1. 以偶合COMT螢光檢測甲硫氨酸腺苷基轉移酵素活性之高效液相層析法的開發 2. HotLig: 以分子表面導向之方法評估蛋白質-配體之交互作用; 1. HPLC method for the determination of methionine adenosyltransferase activity using catechol-O-methyltransferase-coupled fluorometric detection 2. HotLig: A molecular surface-dir
168
0
0
全文
(2) 謝 辭 首先由衷感謝口試委員台北醫學大學陳繼明老師和靜宜大學陳香惠老 師,以及本校黃麗嬌所長、林宗平老師與連金城老師,細心審查本論文並且 給予珍貴的意見與教導。我的指導教授郭盛助教授以及陳勝智教授在我研究 求學階段,相當用心、耐心與包容地指導我,不僅僅是在學術上的啟發,他 們的人格修養更是我人生的導師。 還有研究所內許許多多的好友們,要感謝的人太多了,感謝你們的陪伴 與鼓勵,這裡實在有太多美好的回憶!希望你們都能有更卓越的研究成果以 及璀璨的人生!感謝龔姊多年來的照顧,讓我們許許多多的事務都能順利進 行,希望您青春永駐。還有更要感謝賴建銘老師,我們都會想念您的。 這裡也要感謝泰霙一直以來的支持鼓勵與陪伴,豐富了我人生的色彩。 最後要感恩我的父母親,當我完成學位時,他們都已年近七旬,心中除了不 捨,更有無盡的感恩!.
(3) 中國醫藥大學 藥物化學研究所 博士論文 (一) 指 導 教 授 : 郭盛助 指 導 教 授 : 陳勝智. 教 授 教 授. 以偶合 COMT 螢光檢測甲硫氨酸腺苷基轉移酵素活性之高 效液相層析法的開發. HPLC method for the determination of methionine adenosyltransferase activity using catechol-O-methyltransferase-coupled fluorometric detection. 研究生: 王. 勝. 弘. Wang, Sheng-Hung. 中. 華. 民. 國. 95 年. 6 月.
(4) 目錄 圖目錄 ........................................................................................................... ii 表目錄 .......................................................................................................... iii 中文摘要 ...................................................................................................... iv 英文摘要 ....................................................................................................... v 縮寫 .............................................................................................................. vi 第一章 緒論 ................................................................................................. 1 第一節 MAT 的分子生物學 .................................................................. 1 第二節 Methionine 的代謝路徑及其在醫療上的意義 ......................... 7 第三節 過去 AdoMet 與 MAT 活性之檢測方法................................. 15 第二章 研究動機與目的............................................................................ 22 第三章 實驗器材與方法............................................................................ 23 第一節 實驗材料與儀器..................................................................... 23 第二節 細胞培養與 MAT 酵素製備................................................... 26 第三節 MAT 活性分析方法 ............................................................... 27 第四章 結果與討論.................................................................................... 31 第五章 結論 ............................................................................................... 46 參考文獻 ..................................................................................................... 47 i.
(5) 圖目錄 Scheme 1.. COMT 偶合酵素螢光檢測法之示意圖 ................................ 22. 圖 1.. MAT 所催化的反應 ................................................................. 1. 圖 2.. 老鼠肝細胞 α subunit 模型 (PDB code: 1QM4)..................... 4. 圖 3.. NO 與 GSH 對 MAT 的調節作用 ........................................... 5. 圖 4.. Methionine 的代謝路徑 .......................................................... 8. 圖 5.. 異常甲基轉移酵素調節細胞分裂和細胞分化的角色 ......... 11. 圖 6.. AdoMet 之 isoindole 類螢光化合物形成原理 ..................... 18. 圖 7.. COMT 偶合酵素檢測 MAT 活性之機轉.............................. 34. 圖 8.. MGBG 在 COMT 偶合酵素法中對測定結果之影響........... 36. 圖 9.. COMT 使用量對 scopoletin 生成之影響.............................. 38. 圖 10.. Scopoletin 生成與時間的關係............................................... 39. 圖 11.. COMT 偶合酵素轉換 AdoMet 之效能 ................................. 41. 圖 12.. Scopoletin 檢量線 ................................................................. 42. 圖 13.. MAT 酵素抽取液用量與檢測值之線性關係 ....................... 43. 圖 14.. HL-60 MAT 酵素動力學之 Eadie-Hofstee plots ................... 45. ii.
(6) 表目錄 Table 1.. MAT isozymes 之間的比較 .................................................... 2. iii.
(7) 中文摘要 甲硫氨酸腺苷基轉移酵素 Methionine adenosyltransferase (MAT; EC 2.5.1.6) 是體內代謝中甲基的主要來源,影響範圍包括 DNA、RNA 以及蛋白質與其他 小分子的代謝。然而至今 MAT 酵素的活性依舊只能以放射性同位素法進行測 定。這個研究當中,我們建立了一個高感度,有專一性,又快速簡便的酵素 偶合之螢光液相層析分析法來分析 methionine adenosyltransferase (MAT) 酵素 活性及酵素動力學常數。這個方法利用 catechol-O-methyltransferase (COMT) 將 S-adenosyl-L-methionine (AdoMet) 上的甲基轉移到 esculetin (ECL) 上形成 甲基化的 scopoletin (SPL),經由 n-hexane:ethyl acetate (7:3, v/v) 萃取後,利用 高效液相層析 (HPLC) 配接 Si 60 管柱以及螢光檢測器分析 SPL (ex: 347 nm; em:415 nm),測定 MAT 的活性及酵素動力學常數。SPL 之最低檢測量約為 100 fmol;這個方法僅需要酵素約相當於 2.5 µg 的蛋白質,酵素反應時間可少於 30 分鐘,每個檢品之 HPLC 分析時間約只需 5 分鐘。酵素動力學的研究顯示 HL-60 MAT 呈現了 Hill coefficient 為 0.5 之 negative cooperativity,Km = 6.1 ± 0.3 µM (for methionine),Vmax = 135.4 ± 1.5 nmole AdoMet/mg protein/hr。故此新穎 的 COMT 偶合 HPLC 螢光分析法能成功的應用於測定微量 AdoMet 以及 MAT 酵素活性,並成為放射性同位素法之替代方法。. iv.
(8) 英文摘要 A non-radioactive, sensitive, rapid and specific method for the determination of methionine adenosyltransferase activity has been established. In this method, the methyl group of S-adenosyl-L-methionine was enzymatically transferred to esculetin with the aid of catechol-O-methyltransferase and then the resulting scopoletin was extracted with n-hexane:ethyl acetate (7:3, v/v) and measured by high-performance liquid chromatography with Si 60 column and fluorometric detection with excitation and emission wavelengths at 347 and 415 nm, respectively. The detection limit for scopoletin was about 100 fmol. Using this method to determine MAT activity in HL-60 cells only required about 2.5 µg of protein and the incubation time needed for enzymatic reaction is less than 30 min. The HPLC analysis procedure took only 5 min per sample. The kinetic study showed that MAT in HL-60 cells exhibited negative cooperativity with a Hill coefficient of 0.5. The values of Km and Vmax were 6.1 ± 0.3 µM and 135.4 ± 1.5 nmol AdoMet formed/mg protein/h, respectively.. Key words: methionine adenosyltransferase (MAT); catechol-O-methyltransferase (COMT); S-adenosyl-L-methionine (AdoMet); methylglyoxal bis(guanylhydrazone) (MGBG); esculetin; scopoletin; enzyme-coupled; fluorescence; HPLC; HL-60.. v.
(9) 縮寫 AdoHcy:S-adenosyl-L-homocysteine AdoMet:S-adenosyl-L-methionine ATP:adenosine 5′-triphosphate COMT:catechol-O-methyltransferase DMSO:dimethyl sulfoxide DTT:1,4-dithiothreitol ECL:esculetin GSH:glutathione Hcy:homocysteine L-Met:L-methionine. MAT:methionine adenosyltransferase MATH:MAT isozyme with high Km for methione MATI:MAT isozyme with intermediate Km for methione MATL:MAT isozyme with low Km for methione MATLT:MATL with tumor factor MGBG:methylglyoxal bis-(guanylhydrazone). vi.
(10) MT:Methyltransferase PIPES:piperazine-N-N′-bis(2-ethanesulfonic acid) SAH:S-adenosyl-L-homocysteine SAHH:S-adenoxylhomocysteine hydrolase SAM:S-adenosyl-L-methionine SPL:scopoletin Tris-acetate:tris(hydroxymethyl)aminomethane acetate Tris-HCl:tris(hydroxymethyl)aminomethane hydrochloride. vii.
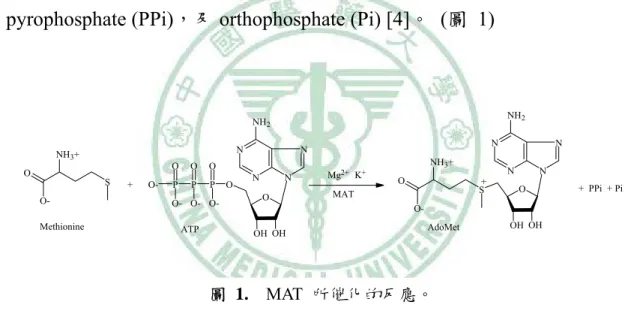
(11) 第一章 緒 論 第一節 MAT 的分子生物學 甲硫氨酸腺苷基轉移酵素 Methionine adenosyltransferase (MAT;SAMS; EC 2.5.1.6),存在於所有的生物細胞中,被認為是生命所必須的酵素之一 [1-3]。它先將 ATP 上的 adenosyl group 轉移到 L-methionine 的硫原子,形成 硫帶正電的 S-adenosyl-L-methionine (AdoMet;SAM),繼而水解三個磷酸根成 為 pyrophosphate (PPi),及 orthophosphate (Pi) [4]。 (圖 1). NH2. NH2 N. NH3+ O. S OMethionine. +. O-. O. O. O. P. P. P. O- O- OATP. O. N. N N. N O. Mg2+ K+. NH3+ O. N + S. MAT. N N O. + PPi + Pi. OOH OH. AdoMet. OH OH. 圖 1. MAT 所催化的反應。. 從細菌乃至到人類,MAT 不論在整體的胺基酸序列還是在構形上,尤其 是活性部位,其胺基酸序列在演化過程中被高度的保存著 [5-10],就拿老鼠 和人類肝臟 α subunit 來比較,序列就有 95% 的相似度 [3]。哺乳動物的 MAT. 1.
(12) 可分為兩大類:肝系列 (liver specific) 的 MAT I/III1 與非肝系列 (non-liver specific) 的 MAT II2,前者來自於基因 MAT1A,後者來自於基因 MAT2A 和 MAT2B (Table 1),MAT1A 表現 α1 subunit,MAT2A 表現 α2 subunit,MAT2B 則表現 β subunit [9,11-16]。. Table 1. MAT isozymes 之間的比較 Catalytic MAT isozyme subunit. MAT I MAT II MAT III. α1 α2 α1. Gene encoding MAT catalytic subunit. MAT1A MAT2A MAT1A. Regulatory subunit. β. Gene encoding MAT regulatory subunit. Subunit composition. MAT2B. (α1)4 (α2, α2′)xβy (α1)2. MAT I/III 與 MAT II 在特性上有一個明顯的區別,就是體外活性分析時, 若有 10 % DMSO 存在,則 MAT I/III 活性會提高 (尤其是 MAT III 活性提高 5 倍),而 MAT II 活性則是被抑制下來 [17]。此外,胎兒肝臟所表現出的 isozyme 是 MAT II,出生後則表現 MAT I/III,這一點和 α-fetoprotein 及 albumin 一樣, 都是肝細胞分化成熟的指標 [18]。. 1 2. MAT I 又稱 MAT α 或 MATI; MAT III 又稱 MAT β 或 MATH。 MAT II 又稱 MAT γ 或 MATL。. 2.
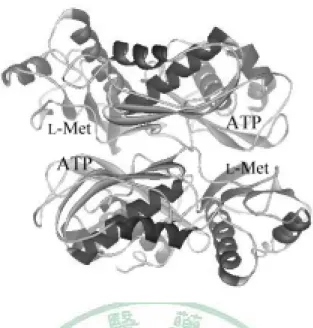
(13) MAT I 由 4 個 α1 組成 homotetramer,MAT III 由 2 個 α1 組成 homodimer, 兩者只存在於成熟的肝細胞 [19-20]。兩個 α subunit 組成 dimer 的作用力較 強,兩個 dimer 組成 tetramer 的作用力較弱 [21],在肝臟中,dimer 與 tetramer 處於平衡狀態 [22],雖然兩者同樣是 α1 所組成,卻表現出不一樣的性質,除 了動力學特性不同外,MAT I 的性質是 nonhydrophobic 而 MAT III 則是 hydrophobic [22-23];MAT I 會被 AdoMet 回饋抑制,而 MAT III 則反被 AdoMet 活化 [24-25]。 事實上,兩個 α subunit 組成 dimer 是 MAT 催化反應的必要條件,如圖 2,當兩個 α subunit 組成 dimer 時,L-Met binding site 與 ATP binding site 對 應疊合,形成了兩個活性部位,此時 MAT 才具有活性 [26];除了兩個活性部 位外,還有另一個凹洞,是相當厭水性的區域,是否有生物分子與之相結合, 是值得研究的主題。. 3.
(14) 圖 2. 老鼠肝細胞 α subunit 模型 (PDB code:1QM4),圖中標示出 L-Met 和 ATP 的結合位置。兩個 α subunit 組成 dimer 時,L-Met binding site 與 ATP binding site 對應組合成一個 binding site,故總共可形成兩個活性部位。. MAT II 則廣泛存在於所有組織細胞,肝臟中也有少量 [9,11-16],事實上, MAT II 的組成目前還不是很了解。其組成包含 β subunit,是一個無催化活性 的調節單位。 在人類,α2′ 則是 α2 轉譯後處理的產物 [27],具酵素活性, 也能被 α2 的抗體辨識,故兩者差異不大。β subunit 和 α2、α2′結合後,會提 高其活性,但也同時使其更容易受到 AdoMet 的回饋抑制;沒有 β subunit 存 在時,其對 methionine 的 Km 為 75 µM,有足夠量的 β subunit 時,其對 methionine 的 Km 為 17 µM [28]。 影響 MAT 活性的因素很多,除了 MAT I/III tetramer 與 dimer 的轉變和 MAT II 的 β subunit 外,還有回饋抑制如 AdoMet、Pi、PPi、polyamine [28-30],. 4.
(15) 不過這些代謝物的抑制作用都不強;在 MAT1A 、MAT2A 基因上的調控則有 DNA methylation 抑 制 轉 錄 , histone acetylation 增 加 轉 錄 [31-32] ; glucocorticoid 亦會促進 MAT1A 基因的轉錄以及增加其 mRNA 的安定性 [33];蛋白質的修飾,則以 S-nitrosylation 最重要,當老鼠肝細胞的 MAT, C121 硫原子被 nitrosylation 後則失去活性 [34-37],而被 nitrosylation 的 MAT,可 被 GSH 再次活化,這表示了 NO 與 GSH 在 MAT 活性調控上的重要性 (圖 3) [37]。. 圖 3. NO 與 GSH 對 MAT 的調節作用 [37]。. 除了受上面因素影響外,另有研究報告指出,當細胞培養時,培養液中 methionine 濃度愈低時,其 MAT 的 Km,Vmax 均相對增加 [38]。 此外,不同 物種來源的 MAT 其 Km、Vmax 都不同,從酵素的胺基酸序列及結構來看,活性 部位差異不大,唯活性部位外有一曲環 (flexible loop),隨物種來源而有不同,. 5.
(16) 被認為是影響其動力學的重要因素。此曲環是一段活躍而不安定的構造,可 以打開或關閉,其構形與溫度、受質存在與否有關 [39],前面所提發生 S-nitrosylation 的 C121 位置就在曲環上面。 由於影響 MAT 酵素動力學特性的因素很多,而且不一樣的研究方法,不 同的酵素純度都影響結果,所以許多研究報告都有不小的差異。一般而言, MAT II 對 methionine 的 Km 為 4~10 µM,MAT I 為 23 µM ~ 1 mM,MAT III 為 215 µM ~ 7 mM [40]。. 6.
(17) 第二節 Methionine 的代謝路徑及其在醫療上的意義 整個 methionine 代謝路徑如圖 4 所示,methionine 經 MAT 作用後產生了 AdoMet,AdoMet 是體內代謝過程中甲基的主要來源,可接受甲基的分子包括 DNA、RNA、proteins、phospholipids、及其他小分子。透過許多不同甲基轉 移酵素的作用,能在不同的分子上甲基化,而進行代謝作用或調節生理功能 [41]。 除了提供甲基外,AdoMet 經過 decarboxylation,能提供 aminopropyl group,而參與了 polyamine (putrescine、spermidine 與 spermine) 的生合成。 在 polyamine 生合成路徑也有一些與細胞分化有關的研究,如 spermidine 的代 謝產物,N1-acetylspermidine 及 N8-acetylspermidine 在誘導 HL-60 細胞分化的 能力上有著極不同的結果,N8-acetylspermidine 在 15 µM 時,即能誘導 80% ~ 90% HL-60 細胞進行分化,而 N1-acetylspermidine 一直到 1.2 mM 都無誘導分 化,也不會抑制細胞生長 [69]。此外,Ornithine decarboxylase 的抑制劑 difluoromethyl ornithine,能誘導 F9 teratocarcinoma stem cells 進行細胞分化 [70],但一般還是認為,polyamine 與細胞增殖是較有關係的 [71-73]。. 7.
(18) 圖 4. Methionine 的代謝路徑 [41]。. 8.
(19) AdoMet 轉移甲基後成為 S-adenosylhomocystein (AdoHcy; SAH),本身是 一種甲基轉移酵素抑制劑, SAHH 的抑制劑會誘導細胞分化,原因可能是 AdoHcy 量提高的緣故 [41]。AdoHcy 經水解後,生成 homocystein (Hcy),近 來發現 Hcy 是個心血管疾病的危險因子,當缺乏葉酸、維生素 B6、B12 等時, Hcy 會無法轉變為 methionine 而堆積,而與動脈粥狀硬化有關 [74-75]。 Hcy 又是 glutathione (GSH) 生合成前驅物,GSH 具有抗氧化功能,而 AdoMet 的量是否因此影響到細胞抗氧化以及老化、凋亡,仍不確定。 其實 AdoMet 還有其他用途,儘管藥效學或藥動學上了解不多,AdoMet 仍被認為對肝臟有保護作用,慢性肝病的 MAT 活性減少,由於 AdoMet 減少 使得 GSH 也減少,而在肝臟疾病治療上,給予 AdoMet 後,肝臟的 GSH 將增 加 [3,76]。還有研究顯示,口服 AdoMet 可能可以通過血腦障蔽 [77],並影響 中樞 monoamine 代謝,在歐洲已用於治療憂鬱症 [78-81]。AdoMet 和軟骨組 織修復有關,也被認為對關節炎有療效 [82]。目前 AdoMet 已為一種健康食 品上市。. 9.
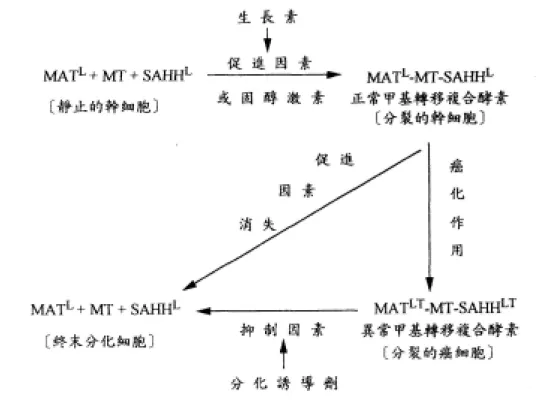
(20) MAT 與癌症的關係,一般認為和 DNA 的甲基化有關。DNA 的甲基化通 常發生在 CpG,使 cytosine 變成 5-methylcytosine,cytosine 甲基化後易突變 為 thymine,可能與腫瘤發生有關。基因的 promoter 若被甲基化,則這段基因 常無法表現 [42-44]。有研究報告認為,若阻斷 DNA 甲基化,或使 DNA 去甲 基化,可能可以活化某些被抑制的基因,進而誘導癌細胞分化並表現細胞功 能,如 DNA 甲基轉移酵素的抑制劑 5-azacytidine 等 [41,45-46]。由以上可知, DNA 的甲基化,在基因表現與癌症發生上扮演相當重要的角色,同時也是藥 物設計的新標靶 [47]。 癌細胞是細胞分裂與分化過程發生錯誤,以致於細胞停留於分裂週期運 轉不息,不能遂行終末分化,所以 Potter 認為細胞分化被阻斷是癌症的最中心 問題 [48]。廖明徵博士發現癌細胞都有異常甲基轉移複合酵素,導致細胞不 能進行分化,因此異常甲基轉移複合酵素是癌症的最根本病因 [49-51]。這種 異常酵素在癌細胞中調節細胞分裂及分化的功能可由圖 5 解詳加說明:. 10.
(21) 圖 5. 異常甲基轉移酵素調節細胞分裂和細胞分化的角色。. 11.
(22) 如圖 5 所示,MAT、MT、SAHH 三個成員酵素在正常細胞中要結合成甲 基轉移複合酵素才能發揮甲基轉移的活性,而這種結合完全依賴外來活化因 子(生長素、固醇激素),當這些活化因子存在時複合酵素活性提高,核糖體的 生產和甲基的複製就可以順利進行,細胞就在分裂週期運轉。然而一旦活化 因子消失,甲基轉移複合酵素失去活性,此時細胞會合成缺甲基的核酸,這 是細胞由分裂轉入分化的關鍵,核糖體不再生產 [52],分化基因得以表達, 細胞被導入終末分化變成不再分裂之終末細胞 [49-51]。癌細胞會產生特異的 蛋白質因子 (specific protein factor) 與 MAT 和 SAHH 結合後再與 MT 結合在 一起,形成 MATLT-MT-SAHHLT,這種異常之甲基轉移複合酵素活性高且非常 穩定,會使甲基化之活性劇增,致使細胞分裂之速度比正常細胞快,不受外 來活化因子控制,能自行維持長期之活性,使癌細胞在分裂週期運轉不息 [49-51]。癌細胞須外來抑制物質 (如 CDA-II) 將甲基轉移複合酵素之癌因子 去除,癌細胞就會像正常細胞進行終末分化。 廖明徵博士進一步地發現所有之癌組織都存在異常甲基轉移複合酵素, 更指出正常組織中不管是否正在進行細胞分裂都沒有異常甲基轉移複合酵素 之存在,所有癌細胞都有異常甲基轉移複合酵素,造成細胞不斷分裂。因此 他們認為能抑制異常甲基轉移複合酵素之活性化合物,才是理想之癌細胞分 化誘導劑。而這種作用在異常甲基轉移複合酵素的癌細胞分化誘導劑,可以. 12.
(23) 從 MATLT 的抑制活性檢測篩選出來,因為當 MATLT 失去腫瘤因子後恢復成 MATL,活性會降低,而且兩者的動力學常數不一樣,MATL 對 methionine 的 Km 為 3 µM,而 MATLT 為 20 µM [49-51]。 從這個理論看來,固醇激素、腫瘤因子與癌細胞分化誘導劑似乎可能作 用在同一個地方,先前提過 MAT 的兩個 α subunit 形成 dimer 後,除了產生兩 個活性部位外,還有一個厭水性區域的凹洞,這裡是否就是固醇激素,腫瘤 因子,與癌細胞分化誘導劑的結合位置,仍需進一步研究。 在癌細胞中通常可發現致癌基因 (oncogenes) 的 hypo-methylation,與抑 癌基因 (suppressor genes) 的 hyper-methylation [53-59],然而造成這種異常的 甲基分布不均,原因仍不清楚。正常細胞的 DNA,其甲基化乃受 maintenance methyltransferase 所催化,而癌細胞中有 de novo methylation 發生,de novo methylation 究竟是由 maintenance methyltransferase 所催化,或是由其他異 常 methyltransferase 催化,則仍屬未知 [47]。癌細胞異常甲基轉移複合酵素中 的 methyltransferase 有何不同,以及它與這些異常甲基分布是否有關,亦有待 研究。 一般來說,MAT 與快速生長的細胞有關,尤其是 MAT II [60-63],也就是 MATL。除了廖明徵博士等人所提的 MATLT 以外,還有許多 MAT 異常表現或 突變的研究報告。如肝癌細胞中,MAT1A 基因沒有表現出來,反而表現 MAT2A. 13.
(24) 基因,其 MAT 活性在正常 methionine 生理濃度下,比正常肝細胞的 MAT 活 性來的高,但卻比較容易被 ethionine 所抑制,因此被認為是有選擇性的標靶 [40]。從 Hypermethioninemia 病人的研究,發現是一種 MAT 先天缺陷疾病, 肝臟中的 MAT1A 基因發生 R264H 突變,使得兩個 α1 subunit 無法形成 dimer, 同時也失去活性 [64-66],MAT 的缺陷甚至也和 brain demyelination 的發生有 關 [67-68]。 以上可知,整個 methionine 代謝路徑影響層面很廣,其中 AdoMet 是這個 路徑的關鍵,這也突顯了 MAT 的重要性。. 14.
(25) 第三節 過去 AdoMet 與 MAT 活性之檢測方法 如前所述,MAT 與許多重要的生理功能與疾病有關,而其活性之測定也 是相關研究所必須的,包括生理檢測以及抑制劑篩選等,然而目前為止 MAT 的活性分析幾乎都是以放射線同位素測定,以廖博士等的方法為例 [83],其 分析的原理為利用 L-[methyl-3H] methionine 當作 MAT 的受質,酵素催化反應 後,產生有放射性的 AdoMet,由於 AdoMet 的硫原子帶正電,故用 P81 陽離 子交換層析試紙吸附後,再以 pH 7 的緩衝液洗去殘留的 L-[methyl-3H] methionine,接著偵測剩餘的放射線量,若需要定量則可以 L-[methyl-3H] methionine 做檢量線定量之。 HPLC 分析法方面,已有不少測定 AdoMet 的方法被建立 [84-92],都是 用以測定組織細胞中 AdoMet 的含量,由於從組織來的細胞數目夠多,通常 UV 偵測器的感度已經足夠。但是如果要將其應用於酵素活性分析,尤其是細 胞來源有限的 culture system cells 之酵素活性分析,或者如血漿中 AdoMet 濃 度低的檢品,這些方法感度都不夠,而且需用到大量檢品,及繁瑣或耗時的 分析程序。其中代表性的方法如 1992 年 Jennifer 等人 [84] 及 1997 年 Carolyn 等人 [85] 所發表,螢光法則由 1998 年 Capdevila 等人 [86] 所提出,還有本 所先前的研究使用 BIACORE 檢測 MAT 與小分子親和力的方法 [112]。這些 方法簡述如下:. 15.
(26) 1. 1992 年 Jennifer 等人之方法: Solid-phase extraction 前處理 ⇓檢品置陽離子交換樹脂 (cation-exchange resin) ⇓以甲醇沖洗 ⇓以 0.05 M potassium dihydrogen phosphate buffer (pH 2.0) 沖洗 ⇓以 0.05 M disodium hydrogen phosphate buffer (pH 11.0) 洗出 AdoMet. HPLC analysis Column: C18 reversed-phase analytical column (100 mm × 4.6 mm) Mobile phase: 0.05 M potassium dihydrogen phosphate buffer (pH 5.7) Flow rate: 1.2 ml/min UV Detector: 254 nm Analysis time: 8 min 此方法用 cation-exchange resin 做前處理,目的在先除去大量雜質,然而 UV 檢測器感度不夠,若要應用到酵素分析上,檢品、試劑用量以及反應時間 均需要大幅度增加。 如果酵素來源是培養的細胞,很難供應大量的酵素蛋 白,而且 AdoMet 不安定,延長反應以及處理的時間會損失更多的 AdoMet。. 16.
(27) 2. 1997 年 Carolyn 等人之方法: ⇓於 0.1 M sodium acetate buffer (pH 6.0) 打破細胞 ⇓trichloroacetic acid 沉澱蛋白質 ⇓離心:25,000g,10 min,5°C ⇓取上清液,再用乙醚萃取除去低極性物質 ⇓水層 0.45 µm 過濾 ⇓檢品 0 ~ 4°C 保存 HPLC analysis Column: ODS,25 × 0.46 cm (5 µm) Mobile phase: 50 mM NaH2PO4 (pH 4.38),10 mM Sodium heptanesulfonate, 20% methanol Flow rate: 0.9 ml/min UV Detector: 254 nm Analysis time: 20 min 這個方法若應用於 MAT 酵素活性分析是有困難的,同樣有感度不夠的問 題,而且酵素反應時添加了許多高濃度物質會干擾 AdoMet 的分析,ion pair reagent 在使用上則較不易控制,試劑配製需更加精準,HPLC 平衡時間需較 長,column 溫度影響也較大。. 17.
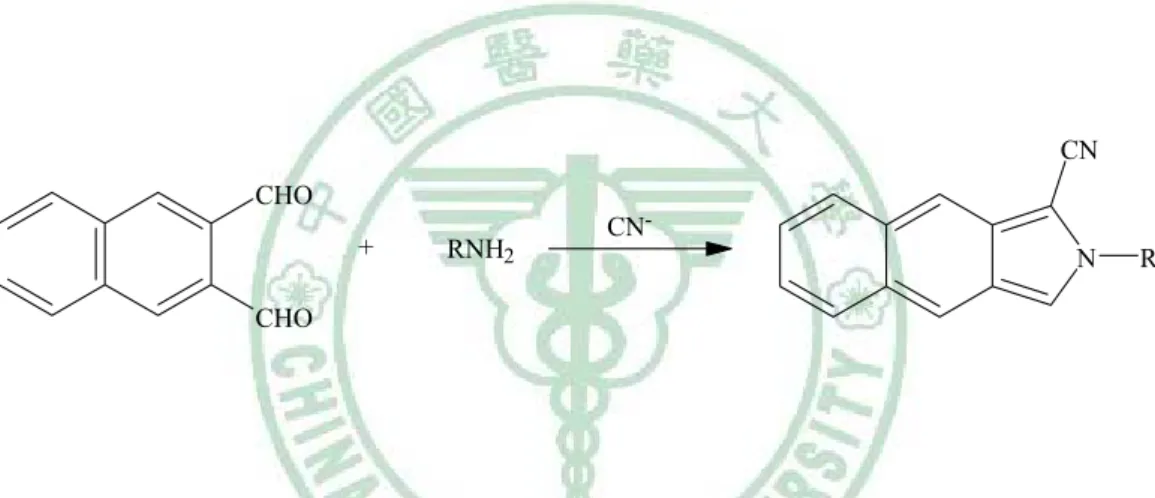
(28) 3. 1998 年 Capdevila 等人之方法 檢品先經過 HPLC,以 C-8 column 初步分離後,再利用 AdoMet 的 amino group 與 naphthalenedialdehyde 及 cyanide 於 pH 9.0 反應 10 分鐘,形成 isoindole 類螢光化合物 (圖 6),接著用 HPLC 以 C-18 column 及螢光檢測器進行分析。 這個方法為螢光分析法,感度較高,測得血漿中 AdoMet 的濃度為 102.7 nM, 然而過程繁瑣是其缺點,而且所使用之試劑毒性較大。. CN CHO +. RNH2. CNN. R. CHO. 圖 6. AdoMet 之 isoindole 類螢光化合物形成原理。1998 年 Capdevila 以 naphthalenedialdehyde 及 cyanide 於 pH 9.0 反應 10 分鐘進行衍生。. 18.
(29) 4. BIACORE - Biomolecular Interaction Analysis BIACORE 是應用表面薄膜共振技術 (surface plasmon resonance, SPR), 去分析生物分子間的相互作用,包括結合與解離的情形,雖然並不是真正的 在檢測 MAT 酵素活性,不過卻也是一種 MAT 與受質,或者與抑制劑之結合 親和力的探討方式。本所先前的研究中,也利用此設備與技術,探討了合成 之 hemoregulatory peptide (HP) 系列衍生物對 MAT 酵素之結合作用[112]。表 面薄膜共振技術是一種光學現象,先將分子鍵結到晶片上面,當另一種分子 加入移動溶液,流經過晶片時,會因結合與未結合呈現不同的訊號,因而能 夠很快速的初步了解,兩分子之間是否可能有結合作用力。. 19.
(30) 在酵素活性的測定方法中,酵素偶合 (enzyme-coupled) 法是在進行酵素 反應時,另外加入一個以上的酵素,將欲檢測酵素之產物轉變成較安定,或 者感度較高,或者較易分離的化合物,再進行分析,是一種間接的分析方法, 亦曾應用於一些酵素的活性分析 [93-102]。 本研究開發了一個新的酵素偶合螢光分析方法,並應用 HPLC 作為定性 定量的工具。我們利用 catechol-O-methyltransferase (COMT) 以及已知的 COMT 螢光受質 esculetin (ECL) [103-106] 來分析 MAT 活性與動力學常數測 定。反應中 MAT 產生的 AdoMet 會被 COMT 轉移甲基給 ECL,接著經 HPLC 分析被甲基化的螢光物質 scopoletin (SPL),進而評估 AdoMet 的產量及 MAT 的活性 (Scheme 1)。這個方法的評估及其可行性、優缺點將於本文陳述討論。. 20.
(31) Scheme 1. Stepwise representation of the COMT-coupled fluorometric assay of MAT activity. The methyl group of AdoMet was transferred to esculetin by coupling with COMT and the formation of scopoletin was measured by HPLC fluorometric analysis. MGBG was used to prevent AdoMet from decarboxylation.. 21.
(32) 第二章 研究動機與目的. 如上所述,MAT 是一個重要的酵素,在整個 methionine 代謝途徑中扮演 了一個關鍵角色,它關係到細胞的增殖分化,基因表現,乃至於牽涉到許多 疾病的發生包括心血管疾病、肥胖、帕金森氏症、老化以及癌症 [107-108]。 廖明徵博士提出的 methylation complex [49-52,83],被認為是造成癌細胞 不能進行終末分化的主因,本實驗室根據此理論基礎,從尿液抽取物以及合 成相關化合物中,希望尋找到作用在此標的之理想有效的癌細胞分化誘導劑。 過去一直以放射線同位素的方法,進行 MATLT 的抑制劑篩選,由於放射 線同位素有環境污染以及對操作者有害的顧慮,而且目前並無一快速簡便, 感度又高的分析方法可用於 MAT 的活性分析與動力學研究,所以著者嘗試開 發螢光分析法,以期應用於 MAT 的相關研究。同時也希望這個方法將來能應 用於臨床診斷以及抗憂鬱藥 AdoMet 的血中濃度分析。. 22.
(33) 第三章 實驗器材與方法 第一節 實驗材料與儀器 (一) 化學藥品 1. 購自 NewTM Life Science Products, INC 者 3. L-[methyl- H] methionine (1 mCi/ml ; 70-85 Ci/mmol). 2. 購自 EcoLume 者 Cocktail 3. 購自 Sigma Chemical Co. (St.Louis,MO) 者 Adenosine 5′-triphosphate Catechol-O-methyltransferase: One unit will catalyze the methylation of 1.0 nmol of protocatechuic acid per h at pH 7.9 at 37°C. Dimethyl sulfoxide 1,4-Dithiothreitol L-Methionine. Methylglyoxal bis-(guanylhydrazone) (MGBG) Tris(hydroxymethyl)aminomethane hydrochloride 4. 購自西德默客 (Merk) 公司者 Ethanol. 23.
(34) Hydrochloric acid Isopropanol Magnesium chloride Methanol Perchloric acid Potassium chloride Potassium dihydrogen phosphate Sodium dihydrogen phosphate Sodium hydroxide Sulphuric acid Tris(hydroxymethyl)aminomethane acetate 5. 購自 Aldrich 者 2,2′-Dithiobis(1-aminonaphthalene) 6. 購自日本東京化成 (TCI) 者 3,4-Dihydroxybenzaldehyde Esculetin Scopoletin Tri-n-butylphosphine 7. 購自 American biorganicx, INC 者 Piperazine-N-N′-bis(2-ethanesulfonic acid) 24.
(35) 8. 購自 Tedia Company, INC 者 Chloroform Ethyl acetate n-Hexane Methanol (二) 實驗儀器 Beckman 放射線自動偵測儀器 Model No: L96500 Homogenizer HP 1050 Series of HPLC Modules HP 1046A Prgrammable Fluorescence Detector HP 3394A integrator Incubator Merck Lichrocart 125-4 Lichrospher Si 60 (5 µm) 離心機 (Hettich zentrifugen) (三) 癌細胞株 HL-60 cell line: American Type Culture Collection (Maryland, USA)。. 25.
(36) 第二節 細胞培養與 MAT 酵素製備 HL-60 細胞懸浮培養於包含 15%胎牛血清之 RPMI 1640 培養液。培養環 境控制維持於溫度 37°C、飽和溼度、以及 5% CO2 [113]。 MAT 酵素溶液的製備,則取 2×107 HL-60 cells,以 1 ml 含 0.25 m 蔗糖 的高張 lysis buffer (0.05 M Tris-acetate, pH 7.0; 1.5 mM MgCl2) 清洗,離心 (4 °C, 1500 rpm, 3 min),倒去上清液,沖洗第二次,離心 (4 °C, 300 g, 3 min), 再以不含蔗糖的低張 lysis buffer (0.05 M Tris-acetate, pH 7.0; 1.5 mM MgCl2) 充分混和後,於冰浴中靜置 5 min,接著在冰浴中以 homogenizer 將細胞打破, 高速離心 (4 °C, 10,000 g, 10 min),小心吸取上清液,置於 1.5-ml 離心管,此 即含 MAT 酵素的抽取液。. 26.
(37) 第三節 MAT 活性分析方法 Method I: Traditional Radioactive Method 測定 MAT 酵素活性之放射性同位素檢測法原則上採用先前的研究方法 [114]。簡單來說,酵素反應液中包含了 10 µl 酵素抽取液 (2.5 µg of protein)、 20 µl 的酸鹼緩衝液 (500 mM PIPES, pH 7.0; 750 mM KCl; 25 mM MgCl2 and 25 mM DTT)、10 µl 的 20 mM ATP、50 µl 的去離子水,以及 10 µl 的 1.2 µΜ L-[methyl-3H]methionine (0.1 mCi/ml) ,最後反應液總體積為 100 µl。空白 試驗則是以不包含 ATP 時之檢測值作為參考值。酵素反應液於 37°C 下反應 30 分鐘,然後移至冰浴中,以 10 µl 之 4 N HClO4 終止酵素反應。經過離心 後 (3,000 g, 1 min),90 µl 的上清液被吸取出來轉移到 P-81 陽離子交換層析 試紙上 (1×1 inch)。然後 P-81 陽離子交換層析試紙使用 100 ml 之 10 mM KH2PO4 清洗十次,以去除含有放射性的受質 (L-Met)。乾燥後,P-81 試紙置 於 5-ml counting vial,同時以 150 µl 的 2 M KCl 將 3H-AdoMet 從試紙上沖洗 出來,加入 3 ml 的閃爍計數液,利用β-counter 檢測其放射性含量。. 27.
(38) Method II: COMT-coupled Method 大致上來說,Method II 的酵素反應條件與 Method I 方法相同,不過不使 用放射性物質,而是使用偶合酵素的方式進行。要配製成酵素反應液 100 µl, 首先加入 10 µl 酵素抽取液 (2.5 µg of protein),20 µl 的酸鹼緩衝液 (500 mM PIPES, pH 7.0; 750 mM KCl; 25 mM MgCl2 and 25 mM DTT) 以及 20 µl 去離 子水, 並加入 10 µl 的 1 mM Methylglyoxal bis-(guanylhydrazone) (MGBG) 水 溶液,用以防止 AdoMet 發生 decarboxylation 反應。接者,為了檢測 AdoMet 之生成,需加入 10 µl 的 50 units COMT 酵素溶液 (50 mM Tris-acetate; pH 7.0, and 1.5 mM MgCl2; 使用前保存在-80°C),以及 10 µl 溶於 isopropanol:water (1:1, v/v) 之 10 mM esculetin。最後,10 µl 的 20 mM ATP 以及 10 µl 的 10 mM L-Met 加入酵素反應液中。空白試驗則是以不包含 ATP 時之檢測值作為參考. 值。酵素反應液經由 Vortex mixer 混合並稍微離心後 (300 g for 1 min),在 37°C 下反應 30 min,移至冰浴中,加入 100 µl boric acid 飽和溶液終止酵素反應。 Boric acid 同時也用於和 esculetin 的 catechol 部位反應形成水溶性鹽類。然後 使用 300 µl 的 n-hexane:ethyl acetate (7:3, v/v) 萃取所生成之 scopoletin。 接著 混合液經過 Vortex 混合 1 min,經過離心 (1,500 g for 2 min) 使之分層,從上 層有機溶媒層取出 40 µl 用於 HPLC 分析,並定量所生成之 scopoletin。. 28.
(39) Calibration Curve for Method II 使用 HPLC 進行分析已知量的 scopoletin,用以製作檢量線,酵素反應在 理想條件下所測得 scopoletin 的量,可代表 AdoMet 之生成量。同時所測得的 scopoletin 定量時,以其回收率進行校正。MAT 酵素活性之表示,使用 units/mg protein,其中 1 unit 的蛋白定義為 1 小時內可生成 1 nmol AdoMet。. Kinetic Properties of MAT in HL-60 Cells MAT 酵素動力學則使用了所建立的 COMT 偶合螢光檢測液相層析法進行 研究。MAT 受質 L-Met 濃度範圍從 1.6 到 1,000 µM,而 ATP 則固定在 2 mM。 所得到的數據以 SigmaPlot 軟體之 Enzyme Kinetics Module (SPSS Science Inc.) 進行分析。MAT 動力學之作圖使用 Eadie-Hofstee plots,並且使用 Hill equation 進行回歸,以求得動力學參數。. HPLC System HPLC 系統使用 HP 1050 Series 之 HPLC Modules,配接 Merck Lichrocart 125-4 Lichrospher Si 60 (5 µm) 分析型管柱,層析結果則以 HP 1046A Programmable Fluorescence Detector (ex 347, em 415) 和 HP 3394A integrator 進 行檢測和記錄。移動相是 dichloromethane : methanol : acetic acid (40:1:0.06,. 29.
(40) v/v/v) 之混合溶液,流速為 1 ml/min。經過系列分析使用後,HPLC 系統則使 用 ethyl acetate : methanol : acetic acid (40:1:0.06, v/v/v) 清洗 1 小時。. Assay of Protein 利用 Bradford 方法 [109],由 bovine serum albumin (BSA) 做標準蛋白溶 液及 Bio-Rad 蛋白質 assay kit 當反應之染料。配置各個已知濃度之標準液,以 分光光度計 (spectrophotometer),設定波長 595 nm,檢測其吸光度,做成檢量 線,再標定欲測定之酵素製劑。. 30.
(41) 第四章 結果與討論 使用 HPLC 以及 UV 檢測器去分析 MAT 酵素活性通常都達不到理想的感 度而無法檢測。此外,AdoMet 相當容易吸潮而且不安定,根據研究,純化過 的 (S,S)-AdoMet 在 37°C、pH 7.5 環境下 4 小時後可能已經衰減 14%。其中分 解 後 可 產 生 adenine 、 S-pentosylmethionine 、. 5′-deoxy-5′-(methylthio). adenosine、homoserine lactone 以及無生理活性的 (R,S)-AdoMet stereoisomer [115]。這也代表著,在分析過程中如果歷經太長時間的酵素反應以及檢品處 理,AdoMet 亦將自行衰減並且產生更多干擾分析的產物。因此,AdoMet 要 準確定量並不容易,而且也不適合作為參照標準品 (reference standard)。要克 服這些困難,則可以利用偶合酵素的方式,將 COMT 加入於 MAT 的酵素反 應中,當 AdoMet 產生後,隨即被 COMT 將其甲基轉移至 esculetin,因而克 服 AdoMet 變質分解而影響定量的問題。此外,所生成的 scopoletin 較安定而 且具有高螢光性質,定性定量上都容易許多,故在最佳化的酵素反應條件下, 採用 scopoletin 當作定量 AdoMet 之參照標準品是較適當的,在應用上也較簡 便。. 31.
(42) Evaluation of the COMT-coupled Reaction 在 COMT 偶 合 的 方 法 中 , 由 於 esculetin 水 溶 解 度 不 佳 , 因 此 溶 於 isopropanol:water (1:1, v/v)。最後酵素反應液裡共含有 5%的 isopropanol,經 過驗證,其對酵素反應沒有抑制作用。參照標準品 scopoletin 溶液平時儲存於 -20°C 中,其安定性至少能維持一個月。Scopoletin 溶解在移動相溶液中,其 螢光之激發波長與放射波長分別為 347 及 415 nm。標準品之 HPLC 分析,使 用 2 pmol scopoletin 以及 20 pmol 的 esculetin。 Scopoletin 的滯留時間約為 2.95 min,而 esculetin 則滯留於 Si 60 管柱 (圖 7A, inset)至少超過 30 min。為 了避免過量的 esculetin 積存於管柱內,加入 100 µl 的 boric acid 飽和溶液,除 了可終止酵素反應外,還具有和 esculetin 反應形成高水溶性鹽類的功能,使 esculetin 在萃取過程中停留於水層。萃取過程中,esculetin 的回收率少於 1% , 而 scopoletin 則約為 80%。 為了確認 Method II 中偶合酵素之機轉,酵素反應分別在 ATP (2 mM)、 esculetin (1 mM) 以及 S-adenosylhomocystein (AdoHcy; 100 µM) 存在與否的 條件下進行分析。AdoHcy 是強效的非專一性 COMT 競爭性抑制劑,能和 AdoMet 競 爭. (圖. 7A) 。 同 樣 的 實 驗 , 也 以 10 µl 的 1.2 µM. 3. L-[methyl- H]methionine (0.1 mCi/ml) 代替 10 µl 的 10 mM 非放射性 L-Met 重. 複驗證 (圖 7B),如此則能利用放射性同位素標定甲基的 L-Met,追蹤其甲基. 32.
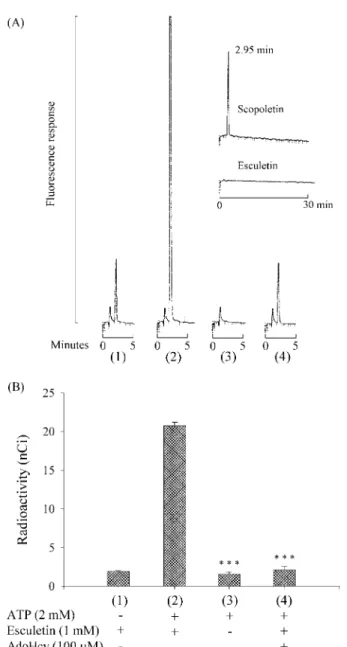
(43) 在 COMT 偶合酵素法測定 MAT 酵素活性過程中的流向。 在 COMT 偶合方法當中,當使用 L-[methyl-3H]methionine 作為 MAT 受質 時,其步驟如 Method II 所描述。但是當反應結束後,改以 300 µl 的 n-hexane:ethyl acetate (7:3, v/v) 進行萃取,100 µl 的上層有機溶液轉移到 5-ml 的計數瓶,加入 3 ml 的閃爍計數液,混合後進行放射性強度的檢測。Scopoletin 的生成量,以放射性強弱表示。 在 COMT 偶合酵素之螢光分析法中,空白實驗酵素反應液沒有加入額外 的 ATP 時 (圖 7A(1)) 有微量的 scopoletin 被檢測到。這可能來自於細胞抽取 液中微量的 AdoMet、ATP 以及部分存在於 esculetin 之雜質。當酵素反應液中 沒有加入 esculetin 時,則也沒有任何 scopoletin 被檢測到 (圖 7A(3))。另外, 當 100 µM AdoHcy 加入酵素反應液中,scopoletin 的生成跟控制組比較,明 顯地被阻斷 (圖 7A(4))。這些結果顯示,scopoletin 的生成,完全同時依賴於 MAT 和 COMT 的 活 性 。 同 樣 的 結 果 也 可 在 使 用 radioactive 3. L-[methyl- H]methionine 當作 MAT 受質之實驗觀察到 (圖 7B)。因此 COMT. 偶合酵素檢測 MAT 酵素活性的機轉可以得到進一部的確認。. 33.
(44) 圖 7. Confirmation of the mechanism of the COMT-coupled reaction in MAT activity determination. The different conditions for each reaction were (1) blank, lacking in ATP, (2) control, MAT activity measured in (A) and (B) was 122 units/mg protein and 0.74 units/mg protein, respectively, (3) lacking in esculetin and (4) in the presence of 100 µM AdoHcy. (A) HPLC chromatograms with fluorometric detection. A 10-µl aliquot of 10 µM L-Met was used as substrate, and the procedure was described in Method II. Inset shows the retention time of 2 pmol of scopoletin was 2.95 min and 20 pmol of esculetin strongly retained in Si 60 column over 30 min. (B) The same experiments were repeated, but a 10-µl aliquot of 1.2 µM 3 L-[methyl- H]methionine (0.1 mCi/ml) was used instead. The other conditions were the same as Method II, but after the reaction was terminated and the reaction mixture was extracted with 300 µl of n-hexane:ethyl acetate (7:3, v/v), 100 µl of upper organic layer was used to count radioactive intensity. Other details are described in the text. Values are represented as mean ± S.D. of three experiments. Significant differences were calculated using Student's unpaired t-test. *** P < 0.001 compared to (2).. 34.
(45) Effect of MGBG on the Determination of MAT Activity in Method I and Method II 在過去許多研究測定 MAT 酵素活性時,使用的傳統分析方法中 ,AdoMet decarboxylase 抑 制 劑 都 沒 有 被 加 入 到 酵 素 反 應 液 裡 。 然 而 AdoMet decarboxylase 勢必存在細胞抽取液當中,而且可能會減少 MAT 活性測定時所 能檢測得到的 AdoMet 之量。所以 MGBG 在這個研究裡,被使用來阻斷 AdoMet decarboxylase 之活性。為了探討 MGBG 對 COMT 偶合酵素法檢測 MAT 酵素活性時之影響,同樣利用 10 µl 的 1.2 µM L-[methyl-3H]methionine (0.1 mCi/ml) 當作 MAT 受質,並分別使用 Method I 以及 Method II 進行分析 比較 (圖 8)。其結果顯示 100 µM 的 MGBG 存在於酵素反應液中時,能有效 阻斷 AdoMet decarboxylase 之活性,而使 COMT 偶合酵素的方法達到檢測的 最大值。同樣的反應條件下,Method II 檢測到的值似乎比 Method I 還要高一 些,這表示 Method I 在清洗 P-81 試紙過程中,難免有 AdoMet 流失掉,這個 實驗也同時看得出,COMT 偶合酵素能夠有效轉換非常微量的 AdoMet。此 外,兩種方法都顯示,在 100 µM 的 MGBG 存在時,所測得之 MAT 酵素活 性,大約是沒有 MGBG 時所測得之兩倍。 很明顯地,在過去的研究方法中, 如果使用了沒有純化的酵素溶液,又沒有阻斷 AdoMet 之 decarboxylation,將 造成很大的誤差。因此,從細胞抽取液中測定 MAT 酵素活性時,使用 MGBG 是不可或缺的。. 35.
(46) 圖 8. The effect of MGBG on the results of COMT-coupled method (Method II) and traditional method (Method I). A 10-µl aliquot of 1.2 µM L-[methyl-3H]methionine (0.1 mCi/ml) was used as substrate for both methods, and the radioactivity of 3H-AdoMet or 3 H-scopoletin was measured. Values are represented as mean ± S.D. of three experiments.. 36.
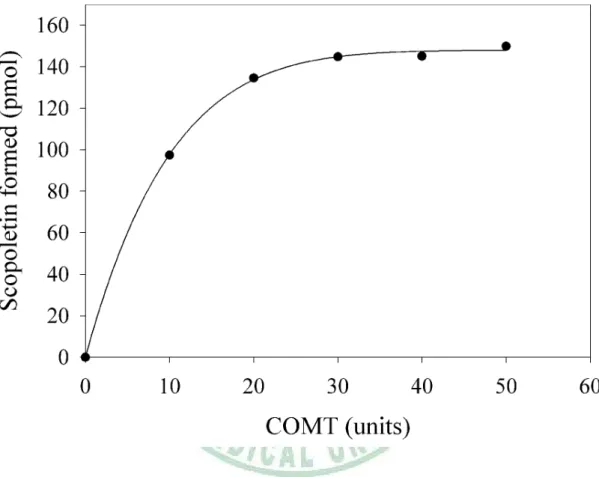
(47) Effect of COMT Concentration and Time-Course of Scopoletin Formation 這裡針對 COMT 使用量以及 scopoletin 生成之 time-course (10-60 min)進 行探討。圖 9 表示 Method II 中使用 50 units 的 COMT 是足量的。Scopoletin 生成之 time-course 顯示 COMT 偶合酵素法中,30-min 的反應時間介於良好 的線性範圍之內 (圖 10)。這些結果顯示,不論是 substrate depletion (L-Met, ATP or esculetin) 或者是 product accumulation (AdoMet or AdoHcy) 在 Method II 條件之分析過程中,都不會造成反應速率的限制與干擾。. 37.
(48) 圖 9. The effect of the quantity of COMT on scopoletin formation.. 38.
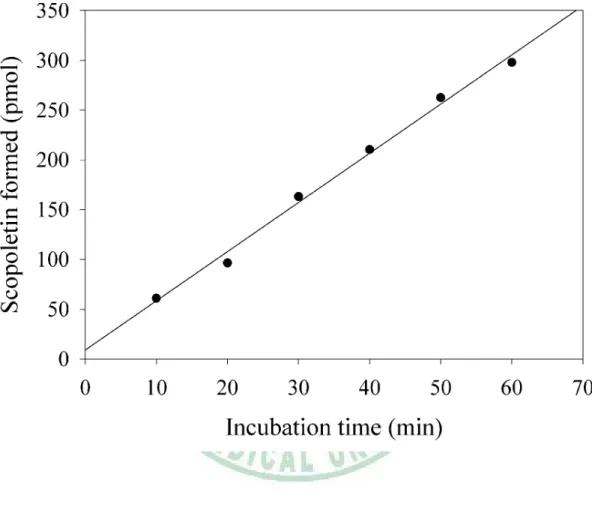
(49) 圖 10. Time-course of scopoletin formation. (R2 = 0.9933).. 39.
(50) Validation of the Conversion Efficiency of COMT-coupled Method Method II 所描述的檢測條件裡,COMT 偶合酵素轉換 AdoMet 的效能以 及 50 units 之 COMT 在 30-min 內所能轉換 (S,S)-AdoMet 量的限制範圍也被 研究探討。由於只有 (S,S)-AdoMet epimer 具有生理功能而當作甲基轉移酵素 的受質,因此,要進行確效前,必須先對標準品 AdoMet 進行 (S,S)-AdoMet 定量,方法是依據 L.K. Revelle 所描述之方法,使用 NMR 定量 [116]。結果 顯示,50 units 的 COMT 至少能夠依線性地有效轉換 50 到 500 pmol 範圍內 的 (S,S)-AdoMet,其 conversion coefficient 為 101 ± 5.6% (圖 11)。. 40.
(51) 圖 11. Validation of the conversion efficiency of COMT-coupled method. The conversion efficiency and the quantity limit of (S,S)-AdoMet were investigated with 50 units of COMT in 30 min. The converting ability of 50 units COMT is linear from 50 to at least 500 pmol of (S,S)-AdoMet with the conversion coefficient of 101 ± 5.6%.. 41.
(52) Determination of MAT Activity MAT 活性之測定,是在理想條件下,以檢測到之 scopoletin 的量進行計算。 已知量的 scopoletin 使用 HPLC 分析得到檢量線,其檢測極小值約為 100 fmol,而且 scopoletin 之螢光強度與其濃度呈現良好的線性關係 (圖 12)。. 圖 12. Scopoletin 檢量線。. Activity and Kinetic Properties of MAT in HL-60 Cells 依據本研究所設定的 COMT 偶合酵素之反應條件,對細胞抽取液中蛋白 質含量以及 MAT 酵素活性測定值之線性關係進行研究,圖 13 顯示, 0.5 到 7.5 µg 的細胞抽取蛋白量和所測得之 MAT 酵素催化速率呈現良好的線性關 係,而本研究使用的蛋白質量介於良好線性範圍之內。. 42.
(53) 圖 13. Linear correlation between enzyme activity and the amount of MAT.. 43.
(54) 接者,利用本研究建立的 COMT 偶合酵素螢光分析法,針對 HL-60 之 MAT 活性以及酵素動力學特性進行研究。結果所得之酵素動力學數據使用 Eadie-Hofstee plots 作圖表示,並且使用 Hill equation 進行回歸分析(v = 135.4s0.5/(6.10.5 + s0.5); v: units/mg protein, s: µM of L-Met) (圖 14)。這些結果顯 示,HL-60 細胞內所含之 MAT 酵素呈現了 negative cooperativity,其 Hill coefficient 為 0.5,這個結果和先前的 MAT II isozyme 的動力學研究相類似 [24]。在 200 µM 的 L-Met 存在下,MAT 達到反應速率的最大值。計算所得之 Vmax 為 135.4 ± 1.5 units/mg protein,和其他正常組織比較起來,高出了許多 (erythrocytes 約 13.9×10-3 units/mg protein;parietal cortex 約 25.72 units/mg protein) [108, 110]。而計算所得之 Km 為 6.1 ± 0.3 µM ,這個數據則和先前研 究 human leukemia cells (3.5-20 µM) 的數據相似 [27]。綜合以上所述,HL-60 細胞中的 MAT,呈現明顯的 negative cooperativity 以及特別高的酵素活性,都 可能表示有異常的 MAT 酵素的表現,或者有其他型式的 MAT 存在於 HL-60 細胞中。. 44.
(55) 圖 14. Eadie-Hofstee plots of kinetic properties of MAT in HL-60 cells. MAT activity was assayed by COMT-coupled fluorometric method. The concentration of ATP was fixed at 2 mM and L-Met was varied from 1.6 to 1,000 µM. The data were analyzed by SigmaPlot software with Enzyme Kinetics Module. The Eadie-Hofstee plots and Hill equation were used for kinetic analyses.. 45.
(56) 第五章 結論 在 MAT 的研究領域上還有很多是未知的,而其在醫藥上也扮演重要角 色。由於過去 MAT 活性分析除了放射線同位素法外,一直缺少一個高感度, 又簡便的分析方法。本研究我們建立了一個酵素偶合螢光檢測之液相層析法 來 分 析 MAT 酵 素 活 性 及 酵 素 動 力 學 常 數 。 這 個 方 法 利 用 catechol-O-methyltransferase 將 AdoMet 上的甲基轉移到 esculetin 上形成 scopoletin,接者利用 HPLC 螢光檢測分析這些被甲基化的生成物,進而檢測 出 MAT 的活性及酵素動力學常數。並測得 HL-60 MAT 的 Km = 6.1 ± 0.3 µM,Vmax = 135.4 ± 1.5 nmole AdoMet/mg protein/hr。 這個方法感度高,對 AdoMet 有專一性,分析時干擾少,過程簡便快速, 花費便宜,故此法在 MAT 活性分析以及酵素動力學研究上,提供了一個較好 的選擇,將來也可試著應用於臨床診斷與抗憂鬱藥 AdoMet 血中濃度監測。. 46.
(57) 參考文獻 1. Tabor, C.W. and Tabor, H. Methionine adenosyltransferase (S-adenosylmethionine synthetase) and S-adenosylmethionine decarboxylase. Adv. Enzymol. Relat. Areas. Mol. Biol., 1984;56:251-282. 2. Kotb, M. and Geller, A.M. Methionine adenosyltransferase: structure and function. Pharmacol. Ther., 1993;59(2):125-143. 3. Mato, J.M., Alvarez, L., Ortiz, P. and Pajares, M.A. S-adenosylmethionine synthesis: molecular mechanisms and clinical implications. Pharmacol. Ther., 1997;73(3):265-280. 4. Mudd, S.H. and Cantoni, G.L. Activation of methionine for transmethylation. III. The methionine-activating enzyme of Bakers′ yeast. J. Biol.Chem., 1958;231:481-492. 5. Markham, G.D., DeParasis, J. and Gatmaitan, J. The sequence of metK, the structural gene for S-adenosylmethionine synthetase in Escherichia coli. J. Biol. Chem., 1984;259(23):14505-14507. 6. Thomas, D. and Surdin-Kerjan, Y.. SAM1, the structural gene for one of the. S-adenosylmethionine synthetases in Saccharomyces cerevisiae. Sequence and expression. J. Biol. Chem., 1987;262(34):16704-16709. 47.
(58) 7. Peleman, J., Saito, K., Cottyn, B., Engler, G., Seurinck, J., Van Montagu, M. and Inze, D.. Structure and expression analyses of the S-adenosyl methionine. synthetase gene family in Arabidopsis thaliana. Gene, 1989;84(2):359-369. 8. Peleman, J., Boerjan, W., Engler, G., Seurinck, J., Botterman, J., Alliotte, T., Van Montagu, M. and Inze, D. of. a. housekeeping. Strong cellular preference in the expression. gene. of. Arabidopsis. thaliana. encoding. S-adenosylmethionine synthetase. Plant. Cell., 1989;1(1):81-93. 9. Alvarez, L., Corrales, F., Martin-Duce, A. and Mato, J.M. Characterization of a full-length cDNA encoding human liver S-adenosylmethionine synthetase: tissue-specific gene expression and mRNA levels in hepatopathies. Biochem. J., 1993;293 (Pt 2):481-486. 10. Horikawa, S. and Tsukada, K.. Molecular cloning and developmental. expression of a human kidney S-adenosylmethionine synthetase. FEBS Lett., 1992;312(1):37-41. 11. Horikawa, S., Ishikawa, M., Ozasa, H. and Tsukada, K.. Isolation of a. cDNA encoding the rat liver S-adenosylmethionine synthetase. Eur. J. Biochem., 1989;184(3):497-501.. 48.
(59) 12. Horikawa, S., Sasuga, J., Shimizu, K., Ozasa, H. and Tsukada, K. Molecular cloning and nucleotide sequence of cDNA encoding the rat kidney S-adenosylmethionine synthetase. J. Biol. Chem., 1990;265(23):13683-13686. 13. Horikawa, S. and Tsukada, K.. Molecular cloning and nucleotide sequence. of cDNA encoding the human liver S-adenosylmethionine synthetase. Biochem. Int., 1991;25(1):81-90. 14. Sakata, S.F., Shelly, L.L., Ruppert, S., Schutz, G. and Chou, J.Y. Cloning and expression of murine S-adenosylmethionine synthetase. J. Biol. Chem., 1993;268(19):13978-13986. 15. De La Rosa, J., Ostrowski, J., Hryniewicz, M.M., Kredich, N.M., Kotb, M., LeGros, H.L. Jr., Valentine, M. and Geller, A.M. Chromosomal localization and catalytic properties of the recombinant alpha subunit of human lymphocyte. methionine. adenosyltransferase.. J.. Biol.. Chem.,. 1995;270(37):21860-21868. 16. Kotb, M., Mudd, S.H., Mato, J.M., Geller, A.M., Kredich, N.M., Chou, J.Y. and Cantoni, G.I. Consensus nomenclature for the mammalian methionine adenosyltransferase. genes. and. gene. products.. Trends. Genet.,. 1997;13(2):51-52.. 49.
(60) 17. van Faassen, H. and Berger, R.. Separation of different forms of. S-adenosylmethionine synthetase by fast protein liquid chromatography. J. Biochem. Biophys. Methods., 1990 Mar;20(3):189-194. 18. Gil, B., Casado, M., Pajares, M.A., Bosca, L., Mato, J.M., Martin-Sanz, P. and Alvarez, L. Differential expression pattern of S-adenosylmethionine synthetase isoenzymes during rat liver development. Hepatology, 1996 Oct;24(4):876-881. 19. Alvarez, L., Mingorance, J., Pajares, M.A. and Mato, J.M. Expression of rat liver S-adenosylmethionine synthetase in Escherichia coli. results in two active oligomeric forms. Biochem. J., 1994;301 ( Pt 2):557-561. 20. Cabrero, C., Puerta, J. and Alemany, S. Purification and comparison of two forms of S-adenosyl-L-methionine synthetase from rat liver. Eur. J. Biochem., 1987;170(1-2):299-304. 21. Takusagawa, F., Kamitori, S. and Markham, G.D. Structure and function of S-adenosylmethionine synthetase: crystal structures of S-adenosylmethionine synthetase with ADP, BrADP, and PPi at 2.8 angstroms resolution. Biochemistry, 1996;35(8):2586-2596. 22. Mingorance, J., Alvarez, L., Pajares, M.A. and Mato, J.M. Recombinant rat. 50.
(61) liver S-adenosyl-L-methionine synthetase tetramers and dimers are in equilibrium. Int. J. Biochem. Cell Biol., 1997;29(3):485-491. 23. Hoffman, J.L. Fractionation of methionine adenosyltransferase isozymes (rat liver). Methods Enzymol., 1983;94:223-228. 24. Sullivan, D.M. and Hoffman, J.L. Fractionation and kinetic properties of rat liver and kidney methionine adenosyltransferase isozymes. Biochemistry, 1983;22(7):1636-1641. 25. Suma, Y., Shimizu, K. and Tsukada, K. Isozymes of S-adenosylmethionine synthetase from rat liver: isolation and characterization. J. Biochem. (Tokyo), 1986;100(1):67-75. 26. Chamberlin, M.E., Ubagai, T., Pao, V.Y., Pearlstein, R.A. and Yang Chou, J. Structural requirements for catalysis and dimerization of human methionine adenosyltransferase I/III. Arch. Biochem. Biophys., 2000;373(1):56-62. 27. Kotb, M. and Kredich, N.M. S-Adenosylmethionine synthetase from human lymphocytes.. Purification. and. characterization.. J.. Biol.. Chem.,. 1985;260(7):3923-3930. 28. Halim, A.B., LeGros, L., Geller, A. and Kotb, M. Expression and functional interaction of the catalytic and regulatory subunits of human methionine. 51.
(62) adenosyltransferase. in. mammalian. cells.. J.. Biol.. Chem.,. 1999;274(42):29720-29725. 29. Wabiko, H., Ochi, K., Nguyen, D.M., Allen, E.R. and Freese, E. Genetic mapping and physiological consequences of metE mutations of Bacillus subtilis. J. Bacteriol., 1988;170(6):2705-2710. 30. Geller, A.M., Legros, H.L., Wherry, K. and Kotb, M.Y. Inhibition of methionine adenosyltransferase by the polyamines. Arch. Biochem. Biophys., 1997;345(1):97-102. 31. Latasa, M.U., Boukaba, A., Garcia-Trevijano, E.R., Torres, L., Rodriguez, J.L., Caballeria, J., Lu, S.C., Lopez-Rodas, G., Franco, L., Mato, J.M. and Avila, M.A. Hepatocyte growth factor induces MAT2A expression and histone acetylation in rat hepatocytes: role in liver regeneration. FASEB J., 2001;15(7):1248-1250. 32. Torre, L., Lopez-Rodas, G., Latasa, M.U., Carretero, M.V., Boukaba, A., Rodriguez, J.L., Franco, L., Mato, J.M. and Avila, M.A. DNA methylation and histone acetylation of rat methionine adenosyltransferase 1A and 2A genes is tissue-specific. Int. J. Biochem. Cell Biol., 2000;32(4):397-404. 33. Gil, B., Pajares, M.A., Mato, J.M. and Alvarez, L. Glucocorticoid regulation. 52.
(63) of hepatic S-adenosylmethionine synthetase gene expression. Endocrinology, 1997;138(3):1251-1258. 34. Castro, C., Ruiz, F.A., Perez-Mato, I., Sanchez del Pino, M.M., LeGros, L., Geller, A.M., Kotb, M., Corrales, F.J. and Mato, J.M. Creation of a functional S-nitrosylation site in vitro by single point mutation. FEBS Lett., 1999;459(3):319-322. 35. Perez-Mato, I., Castro, C., Ruiz, F.A., Corrales, F.J. and Mato, J.M. Methionine adenosyltransferase S-nitrosylation is regulated by the basic and acidic amino acids surrounding the target thiol. J. Biol. Chem., 1999;274(24):17075-17079. 36. Ruiz, F., Corrales, F.J., Miqueo, C. and Mato, J.M. Nitric oxide inactivates rat hepatic methionine adenosyltransferase In vivo by S-nitrosylation. Hepatology, 1998;28(4):1051-1057. 37. Corrales, F.J., Ruiz, F. and Mato, J.M. In vivo regulation by glutathione of methionine adenosyltransferase S-nitrosylation in rat liver. J. Hepatol., 1999;31(5):887-894. 38. Gomes Trolin, C., Ekblom, J. and Oreland, L. Regulation of methionine adenosyltransferase catalytic activity and messenger RNA in SH-SY5Y. 53.
(64) human neuroblastoma cells. Biochem. Pharmacol., 1998;55(5):567-571. 39. Fu, Z., Hu, Y., Markham, G.D. and Takusagawa, F.. Flexible loop in the. structure of S-adenosylmethionine synthetase crystallized in the tetragonal modification. J. Biomol. Struct. Dyn., 1996;13(5):727-739. 40. Cai, J., Sun, W.M., Hwang, J.J., Stain, S.C. and Lu, S.C. S-adenosylmethionine. synthetase. in. human. liver. cancer:. Changes in molecular. characterization and significance. Hepatology, 1996;24(5):1090-1097. 41. Chiang, P.K., Gordon, R.K., Tal, J., Zeng, G.C., Doctor, B.P., Pardhasaradhi, K. and McCann, P.P. S-adenosylmethionine and methylation. FASEB J., 1996;10:471-480. 42. Tajima, S. and Suetake, I. Regulation and function of DNA methylation in vertebrates. Biochem. (Tokyo), 1998;123(6):993-999. 43. Schmutte, C. and Jones, P.A. Involvement of DNA methylation in human carcinogenesis. Biol. Chem., 1998;379(4-5):377-388. 44. Zingg, J.M. and Jones, P.A. Genetic and epigenetic aspects of DNA methylation on genome expression, evolution, mutation and carcinogenesis. Carcinogenesis, 1997;18(5):869-882. 45. Momparler, R.L., Cote, S. and Eliopoulos, N.. Pharmacological approach for 54.
(65) optimization of the dose schedule of 5-aza-2′-deoxycytidine (Decitabine) for the therapy of leukemia. Leukemia, 1997;11(2):175-180. 46. Tutt Landolfi, M.M., Scollay, R. and Parnes, J.R. Specific demethylation of the CD4 gene during CD4 T lymphocyte differentiation. Mol. Immunol., 1997;34(1):53-61. 47. Bender Christina, M., Zingg Jean-Marc and Jones Peter, A. methylation as a target for drug design.. DNA. Pharmaceutical Research,. 1998;15(2):175-187. 48. Potter, V.R., Phenotypic diversity in experimental hepatoma concept of partially blocked ontogeny. Br. J. Cancer., 1978;38:1. 49. Liau, M.C., Chang, C.F. and Giovanella, B.C. Demonstration of an altered S-adenosylmethionine synthetase in human malignant tumors xenografted into athymic nude mice. J. Natl. Cancer Inst., 1980;64:1071. 50. Liau,. M.C.,. Chang,. C.F.,. Saunders,. G.F.. and. Tsai,. Y.H.. S-adenosylhomocysteine hydrolases as the primary target enzymes in androgen regulation of methylation complexes. Arch. Biochem. Biophys., 1981;208:261. 51. Liau, M.C. and Burzynski, S.R. Altered methylation complex isozymes as. 55.
(66) selective targets for cancer chemotherapy. Drugs Exp. Clin. Res., 1986;12 Suppl 1:77-86. 52. Liau, M.C., Hunt, M.E. and Hurlbert, R.B. Role of ribosomal RNA methylases in the regulation of ribosome production. Biochem., 1976, 15:3158. 53. Hanada, M., Delia, D., Aiello, A., Stadtmauer, E. and Reed, J.C. bcl-2 gene hypomethylation and high-level expression in B-cell chronic lymphocytic leukemia. Blood, 1993;82(6):1820-1828. 54. Crossen, P.E. and Morrison, M.J. Hypermethylation of the M27beta (DXS255) locus in chronic B-cell leukaemia. Br. J. Haematol., 1998;100(1):191-193. 55. Iravani, M., Dhat, R. and Price, C.M. Methylation of the multi tumor suppressor gene-2 (MTS2, CDKN1, p15INK4B) in childhood acute lymphoblastic leukemia. Oncogene, 1997;15(21):2609-2614. 56. Eguchi,. K.,. Kanai,. Y.,. Kobayashi,. K.. and. Hirohashi,. S.. DNA. hypermethylation at the D17S5 locus in non-small cell lung cancers: its association with smoking history. Cancer Res., 1997;57(21):4913-4915. 57. Wicki, R., Franz, C., Scholl, F.A., Heizmann, C.W. and Schafer, B.W. Repression of the candidate tumor suppressor gene S100A2 in breast cancer is. 56.
(67) mediated. by. site-specific. hypermethylation.. Cell. Calcium,. 1997;22(4):243-254. 58. Martin, V., Ribieras, S., Song-Wang, X., Rio, M.C. and Dante, R. Genomic sequencing indicates a correlation between DNA hypomethylation in the 5′ region of the pS2 gene and its expression in human breast cancer cell lines. Gene, 1995;157(1-2):261-264. 59. Makos, M., Nelkin, B.D., Lerman, M.I., Latif, F., Zbar, B. and Baylin, S.B. Distinct hypermethylation patterns occur at altered chromosome loci in human. lung. and. colon. cancer.. Proc.. Natl.. Acad.. Sci.. USA,. 1992;89(5):1929-1933 60. Huang, Z.Z., Mao, Z., Cai, J. and Lu, S.C. Changes in methionine adenosyltransferase during liver regeneration in the rat. Am. J. Physiol., 1998;275(1 Pt 1):G14-G21. 61. Cai, J., Mao, Z., Hwang, J.J. and Lu, S.C. Differential expression of methionine adenosyltransferase genes influences the rate of growth of human hepatocellularcarcinoma cells. Cancer Res., 1998;58(7):1444-1450. 62. Frago, L.M., Gimenez, A., Rodriguez, E.N. and Varela-Nieto, I. Pattern of methionine adenosyltransferase isoenzyme expression during rat liver. 57.
(68) regeneration after partial hepatectomy. FEBS Lett., 1998;426(3):305-308. 63. Newman, E.B., Budman, L.I., Chan, E.C., Greene, R.C., Lin, R.T., Woldringh, C.L. and D′Ari, R.. Lack of S-adenosylmethionine results in a cell division. defect in Escherichia coli. J. Bacteriol., 1998;180(14):3614-3619. 64. Chamberlin, M.E., Ubagai, T., Mudd, S.H., Levy, H.L. and Chou, J.Y. Dominant inheritance of isolated hypermethioninemia is associated with a mutation in the human methionine adenosyltransferase 1A gene. Am. J. Hum. Genet., 1997;60(3):540-546. 65. Nagao, M. and Oyanagi, K.. Genetic analysis of isolated persistent. hypermethioninemia with dominant inheritance. Acta. Paediatr. Jpn., 1997;39(5):601-606. 66. Mato, I.P., Sanchez del Pino, M.M., Chamberlin, M.E., Mudd, S.H., Mato, J.M. and Corrales, F.J. Biochemical basis for the dominant inheritance of hypermethioninemia associated with the R264H mutation of the MAT1A gene. A monomeric methionine adenosyltransferase with tripolyphosphatase activity. J. Biol. Chem., 2001;276(17):13803-13809. 67. Surtees, R., Leonard, J. and Austin, S. Association of demyelination with deficiency of cerebrospinal-fluid S-adenosylmethionine in inborn errors of. 58.
(69) methyl-transfer pathway. Lancet., 1991;338 (8782-8783):1550-1554. 68. Chamberlin, M.E., Ubagai, T., Mudd, S.H., Wilson, W.G., Leonard, J.V. and Chou, JY. Demyelination of the brain is associated with methionine adenosyltransferase I/III deficiency. J. Clin. Invest., 1996;98(4):1021-1027. 69. Snyder, S.W., Egorin, M.J. and Callery, P.S. Induction of human leukemia cell differentiation by regiospecifically acetylated spermidines. Biochem. Biophys. Res. Commun., 1991;180(2):591-596. 70. Bjersing, J.L., Brorsson, A. and Heby, O. Increased expression of c-jun, but not retinoic acid receptor beta, is associated with F9 teratocarcinoma stem cell differentiation induced by polyamine depletion. J. Cell. Biochem., 1997;67(3):378-385. 71. Luk, G.D., Civin, C.I., Weissman, R.M. and Baylin, S.B. Ornithine decarboxylase: essential in proliferation but not differentiation of human promyelocytic leukemia cells. Science, 1982;216(4541):75-77. 72. Sugiura, M., Shafman, T., Mitchell, T., Griffin, J. and Kufe, D. Involvement of spermidine in proliferation and differentiation of human promyelocytic leukemia cells. Blood, 1984;63(5):1153-1158. 73. Riscoe, M.K., Schwamborn, J., Ferro, A.J., Olson, K.D. and Fitchen, J.H.. 59.
(70) Inhibition of growth but not differentiation of normal and leukemic myeloid cells by methylthioadenosine. Cancer Res., 1987;47(14):3830-3834. 74. Sydow, K. and Boger, R.H. Homocysteine, endothelial dysfunction and cardiovascular risk: pathomechanisms and therapeutic options.. Z. Kardiol.,. 2001;90(1):1-11. 75. Kaletha, K., Chodorowski, Z., Sein-Anand, J., Gazda, M. and NagelStarczynowska, G. Homocysteine as a risk factor for atherosclerosis. Przegl. Lek., 2000;57(10):591-595. 76. Lu, S.C. S-Adenosylmethionine. The International Journal of Biochemistry & Cell Biology., 2000;32(4):391-395. 77. Bottiglieri, T., and Hyland, K. S-adenosylmethionine levels in psychiatric and neurological disorders: a review. Acta. Neurol. Scand., 1994;154:19-26. 78. Fava, M., Giannelli, A., Rapisarda, V., Patralia, A. and Guaraldi, G.P. Rapidity. of. onset. of. the. antidepressant. effect. of. parenteral. S-adenosyl-L-methionine. Psychiatry Res., 1995;56(3):295-297. 79. Bottiglieri, T., Hyland, K. and Reynolds, E.H. The clinical potential of ademetionine(S-adenosylmethionine). in. neurological. disorders.. Drugs,. 1994;48(2):137-152.. 60.
(71) 80. Bressa,. G.M.. S-adenosyl-l-methionine. (SAMe). as. antidepressant:. meta-analysis of clinical studies. Acta. Neurol. Scand. Suppl., 1994;154:7-14. 81. Bottiglieri, T., Godfrey, P., Flynn, T., Carney, M.W., Toone, B.K. and Reynolds, E.H. Cerebrospinal fluid S-adenosylmethionine in depression and dementia: effects of treatment with parenteral and oral S-adenosylmethionine. J. Neurol .Neurosurg. Psychiatry, 1990;53(12):1096-1098. 82. Schumacher, H.R. Jr. Osteoarthritis: the clinical picture, pathogenesis, and management with studies on a new therapeutic agent, S-adenosylmethionine. Am. J. Med., 1987;83(5A):1-4. 83. Liau, M.C., Chang, C.F. and Becker, F.F. Alteration of S-adenosylmethionine synthetasesduring chemical hepatocarcinogenesis and in resulting carcinomas. Cancer Res., 1979;39(6 Pt 1):2113-2119. 84. Jennifer, Lagendijk, Johan, B., Ubbink, W.J. and Hayward, Vermaak Quantification of erythrocyte S-adenosyl-L-methionine levels and its application in enzyme studies.. J. Chromatogr., 1992;576:95-101 Biomedical. Applications. 85. Carolyn K. Wise, Craig A. Gooney, Syed F. Ali and Lionel A. Poirier Measuring S-adenosylmethionine in whole blood, red blood cells and cultured. 61.
(72) cells using-a fast preparation method and high performance liquid chromatography.. J. Chromatogr. B, 1997;696:145-152.. 86. Capdevila, A. and Wagner, C. Measurement of plasma S-adenosylmethionine and S-adenosylhomocysteine as their fluorescent isoindoles. Anal. Biochem., 1998;264(2):180-184. 87. She, Q.B., Nagao, I., Hayakawa, T. and Tsuge, H. A simple HPLC method for the determination of S-adenosylmethionine and S-adenosyl- homocysteine in rat tissues: the effect of vitamin B6 deficiency on these concentrations in rat liver. Biochem. Biophys. Res. Commun., 1994;205(3):1748-1754 88. Hamedani, M.P., Valko, K., Qi, X., Welham, K.J. and Gibbons, W.A. Two-dimensional high-performance liquid chromatographic method for assaying S-adenosyl-L-methionine and its related metabolites in tissues. J Chromatogr., 1993;619(2):191-198. 89. Valko, K., Hamedani, M.P., Ascah, T.L. and Gibbons, W.A. A comparative study. of. the. reversed-phase. HPLC. retention. behaviour. of. S-adenosyl-L-methionine and its related metabolites on Hypersil ODS and Supelcosil. LC-ABZ. stationary. phases.. J.. Pharm.. Biomed.. Anal.,. 1993;11(4-5):361-366.. 62.
(73) 90. Bottiglieri, T.. Isocratic high performance liquid chromatographic analysis of. S-adenosylmethionine and S-adenosylhomocysteine in animal tissues: the effect of exposure to nitrous oxide. Biomed. Chromatogr., 1990;4(6):239-241. 91. Wagner, J., Claverie, N. and Danzin, C. A rapid high-performance liquid chromatographic procedure for the simultaneous determination of methionine, ethionine, S-adenosylmethionine, S-adenosylethionine, and the natural polyamines in rat tissues. Anal. Biochem., 1984;140(1):108-116. 92. Wagner, J., Danzin, C., Huot-Olivier, S., Claverie, N. and Palfreyman, M.G. High-performance liquid chromatographic analysis of S-adenosylmethionine and its metabolites in rat tissues: interrelationship with changes in biogenic catechol. levels. following. treatment. with. L-dopa.. J.. Chromatogr.,. 1984;290:247-262. 93. Gutheil, W.G., Stefanova, M.E. and Nicholas, R.A. Fluorescent coupled enzyme assays for D-alanine: application to penicillin-binding protein and vancomycin activity assays. Anal. Biochem., 2000;287(2):196-202. 94. Yasmineh, W.G., Chung, M.Y. and Caspers, JI. Determination of serum catalase activity on a centrifugal analyzer by an NADP/NADPH coupled enzyme reaction system. Clin. Biochem., 1992;25(1):21-27.. 63.
(74) 95. Alon, R., Bayer, E.A. and Wilchek, M. Related Articles. A coupled enzyme. assay for measurement of sialidase activity. J. Biochem. Biophys. Methods., 1991;22(1):23-33. 96. Van Lente, F. and Pepoy, M. Coupled-enzyme determination of catalase activity in erythrocytes. Clin. Chem., 1990;36(7):1339-1343. 97. Hirashima, Y., Farooqui, A.A. and Horrocks, L.A. Related Articles Fluorimetric coupled enzyme assay for lysoplasmalogenase activity in liver. Biochem. J., 1989;260(2):605-608. 98. Tsai, S.F., Bishop, D.F. and Desnick, R.J. Coupled-enzyme and direct assays for uroporphyrinogen III synthase activity in human erythrocytes and cultured lymphoblasts. Enzymatic diagnosis of heterozygotes and homozygotes with congenital erythropoietic porphyria. Anal. Biochem., 1987;166(1):120-133. 99. Teruel, J.A., Tudela, J., Fernandez-Belda, F., Garcia-Carmona, F., Garcia-Canovas, F. and Gomez-Fernandez, J.C. A kinetic study of the irreversible inhibition of an enzyme measured in the presence of coupled enzymes.. Fluorescein. isothiocyanate. as. inhibitor. of. the. adenosinetriphosphatase activity from sarcoplasmic reticulum. Biochim.. 64.
(75) Biophys. Acta., 1986;869(1):8-15. 100. Ozer, N.. A new enzyme-coupled spectrophotometric method for the. determination of arginase activity. Biochem. Med., 1985;33(3):367-371. 101. Wilkinson, A.P., Rhodes, M.J. and Fenwick, G.R. Determination of myrosinase (thioglucoside glucohydrolase) activity by a spectrophotometric coupled enzyme assay. Anal. Biochem., 1984;139(2):284-291. 102. Giampietro, P.F. and Desnick, R.J. Determination of delta-aminolevulinate dehydratase activity by a specific fluorometric coupled-enzyme assay. Anal. Biochem., 1983;131(1):83-92. 103. Nohta, H., Noma, S. and Ohkura, Y. Assay for Catechol-O- methyltransferase in erythrocytes using a new fluorogenic substrate, 2-(3,4-Dihydroxyphenyl) naphtho[1,2-d]thiazole. J. Chromatogr., 1984; 304:93-100. 104. Nohta, H., Noma, S. and Ohkura, Y. High-performance liquid chromatographic assay for Catechol-O-methyltransferase in human and rat erythrocyte membrane and soluble fractions, and rat tissues. Chem. Pharm. bull., 1986;34(11):4687-4693. 105. Muller-Enoch, D., Seidl, E. and Thomas, H. [6.7-Dihydroxycoumarin (Aesculetin) as a substrate for catechol-O-methyltransferase (author′s transl)].. 65.
(76) Z. Naturforsch. [C], 1976;31(5-6):280-284. 106. Thomas, H., Veser, J. and Muller-Enoch, D. [A sensitive fluorometric determination of catechol methyltransferase activity (author′s transl)]. Hoppe. Seylers. Z. Physiol. Chem., 1976;357(10):1347-1351. 107. Hoffman, R.M. Methioninase: a therapeutic for diseases related to altered methionine metabolism and transmethylation: cancer, heart disease, obesity, aging, and Parkinson′s disease. Hum. Cell, 1997;10(1):69-80. 108. Cheng, H., Gomes-Trolin, C., Aquilonius, S.M., Steinberg, A., Lofberg, C., Ekblom, J. and Oreland, L. Levels of L-methionine S-adenosyltransferase activity in erythrocytes and concentrations of S-adenosylmethionine and S-adenosylhomocysteine in whole blood of patients with Parkinson′s disease. Exp. Neurol., 1997;145(2 Pt 1):580-585. 109. Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 1976;72,248-254. 110. Trolin, C.G., Lofberg, C., Trolin, G. and Oreland, L. Brain ATP:L-methionine S-adenosyltransferase (MAT), S-adenosylmethionine (AdoMet) and S-adenosylhomocysteine (SAH): regional distribution and age-related changes.. 66.
(77) Eur. Neuropsychopharmacol, 1994;4(4):469-477. 111. Ekegren, T., Aquilonius, S.M. and Gomes-Trolin, C. A comparative study of methionine adenosyltransferase activity and regional distribution in mammalian spinal cord. Biochem. Pharmacol., 2000;60(3):441-445. 112. Tu, Y.S., Huang, L.J., Hsu, M.H., Wang, S.H., Lai, J.M., Yeh, C.C., Liau, M.C. and Lung, F.D. Synthesis of hemoregulatory peptide (HP) derivatives as inhibitors of the interaction between methionine adenosyl transferase and methionine. The Chin. Pharm. J., 2000;52:139-149. 113. Gallagher, R., Collins, S., Trujillo, J., McCredie, K., Ahearn, M., Tsai, S., Metzgar, R., Aulakh, G., Ting, R., Ruscetti, F., and Gallo, R. Characterization of the continuous, differentiating myeloid cell line (HL-60) from a patient with acute promyelocytic leukemia. Blood, 1979; 54:713-733. 114. Liau, M. C., Lin, G. W., and Hurlbert, R. B. Partial purification and characterization of tumor and liver S-adenosylmethionine synthetases. Cancer Res., 1977;37:427-435. 115. Hoffman, J. L. Chromatographic analysis of the chiral and covalent instability of S-adenosyl-L-methionine. Biochemistry, 1986;25:4444-4449. 116. Revelle, L. K., d′Avignon, D. A., Reepmeyer, J.C. and Zerfing, R. C.. 67.
(78) Stability-indicating proton nuclear magnetic resonance spectroscopic method for determination of S-adenosyl-L-methionine in tablets. J. AOAC Int., 1995;78:353-358.. 68.
(79) 中國醫藥大學 藥物化學研究所 博士論文 (二) 指 導 教 授 : 郭盛助 指 導 教 授 : 陳勝智. 教 授 教 授. HotLig: 以分子表面導向之方法評估蛋白質-配體之交互作用. HotLig: A molecular surface-directed approach to scoring protein-ligand interactions. 研究生: 王. 勝. 弘. Wang, Sheng-Hung. 中. 華. 民. 國. 95 年. 6 月.
數據

![圖 3. NO 與 GSH 對 MAT 的調節作用 [37]。](https://thumb-ap.123doks.com/thumbv2/9libinfo/8930056.267244/15.892.275.602.473.783/圖3NO與GSH對MAT的調節作用37.webp)
+7
![圖 4. Methionine 的代謝路徑 [41]。](https://thumb-ap.123doks.com/thumbv2/9libinfo/8930056.267244/18.892.162.743.248.874/圖4Methionine的代謝路徑41.webp)
相關文件
而在後續甲烷化反應試驗方面,以前段經厭氧醱酵產氫後之出流水為進流基 質。在厭氧光合產氫微生物方面,以光合作用產氫細菌中產氫能力最好的菌株 Rhodopseudomonas palustris
一般而言,物質的黏度與流體間的凝聚 力和分子間的動量轉移率有關。液體分子與
一般而言,物質的黏度與流體間的凝聚 力和分子間的動量轉移率有關。液體分子與
在上 一節中給出了有單位元的交換環 R 上的模的定義以及它的一些性質。 當環 R 為 體時, 模就是向量空間, 至於向量空間中的部分基本概念與定理, 有些可以移植到模上來。 例如 子
Rapiacta 因不經肝代謝,故透過 CYP 機轉與其他藥物發生 交互作用之可能性應該很低,就目前所知的排除途徑以及從 體外試驗可推知 Rapiacta 並不會誘導或抑制 CYP 450。 1)
是屬於層次較高之檢測工作。由此類工作的特質 是屬於層次較高之檢測工作。由此類工作的特質 來看,其應屬於非破壞評估(Non-Destructive
Ø 该类抑制剂与 COX-2 的共晶结构表明,甲磺酰基或氨磺 酰基可作用于 COX-2 通道上由缬氨酸
一、對抗生素Meropenem Trihydrate 具抗藥性(R 菌)和不具抗藥性(S 菌)的 Acinetobacter baumannii 菌在蛋白質電泳實驗中,比較蛋白質表現的差異。(文中所用代號: 「R
