1)行 政 院 農 業 委 員 會 林 業 試 驗 所 森 林 保 護 組 ,100台北市南海路53號 Division of Forest Protection, Taiwan Forestry Research Institute. 53 Nanhai Rd., Taipei 100, Taiwan.
2)國 立 台 灣 大 學 動 物 學 系 ,106台北市羅斯福路四段1號 Department of Zoology, National Taiwan University. 1, Roosevelt Rd., Sec. 4, Taipei, Taiwan 106.
3)目 前 地 址 : 國 立 台 灣 大 學 生 態 學 與 演 化 生 物 學 研 究 所 Present address: Institute of Ecology and Evolutionary Biology, National Taiwan University.
4)通訊作者 Corresponding author, e-mail:[email protected]
2003年1月送審 2003年7月通過 Received January 2003, Accepted July 2003.
台灣北部繁殖鳥類之海拔分布型態
葛兆年
1,4)李培芬
2,3)摘 要
本研究探討台灣北部陸域繁殖鳥類沿海拔梯度的變化型態。在台灣北部陸域海拔7.7至3,423 m的 101個樣點,調查所得之繁殖鳥類密度資料,以降趨對應分析(DCA)及群集分析結果,將台灣北部陸域 繁殖鳥類分為6個群聚。海拔梯度為決定群聚類型的重要因子。中海拔的兩群聚在鳥種數及多樣性指標 皆顯著大於高與低海拔群聚,顯示中海拔鳥類群聚的種類豐富度及多樣性較高。在涵括3,400 m的海拔 梯度上,鳥種豐富度在中海拔出現高峰,往高、低海拔逐漸下降。分布中點愈靠近海拔兩端的鳥種, 其海拔幅寬愈小;海拔幅寬較大的鳥種,其分布中點較靠近海拔梯度的中間帶,故支持以mid-domain effect來解釋台灣北部繁殖鳥類種豐富度的駝型分布。 關鍵詞:台灣、鳥類群聚、海拔梯度、種豐富度、多樣性。 葛兆年、李培芬。2003。台灣北部繁殖鳥類之海拔分布型態。台灣林業科學18(4):349-61。 Research paperElevational Gradients in Breeding Birds in Northern Taiwan
Chao-Nien Koh,
1,3)Pei-Fen Lee
2)【
Summary】
Patterns of breeding birds along an elevational gradient were investigated in northern Taiwan.
According to DCA and cluster analysis, 101 sampling sites of breeding birds were clustered into
6 communities from 7.7 to 3,423 m of elevation. The elevational gradient is an important factor in
determining the community types. Two mid-elevation communities had higher species number and
diversity index, reflecting that they have higher species richness and diversity than those of low or
high elevations. Along the 3,400- m elevational gradient, species richness peaked at intermediate
elevations and declined toward either high or low elevations. Species with midpoints closer to
high or low elevations had smaller elevational ranges, while species with correspondingly larger
elevational ranges had midpoints increasingly nearer the central portion of the elevational gradient.
The hump-shaped distribution of species richness along the elevational gradient, with a peak at
intermediate elevations, can be explained by the mid-domain effect.
Key words: Taiwan, bird community, elevational gradient, species richness, diversity.
Koh CN, Lee PF. 2003. Elevational gradients in breeding birds in northern Taiwan. Taiwan J For Sci
18(4):349-61.
緒 言
長期以來,生物的分布型態一直是生態學 和生物地理學的研究焦點,藉由觀察了解生物 的分布型態,進而找出其背後的成因與機制, 不僅具有學術研究意義,同時亦可做為經營管 理的基礎,以及預測未來環境變遷時對生物的 影響。 台灣因地理位置的特殊與地形的複雜,孕 育豐富的鳥類相。在五百多種鳥類當中,有一 百五十餘種為留鳥,這些留鳥大多棲息於森林 內,其分布因海拔而有所差異。Kano (1940) 首先描述雪山地區鳥類的海拔垂直分布及其棲 息環境;Jai (1977)台灣中部山區鳥類分為低海 拔、中海拔及高海拔帶;Shiu (1995)研究台灣 中海拔山區森林鳥類群聚與環境的關係,指出 海拔高度為影響鳥類群聚的主要因子;Huang (2001)研究北部陸域鳥類群聚與環境因子的關 係,亦認為影響鳥類空間分布最主要的因子為 海拔/溫度梯度。上述研究說明台灣陸域鳥類沿 海拔梯度確實可能有一定的分布型態。 近 年 來Brown (1995)提出巨觀生態學的 理論,以大量可靠的生物分布資料做為分析依 據,找出生物分布的特徵,並加以解釋,乃屬 於大尺度的研究方法。此方法可以突破以往傳 統野外實驗的尺度限制,從較大尺度找出以往 未發現的生態特質(Brown 1995, Blackburn and Gaston 1998)。台灣地區以往較缺乏涵蓋完整海 拔梯度的鳥類調查與研究(Huang 2001),因此 本研究採用大尺度的研究方法,來探討台灣北 部地區陸域繁殖鳥類在整個海拔帶上的分布型 態,包括鳥類群聚的分類及各群聚的總密度、 種豐富度、多樣性及均勻度。 在 講 求 生 物 多 樣 性 保 育 的 今 天 , 已 有 許 多生態學者研究不同分類的生物種豐富度沿海 拔梯度的分布,大致獲得兩種分布型態:一是 種豐富度隨海拔梯度上升呈線性遞減(Stevens 1992),另一是種豐富度沿海拔梯度上升呈駝型 分布(Colwell and Hurtt 1994, Rahbek 1997), 這兩種分布型態分別獲得許多不同的研究結果 支持。Stevens (1992)除了認為種豐富度有隨 海拔梯度上升呈線性遞減的型態,更進一步以 Rapoport’s altitudinal rule(以下簡稱Rapoport’s rule) 來解釋此型態背後的機制,Rapoport’s rule是指 物種的海拔幅寬與梯度有正相關的關係,低海 拔的物種因幅寬較窄,棲地間的物種補充效應 (rescue effect)較容易發生,因此低海拔棲地有 較高的種豐富度。而Colwell and Lees (2000)則 提出物種之分布幅寬對其分布中點有一個等邊 三角形的模式:假設緯度或海拔梯度有兩個限 制分布的邊界(南極與北極或海邊與山頂),在界 限之內,物種沿緯度或海拔梯度的分布位置及 分布範圍均為隨機事件時,在兩端邊界之物種 會有最小分布範圍(等邊三角形底邊兩端點);而 分布中點在地理中央之物種,則會有最大分布 範圍(等邊三角形頂點),所有物種的分布中點都 落在此等邊三角形中,因此分布中點靠近邊界 的物種會有較小分布範圍,物種間分布重疊較 小,相對而言,趨近中間地帶則有較大的物種 重疊,故種豐度必然在空間梯度的中間地帶達 到最高,此現象稱為mid-domain effect。本研究 乃利用台灣北部陸域繁殖鳥類的資料,探討物 種的豐富度沿海拔梯度有何種變化型態,其是否 可以物種的海拔幅寬來加以解釋。材 料 與 方 法
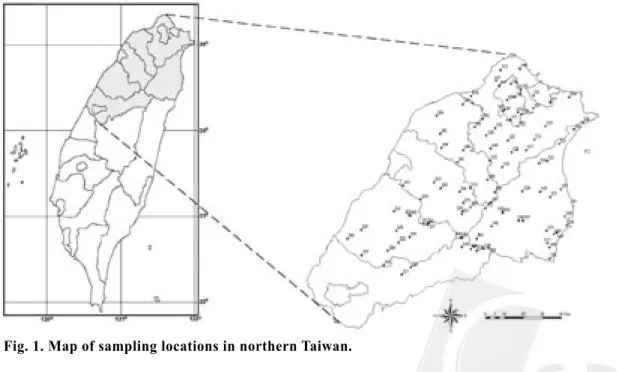
一、調查地區 本研究以台灣北部地區為調查範圍,涵蓋 北台灣六個縣市,包括:台北縣市、宜蘭縣、桃園縣、新竹縣、苗栗縣,整個研究區域海拔 跨幅超過3,000 m,其中3,000 m以上之高山均 位於本研究地區之南端,已由新竹、苗栗及宜 蘭等縣橫跨至台中縣北境,故有部份調查地點 延伸至台中縣北境(Fig. 1)。 二、調查樣點之選取設置 我 們 選 擇 鳥 類 調 查 樣 點 的 方 式 , 參 考 地 理資訊系統之生態分區(Su 1992)、海拔高度分 布、植被分布、道路交通等分布狀況,依照面 積比例,盡可能均勻選擇調查樣點,共選取101 個樣點(Fig. 1),於每一樣點設一穿越線,沿線 設置5-10個取樣站,取樣站之間至少相距150 m,共設置978個取樣站。 三、調查方法 於2001及2002年2月下旬至7月中旬之鳥類 繁殖季,採用不定半徑圓圈法(Reynolds et al. 1980),至各調查點調查鳥類族群密度。調查 只在天氣良好時進行,自日出至日出後3 h之間 完成。每一樣點調查一次,每一取樣站停留6 min,依序記錄目擊或聽到的鳥種、數量、距離 (小於或大於50 m)及取樣站的海拔高度。記錄 之鳥種以陸域性繁殖鳥類為主,外來種、過境 鳥、持續於空中飛行之鳥種則不予記錄。 四、資料分析 各 樣 點 之 鳥 類 族 群 密 度 以 下 列 公 式 計 算 (Reynolds et al. 1980): D = n*10000 ; π*r2*C*P 其中D:鳥類族群密度(隻數/ha),n:特定基礎 半 徑 內 所 記 錄 之 總 隻 數 ,r:特定基礎半徑, 在此半徑內大部分的鳥隻皆可察覺到,單位為 m,C:調查次數,p:取樣站數目。 密 度 計 算 時 , 平 均 各 取 樣 站 的 鳥 類 密 度 資料,得各樣點之鳥類族群密度,經平方根之 數值轉換,先去除記錄樣點少於三個之稀有鳥 種 (Shiu 1995),採用PC-ORD vers. 4 (McCune and Mefford 1999)進行降趨對應分析(detrended correspondence analysis, DCA) (Gauch 1982), 找出鳥類群聚的主要變異梯度,以及各樣點在 鳥類群聚之主要變異梯度軸上的分布序列值。
以各樣點之鳥類群聚密度為特徵值,進行 群集分析(cluster analysis) (Gauch 1982),利用 鳥類密度資料以relative Euclidean distance計算 各樣點間之相異度矩陣(Ludwig and Reynolds
1988),再以UPGMA (unweighted pair-group method using arithmetic averages)方法,連結各
樣點成一樹狀圖。將群集分析與DCA之結果綜
合比較,以決定鳥類群聚類型之分類。
對 各 樣 點 之DCA第一軸分布序列值及海
拔高度做迴歸分析;以變方分析及鄧肯氏多變 域測驗Duncan’s new multiple range test (Zar 1999)檢定以下各數值之差異:各群聚之DCA 第一軸分布序列值、海拔高度、總密度、鳥種
數、Shannon多樣性指數(H’)及Shannon均勻度
指數(E) (Magurran 1988, Krebs 1999)。計算公 式如下: H’ = ─
Σ
Pilog2Pi; E = H’ = H’ ; H max log2S 其 中S:各群聚中所記錄到之鳥種種數,Pi: 各群聚中第i種鳥種所佔的數量百分比,Hmax = log2S。結 果
2001及2002年兩年之繁殖季共調查101個 樣點(Table 1),刪除少於3個樣點的稀有鳥種 i=1 sTable 1. Elevations of 101 sampling sites in northern Taiwan and their geographical
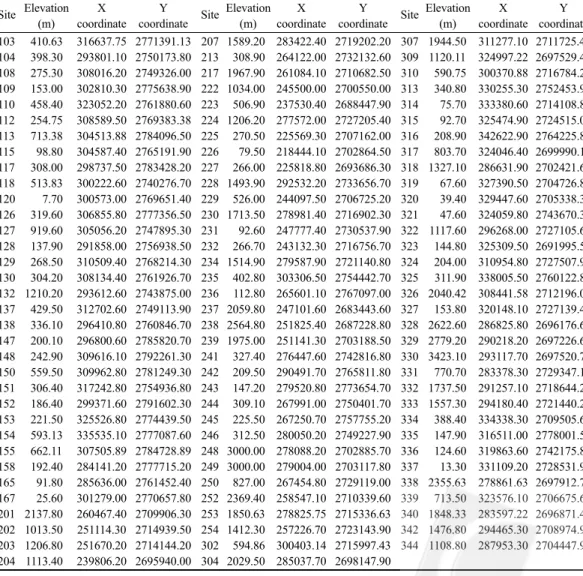
positions using the TM2 coordinate system
Site Elevation X Y Site Elevation X Y Site Elevation X Y (m) coordinate coordinate (m) coordinate coordinate (m) coordinate coordinate 103 410.63 316637.75 2771391.13 207 1589.20 283422.40 2719202.20 307 1944.50 311277.10 2711725.40 104 398.30 293801.10 2750173.80 213 308.90 264122.00 2732132.60 309 1120.11 324997.22 2697529.44 108 275.30 308016.20 2749326.00 217 1967.90 261084.10 2710682.50 310 590.75 300370.88 2716784.25 109 153.00 302810.30 2775638.90 222 1034.00 245500.00 2700550.00 313 340.80 330255.30 2752453.90 110 458.40 323052.20 2761880.60 223 506.90 237530.40 2688447.90 314 75.70 333380.60 2714108.80 112 254.75 308589.50 2769383.38 224 1206.20 277572.00 2727205.40 315 92.70 325474.90 2724515.00 113 713.38 304513.88 2784096.50 225 270.50 225569.30 2707162.00 316 208.90 342622.90 2764225.80 115 98.80 304587.40 2765191.90 226 79.50 218444.10 2702864.50 317 803.70 324046.40 2699990.10 117 308.00 298737.50 2783428.20 227 266.00 225818.80 2693686.30 318 1327.10 286631.90 2702421.60 118 513.83 300222.60 2740276.70 228 1493.90 292532.20 2733656.70 319 67.60 327390.50 2704726.80 120 7.70 300573.00 2769651.40 229 526.00 244097.50 2706725.20 320 39.40 329447.60 2705338.30 126 319.60 306855.80 2777356.50 230 1713.50 278981.40 2716902.30 321 47.60 324059.80 2743670.30 127 919.60 305056.20 2747895.30 231 92.60 247777.40 2730537.90 322 1117.60 296268.00 2727105.60 128 137.90 291858.00 2756938.50 232 266.70 243132.30 2716756.70 323 144.80 325309.50 2691995.50 129 268.50 310509.40 2768214.30 234 1514.90 279587.90 2721140.80 324 204.00 310954.80 2727507.90 130 304.20 308134.40 2761926.70 235 402.80 303306.50 2754442.70 325 311.90 338005.50 2760122.80 132 1210.20 293612.60 2743875.00 236 112.80 265601.10 2767097.00 326 2040.42 308441.58 2712196.08 137 429.50 312702.60 2749113.90 237 2059.80 247101.60 2683443.60 327 153.80 320148.10 2727139.40 138 336.10 296410.80 2760846.70 238 2564.80 251825.40 2687228.80 328 2622.60 286825.80 2696176.60 147 200.10 296800.60 2785820.70 239 1975.00 251141.30 2703188.50 329 2779.20 290218.20 2697226.60 148 242.90 309616.10 2792261.30 241 327.40 276447.60 2742816.80 330 3423.10 293117.70 2697520.70 150 559.50 309962.80 2781249.30 242 209.50 290491.70 2765811.80 331 770.70 283378.30 2729347.10 151 306.40 317242.80 2754936.80 243 147.20 279520.80 2773654.70 332 1737.50 291257.10 2718644.20 152 186.40 299371.60 2791602.30 244 309.10 267991.00 2750401.70 333 1557.30 294180.40 2721440.20 153 221.50 325526.80 2774439.50 245 225.50 267250.70 2757755.20 334 388.40 334338.30 2709505.60 154 593.13 335535.10 2777087.60 246 312.50 280050.20 2749227.90 335 147.90 316511.00 2778001.50 155 662.11 307505.89 2784728.89 248 3000.00 278088.20 2702885.70 336 124.60 319863.60 2742175.80 158 192.40 284141.20 2777715.20 249 3000.00 279004.00 2703117.80 337 13.30 331109.20 2728531.90 165 91.80 285636.00 2761452.40 250 827.00 267454.80 2729119.00 338 2355.63 278861.63 2697912.75 167 25.60 301279.00 2770657.80 252 2369.40 258547.10 2710339.60 339 713.50 323576.10 2706675.60 201 2137.80 260467.40 2709906.30 253 1850.63 278825.75 2715336.63 340 1848.33 283597.22 2696871.44 202 1013.50 251114.30 2714939.50 254 1412.30 257226.70 2723143.90 342 1476.80 294465.30 2708974.90 203 1206.80 251670.20 2714144.20 302 594.86 300403.14 2715997.43 344 1108.80 287953.30 2704447.90 204 1113.40 239806.20 2695940.00 304 2029.50 285037.70 2698147.90
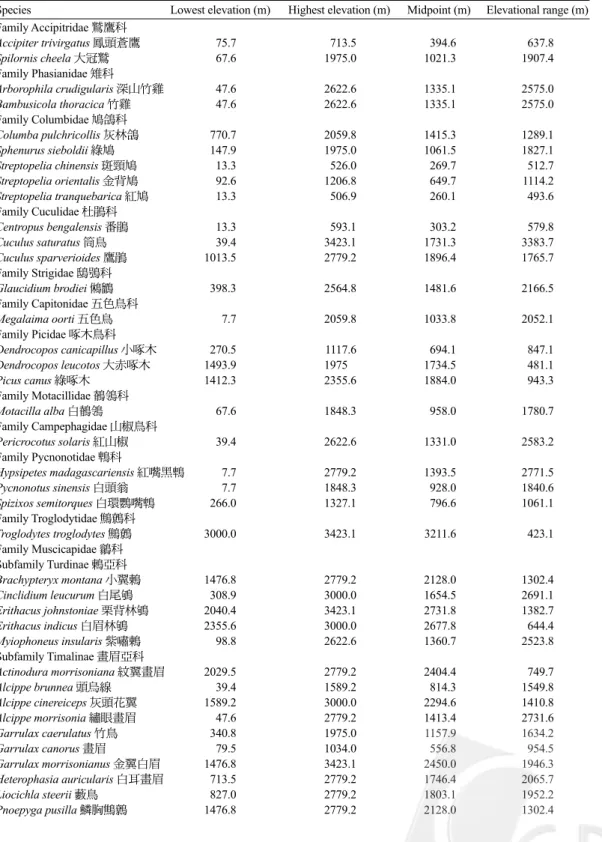
Table 2. Lowest elevations, highest elevations, midpoints, and elevational ranges of species
of terrestrial breeding birds surveyed in northern Taiwan from 2001 to 2002. Latin and
Chinese names of species follow Wang et al. (1991)
Species Lowest elevation (m) Highest elevation (m) Midpoint (m) Elevational range (m) Family Accipitridae 鷲鷹科 Accipiter trivirgatus 鳳頭蒼鷹 75.7 713.5 394.6 637.8 Spilornis cheela 大冠鷲 67.6 1975.0 1021.3 1907.4 Family Phasianidae 雉科 Arborophila crudigularis 深山竹雞 47.6 2622.6 1335.1 2575.0 Bambusicola thoracica 竹雞 47.6 2622.6 1335.1 2575.0 Family Columbidae 鳩鴿科 Columba pulchricollis 灰林鴿 770.7 2059.8 1415.3 1289.1 Sphenurus sieboldii 綠鳩 147.9 1975.0 1061.5 1827.1 Streptopelia chinensis 斑頸鳩 13.3 526.0 269.7 512.7 Streptopelia orientalis 金背鳩 92.6 1206.8 649.7 1114.2 Streptopelia tranquebarica 紅鳩 13.3 506.9 260.1 493.6 Family Cuculidae 杜鵑科 Centropus bengalensis 番鵑 13.3 593.1 303.2 579.8 Cuculus saturatus 筒鳥 39.4 3423.1 1731.3 3383.7 Cuculus sparverioides 鷹鵑 1013.5 2779.2 1896.4 1765.7 Family Strigidae 鴟鴞科 Glaucidium brodiei 鵂鶹 398.3 2564.8 1481.6 2166.5 Family Capitonidae 五色鳥科 Megalaima oorti 五色鳥 7.7 2059.8 1033.8 2052.1 Family Picidae 啄木鳥科 Dendrocopos canicapillus 小啄木 270.5 1117.6 694.1 847.1 Dendrocopos leucotos 大赤啄木 1493.9 1975 1734.5 481.1 Picus canus 綠啄木 1412.3 2355.6 1884.0 943.3 Family Motacillidae 鶺鴒科 Motacilla alba 白鶺鴒 67.6 1848.3 958.0 1780.7 Family Campephagidae 山椒鳥科 Pericrocotus solaris 紅山椒 39.4 2622.6 1331.0 2583.2 Family Pycnonotidae 鵯科 Hypsipetes madagascariensis 紅嘴黑鵯 7.7 2779.2 1393.5 2771.5 Pycnonotus sinensis 白頭翁 7.7 1848.3 928.0 1840.6 Spizixos semitorques 白環鸚嘴鵯 266.0 1327.1 796.6 1061.1 Family Troglodytidae 鷦鷯科 Troglodytes troglodytes 鷦鷯 3000.0 3423.1 3211.6 423.1 Family Muscicapidae 鶲科 Subfamily Turdinae 鶇亞科 Brachypteryx montana 小翼鶇 1476.8 2779.2 2128.0 1302.4 Cinclidium leucurum 白尾鴝 308.9 3000.0 1654.5 2691.1 Erithacus johnstoniae 栗背林鴝 2040.4 3423.1 2731.8 1382.7 Erithacus indicus 白眉林鴝 2355.6 3000.0 2677.8 644.4 Myiophoneus insularis 紫嘯鶇 98.8 2622.6 1360.7 2523.8 Subfamily Timalinae 畫眉亞科 Actinodura morrisoniana 紋翼畫眉 2029.5 2779.2 2404.4 749.7 Alcippe brunnea 頭烏線 39.4 1589.2 814.3 1549.8 Alcippe cinereiceps 灰頭花翼 1589.2 3000.0 2294.6 1410.8 Alcippe morrisonia 繡眼畫眉 47.6 2779.2 1413.4 2731.6 Garrulax caerulatus 竹鳥 340.8 1975.0 1157.9 1634.2 Garrulax canorus 畫眉 79.5 1034.0 556.8 954.5 Garrulax morrisonianus 金翼白眉 1476.8 3423.1 2450.0 1946.3 Heterophasia auricularis 白耳畫眉 713.5 2779.2 1746.4 2065.7 Liocichla steerii 藪鳥 827.0 2779.2 1803.1 1952.2 Pnoepyga pusilla 鱗胸鷦鷯 1476.8 2779.2 2128.0 1302.4
Continue Pomatorhinus erythrogenys 大彎嘴 47.6 1975.0 1011.3 1927.4 Pomatorhinus ruficollis 小彎嘴 13.3 2040.4 1026.9 2027.1 Stachyris ruficeps 山紅頭 39.4 2779.2 1409.3 2739.8 Yuhina brunneiceps 冠羽畫眉 803.7 2779.2 1791.5 1975.5 Yuhina zantholeuca 綠畫眉 47.6 1850.6 949.1 1803.0 Subfamily Paradoxornithinae 鸚嘴亞科 Paradoxornis webbianus 粉紅鸚嘴 67.6 1713.5 890.6 1645.9 Subfamily Sylviinae 鶯亞科 Abroscopus albogularis 棕面鶯 266.7 2622.6 1444.7 2355.9 Bradypterus seebohmi 褐色叢樹鶯 1327.1 3423.1 2375.1 2096.0 Cettia acanthizoides 深山鶯 1944.5 3423.1 2683.8 1478.6 Cettia fortipes 小鶯 308.9 2369.4 1339.2 2060.5 Cisticola exills 黃頭扇尾鶯 79.5 506.9 293.2 427.4 Cisticola juncidis 棕扇尾鶯 266.0 827.0 546.5 561.0 Prinia flaviventris 灰頭鷦鶯 13.3 1514.9 764.1 1501.6 Prinia polychroa 斑紋鷦鶯 79.5 1206.8 643.2 1127.3 Prinia subflava 褐頭鷦鶯 13.3 1713.5 863.4 1700.2 Regulus goodfellowi 火冠戴菊鳥 2137.8 3423.1 2780.5 1285.3 Subfamily Muscicapinae 鶲亞科 Ficedula hyperythra 黃胸青鶲 1117.6 2779.2 1948.4 1661.6 Hypothymis azurea 黑枕藍鶲 47.6 1476.8 762.2 1429.2 Muscicapa ferruginea 紅尾鶲 1034.0 3423.1 2228.6 2389.1 Niltava vivida 黃腹琉璃 306.4 2622.6 1464.5 2316.2 Family Aegithalidae 長尾山雀科 Aegithalos concinnus 紅頭山雀 1034.0 2779.2 1906.6 1745.2 Family Paridae 山雀科 Parus ater 煤山雀 1514.9 3423.1 2469.0 1908.2 Parus holsti 黃山雀 1034.0 1975.0 1504.5 941.0 Parus monticolus 青背山雀 1113.4 2779.2 1946.3 1665.8 Parus varius 赤腹山雀 312.5 1850.6 1081.6 1538.1 Family Sittidae 鳾科 Sitta europaea 茶腹鳾 1493.9 2779.2 2136.6 1285.3 Family Dicaeidae 啄花鳥科 Dicaeum concolor 綠啄花鳥 268.5 1975.0 1121.8 1706.5 Dicaeum ignipectus 紅胸啄花鳥 803.7 2779.2 1791.5 1975.5 Family Zosteropidae 繡眼科 Zosterops japonica 綠繡眼 7.7 1206.2 607.0 1198.5 Family Fringillidae 雀科 Carpodacus vinaceus 朱雀 2040.4 3423.1 2731.8 1382.7 Pyrrhula nipalensis 褐鷽 1013.5 2622.6 1818.1 1609.1 Family Ploceidae 文鳥科 Lonchura punctulata 斑文鳥 13.3 506.9 260.1 493.6 Lonchura striata 白腰文鳥 13.3 1589.2 801.3 1575.9 Passer montanus 麻雀 7.7 1327.1 667.4 1319.4 Family Oriolidae 黃鸝科 Oriolus traillii 朱鸝 144.8 594.9 369.8 450.1 Family Dicruridae 卷尾科 Dicrurus aeneus 小卷尾 39.4 1589.2 814.3 1549.8 Dicrurus macrocercus 大卷尾 13.3 309.1 161.2 295.8 Family Corvidae 鴉科 Corvus macrorhynchos 巨嘴鴉 67.6 3423.1 1745.4 3355.5 Dendrocitta formosae 樹鵲 7.7 1589.2 798.5 1581.5 Garrulus glandarius 松鴉 590.8 2779.2 1685.0 2188.4 Nucifraga caryocatactes 星鴉 2137.8 3423.1 2780.5 1285.3 Urocissa caerulea 台灣藍鵲 47.6 513.8 280.7 466.2
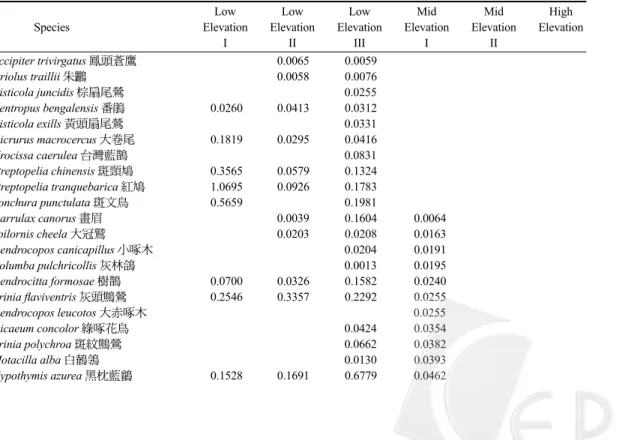
Table 3. Average density (no./ha) of each bird species, the score on the first axis of DCA,
elevation, and community parameters of each community in northern Taiwan
Low Low Low Mid Mid High
Species Elevation Elevation Elevation Elevation Elevation Elevation
I II III I II Accipiter trivirgatus 鳳頭蒼鷹 0.0065 0.0059 Oriolus traillii 朱鸝 0.0058 0.0076 Cisticola juncidis 棕扇尾鶯 0.0255 Centropus bengalensis 番鵑 0.0260 0.0413 0.0312 Cisticola exills 黃頭扇尾鶯 0.0331 Dicrurus macrocercus 大卷尾 0.1819 0.0295 0.0416 Urocissa caerulea 台灣藍鵲 0.0831 Streptopelia chinensis 斑頸鳩 0.3565 0.0579 0.1324 Streptopelia tranquebarica 紅鳩 1.0695 0.0926 0.1783 Lonchura punctulata 斑文鳥 0.5659 0.1981 Garrulax canorus 畫眉 0.0039 0.1604 0.0064 Spilornis cheela 大冠鷲 0.0203 0.0208 0.0163 Dendrocopos canicapillus 小啄木 0.0204 0.0191 Columba pulchricollis 灰林鴿 0.0013 0.0195 Dendrocitta formosae 樹鵲 0.0700 0.0326 0.1582 0.0240 Prinia flaviventris 灰頭鷦鶯 0.2546 0.3357 0.2292 0.0255 Dendrocopos leucotos 大赤啄木 0.0255 Dicaeum concolor 綠啄花鳥 0.0424 0.0354 Prinia polychroa 斑紋鷦鶯 0.0662 0.0382 Motacilla alba 白鶺鴒 0.0130 0.0393 Hypothymis azurea 黑枕藍鶲 0.1528 0.1691 0.6779 0.0462
Fig. 2. Dendrogram from a cluster analysis
based on bird densities of 101 sites,
utilizing relative Euclidean distances and
the unweighted pair-group method using
arithmetic averages.
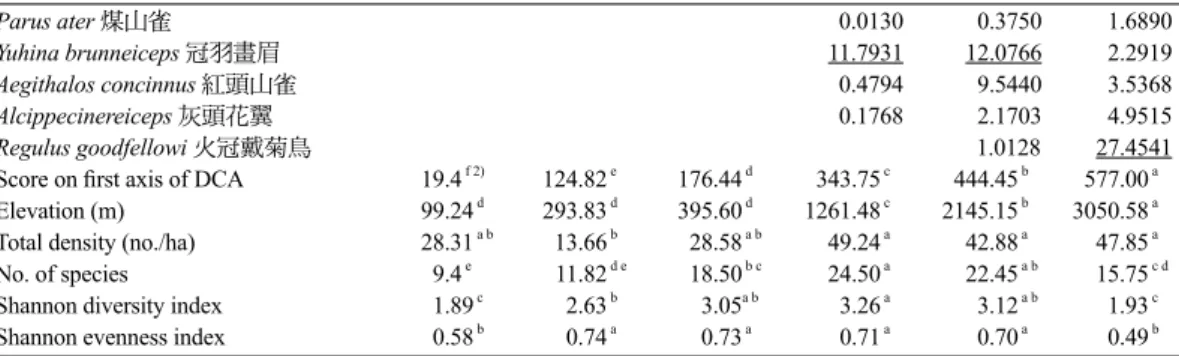
後,共計有22科80種鳥類(Table 2)。以所有樣 點之鳥類密度進行DCA運算,並配合群集分析 結果,將所有樣點分為6群聚(Fig. 2),以變方分 析檢定6個群聚之各樣點在DCA第一軸的分布序 列值(Table 3),6個群聚間確實有顯著差異,再 以鄧肯氏多變域測驗檢定兩兩群聚之差異,任 意兩群聚間皆有顯著差異。以各樣點的海拔高 度為自變項、各樣點在DCA第一軸的分布序列 值為應變項進行迴歸分析,海拔高度可解釋各 樣點在DCA第一軸的分布序列值之92.8%的變 異(p < 0.0001),顯示海拔梯度為影響鳥類群聚 變異之重要因子。對照海拔高度後,第1、2、 3群聚之海拔高度互有重疊,但均在1,000 m以 下(稱為低海拔I、II、III群聚),第4群聚之樣點 介於1,000-1,700 m間(稱為中海拔I群聚),第5群 聚之樣點介於1,700-2,700 m間(稱為中海拔II群 聚),第6群聚之海拔樣點均在2,700 m以上(稱為 高海拔群聚)。 6個群聚中各鳥種的平均密度如Table 3。各 群聚的海拔高度、總密度、鳥種數、多樣性指Continue Streptopelia orientalis 金背鳩 0.4210 0.0509 Spizixos semitorques 白環鸚嘴鵯 0.0637 0.0715 Pycnonotus sinensis 白頭翁 4.8330 3.9806 2.4754 0.1314 Pomatorhinus erythrogenys 大彎嘴 0.0188 0.1703 0.1318 Dicrurus aeneus 小卷尾 0.0591 0.2661 0.1667 Zosterops japonica 綠繡眼 5.5881 1.3585 2.1180 0.3183 Lonchura striata 白腰文鳥 0.8488 0.5517 0.5955 Alcippe brunnea 頭烏線 0.1981 1.5164 1.3469 Garrulax caerulatus 竹鳥 0.2806 1.7058 Passer montanus 麻雀 16.4108 1) 0.9646 0.7357 2.4758 Glaucidium brodiei 鵂鶹 0.0002 0.0045 0.0022 Megalaima oorti 五色鳥 0.1337 0.2564 0.3890 0.2739 0.0029 Sphenurus sieboldii 綠鳩 0.0079 0.0393 0.0058 Prinia subflava 褐頭鷦鶯 0.2801 0.3022 0.1706 0.0116 Picus canus 綠啄木 0.0076 0.0192 Arborophila crudigularis 深山竹雞 0.0116 0.0216 0.0942 0.0217 Bambusicola thoracica 竹雞 0.0825 0.4603 0.1631 0.0289 Parus holsti 黃山雀 0.1061 0.0347 Myiophoneus insularis 紫嘯鶇 0.0752 0.0365 0.0399 Pomatorhinus ruficollis 小彎嘴 0.2339 1.4186 1.8632 0.5110 0.0630 Yuhina zantholeuca 綠畫眉 0.0231 0.6140 0.1280 0.0752 Parus varius 赤腹山雀 0.0613 0.1140 0.0804 Pericrocotus solaris 紅山椒 0.0472 0.3086 0.3697 0.2805 Paradoxornis webbianus 粉紅鸚嘴 0.1286 0.5659 0.3858 Pyrrhula nipalensis 褐鷽 0.2555 0.3898 Abroscopus albogularis 棕面鶯 0.2164 1.7339 0.5633 Cettia fortipes 小鶯 0.1019 0.6140 0.7167 Niltava vivida 黃腹琉璃 0.0242 1.1378 0.8874 Cuculus sparverioides 鷹鵑 0.0282 0.0127 0.0020 Corvus macrorhynchos 巨嘴鴉 0.0034 0.0139 0.0040 0.0080 Ficedula hyperythra 黃胸青鶲 0.0907 0.2604 0.0318 Garrulus glandarius 松鴉 0.0253 0.1174 0.0637 Parus monticolus 青背山雀 0.4490 0.5604 0.0637 Cinclidium leucurum 白尾鴝 0.0662 2.5723 0.8218 0.0637 Heterophasia auricularis 白耳畫眉 0.0078 2.9512 0.8012 0.0650 Cuculus saturatus 筒鳥 0.0569 0.0516 0.0628 0.0497 0.0796 Dicaeum ignipectus 紅胸啄花鳥 0.0071 0.4539 0.4099 0.0884 Muscicapa ferruginea 紅尾鶲 0.1415 0.6618 0.0884 Carpodacus vinaceus 朱雀 0.0096 0.0955 Sitta europaea 茶腹鳾 0.0227 0.1240 0.1137 Actinodura morrisoniana 紋翼畫眉 0.1408 0.1273 Nucifraga caryocatactes 星鴉 0.0032 0.0116 0.1592 Stachyris ruficeps 山紅頭 0.0255 1.0793 1.7728 2.0005 0.5980 0.1592 Pnoepyga pusilla 鱗胸鷦鷯 0.1832 0.4707 0.2546 Troglodytes troglodytes 鷦鷯 0.2865 Bradypterus seebohmi 褐色叢樹鶯 0.0247 0.1452 0.3086 Erithacus johnstoniae 栗背林鴝 0.2440 0.3183 Hypsipetes madagascariensis 紅嘴黑鵯 0.6496 1.4793 2.6052 1.7361 0.3248 Garrulax morrisonianus 金翼白眉 0.0041 0.5349 0.5359 Cettia acanthizoides 深山鶯 0.3509 0.6496 Erithacus indicus 白眉林鴝 0.1071 0.6684 Brachypteryx montana 小翼鶇 0.1082 0.9250 0.7639 Liocichla steerii 藪鳥 0.0255 7.0449 3.7426 1.2733 Alcippe morrisonia 繡眼畫眉 0.2366 8.3255 5.6716 2.7780 1.2733
標、均勻度指標(Table 3),分別以變方分析檢 定其差異,各變項在6個群聚間均有顯著差異, 再以鄧肯氏多變域測驗對各群聚做兩兩比較: 高海拔群聚的海拔高度顯著大於其他5群聚,中 海拔II群聚的海拔高度顯著大於其他4群聚,中 海拔I群聚的海拔高度顯著大於其他3群聚,但 低海拔I、II、III群聚間則無顯著差異;中海拔 I、II群聚及高海拔群聚的鳥類族群總密度顯著 大於低海拔II群聚,但與低海拔I、III群聚無顯 著差異;中海拔I群聚的鳥種數與中海拔II群聚 無顯著差異,但顯著大於其他群聚;中海拔I群 聚的多樣性指數與中海拔II群聚及低海拔III群 聚無顯著差異,但顯著大於低海拔I、II群聚及 高海拔群聚;高海拔群聚及低海拔I群聚的均勻 度指數無顯著差異,但顯著小於其他群聚。 本研究各樣點的鳥種數與其海拔高度的關 係如Fig. 3,自海拔7.7-1,400 m,鳥種數與海拔 高度有顯著的正相關(r2 = 0.206, p = 0.00004), 但自海拔1,400 m起,鳥種數沿海拔梯度呈平緩 且略下降趨勢,鳥種數與海拔高度成負相關(r2 = 0.274, p = 0.0072),因此鳥種數在整個海拔梯 度上乃形成駝型分布。 以分布中點法(Rohde et al. 1993)檢驗各鳥 種之海拔分布中點值與海拔幅寬(Table 2)的關 係,刪除少於5個樣點的鳥種後,自海拔高度約 100至1,500 m,似有如Rapoport’s rule所認定之 物種分布之海拔愈高,其所佔有的海拔幅寬愈 大之趨勢(Fig. 4),但自海拔1,500 m起,此上升 趨勢消失,至海拔2,500 m以後,海拔幅寬略有 沿海拔分布中點值下降的味道。將海拔幅寬取
Fig. 3. Number of breeding bird species
along an elevation gradient in northern
Taiwan. The fit lines were calculated by
linear regressions that corresponded to the
ranges of elevation below 1400 m (r
2=0.206)
and above 1,400 m (r
2=0.274).
Continue Parus ater 煤山雀 0.0130 0.3750 1.6890 Yuhina brunneiceps 冠羽畫眉 11.7931 12.0766 2.2919 Aegithalos concinnus 紅頭山雀 0.4794 9.5440 3.5368 Alcippecinereiceps 灰頭花翼 0.1768 2.1703 4.9515 Regulus goodfellowi 火冠戴菊鳥 1.0128 27.4541Score on first axis of DCA 19.4 f 2) 124.82 e 176.44 d 343.75 c 444.45 b 577.00 a Elevation (m) 99.24 d 293.83 d 395.60 d 1261.48 c 2145.15 b 3050.58 a Total density (no./ha) 28.31 a b 13.66 b 28.58 a b 49.24 a 42.88 a 47.85 a No. of species 9.4 e 11.82 d e 18.50 b c 24.50 a 22.45 a b 15.75 c d Shannon diversity index 1.89 c 2.63 b 3.05a b 3.26 a 3.12 a b 1.93 c Shannon evenness index 0.58 b 0.74 a 0.73 a 0.71 a 0.70 a 0.49 b 1)Density (no./ha) of dominant species (underlined) in each community.
2)Means followed by different letters within columns significantly differ (by Duncan’s new multiple range test, p < 0.01).
對數值後,海拔分布中點值與海拔幅寬對數值 之二次迴歸模式為log Range = 3.1999 - 0.0029 × Midpoint + 0.2264 × Midpoint2 (r2 = 0.798)。
討 論
台 灣 北 部 陸 域 繁 殖 鳥 類 可 分 為 低 海 拔I、 II、III、中海拔I、II及高海拔等6個群聚,海拔 梯度是影響鳥類群聚差異之重要因子,因此可 以海拔梯度做為鳥類群聚的分界,本研究之低海拔三群聚在1,000 m以下,中海拔兩群聚介於 1,000-2,700 m,高海拔群聚在2,700 m以上,此 結果與Jai (1977)、Huang (2001)之研究相較: Jai (1977)將台灣鳥類分成小於1,000 m的低海 拔帶、1,000-2,300 m的中海拔帶及超過2,300 m的高海拔帶,Huang (2001)則以900 m做為 北 部 繁 殖 鳥 類 之 低 中 海 拔 分 界 , 但 其 研 究 中 2,100-2,800 m之調查樣點僅有1 個,故無法明 確指出北部繁殖鳥類之中高海拔分界。本研究 結果與Jai (1977)和Huang (2001)之低中海拔分 界相似,而與前者之中高海拔分界有所不同, 可能是因為本研究地區局限於台灣北部,所得 到的群聚分布類型乃屬於地區性的現象,故不 同於Jai (1977)之全省調查結果;但也可能是因 為本研究依照面積比例取樣,故高海拔的樣點 數稍嫌不足,使得鳥類群聚在中高海拔處出現 不連續現象,因而拉大了2,700 m以上樣點與其 他較低樣點的差異程度,因此需要累積更多的 調查資料,才能了解差異的原因。至於低海拔 3群聚之海拔分界不明顯,Huang (2001)的研究 曾指出對低海拔鳥類群聚最重要的影響因子為 植被覆蓋狀況,其次才為海拔梯度,此應可以解 釋低海拔三群聚在海拔上無明顯差異的現象。 鳥類群聚除了受海拔梯度的影響之外,亦 有研究者認為植群係重要影響因子(Jai 1977, Ding 1993)。Su (1984)以溫度梯度將台灣山地 植群帶依低至高溫劃分為高山帶、冷杉帶、鐵 杉雲杉帶、櫟林帶、楠櫧林帶及榕楠林帶,各 帶 大 致 分 布 在 一 固 定 海 拔 段 。 若 以Su (1984) 所 列 之 台 北 木 柵 、 烏 來 山 區 、 宜 蘭 哈 盆 至 雙 連 埤 、 太 平 山 、 大 元 山 、 拉 拉 山 北 及 南 坡 、 南 湖 大 山 北 坡 及 大 霸 尖 山 北 坡(以上等地皆在 本 研 究 範 圍 內)之植群及海拔分布範圍來看, 則 本 研 究 地 區 內 之 植 群 分 布 大 致 為 : 高 山 帶 在3,100-3,700 m間、冷杉林帶在2,600-3,400 m間、鐵杉雲杉林帶在2,000-2,800 m間、櫟 林帶上層在1,800-2,200 m間、櫟林帶下層在 1,200-1,800 m間、楠櫧林帶在300-1,200 m間、 榕楠林帶低於300 m,以其與本研究地區之鳥類 群聚相對照:高海拔鳥類群聚約分布在高山帶 及冷杉林帶、中海拔II鳥類群聚約分布在鐵杉 雲杉林帶及櫟林帶上層、中海拔I鳥類群聚較接 近櫟林帶下層、低海拔三群聚大致分布在楠櫧 林帶及榕楠林帶,故本研究之鳥類群聚更迭與 其整個區域之植被變動可謂大致互相吻合。Jai (1977)認為其研究中,中海拔鳥類群聚至高海 拔鳥類群聚所發生的鳥類組成更動與植被變化 一致,在該海拔高度範圍內,植被由針闊葉混 淆林轉為針葉林;但在低海拔鳥類群聚至中海 拔鳥類群聚,其間之變動與植被的變化卻不一 致,即植被未有明顯的林型變化,這可能是因 為其調查範圍涵括全台灣,而全台灣的植被與 海拔高度的關係會因區域的不同而有所差異, 故由全台灣的尺度來看鳥類群聚與植群的關係 時,似乎無法得到明確的結果,反之,在較小 的尺度下應可看出鳥類群聚與植被間的關係, 如Ding (1993)研究玉山地區成熟林之鳥類群 聚,在不同植群內做鳥類調查,結果發現鳥類 群聚依不同植群而劃分為低層闊葉林、高層闊 葉林、針闊葉混合林、雲杉林、鐵杉冷杉林及 圓柏灌叢等六個群聚。因此研究尺度的差異,可 能會對鳥類群聚與植群的關係產生不同的結論。 比 較 各 群 聚 的 鳥 類 總 密 度 , 中 海 拔I群聚 的鳥類總密度顯著大於低海拔II、III群聚,其 他群聚則介於其間;大體而言,中、高海拔地 區可以支持較大的鳥類族群密度,乃較適合鳥 類生存的環境。中海拔群聚的鳥種數顯著大於
Fig. 4. Midpoint plot of elevational ranges
of breeding birds in northern Taiwan.
其他群聚,其鳥類多樣性指標亦顯著大於其他 群聚(惟與低海拔III群聚無顯著差異),表示中 海拔地區的鳥類群聚有較為豐富且多樣性較高 的鳥種。均勻度指標在低海拔II、III群聚及中 海拔群聚皆無顯著差異,但在高海拔群聚及低 海 拔I群聚顯著較小,可能是分別有特別優勢 的鳥種如麻雀(Passer montanus)及火冠戴菊鳥
(Regulus goodfellowi) (Table 3),造成均勻度的 降低;相對來說,其他群聚的鳥種就較為平衡。 一 般 公 認 種 豐 富 度 沿 緯 度 梯 度 呈 遞 減 的
變化(Begon et al. 1996),許多研究更認為種豐
富度在海拔梯度上有類似緯度梯度的變化,即
沿海拔梯度呈線性遞減的趨勢(Stevens 1992,
Begon et al. 1996),但是Rahbek(1995, 1997) 與 其他人(Lees et al. 1999, Colwell and Lees 2000) 認為種豐富度隨海拔上升在中海拔區到達高峰 的駝型分布應該比遞減分布更為常見。以本研 究結果而言,鳥種數沿海拔梯度的分布雖有線 性遞增的趨勢,但在中海拔1,000-2,000 m間達 到高峰,往海拔2,000 m以上則遞減,此現象符 合Rahbek所提出的駝型分布,但不論是海拔前 段的上升或後段的下降直線,其對於鳥種數的 預測能力都不高,這或許是由於各調查點之自 然度(或受人為干擾的程度)可能相差很多,因而 造成鳥種數的變異度很大,未來必須再蒐集各調 查點的環境背景資料,以求得較佳之迴歸模式。 種 豐 富 度 在 海 拔 梯 度 上 呈 駝 型 分 布 , 有 許多可能的解釋機制(Begon et al. 1996),除了 mid-domain effect之外,本研究另就生產力假 說及人類活動之干擾來說明:(一)生產力假說: 種豐富度與地表植被生產力的關係可能是線性 遞增(MacArthur 1972, Brown 1988)或種豐度在 中度生產力達到高峰(Rosenzweig and Abramsky 1993, Rosenzweig 1995),Ding (2001)認為台灣 鳥類種豐富度與地表植被生產力呈正相關,地 表植被生產力在中海拔帶最高,故可解釋鳥類 種豐富度在中海拔有高峰的現象。(二)人類活動 的干擾:人為干擾會造成地表植被及景觀的改 變,進而影響鳥類的分布範圍。如偏好森林棲 地的鳥類可能因平地及低海拔的過度開發,而 造成其海拔分布下限向上退縮,因此降低平地 及低海拔的鳥種豐富度,另一方面,中海拔地 區也因森林砍伐、農墾及聚落發展等現象,使 得低海拔農耕地的優勢鳥種向上入侵中海拔, 因而增加了中海拔區的鳥種豐富度。故此項因 素應可解釋鳥類種豐富度由中海拔往低海拔遞 減的趨勢。 本 研 究 中 鳥 種 分 布 之 海 拔 與 其 海 拔 幅 寬 之 關 係 , 以 分 布 中 點 法 所 得 之 結 果 : 自 海 拔1 0 0 - 1 , 5 0 0 m , 鳥 種 之 海 拔 幅 寬 似 有 隨 分 布 之 海 拔 高 度 逐 漸 上 升 的 趨 勢 , 在 中 海 拔 約 1,500-2,500 m區間,鳥種的海拔幅寬沒有明顯 趨勢,至海拔約2,500 m後則呈下降趨勢,這樣 的結果不符合Rapoport’s rule,該原則指明物種 的海拔高度與其海拔幅寬有正相關的關係,亦 即物種的海拔高度愈高,其分布的海拔幅寬愈 大。雖然一些研究曾得到類似Rapoport’s rule的
結果(Fleishman et al. 1998, Sanders 2002),並 且以此原則當作是種豐富度在海拔梯度呈線性
遞減的可能原因之一(Stevens 1992),但也有學
者指出Stevens (1992)所展示的資料中,其實有 部分顯示出物種分布的海拔幅寬在中海拔段有 高峰(Colwell and Hurtt 1994, Rahbek 1997)。 本研究結果較符合Colwell and Lees (2000)所提 之三角形模式:分布中點在海拔較低或較高的 鳥種有較小的海拔幅寬,分布中點在中海拔地 區的鳥種,則有部份鳥種有較大的海拔幅寬。 Jai (1977)自海平面至接近4,000 m的高山進行台 灣陸棲繁殖鳥類的調查,在中部山區記錄124種 台灣陸棲繁殖鳥類於繁殖期的海拔分布高度, 所有鳥種的海拔幅寬沿其海拔分布中點值的型 態(Fig. 5),亦與本研究結果有類似之處:自平 地至海拔高度1,000 m,鳥種的海拔幅寬沿其海 拔分布中點值呈上升趨勢,但自1,000 m起,鳥 種的海拔分布幅度散布在900至2,800 m之寬度 間,直至3,000 m後才有下降趨勢;亦即分布中 點愈靠近海拔兩端的鳥種,其海拔幅寬愈小, 而分布中點靠近海拔中段的鳥種,則有部份鳥 種具有較大的海拔幅寬,故此項結果亦大致符 合Colwell and Lees (2000)所提之等邊三角形模 式,足見該模式確實可以描述台灣局部地區鳥 種之海拔分布與幅寬的關係。因此本研究認為
Colwell and Lees (2000)所論述之mid-domain effect - 物種因受地理邊界限制其分布範圍,而 造成物種豐度在中間地帶產生高峰,較能夠解 釋台灣北部繁殖鳥類豐富度在中海拔地區有高 峰的現象;而Rapoport’s rule在本研究中則無法 獲得支持。但是若將冬季鳥類分布資料涵蓋進 來,原來由中海拔往兩端下降的三角形模式是 否可能改變成Rapoport’s rule的線性曲線或其他 曲線,則有待更多的調查研究。