國立台灣大學醫學院生物化學暨分子生物學研究所 碩士論文
Graduate Institute of Biochemistry and Molecular Biology College of Medicine
National Taiwan University Master Thesis
鑑定幽門螺旋桿菌中之生物標記分子及 開發胃癌診斷之膜蛋白質晶片
Identification of Potential Biomarkers from Helicobacter pylori and Development of Membrane Protein Chips for
Gastric Cancer Diagnosis
黃翔鈴
Hsiang-Ling Huang
指導教授:周綠蘋 博士 Adviser: Lu-Ping Chow, Ph.D.
中華民國 101 年 7 月
國立臺灣大學碩士學位論文 口試委員會審定書
鑑定幽門螺旋桿菌中之生物標記分子及 開發胃癌診斷之膜蛋白質晶片
Identification of Potential Biomarkers from
Helicobacter pylori and Development of Membrane Protein Chips for Gastric Cancer Diagnosis
本論文係黃翔鈴君(學號 R99442010)在國立臺灣大學生
物化學暨分子生物學研究所完成之碩士學位論文,於民國 101
年 7 月 28 日承下列考試委員審查通過及口試及格,特此證明
謝誌
碩班兩年似乎很快就過去了,但在這期間卻學到了許多這個領域相關的專業知識
與技術,更學到了如何找出問題並且自己試著解決以及磨練了口頭報告的技巧,經歷 了大學以前不曾體驗過的科學家生活,這兩年來真的是一個很棒的經驗!首先一定要感 謝周綠蘋老師的教導,當初剛考進研究所時老師馬上答應收我為指導學生時真的讓我 十分感動,且老師給我的第一印象到現在永遠都是笑容可掬,並且告訴我們每一天都 應該以樂觀的態度去面對每一天的事情,讓我十分欽佩。在研究領域上,每次meeting 時和老師討論都能慢慢引導我們思考研究題目的重要意義與方向,老師在報告與研究 時適時的提醒及建議讓我的碩士生涯獲益良多,感謝周老師,沒有老師,今天就無法 這樣讓我對自己的碩士學位感到自豪!再來還要感謝陳健生老師,謝謝陳老師能讓我在 中央的實驗室完成晶片相關的實驗,並且在實驗及口試時給予許多建議與教導,老師 的鼓勵更讓我感動許久;另外要感謝口委老師黃楓婷老師以及之前 committee 時的繆希 椿老師,謝謝老師們在口試及committee 時的教導與鼓勵,讓我能順利完成碩士學業。
碩士生活中還要感謝的就是 R941 的大家,首先感謝九樓的郁鈴與玟瑾,謝謝妳 們在我碩士生活時與我朝夕相處,有妳們的陪伴讓我的生活每天都很歡樂,還能一起 娛樂、討論實驗、一起努力,希望我們以後還能是永遠的好朋友!!還要感謝函君學姊 和培芬學姊與我討論實驗,和妳們聊天總是讓我開懷大笑。再來要感謝基因體的景光 學長與文雅學姊,謝謝你們在我做clone 時給予我非常多很棒的經驗與建議;感謝成州 學長、昭圻學長及治達學長在實驗上給我的建議,幫助我能順利完成碩士研究,也謝 謝可愛的玫君和漂亮的靖雅兩位學妹,實驗室有妳們就更增添了青春的氣息;感謝家錚 學姊、欣潔學姊、李豪學長及文麒學長,雖然不能常常見到你們,可是每次見面時討 論實驗或暢快聊天都讓我好開心;感謝已畢業的竺陵學姊、韻如與瑩竹學姊,除了與你 們討論實驗以外,我更喜歡大家一起出去喝下午茶!感謝瑞傑學長與書宇學姊,你們認 真做實驗的態度一直是我學習的典範。還要感謝同實驗室的好同學方瑩與宗錡,和你 們討論實驗與互相關心讓我在碩士時不用擔心會忘記做哪些事情,也感謝碩班的好朋 友哈哈與心瀅和其他所有的碩班同學,一起上課聊天辦活動加油打氣讓我知道有許多 人與我一起共同努力著。謝謝所有在學校的你們的陪伴讓我不會忘記這段精彩的生活。
還要感謝中央大學陳健生老師實驗室的柏中學長及實驗室的大家,謝謝你們能耐心的 帶我完成晶片的實驗,更要感謝柏中學長的幫忙,並祝福學長在德國一切順利!
最後要感謝爸媽還有姊姊,支持著我一路念書到現在,尤其每當我做實驗很累時 回到家能看到你們給我的溫暖,就讓我更有動力繼續加油,尤其最後在寫論文時,感 謝爸媽的體諒與幫忙,沒有你們我無法完成這一切;還要感謝小威,從大學到現在你的 支持、陪伴與付出,不是言語就能形容的,我是如此幸運才能遇到你,能彼此互相鼓 勵與體諒幫忙,才能讓我一路走到現在,所有歡笑與淚水都有你在身旁一起分享,尤 其在我忙碌與痛苦時更總是能依靠你,謝謝你。
總之感謝所有的大家,要感謝的人太多了,就謝天吧!
摘要
幽門螺旋桿菌為人類的病原菌,是一種寄生於人類消化道的革蘭氏陰性菌,
並且造成了胃發炎、胃潰瘍、十二指腸潰瘍及胃癌等消化道疾病。有趣的是,臨 床統計結果發現受幽門螺旋桿菌感染的病人其得到十二指腸潰瘍或是胃癌是兩條 分歧的臨床走向。而之前有一些研究顯示幽門螺旋桿菌的膜蛋白對於菌的致病性 及和宿主之間的辨認都扮演了重要的角色,因此我們的研究主要就是希望能從幽 門螺旋桿菌的膜蛋白質中找到胃癌相關的生物標記分子,並期望能發展出一個新 的平台以應用於臨床上,幫助簡易且快速的診斷出受幽門螺旋桿菌感染的胃癌病 患,並能與十二指腸潰瘍病患做區隔。
過去實驗室比較從胃癌及十二指腸潰瘍病人體內分離出來的幽門螺旋桿菌菌 株之膜蛋白質體,我們將目標放在於胃癌菌株中表現量高於十二指腸潰瘍菌株的 蛋白質,並鑑定到了57 個蛋白質。而在我們的實驗中,我們發現了四個膜蛋白具 有潛力能做為生物標記分子,這四個膜蛋白為: outer inflammatory protein (OipA), adherence-associated lipoprotein A (AlpA), sialic acid-binding adhesin (SabA)與 blood group antigen binding adhesin (BabB),這四個蛋白都被報導為黏著因子,與幽門螺 旋桿菌貼附於胃表皮細胞及胃黏膜相關。我們將這四個膜蛋白基因進行質體建構 並且純化出重組蛋白,再利用西方墨點法將重組蛋白與正常人或胃發炎、十二指 腸潰瘍及胃癌病人的血清進行反應,結果顯示OipA, AlpA, SabA 及 BabB 這四個膜 蛋白在胃癌這組中的免疫反應程度都高於十二指腸潰瘍的組別,且有顯著差異,
經過更進一步的統計後發現OipA, SabA 與 BabB 這三個膜蛋白與胃癌病人有高度 相關性。此外,我們還將四個膜蛋白合併在一起進行檢測發現到隨著病人辨認到 的抗原膜蛋白的個數增加,能區分胃癌與十二指腸潰瘍病人的效果更加顯著,勝 算比從只認到一個抗原的0.75 (0.26–2.15),到認到兩個抗原的 3.00 (0.95–9.53),再 到認到三個抗原或是全部抗原都有認到的6.33 (1.29–31.1)。從以上的結果我們認為,
我們所選擇的這四個膜蛋白是具潛力的胃癌相關之幽門螺旋桿菌生物標記分子。
接著,我們為了發展出一個能在臨床上快速又方便的診斷方法,我們選擇以 蛋白質晶片為基礎並測試了適合膜蛋白點印於晶片上的條件,以製做出胃癌相關 之幽門螺旋桿菌膜蛋白質抗原晶片,並且以正常人或十二指腸潰瘍及胃癌的病人 血清與膜蛋白晶片進行反應,我們發現到膜蛋白晶片的結果與前一部份以西方墨 點法檢測的結果相符合,OipA, SabA 及 BabB 確實能夠將胃癌的病人與十二指腸 潰瘍病患及正常人區分開來。未來我們希望能夠利用我們發展的胃癌相關之幽門 螺旋桿菌膜蛋白質抗原晶片去偵測病人血清內多種抗體的反應程度,並期望此平 台及結合複合型生物標記分子的概念能有效應用於臨床診斷上。
關鍵字:幽門螺旋桿菌,胃癌,黏著因子,生物標記分子,膜蛋白質晶片
Abstract
Helicobacter pylori (H. pylori), a human pathogen, is a gram-negative bacterium that colonizes in the host gastroduodenal tract, causing some variant diseases like gastritis, gastric ulcer (GU), duodenal ulcer (DU) and gastric cancer (GC). Interestingly, DU and GC are considered as clinically divergent events. Some previous studies suggested that the outer membrane proteins of H. pylori played important roles in pathogenesis and host recognition. Thus, we aimed to focus on discovering the outer membrane proteins of H.
pylori as GC-related biomarkers, and to develop a new platform which is rapid and easy for diagnosis of screening GC patients from DU patients infected by H. pylori.
In our previous study, we compared the membrane proteome of two clinical H. pylori strains, one strain from GC patients and the other strain from DU patients. We identified 57 proteins which the expression levels of GC strain were higher than that of DU strain. In this study, we found four membrane proteins as potential biomarkers. The four proteins were outer inflammatory protein (OipA), adherence-associated lipoprotein A (AlpA), sialic acid-binding adhesin (SabA) and blood group antigen binding adhesin (BabB).
These candidates were reported to be responsible for the adhesion of H. pylori to the host gastric epithelium and mucosa. We cloned and purified the recombinant membrane proteins. Using immunoblot, the purified proteins were probed with serum samples from gastritis, DU, GC patients and normal controls. The results revealed that OipA, AlpA, SabA and BabB showed significantly higher immuno-reactivity in GC patients than that in DU patients, and OipA, SabA and BabB were strongly related to GC. In addition, we found that the use of multiple antigens improved the discrimination between patients with GC and those with DU as the odds ratios increased from 0.75 (0.26–2.15) for seropositivity for any one antigen alone to 3.00 (0.95–9.53) for two of the four antigens and to 6.33 (1.29–31.1) for three antigens or all four antigens. These results suggested that
these adhesion molecules we selected may be potential H. pylori GC-associated biomarkers.
Moreover, to develop a faster and more convenient platform for diagnosis, we tested the condition for printing the purified recombinant membrane proteins onto glass slides to create GC-related antigen membrane protein chip and the chip was incubated with serum from DU, GC patients and normal controls. The results were significantly consistent with our results of immunoblot that OipA, SabA and BabB were potential biomarkers for specifically distinguishing GC from DU and healthy controls. In the future, we expect to use GC-related antigen membrane protein chip to detect antibody patterns in sera of patients and the membrane protein chip combined with the idea of multiple biomarkers can be used for diagnostic application.
Keywords: Helicobacter pylori, gastric cancer, adhesion molecule, biomarker, membrane protein chip
縮寫
ACN Alp APS Bab β-Me BSA CagA CBB CHAPS DTT DU EDTA Fla GC GS(+) GS(-) HRP IgG IL IPTG Kan kDa
LC-MS/MS LPS
Mw OD600
Oip PAGE PBS PVDF rpm Sab SDS TEMED TFA
acetonitrile
adherence-associated lipoprotein ammonium persulfate
blood group antigen binding β-Mercaptoethanol
bovine serum albumin cytotoxin-associated gene A coomassie brilliant blue
3-((3-cholamidopropyl)dimethylammonio)-1-propane-sulfonate dithiothreitol
duodenal ulcer
ethylenediamineteraacetic acid flagellin
gastric cancer
gastritis (infected by Helicobacter pylori) gastritis (not infected by Helicobacter pylori) horseradish peroxidase
immunoglobulin G interleukin
isopropyl-D-thiogalactopyranoside kanamycin
kilo Dalton(s)
liquid chromatography tandem mass spectrometry lipopolysaccharide
molecular weight
optical density at 600 nm outer inflammatory protein
polyacrylamide gel electrophoresis posphate-buffered saline
polyvinylidene difluoride revolution per minute sodium dodecylsulphate sialic acid-binding
N,N,N’,N”-tetramethylene diamine trifluoroacetic acid
TN TNT Tris Vac
tris and NaCl buffer
tris , NaCl and Tween buffer
tris(hydroxymethyl)-aminomethane vacuolating cytotoxin
目錄
口試委員會審定書 ... i
謝誌 ... ii
摘要 ... iii
Abstract ... v
縮寫 ... vii
第一章 導論 ... 1
第一節 幽門螺旋桿菌的介紹與研究 ... 1
1.1 幽門螺旋桿菌的型態與特徵 ... 1
1.2 幽門螺旋桿菌於流行病學(epidemiology)上之角色 ... 2
1.3 幽門螺旋桿菌所引發之宿主免疫反應與相關疾病 ... 3
1.4 幽門螺旋桿菌之致病因子(virulence factors) ... 7
1.5 幽門螺旋桿菌之膜蛋白(outer membrane protein, OMP)與黏著因子 (adhesin)的重要性 ... 10
第二節生物標記分子(biomarker)與蛋白質微陣列晶片(protein microarray chip)之應用 ... 14
2.1 生物標記分子 ... 14
2.2 蛋白質微陣列晶片 ... 15
第三節 研究動機、目的與策略 ... 17
3.1 研究動機 ... 17
3.2 研究目的 ... 18
3.3 研究策略 ... 18
第二章 實驗材料 ... 20
第一節 幽門螺旋桿菌菌株 ... 20
第二節 血清樣本 ... 20
第三節 大腸桿菌及質體 ... 20
第四節 酵素 ... 20
第五節 抗體 ... 21
第六節 藥品 ... 21
第七節 試劑組 ... 23
第八節 重要儀器 ... 24
第三章 實驗方法 ... 26
第一節 幽門螺旋桿菌標的基因之質體建構(plasmid construction)與重組 膜蛋白質的表現與純化 ... 26
1.1 標的基因之質體建構 ... 26
1.2 重組膜蛋白質之表現與純化 ... 31
1.3 十二烷基磺酸鈉-聚丙烯醯胺膠電泳法 (Sodium dodecyl sulfate -polyacrylamide gel electrophoresis, SDS-PAGE ) ... 34
1.4 以質譜儀鑑定重組膜蛋白質之身份 ... 38
第二節 幽門螺旋桿菌重組膜蛋白質之免疫特性分析 ... 39
2.1 西方墨點法 (Western blotting) ... 39
2.2 統計分析 (Statistical analysis) ... 41
第三節 開發胃癌相關之幽門螺旋桿菌抗原膜蛋白質微陣列晶片 (membrane protein microarray chip) ... 41
3.1 膜蛋白質樣品的處理 (Sample preparation) ... 42
3.2 點印蛋白質樣品至晶片表面 (Printing of protein chips) ... 42
3.3 蛋白質晶片試驗 (Protein chip assay) ... 43
3.4 晶片影像掃描及分析 (Chip image scanning and analysis) ... 44
第四章 實驗結果 ... 46
第一節 幽門螺旋桿菌標的基因之質體建構與重組膜蛋白質的表現與純化 .. 46
1.1 膜蛋白質 OipA, AlpA, SabA 及 BabB 之基因質體建構 ... 46
1.2 膜蛋白質 OipA, AlpA, SabA 及 BabB 的表現與純化 ... 46
第二節 幽門螺旋桿菌重組標的膜蛋白質之免疫特性分析 ... 47
2.1 膜蛋白質 OipA, AlpA, SabA 及 BabB 之免疫原性(immunogenicity) 測試 ... 47
2.2 膜蛋白質 OipA, AlpA, SabA 及 BabB 之免疫特性分析 ... 48
第三節 胃癌相關之幽門螺旋桿菌抗原膜蛋白質微陣列晶片(membrane protein microarray chip)平台的開發 ... 51
3.1 膜蛋白質晶片之最佳化條件測試 ... 51
3.2 胃癌相關之幽門螺旋桿菌抗原膜蛋白質晶片與血清的反應 ... 53
第四節 實驗結果總結 ... 54
第五章 討論 ... 56
第一節 胃癌相關之幽門螺旋桿菌膜蛋白生物標記分子與臨床診斷之意義 .. 56
第二節 胃癌相關之幽門螺旋桿菌抗原膜蛋白晶片與臨床診斷之意義 ... 60
第三節 未來展望 ... 62
第六章 參考文獻 ... 63
圖表 ... 70
附錄 ... 95
第一章 導論
現今許多人有腸胃道的疾病,而對於疾病的發生原因,早期的人們普遍認為 是由於環境因子例如外在壓力、衛生條件及生活方式等所造成,大家並不認為腸 胃疾病會與細菌有關,畢竟像胃部這樣極酸的環境下是很難讓生物生長於其中的,
一直到1982 年,澳洲的 Barry J. Marshall 醫師與病理學家 Robin J. Warren 將幽門 螺旋桿菌(Helicobacter pylori, H. pylori)從胃炎(gastritis)及消化性潰瘍 (peptic ulceration)的病人體內分離出來並且首次成功體外培養,並以自己為對象進行人體 試驗,證實了幽門螺旋桿菌確實會導致胃炎的發生,他們提出了幽門螺旋桿菌的 寄生與胃部疾病有關 [1, 2],之後許多的科學家也慢慢研究出幽門螺旋桿菌致病 的各種機制,從此人們才普遍認同腸胃疾病的發生和幽門螺旋桿菌的感染有著密 不可分的關係。幽門螺旋桿菌的研究可說是消化醫學與臨床研究上的一大進展 [2],Marshall 與 Warren 也因為這些偉大的研究於 2005 年獲得了諾貝爾生理醫學 獎。由於研究幽門螺旋桿菌的致病性對人類十分重要,因此探討幽門螺旋桿菌的 相關問題是生物醫學領域中主要的課題。
第一節 幽門螺旋桿菌的介紹與研究
1.1 幽門螺旋桿菌的型態與特徵
幽門螺旋桿菌是一種生長緩慢呈螺旋狀型態的革蘭氏陰性菌(gram-negative bacteria),喜好生長於微嗜氧 (microaerophilic)的環境下,菌體長約 2~4 μm,寬介 於0.5~1.0 μm 之間,且尾部具有四到六根鞭毛(flagella),它可以適應的 pH 值範圍 非常廣,而最適合生長的pH 值是在 6.9~8 之間 [3],另外有研究指出,幽門螺旋 桿菌即使在pH 2 的極酸性環境下,也可存活約 30 分鐘 [4],但由於幽門螺旋桿 菌可以透過一些獨特的的機制去適應胃中強酸的特殊環境,使它能長久寄生在人 體內胃黏膜的黏液層(mucus gel layer)及胃黏膜的上表皮細胞(superficial
epithelium)之間 [5],也因此能成為人類胃部內唯一的優勢種(single dominant species) [6]。
幽門螺旋桿菌的基因體序列於 1997~1999 年完成,大小為 1.67 Mb,其中 91%
為coding region,可編碼出 1590 個 putative protein,基因體序列中還包含了五個 G+C 特殊組成的區域,當時的科學家更透過與其他已知細菌基因間的比較去推測 出幽門螺旋桿菌其代謝及適應環境的機制及致病性 [7-9]。此外,比較兩個不同臨 床菌株的基因體序列顯示出除了有約6%~7%的蛋白質表現具有菌株特異性(strain specificity)以外,幽門螺旋桿菌具有高度的保守性(conservation) [8]。目前幽門螺 旋桿菌的基因資料庫(database)如 National Center for Biotechnology Information (NCBI)內已提供許多幽門螺旋桿菌基因的資訊。
1.2 幽門螺旋桿菌於流行病學(epidemiology)上之角色
人類是幽門螺旋桿菌主要的宿主,感染通常發生於幼童時期 [10],從近期區 域性的研究發現到學齡前到剛上學的孩童約有 50%~60%的人有被感染幽門螺旋 桿菌,每年更約有 5%新的孩童宿主產生 [11]。幽門螺旋桿菌主要傳染途徑透過 人傳染給人(person to person)的方式,並經口對口(oral-oral)(例如因接觸到嘔吐物 或唾液而接觸到幽門螺旋桿菌)或糞口(fecal-oral)傳染,一般就是透過遭受污染 的食物及水傳染,例如飲用水中被汙染到糞便 [10]。有研究顯示同一家庭裡,若 父母為帶原者,小孩也是帶原者的機率很高,顯示家族間的互相傳染對幽門螺旋 桿菌的傳染是很重要的危險因子(risk factor) [12, 13]。幽門螺旋桿菌感染的主要因 素為環境衛生條件、家族、種族以外 [13],其他影響幽門螺旋桿菌感染的因素有:
宿主血型的不同(ABO blood type)影響細胞表面醣類的修飾、菸酒的使用、飲 食缺乏維生素及微量元素、高鹽和致癌物質的過度攝取以及遺傳因子的不同,都 會影響其感染性,且一旦感染後宿主便會終身帶原除非經過治療 [14]。
幽門螺旋桿菌感染現今受到全球重視,而其盛行率(prevalence)有著地域性的
差別:在未開發國家或開發中國家感染幽門螺旋桿菌的盛行率較已開發國家為高 [10]。至 2011 年之前有關區域性幽門螺旋桿菌盛行率的研究,其中發現到如美國 整個國家因為種族的不同其感染率為7.5%~79.0%,顯示一個國家間感染率也深受 其他因素影響而有極大差異,而非洲國家約41.3%~ 91.3%,其盛行率在全世界國 家當中是偏高的,另外大部分亞洲國家感染率約 50%~70%,但如西伯利亞高達 90%,而馬來西亞只有 19% [13]。之前有文獻指出其感染率在同一個國家內,其 盛行率隨著年齡增加而增加 [15];與社經地位則有成反比的趨勢 [15, 16]。而在 台灣地區根據 2000 年初左右的調查結果,年齡為 1 至 74 歲的人口中感染率為 54.4%,其中男性感染率為 53.7%,女性為 55.2%,男女比例相當,感染率則隨著 年齡增長而增加,而不同種族間也有差異性,例如台灣地區閩南人的感染率較客 家人高出許多,原住民則最高 [17]。近年由於對於幽門螺旋桿菌感染的重視與積 極處理,研究顯示全球盛行率整體而言有下降的趨勢 [13, 18]。
1.3 幽門螺旋桿菌所引發之宿主免疫反應與相關疾病 1.3-1 宿主免疫反應
感染幽門螺旋桿菌後,其菌體與毒性因子和宿主細胞交互作用的結果,普遍 引發宿主細胞一系列免疫反應。首先當幽門螺旋桿菌和胃上皮細胞(epithelial cell) 接觸後,pattern recognition receptors (PRRs)如 toll-like receptors (TLRs)會辨認菌體 上的pathogen-associated molecular patterns (PAMPs),並使細胞產生 NF-κB 所引發 的訊息傳遞並釋放interleukin (IL)-8、tumor necrosis factor (TNF-α)及其他細胞激素
(cytokine) [2, 19],接著一些趨化細胞激素(chemokine)促使原本在血液中的嗜 中性白血球(neutrophils)從血管中進入組織中,連同幽門螺旋桿菌毒性因子的刺激 下,使得單核球細胞(monocyte)、巨噬細胞(macrophage)及樹突細胞(dendritic cell)
聚集 [20],接著它們釋放許多細胞激素如IL-6、IL-10 及 IL-12 等引發後天性免疫 反應(adaptive immunity)如活化 T-helper 1 (Th1) cell 及 B cell。在針對幽門螺旋
桿菌的反應中主要是由 T-helper 1 (Th1)細胞參與在其中,並且釋放 interferon-γ (IFN-γ)、IL-2、IL-10 等細胞激素以強化巨噬細胞及其他 T cell 幫助毒殺幽門螺旋 桿菌 [19, 20]。
然而幽門螺旋桿菌可以透過許多方式避免自己被免疫反應清除掉,例如有研 究指出其脂多醣體(lipopolysaccharide, LPS)上有類似人類blood group antigen的結 構,可能就是幽門螺旋桿菌用來逃過免疫系統攻擊的方式 [21],另外幽門螺旋桿 菌會釋放一些毒性因子幫助抑制T cell的功能而降低免疫反應 [19]。當人體無法把 幽門螺旋桿菌清除乾淨,其所引發的免疫反應會一直持續,且過程中原本宿主用 來抗菌的因子也會造成免疫細胞和胃上皮細胞的損傷使得細胞凋亡(apoptosis),另 外某些病人受感染後會產生自體免疫抗體,攻擊胃中分泌胃酸的細胞parietal cell 的H+/K+-ATPase,造成胃體部的萎縮(corpus/ body region atrophy) [20],這些最終造 成了某些疾病症狀的產生。
1.3-2 幽門螺旋桿菌之相關疾病與分歧的臨床結果
在被幽門螺旋桿菌感染的帶原者中,大多數的帶原者不會有症狀的產生,或 是產生慢性胃炎(chronic gastritis)這類症狀較輕微的疾病,但有約 10%~15%的帶 原者會得到十二指腸潰瘍(duodenal ulcer),另外更有少部份約 2%~5%的帶原者 會得到胃潰瘍(gastric ulcer)並且有可能進一步演變成胃癌 [2, 22]。根據研究,幽 門螺旋桿菌感染胃中不同的位置,會導致不同的病症:通常幽門螺旋桿菌若主要感 染胃竇部(antrum region),促使免疫細胞產生一些細胞激素(如 1.3-1 所述)會使得 荷爾蒙somatostatin 的產量下降以及 gastrin 的上升,而 gastrin 並經由血管運送至 胃體並刺激parietal cell 分泌胃酸,最後造成胃體胃酸過多而容易走向十二指腸潰 瘍;相反的,若幽門螺旋桿菌主要感染在胃體部 (corpus/ body region),免疫細胞 產生的細胞激素會直接抑制parietal cell 產酸的能力而造成胃中 pH 值上升,並造 成parietal cell 的損傷及胃體萎縮,此時則較容易出現胃潰瘍症狀,甚至有機會走
向intestinal metaplasia、dysplasia 而導致胃癌 [2, 20, 23]。
根據International Agency for Research on Cancer (IARC)最近期 2008 年的統計 資料顯示,胃癌在全世界癌症排行榜上是排名第四,而癌症死亡率則排名第二 [24],足以可見研究胃癌相關議題是非常重要的。胃癌中有九成屬於胃腺癌 (adenocarcinomas),其他型如胃淋巴瘤(lymphomas)則佔較少發生率 [25],有許多 原因會造成胃癌的發生,常見的原因如抽菸、飲食、家族遺傳史與生活型態等等 [26],而幽門螺旋桿菌無疑是其中一個十分重要的危險因子,之前的文獻顯示被 幽門螺旋桿菌感染的人比未被感染的人其罹患胃腺癌(非賁門區域(non-cardia regions))的機率高出六~八倍 [27, 28],胃癌的發生與幽門螺旋桿菌的盛行率也有 一定程度的相關 [29],更有研究指出在得胃癌的人當中約有 70%的人是有被感染 幽門螺旋桿菌的 [30],也因此 World Health Organization (WHO)在 1994 年將幽門 螺旋桿菌列為人類第一型致癌因子 [31]。
有趣的是,根據臨床統計的結果發現到,病人得到十二指腸潰瘍或得到胃癌,
這兩條路是分歧的,也就是說臨床上十二指腸潰瘍的病人其終生幾乎不會罹患胃 癌,而胃癌病患也幾乎不會再得到十二指腸潰瘍,這兩種病同時存在於同一個人 身上的機率接近於0 (0.1%~1.7%) [32-34],而這兩種疾病的共同危險因子正是幽 門螺旋桿菌。為什麼會造成這樣的現象目前還無法完全解釋清楚,除了一些說法 像是使用藥物導致胃酸改變的情況而造成最後不同的疾病,或是飲食生活方式不 同以外,科學家們還認為和不同幽門螺旋桿菌菌株(strain)的感染、宿主差異與環 境因子有很大的關係,宿主差異像是宿主本身的基因型、對幽門螺旋桿菌感染後 基因的感受性(genetic susceptibility)等等,而不同幽門螺旋桿菌菌株其基因型態的 表現有一些差異,表現的致病因子也有所不同,過去就有一些報導指出某些基因 的表現會導致胃癌又或某些基因則會導致十二指腸潰瘍,可能因受到帶有不同基 因的菌株感染而走向不同的疾病,另外是否也因不同的菌株喜好感染胃部不同的 地方(胃竇部或胃體,如上面所述)而導致不同疾病,雖然這些因素是在複雜交互
作用的情況下才能演變成最後的結果,但這個議題仍值得深入研究 [32, 35]。
1.3-3 幽門螺旋桿菌之診斷(diagnosis)與治療
欲得知胃中是否有幽門螺旋桿菌之感染,方法大致可分為兩大類:侵襲性與 非侵襲性,侵襲性是利用胃部內視鏡採檢的胃黏膜,做組織學染色、細菌培養或 直接以聚合酶連鎖反應(polymerase chain reaction, PCR)方法偵測幽門螺旋桿菌特 有的基因,及快速尿素酶法 (rapid urease test),此方法有很多種發展的產品,都 是去測胃部檢體是否含有幽門螺旋桿菌重要的酵素--尿素酶,若有則能將尿素分 解成氨氣,並利用酸鹼質的改變偵測。快速尿素酶法不需要慢慢培養細菌,比其 他種侵襲性的方法更為快速也較便宜。非侵襲性檢驗通常較不影響受試者,靈敏 度及特異性也都很高,包含了血清學(serology)、13C 或 14C 標定之二氧化碳尿素 呼吸測試法(urea breath test, UBT)及糞便檢體抗原檢查(Helicobacter pylori stool antigen, HpSA Test) 。 血 清 學 會 利 用 酵 素 連 結 免 疫 吸 附 分 析 ( enzyme-linked immunosorbent assay, ELISA)或西方墨點法 (Western blotting)偵測抗幽門螺旋桿 菌菌體或蛋白的抗體;呼吸測試法原理為若有感染幽門螺旋桿菌,受試者先喝入含 尿素的液體,而菌體的尿素酶可水解尿素產生具標定的二氧化碳,這些二氧化碳 經受試者呼吸道呼出後可偵測到同位素;糞便檢驗利用酵素免疫試驗 (enzymatic immunoassay)檢驗糞便檢體中幽門螺旋桿菌抗原存在與否。UBT 和 HpSA Test 的 好處是他們適用於診斷和評估幽門螺旋桿菌治療情況,能偵測到人體內活菌的存 在,通常在幽門螺旋桿菌療程結束7 天後若這些檢驗仍呈現陽性,則表示治療失 敗,而血清學因為只要感染過幽門螺旋桿菌,其抗體會一直存在,所以此法只能 用來檢測受試者是否有感染過,無法評估治療情況 [36]。
目前對於幽門螺旋桿菌的治療會投以三合一療法:合併鉍鹽(bismith)、抗菌劑 metronidazole與抗生素像是amoxicillin,若是四合一療法會再合併氫離子幫浦抑制 劑(proton pump inhibitors, PPIs),鉍鹽的作用機制目前並不清楚,只知道幽門螺旋
桿菌對此化合物十分敏感。通常藥物治療之後可有效清除幽門螺旋桿菌,但許多 菌已經有抗藥性造成藥物治療無法100%達到效果 [2]。傳統上針對消化性潰瘍的 藥物一類是對抗胃酸分泌,另一類是增加胃黏膜的防禦力,但近年來了解到幽門 螺旋桿菌是造成消化性潰瘍的主因後,治療方向大多轉變成以消除幽門螺旋桿菌 為主,並給予非類固醇類抗發炎藥劑(non-steroidal anti-inflammatory drugs, NSAIDs),
能有效阻止潰瘍的發展 [37]。
1.4 幽門螺旋桿菌之致病因子(virulence factors)
幽門螺旋桿菌的致病因子已有許多相關研究,藉由菌體的結構或分泌的蛋白 等致病因子(virulence factors)可幫助其存活、移動並且生長在胃部的黏膜層,這些 致病因子可幫助研究者了解幽門螺旋桿菌的致病機制、檢驗幽門螺旋桿菌的感染,
甚至可能做為潛在的藥物標的,以下簡述較為主要的致病因子,而其中一項致病 因子「膜蛋白與黏著因子(adhesin)」將在論文 1.5 敘述。
1.4-1 尿素酶(urease)
幽門螺旋桿菌由食道進入胃部時必須存活於強酸環境,菌體可利用氫離子調 控尿素通道(H+-gated urea channel)—UreI 從外界獲取尿素,並以尿素酶將尿素分 解並產生氨離子及二氧化碳,此氨離子可以中和胃酸,而在菌體周圍產生一中性 的環境,使幽門螺旋桿菌可暫時存活於胃部 [38, 39]。目前被分離出來的所有幽 門螺旋桿菌菌株中都有尿素酶的存在,但其表現量可能有些微的差異 [40],且研 究指出,若將尿素酶基因剔除,幽門螺旋桿菌則失去生長於胃部的能力,並縮短 存活於胃部時間,因此尿素酶對幽門螺旋桿菌之存活扮演重要的角色 [41]。
1.4-2 鞭毛(flagella)
幽門螺旋桿菌尾部具有4~6 根鞭毛,可藉由鞭毛來進行移動,雖然它可在強
酸條件下存活,但菌體無法在這樣的環境下停留太久,當幽門螺旋桿菌進入胃腔 時,胃腔內為極酸的環境,此時幽門螺旋桿菌先以上述所說的尿素酶中和胃酸,
接著它可被胃黏膜的黏液層(mucus gel layer)中高濃度的尿素與重碳酸根離子 (bicarbonate)所趨化,再利用鞭毛和其螺旋結構幫助菌體穿透黏液層,才能穩定寄 生於pH 值比較接近中性的胃表皮與胃黏液層之間(pH 5.5) [42, 43],不過也有研 究指出幽門螺旋桿菌並非受尿素與重碳酸根離子所趨化,而是透過胃黏液層的pH 梯度(gradient) 所趨化 [44]。
1.4-3 脂多醣體(lipopolysaccharide, LPS)
先前的文獻指出,幽門螺旋桿菌的脂多醣體致死毒性(lethal toxicity)及引發的 免疫反應都較其他革蘭氏陰性菌低 [45, 46],所以幽門螺旋桿菌 LPS 並非直接嚴 重傷害宿主細胞,但是由於其LPS 上的 O-抗原(O-antigen)有果醣基化(fucosylation) 模擬出類似人類血型抗原(blood group antigen)--Lewis 抗原的構型,藉此幽門螺旋 桿菌可以逃避人體免疫系統的辨認,延長其感染的時間 [2],且有研究提出了這 些類似Lewis 抗原的 LPS 可能使宿主產生了自體免疫抗體(autoantibody),這些抗 體會去攻擊分泌胃酸的parietal cell 使細胞損傷,胃酸分泌減少最後可能就導致胃 癌 [21]。
1.4-4 Vacuolating toxin A (VacA)
約有50%的幽門螺旋桿菌菌株會分泌出有活性的 VacA 蛋白,且許多研究指 出VacA 與幽門螺旋桿菌造成消化性潰瘍或胃癌有很大的關係 [2, 47],VacA 被分 泌出來後有一部份會黏附在幽門螺旋桿菌外膜 [48],當菌體接觸胃上皮細胞後,
VacA 會聚合成複合體,在上皮細胞表面形成陰離子選擇通道(anion-selective channel),此通道可以讓重碳酸根(bicarbonate)、尿素與有機陰離子(organic anions) 從細胞質內運出細胞外,提供幽門螺旋桿菌生長所需的營養 [49]。而 VacA 也會
經由細胞之內嗜作用(endocytosis)進入細胞,插入內嗜小體(endosome)膜上並形成 孔洞,造成內嗜小體內陰離子組成的改變,最後在細胞內形成一個大空泡並使細 胞因滲透壓改變而脹破 [50]。此外 VacA 還可以改變細胞的 tight junction 使通透 性增加,許多小分子(鐵、鎳、離子、胺基酸等)因此可以送出細胞外,供給幽門 螺旋桿菌營養,而有文獻也指出VacA 可通過 tight junction 並抑制宿主 T cell 的活 性,另外VacA 也可進入細胞內使上皮細胞中的細胞色素 c (cytochrome c)從粒線 體釋放,活化細胞凋亡 [2, 50]。VacA 被許多科學家研究,但大多為體外研究(in vitro),體內研究(in vivo)的證據還不是很多。
1.4-5 Cytotoxin-associated gene A (CagA)
CagA 是幽門螺旋桿菌中最為被廣為研究也是最主要的致病因子,且目前並 沒有同源蛋白(homologous protein)在其他細菌中被發現 [50],這說明了 CagA 在 幽門螺旋桿菌中的獨特性。CagA 蛋白是由基因 cagA 所表現,而 cagA 位於幽門 螺旋桿菌中的cytotoxin-associated gene pathogenicity island(cag PAI),cag PAI 長度約40 kb,是由一群特殊基因群所組成,其中除了 cagA 以外還有許多負責表 現第四型分泌系統(type IV secretion system, TFSS)的基因,研究認為幽門螺旋 桿菌可藉此系統將 CagA 蛋白或其他致病因子直接注入宿主細胞中 [50, 51]。
CagA 進入表皮細胞後除了會刺激細胞分泌 IL-8 並活化 proinflammatory 訊息傳遞 外 [52],蛋白的 c 端(c-terminus)還會被宿主細胞的一些 kinase 如 Src 或 c-abl 磷酸 化,磷酸化後的 CagA 會在細胞膜附近並可參與在宿主細胞內的訊息傳遞,就像 細胞內一些receptor tyrosine kinase (RTK)的功能並且影響到細胞骨架(cytoskeleton) 的組成與細胞的移動力,像是經由CagA 本身 SH2(Src homology region 2)domain 和細胞的Csk(C-terminal Src kinase)結合,導致 Src 失去活性無法對 cortactin 進 行磷酸化,去磷酸化的cortactin 和 F-actin 結合後會改變細胞骨架使細胞產生蜂鳥 狀型態(hummingbird phenotype),或是當磷酸化的 CagA 和細胞膜附近的 Grb2、
SoS、Ras 進行交互作用後,會引發下游 MAPK 的訊息傳導,最後導致細胞型態 的改變;此外 CagA 和蛋白質磷解酶(phosphatase) SHP-2 結合後,還會活化 c-met,
而c-met 不正常的活化則和細胞癌化有關 [50, 53]。若 CagA 沒有被磷酸化,其也 會影響與 cell junction 相關的蛋白如 ZO-1、JAM、E-cadherin 等,使細胞形變 (transformation)為接近癌細胞的型態 [50]。
約有 50%~70%的幽門螺旋桿菌菌株帶有 cag PAI 基因,稱為 CagA+,由於 CagA 劇烈影響宿主細胞,CagA+菌株被認為容易造成消化性潰瘍或胃癌等嚴重的 疾病,而也確實有研究發現到感染CagA+菌株的病人較容易發生胃癌的徵兆 [54],
然而上述情形只符合西方國家,在亞洲國家病人身上分離出來的幽門螺旋桿菌菌 株皆為CagA+,與臨床之結果並沒有太大的關聯性 [2]。
1.5 幽門螺旋桿菌之膜蛋白(outer membrane protein, OMP)與黏著因子(adhesin) 的重要性
之前的文獻分析出幽門螺旋桿菌的基因當中,有許多負責表現膜蛋白的基因,
科學家將他們分為五大平行同源(paralogous)基因的家族(family),其中一類最為龐 大也最為主要的基因家族包含了21 個 Hop 與 12 個 Hor 膜蛋白(Hop: H. pylori outer membrane protein, Hor: Hop-related protein),共 33 個膜蛋白,這家族包含了許多 和細菌貼附(adhesion)於胃表皮細胞相關的蛋白,而其他四個家族的膜蛋白則包含 了一些膜孔蛋白(porin)、和運輸鐵相關的運輸蛋白(transporter)、和鞭毛相關的蛋 白以及其他功能未知的蛋白 [2, 55]。
和細菌貼附相關的蛋白就稱為黏著因子(adhesin),黏著因子也是致病因子的 一種,主要原因為當菌體進入胃腔中時,胃腔的強酸環境約為pH 1~2,幽門螺旋 桿菌在此環境下只能存活30 分鐘,這時會配合論文 1.4 中介紹過的尿素酶與鞭毛 幫助菌體快速移動至胃黏液層,而胃黏液層的 pH 環境才適合幽門螺旋桿菌在這 裡長久生存,之後細菌必須緊緊貼附於胃表皮細胞表面,並且繁殖形成菌落
(colonization),最後才有可能對宿主細胞造成影響或傷害;再者,由於胃壁可能時 常有蠕動或胃酸分泌的情況,幽門螺旋桿菌必須以黏著因子貼附才能讓它們免於 被清除的命運,因此黏著因子對幽門螺旋桿菌在胃中的生存是不可或缺的 [50, 56]。另外,幽門螺旋桿菌的黏著因子也被報導和疾病或是發炎反應有關 [50],以 下簡述幾個被研究認為是黏著因子的蛋白。
1.5-1 Sialic acid-binding adhesin (SabA, HopP)
人類胃細胞會表現血型抗原(blood group antigen)中的Lewis抗原,而SabA被研 究會和有不同醣化修飾的Lewis x抗原和Lewis a抗原(sialyl-Lex和sialyl-Lea)結合,最 早是發現到一些sialyl-Lewis抗原會和幽門螺旋桿菌結合,接著將會和sialyl-Lewis 抗原結合的蛋白純化出來而找到,並命名為sialic acid-binding adhesin, SabA [57]。
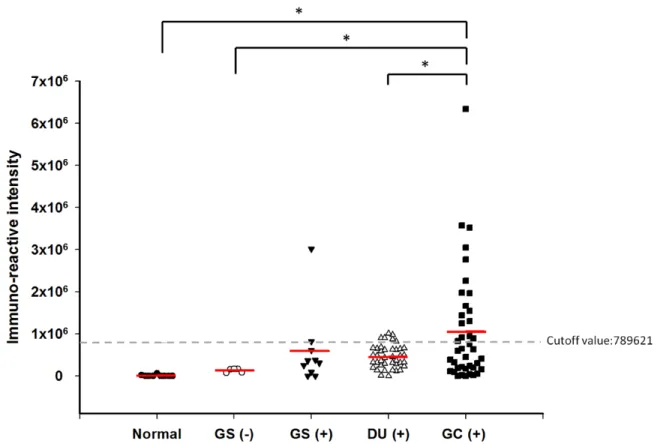
有研究發現帶有sialyl-Lex及sialyl-Lea抗原和受幽門螺旋桿菌感染造成的胃發炎及 胃癌有關 [2],另外在美國也有研究指出SabA的表現和嚴重的intestinal metaplasia、
胃體萎縮及胃癌發生有高度相關 [58],顯示了SabA確實是一個幽門螺旋桿菌的致 病因子。另外由於嗜中性白血球表面也有sialyl-Lewis抗原,因此SabA也會和其結 合並且活化嗜中性白血球,活化的嗜中性白血球會釋放出氧自由基而傷害胃表皮 細胞 [59]。SabA的表現會受胃中環境如pH值影響而改變其表現量,另外也有研究 指出SabA會和細胞外基質蛋白(extracellular matrix protein) laminin結合 [2, 58]。
1.5-2 Blood group antigen binding adhesin (BabA, HopS)
BabA是另一個功能較確立的黏著因子,會和人類胃細胞中的血型抗原Lewis b 抗原(Leb)結合,最早也和SabA的發現方式相似。研究發現到BabA的基因有兩個不 同的對偶基因(allele),為babA1和babA2,但由於babA1的signal peptide不完全所以 不會表達蛋白,只有babA2會表達有活性的蛋白 [60]。有許多研究指出臨床病人中 感染有表現BabA的幽門螺旋桿菌,其和嚴重的消化性潰瘍與胃癌有非常高度的相
關,當有BabA表現時,病人有較嚴重的免疫反應和發炎情況 [2, 61, 62],另外文 獻顯示當以基因轉殖老鼠(transgenic mice)轉殖人類的Leb抗原後並感染幽門螺旋桿 菌,比沒有轉殖的老鼠有更為嚴重的發炎情況 [63],這些都代表了幽門螺旋桿菌 透過BabA貼附宿主細胞並導致較嚴重的疾病。通常BabA的表現和vacA及cagA基因 有高度相關性,另外也有研究指出幽門螺旋桿菌會先透過BabA和細胞貼附,接著 再透過SabA和sialyl-Lex及sialyl-Lea的貼附,使幽門螺旋桿菌造成持續發炎的效果 [2, 57]。
babA有同源基因為babB,兩者之基因序列在5’端與3’端極為相似,但中間的序 列則不太相同,babB表現的蛋白BabB目前功能還不是很清楚,但有研究顯示有些 菌株含有不會表現蛋白的babA1基因,此基因可以在某些時後插入babB基因當中,
而這個babA/babB複合基因體的蛋白則可和Leb結合,另外研究也發現從病人身上分 離出來的菌株其babA與babB基因的基因型組合十分繁雜,不同菌株有不同的組合,
因此科學家認為babA或babB受到菌的精密調控以反應在宿主身體內發生的事件 [2, 64]。
1.5-3 Outer inflammatory protein (OipA, HopH)
OipA最早被發現可以刺激細胞分泌IL-8,產生前發炎(proinflammatory)反應,
且其在CagA+的幽門螺旋桿菌裡,刺激的效果更為強大 [65],分析臨床病人菌株的 oipA基因發現oipA和cagA基因的表現情況是互相關聯的,且被認為和十二指腸潰瘍 及胃癌有關 [2, 65]。之後的體外研究顯示,突變OipA的基因使幽門螺旋桿菌貼附 效果及菌落形成能力下降,因此科學家認為OipA和細菌貼附有關 [66],另外透過 oipA基因突變和野生型(wild type) oipA相比的實驗可以發現到OipA會造成細胞的 focal adhesion kinase (FAK)活化,進一步使Src活化後最後導致細胞骨架的改變,這 可能是幽門螺旋桿菌感染後造成細胞型態改變的一個原因 [67]。然而和OipA結合 的宿主蛋白還未被發現,有研究指出OipA可能可以使表皮細胞生長因子受器
(epidermal growth factor receptor, EGFR)磷酸化並活化下游Akt訊息傳遞 [68]。
1.5-4 Adherence-associated lipoprotein A (AlpA, HopC)
AlpA最早被發現是由於科學家將其基因突變後發現到幽門螺旋桿菌貼附於胃 組織的能力下降,且發現到其蛋白質N端帶有脂蛋白(lipoprotein)的signal sequence,
因而取名為adherence-associated lipoprotein A, AlpA [69],之後的研究包括以突變 alpA基因的幽門螺旋桿菌與細胞或活體動物共同陪養都發現到細菌貼附及形成菌 落的能力下降,且和野生型相比發現刺激細胞分泌的IL-6下降,顯示AlpA可能也 會造成前發炎反應 [70, 71]。有文獻顯示AlpA會和宿主的laminin結合 [72]。
幽門螺旋桿菌還有其他的黏著因子被科學家們研究,如下表的其他蛋白:
【Table 1】Clin Microbiol Rev, 19: 449 (2006)
第 二 節 生 物 標 記 分 子 (biomarker) 與 蛋 白 質 微 陣 列 晶 片 (protein microarray chip)之應用
2.1 生物標記分子
2.1-1 生物標記分子簡介
生物個體在發育生長的過程中,隨著生理功能的變化,體內如mRNA、蛋白 質、荷爾蒙等生物分子也會有不同程度的增減,此外如細胞受到刺激,造成DNA 產生突變或修飾、受病原體感染,或是當細胞受損嚴重,讓蛋白質轉移至血液或 其他體液(如胸水、腹水),上述產生質變或量變的生物分子可以反應出生物個體 當下所處的生理狀況,因此稱為生物標記分子,生物標記分子的發現可以幫助早 期診斷、疾病進展的了解、預後(prognosis)情形及發展治療藥物 [73]。常見的應 用 如 肝 臟 發 炎 時 造 成 肝 細 胞 壞 死 受 損 , 因 而 讓 肝 細 胞 內 蛋 白 質 如 glutamic oxaloacetic transaminase (GOT)及 glutamic pyruvic transaminase (GPT)兩種酵素轉 移至血清中而能被偵測到,藉由偵測這兩種酵素在血清中的量可了解肝受損情形。
此外如女性懷孕時,human chorionic gonadotropin (hCG)的分泌量增加而能夠在血 液和尿液中被偵測到,此原理也廣泛應用到目前市面上常見的驗孕器材中。
生物標記分子通常以血液及體液如尿液、唾液、腹水等生物材料來分析,雖 然有十分大量的科學研究報導生物標記分子,但真正可應用於臨床的非常少,主 要原因為個體的差異性極大,近年來由於單一生物標記分子其臨床上的靈敏度 (sensitivity)與專一性(specificity)不夠高,逐漸朝向複合型生物標記分子(multiple marker)的開發,期望透過多個生物標記分子的篩選,可避免個體上之差異,提高 在臨床上應用的準確度 [74]。
2.1-2 幽門螺旋桿菌及相關疾病之生物標記分子
幽門螺旋桿菌及相關疾病之相關生物標記分子有許多研究結果,文獻大多以 直接從病人身上分離出菌株並以 PCR 方法偵測幽門螺旋桿菌中較為主要的幾個
致病因子的基因型,再和未被感染幽門螺旋桿菌的正常人做比較並以統計計算,
另一種方式是病人血清學偵測是否帶有幽門螺旋桿菌相關因子的抗體,或是以多 株抗體(polyclonal antibody)去偵測菌株身上蛋白量的表現,不論用血清學或多株 抗體方法都必須自行純化或分離出幽門螺旋桿菌蛋白,且市面上幽門螺旋桿菌蛋 白的抗體種類甚少,因此目前相關生物標記分子的研究大多會以分析菌的基因型 為主。針對診斷是否有感染幽門螺旋桿菌已於論文 1.3-3 中論述,然而感染後所 發生的疾病相關生物標記分子目前還未能真正應用於臨床診斷。常見的分析目標 如尿素酶UreA 或 UreB、VacA、CagA、鞭毛蛋白 FlaA 或 FlaB、及各種黏著因子,
有研究發現到中國地區的幽門螺旋桿菌全都有表達UreB,且不管是胃炎或是消化 性潰瘍的病人血清中都帶有CagA 抗體 [75],而在歐洲地區,相較於慢性胃炎的 病人,較多消化性潰瘍病人身上的菌株表現了CagA,且發現 CagA 及熱休克蛋白 (heat shock protein) GroEL 和胃癌病人有高度相關 [76, 77],而美國研究也發現 CagA、 SabA 及 OipA 和胃癌病人相關 [58],可以看見由於區域性臨床幽門螺旋 桿菌菌株的的差異性很大,雖然如 CagA 的致病機制已經較為了解,但仍無法成 為能普遍應用的生物標記分子,因此目前這方面的生物標記分子還未普及化。
2.2 蛋白質微陣列晶片 2.2-1 蛋白質微陣列晶片簡介
蛋白質微陣列晶片或稱為蛋白質晶片(protein chip)的概念源自於 DNA 晶片,
將多個樣品以微陣列方式點在玻片上以形成生物探針(probe),接著將待分析之物 質和玻片上樣品反應雜交(hybridization),並且能快速同時去分析多個樣品反樣強 弱的訊號,其優點為由於點在晶片上的每個樣品大小只有150 ~ 200 μm,因此樣 品量每平方公分可高達1600 個點,且樣品消耗量極少,再配合晶片的點樣通常是 以自動化的方式製作晶片,因此能快速產生多個晶片,每個晶片之上又有多個數 量的樣品,達到多重複合式(multiplex)且高通量(high-throughput)的目的。然而比
起DNA 晶片,蛋白質晶片的困難在於每一個蛋白質的製備純化比起 DNA 或 RNA 的製備還要繁複許多,且由於每個蛋白質的特性如帶電性、溶解度、穩定度等等 都不盡相同,更增加了蛋白質晶片製作的困難性,然而不能否認的是,蛋白質比 起核酸更能真實反應生物的生理狀況,因此蛋白質晶片的確能帶來更多的資訊 [78]。
蛋白質晶片的表面材質有許多種,這些材質可以幫助抓住或吸附蛋白質使其 被固定在晶片表面,並且有防止蛋白蒸發及幫助蛋白質聚集的特性,常見的如醛 基(aldehyde)、環氧樹脂(epoxy)及 poly-L-lysine 等,這些材質可以抓住蛋白質並固 定蛋白,而另外有 nitrocellulose 或一層膠的材質能吸附住蛋白質以固定化 [78, 79]。
2.2-2 蛋白質微陣列晶片之應用
蛋白質晶片可應用在許多科學研究上,例如科學家建立了酵母菌蛋白質體 (proteome)晶片,將酵母菌中 80%的蛋白都純化並點於晶片上,接著將 calmodulin 或科學家想研究的脂質加入到晶片上和蛋白反應,觀察蛋白-蛋白或蛋白-脂質的 交互作用(protein-protein interactions and protein-lipid interactions) [80],另外也有文 獻是建立了大腸桿菌蛋白質體(proteome)晶片,先建立了高通量純化大腸桿菌蛋白 質的方法,接著將4,256 個蛋白點至晶片上,除了可以研究 DNA 損傷的分子機制 以外,更可以利用病人的血清和晶片反應,找出疾病相關的生物標記分子 [81, 82],
此外還有科學家建立了人類蛋白質體晶片以及幽門螺旋桿菌蛋白質發展成的晶片,
並且也以病人血清做為材料去找尋或驗證疾病的生物標記分子 [83, 84],從以上 例子可以知道蛋白質晶片的應用很廣,可用來研究各種生物分子的交互作用,或 是用來幫助找尋疾病的生物標記分子。
第三節 研究動機、目的與策略
3.1 研究動機
如同論文前面所述,幽門螺旋桿菌和胃發炎、十二指腸潰瘍及胃癌都有高度 相關性,然而被感染的病人最終會發展成十二指腸潰瘍或是胃癌卻是兩條分歧的 臨床結果,即得到其中一種疾病的病人再得到另一種疾病的機率幾乎為 0,科學 家們普遍認為其中一個原因可能就是由於感染了不同幽門螺旋桿菌菌株的結果,
而從前人的研究中可以發現到幽門螺旋桿菌菌株的基因型有高度的變異性,其蛋 白質及致病因子的表現情況也會有很大的差異,因此我們可以假設胃癌病人及十 二指腸潰瘍病人可能是受到不同幽門螺旋桿菌菌株的感染,並且受其不同致病因 子表現情況的影響,最後才導致不同的臨床走向。另外,目前受幽門螺旋桿菌感 染後相關疾病的生物標記分子尚未被科學家所確立,若是假設成不同菌株造成病 人走向十二指腸潰瘍或走向胃癌,可以去研究造成這兩種疾病的兩種不同菌株之 間蛋白質表現量的差異,比較差異後更可以幫助找尋十二指腸潰瘍專一性或是胃 癌專一性的生物標記分子。另外在臨床應用中,如何能快速診斷受幽門螺旋桿菌 感染的病人當中哪些是得胃癌的高危險群或者哪些是得到十二指腸潰瘍的高危險 群,也是值得探討的方向。而這些疾病當中,胃癌的議題更是刻不容緩,因此找 尋幽門螺旋桿菌相關胃癌的專一性生物標記分子,以及發展方法以期望能快篩出 受幽門螺旋桿菌感染而造成胃癌的病患,會是我們主要的目標。
而從論文前面的介紹中也論述過幽門螺旋桿菌膜蛋白及黏著因子的重要性,
膜蛋白是最有可能和宿主直接接觸的區域,且膜蛋白也可能和宿主辨認(host recognition)的功能相關,尤其膜蛋白在菌體表面的特性,讓這些蛋白成為尋找病 菌生物標記分子以及發展疫苗領域內很好的選擇 [85],因此我們較感興趣的部份 為幽門螺旋桿菌膜蛋白的差異。雖然目前有一些幽門螺旋桿菌膜蛋白和相關疾病 的研究,但大多以菌株的基因型和臨床結果做比較 [76],較少從蛋白質層面去探 討,去研究菌中蛋白的表現量或是偵測病人血清中幽門螺旋桿菌蛋白質的抗體表
現量應該更能提高準確性。
3.2 研究目的
綜合以上的研究動機,我們的研究目的有以下兩項:
1. 比較十二指腸潰瘍病人或胃癌病人身上分離出來的兩種幽門螺旋桿菌菌株其 膜蛋白質體(membrane proteome)的表現差異,期望從這些差異當中找出和胃癌 相關之具潛力的膜蛋白生物標記分子。
2. 我們期望能發展出一個新的應用平台,可以在臨床上幫助快速又簡易的診斷,
並且能篩檢及區分出受幽門螺旋桿菌感染的個體當中發展成十二指腸潰瘍或 胃癌的病人。
3.3 研究策略
本實驗室先前為了比較十二指腸潰瘍(簡稱 DU)病人及胃癌(簡稱 GC)病人身 上分離出來的兩種幽門螺旋桿菌菌株(簡稱 DU strain 及 GC strain)的膜蛋白質體差 異,因此利用了16-BAC/SDS-PAGE 的二維電泳(two-dimensional gel electrophoresis) 系 統 來 分 析 膜 蛋 白 , 此 種 二 維 電 泳 與 傳 統 二 維 電 泳 的 差 異 在 於 , 16-BAC/SDS-PAGE 的 第 一 維 電 泳 是 將 蛋 白 質 以 16-BAC (benzylhexadecyl- dimethylammonium chloride)這種清潔劑(detergent)溶解蛋白,並且以蛋白質分子 量大小將蛋白在電泳中區分開來,並非傳統二維電泳當中以蛋白質等電點高低來 區分,先前研究也指出由於此種二維 電泳系統能幫助溶解較疏水性的蛋白 (hydrophobic protein),因此適合用於分析膜蛋白這類較難以溶解的蛋白 [86]。而 本實驗室先前以二維電泳比較蛋白質的表現量差異後,選取GC strain 表現量大於 DU strain 表現量的蛋白(Supplementary S1、Supplementary S2),並將膠體的蛋 白點切割下來後經膠體原位酵素切割 (in-gel digestion),再將這些表現量有差異的 蛋白以nano-LC MS/MS 鑑定身份,其結果於附錄的 Table S1,結果顯示鑑定到的
蛋白質一半以上為膜蛋白,再次證明了16-BAC/SDS-PAGE 用於分析膜蛋白的可 行性(Supplementary S3)。
而本實驗將會根據實驗室先前研究的結果: Table S1 鑑定到的蛋白進行文獻 搜尋,並且選出四個標的膜蛋白分別為OipA、AlpA、SabA 及 BabB 這四個幽門 螺旋桿菌的黏著因子,接著我們會將這四個黏著因子的重組蛋白質(recombinant protein)以 8M 尿素幫助溶解並純化出來後,利用西方墨點法(Western blotting),分 別以正常人、胃發炎無感染幽門螺旋桿菌者、胃發炎有感染幽門螺旋桿菌者、十 二指腸潰瘍病人及胃癌病人的血清去反應,觀察其免疫特異性,之後以統計分析 結果,並嘗試加入複合型生物標記分子的概念,期望找尋出胃癌相關之幽門螺旋 桿菌生物標記分子。
最後,我們會將這四種膜蛋白黏著因子蛋白點印至晶片上,由於本實驗室先 前的研究中曾將溶於尿素中的幽門螺旋桿菌蛋白點至晶片上 [84],但其點樣良率 並不高,因此我們會測試並試圖找尋出較適合這四個溶於8M 尿素緩衝夜(buffer) 的膜蛋白其點樣的最佳條件,之後再以正常人及病人血清和蛋白質晶片反應,觀 察其反應趨勢是否與西方墨點法的結果一致,期望能發展出胃癌相關之幽門螺旋 桿菌抗原膜蛋白質晶片,並能幫助臨床上的診斷。
第二章 實驗材料
第一節 幽門螺旋桿菌菌株
選用分離自臨床胃癌病人與十二指腸潰瘍病人身上之幽門螺旋桿菌菌株,由 臺大醫院內科提供,培養於臺大醫院。
第二節 血清樣本
腸胃道疾病病人之血清提供自臺大醫院內科,依據病人臨床症狀分類為十二 指腸潰瘍(duodenal ulcer)、胃癌(gastric cancer)以及非上述兩種病症之胃炎 (gastritis);病人血清均經過幽門螺旋桿菌血清學檢驗為陽性,而胃炎病人血清則 有分為陰性或陽性。另外,正常人血清定義為臨床上無腸胃道疾病症狀,並經過 幽門螺旋桿菌血清學檢驗確定為陰性之血清。
第三節 大腸桿菌及質體
品名 廠牌 E. coli Host Strain, DH5α RBC Bioscience
E. coli Host Strain, BL21 Promega Plasmid pET28a Novagene
第四節 酵素
品名 廠牌 Restriction enzyme New England Biolabs
T4 DNA ligase New England Biolabs Trypsin Promega
Lysozyme Sigma
第五節 抗體
初級抗體 廠牌 Mouse anti-His tag antibody Santa Cruz
次級抗體 廠牌 HRP-conjugated rabbit anti-mouse IgG Millipore
HRP-conjugated goat anti-human IgG Jackson ImmunoResearch Laboratories
Unconjugated rabbit anti-human IgA + IgG + IgM
Jackson ImmunoResearch Laboratories
Cy3-conjugated rabbit anti-mouse IgG Jackson ImmunoResearch Laboratories
Cy3-conjugated rabbit anti-human IgA + IgG + IgM
Jackson ImmunoResearch Laboratories
第六節 藥品
品名 廠牌
2-mercaptoethanol Merck 10X ligation buffer New England Biolabs
10X NEB buffer New England Biolabs
Acetic acid Merck
Acetonitrile, ACN Merck
Acrylamide/N, N’-methylene-bis-acrylamide Amersham Bioscience Ammonium persulfate, APS USB
Bovine Serum Albumin, BSA Merck
Bromophenol blue, BPB Pharmacia Biotech
Calcium chloride, CaCl2 Sigma
CBB-250 Amersham Bioscience
Chelating SepharoseTMFast Flow Amersham Bioscience
EDTA Sigma Ethanol Sigma Ethidium Bromide Sigma
Fast green Sigma
Formic acid Merck
Glycerol Merck Glycine Merck Hydrogen chloride, HCl Merck
Imidazole Sigma IPTG Sigma Kanamycin Sigma
Magnesium chloride, MgCl2 Sigma
Methanol Merck
Nickel chloride ,NiCl2 Sigma
PBS Gibco SDS Amresco Sodium chloride, NaCl Sigma
Sodium hydride, NaOH Sigma
TAE buffer Amresco
TEMED Merck Tris Merck Tween-20 Sigma Urea USB
第七節 試劑組
品名 廠牌 KAPA HiFiTM PCR kit KAPA Biosystems 660 nm Protein Assay Reagent Pierce
Bio-100TM DNA Ladder(range 100bp ~ 3 kbp) Promega Gel/PCR DNA Fragments Extraction Kit Geneaid
High Speed Plasmid mini Kit Geneaid ImmobilonTM Western Chemiluminescent
HRP substrate
Millipore Low Range Protein Marker Bio-Rad
PUREGENE DNA purification kit From 臺大醫院內科楊 智欽醫師
第八節 重要儀器
儀器 廠牌型號 Dry bath incubator Major science
Electrophoresis power supply 301/1001 Amersham Bioscience ESI Q-TOF tandem mass spectrometry QSTARTM XL system GeneAmp PCR system 2700 Applied Biosystem
Himac CF15R centrifuge Hitachi Hotplate Stirrer JLab Tech ImageQuant 300 Amershan Kubota 1020 centrifuge Kubota
Liquidator96 RAININ
Luxscan 10K Microarray Scanner CapitalBio Maxi Dry Plus (SpeedVac) Heto Holten
Micro-centrifuge Bertec Microfuge R centrifuge Beckman Coulter
Multichannel pipett RAININ Nano LC system LC packing NVT 90 ultraspeed rotor, for Backman Centrifuge Beckman Coulter
Orbital shaker TKS
pH meter InLab
Pipett Gilson SlideWasher 8 Slide Clean-up Station CapitalBio
SmartArrayer 136 CapitalBio
SpectraMax® M5 multi-detection reader Molecular Devices TM-325 autoclave Tomin
Ultracentrifuge Beckman Coulter
Ultrasonicator Sonics & Materials, Inc Ultrospec 3000 Spectrophotometer Amersham Biosciences Vortex-2 Genie Scientific Industries
第三章 實驗方法
第一節 幽門螺旋桿菌標的基因之質體建構(plasmid construction)與重組膜 蛋白質的表現與純化
1.1 標的基因之質體建構
1.1-1 抽取幽門螺旋桿菌之 genomic DNA (Extraction of genomic DNA )
先以棉棒刮一個培養基(plate)的菌量並收集至裝有 1.5mL ddH2O 的 eppendorf 中,再以7000g 轉速離心 5min 後去除上清液,沉澱的 pellet 即是菌體,可置於-20℃
中保存或繼續以下步驟,接著使用PUREGENE DNA purification kit 抽取幽門螺旋 桿菌之genomic DNA,先加 600μL 的 cell lysis solution,輕輕混勻後置於 80℃, 5min 以破菌完全,之後回到室溫,再加入3μL 的 RNase A,在 37℃下 incubate 60min 以去掉RNA,再來加入 200μL 的 protein precipitation solution on ice 5min,再離心 15000g, 1min, 4℃以沉澱蛋白質,之後取上清液再和 isopropanol 600μL 混勻,並 一樣以15000g, 1min, 4℃離心後倒去上清液,最後先加入 600μL 100%酒精洗 pellet,
離心後倒去上清再換成600μL 70%酒精洗 pellet,離心 13000g, 10min, 4℃後去掉 上清,pellet 晾乾(可 overnight),最後加入 50μL 的 DNA hydration solution(或 ddH2O) 並於65℃下 incubate 1hr 回溶 DNA,DNA 可放置於-20℃中保存。
1.1-2 聚合酶連鎖反應 (Polymerase Chain Reaction, PCR)
利用聚合酶連鎖反應放大標的基因,使用KAPA HiFiTM PCR kit,並以上述抽 取之幽門螺旋桿菌genomic DNA 做為模板(template),其反應時使用配方如下,一 次實驗25μL 兩管,共 50μL,:
*使用的primer 經過 SignalP 4.0 Server 軟體預測蛋白之 signal peptide 後,去除 signal sequence 以設計 primer,序列附於附錄 Table S2
充分混合後,開始聚合酶連鎖反應,四個基因個別的反應條件如下:
聚合酶連鎖反應完成之後,利用1%洋菜膠體電泳分析 OipA, AlpA, SabA 及 BabB 之PCR 產物。
1.1-3 洋菜凝膠電泳 (Agarose gel electrophoresis)
配置 1%之洋菜膠體,取 1 g agarose 加入 1X TAE buffer 至 100 mL (20X TAE buffer 配方如下),放入微波爐加熱 3 至 5 分鐘沸騰使 agarose 完全溶解,將些微 冷卻之溶液注入預先架設好之電泳模板中,置於室溫下使其冷卻凝固,之後將膠 體放在電泳槽中,並加入1X TAE buffer 蓋過膠體做為電泳時的緩衝液。將 6X DNA
成份 體積(μL)
KAPA HiFi DNA polymerase DNA Template
*Primer forward (10 μM)
*Primer reverse (10 μM) dNTP (10 mM)
5X KAPA HiFi fidelity buffer Sterile water
0.5 1 0.75 0.75 0.75 5 16.25 Total volume 25
loading dye 和 PCR 產物混勻後注入膠體樣品槽中,以電壓 110 伏特(V)、電流 400 毫安培(mA)下進行 30 分鐘電泳,電泳結束後,將膠體浸入染色液(EtBr 濃度: 0.5 μg/mL )中 5~10 分鐘,以清水清洗數次後置於 ImageQuant 300 儀器中以 UV 照射 進行DNA 影像分析,並利用 IQuant Capture 300 進行影像擷取。
20X TAE buffer
EDTA 15 g
Tris-Base 194 g
NaOH 8 g
Acetic acid 80 mL 加ddH2O 至 2L
1.1-4 DNA 電泳產物之回收 (Gel extraction)
用 酒 精 擦 拭 過 刀 片 後 將 要 回 收 的 DNA 片段從洋菜膠體上切下,裝入 eppendorf 中並秤出洋菜膠體之重量,之後使用 Gel/PCR DNA Fragments Extraction Kit 進行 PCR 產物回收,而回收之 DNA 儲存於-20 ℃。
1.1-5 質體 DNA 的製備與微量抽取 (Extraction of plasmid DNA)
實驗使用的 pET28a 質體原轉型於 E. coli JM109 中保存,所以先將帶有 pET28a 的 JM109 菌養開,以過火殺菌之 loop 沾 JM109 菌液並將菌液畫開在含有 Kanamycin (pET28a 帶有 Kanamycin-resistant 基因,而抗生素以 LB 體積之五千分 之一的比例加入50 mg/mL 之 Kanamycin)之 LB 培養基上,置於 37℃培養箱中培 養16~18 小時,之後挑出單一菌落(single colony)並於 5mL 的 LB 液中在 37℃下震 盪培養16~18 小時(LB 也含有 Kanamycin),菌液再使用 High Speed Plasmid mini Kit 抽取pET28a 質體,抽出的質體儲存於-20 ℃。
1.1-6 限制酶水解 (Reaction of restriction enzyme)
將準備好的 pET28a 質體與 PCR 產物回收的 DNA 進行限制酶水解,反應配 方如下:
在37℃下反應 2 小時,之後加入 calf intestinal alkaline phosphatase (CIP) 1mL, 1 小 時37℃以防止自我接合(self-ligation),然後加入 6X DNA loading dye 後,利用 DNA 電泳技術分析其DNA 片段,最後使用 Gel/PCR DNA Fragments Extraction Kit 將 DNA 片段回收,回收之 DNA 儲存於-20 ℃。
1.1-7 與 pET28a 質體之接合反應 (Ligation with pET28a plasmid)
將被限制酶水解後的四個DNA 片段與質體 pET28a 進行酵素結合反應,DNA 片段與pET28a 各加入 50 ng 來進行反應,反應條件如下:
成份 比例
質體或DNA 片段 10X NEB buffer
10X BSA BamHⅠ
XhoⅠ Sterile water
1000~2000 ng 5μL 5μL 1μL 1μL 補體積 Total volume 50μL
將混合好的樣品於 16 ℃下培養 20 小時,接著轉型至勝任細胞(competent cells) E. coli DH5α 菌株。
1.1-8 轉型作用 (Transformation)
將勝任細胞 DH5α 和接合反應後的產物混勻後置於冰上 15 分鐘,之後於 42℃
下進行熱休克效應(heat shock) 45 sec,再置於冰上 2 分鐘,接著加入 1 mL LB 液 並於37 ℃下培養 1 小時使細菌恢復並增殖,培養過後的菌液以 5000 rpm,10 分 鐘之離心狀態離心後,去除上清液至剩下 50 μL,輕拍離心管管壁使離心之菌體 均勻懸浮其中,再以過火殺菌之玻璃棒將菌液均勻塗在含有Kanamycin (以 LB 體 積之五千分之一的比例加入50 mg/mL 之 Kanamycin)之 LB 培養基上,置於 37℃
培養箱中培養16~18 小時。
1.1-9 以 colony PCR 之方式篩選轉型基因庫 (Colony PCR screening)
已滅菌之離心管加入 10 μL 之滅菌水,用 tip 尖端沾取 LB 培養基上之單一菌 落後加入離心管之滅菌水中,並以 1μL 之含有菌落的滅菌水做為模板,再以 pET28a 上之 T7 與 T7 terminator 的序列做為 primer 以及論文中 1.1-2 提及之四個 基因上之專一性primer 來進行 PCR 反應,T7 與 T7 terminator primer 之 annealing
成份 比例
質體pET28a DNA 片段 T4 DNA ligase (1 U/μL)
10X ligation buffer Sterile water
50 ng 50 ng 1μL 1μL 補體積 Total volume 10μL
temperature 為 50℃。PCR 產物以 1%之洋菜膠體電泳分析,選擇有增殖出正確 DNA 分子量之菌株,這些菌株即帶有成功建構的含標的基因之質體,將這些菌株培養 後進行微量質體DNA 之抽取,質體保存於-20 ℃,
1.1-10 DNA 定序 (DNA sequencing)
萃取之質體 DNA 送交台大醫學院第一共同儀器中心進行 DNA 定序,確認其 DNA 序列無誤後,由於勝任細胞 DH5α 不適合用來表現與純化重組蛋白,因此將 質體再進行轉型作用至另一種勝任細胞 E. coli BL21 菌株。
1.1-11 製備勝任細胞 (Preparation of competent cells)
要製備勝任細胞 BL21 菌株,首先取 BL21 單一菌落接種至 2 mL 培養液中震 盪培養16~18 小時,取 1.5 mL 之菌液接種至 150 mL LB 培養液中(1:100 放大),
於37℃震盪培養至菌液之 OD600讀值為0.3 到 0.4 之間,之後以 5000 rpm,10 分 鐘,4℃之離心狀態下將菌體沉澱,去除上清液後,以下步驟全置於冰上操作,將 75 mL 之滅菌過的 100 mM MgCl2溶液加入並輕輕拍打使沉澱的菌體均勻懸浮,
之後置於冰上10 分鐘,離心再將菌體沉澱,去掉上清液,接著再加入 75 mL 之 滅菌過的100 mM CaCl2溶液一樣將菌體均勻懸浮後置於冰上15 分鐘,之後一樣 離心完後去掉上清液,最後以和原菌液比100:1 的 100 mM CaCl2含有20%甘油 (glycerol)的溶液回溶菌體即製備完成。將成功建構的含標的基因之質體經轉型作 用送入BL21 後,即可利用 BL21 菌株表現標的基因之重組蛋白質,而這些 BL21 可加入甘油(最後濃度為 10~15%)混勻後將菌種保存於-80℃。
1.2 重組膜蛋白質之表現與純化
1.2-1 利 用 大 腸 桿 菌 BL21 表 現 重 組 膜 蛋 白 質 (Expression of recombinant membrane proteins)
將大腸桿菌 BL21 菌株以一百分之一之比例加入 1 L 之 LB 培養液中(含有 Kanamycin),在 37℃下震盪培養至菌液之 OD600吸光值讀值落於0.5 至 0.7 之間,
再加入1 mM IPTG 分別使帶有四個標的基因(OipA, AlpA, SabA 及 BabB)的 BL21 誘導表現這四個重組蛋白質,經過4 小時之培養後,利用 JA-14 rotor 以 6000rpm,
10 分鐘,4℃之離心條件下將菌體在離心瓶中離心,去除上清液後加入含有 lysozyme (1 mg/mL)之 binding buffer 40 mL 使之再懸浮(binding buffer 配方如下),
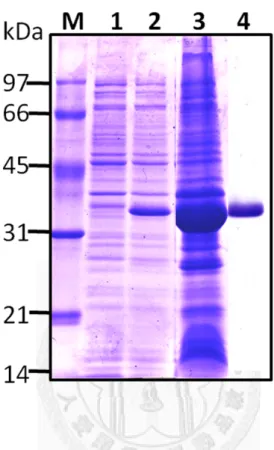
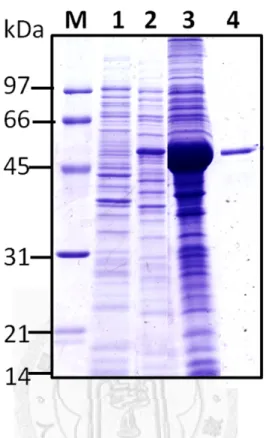
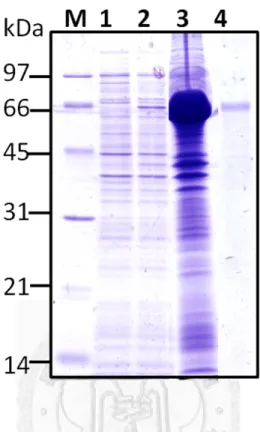
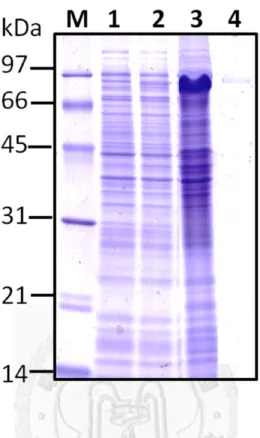
然後放入-80℃冰箱中冷凍解凍各三次以幫助破菌,接著放在冰上利用超音波儀器 (Ultrasonicator)震盪 0.5 sec、暫停 0.5 sec,持續四分鐘,循環 6~7 次,將細菌之外 膜徹底打碎,過程中要注意溫度不能過熱以免破壞蛋白質。最後使用JA-14 rotor 以13000 rpm,4 ℃離心 15 分鐘後,分別收集上清液與沉澱部份,經由本篇論文 第三章1.3 的 SDS-PAGE 方法檢測重組蛋白質的表現量與表現情形。
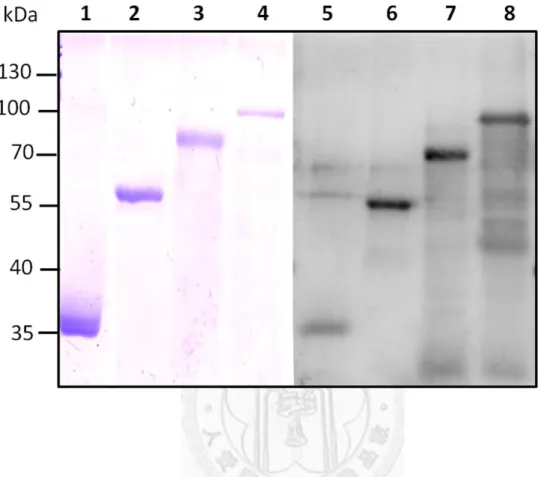
。
1.2-2 重組膜蛋白質之純化 (Purification of recombinant membrane proteins) 由於表現出的重組蛋白質大部份位在沉澱也就是包含體(includsion body)中,
於是將以13000 rpm 於 4℃離心 15 分鐘收集到的沉澱部份,加入含有 8M 尿素(urea) 的binding buffer 40 mL,由於在 4℃下 urea 容易結晶而影響其效果,所以要在室 溫或不低於4℃的低溫下作用 1 小時溶解沉澱物,以利之後純化。
Binding buffer
Tris-HCl 20 mM
NaCl 500 mM
Immidazole 5 mM
pH7.9