CXCR4趨化激素受體的表現促進基底細胞癌的腫瘤發生; Expression of CXC Chemokine Receptor CXCR4 Enhances Basal Cell Carcinoma Tumorigenesis
72
0
0
全文
(2) 中文摘要 除了能夠引導白血球的趨化行為外,趨化激素與其受體也扮演了 促使腫瘤生長與轉移的角色,如腫瘤細胞的增生、凋亡與不正常的血 管增生。本篇研究在於探討趨化受體與表皮基底細胞癌化過程的相關 性。研究顯示,CXCR4 趨化受體在人類基底細胞癌株或患者病灶處所 取下的基底細胞癌組織樣品內存有高度的表現量。此外,我們更假設 在 CXCR4 趨化受體過度表現下將會促使表皮基底細胞的癌化。藉由反 轉錄病毒載體轉染技術,CXCR4 受體蛋白基因被轉染至表皮基底細胞 癌(CXCR4-BCC 細胞)並經由體外細胞增殖實驗,顯示 CXCR4-BCC 細胞 在低血清培養下以 CXCR4 受體接受分子 CXCL12 處理,會導致 CXCR4-BCC 細胞的增生。在細胞凋亡試驗下,CXCL12 趨化激素也可以 防止 CXCR4-BCC 細胞進行程序性死亡,但此現象卻受到專一性的 CXCR4 阻斷生? ,T22 peptide 作拮抗性的中和而阻斷。在體外血管 新生試驗方面,添加以 CXCL12 趨化激素處理 CXCR4-BCC 細胞的培養 液,能夠促使人類內皮細胞管狀形成,此現象亦可受到 T22 peptide 作拮抗性的阻斷。另外,我們將 CXCR4-BCC 細胞以皮下注射進入裸鼠 進行異種移植而致腫瘤形成,然而 T22 的添加也確實能造成腫瘤的消 1.
(3) 減。 因此, CXCR4 受體蛋白的表現或許與表皮基底細胞的癌化發生 過程有關, 而阻斷 CXCR4 受體蛋白的活化或許可成為未來治療此類 癌症的方向之一。. 2.
(4) 英文摘要 Beside the well-known capacity to regulate leukocyte trafficking,. chemokines and their receptors also. play. important roles in the regulation of cell proliferation, apoptosis, and angiogenesis, which may enhance tumor growth or metastasis. Herein we investigated the possible involvements of chemokine receptors in the pathogenesis of cutaneous basal cell carcinoma (BCC), the most common human cancer. We found high expression of chemokine receptor CXCR4 in a human BCC cell line and a subset of tissue samples from BCC lesions. Furthermore, we addressed whether overexpression of CXCR4 may alter BCC tumor progression. By retroviral transduction, CXCR4 gene. was. transferred. into. BCC. cells. (CXCR4-BCC). and. functionally expressed. In cell proliferation assay in vitro, CXCR4-BCC cells were cultured under low serum concentration and subject to treatments with the ligand for CXCR4, CXCL12. CXCL12 treatments resulted in significant increase of CXCR4-BCC proliferation, which could be neutralized by anti-CXCR4 3.
(5) monoclonal antibodies. In apoptosis-resistance assay in vitro, treatments with CXCL12 decreased apoptosis level in CXCR4-BCC, which could be negated by CXCR4-blocking peptide, T22. In angiogenesis assay in vitro, conditioned medium from CXCR4-BCC cells pre-incubated with CXCL12 could enhance tubule formation of human endothelial cells, which could be reversed by T22 addition. Moreover, xenograft tumor transplants by s.c. injections of CXCR4-BCC cells yielded significant tumor progression in nude mice, whereas additional serial injections of T22 resulted in significant tumor regression. Thus, CXCR4 expression may play critical roles in BCC tumorigenesis, and functional blockade of CXCR4 could be a potentially promising therapeutic strategy.. 4.
(6) 致謝辭 在兩年的研究所求知階段,特別感謝所內師長的細心教導。尤其 是指導教授吳孟澤老師對論文的細心教導指正,及共同指導教授周寬 基老師在第一年的研究所階段,針對研究方法及基礎醫學研究方面扎 實的訓練。使我了解像吳教授•周教授一樣,從事一個臨床醫學的研 究,所必須具備的嚴謹及全心投入的態度。這是我所必須努力學習的 態度。並且對於在求學階段,中國醫藥大學附設醫院皮膚科的所有同 仁對我的協助,使我能順利完成學業感到感激。最後感謝我的家人對 我的支持,使我能全心的求知。而能達成一個階段性的成就。謝謝所 有上述的師長•朋友•家人,及所有在我就學階段協助過我的人。. 5.
(7) 目錄 摘要. --------------------------------------1. 致謝辭. ------------------------------------5. 第一章. 前言. 第二章. 第三章. 第四章. 第五章. -----------------------------8. 第一節. 研究緣起. -------------------8. 第二節. 研究目的. -------------------10. 文獻探討. -------------------------11. 第一節. 趨化激素與其受體之定義. 第二節. 皮膚基底細胞癌之定義. 研究架構與研究設計. -----11 -------16. ---------------18. 第一節. 研究設計. -------------------18. 第二節. 研究架構. -------------------19. 第三節. 研究假說. -------------------22. 研究材料及統計方法. ---------------23. 第一節. 研究材料及資料收集方法. 第二節. 研究變項及操作型定義. 第三節. 資料處理及分析方法. 研究結果. -----23 -------24. ---------32. -------------------------33 6.
(8) 第六章. 討論. -----------------------------52. 第一節. 結果討論. -------------------52. 第二節. 研究限制. -------------------57. 第七章. 結論與建議. -----------------------58. 第一節. 結論. -----------------------58. 第二節. 建議. -----------------------59. 參考文獻及附錄 簡單自述. ----------------------------60. ----------------------------------70. 7.
(9) 第一章 前言 第一節 研究緣起. 趨化激素為含有四個胱氨酸固定保留區位的類細胞激素小分子 並具有結合某一 G protein-coupled 受體蛋白的能力 (1)。 除了 擁有能夠引導白血球趨化性的能力外並與感染與發炎現象有關。事 實上,趨化激素與其受體在生物體內扮演廣泛的角色(1, 2)。CXCR4 受體蛋白與其結合分子, CXCL12 (即 stromal-derived factor-1, SDF-1)是此次研究的主題。 除了同樣擁有能夠引導白血球趨化性的 能力與感染與發炎現象有關外,CXCR4 受體蛋白也被發現廣泛地表現 於造血細胞 (3),血管內皮細胞 (4),神經元,微神經膠細胞與星 狀細胞中 (5),與人類後天免疫缺乏病毒(HIV-1) 的感染(幫助 HIV-1 病毒進入 T 細胞)、造血、胚胎形成、器官形成與血管新生有著相當 大的關聯。 除此之外, CXCR4 受體蛋白也與腫瘤形成有關(8-13)。 Muller 等人 (8) 早期已證明 CXCR4 受體蛋白在乳癌細胞中有高度表現,且 發現與其接合之 CXCL12 趨化激素在乳癌細胞易進行轉移的部位存有 較高的濃度。Muller 等人更進一步以體內實驗證明 CXCR 受体蛋白在 單株抗體拮抗中和後,可以明顯地減低乳癌細胞轉移至鄰近的淋巴 8.
(10) 結與肺臟器官,此現象指出乳癌細胞受到 CXCR4 受體蛋白的影響而 有選擇性地轉移至某特定器官。為了釐清 CXCR4 在癌細胞轉移的角 色, Murakami 等人 (9) 也以體內實驗證明 CXCR4 的表現,可以有 選擇性地促使黑色素細胞轉移至肺臟器官並顯示 CXCR4 受體蛋白可 促使腫瘤吸附至內皮細胞與促進腫瘤細胞的生長。 除了乳癌與黑色 素細胞, CXCR4 受體蛋白也被發現在不同癌細胞中表現 (前列腺, 胰臟,肺臟,與腦) 並與癌化發生有關,包括腫瘤的生長,遷移, 吸附與引導血管新生 (10-13)。 表皮基底細胞癌是一種人類好發型的惡性腫瘤,其屬於表皮腫瘤 細胞局部之浸潤與不正長之血管新生, 且發生率持續上升 (14, 15)。 雖然影響表皮基底細胞進行癌化的病理因子有很多,包括 UV 光的暴露 (16), IL-6 的傳導 (17), 及與胚胎發生有關的 Sonic hedgehog 生化路徑中某些基因的變異(18,19,20)等,但實際上表 皮基底細胞腫瘤的形成仍然有許多未知。. 9.
(11) 第二節 研究目的. 為了證明趨化激素受體扮演了表皮基底細胞腫瘤化的過程,本 次實驗使用人類表皮基底細胞株並分析其趨化激素受體之表現量。我 們證實了 CXCR4 受體蛋白在表皮基底細胞株與腫瘤組織檢體樣品內 有高度的表現量。藉由體外實驗證明,經反轉錄病毒載體轉染 CXCR4 基因之表皮基底細胞能促使腫瘤細胞的增殖、防止細胞本身走向程序 性死亡、造成細胞遷移與引導血管新生。 更重要的是,裸鼠被施打 CXCR4 基因轉染之表皮基底細胞後,體內發生癌化,腫瘤細胞浸潤與 不正常之血管新生。此現象可經由專一性的 CXCR4 阻斷生? , T22, 的拮抗而消除。 (21)。 因此, CXCR4 受體蛋白的表現對於表皮基 底細胞腫瘤化的發生過程扮演了極重要的角色。. 10.
(12) 第二章文獻探討 第一節 趨化激素與其受體之定義. I.趨 化 激 素 與 其 受 體 之 命 名 及 結 構 (1) 趨化激素為類似細胞激素的趨化蛋白,具有共通的構造,能 吸引白血球的趨附。 (2) 趨化激素之結構為具有四個胱胺酸 ( cysteins ) 在胺基端 的前兩個 cysteins 分別與第二,第四個 cysteins 形成雙硫 鍵,而造成其具有共同特色之三維摺疊結構。 (3) 趨化激素與細胞激素之異同點如下: j 都是經由白血球或組織細胞藉由 paracrine 或 autocrine 所產生的蛋白質。 k但趨化激素分子較小(約 8-10 kd) 需透過其相對應 G-protein coupled 受體來產生作用。 (4) 趨化激素依據它前兩個胱胺酸的空間排列,可分成四種次類: j CXC chemokines:一個胱胺酸接一個非保留的胺基酸再連接 胱胺酸。 k CC chemokines: 二個胱胺酸連接,中間沒有額外附加之胺 11.
(13) 基酸。 l C chemokines: 無連接之胱胺酸。 m CXXXC: 二個胱胺酸中間隔三個胺基酸。 (5) 趨化激素受體 (chemokine receptor): 是一個 G-protein coupled,具有七個穿透胞膜蛋白的受體, 可與 chemokines 相結合 (6) 目前之命名傾向以數字為系統性之命名原則,例如:本研究 之 CXCL 12 中 CXC 表 chemokines, L 表 ligand, 12 是數字命 名編號。而 CXCR 4,CXC 表示 chemokines 趨化激素, R 表示 receptor 受體。. 12.
(14) II. 趨化激素與其受體之作用. 趨化激素可結合在其相對應之趨化激素受體,經由訊息傳導路 徑,而使細胞產生趨化作用 (chemotaxis)。 目前己知有以下之作用: (1) 調控白血球的趨化性: j使白血球從血管趨附至血管內皮進而趨附至特定之發炎部位 k調節 T 細胞之移行未經抗原活化之 T 細胞,藉由表現樹突細 胞的抗原使 T 細胞活化成為記憶性 T 細胞(memory T cell), 而 藉由趨化激素之作用而易移行至周邊特定發炎位置。 l促使樹突細胞移行至局淋巴組織:抗原表現細胞,包括樹突 細胞等是在免疫反應中的關鍵步驟,可將抗原帶至局部淋巴結,促 使淋巴細胞辨識抗原。趨化激素在其中扮演了關鍵性的角色。 趨化激素能導引樹突細胞至局部之淋巴結,而在淋巴結中又用 各種趨化激素之作用使 T 細胞趨近辨識抗原。 (2)感染性疾病方面: 微生物可能產生類似趨化激素或趨化激素受體之蛋白質,而與宿 主細胞結合。如:kaposis sarcoma 的 ( human herpes virus-8) 13.
(15) 的 GPCR 受體能與多種 chemokines 結合,而融入宿主細胞,甚至活 化或啟動 kaposis 肉瘤的生長。 (3)調控腫瘤生長方面: j趨化激素透過各種不同的方式來調控腫瘤的生長及人體的免 疫反應。 例如,造成血管生成、抗細胞凋亡、促進細胞分裂、誘導免疫 反應以抑制腫瘤細胞等。 在惡性黑色素瘤方面,趨化激素 CXCL1, CXCL2, CXCL8 等可以 促進腫瘤的生長。而 CXCL8 又可促使惡性黑色素瘤細胞分泌 MMP-2 ( type Ⅳcollagenae)。而促進腫瘤細胞的侵犯及轉移能力。腫瘤 細為何易轉移至特定器官。. 14.
(16) III. 在 腫 瘤 生 長 方 面 CXCR4 之 研 究 現 況. (1). CXCR4 及 CCR7 在乳癌腫瘤上有較正常乳房組織高度的表 (2). CXCR4 之接合趨化激素 CXCL12 在乳癌腫瘤轉移器官上有較強表 現,此二點似乎暗示著 CXCR4 和乳癌之腫瘤生長及轉移有相關。 (3). CXCR4 也被發現在其他腫瘤上,例如:卵巢癌、胰臟癌、肺癌、 腦癌等。 (4). Muller 等人發現若用抗體中和 CXCR4 則乳癌細胞則不易在富有 CXCL12 之肺組織上生長。 (5). 在惡性黑色素瘤方面也有較正常組織高度表現 CXCR4 受體。. 15.
(17) 第二節 皮膚基底細胞癌之定義. 皮膚基底細胞癌是最常見的皮膚癌,可能也是最常見之一種癌 症,它通常造成局部之侵犯、破壞,但絕少轉移。 臨床表現: (1) 好發在 40 歲以上之人,男性略多於女性,白種人發生率高於 黃種人或黑人。 (2) 致病因素:包括皮膚較白者,長期陽光曝曬,或年輕時曾有 大量之陽光曝曬,尤其是 14 歲青春期之前。 (3) 好發部位:通常為單一病,灶好發在臉,部若腫瘤位在眼內 眥,外眥,鼻唇溝,日後通常較危險,對周遭組織破壞較大。 (4) 分為四型: j結節型(Nodular) :膚色或珍珠色之丘疹或結節其上可有血 管擴張。 k潰瘍型 (ulcering):為一個囓齒狀之潰瘍,中間有一黑色 痂皮。 l硬皮型(sclerosing):皮膚硬化,類似疤痕的表現。 m表淺型(superficial):為較薄,表淺之紅色到黑色斑塊, 16.
(18) 可能為多發性,其上可能有血管擴張現象。 (5) 病理上可見一增生性非典型之基底細胞,呈現卵圓型深藍色 之細胞,在腫瘤邊緣之細胞呈現柵狀排列。 (6) 治療上,一般以單純之手術切除即可,冷凍治療或電燒也有 不錯的療效。但在危險區域,則須注意邊緣或深部侵犯,再施 行大範圍之切除。其他像局塗抹 5-fluorouracil 軟膏或 imiquimod 軟膏也可應用在較表淺之腫瘤,因絕少轉移,故只 要針對侵犯部位徹底切除,預後都不錯。. 17.
(19) 第三章 研究架構與研究設計 第一節 研究設計. 為探討趨化激素受體 CXCR4 的表現和基底細胞癌之調控作用 關係,我們運用一系列之體外及體內試驗來證實我們的想法。. 18.
(20) 第二節 研究架構. 1. 首先,確認基底細胞癌上具有 CXCR4,我們用以下之方法來證 實: (1)作 quantitative RT-PCR 來證實基底細胞癌細胞株(BCC cell lines)有趨化激素受體 CXCR4,且跟其他之正常細胞株比較是否 其含量明顯增多。 (2)在基底細胞癌的組織切片上,以免疫組織化學方法測定是否 有趨化激素受體 CXCR4。 (3)再於基底細胞癌組織切片上,用原位雜交之方法看是否有趨 化激素受體 CXCR4。 2. 接著,利用以下方法確定 CXCR4 的表現和基底細胞癌之腫瘤生成 是否有關 (1) 利用轉殖技術,將基底細胞癌細胞株(BCC cell lines)轉殖 成為高度表現 CXCR4 之 BCC-cell line (2) 具有 CXCR4 能促進基底細胞癌增殖, 比較有轉殖 CXCR4 之 CXCR4-BCC 細胞株和沒有轉殖 CXCR4 之 pLNCX2-BCC 細胞株那一個較能增殖。 19.
(21) (3) 具 CXCR4 能抵抗細胞凋亡, 比較有轉殖 CXCR4 之 CXCR4-BCC 細胞株和沒有轉殖 CXCR4 之 pLNCX2-BCC 細胞株那一個能保護細胞,不會自我凋亡 (apoptosis) 。 (4) 具有 CXCR4 之基底細胞癌能有較佳之移行及侵犯能力。 j 我們利用二維空間的移行,來判別有無 CXCR4 之轉殖對 細胞移行之影響。 k 我們用細胞外基質薄膜的侵犯能力,來比較有無 CXCR4 轉殖的差異。 (5) 血管新生能力: 比較有無轉殖 CXCR4 之 BCC-cell line 是否較能促進血管內 皮細胞形成管狀構造,且能分泌較多之血管生成因子。 3. 腫瘤之生長需很多方面因子促成,因此我們用間接及直接證據來 證明 CXCR4 之能促進基底細胞癌之生長。 (1)間接證據: 利用體外試驗中增殖 ( proliferation),抗凋亡能力 (antiapoptosis),移行(migration),侵犯(invasion),血管新 生(angiogenesis), 能力之增強,來表示 CXCR4 能促進 BCC 腫 瘤生成。 20.
(22) (2)直接證據: 利用體內實驗 (in vivo)中,觀察轉殖 CXCR4-BCC cell line 接種到裸鼠之皮下直接觀察腫瘤的大小是否有不同,形態是否有 血管新生,來判斷 CXCR4 是否能促進基底細胞癌之生長。. 21.
(23) 第二節 研究假說. 1. 基底細胞癌上具有較正常細胞多之趨化激素受體 CXCR4 2. 基底細胞癌之腫瘤生長和趨化激素受體 CXCR4 之表現有關 3. 趨化激素受體 CXCR4 之高度表現能促進基底細胞癌之增殖,抗凋 亡能力,細胞移行,侵犯能力,血管新生能力 4. 趨化激素受體 CXCR4 表現較強之基底細胞癌有較佳之腫瘤生長 能力 5. 能拮抗趨化激素受體 CXCR4 表現之胺基酸,間接造成基底細胞癌 之生長抑制,不管是增殖,抗凋亡,細胞移行,侵犯,血管新生 等方面之能力. 22.
(24) 第四章 研究材料及統計方法 第一節 研究材料及資料收集方法. 細胞株, 實驗動物與試劑. 人類 BCC-1/KMC 細胞株由高雄醫學院教授蔣連財 贈予,其分離取得之方式可參考文獻 17, 22。細胞以含有 10% 之小牛血清 RPMI 1640 培養基培養。 8-12 週大之母裸鼠由國家實 驗動物中心提供 (台北, 台灣), 並由動物實驗管理小組核可動物 實驗的進行。人類黑色素瘤細胞株 A2058, 鱗狀上皮瘤細胞株 A431 與正常纖維母細胞 CCD-966SK, 由新竹生物資源保存及研究中心 細胞庫所購得。人類角質細胞與黑色素細胞之分離取得與培養可參考 文獻 23。人類永生血管內皮細胞(HMEC-1)獲 Dr。FJ 贈與,其培養 方式可見文獻 25。人類 CXCR4 單株抗體與免疫球蛋白向 R&D 公 司購得。趨化激素 CXCL12 向 Peprotech, Rochkyhill, NJ 購得。 CXCR4 受體拮抗月生? ,T22 (NH2-RRWCYRKCYKGYCYRKCR-COOH)由 United Biochemical 23.
(25) Research 公司合成提供,純度大約為 95%。. 24.
(26) 第二節 研究變項及操作型定義. 反轉聚合酵素反應定量試驗. 以 Qiagen 公司之 RNeasy 試劑組萃取表皮基底細胞 BCC-1/KMC 之 RNA,再以此 RNA 為模板,利用反轉聚合酵素製做出 cDNA。 以 ABI 公司出廠之 PRISM 7000 DNA Sequence Detection System 與 SYBR Green 對反轉聚合酵素反應產物進行即時定量偵測。並以人 類 glyceraldehyde 3 -phosphate dehydrogenase (G3PDH)作為內部定量 控制組。使用 Primer Express software( Applied Biosystems)設 計 之 趨 化 激 素受 體 (CCR1-CCR10 、 CXCR1-CXCR6) , 趨 化 激 素 CXCL12 , 內 皮 生 長 因 子 (VEGF) ,VEGF-B, VEGF-C, VEGF-D , cycloxygenase-2 (COX-2) 與 G3PDH 等的特殊初始子(primer)。. 25.
(27) 免疫組織化學與原位雜交分析. 表皮基底腫瘤組織細胞樣品以 antihuman CXCR4 之單株抗體、 (clone #44716,老鼠的 IgG2B) 定之. istotype mouse IgG 2B、 生物素標. antimouse 二 級 抗 體 ,. 與 streptavidin-horseradish. peroxidase enzyme (後者向 Zymed, South San Francisco, CA 取 得)標定並以 DAB(diaminobenzidine, from DAKO, Carpinteria, CA)染色、蘇木素復染。原位雜交反應偵測人類 CXCR4 mRNA 的方法 可 參 考 文 獻 27 。 所 使 用 之 生 物 素 標 定 人 類 CXCR4 antisense oligonucleotides 的序列如下: 5’-GTAACCCATGACCAGGATGACCAATCCATT-3’(195-166), 5’-AGACTGATGAAGGCCAGGATGAGGACACTG-3’(395-366), 5’-ACTGGAACACAACCACCCACAAGTCATTGG-3’(601-572), 5’-CCAGGAGGATGAAGGAGTCGATGCTGATCC-3’(802-773), 與 5’-GCTGGAGTGAAAACTTGAAGACTCAGACTC-3’(1056-1027)。. 26.
(28) 以 逆 轉 錄 病 毒 載 體 轉 染 CXCR4 基 因. 萃取 BCC-1/KMC 細胞中的 RNA 並以反轉錄酵素產生 cDNA,使用 酵素聚合反應與初始子對 (5′- GGAAGATCTATGGAGGGGATCAGTATATA-3′ 與 5′-CCCAAGCTTTTAGCTGGAGTGAAAACTTG -3′, 具有 Bgl II 與 Hind III 限制酵素切位) 產生人類 CXCR4 的 cDNA。 之後 CXCR4 cDNA 以 酵素剪切接入 pLNCX2 retroviral 載體 (Clontech, Palo Alto, CA)。之後將 pLNCX2-CXCR4 與 不含 CXCR4 基因之 pLNCX2 質體轉殖 進入 BCC-1/KMC 表皮基底細胞癌,而產生 CXCR4 基因之表皮基底細胞 癌 (CXCR4-BCC) 與 不含 CXCR4 基因之表皮基底細胞癌(pLNCX2-BCC) 二種細胞 (28)。 另外,為了能得到 CXCR4 表現量高的 CXCR4-BCC 細胞株,我們選擇使用含有 FITC-conjugated anti-human CXCR4 單 株抗體 (clone 12G5)與 anti-FITC 之磁力珠(Miltenyi Biotec, Auburn, CA)分離取得同質性高的 CXCR4-BCC 細胞。 此外,培養時 添加 G418 (GIBCO)至培養基內以維持 CXCR4-BCC 細胞中 CXCR4 受體 蛋白的表現量。. 27.
(29) 細胞增殖試驗. 細 胞 增 殖 試 驗 方 法 可 參 考 Murakami 等 人 的 文 獻 (9) 。 pLNCX2-BCC細胞株與CXCR4-BCC細胞株在正常血清濃度(10% FCS)或 低濃度血清(0.5% FCS)下,以 3×104 /孔細胞密度種植在96-孔培養盤 中。 依指定之實驗組或對照組分別添加anti-human CXCR4 單株抗體 (clone #44716) 、mouse IgG 2B isotype或 PBS後,再添加CXCL12 (500 ng/ml)處理24或48小時(圖 3),之後以市售WST-1試劑組偵測腫瘤細 胞增殖的程度 (Quick cell 細胞增殖試劑組, BioVision, Mountain View, CA) (29)。. 28.
(30) 細胞凋亡試驗. pLNCX2-BCC 細胞株與 CXCR4-BCC 細胞株以指定的試劑,500 ng/ml CXCL12 或 CXCR4-blocking peptide T22 (1µg/ml),處理 24 小 時後, 接著經 UVB 照射(25 millijoule/cm 2) 以誘導細胞凋亡(圖 4)。 之後細胞凋亡的程度以市售之 BD ApoAlert caspase-3 試驗培養盤 (Clontech) 偵測其 caspase-3 的活性。 簡而言之, BCC 細胞 lysates (2 × 105 細胞) 加入表面已覆蓋 caspase-3 受質 (Asp-Glu-Val-Asp or DEVD)連結螢光物質(7-amino-4-methyl coumarin, AMC)的 96 孔 培養盤,一旦受質經由 caspase-3 水解後,其所釋放的螢光物質 (7-amino-4-methyl coumarin, AMC)可經由 380 nm 波長光源激發後 釋放出 460 nm 波長,再以讀取儀偵測。. 29.
(31) 細胞移行與侵犯試驗. 二維遷移試驗方法可參考文獻 13。 簡而言之,將表皮基底細胞 癌種於培養盤中至長滿,之後以刮取器將每個培養孔挖掉約 2mm 寬區 域的細胞後,將原先的培養基置換成有添加 CXCL12 (500 ng/ml)或 T22 (1µg/ml) 的 培 養 液 ( 圖 5) 。 培 養 48 小 時 後 , 以 CCD (charge-coupled device) camera 與 image analysis software 定 出移行細胞之原始寬度並減去剩餘寬度的值來計算細胞遷移的距離 (PhotoImpact 7。0, Ulead Systems Inc。 Taipei, Taiwan)。 另 外,使用表面覆蓋有細胞基質的 Boyden chamber 來進行細胞侵犯試 驗(Matrigel, from BD Biosciences Discovery Labware, Bedford, MA)。侵犯程度以細胞穿透細胞基質膜的數量決定(10)。. 30.
(32) 血管新生體外試驗. 105 pLNCX2-BCC 與 CXCR4-BCC 細胞培養於 6-well 培養盤中, 依 實驗組或控制組選擇是否處理 CXCL12 趨化激素。如圖 6,在某些指 定的實驗組或控制組添加 T22。 培養 24 小時後, 收集各組之培養 液,並使用含有 3,000 kD cut-off 濾膜之 Centricon 離心管做離 心濃縮(~4 倍) (Millipore, Billerica, MA)。 同時, 將 10,000 HMEC-1 細 胞 種 植 於 表 面 受 Matrigel 處 理 的 96- 孔 盤 中 (BD Biosciences Discovery Labware) , 之 後 再 加 入 濃 縮 後 培 養 過 pLNCX2-BCC 細胞或 CXCR4-BCC 細胞的培養液(250 µl)(圖 6)。 培養 8 小時後, 觀察其管狀形成的長度 (30)。. 31.
(33) 癌細胞異種移植試驗. 以胰蛋白水解酵素將 BCC-1/KMC、pLNCX2-BCC 與 CXCR4-BCC 細 胞脫離培養瓶壁後,以 PBS 漂洗。 將其以 5×105 細胞/100 µl PBS 濃度分別皮下注射進入三組裸鼠 (day 0, 三組, n=5)。 另外一組 動物實驗是在裸鼠的 CXCR4-BCC 細胞注射部位,再以每天ㄧ次的方 式,施打 CXCR4-blocking peptide T22 (2 µg/鼠 ? 日劑量; day 0 – day 5)。 腫瘤生長的大小程度以卡尺測量其長、寬與厚度。腫瘤大 小測量公式為長度×寬度×厚度×0。5236(17)。. 32.
(34) 第三節 資料處理及分析方法. 實驗術據以平均值 ± 標準差呈現 。 體外實驗的數據以 Student’s t tests (two-sided, parametric)分析, Wilcoxon test 則用於分析體內腫瘤生長程度。 以 P < 0.05 決定統計上的差異。 同性實驗皆重複三次以上。. 33.
(35) 第五章 研究結果 人 類 表 皮 基 底 細 胞 表 現 CXCR4 受 體 蛋 白. 為了證明是否趨化激素受體影響表皮基底細胞的腫瘤發生, 本 次實驗先以 qPCR 偵測 CCR1−CCR10 與 CXCR1−CXCR6 趨化激素受體在 BCC-1/KMC 細胞株中的表現量。 如圖 1A 顯示,比起其它趨化激素 受體,CXCR4 受體蛋白在 BCC -1/KMC 細胞株中的表現量是 CCR10 受 體蛋白表現量的 72 倍。另外,BCC-1/KMC 細胞則未被偵測到有 CXCL12 的表現 (圖 1A)。我們比較 BCC-1/KMC 細胞與其它皮膚癌細胞株 (melanoma, A2058 與 squamous cell carcinoma, A431)或正常表 皮細胞(角質細胞、黑色素細胞與纖維母細胞) 的 CXCR4 表現量,發 現 BCC-1/KMC 細胞比起其他細胞有較高的 CXCR4 表現量 (~500 倍 ~3000 倍)(圖 1B)。 此外,我們也以免疫墨點法試驗偵測 BCC-1/KMC 細胞中 CXCR4 受體蛋白的表現(101 kDa, 圖 1C) (31)。 ? 了偵測 CXCR4 受體蛋白是否會在表皮基底腫瘤組織細胞中表現, 我們將表皮基底組織樣品 進行免疫化學法試驗。 以 Anti-CXCR4 單 株抗體結合 DAB 染劑的方式 (呈褐色) 染色表皮基底組織細胞樣品 34.
(36) (7 in 12),如圖 1D。 以 isotype IgG 所做的對照組則無呈色反應。 同時我們也使用 CXCR4 anti-sense oligonucleotide 探針進行原位 雜交實驗證明表皮基底腫瘤細胞有 CXCR4 mRNA 的表現(圖 1E)。. 35.
(37) Relative CXCR4 expression. B. D BCC 1.6. MM SCC KC. 3169. 1000. 100. 10 6.4. 1.8 1. 1 FC. E. 36. 10000. C. CX CL 12. CX CR 6. CX CR 5. CX CR 4. CX CR 3. CX CR 2. CX CR 1. CC R1 0. CC R9. CC R8. CC R7. CC R6. CC R5. CC R4. CC R3. CC R2. CC R1. Relative expression by BCC. A. 6000. 4000. 2000. 0.
(38) 圖 1。 表皮基底細胞可表現 CXCR4 受體蛋白。 圖 A, 以定量 聚 合 酵 素 反 應分 析 BCC-1/KMC 細 胞 中 趨 化 激 素 受 體 蛋 白 與 CXCL12 趨化激素蛋白的表現量。 圖 B,以定量聚合酵素反應偵測 表皮基底細胞,黑色素細胞 (MM), 鱗狀細胞瘤 (SCC),纖維細胞 (FB) 與角質細胞 (KC)中 CXCR4 受體蛋白的相對表現量。圖 C,以 免疫墨點法偵測人類 CXCR4 (hCXCR4)受體蛋白之表現。圖 D,以 antihuman CXCR4DAB 染劑染色人類表皮基底腫瘤組織細胞。圖 E, 以原位雜交法偵測表皮基底細胞內 CXCR4 mRNA 的表現量。. 37.
(39) 以 逆 轉 錄 病 毒 載 體 轉 染 CXCR4 基 因 至 表 皮 基 底 細 胞 。. 為了證明單獨的 CXCR4 受體蛋白可以促進表皮基底細胞的腫瘤 化 , 我們使用逆轉錄病毒載體, pLNCX2, 將 CXCR4 基因轉染至 BCC-1/KMC 細胞中。之後以含有 FITC-conjugated anti-human CXCR4 單株抗體與 anti-FITC 之磁力珠(Miltenyi Biotec, Auburn, CA) 分離取得同質性高的 CXCR4-BCC 細胞(圖 2A)。經過 CXCR4 基因轉染 後的 CXCR4-BCC 細胞再以 qPCR 偵測後,發現比起野生型 BCC-1/KMC 與 pLNCX2-BCC 細胞有較高的 CXCR4 表現量,如圖 2B。鈣離子灌流 實驗方式可參考文獻 28,此試驗是為了證明 CXCR4-BCC 細胞所產生 之 CXCR4 受體蛋白具有功能性,如圖 2C 所示。. 38.
(40) A. B C. 1.4. 60.1. 1.3. 60. Ratio. Relative CXCR4 expression. 80. 40. CXCL12. 1.2 1.1 1. 20. 1. 0.9. 3.6. 0. 100. 200. 300. 0. BCC. CXCR4-BCC pLNCX2-BCC. Time (s). 圖 2。 將 CXCR4 基因經由反轉錄病毒轉染至 BCC-1/KMC 表皮基底 細胞株。 圖 A, CXCR4 基因轉染之表皮基底細胞經磁力株篩選後, 再以 FITC 螢光染劑標定的 anti-human CXCR4 (hCXCR4)單株抗體作 染色。 FSC, 前向散射。 圖 B, 以定量聚合酵素反應分析 BCC-1/KMC (表皮基底細胞), CXCR4-BCC 細胞與 pLNCX2-BCC 細胞內之 CXCR4 相 對表現量。圖 C,鈣離子灌流試驗,以 fura-2-acetoxymethyl ester 標定 CXCR4-BCC 細胞並以含 500ng CXCL12/ml 之鈣緩衝液處理細胞 (45)。. 39. 400.
(41) 體外實驗證明在低血清濃度下,CXCL12 受體蛋白促 CXCR4-BCC 細胞 增殖. 為了證明 CXCR4 受體蛋白的表現能夠促使表皮基底細胞腫瘤的 生長, 將 pLNCX2-BCC 與 CXCR4-BCC 細胞培養於正常濃度(10% FCS) 或低濃度(0。5% FCS)血清下,觀察 CXCL12 趨化激素的存在與缺乏下 所造成腫瘤細胞的增殖現象。在正常血清濃度下,不論有無 CXCL12 趨化激素的存在,CXCR4-BCC 與 pLNCX2-BCC 細胞都有相同的增殖率。 然而,比起只有 PBS 處理的對照組,以 CXCL12 處理的實驗組有明顯 CXCR4-BCC 細胞增殖的現象 (24 小時 ~1。2 倍, P=0。027; 48 小時, ~2。2 倍, P=0。004) (圖 3)。 以 CXCL12 處理所造成的生長現象 會受到 anti-CXCR4 單株抗體的處理而消減 (300 µg/ml) (圖 3)。 在 低血清濃度下 CXCL12 並不會明顯地促使 pLNCX2-BCC 細胞的增殖 (P=0。4, data not shown)。 因此, 表皮基底腫瘤細胞處於低營 養供給的情況下,可藉 CXCL12 與 CECR4 受體蛋白所產生的交互作用 而促使其生長。. 40.
(42) 24 h. Cell number. 12000. 48 h ∗. 10000 8000 6000 ♦. 4000 2000 0 PBS. CXCL12. CXCL12 + anti-CXCR4. CXCL12 + mouse IgG2B. 圖 3。 經體外實驗證明在低血清濃度培養下,CXCL12 促使 CXCR4-BCC 細胞的增殖。在 anti-CXCR4 單株抗體或 isotype 單株抗體存在下, CXCR4-BCC 細胞(3 × 104 細胞/孔;三重複) 培養於 0.5% 小牛血清並 處理 500 ng/ml 的 CXCL12。在指定的時間點內以 WST-1 試驗方法偵 測計數存活的細胞。 At 24 h: ♦, P=0.027 versus PBS-treated 細胞。 At 48 h: ∗, P= 0.004. (all versus PBS-treated 細胞)。. 41.
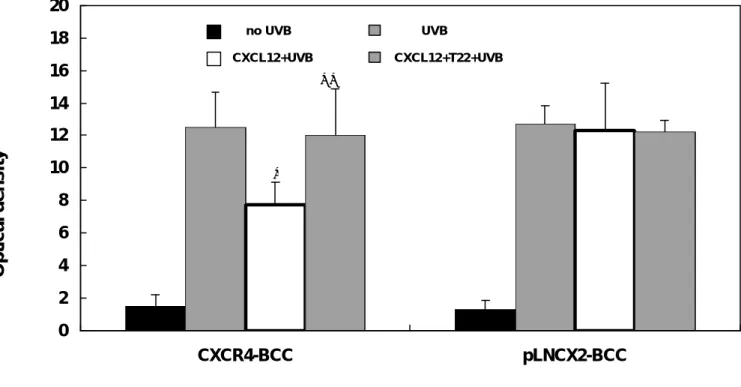
(43) 體外實驗證明在 UVB 的照射下, CXCL12/CXCR4 二者的交互作用可以 防 止 CXCR4-BCC 細 胞 進 行 細 胞 凋 亡. 為了證明 CXCR4 受體蛋白所引起的訊息傳導可以防止表皮基底 細胞走向程序性死亡,我們將 pLNCX2-BCC 細胞 與 CXCR4-BCC 細胞 暴露於 UVB 的照射以誘發其細胞凋亡。 如圖 4,在有 CXCL12 的前 處理下, 大約有 38%(P=0。04)的 CXCR4-BCC 細胞可免於細胞凋亡, 然而,此現象卻受到 CXCR4-blocking T22 peptide 的阻斷。受 UVB 照射之對照組 pLNCX2-BCC 細胞則不受 CXCL12 趨化激素或 T22 的影響 (圖 4)。 因此,CXCL12/CXCR4 二者的交互作用可以防止表皮基底細 胞走向凋亡。. 42.
(44) 20 18. no UVB. UVB. CXCL12+UVB. 16. CXCL12+T22+UVB ∗∗. 14. Optical density. 12 10. ∗. 8 6 4 2 0 CXCR4-BCC. pLNCX2-BCC. 圖 4。 經 體 外 實 驗 證 明CXCR4/CXCL12趨化性的交互作用可阻斷 CXCR4-BCC細胞的凋亡。 在T22存在或缺乏下,以500 ng/ml CXCL12 處理CXCR4-BCC細胞與pLNCX2-BCC細胞。之後在螢光染劑(AMC)釋放 460 nm波長下以光學密度偵測caspase-3的活性。 CXCR4-BCC細胞: *, P=0.04; **, P=0。86, versus UVB-irradiated cells without CXCL12 and T22.. 43.
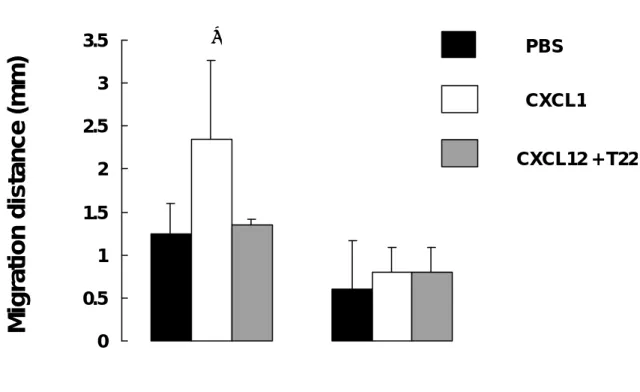
(45) CXCR4-表 皮 基 底 細 胞 移 行 與 侵 犯 之 體 外 實 驗. 為了證明 CXCR4 蛋白的表現會引起表皮基底細胞的遷移,以二維 遷移試驗偵測 CXCR4-BCC 細胞在有無 CXCL12 趨化激素存在下的移動 現象。 如圖 5, 在 CXCL12 趨化激素存在下,有 88% CXCR4-BCC 細 胞會產生移行(P<0。05)。以 T22 與 CXCL12 共同處理下,產生移行 CXCR4-BCC 細胞減少。 CXCL12 的處理並不會明顯地增加 pLNCX2-BCC 細胞的移行(P=0.7)。另外,在癌細胞侵犯試驗下,並未觀察到受 CXCL12 處理的 CXCR4-表皮基底細胞、pLNCX2-表皮基底細胞與未受 CXCL12 處理的對照組細胞有任何結果上的差異。. 44.
(46) Migration distance (mm). 3.5. ∗. PBS. 3 CXCL1 2.5 CXCL12 + T22. 2 1.5 1 0.5 0. 圖 5。 經體外實驗證明CXCR4/CXCL12趨化性的交互作用促進表皮基 底細胞的移行。 在有CXCL12的存在或缺乏下,以二維移行試驗觀察 CXCR4-BCC與 pLNCX2-BCC細胞所產生的細胞遷移。並依據實驗組或對 照組來判別是否添加T22。∗ P<0.05, versus PBS-treated cells.. 45.
(47) 體外實驗顯示 CXCL12/CXCR4 路徑可促使 CXCR4-BCC 細胞進行血管新 生. 為了證明 CXCR4 的表現能夠促使腫瘤細胞周邊血管新生,將培 養 pLNCX2-BCC 細胞或 CXCR4-BCC 細胞的培養液濃縮並處理人類內皮 細胞。與對照組比較下(沒有 CXCL12 趨化激素的處理) (圖 6A 與 D; ~2。19 倍, P<0。05),有添加 CXCL12 趨化激素的 CXCR4-BCC 細胞 培養液有明顯地促使 HMEC-1 細胞管狀發生現象(圖 6B 與 D,) ,然 而,T22 peptide 的添加則可以拮抗中斷此現象,如圖 6C 與 D。無 添 加 CXCL12 趨化激素的 pLNCX2-BCC 細 胞培養液則不具有促使 HMEC-1 細胞管狀發生(圖 6D, P=0。086)。因此,CXCR4-BCC 細胞 在 CXCR4/CXCL12 二者交互作用後能夠促使腫瘤細胞產生引發周邊血 管新生的可溶性物質。為了證明 CXCR4/CXCL12 傳導路徑可以促進某 些血管新生因子的表現,我們將受到 CXCL12 前處理的 pLNCX2-BCC 與 CXCR4-BCC 細胞做 VEGF 的 qPCR 分析。 如圖 6E, 24 小時 後,VEGF 的表現量在 CXCR4-BCC 細胞與 pLNCX2-BCC 細胞皆有上升 (CXCR4-BCC 細胞, ~2.5 倍; pLNCX2-BCC 細胞, ~1.64 倍)。然而,CXCL12 的 處理並不能促使其它血管新生因子 (VEGF-B, VEGF-C, VEGF-D 與 46.
(48) COX-2, data not shown)的表現。 此外,VEGF 表現量增加的現象 可在添加 T22 peptide 後而被消除 (圖 6E),此結果可被認為表皮基 底細胞中 VEGF 的表現對 CXCR4R/CXCL12 所產生的訊息傳導有依賴 性。. 47.
(49) A. B. C. D. PBS. Tubule formation (mm). 1800 1600. ∗. CXCL12. 1400 1200. CXCL12 + T22. 1000. ∗∗. 800 600 400 200 0. CXCR4-BCC. pLNCX2-BCC. CXCL12 stimulation enhance the growth of human endothelial cells. 48.
(50) E 7.00 6.00. Relative expression. PBS 5.00. CXCL1 4.00. CXCL12 + T22. 3.00 2.00 1.00 0.00 1. 2. CXCR4-BCC. pLNCX2-BCC. CXCL12 stimulation enhance VEGF expression in CXCR4-BCC. 圖 6。 體外實驗證明CXCR4 蛋白的表現促表皮基底細胞血管新生。 HMEC-1 細胞接種於以 Matrigel覆蓋的培養盤上,之後再以培養過 CXCR4-BCC細胞之培養液處理(圖A-C,顯示HMEC-1 細胞的管狀發生) 與 pLNCX2-表皮基底細胞in various conditions。 A, 培養過 CXCR4-BCC細胞之培養液處理HMEC-1 細胞但無添加CXCL12。 B, 培 養過CXCR4-BCC細胞之培養液處理HMEC-1 細胞且以CXCL12做前處理。 C, 培養過CXCR4-BCC細胞之培養液處理HMEC-1 細胞且以CXCL12與 T22做前處理。 D, HMEC-1 細胞在培養8小時候有管狀形成。∗ P < 0.05 49.
(51) versus HMEC-1 cells incubated with control (PBS only) conditioned medium. ∗∗ P= 0.086 圖 E,細胞在有 CXCL12 的存在或缺乏下,以 RT-PCR 定 量 CXCR4-BCC 與 pLNCX2-BCC 細胞內 VEGF 蛋白的表現量。並依據實 驗組或對照組來判別是否添加 T22。. 50.
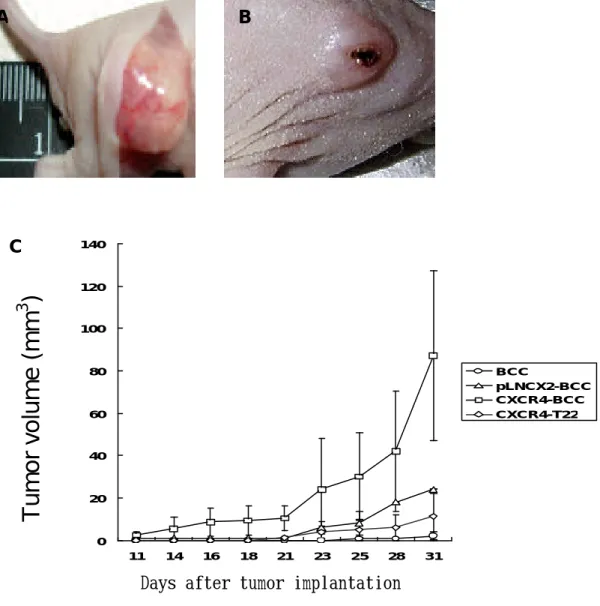
(52) 體內實驗證明表皮基底細胞經 CXCR4 基因轉染後,其表現之 CXCR4 受 體蛋白能夠促表皮基底細胞腫瘤化. 為了證明是否 CXCR4 的表現可以促使表皮基底細胞腫瘤的發 生,我們將 BCC-1/KMC、 pLNCX2-BCC 與 CXCR4-BCC 細胞皮下注射進 入裸鼠。 在第 31 天觀察期結束後,以 CXCR4-BCC 細胞注射之裸鼠比 注射 pLNXC2-BCC 細胞的裸鼠具有高腫瘤負荷 (~3。6-fold, P< 0。 05), 如圖 7A。 當注射 CXCR4-BCC 細胞近入裸鼠體內後,發現有不 正常的血管新生 (圖 7B) 與潰瘍的現象 (圖 7C), 如同人類的表皮 基底細胞腫瘤。 而且, CXCR4-BCC 腫瘤細胞的生長可受到連續 T22 的施打注射而受到抑制(圖 7A),且透過檢視腫瘤小鼠的內臟器官(肺 臟,肝臟與腸胃道)與淋巴結, 並未發現有任何轉移。這些結果指出 CXCR4 的表現量能夠促使表皮基底腫瘤細胞的生長及其週邊血管的 新生。. 51.
(53) A. B. C. 140. Tumor volume (mm3). 120. 100. 80. BCC pLNCX2-BCC CXCR4-BCC CXCR4-T22. 60. 40. 20. 0 11. 14. 16. 18. 21. 23. 25. 28. 31. Days after tumor implantation. 圖 7。 體內試驗證明 CXCR4 受體蛋白的表現促使表皮基底細胞的癌 化發生。 將 BCC-1/KMC, pLNCX2-BCC 與 CXCR4-BCC 三種細胞以 5×105 /100 µl PBS 的密度皮下注射進入裸鼠。(重複實驗三次, n=5)。 圖 A-B, 體內實驗顯示 CXCR4-BCC 腫瘤細胞造成血管新生形成過程(A) 與腫瘤發生部位潰瘍的情形(B)。 圖 C,注射表皮基底腫瘤細胞後之 腫瘤負荷。P < 0.05. 52.
(54) 第六章 討論 第一節 結果討論. 表皮基底腫瘤細胞生長的分子機制仍有許多未明。 最近的研究 報告顯示腫瘤細胞的生長與轉移或許與趨化激素與其受體有著密切 的關聯。於是, 我們假設某種趨化激素受體可能與表皮基底細胞腫 瘤化有關。經由實驗,我們發現人類表皮基底細胞株擁有高度表現 CXCR4受體蛋白的能力。此外,我們認為只有在表皮基底腫瘤組織的 某一區塊能夠表現CXCR4受体蛋白,這說明腫瘤組織內細胞的異質 性。我們的假設是人類皮膚所具有之樹突細胞與內皮細胞(32)能夠表 現並分泌CXCL12趨化激素,在以此旁分泌的方式下與BCC-1/KMC 細胞 表面上的CXCR4趨化激素受體蛋白產生訊息傳遞上的交互作用。 藉由逆轉錄病毒載體轉染的方式,我們證明表皮基底細胞有高度的 CXCR4表現並歸納出其促使表皮基底細胞腫瘤化的一些機制。首先, 在低血清濃度下,CXCR4 的表現可以促使表皮基底細胞的增殖,相關 的其他文獻也證實CXCR4與一些正常或腫瘤細胞的生長有關(9, 13, 33, 34)。 此現象說明了CXCR4/CCL12 訊息傳導能夠活化另外二個 53.
(55) 與細胞生長有關的訊息傳遞路徑,phosphatidylinostiol 3-kinase (PI3K). 與. mitogen-activated. protein. kinase. (MAPK). pathway(35-37)。 此外, 我們也證實CXCR4 的表現量可以防止UVB 照射後的腫瘤細胞走向細胞凋亡。此結果與其它研究報告一致(13, 33)。在生長因子缺乏下,CXCR4/CXCL12的訊息傳遞作用作可以使腫 瘤細胞免於走向死亡且Akt (即protein kinase B) 的生化路徑也影 響著受CXCR4 調控的細胞 (13)。 最近一些體外實驗指出CXCR4/CXCL12的訊息傳導作用可能對腫瘤 引起之血管新生有關 (11)。 內皮細胞本身可以同時表現CXCR4受體 蛋白與CXCL12趨化激素(32),當CXCR4受體蛋白受體與CXCL12結合 後,不只引起攜帶CXCR4受體蛋白的內皮細胞產生趨化反應,同時也 引起VEGF的釋放(38)。 事實上,由內皮細胞所釋放的VEGF,除了可 以誘發血管新生外,同時也可以回饋促進內皮細胞本身表達CXCR4 與 CXCL12的表現(39-41),因此造成一種放大的生物機制迴圈反應。然 而, 我們想釐清的是,在腫瘤與內皮細胞基質中存在何種訊息上的 交互作用,而腫瘤細胞是否藉由趨化激素的表現而影響內皮細胞進行 血管的新生。事實上,我們以體外實驗證明腫瘤細胞中CXCR4的表現 與 血 管 新 生 有 關 。 除 此 之 外 , 我 們 更 進 ㄧ 步 發 現腫 瘤 細 胞 中 CXCR4/CXCL12 的訊號傳遞路徑能引發VEGF的表現,並因VEGF的釋放 54.
(56) 而引起血管新生。此外, 我們藉由CXCR4-BCC細胞異種移植進入裸鼠 的體內實驗,更清楚觀察到血管新生的現象(圖7B)。因此,腫瘤細胞 內CXCR4的表現及訊息傳遞可與內皮細胞產生訊息上的交換而促使血 管新生。 CXCR4受體蛋白是否影響腫瘤遷移與浸潤仍有爭議。 Taichman 等人 (10) 研究揭示前列腺癌細胞株能夠表現CXCR4受體蛋白並透過 此蛋白促使此類腫瘤細胞的浸潤以及內皮細胞的遷移。相反地, Zhou 等人 (13) 則認為CXCL12趨化激素的存在下並不改變帶有CXCR4蛋白 神經膠質瘤細胞的移動與浸潤。本次研究結果也顯示,表皮基底細胞 對於CXCR4受體蛋白是否造成表皮基底細胞的遷移或浸潤,其結果有 著相當大的差異及矛盾。二維遷移實驗的結果雖然顯示CXCR4的表現 可以造成表皮基底細胞的遷移,但在浸潤試驗中卻顯示CXCR4-BCC細 胞在浸潤的程度上並不具有與對照組有太大的區別。雖然如此,我們 假設主要的原因,可能是由於不同腫瘤細胞彼此擁有的異質性,故 CXCR4受體蛋白也以不同的方式影響不同腫瘤的發生。 有待釐清的是,腫瘤細胞如何改變自己產生CXCR4蛋白以促進本身 的腫瘤化。最近, Staller 等人 (42) 在腎清晰腫瘤細胞上(renal clear cell carcinoma) 證 明 一 種 內 在 腫 瘤 抑 制 蛋 白 ,. von. Hippel-Lindau protein(VHL), 能夠藉由其本身的去活化而使得活 55.
(57) 化一種於大部份腫瘤細胞皆會大量表現並改變腫瘤細胞生長的轉錄 因子,hypoxia-inducible factor (HIF),進而促使CXCR4受體蛋白 的表現(43)。同樣地,屬於Sonic hedgehog 訊息傳遞路徑中的PTCH 腫瘤抑制基因的變異也會影響表皮基底細胞病理發生的過程(18), 事實上,CXCR4/CXCL12 與 Sonic hedgehog生化路徑同樣不只是影響 胚胎形成(7, 20),也與表皮基底腫瘤細胞的發生有關(18, 19), 此 也說明Sonic hedgehog 與 CXCR4彼此的訊息傳導路徑有著某種程度 上的關聯。 此外,先前文獻報告指出在腎清晰腫瘤細胞內, PI3K 與 MAPK 訊息路徑的活化可使VHL蛋白去活而致HIF蛋白的合成,最後造 成CXCR4 表現量的上升(42) (41, 44),然而CXCR4 表現量的上升又 可誘發PI3K 與 MAPK 訊息傳遞 (35-37)。如此週而復始的現象,我 們稱為一種放大的生物機制正迴饋反應(35-37, 41, 42, 44) 本次研究的結果,認為表皮基底細胞癌也屬於受到 CXCR4 受體 蛋白所影響的腫瘤之一, 並證明 CXCR4 在人類表皮基底細胞中有高 度的表現;在體外實驗方面,CXCR4 的表現能促使表皮基底細胞增 殖,阻止細胞凋亡,並影響其遷移與促進不正常的血管新生, 而體 內實驗更顯示攜帶 CXCR4 受體白的表皮基底細胞能夠產生腫瘤化現 象。 此外,經體內試驗發現,CXCR4 受體蛋白訊息傳遞的受阻可使 表皮基底腫瘤細胞消失,如此阻斷 CXCR4 受體蛋白的活化或許可成為 56.
(58) 未來治療此類癌症的方向之一。. 57.
(59) 第二節 研究限制. 1. 本研究著重在腫瘤生長本身方面,對於他與人體免疫力之間的交 互作用則無太多著墨。 2. 因此,對於巨噬細胞在基底細胞癌的角色,T 細胞,NK 細胞 (natural killer cells),樹突細胞(dendritic cells)等浸 潤細胞對基底細胞癌的影響須進一步評估。 3. 本研究也無法確立為何臨床上的基底細胞癌較不易遠處轉移。此 點或許與對白血球等細胞的趨附能力及免疫力有關。 4. 雖然,有無 CXCR4 似乎對基底細胞癌的侵犯能力並無明顯不同, 但因只是體外試驗,CXCR4 在基底細胞癌的侵犯能力中的角色還 須進一步釐清。. 58.
(60) 第七章 結論與建議 第一節 結論. 趨化激素透過各種不同的機轉調控腫瘤的形成:(1)直接控制腫 瘤之生長;(2)間接透過免疫反應來調整 在本研究中,我們從體外實驗中確證基底細胞癌中有趨化激素受體 CXCR4 的存在,且它的表現促進基底細胞癌的增殖、抗凋亡作用、血 管新生作用,細胞移行能力。從裸鼠的體內實驗中,我們觀察到了腫 瘤的增長,血管增生受到趨化激素受體 CXCR4 的高度表現而增進。 因此,若能找到一個有效又安全的方法可以拮抗趨化激素受體 CXCR4 的表現,相信可以達到極佳的治療癌症的效果。例如:我們可 以利用趨化激素的調節免疫反應的能力,來促進樹突細胞或作用 T 細 胞的活化,進而抑制趨化激素受體的表現,達成消滅癌細胞的能力。 類似癌症疫苗般,藉由自體的免疫力達成控制癌症腫瘤的生長。 所以,若能更進一步的了解趨化激素如何透過免疫反應來調節腫瘤 細胞生長的機制,相信對於癌症的治療會有更大的突破。. 59.
(61) 第二節 建議. CXCR4 在基底細胞癌的腫瘤生長方面的角色,經由本實驗,已可 獲得證實,但基底細胞癌的轉移及侵犯方面,似乎無法由內本實驗獲 得相關結論。 因此,下一步的實驗方面,或可著重在 CXCR4 在基底細胞癌中和 皮膚周遭組織及免疫細胞之間的相對關係。 (1). 巨噬細胞趨附是否會因基底細胞癌中 CXCR4 或其他組織. 分泌 CXCL12 之影響,而產生生長因子,血管生成因子或基 質間質蛋白? (matrix metalloprotein)等。進而有所增 進或抑制。 (2). 樹突細胞的趨化作用,在基底細胞癌中的角色為何?樹突. 細胞(dendritic cells)的趨化作用與 CXCR4 或 CXCL12 是 否有關? 基底細胞癌之所以易侵犯局部組織而不易轉移,樹突細胞在 此是否扮演關鍵角色,都值得我們更深入探討。 所以,我們的下一個方向似乎可著重在基底細胞癌、趨化激 素及免疫反應三者之間的相對應關係做進一步研究。 60.
(62) 參考文獻及附錄. 1.. Baggiolini, M. Chemokines in pathology and medicine. J Intern Med, 250: 91-104, 2001.. 2.. Rossi, D. and Zlotnik, A. The biology of chemokines and their receptors. Annu Rev Immunol, 18: 217-242, 2000.. 3.. Bleul, C. C., Wu, L., Hoxie, J. A., Springer, T. A., and Mackay, C. R. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes. Proc Natl Acad Sci U S A, 94: 1925-1930, 1997.. 4.. Gupta, S. K., Lysko, P. G., Pillarisetti, K., Ohlstein, E., and Stadel, J. M. Chemokine receptors in human endothelial cells. Functional expression of CXCR4 and its transcriptional regulation by inflammatory cytokines. J Biol Chem, 273: 4282-4287, 1998.. 5.. Hesselgesser, J., Halks-Miller, M., DelVecchio, V., Peiper, S. C., Hoxie, J., Kolson, D. L., Taub, D., and Horuk, R. CD4-independent association between HIV-1 gp120 and CXCR4: functional chemokine receptors are expressed in human neurons. Curr Biol, 7: 112-121,. 61.
(63) 1997. 6.. Murdoch, C. CXCR4: chemokine receptor extraordinaire. Immunol Rev, 177: 175-184, 2000.. 7.. Horuk, R. Chemokine receptors. Cytokine Growth Factor Rev, 12: 313-335, 2001.. 8.. Muller, A., Homey, B., Soto, H., Ge, N., Catron, D., Buchanan, M. E., McClanahan, T., Murphy, E., Yuan, W., Wagner, S. N., Barrera, J. L., Mohar, A., Verastegui, E., and Zlotnik, A. Involvement of chemokine receptors in breast cancer metastasis. Nature, 410: 50-56, 2001.. 9.. Murakami, T., Maki, W., Cardones, A. R., Fang, H., Tun Kyi, A., Nestle, F. O., and Hwang, S. T. Expression of CXC chemokine receptor-4 enhances the pulmonary metastatic potential of murine B16 melanoma cells. Cancer Res, 62: 7328-7334, 2002.. 10.. Taichman, R. S., Cooper, C., Keller, E. T., Pienta, K. J., Taichman, N. S., and McCauley, L. K. Use of the stromal cell-derived factor-1/CXCR4 pathway in prostate cancer metastasis to bone. Cancer Res, 62: 1832-1837, 2002.. 11.. Koshiba, T., Hosotani, R., Miyamoto, Y., Ida, J., Tsuji, S.,. 62.
(64) Nakajima, S., Kawaguchi, M., Kobayashi, H., Doi, R., Hori, T., Fujii, N., and Imamura, M. Expression of stromal cell-derived factor 1 and CXCR4 ligand receptor system in pancreatic cancer: a possible role for tumor progression. Clin Cancer Res, 6: 3530-3535, 2000. 12.. Burger, M., Glodek, A., Hartmann, T., Schmitt-Graff, A., Silberstein, L. E., Fujii, N., Kipps, T. J., and Burger, J. A. Functional expression of CXCR4 (CD184) on small-cell lung cancer cells mediates migration, integrin activation, and adhesion to stromal cells. Oncogene, 22: 8093-8101, 2003.. 13.. Zhou, Y., Larsen, P. H., Hao, C., and Yong, V. W. CXCR4 is a major chemokine receptor on glioma cells and mediates their survival. J Biol Chem, 277: 49481-49487, 2002.. 14.. Miller, D. L. and Weinstock, M. A. Nonmelanoma skin cancer in the United States: incidence. J Am Acad Dermatol, 30: 774-778, 1994.. 15.. Gloster, H. M., Jr. and Brodland, D. G. The epidemiology of skin cancer. Dermatol Surg, 22: 217-226, 1996.. 16.. Gailani, M. R., Leffell, D. J., Ziegler, A., Gross, E. G., Brash, D. E., and Bale, A. E. Relationship between sunlight exposure and a key genetic alteration in basal cell carcinoma. J Natl Cancer Inst,. 63.
(65) 88: 349-354, 1996. 17.. Jee, S. H., Shen, S. C., Chiu, H. C., Tsai, W. L., and Kuo, M. L. Overexpression of interleukin-6 in human basal cell carcinoma cell lines increases anti-apoptotic activity and tumorigenic potency. Oncogene, 20: 198-208, 2001.. 18.. Unden, A. B., Zaphiropoulos, P. G., Bruce, K., Toftgard, R., and Stahle-Backdahl, M. Human patched (PTCH) mRNA is overexpressed consistently in tumor cells of both familial and sporadic basal cell carcinoma. Cancer Res, 57: 2336-2340, 1997.. 19.. Oro, A. E., Higgins, K. M., Hu, Z., Bonifas, J. M., Epstein, E. H., Jr., and Scott, M. P. Basal cell carcinomas in mice overexpressing sonic hedgehog. Science, 276: 817-821, 1997.. 20.. Taipale, J. and Beachy, P. A. The Hedgehog and Wnt signalling pathways in cancer. Nature, 411: 349-354, 2001.. 21.. Murakami, T., Nakajima, T., Koyanagi, Y., Tachibana, K., Fujii, N., Tamamura, H., Yoshida, N., Waki, M., Matsumoto, A., Yoshie, O., Kishimoto, T., Yamamoto, N., and Nagasawa, T. A small molecule CXCR4 inhibitor that blocks T cell line-tropic HIV-1 infection. J Exp Med, 186: 1389-1393, 1997.. 64.
(66) 22.. Chiang, L. C., Chiang, W., Yu, H. S., Sheu, H. M., and Chen, H. Y. Establishment a nd characterization of a continuous human basal cell carcinoma cell line from facial skin (I) cytological behavior of early passages. Gaoxiong Yi Xue Ke Xue Za Zhi, 10: 170-176, 1994.. 23.. Yu, H. S., Kao, C. H., and Yu, C. L. Coexistence and relationship of antikeratinocyte and antimelanocyte antibodies in patients with non-segmental-type vitiligo. J Invest Dermatol, 100: 823-828, 1993.. 24.. Ades, E. W., Candal, F. J., Swerlick, R. A., George, V. G., Summers, S., Bosse, D. C., and Lawley, T. J. HMEC-1: establishment of an immortalized human microvascular endothelial cell line. J Invest Dermatol, 99: 683-690, 1992.. 25.. Lamouille, S., Mallet, C., Feige, J. J., and Bailly, S. Activin receptor-like kinase 1 is implicated in the maturation phase of angiogenesis. Blood, 100: 4495-4501, 2002.. 26.. Saeki, H., Wu, M. T., Olasz, E., and Hwang, S. T. A migratory population of skin-derived dendritic cells expresses CXCR5, responds to B lymphocyte chemoattractant in vitro, and co-localizes to B cell zones in lymph nodes in vivo [In Process Citation]. Eur. 65.
(67) J Immunol, 30: 2808-2814, 2000. 27.. Hsu, N. Y., Ho, H. C., Chow, K. C., Lin, T. Y., Shih, C. S., Wang, L. S., and Tsai, C. M. Overexpression of dihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancer. Cancer Res, 61: 2727-2731, 2001.. 28.. Wiley, H. E., Gonzalez, E. B., Maki, W., Wu, M. T., and Hwang, S. T. Expression of CC chemokine receptor-7 and regional lymph node metastasis of B16 murine melanoma. J Natl Cancer Inst, 93: 1638-1643, 2001.. 29.. Ishiyama, M., Tominaga, H., Shiga, M., Sasamoto, K., Ohkura, Y., and Ueno, K. A combined assay of cell viability and in vitro cytotoxicity with a highly water-soluble tetrazolium salt, neutral red and crystal violet. Biol Pharm Bull, 19: 1518-1520, 1996.. 30.. Donovan, D., Brown, N. J., Bishop, E. T., and Lewis, C. E. Comparison of three in vitro human 'angiogenesis' assays with capillaries formed in vivo. Angiogenesis, 4: 113-121, 2001.. 31.. Lapham, C. K., Romantseva, T., Petricoin, E., King, L. R., Manischewitz, J., Zaitseva, M. B., and Golding, H. CXCR4 heterogeneity in primary cells: possible role of ubiquitination.. 66.
(68) J Leukoc Biol, 72: 1206-1214, 2002. 32.. Pablos, J. L., Amara, A., Bouloc, A., Santiago, B., Caruz, A., Galindo, M., Delaunay, T., Virelizier, J. L., and Arenzana-Seisdedos, F. Stromal-cell derived factor is expressed by dendritic cells and endothelium in human skin. Am J Pathol, 155: 1577-1586, 1999.. 33.. Broxmeyer, H. E., Kohli, L., Kim, C. H., Lee, Y., Mantel, C., Cooper, S., Hangoc, G., Shaheen, M., Li, X., and Clapp, D. W. Stromal cell-derived factor-1/CXCL12 directly enhances survival/antiapoptosis of myeloid progenitor cells through CXCR4 and G(alpha)i proteins and enhances engraftment of competitive, repopulating stem cells. J Leukoc Biol, 73: 630-638, 2003.. 34.. Barbero, S., Bonavia, R., Bajetto, A., Porcile, C., Pirani, P., Ravetti, J. L., Zona, G. L., Spaziante, R., Florio, T., and Schettini, G. Stromal cell-derived factor 1alpha stimulates human glioblastoma cell growth through the activation of both extracellular signal-regulated kinases 1/2 and Akt. Cancer Res, 63: 1969-1974, 2003.. 35.. Ganju, R. K., Brubaker, S. A., Meyer, J., Dutt, P., Yang, Y., Qin,. 67.
(69) S., Newman, W., and Groopman, J. E. The alpha-chemokine, stromal cell-derived factor-1alpha, binds to the transmembrane G-protein-coupled CXCR-4 receptor and activates multiple signal transduction pathways. J Biol Chem, 273: 23169-23175, 1998. 36.. Vossler, M. R., Yao, H., York, R. D., Pan, M. G., Rim, C. S., and Stork, P. J. cAMP activates MAP kinase and Elk-1 through a B-Rafand Rap1-dependent pathway. Cell, 89: 73-82, 1997.. 37.. Katso, R., Okkenhaug, K., Ahmadi, K., White, S., Timms, J., and Waterfield, M. D. Cellular function of phosphoinositide 3 -kinases: implications for development, homeostasis, and cancer. Annu Rev Cell Dev Biol, 17: 615-675, 2001.. 38.. Mirshahi, F., Pourtau, J., Li, H., Muraine, M., Trochon, V., Legrand, E., Vannier, J., Soria, J., Vasse, M., and Soria, C. SDF-1 activity on microvascular endothelial cells: consequences on angiogenesis in in vitro and in vivo models. Thromb Res, 99: 587-594, 2000.. 39.. Salcedo, R., Wasserman, K., Young, H. A., Grimm, M. C., Howard, O. M., Anver, M. R., Kleinman, H. K., Murphy, W. J., and Oppenheim, J. J. Vascular endothelial growth factor and basic fibroblast growth factor induce expression of CXCR4 on human endothelial cells:. 68.
(70) In vivo neovascularization induced by stromal-derived factor-1alpha. Am J Pathol, 154: 1125-1135, 1999. 40.. Salvucci, O., Yao, L., Villalba, S., Sajewicz, A., Pittaluga, S., and Tosato, G. Regulation of endothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1. Blood, 99: 2703-2711, 2002.. 41.. Bachelder, R. E., Wendt, M. A., and Mercurio, A. M. Vascular endothelial growth factor promotes breast carcinoma invasion in an autocrine manner by regulating the chemokine receptor CXCR4. Cancer Res, 62: 7203-7206, 2002.. 42.. Staller, P., Sulitkova, J., Lisztwan, J., Moch, H., Oakeley, E. J., and Krek, W. Chemokine receptor CXCR4 downregulated by von Hippel-Lindau tumour suppressor pVHL. Nature, 425: 307-311, 2003.. 43.. Semenza, G. L. Targeting HIF-1 for cancer therapy. Nat Rev Cancer, 3: 721-732, 2003.. 44.. Zhong, H., Chiles, K., Feldser, D., Laughner, E., Hanrahan, C., Georgescu, M. M., Simons, J. W., and Semenza, G. L. Modulation of hypoxia-inducible factor 1alpha expression by the epidermal growth factor/phosphatidylinositol 3-kinase/PTEN/AKT/FRAP pathway in. 69.
(71) human prostate cancer cells: implications for tumor angiogenesis and therapeutics. Cancer Res, 60: 1541-1545, 2000. 45.. Tiffany, H. L., Lautens, L. L., Gao, J. L., Pease, J., Locati, M., Combadiere, C., Modi, W., Bonner, T. I., and Murphy, P. M. Identification of CCR8: a human monocyte and thymus receptor for the CC chemokine I-309. J Exp Med, 186: 165-170, 1997.. 70.
(72) 簡單自述 我出生於台中縣豐原市,排行家裡的老四。小學中學畢業於豐原 國小、豐原國中。高中就讀台中一中,大學也是就讀母校中國醫藥學 院。畢業後曾在台北國泰醫院內科短暫行醫,之後於中國醫藥大學附 設醫院皮膚科完成專科醫師訓練。現在為附設醫院皮膚科主治醫師。. 71.
(73)
數據


+3

Outline
相關文件
E-B3 具備藝術創作與欣賞的基 本素養,促進多元感官的發 展,培養生活環境中的美感體 驗。. 音樂與
作」的基本語文素養,並具有 生活所需的基礎數理、肢體及 藝術等符號知能,能以同理心 應用在生活與人際溝通。.. 英語
(3) Day 6, 04Nov2015, subject 5120001:病人轉用 口服抗生素若為 pathogen based therapy,依計 畫要求應先提供細菌培養報告並取得medical
而在後續甲烷化反應試驗方面,以前段經厭氧醱酵產氫後之出流水為進流基 質。在厭氧光合產氫微生物方面,以光合作用產氫細菌中產氫能力最好的菌株 Rhodopseudomonas palustris
the lymphocyte function-associated antigen 1, or LFA-1, was so named because antibodies recognizing this structure interfere with lymphocyte cell adhesion events and
SF11310 張基晟 比較 Ipilimumab 加上 Etoposide/Platinum 與 Etoposide/Platinum
• 是細胞不正常增生,且這些增生的細胞可
當時,許多科學家認為“活”的酵素(如酵母細胞)與“沒 有生命”的胃消化液是不同的東西(生機論),但德國生 理學家Kühne則提出後者是所謂的“酵素”
