鍶(Sr)和氧(O)同位素分析,並與鄰近區域的其它環境或 其它遺址之鍶、氧同位素代表值做比較,或許有助於明確推論這些個體的來源區。 二、文化或社會結構帶來的個體攝食差異。 此二個體的 δ15N 值低於其他個體,代表其飲食中有較多植物類食物的貢獻。此一 現象或許暗示圓山人群有社會分化或性別差異的特徵,導致不同地位或不同性別的個 體具有特定的取食行為。社會分層或性別差異造成的食物取得差異現象也曾出現在其 它史前社會中,經常表現為地位較高者或男性有較多的機會獲得動物性蛋白質;而地 位較低者或女性則較常以植物做為食物(例如Ambrose et al. 2003)。可惜的是,現有 的考古材料無法用以判定圓山文化的社會中是否有地位高低或性別的差異。除了M2 有 疑似陪葬品的一個石斧外,圓山遺址的其它個體均無陪葬品出土(石璋如 1954);而 墓葬的體質保存狀況也不足提供性別的判定。無法排除這兩個個體也可能有著與他人 不同的信念或個人攝食偏好,因此特地選擇植物類食物為主要取食對象,如同現代的 素食主義者。但這類推論已超出本研究所能討論的範圍。 三、這些個體在年代上與其他個體有所不同。 M1 – M5 為圓山遺址在 1953、54 的發掘工作中所判斷出來的五個個體(石璋如 1954),而 M6 是本研究在人骨整理與取樣過程中另外辨識出來的個體。米田穣等 (2008)所分析個體並無考古出土脈絡資訊。可以確定的是 M1 – M6 皆出土於貝塚內, 並無以棺槨埋葬。這些骨骼無法完全排除可能受到外力擾動而偏離其原始埋葬層位, 例如M4 和 M5 的骨骼在出土時皆呈頭下腳上的姿態,且與山坡斜度吻合;代表有被移 動過的可能性。而M5 頭骨被壓扁,軀幹因關節鬆弛變長,更被判定可能因地震而移動 (石璋如 1954)。另外,貝塚分上層灰土和下層黃土的堆積差異,墓葬出土的堆積深 度與層位略有差異(石璋如 1954)。從這些發掘記錄來看,無法肯定這些個體是否都 屬於同一個層位或同一個時期。換言之,雖然這些個體仍明確屬於圓山文化,但因為 圓山文化的年代範圍很寬,這些個體可能有生存時間早晚的區別,而分別代表圓山文
化中不同時期。因此他們雖皆屬於同一文化的人群,但一個群體中的攝食體系卻可能 隨著時間發生變化。未來可進一步針對圓山人骨進行放射碳定年分析,或可釐清這些 人骨的生存年代,進一步檢視圓山文化中不同時期的取食特性。
結論與展望
本研究分析圓山遺址的20 個人骨和獸骨的骨膠原碳、氮同位素組成。其中 14 個人 骨樣本和3 個獸骨樣本通過骨膠原品質評估。14 個人骨分屬於六個(M1 – M6)或六個 以上的個體。除了M6 外,其他五個個體的骨膠原同位素組成指出圓山人的飲食主要包 括 C3食物源(C3植物和以 C3類食物為食的動物)以及海洋資源(以貝類為主);C4 源食物的貢獻相對較小。此一結果與考古器物研究和生態遺留分析所獲得的結果相吻 合。人骨同位素和考古學證據共同說明圓山人的生業方式為多樣性策略: 獵捕魚、豬、 鹿和其它動物、採集貝類、並耕作稻米等 C3類植物。人骨同位素的分析成果不僅能協 助檢視並支持早期考古學研究對圓山人的攝食與生業策略的復原結果,更將理解細緻 化;甚至提供了個體差異資訊,以供其他文化社會面向的進一步探討。 雖然本研究對圓山人的同位素攝食研究有了初步的成果,但仍有許多疑問待釐 清,這將需進一步的分析工作,包括: 一、現有獸骨樣本數量仍太少,無法建立起明確的圓山遺址同位素基礎值以供人 骨膠原同位素組成做詳細比較。未來需取樣更多的豬、鹿和魚骨作進一步的分析,同 時也當包含不同的動物種類。這些分析結果將能提供關於圓山人攝食特徵的詳細資 訊。 二、欲評估植物類食物(如稻米和小米)在圓山人飲食中的重要性,需要針對遺址 出土的植物遺留進行碳、氮同位素分析。但圓山遺址現有的考古發掘僅出土少量碳化 稻米粒遺留,而並未發現小米或其他植物遺留。有可能是因小米的確未被圓山人所種 植使用,或者未能被留存下來,也或許在發掘當時未能掏洗出。無論原因為何,現有 的考古學證據指出除稻米外並無法確定到底圓山人曾食用哪些植物,以及各種植物在 圓山人飲食中的重要性又是如何。可能的解決方法是嘗試從臺灣其它曾出土稻米、小 米和其它植物遺留的遺址中採集樣本以進行植物同位素組成的分析,並配合圓山人骨 膠原同位素數據,或許能幫助釐清小米、稻米或其它植物在圓山人飲食中的重要程 度。三、針對圓山遺址所出土的人骨進行進一步的分析工作,例如人骨碳定年分析將 有助於比較每個個體之間的年代關係。此外如骨頭礦物中的鍶和氧同位素分析也能幫 助判斷個體是否為外來者,並推測可能的來源地區。 四、針對圓山文化其它遺址的人骨,或同時代不同文化的人骨進行同位素研究。 這將有利於比較新石器中期圓山文化居住於不同地理位置人群的生業策略差異、或不 同文化人群的攝食偏好,進而探討人類與環境的互動關係以及人類對環境的利用和適 應形式。
致 謝
本研究感謝臺灣大學海洋研究所的曾靜宜小姐、林其郁小姐和劉玲雯小姐協助樣 本的元素和同位素分析工作;海研所蘇志杰教授提供冷凍乾燥機;計畫相關工作人員 在人骨與獸骨的清理與紀錄工作上的協助。另感謝三位審查者以及編輯的寶貴建議和 提問。本研究特別感謝蔣經國基金會對本研究的經費支持。附 註
1. 成岩同位素置換作用係指骨頭樣本經埋藏後,其原有的生物性同位素訊號因骨頭分 解而損失,或遭環境同位素置換而改變。 2. 由於 C3化石燃料(貧13C)的燃燒,使得現代大氣中的二氧化碳 δ13C 值會比工業革 命前低,此效應在現代海洋樣本中為0.5‰,在現代陸地樣本中為 1.5 ‰(Marino and McElroy 1991; Guilderson et al. 1998)。換言之,所有現代樣本的 δ13C 量測值必須加 上此效應值,才等同於未受到此效應影響的古代樣本δ13C 值。引用書目
石璋如
朱正宜 1990 〈臺灣地區碳十四年代數據輯〉。《田野考古》1(1):95-122。 米田穣、向井人史、蔡錫圭 2008 〈台湾先史時代遺跡か か出土し し古人骨 ,近代と ブ ブ ブ人骨に に に に炭素・ 窒素同位体分析〉。Anthropological Science 116(2): 161-170。 宋文薰 1954 〈圓山貝塚民族的生產方式〉。《臺北文物》3(1):2-7。 宋文薰、張光直 1964 〈圓山文化的年代〉。《國立臺灣大學考古人類學刊》23/24:1-11。 宋文薰、尹建中、黃士強、連照美、臧振華、陳仲玉、劉益昌 1992 《臺灣地區重要考古遺址初步評估–第一階段研究報告》。中國民族學會專案研 究叢刊(一)。內政部委託中國民族學會。 李匡悌 1995 《恆春半島史前海岸聚落的比較研究:以龜山史前遺址和鵝鑾鼻第二史前遺址 為例》。國立海洋生物博物館籌備處委託中央研究院歷史語言研究所。 李作婷、鄭建文 2008 〈鵝鑾鼻第二史前遺址的植矽石分析 史前稻作農業的相關考察〉。「2008 年度 台灣考古工作會報摘要集」宣讀論文,中央研究院歷史語言研究所,3 月 28-29 日。 李政益、林秀嫚 2015 〈烏山頭遺址人骨骨膠穩定碳氮同位素之解析〉。刊於《骨說史前人》。林秀嫚 編,頁65-72。臺東:國立臺灣史前文化博物館。 李娜莉 2003 《論圓山文化的生產模式與經濟策略:以圓山遺址的動物遺留為例》。國立臺 灣大學人類學系碩士論文。 連照美 1988 〈臺北圓山遺址現況調查研究報告〉。《臺北文獻》83:1-47。
陳光祖 2004 〈台灣科技考古研究工作的回顧〉。《田野考古》8:15-44。 郭素秋 2002 〈「植物園文化」探析〉。《文與哲》1:273-332。 張光直 1957 〈圓山出土的一顆人齒〉。《考古人類學刊》9/10:146-148。 黃士強 1989 〈臺北市圓山遺址第二地點試掘報告〉。《國立臺灣大學考古人類學刊》45: 20-65。 1991 《圓山遺址中山三十三號道路試掘與評估》。臺北市民政局委託國立臺灣大學 人類學系。 1992 《第一級古蹟圓山遺址範圍試掘評估報告》。內政部委託國立臺灣大學人類學 系。 黃士強、劉益昌 1999 《圓山遺址史蹟公園範圍區考古發掘研究計畫》。臺北市立兒童育樂中心委託 國立臺灣大學人類學系。 曾美惠、劉平妹 1999 〈臺北盆地兩萬年來孢粉組合與古環境初探〉。《中央地質調查所特刊》11: 159-179。 趙榮台、方國運 1988 《臺灣野豬之生態與行為研究》。臺北:行政院農業委員會。 劉益昌 2000 〈圓山文化年代檢討—兼論臺北盆地史前文化變遷〉。刊於《北臺灣鄉土文化 學術研討會論文集》。周惠民編,頁71-112。臺北:國立政治大學歷史學系。 Ambrose, Stanley H.
1993 Isotopic Analysis of Paleodiet: Methodological and Interpretive Considerations. In Investigations of Ancient Human Tissue: Chemical Analyses in Anthropology. Mary
K. Sandford, ed. Pp. 59-130. Langhorne: Gordon and Breach.
2000 Controlled Diet and Climate Experiments on Nitrogen Isotope Ratios of Rats. In Biogeochemical Approaches to Paleodietary Analysis. Stanley H. Ambrose and M. A. Katzenberg, eds. Pp. 243-259. New York: Kluwer Academic/Plenum Publishers. DOI: 10.1007/0-306-47194-9_12
Ambrose, Stanley H., Jane Buikstra, and Harold W. Krueger
2003 Status and Gender Differences in Diet at Mound 72, Cahokia, Revealed by Isotopic Analysis of Bone. Journal of Anthropological Archaeology 22: 217-226. DOI: 10.1016/S0278-4165(03)00036-9
Beasley, Melanie M., Eric J. Bartelink, Lacy Taylor, and Randy M. Miller
2014 Comparison of Transmission FTIR, ATR, and DRIFT Spectra: Implications for Assessment of Bone Bioapatite Diagenesis. Journal of Anthropological Archaeology 46: 16-22. DOI: 10.1016/j.jas.2014.03.008
Chisholm, Brian S., D. Erle Nelson, and Henry Schwarcz
1982 Stable Carbon Isotope Ratios as a Measure of Marine Versus Terrestrial Protein in Ancient Diets. Science 216: 1131-1132. DOI: 10.1126/science.216.4550.1131 DeNiro, Michael J.
1985 Postmortem Preservation and Alteration of in Vivo Bone Collagen Isotope Ratios in Relation to Palaeodietary Reconstruction. Nature 317: 806-809. DOI: 10.1038/317806a0
DeNiro, Michael J., and Samuel Epstein
1978 Influence of Diet on the Distribution of Carbon Isotopes in Animals. Geochimica et Cosmochimica Acta 42: 495-506. DOI: 10.1016/0016-7037(78)90199-0
1981 Influence of Diet on the Distribution of Nitrogen Isotopes in Animals. Geochimica et Cosmochimica Acta 45: 341-351. DOI: 10.1016/0016-7037(81)90244-1
DeNiro, Michael J., and Stephen Weiner
1988 Chemical, Enzymatic and Spectroscopic Characterization of “Collagen” and other Organic Fractions from Prehistoric Bones. Geochimica et Cosmochimica Acta 52:
2197-2206. DOI: 10.1016/0016-7037(88)90122-6 Farquhar, Graham D., Jim R. Ehleringer, and K. T. Hubick
1989 Carbon Isotope Discrimination and Photosynthesis. Annual Review in Plant Physiology and Plant Molecular Biology 40: 503-537. DOI: 10.1146/annurev.pp. 40.060189.002443
Guilderson, Thomas P., Danie P. Schrag, Michaele Kashgarian, and John Southon
1998 Radiocarbon Variability in the Western Equatorial Pacific Interred from a High-Resolution Coral Record from Nauru Island. Journal of Geophysical Research 103: 24641-50. doi: 10.1029/98JC02271
Hedges, Robert E. M., John G. Clement, C. David, L. Thomas, and Tamsin C. O’connell 2007 Collagen Turnover in Adult Femoral Mid-Shaft: Modeled from Anthropogenic
Radiocarbon Tracer Measurements. American Journal of Physical Anthropology 133: 808-816. doi: 10.1002/ajpa.20598
Hedges, Robert E. M., and Linda M. Reynard
2007 Nitrogen Isotopes and the Trophic Level of Humans in Archaeology. Journal of Archaeological Science 34: 1240-1251. doi: 10.1016/j.jas.2006.10.015
Kinoston, Rebecca, Hallie Buckley, Andrew Gray, Ben Shaw, and Herman Mandui
2013 Exploring Subsistence and Cultural Complexes on the South Coast of Papua New Guinea Using Palaeodietary Analyses. Journal of Archaeological Science 40: 904-913. doi: 10.1016/j.jas.2012.09.002
Krigbaum, John
2003 Neolithic Subsistence Patterns in Northern Borneo Reconstructed with Stable Carbon Isotopes of Enamel. Journal of Anthropological Archaeology 22: 292-304. doi: 10.1016/S0278-4165(03)00041-2
Lee-Thorp, Julia A.
2008 On Isotopes and Old Bones. Archaeometry 50: 925-950. doi: 10.1111/j.1475-4754. 2008.00441.x
Lee-Thorp, Julia A., Matt Sponheimer
2003 Three Case Studies Used to Reassess the Reliability of Fossil Bone and Enamel Isotope Signals for Paleodietary Studies. Journal of Archaeological Science 22: 208-216. doi: 10.1016/S0278-4165(03)00035-7
Lubell, David, Mary Jackes, Henry Schwarcz, Martin Knyf, and Christopher Meiklejohn 1994 The Mesolithic-Neolithic Transition in Portugal: Isotopic and Dental Evidence of
Diet. Journal of Archaeological Science 21: 201-216. doi: 10.1006/jasc.1994.1022 Marino, Bruno D., and Michael B. McElroy
1991 Isotopic Composition of Atmospheric CO2 Inferred from Carbon in C4 Plant Cellulose. Nature 349: 127-131. doi: 10.1038/349127a0
Minagawa, Masao, and Eitaro Wada
1984 Stepwise Enrichment of 15N Along Food Chains: Further Evidence and the Relation between Δ15N and Animal Age. Geochimica et Cosmochimica Acta 48: 1135-1140. doi: 10.1016/0016-7037(84)90204-7
O'Connell, Tamsin C., C. J. Kneale, N. Tasevska, and G. G. C. Kuhnle
2012 The Diet-Body Offset in Human Nitrogen Isotopic Values: A Controlled Dietary Study. American Journal of Physical Anthropology 149: 426-434. doi: 10.1002/ajpa.22140
Polet, Caroline, and M. Anne Katzenberg
2003 Reconstruction of the Diet in a Mediaeval Monastic Community from the Coast of Belgium. Journal of Archaeological Science 30: 525-533. doi: 10.1016/S0305-4403 (02)00183-8
Pollard, A. Mark, and Carl Heron, eds.
2008 Archaeological Chemistry. 2nd edition. Cambridge: The Royal Society of Chemistry.
Pollard, A. Mark, Peter Ditchfield, James S. O. McCullagh, T. G. Allen, M. Gibson, C. Boston, Sharon Clough, Nicolas Marquez-Grant, and R. A. Nicholson
Inhumation from Gravesend, Kent, UK. American Journal of Physical Anthropology 146: 446-456. doi: 10.1002/ajpa.21602
Richards, Mike P., B. Fuller, and T. Molleson
2006 Stable Isotope Palaeodietary Study of Humans and Fauna from the Multi-Period (Iron Age, Viking and Late Medieval) Site of Newark Bay, Orkney. Journal of Archaeological Science 33: 122-131.
Schoeninger, Margaret J., Michael J. DeNiro, and H. Tauber
1983 Stable Nitrogen Isotope Ratios of Bone Collagen Reflect Marine and Terrestrial Components of Prehistoric Human Diet. Science 220: 1381-1383. doi: 10.1126/science.6344217
Schoeninger, Margaret J., and Michael J. DeNiro
1984 Nitrogen and Carbon Isotopic Composition of Bone Collagen from Marine and Terrestrial Animals. Geochimica et Cosmochimica Acta 48: 625-639. doi: 10.1016/0016-7037(84)90091-7
Schoeninger, Margaret J.
2011 Diet Reconstruction and Ecology Using Stable Isotope Ratios. In A Companion to Biological Anthropology. Clark Spencer Larsen, ed. Pp. 445-464. Chichester: Wiley-Blackwell. doi: 10.1002/9781444320039.ch25
Schwarcz, Henry P., and Margaret J. Schoeninger
2012 Stable Isotopes of Carbon and Nitrogen as Tracers for Paleo-Diet Reconstruction. In Handbook of Environmental Isotope Geochemistry, Advances in Isotope Geochemistry. Mark Baskaran, ed. Pp. 725-742. Berlin: Springer-Verlag. doi: 10.1007/978-3-642-10637-8_34
Sealy, Judith C., Nikolaas J van der Merwe, Julia A Lee-Thorp, and John L. Lanham
1987 Nitrogen Isotopic Ecology in Southern Africa: Implications for Environmental and Dietary Tracing. Geochimica et Cosmochimica Acta 51: 2707-2717. doi: 10.1016/0016-7037(87)90151-7
Stuiver, Minze, Paula Reimer, and R. Reimer
2005 CALIB Radiocarbon Calibration, version 5.0. University of Washington Quaternary Isotope Lab, http://radiocarbon.pa.qub.ac.uk/calib/, accessed 25 September 2014. Tan, K.
1934 The Variation of Corbicula Maxima Prime between Contemporary Specimen and Specimen from Yuan-Shan Midden. The Venus 4(5): 289-302.
Tauber, Henrik
1981 13C Evidence for Dietary Habits of Prehistoric Man in Denmark. Nature 292: 332-333.
Tieszen, Larry L., and Tim Fagre
1993 Effect of Diet Quality and Composition on the Isotopic Composition of Respiratory CO2, Bone Collagen, Bioapatite, and Soft Tissues. In Prehistoric Human Bone: Archaeology at the Molecular Level. J. B. Lambert and G. Grupe, eds. Pp. 121-155. Berlin: Springer-Verlag.
Tsang, Cheng-hwa
1992 Archaeology of Peng-hu Islands. Special Publications no. 95, Pp. 492. Taipei: Institute of History and Philology, Academia Sinica.
van der Merwe, Nikolaas J., J. C. Vogel
1978 13C Content of Human Collagen as a Measure of Prehistoric Diet in Woodland North America. Nature 276: 815-816. doi: 10.1038/276815a0
Yoneda, Minoru, Ryo Suzuki, Yasuyuki Shibata, Masatoshi Morita, Tomohiro Sukegawa, Nobuo Shigehara, and TakeruAkazawa
2004 Isotopic Evidence of Inland-Water Fishing by a Jomon Population Excavated from the Boji Site, Nagano, Japan. Journal of Archaeological Science 31: 97-107. doi: 10.1016/S0305-4403(03)00103-1
Zhao, Zhijun
2011 New Archaeobotanic Data for the Study of the Origins of Agriculture in China. Current Anthropology 52: S295–S306. doi: 10.1086/659308