國立臺灣大學公共衛生學院環境衛生研究所 碩士論文
Graduate Institute of Environment Health College of Public Health
National Taiwan University Master Thesis
活性棘阿米巴原蟲定量方法開發與環境暴露初探 Method Development for Quantifying Viable Acanthamoeba and Exposure Assessment in Various
Environments
郭忠龍
Chung-Long Kuo
指導教授:張靜文 博士 Advisor: Ching-Wen Chang, Ph.D.
中華民國 102 年 1 月
Jan, 2013
i
致謝
謝謝我的指導老師張靜文老師,這兩年半我在您的身上學到太多東西,不僅 是邏輯思考,處理事情以及做人處世上亦是,對於做事情絕不馬虎的態度更是我 畢生的目標。
謝謝口試委員慈大公衛所曾俊傑老師、逢甲環工所李書安老師給我在研究上 的建議,讓我的論文更加完整。謝謝台大農化系陳尊賢老師、張顥嚴同學、慈大 微免所俞美萱老師、師丈倪禮豐先生讓我學到了土壤學的相關知識以及提供研究 上的幫忙。謝謝恆春鎮農會張秀桃股長、車城鄉農會林茂盛股長及洋蔥田農民、
外婆康擇連、台大農場李建輝先生及鍾秀媚女士、迪化汙水處理廠操作部門郭紫 因組長、農業試驗所陳柱中學長、台大生態所胡哲明老師、台大園藝所張祖亮老 師,讓我在採樣跟撰寫論文時提供了很大的幫忙。
謝謝洪佩瑜學姐、張維珍學姐、呂聆文學姐、李星儀學姐對我在實驗及研究 上的指導。謝謝我的實驗室好伙伴許雅婷與洪念慈、實驗室學弟妹楊佳蓉、王儷 瑾、林楚雲、徐藝瑋、胡凱鈞在實驗上的幫助及環衛所翁嘉陽學長、白芳瑜學姐、
張瓊文學姐跟IEH 張善為粉絲團的所有同學,及我的好鄰居學弟妹們的加油打氣。
謝謝偉大航道的所有夥伴,每次出遊都讓我神清氣爽!論文煩惱一掃而空!
謝謝老大、大姊張雨慧,做好自己是身為人的第一步,我絕對不會忘記的。
最後謝謝我的爸媽、姊姊阿密、三位弟弟以及弟媳蝦米外加姪女晴晴,還有 在家裡工作的叔叔們,謝謝你們在我離家這七年都很平安,可以讓我完成自己的 學業。
102.01.17 郭忠龍
ii
摘要
棘阿米巴原蟲(Acanthamoeba spp.)為自由型阿米巴原蟲(free-living amoeba, FLA)的一種,存在於土壤、空氣以及人工與自然水體當中。暴露於活性棘阿米巴 原蟲可感染棘阿米巴角膜炎(Acanthamoeba keratitis ,AK)、棘阿米巴皮膚炎
(Cutaneous acanthamebiasis)以及肉芽腫性阿米巴腦膜炎(Granulomatous amoebic encephalitis, GAE),目前在環境當中,學術研究多屬定性之棘阿米巴原蟲檢出報告,
以致難以量化各式環境之暴露風險。本研究嘗試建立以ethidium monoazide (EMA) 搭配即時定量聚合酶連鎖反應(real-time quantitative polymerase chain reaction, qPCR)方法,以定量土壤、空氣以及水體中活性棘阿米巴原蟲,並分別於洋蔥田 (N=12)、蔬果/香草植物田(N=36)及水稻田(N=5)採集表層土壤,於洋蔥田中央進行 空氣採樣(N=12),於冷卻水塔(N=6)、廢水處理場(N=3)、農田溝渠(N=6)、水稻田 (N=6)以及生物實驗室洗眼站(N=4)採集水體,再以所建立之 EMA-qPCR 及 qPCR 方法定量樣本中活性與總棘阿米巴原蟲濃度。土壤採樣同時也記錄當時土壤結構 (鬆軟、堅硬但有破壞、堅硬)以及植被覆蓋狀況(無植物、有植物),並於實驗室分 析土壤的pH 值、含水量及異營性細菌濃度,另於空氣採樣時同步監測空氣中溫度、
相對濕度及風速,本研究將土壤、空氣環境因子與活性及總棘阿米巴原蟲濃度進 行統計分析,以評估影響土壤與空氣中活性與總棘阿米巴原蟲濃度之顯著相關因 子。
在開發定量活性棘阿米巴原蟲部分,本研究將未受熱與受熱棘阿米巴原蟲囊 體分別以0、2.3、23 μg/mL EMA 或 PMA 處理 5 min,再以 500W 鹵素燈距離 15 cm 光照20 min 後以 qPCR 定量,數據結果顯示,經 2.3 μg/mL EMA 或 PMA 處理後,
並不影響未受熱原蟲囊體準確定量,且可使受熱原蟲囊體下降約3 log DNA quantity,另外 EMA-qPCR 與 PMA-qPCR 在定量未受熱與受熱原蟲囊體之 DNA 上,
經魏克森等級和檢定(Wilcoxon rank sums test)顯示無統計顯著差異(P=1.00)。
iii
在定量土壤中活性棘阿米巴原蟲部分,本研究將未受熱與受熱原蟲添加至 1X-300X 滅菌土壤稀釋液當中,並以 0、2.3、23、46 μg/mL EMA 處理 5 min 再以 500W 鹵素燈距離 15 cm 或 LED 燈(PhAST Blue)光照 20 min 後以 qPCR 定量,結 果發現50X 土壤稀釋液搭配 2.3 μg/mL EMA 處理不影響未受熱原蟲準確定量亦可 使受熱原蟲下降約4-6 log DNA quantity,且鹵素燈與 LED 燈對於定量未受熱與受 熱棘阿米巴原蟲之能力經魏克森等級和檢定無統計上顯著差異(P=0.53)。
洋蔥田表層土壤活性棘阿米巴原蟲平均濃度(3.92×104 cells/g dry wt.)顯著高於 蔬果/香草植物田(1.58×104 cells/g dry wt.)及水稻田(6.57×103 cells/g dry wt.)。在總棘 阿米巴原蟲方面,洋蔥田平均濃度(8.13×104 cells/g dry wt.)亦顯著高於蔬果/香草植 物田(3.23×104 cells/g dry wt.)及水稻田(1.24×104 cells/g dry wt.)。
洋蔥田空氣中活性與總棘阿米巴原蟲平均濃度分別為1.30 與 3.38 cells/m3,而 經魏克森等級和檢定後,採收期總原蟲濃度(4.73 cells/m3)大於採收後(0.42
cells/m3)。
廢水處理廠活性與總棘阿米巴原蟲濃度以曝氣池水最高,分別為6.33×105與 6.71×105 cells/L;進水管處之活性與總原蟲濃度次之,分別為 4.52×104與5.78×104 cells/L,放流管處之活性與總原蟲濃度最低(5.23×102與1.20×103 cells/L)。冷卻水 塔中活性與總棘阿米巴原蟲濃度為3.02×102與4.13×102 cells/L。農田環境中,水稻 田水樣之活性與總棘阿米巴原蟲平均濃度分別為2.67×104與3.86×104 cells/L,農田 溝渠水樣則降低至2.16×103與4.33×103 cells/L。在實驗室洗眼站樣本則未檢出。
以斯皮爾曼等級相關(Spearman's rank correlation)分析表層土壤中活性與總棘 阿米巴原蟲濃度與土壤環境因子(土壤 pH 值、土壤含水量、土壤異營性細菌濃度、
土壤結構以及植被覆蓋)之相關性,發現 pH 值(5.41-7.60)與土壤表層活性及總棘阿 米巴原蟲濃度成正相關(r=0.60, P<0.0001;r=0.43, P=0.001);土壤異營性細菌濃度 (2.21×107-7.74×108 CFU/g dry wt.)也與活性及總原蟲濃度呈正相關(r=0.33, P=0.02;
iv
r=0.39, P=0.004);而土壤含水量(2.19-58.23%)與活性及總原蟲濃度呈現負相關 (r=-0.4, P=0.003;r=-0.37, P=0.006)。
以多元線性逐步迴歸(multiple linear regression with stepwise procedure)分析表 層土壤中活性與總棘阿米巴原蟲濃度之影響因子,顯示pH 值(5.41-7.60)增加可影 響活性與總棘阿米巴原蟲濃度(β=21322, P<0.0001 和 β=37409, P=0.007),而土壤異 營性細菌濃度(2.21×107-7.74×108 CFU/g dry wt.)增加亦有同樣作用(β=0.000064, P=0.001 和 β=0.00017, P=0.001)。
以斯皮爾曼等級相關分析空氣中活性與總棘阿米巴原蟲濃度(0.11-3.61 與 0.20-6.99 cells/m3)、土壤表層活性與總原蟲濃度及環境因子(土壤 pH 值、土壤含水 量、土壤異營性細菌濃度、土壤結構以及植被覆蓋、空氣溫度、空氣相對溼度、
空氣風速)之相關性時,發現僅空氣中總棘阿米巴原蟲濃度與土壤植被覆蓋呈現正 相關(r=0.52, P=0.08),顯示農田表面有植被覆蓋時,空氣中總棘阿米巴原蟲濃度較 高。
本研究開發活性棘阿米巴原蟲之定量監測方法,並將其應用在土壤、空氣、
水體當中以得知棘阿米巴原蟲濃度,並嘗試利用統計方法從土壤及空氣環境因子 中找出影響棘阿米巴原蟲濃度變化的因子,然而本研究是偵測 Acanthamoeba spp.,
包含了致病性及非致病性,需要有研究去說明致病性與非致病性棘阿米巴原蟲在 不同環境中的比例,方能清楚知道人在環境當中的暴露風險為何。
關鍵字:棘阿米巴原蟲、核酸染劑、及時定量聚合酶連鎖反應、土壤、空氣、水 體
v
Abstract
Acanthamoeba spp. are free-living amoebae and ubiquitous in a wide variety of
natural habitats and human-made environment, including soil, air and water. Exposure to viable Acanthamoeba may cause Acanthamoeba keratitis (AK), cutaneous
acanthamebiasis and lethal granulomatous amoebic encephalitis (GAE) in human.
Previous studies have focused on detection of Acanthamoeba from various environments. Limited information on quantitative data makes the assessment of exposure risk difficult. Hence, we have adopted ethidium monoaziede (EMA) coupled with real-time quantitative polymerase chain reaction (qPCR) to quantify viable Acanthamoeba in soil, air and water environmental. Further, we applying EMA-qPCR
and qPCR to quantity viable and total Acanthamoeba in soil which seeded with onion, vegetable/Herb and rice. Air samplings were also conducted in onion farmlands. Water samplings were conducted in cooling tower, wastewater treatment plant, rice fields, farmland ditch, as well as eyewash stations in biological laboratory. Environmental factors in soil sample including water content, pH, heterotrophic plate count, structure and vegetation cover. Temperature, wind vecocity and relative humidity in air were meansure in order to determine how these factors affecting the viable and total Acanthamoeba in farmland soil and air.
In the present study, alive and heat-inactivated Acanthamoeba cyst were treated with 0 (control)、2.3、23 μg/mL EMA or PMA for 5 min followed by 500W halogen light exposure for 20 min. After DNA extraction and qPCR, DNA quantity in samples was determined. We found that 2.3 μg/mL EMA or PMA didn’t affect the avlie cyst quantification, while the heat-inactivated cyst decreaed about 3 log DNA quantity. There is no statictical. The result indicated that there is no statistically significant difference
vi
between EMA-qPCR and PMA-qPCR in quantifying alive and heat-inactivated Acanthamoeba cyst (P=1.00).
In the part of quantifing Acanthamoeba in soil by EMA-qPCR, alive and heat-inactivated Acanthamoeba were spiked to 1X-300X autocalved soil dilution sulrries and treated with 0 (control)、2.3、23、46 μg/mL EMA for 5 min followed by 500W halogen light exposure or LED light (PhAST Blue) for 20 min. We found that incubated 2.3 μg/mL EMA for 5 min with a 500W halogen light exposure or PhAST blue for 20 min can discriminate between alive and heat-inactivated Acanthamoeba in 50 fold dilution soil slurries. There is no statistically significant difference between the use of halogen light and PhAST blue in quantifying alive and heat-inactivated
Acanthamoeba cyst (P=0.53).
In exposure assessment, the mean concentration of viable Acanthamoeba (3.92×104 cells/g dry wt.) in onion farmland soil is statistical higher than
vegetable/Herb farmland (1.58×104 cells/g dry wt.) and rice field (6.57×103 cells/g dry wt.), while the mean concentration of total Acanthamoeba (8.13×104 cells/g dry wt.) in onion farmland soil is statistical higher than vegetable/Herb farmland (3.23×104 cells/g dry wt.) and rice field (1.24×104 cells/g dry wt.).
The levels of viable Acanthamoeba concentration in air were between 0.11 and 3.61 cells/m3, while total Acanthamoeba were at 0.20 to 6.99 cells/m3. We found the total Acanthamoeba concentration (4.73 cells/m3) during harvest is statistical higher than the period after harvest (0.42 cells/m3).
The level of viable Acanthamoeba in wastewater treatment plant ranged from 5.23×102 to 6.33×105 cells/L, while total those of total Acanthamoeba were from 1.20×103 to 6.71×105 cells/L. The viable and total Acanthamoeba contentration in cooling tower is 3.02×102 and 4.13×102 cells/L. In farmland envorinment, the viable
vii
and total Acanthamoeba contentration in rice field is 2.67×104 and 3.86×104 cells/L, while the contentration in farmland ditch is 2.16×103 and 4.33×103 cells/L. The sample from eyewash station is non-detected.
Spearman's rank correlation analyzed the association between soil environmental factiors and viable/total Acanthamoeba concentration in soil. We found pH and the concentration of heterotrophic plate count is positive association with viable/total Acanthamoeba concentration in soil (PH: r=0.60, P<0.0001; r=0.43, P=0.001, HPC:
r=0.33, P=0.02; r=0.39, P=0.004), while water content is negative association (r=-0.4, P=0.003; r=-0.37, P=0.006).
Multiple linear regressions with stepwise procedure model analyzed the association between soil environmental factiors and viable/total Acanthamoeba concentration in soil.
Viable and total Acanthamoeba concentration were positively associate with pH (β=21322, P<0.0001 and β=37409, P=0.007).On the other hand, the concentration of heterotrophic plate count also had positive association with viable and total
Acanthamoeba concentration (β=0.000064, P=0.001 and β=0.00017, P=0.001).
Analyzing Spearman's rank correlation coefficient among viable/total
Acanthamoeba concentration in air and soil and the environmental facors, we found
total Acanthamoeba concentration in air were positive association with vegetation cover (r=0.52, P=0.08) suggested that when the farmland cover with vegetation, the total Acanthamoeba concentration in air will be high.
Keywords: Acanthamoeba, DNA intercalating dyes、Real-time quantitative PCR、soil、
air、water
viii
目錄
致謝 ... i
摘要 ... ii
Abstract ... v
目錄 ... viii
圖次 ... xiv
表次 ... xvii
第一章 前言 ... 1
1.1 研究背景 ... 1
1.2 文獻回顧 ... 1
1.2.1 棘阿米巴原蟲(Acanthamoeba spp.) ... 1
1.2.2 棘阿米巴原蟲導致之疾病及與土壤之相關性 ... 2
1.2.3 棘阿米巴原蟲於土壤中之流佈及其影響因子 ... 6
1.2.4 棘阿米巴原蟲於空氣中之流佈及其影響因子 ... 9
1.2.5 棘阿米巴原蟲於水體中之流佈 ... 14
1.2.6 環境棘阿米巴原蟲之偵測與定量方法 ... 15
第二章 研究目的 ... 18
第三章 研究架構 ... 19
第四章 材料與方法 ... 22
4.1 微生物菌種 ... 22
4.1.1 Acanthamoeba castellanii (ATCC30234) ... 22
ix
4.1.1.1 滋養體 ... 22
4.1.1.2 囊體 ... 22
4.1.2 Escherichia coli (ATCC25922) ... 22
4.2 試劑與緩衝液 ... 22
4.2.1 製備 ATCC medium 712 ... 22
4.2.2 製備 Page’s Amoeba Saline ... 23
4.2.3 製備囊化培養液(Encyst medium) ... 24
4.2.4 製備 TE buffer ... 24
4.2.5 製備 Nutrient agar (NA) ... 24
4.2.6 製備 R2A agar ... 25
4.2.7 製備 Non-Nutrient agar (NNA) ... 25
4.2.8 製備 Heat-killed E.coli NNA plate ... 25
4.3 方法開發 ... 26
4.3.1 棘阿米巴原蟲囊體核酸染劑之選擇 ... 26
4.3.1.1 置備已知濃度的未受熱與受熱棘阿米巴原蟲囊體懸浮液 ... 26
4.3.1.2 使用 Heat-killed E.coli NNA plate 確認未受熱與受熱囊體之活 性 ... 26
4.3.1.3 EMA 與 PMA 核酸染劑之配置 ... 26
4.3.1.4 EMA 與 PMA 核酸染劑處理 ... 27
4.3.1.5 DNA 萃取 ... 27
4.3.1.6 Real-time quantitative PCR (qPCR) ... 27
4.3.1.7 製備棘阿米巴原蟲 DNA 標準品及檢量線 ... 28
4.3.1.8 定量指標、ND 之定義及處理 ... 29
x
4.3.2 EMA-qPCR 定量土壤中活性棘阿米巴原蟲之方法開發 ... 30
4.3.2.1 未滅菌土壤添加未受熱與受熱棘阿米巴原蟲進行不同濃度 EMA 與鹵素燈處理 ... 30
4.3.2.1.1 製備已知濃度的未受熱與受熱原蟲懸浮液 ... 30
4.3.2.1.2 未受熱與受熱原蟲土壤混合液配置 ... 30
4.3.2.1.3 核酸染劑處理 ... 30
4.3.2.1.4 DNA 萃取、DNA 稀釋、qPCR 及定量指標 ... 31
4.3.2.2 滅菌土壤經過 1X-300X 稀釋後添加未受熱與受熱原蟲進行不 同濃度EMA 與鹵素燈處理 ... 31
4.3.2.2.1 製備已知濃度的未受熱與受熱原蟲懸浮液 ... 31
4.3.2.2.2 配置 1X-300X 土壤稀釋液 ... 32
4.3.2.2.3 1X-300X 土壤稀釋液之總懸浮固體測量 ... 32
4.3.2.2.4 未受熱與受熱原蟲土壤稀釋混合液配置 ... 32
4.3.2.2.5 核酸染劑處理 ... 33
4.3.2.2.6 DNA 萃取、DNA 稀釋、qPCR 及定量指標 ... 33
4.3.2.3 滅菌土壤經過 10X-200X 稀釋後添加未受熱與受熱原蟲進行 不同濃度EMA 與 PhAST Blue 處理 ... 34
4.3.2.3.1 製備已知濃度受熱與未受熱棘阿米巴原蟲 ... 34
4.3.2.3.2 配置 1X-300X 土壤稀釋液 ... 34
4.3.2.3.3 未受熱與受熱原蟲土壤稀釋混合液配置 ... 34
4.3.2.3.4 核酸染劑處理 ... 35
4.3.2.3.5 DNA 萃取、DNA 稀釋、qPCR 以及定量指標 ... 36
4.3.3 生物氣膠採樣器比較 ... 36
4.3.3.1 採樣策略 ... 36
4.3.3.2 分析方法 ... 37
xi
4.4 環境驗證與暴露初探 ... 38
4.4.1 土壤 ... 38
4.4.1.1 採樣策略 ... 39
4.4.1.2 分析方法 ... 41
4.4.2 空氣 ... 42
4.4.2.1 採樣策略 ... 42
4.4.2.2 分析方法 ... 43
4.4.3 水體 ... 44
4.4.3.1 採樣策略 ... 44
4.4.3.2 分析方法 ... 45
4.5 土壤與空氣環境因子測量 ... 47
4.5.1 土壤環境因子測量 ... 47
4.5.1.1 土壤含水量測定 ... 47
4.5.1.2 土壤 pH 值測定 ... 47
4.5.1.3 土壤異營性細菌濃度測定 ... 47
4.5.1.4 土壤植被覆蓋判定 ... 48
4.5.1.5 土壤結構判定 ... 49
4.5.2 空氣環境因子測量 ... 50
4.7 資料分析 ... 51
4.7.1 總和稀釋倍數 ... 51
4.7.2 統計分析 ... 51
第五章 結果 ... 53
5.1 方法開發 ... 53
xii
5.1.1 棘阿米巴原蟲囊體核酸染劑之選擇 ... 53
5.1.2 EMA-qPCR 定量土壤中活性棘阿米巴原蟲之方法開發 ... 57
5.1.3 生物氣膠採樣器比較 ... 65
5.1.4 總和稀釋倍數 ... 67
5.2 環境驗證 ... 69
5.3 暴露初探 ... 71
5.3.1 土壤中總與活性棘阿米巴原蟲濃度 ... 71
5.3.2 空氣中總與活性棘阿米巴原蟲濃度 ... 80
5.3.3 水體中總與活性棘阿米巴原蟲濃度 ... 83
5.3.4 土壤中環境因子測量及相關分析 ... 86
5.5.5 空氣中環境因子測量及相關分析 ... 103
第六章 討論 ... 108
6.1 方法開發 ... 108
6.1.1 棘阿米巴原蟲囊體核酸染劑之選擇 ... 108
6.1.2 EMA-qPCR 定量土壤中活性棘阿米巴原蟲之方法開發 ... 109
6.1.3 生物氣膠採樣器比較 ... 113
6.2 暴露初探 ... 113
6.2.1 土壤中棘阿米巴原蟲濃度與相關因子 ... 113
6.2.2 空氣中棘阿米巴原蟲濃度與相關因子 ... 115
6.2.3 水體中棘阿米巴原蟲濃度 ... 117
6.3 未來研究建議 ... 119
xiii
第七章 結論 ... 120
第八章 參考文獻 ... 122
附錄 ... 132
口試委員提問與回答 ... 148
xiv
圖次
圖一:方法開發之架構圖 ... 20
圖二:暴露初探與環境驗證之架構圖 ... 21
圖三:樣本於PhAST Blue 之位置 ... 35
圖四:PhAST Blue 之使用 ... 35
圖五:屏東縣車城鄉及恆春鎮洋蔥田(A)、台灣大學園藝農場蔬果/香草植物田(B)、 台灣大學農藝農場水稻田(C)之土壤採樣點位 ... 40
圖六:空氣採樣器位於田中之擺設 ... 43
圖七:土壤植被覆蓋率判定:(A)無植物、(B)有植物 ... 48
圖八:土壤結構判定- 堅硬(A)、堅硬但有破壞(B)、鬆軟(C) ... 49
圖九:未受熱(○)與 95℃受熱 20 min (●)棘阿米巴原蟲囊體培養於 Heat-killed Escherichia coli NNA plate 14 日之 Clean zone 直徑(n=3) ... 53
圖十:以200X 倒立式顯微鏡觀察經 Heat-killed Escherichia coli NNA plate 培養 6 日之未受熱(A)與 95℃受熱 20 min (B)棘阿米巴原蟲囊體 ... 54
圖十一:以200X 倒立式顯微鏡觀察經 Heat-killed Escherichia coli NNA plate 培養 14 日之未受熱(A)與 95℃受熱 20 min (B)棘阿米巴原蟲囊體 ... 54
圖十二:未受熱( )與 95℃受熱 20 min ( ) 棘阿米巴原蟲囊體經 EMA(A) 與PMA(B)處理並以 qPCR 分析之 DNA 量(n=3) (圖中相同英文字母記號者表 示經Scheffe's test 事後檢定未達統計顯著差異(P≧0.05)) ... 56
圖十三:未受熱(Unheated)與 75℃受熱 20 min (Heated)棘阿米巴原蟲於未滅菌且未 稀釋處理之土壤液中經0.0( )、2.3( )、23.0( )、46.0( )與 76.7( ) μg/mL EMA 處理並進行暗反應 5 或 10 min 後以 qPCR 所得 DNA 量(n=1) (*:樣本在 DNA 萃取步驟中流失) ... 57 圖十四:不稀釋(1X)與各稀釋倍數(10-300X)滅菌土壤稀釋液中總懸浮固體物濃
xv
度(n=3) ... 59 圖十五:不稀釋(1X)與各稀釋倍數(10-300X)滅菌土壤液中未受熱(A)與 75℃受 熱20 min(B)之棘阿米巴原蟲經 0( )、2.3( )、23.0( )與 46.0( ) μg/mL EMA 與鹵素燈處理並以 qPCR 分析之 DNA 量(n=3) (相同英文字母記 號者表示經Scheffe's test 事後檢定其 DNA 量未達統計顯著差異(P≧0.05)) 61 圖十六:10-200X 滅菌土壤稀釋液中未受熱(A)與 75℃受熱 20 min(B)之棘阿米巴
原蟲經0( )、2.3( )、23.0( ) μg/mL EMA 與 PhAST blue 處理並以 qPCR 分析之 DNA 量(n=3) (相同英文字母記號者表示經 Scheffe's test 事後檢 定其DNA 量未達統計顯著差異(P≧0.05)) ... 64 圖十七:MAS-100 (A)與 IOM (B)於洋蔥田中之架設 ... 65 圖十八:(A)採收期(101.3)與(B)採收後(101.4)之洋蔥田植栽狀況... 71 圖十九:蔬果田( , N=26)、香草植物田( , N=10)、蔬果香草植物田( ,
N=36)之土壤表層活性棘阿米巴原蟲濃度 ... 75 圖二十:蔬果田( , N=26)、香草植物田( , N=10)、蔬果香草植物田( ,
N=36)之土壤表層總棘阿米巴原蟲濃度 ... 76 圖二十一:洋蔥田(N=12)、蔬果/香草植物田(N=36)與水稻田(N=5)土壤表層活性
( )與總( )棘阿米巴原蟲濃度(柱狀圖中有相同英文字母記號者,表示 經Scheffe's test 事後檢定,其原蟲濃度未達統計顯著差異(P≧0.05) (a:經 Wilcoxon signed-rank test 檢定各農田總棘阿米巴原蟲與活性棘阿米巴原蟲濃 度之P 值) ... 78 圖二十二:不同地點(恆春 N=7、車城 N=5)與採收時期(採收期 N=9、採收後 N=3) 之洋蔥田空氣中活性( )與總( )棘阿米巴原蟲濃度(柱狀圖中有相同英 文字母記號者,表示經Wilcoxon rank sum test 檢定,其原蟲濃度未達統計顯 著差異(P≧0.05)) (a:經 Wilcoxon signed-rank test 檢定各農田總棘阿米巴原蟲 與活性棘阿米巴原蟲濃度之P 值) ... 82
xvi
圖二十三:冷卻水塔(N=1)、廢水處理場-進水管(N=1)、廢水處理場-曝氣池(N=1)、
廢水處理場-放流管(N=1)、水稻田(N=6)、農田溝渠(N=6)之水體中活性( ) 與總( )棘阿米巴原蟲濃度(a:經 Wilcoxon signed-rank test 檢定總棘阿米巴 原蟲與活性棘阿米巴原蟲濃度之P 值) ... 85 圖二十四:蔬果/香草植物田土壤表層活性棘阿米巴原蟲濃度與土壤 pH 值之散佈
圖 (N=36) ... 94 圖二十五:蔬果/香草植物田土壤表層活性棘阿米巴原蟲濃度與土壤結構之散佈圖
(N=36) ... 94 圖二十六:水稻田土壤表層活性棘阿米巴原蟲濃度與土壤異營性細菌濃度之散佈 圖(N=5) ... 96 圖二十七:所有農田土壤表層活性(A)與總(B)棘阿米巴原蟲濃度與土壤 pH 值之散 佈圖 (N=53) ... 98 圖二十八:所有農田土壤表層活性(A)與總(B)棘阿米巴原蟲濃度與土壤異營性細
菌濃度之散佈圖 (N=53) ... 99 圖二十九:所有農田土壤表層活性(A)與總(B)棘阿米巴原蟲濃度與土壤含水量之
散佈圖 (N=53) ... 100 圖三十:洋蔥田空氣中總棘阿米巴原蟲濃度與土壤植被覆蓋(1:無植物;2:有植
物)之散佈圖(N=12) ... 107
xvii
表次
表一:棘阿米巴原蟲感染之前置因子 ... 4
表二:土壤環境中棘阿米巴原蟲監測 ... 7
表三:空氣中棘阿米巴原蟲採樣方法 ... 11
表四:空氣中棘阿米巴原蟲之影響因子 ... 12
表五:屏東縣車城鄉與恆春鎮洋蔥田之採樣點經緯度座標 ... 38
表六:台灣大學園藝農場蔬果/香草植物田之採樣點經緯度座標 ... 39
表七:台灣大學農藝農場水稻田之採樣點經緯度座標 ... 39
表八:以不同生物氣膠採樣器於洋蔥田採樣並以qPCR 定量空氣中總與活性棘阿 米巴原蟲之Cycle threshold (Ct)值與濃度(n=1) ... 66
表九:不同介質與採樣地點之樣本於qPCR 分析之總和稀釋倍數 ... 68
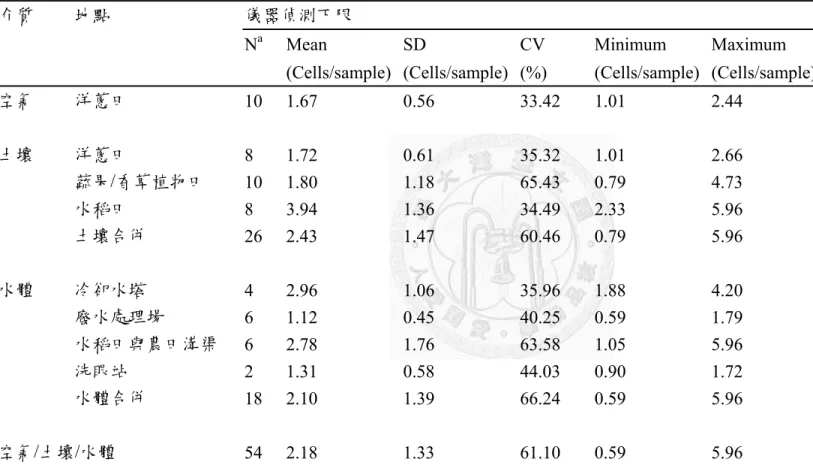
表十:不同環境介質中棘阿米巴原蟲濃度及活性比 ... 70
表十一:不同地點與採收期間洋蔥田土壤表層活性與總棘阿米巴原蟲濃度 ... 72
表十二:台大園藝農場蔬果/香草植物田之植栽狀況(101.7) ... 74
表十三:台大農藝農場水稻田之植栽狀況(101.7) ... 77
表十四:不同類型農田土壤中活性與總棘阿米巴原蟲濃度 ... 79
表十五:不同地點與採收期間洋蔥田空氣中活性與總棘阿米巴原蟲濃度 ... 81
表十六:不同水體環境陽性檢出樣本之活性與棘阿米巴原蟲濃度 ... 84
表十七:洋蔥田、蔬果/香草植物田與水稻田土壤環境因子測量結果 ... 87
表十八:恆春與車城洋蔥田土壤中環境因子測量結果 ... 89
表十九:採收期與採收後洋蔥田土壤環境因子量測結果 ... 91
表二十:洋蔥田土壤表層棘阿米巴原蟲濃度與環境因子之Spearman's rank correlation coefficients (r) (N=12) ... 92
表二十一:蔬果/香草植物田土壤表層棘阿米巴原蟲濃度與環境因子之 Spearman's rank correlation coefficients (r) (N=36) ... 93
xviii
表二十二:水稻田土壤表層棘阿米巴原蟲濃度與環境因子之Spearman's rank correlation coefficients (r) (N=5) ... 95 表二十三:所有農田土壤表層總與活性棘阿米巴原蟲濃度與土壤環境因子之
Spearman's rank correlation coefficients (r) (N=53) ... 97 表二十四:以Multiple linear regression with stepwise procedure 評估影響農田土壤
表層棘阿米巴原蟲濃度及其活性比之環境因子(N=53) ... 102 表二十五:恆春與車城洋蔥田空氣中環境因子 ... 103 表二十六:採收期與採收後洋蔥田空氣中環境因子 ... 104 表二十七:洋蔥田空氣中棘阿米巴原蟲濃度與土壤表層原蟲及其他環境因子之
Spearman’s rank correlation coefficients (r) (N=12) ... 106
1
第一章 前言
1.1 研究背景
棘阿米巴原蟲(Acanthameoba spp.)為一單細胞生物,廣泛存在於環境當中,可 導致棘阿米巴角膜炎與肉芽腫性阿米巴腦膜炎。目前在各環境當中,土壤與空氣 中監測之相關文獻主要採培養法配合型態學鑑定或者以聚合酶連鎖反應
(polymerase chain reaction, PCR)進行定性檢測,雖然知道其存在於環境當中,其定 量資料,無法評估暴露風險。水體已有培養法配合形態學鑑定或以即時定量聚合 酶連鎖反應(real-time quantitative Polymerase chain reaction, qPCR)進行定量,但 qPCR 無法分別死菌與活菌造成水體中棘阿米巴原蟲濃度的高估。
本研究以qPCR 為基礎並結合 ethidium monoazide (EMA) (EMA-qPCR)建立定 量環境中活性棘阿米巴原蟲之方法後,在土壤、空氣、水體當中進行環境驗證與 暴露初探,同時探討土壤跟空氣中棘阿米巴原蟲濃度與環境因子之相關性,試著 找出在土壤與空氣中影響棘阿米巴原蟲濃度的顯著相關環境因子。
1.2 文獻回顧
1.2.1 棘阿米巴原蟲(Acanthamoeba spp.)
棘阿米巴原蟲為自由型阿米巴原蟲(free-living amoeba, FLA)的一種,廣泛存在 自然環境當中,於土壤(Kawaguchi et al., 2009; Lorenzo-Morales et al., 2005 ;
Rezaeian et al., 2008)、以及大氣(Kingston & Warhurst, 1969;Mulec et al., 2012;
Rivera et al., 1987;Rivera et al., 1994;Rogerson & Detwiler, 1999)、自然以及人工 水體(Chang & Wu, 2010;Chang et al., 2010;Gianinazzi et al., 2009;Kilvington, 2004;
Hsu et al., 2009)中都有文獻顯示曾經分離出棘阿米巴原蟲。棘阿米巴原蟲具有兩種 型態:滋養體(trophozoite)及囊體(cyst)。當處於適合生長的環境時,它會呈現滋養 體並進行攝食及分裂,而其型態依物種或基因型的不同,直徑大小介於12-35 μm,
2
其表面會伸出手指狀的偽足(acnthopodia)以幫助移動及攝食,於細胞內部則具有中 央核仁之囊泡狀細胞核(Khan, 2006)。當環境惡劣時(例:缺乏食物來源、乾燥、極 端溫度或pH 值),會轉變成囊體而進入休眠狀態,直徑大小介於 5-20 μm (Khan, 2006),並具有多皺紋外層及多邊形或圓形內層所組成的雙層外壁結構(Visvesvara et al., 2007)。囊體型態的原蟲可以抵抗惡劣的環境包括 80℃以上高溫(Storey et al., 2004)、殺蟲劑(biocide) (Lee et al., 2007;Lloyd et al., 2001)、氯(King et al., 1988)以 及抗生素(Lloyd et al., 2001;Turnera et al., 2000)。另一方面,棘阿米巴原蟲也是一 些人類致病菌的自然宿主,如:退伍軍人菌(Legionella spp.) (Rowbotham, 1980)以 及幽門螺旋桿菌(Helicobacter pylori) (Winiecka-Krusnell et al., 2002)。Greub &
Raoult (2004)的回顧文獻中則指出,於棘阿米巴原蟲體內複製增生後之退伍軍人菌,
其移動性、毒性以及耐藥性均較高,進而增加人類感染此病原細菌的風險。
1.2.2 棘阿米巴原蟲導致之疾病及與土壤之相關性
棘阿米巴原蟲除了為人類部分致病菌的自然宿主外,也可透過眼睛及皮膚傷 口接觸,或以吸入的方式進入人體而造成感染,如導致眼睛劇烈疼痛及失明的棘 阿米巴角膜炎(Acanthamoeba keratitis, AK)、使皮膚潰爛的棘阿米巴皮膚炎
(Cutaneous acanthamebiasis)、以及高致死率的肉芽腫性阿米巴腦膜炎
(Granulomatous amebic encephalitis, GAE)。被棘阿米巴原蟲感染的人數日益增加,
Diaz (2011)的文獻指出,目前世界上每年有 10,000 件感染棘阿米巴角膜炎的案例,
且至2011 年為止全球至少有 250 件肉芽腫性阿米巴腦膜炎的病例出現。Chen 等人 (2004)則指出,根據國立台灣大學附設醫院 1987 年至 2001 年的診斷紀錄,所有微 生物導致角膜炎而需進行角膜置換術的病患當中有13.9%是棘阿米巴原蟲造成的 感染。在2009 年台灣也出現了第一個感染肉芽腫性阿米巴腦膜炎的病例,此個案 是一位63 歲的男性農夫,在農田工作時不小心跌入農田溝渠,嗆入泥水而導致感 染(Sheng et al., 2009)。
3
回顧土壤相關活動與棘阿米巴原蟲感染的病例報告,彙整如表一,發現國內 外的病例當中(1)感染棘阿米巴原蟲之病例的職業當中農夫佔 30-100% (Ma et al., 1981;Srinivasan et al., 2003;Manikandan et al., 2004;MBBS & Mahgoub, 2006;
Babu & Murthy, 2007;Sun et al., 2006;Shi et al., 2009;Tien & Sheu, 1999;Sheng et al., 2009),顯示農夫確實有被棘阿米巴原蟲感染之風險。(2) 30%~ 92%感染棘阿米 巴角膜炎的病例有眼睛創傷的紀錄(Ma et al., 1981;Radford et al., 1998;Sharma et al., 2000;Srinivasan et al., 2003;Manikandan et al., 2004;Sun et al., 2006;Shi et al., 2009;Tien & Sheu, 1999;Chen et al., 2004;Lin et al., 2009),顯示創傷是感染棘阿 米巴角膜炎的前置因子之一,其中造成創傷的物體包括植物(vegetative matter) (Ma et al., 1981;Sharma et al., 2000;Manikandan et al., 2004;Sun et al., 2006;Shi et al., 2009)、石頭(Sharma et al., 2000;Manikandan et al., 2004)、粉塵(Sharma et al., 2000;
Manikandan et al., 2004;Sun et al., 2006;Shi et al., 2009)、昆蟲(Manikandan et al., 2004)、金屬(Manikandan et al., 2004)、非金屬(Srinivasan et al., 2003)等。而在 Sharma 等人(2000)的研究當中提到,病患若非隱形眼鏡配戴者,醫生常會誤判是真菌造成 角膜炎感染,而不認為感染棘阿米巴角膜炎,誤診的結果通常會導致比較不好的 預後。
總結上述,從事土壤相關活動的人有感染棘阿米巴原蟲的風險。若是在農田 工作,其暴露來源包括空氣、土壤及水體。以下分別針對土壤、空氣及水體進行 棘阿米巴原蟲的流佈情形之文獻回顧,並針對土壤及空氣環境因子探討影響其流 佈的相關性。
4
表一:棘阿米巴原蟲感染之前置因子
創傷
國家 病例 疾病 農夫 (%) 數量(%) 描述 參考文獻
美國 10 AK 3(30%) 6(60%) 綑綁大麥時眼睛遭到侵入,使用不乾淨的水清洗 Ma et al., 1981
英國 同上 AK 同上 同上 右眼遭到樹枝侵入 Ma et al., 1981
德國 同上 AK 同上 同上 使用電鋸切木頭時,眼睛遭到木屑侵入 Ma et al., 1981
英國 12 AK-非隱形眼 鏡配戴者
未知 11(92%) 花園(4)、營建工地(3)、牧場(2)、釣魚(1)、航海(1) Radford et al., 1998
印度 39 AK-非隱形眼 鏡配戴者
未知 15(38.5%) 15 位病人的眼睛有遭到植物、石頭或粉塵侵入,其
中3 人的眼睛遭物體侵入後觸碰到不乾淨的水
Sharma et al., 2000
印度 103 AK-非隱形眼 鏡配戴者
36(34.9%) 52(50.5%) 52 個病人的眼睛遭到非金屬物體入侵 Srinivasan et al., 2003
印度 33 AK-非隱形眼 鏡配戴者
16(50%) 18(56.3%) 樹枝(5)、粉塵(5)、水稻葉(2)、昆蟲(2)、木柴(1)、釘 子(1)、有機物質 (1)、石灰物質(1)
Manikandan et al., 2004
蘇丹 6 AK 3(50%) 未知 MBBS & Mahgoub,
2006
英國 1 AK 1(100%) 未有創傷紀錄 Babu & Murthy,
2007 中國 20 AK 6(30%) 6(30%) 角膜塑形術(8)、軟式隱形眼鏡(4)、眼睛有遭到植物、
粉塵入侵或發病前有暴露到游泳池的水(6)、無明確 風險因子(2)
Sun et al., 2006
5
中國 22 AK 13 (59%) 20(91%) 植物或粉塵(13)、家禽飼養(6)、石油職業暴露(1)、
無明顯因子(2)
Shi et al., 2009 臺灣 3 AK+真菌感染 1(33%) 1(33%) 在泥水坑工作時右眼遭到外物入侵 Tein and Sheu,
1999
臺灣 13 AK 未知 1(7.7%) 遭到非植物的外物入侵 Chen et al., 2004
臺灣 11 AK 未知 2(5.5%) 在學校營建工地,眼睛遭到外物入侵 Lin et al., 2008
臺灣 1 GAE 1(100%) 1(100%) 未提及是否創傷,摔入水溝吸嗆到泥水 Sheng et al., 2009
6
1.2.3 棘阿米巴原蟲於土壤中之流佈及其影響因子
回顧土壤中棘阿米巴原蟲環境監測之相關研究(表二),其陽性檢出率從 37%至 100%,顯示棘阿米巴原蟲廣泛地存在於在各類土壤當中。而過去研究皆探討其陽 性檢出率,但並未提到其相關定量數據(Sawyer, 1989;Rodriguze-Zaragozai & Garcia, 1997;Khan & Paget., 2002;Robinson et al., 2002;Booton et al., 2004;Brown &
Smirnov, 2004;Tsvetkova et al., 2004;Kilic et al., 2004;Lorenzo-Morales et al., 2005;
Rodriguez-Zaragoza et al., 2005;Bamforth, 2007;Ertabaklar et al., 2007;Lara et al., 2007;Bamforth, 2008;Rezaeian et al., 2008;Rivera & Adao, 2008;Kawaguchi et al., 2009;Matsuo et al., 2010),另外也發現國內目前沒有土壤棘阿米巴原蟲之流佈研 究。
土壤採樣環境包括農田(Sawyer, 1989;Rodriguez-Zaragoza et al., 2005)、花園 (Ertabaklar et al., 2007)、海灘(Sawyer, 1989;Booton et al., 2004;Lorenzo-Morales et al., 2005)、公園(Khan & Paget., 2002;Bamforth, 2007;Rezaeian et al., 2008;
Kawaguchi et al., 2009;Matsuo et al., 2010)、花盆(Kilic et al., 2004)、草原(Brown &
Smirnov, 2004)、遭到 PAH 汙染的土壤(Lara et al., 2007)等等。
而檢出之分析方法大部分是使用培養法搭配型態學鑑定(Sawyer, 1989;
Rodriguze-Zaragozai & Garcia, 1997;Robinson et al, 2002;Booton et al., 2004;Brown
& Smirnov, 2004;Tsvetkova et al., 2004;Kilic et al., 2004;Lorenzo-Morales et al., 2005;Rodriguez-Zaragoza et al., 2005;Bamforth, 2007;Ertabaklar et al., 2007;Lara et al., 2007;Bamforth, 2008;Rezaeian et al., 2008),然也有少部分研究使用 PCR 進 行棘阿米巴原蟲鑑定(Khan & Paget., 2002;Kilic et al., 2004;Lorenzo-Morales et al., 2005;Lara et al., 2007;Rivera & Adao, 2008;Kawaguchi et al., 2009;Matsuo et al., 2010)。
7
表二:土壤環境中棘阿米巴原蟲監測
國家 採樣地點 分析方法/鑑定 重要結果 參考文獻
美國 農田、山溝、海灘 培養法/型態學 Acanthamoeba 樣本檢出率為 100% (7/7) Sawyer, 1989
墨西哥 山谷 培養法/型態學 Acanthamoeba 在所有檢出的 amobea 當中佔 9.4% Rodriguze-Zaragozai & Garcia, 1997
英國 公園、荒地…等 PCR Acanthamoeba 在土壤樣本檢出率為 100% (10/10) Khan & Paget, 2002 澳洲 各地乾旱土壤 培養法/型態學 Acanthamoeba 在土壤樣本檢出率, group 2 為 92%
(24/26)、group 3 為 88% (23/26)
Robinson et al., 2002
美國 海灘 培養法/型態學 Acanthamoeba 沙子中檢出率為 38% (23/60) Booton et al., 2004
蘇格蘭 草原 培養法/型態學 檢出含有棘狀偽足的naked amoeba 在所有樣本中佔 84%
(126/150)
Brown & Smirnov, 2004
保加利亞 不同類型土壤 培養法/型態學 Acanthamoeba 在土壤樣本檢出率為 100% (11/11)、在沙 土樣本檢出率為91.66% (22/24)
Tsvetkova et al., 2004
土耳其 醫院中花盆 培養法/型態學/PCR Acanthamoeba 在花盆土壤檢出率為 100% (28/28) (16/16) Kilic et al., 2004 西班牙 海灘, 全島各地土壤 培養法/型態學/PCR Acanthamoeba 在海灘檢出率為 42% (21/50), 在土壤檢
出率為37.7% (43/114)
Lorenzo-Morales et al., 2005
以色列 試驗農場 培養法/型態學 Acanthamoeba 為最常檢出的屬之一 Rodriguez-Zaragoza et al., 2005 美國 森林公園 培養法/型態學 在15 個採樣點當中有 14 採樣點檢出 Acanthamoeba Bamforth, 2007
土耳其 花園 培養法/型態學 花園土壤樣本中100%檢出 Acanthamoeba (1/1) Ertabaklar et al., 2007 西班牙 遭到PAH 汙染的土壤 培養法/型態學/PCR 在遭到PAH 污染的土壤樣本當中都有檢出 Acanthamoeba Lara et al., 2007 美國 高原 培養法/型態學 在5 種不同類型的土壤當中皆有檢出 Acanthamoeba Bamforth, 2008 伊朗 眼科中心, 公園 培養法/型態學 Acanthamoeba 在土壤中檢出率為 100% (5/5) Rezaeian et al., 2008 菲律賓 菲律賓各地 PCR Acanthamoeba 在土壤檢出率為 100% (10/10) Rivera & Adao, 2008 日本 公園 PCR Acanthamoeba 在公園土壤檢出率為 97% (69/71) Kawaguchi et al., 2009 日本 公園 PCR Acanthamoeba 在公園土壤中檢出率為 92% (69/75) Matsuo et al., 2010
8
目前並無任何研究直接探討影響棘阿米巴原蟲在土壤流佈的因子,故以Type 1 amoeba (例:Acanthamoeba、Filamoeba )或以 amoeba 進行文獻回顧。
Rodriguez-zaragoza & Garcia (1997)於墨西哥沙漠之仙人掌 Escontriu chiotilla 附近土壤進行採樣,並以MPN 方法定量土壤中 amoeba,作者發現雨季表層 10 與 30 cm 土壤中 amoeba 濃度為乾季的 2-3 倍與 1 倍;而在表層 10 cm 土壤中 amoeba 濃度則大於表層30 cm,符合原蟲的生存範圍在表層 15 cm 內的假設。
Rodriguez-Zaragoza 等人(2005)在以色列内蓋夫沙漠當中灌木 Hammada scoparia 和 Atriplex halimus 附近進行土壤採樣,並以 MPN 搭配顯微鏡觀察定量土
壤中amoeba,作者發現濕季的總原蟲量顯著高於乾季,另外發現 Type 1 amoeba 相較於Type 2、3、4 可以正常生活在極端的環境當中(作者並未說明何謂極端環境),
且可以適應含水量較低(本研究範圍約於 1.4-9%)的環境。
Rodriguez-Zaragoza 等人(2007)在美國新墨西哥州分水嶺的螞蟻窩進行土壤採 樣,並以MPN 搭配顯微鏡觀察定量土壤中 amoeba。作者發現,相對於沒有螞蟻 存在的土壤當中可發現不同型態的amoeba,有 Pogonomyrmex rugosus 和
Myrmecocystus depilis 這兩種螞蟻存在的土壤擁有較低的含水量(本研究範圍約於
2-6%)以及有機物質(本研究範圍約於 1-2.5%),且只會出現 Type 1 amobeae。
總結上述,目前還沒有土壤中棘阿米巴原蟲的定量資料,而影響Type 1 amoeba (例:Acanthamoeba、Filamoeba)或 amoeba 在土壤中流佈的環境因子包括季節、土 壤含水量、土壤深度以及土壤有機物質含量。
9
1.2.4 棘阿米巴原蟲於空氣中之流佈及其影響因子
整理過去探討空氣中棘阿米巴原蟲相關文獻,將採樣分析法整理於表三,而 其影響其空氣中流佈因子則整理於表四。
在採樣分析方法部分,文獻顯示其採樣介質包括濾紙(Rogerson & Detwiler, 1999)、培養基(Kingston & Warhurst, 1969)以及液體(Rodriguez-Zaragoza &
Magana-Becerra, 1997;Rivera et al., 1994;Rivera et al., 1987)三類,採樣高度在 100- 1100 cm 之間,採樣流量介於 15- 570 LPM 之間,配合採樣時間(10-1525 min),其 採樣體積介於978- 24400 L 之間,至於分析方法皆是使用培養法搭配型態學或 PCR
鑑定,其中有1 篇文獻進一步配合採樣體積以計算空氣中棘阿米巴原蟲濃度
(Kingston and Warhurst, 1969)。
Rogerson & Detwiler (1999)使用 0.8-2 μm 及 2-20 μm 濾紙在校園草地採樣,在 所有分離出的22 株原蟲當中,有 28%為 naked amoeba,其中最常分離出的屬為 Acanthamoeba,而空氣中平均原蟲濃度為 0.25 cyst/ m3,另外作者指出在風速較強 時空氣中會有較多的土壤顆粒以及原蟲,且雨季時可洗掉空氣中的原蟲,並增加 土壤的溼度使得原蟲難以揚起;而空氣中原蟲種類也可以在土壤中發現,顯示此 研究大部分的原蟲是來自於土壤。
Rodriguez-Zaragoza & Magana-Becerra (1997)則使用 50 mL sterilized saline solution 搭配 Impinger 在都市、郊區及農村離地面 200 cm 處進行 60 min 採樣,採 樣體積共978 L,結果發現,在 23 個 Acanthamoeba strains 中,有超過 60%來自都 市及郊區或是來乾熱的季節。作者認為都市以及郊區有這種高檢出率,其原因是 垃圾直接傾倒於街上或者地上沒有鋪路(Lack of pavement)。
Rivera 等人(1994)使用 Bold's Basal Medium 搭配 Impinger 在墨西哥 4 個靠近氣 象監測站的地點,距離地面200、500 cm 進行 150 min 採樣,採樣體積共 2940 L,
作者發現空氣中阿米巴原蟲來源為土壤,且風速(本研究範圍約介於 2-5.4 m/s)、SO2
(本研究範圍約介於 0.05-0.1 ppm)與阿米巴原蟲檢出數量呈正相關(r=0.79、P<0.05;
10
r=0.73、P<0.05),相對溼度(本研究範圍約介於 25-47%)、溫度(本研究範圍約介於 18-23.5℃)跟阿米巴原蟲檢出數量無相關性(r=-0.61、P>0.05;r=-0.03、P>0.05),而 影響阿米巴原蟲高檢出率及生物多樣性的原因可能為沙塵暴頻率較高、靠近開放 式的垃圾場及大面積裸土。
Rivera 等人(1987)使用 50 mL sterilized straw infusion (4:100; pH 7.0)搭配 Impinger 在大學、Cuemanco 運河附近距離地面 100-1100 cm 進行 60 min 採樣,採 樣體積共900 L,作者發現 3 個空氣樣本中皆檢出囊體型態的棘阿米巴原蟲。
Kingston & Warhurst (1969)的研究當中,使用 Silt sampler 與塗有 Klebsiella aerogenes 的培養基於實驗室、有換氣設備的病房、室外進行 10 min 採樣,採樣體
積共5700 L,作者發現在室外空氣平均棘阿米巴原蟲濃度為 0.055 cells/m3。 總結上述,目前尚未有研究在農田環境中定量空氣中棘阿米巴原蟲,而空氣 中棘阿米巴原蟲相關文獻大部分則以定性為主,只有一篇文獻以培養法進行定量。
而在影響因子部分,雖有文獻指出空氣中阿米巴原蟲來自於土壤,但是尚未有研 究證明空氣與土壤中棘阿米巴之間的相關性,而影響棘阿米巴原蟲在空氣中分布 之因子包括風速、雨、相對溼度、SO2、溫度、沙塵暴、車輛、靠近垃圾場或大面 積裸土等等。
11
表三:空氣中棘阿米巴原蟲採樣方法
NA:不適用
國家 採樣器 採樣介質 採樣地點 採樣高
度(cm)
採樣流量 (LPM)
採樣時間 (min)
採樣體積 (L)
分析/鑑定 參考文獻 美國 Model 200
virtual impactor
0.8-2 μm, 2-20 μm 濾紙
學校草地 140 16 356-1525 5700-24400 培養/型態學 Rogerson &
Detwiler, 1999 墨西哥 Impinger 50 mL Sterilized
saline solution
都市、郊區、
農村
200 16.3 60 978 培養/型態學 Rodriguze-Zarag oza &
MaganaBecerra, 1997
墨西哥 Impinger Bold'sBasal Medium
4 個靠近氣象 監測站的地點
200, 500 19.6 150 2940 培養/型態學 Rivera et al., 1994
墨西哥 Impinger 50 mL Sterilized straw infusion (4:100; pH 7.0)
大學、
Cuemanco 運 河附近
100-1100 15 60 900 培養/型態學 Rivera et al., 1987
英國 Slit sampler 塗有 K. aerogenes 的培養基
實驗室、有換 氣設備的病 房、室外
未說明 570 10 5700 培養/型態學 Kingston &
Warhurst, 1969 英國 Settle plates 塗有 K. aerogenes
的培養基
實驗室、有換 氣設備的病 房、室外
未說明 NA 420 NA 培養/型態學 Kingston &
Warhurst, 1969
12
表四:空氣中棘阿米巴原蟲之影響因子
環境因子 採樣地點 重要結果 參考文獻
土壤 學校草地 空氣中原蟲種類也可以在土壤中發現,顯示此研究大部分
的原蟲是來自於土壤
Rogerson & Detwiler, 1999
4 個靠近氣象監測站的地點 空氣中的阿米巴原蟲來源是土壤 Rivera et al., 1994
風 學校草地 在風速較強的時候空氣中會有較多的土壤顆粒以及原蟲 Rogerson & Detwiler, 1999 4 個靠近氣象監測站的地點 風速與阿米巴原蟲檢出數量呈正相關(r=0.79;P<0.05) Rivera et al., 1994
雨 學校草地 雨可以洗去空氣中的原蟲,並提高土壤的溼度防止風將土
壤中的原蟲揚起
Rogerson & Detwiler, 1999
相對溼度 都市、郊區、農村 超過60%的棘阿米巴原蟲來自最熱以及最乾燥的月份 (4
月-5 月)
Rodriguez-Zaragoza &
Magana-Becerra, 1997 4 個靠近氣象監測站的地點 在低相對濕度時,棘阿米巴原蟲的檢出率最高 Rivera et al., 1994 4 個靠近氣象監測站的地點 相對溼度與阿米巴原蟲檢出數量無相關性(r=-0.61;
P>0.05)
Rivera et al., 1994
SO2 4 個靠近氣象監測站的地點 SO2與阿米巴原蟲檢出數量呈正相關(r=0.73;P<0.05) Rivera et al., 1994
溫度 都市、郊區、農村 超過60%的棘阿米巴原蟲來自最熱以及最乾燥的月份 (4
月-5 月)
Rodriguez-Zaragoza &
Magana-Becerra, 1997
13
4 個靠近氣象監測站的地點 溫度與阿米巴原蟲檢出數量無相關性(r=-0.03;P>0.05) Rivera et al., 1994
沙塵暴 4 個靠近氣象監測站的地點 阿米巴原蟲高檢出率及生物多樣性是因為沙塵暴頻率較
多的關係
Rivera et al., 1994
車輛 4 個靠近氣象監測站的地點 阿米巴原蟲高檢出率及生物多樣性是因為此區域有大型
車頻繁經過造成微生物揚起
Rivera et al., 1994
垃圾場 都市、郊區、農村 都市以及郊區擁有較高檢出率是因為垃圾直接傾倒於街
上
Rodriguez-Zaragoza &
Magana-Becerra, 1997 4 個靠近氣象監測站的地點 阿米巴原蟲高檢出率及生物多樣性是因為此區域靠近開
放式的垃圾場
Rivera et al., 1994
大面積裸土 都市、郊區、農村 都市以及郊區擁有較高檢出率是因為沒有路面(lack of
pavement)
Rodriguez-Zaragoza &
Magana-Becerra, 1997 4 個靠近氣象監測站的地點 阿米巴原蟲高檢出率及生物多樣性是因為此區域有大面
積的裸土
Rivera et al., 1994
14
1.2.5 棘阿米巴原蟲於水體中之流佈
過去研究曾從游泳池(Gianinazzi et al., 2009)、自來水(Kilvington, 2004)、溫泉 (Hsu et al., 2009)、冷卻水塔(Declerck et al., 2007)、瓶裝水以及汙水(Tsvetkova et al., 2004)、實驗室洗眼站(Paszko-Kolva et al., 1998)、水稻田(Liang et al., 2010)分離出 棘阿米巴原蟲
在定量研究上,Behets等人(2007)在比利時發電廠之冷卻水塔以most probable number (MPN)方法在44℃定量Thermophilic Acanthamoeba,結果發現,發電廠冷卻 水塔可培養性棘阿米巴原蟲濃度介於0-452 cells/L之間。Chang等人(2010)於台灣護 理之家冷卻水塔以qPCR方法定量總棘阿米巴原蟲,其平均濃度為546.7 (3.4-5034.0) cells/L,另Chang & Wu (2010)在台灣冷卻水塔以qPCR定量總棘阿米巴原蟲,其平 均濃度為1495 (118-4211.0) cells/L。
總結上述,已有多篇文獻棘從不同水體當中分離出阿米巴原蟲,但其定量資 訊卻只限於冷卻水塔當中。另外,雖然有研究在水稻田分離出棘阿米巴原蟲(Liang et al., 2010),但其濃度目前仍未知,難以量化農夫在工作環境之暴露風險。
15
1.2.6 環境棘阿米巴原蟲之偵測與定量方法
分離環境樣本中棘阿米巴原蟲之傳統方法,是將樣本於前處理後放置於覆有 大腸桿菌(Escherichia coli)之非營養培養基上(non nutrient agar, NNA),經數日培養、
分離純化等步驟之後,於顯微鏡下觀察(Booton et al., 2004)或利用 PCR(Schroeder et al., 2001)等技術進行棘阿米巴原蟲之鑑定。但是以培養搭配顯微鏡或 PCR 等方法 會有耗時、主觀判定差異以及無法提供量化數據等缺點(Qvarnstrom et al., 2006)。
在定量環境棘阿米巴原蟲上,過去文獻使用MPN 並搭配上述之培養及分離純 化方法去定量環境中的棘阿米巴原蟲(Behets et al., 2007),仍然有耗時、操作流程 複雜(Chang et al., 2010)以及主觀判定的缺點。Qvarnstrom 等人(2006)以分子生物技 術為基礎,開發針對棘阿米巴原蟲18S rRNA gene 之 qPCR 方法,而目前也有文獻 將此方法應用於定量環境水體中之棘阿米巴原蟲(Chang & Wu, 2010;Chang et al., 2010)。然而,qPCR 雖然具有特異性、敏感性及快速等特點,但它還是有兩個缺 點,一是環境中可能有PCR 之抑制物存在,會干擾 PCR 正常放大;二是無法分辨 活菌跟死菌,造成環境中棘阿米巴原蟲濃度的高估。
環境樣本中可能含有腐植質(humic substances)或黃腐酸(fulvic acid),這些物質 可在DNA 萃取過程中與 DNA 一起被萃取出來,並干擾 qPCR 正常放大 DNA 片段,
而影響qPCR 之定量結果(Wilson, 1997)。目前排除 qPCR 抑制物干擾之方法,包括 使用稀釋DNA (Wilson, 1997)、於 PCR 試劑中加入牛血清白蛋白(bovine serum albumin, BSA),其可以減輕抑制物干擾聚合酶鍵結的問題(Kreader, 1996)以及將萃 取的DNA 再次進行純化等方法(LaMontagne et al., 2002)。而 DNA 稀釋雖具有方法 簡單、不需額外繁雜步驟等優點,但是有可能會因DNA 過度稀釋而導致 DNA 濃 度低於qPCR 偵測下限(Wilson, 1997),但實際上 DNA 稀釋應用廣泛,並證實可解 決於定量環境中棘阿米米巴原蟲時,抑制物干擾qPCR 放大的問題 (Chang et al.
2010; Chang and Wu., 2010)。
16
近年來ethidium monoazide (EMA) 與 propidium monoazide (PMA)廣泛使用於 qPCR 技術當中(Bae & Wuertz, 2009;Brescia et al., 2009;Taskin et al., 2011;Varma et al., 2009;Wagner et al., 2008;Pisz et al., 2007),這兩種核酸染劑可藉由細胞是否 完整來區別死菌與活菌,核酸染劑會進入細胞膜破損的微生物,且經由光照後與 DNA 產生共價鍵結,抑制後續的 PCR 放大;而細胞膜完整的微生物擁有抵抗核酸 染劑的屏障,未與DNA 產生共價鍵結之核酸染劑經光照後則會被水解(Nogva et al., 2003)。然而核酸染劑在不同微生物之間有不同的處理方式,故必須最佳化其染劑 種類及濃度、反應時間、光源種類與光照時間等。
在光源種類當中,近年來LED 燈成為鹵素燈的另一種替代燈源,Vesper 等人 (2009)係第一篇以 LED 燈進行核酸染劑處理的研究,作者將未受熱與受熱的真菌 過濾於濾紙上並以PMA 與 PBS(控制組)處理後,經 LED 燈系統光照 10 min,並以 qPCR 定量,結果顯示,經 PMA 處理的受熱真菌,其 Ct 值與控制組相差約 6-9,
證實此方法可應用於區分未受熱與受熱真菌。此LED 燈的最佳波長於 470 nm 附 近(範圍介於 445-485 nm)擁有較窄且平均的波長分布,且使用菲涅爾透鏡(Fresnel lens)進行光的集中,後來此 LED 系統商品化為 PhAST Blue 系統(GenIUL, Barcelona, Spain),並應用於近期研究當中(Agustı et al., 2010;Elizaquível et al., 2012;Fittipaldi et al., 2011;Sánchez et al., 2011)。相對於鹵素燈擁有大範圍波長(300-800 nm)、產 熱、需要手動進行處理,PhAST Blue 擁有較窄的波長(464-476 nm)、不產熱以及自 動化等優點(此儀器放出的藍光最佳波長位於 465 nm 附近,而 EMA 與 PMA 核酸 染劑的所需要的最大吸收波長為456 和 464 nm) (Fittipaldi et al., 2012),且樣本與 燈源距離固定(<2 cm),不會造成手動誤差,而目前尚未有研究評估鹵素燈和 PhAST Blue 對於核酸染劑的處理是否造成差異。
回顧近年來使用核酸染劑搭配qPCR 定量活性棘阿米巴原蟲相關文獻,呂氏 (2010)開發以 2.3 μg/mL (5.5 μM) EMA 或(4.5 μM)PMA 處理 5 min 搭配 500W 鹵素 燈距離15 cm 光照 20 min 後進行 qPCR 可區別未受熱與經 75℃受熱之棘阿米巴原
17
蟲滋養體,並以顯微鏡搭配23 μg/mL EMA 和 PMA 觀察經染色後的棘阿米巴原蟲 滋養體,確認核酸染劑確實進到受熱棘阿米巴原蟲滋養體當中。Fittipaldi 等人(2011) 則發現以200 μM PMA 處理 30 min 後,使用 PhAST Blue 光照 15 min 搭配 qPCR,
可區別未受熱與經121℃滅菌 20 min 之棘阿米巴原蟲滋養體與囊體。
檢視兩篇研究之間的差異可以發現,呂氏(2010)雖以 EMA、PMA 兩種核酸染 劑評估棘阿米巴原蟲滋養體之適用性,卻未測試囊體的部分;反觀Fittipaldi 等人 (2011)雖以棘阿米巴原蟲滋養體以及囊體進行測試,但只有評估 PMA,而目前尚 未有人將EMA-qPCR 或 PMA-qPCR 應用於定量環境中之棘阿米巴原蟲。另外呂氏 (2010)與 Fittipaldi 等人(2011)所開發之 EMA-qPCR 或 PMA-qPCR 方法,雖可以區 分未受熱與受熱之棘阿米巴原蟲,但若將其直接應用在土壤樣本上,可能會有基 質干擾的問題存在。Pizs 等人(2007)認為,因 EMA 帶有正電,而土壤有較多的陽 離子交換位置(cation exchange site),此時土壤基質就會干擾 EMA 與我們想要觀察 的菌種結合,造成活菌高估的情況。.
總結上述,目前文獻雖有利用棘阿米巴原蟲滋養體以及囊體進行核酸染劑評 估,但缺少囊體對於EMA 之評估結果,且尚未有研究使用 EMA-qPCR 或
PMA-qPCR 進行環境棘阿米巴原蟲之監測,另外若要將此類技術應用於土壤樣本,
必須審慎處置土壤基質對於EMA 作用的干擾並避免出現 qPCR 之抑制作用。
18
第二章 研究目的
棘阿米巴原蟲廣泛地存在於環境當中,但目前對於其在土壤、空氣以及水體 的定量資訊有限,無法評估人在各環境中之暴露風險。有鑑於此,本研究主要目 的如下:
1. 方法開發:
(1) 選擇定量棘阿米巴原蟲囊體最佳核酸染劑濃度
(2) EMA-qPCR 定量土壤中活性棘阿米巴原蟲之方法開發 (3) 生物氣膠採樣器之比較
(4) 空氣、土壤以及水體當中qPCR 之抑制物處置 2. 環境驗證:
(1) 檢視EMA-qPCR 於空氣、土壤以及水體當中之適用性。
3. 暴露初探:
(1) 探討空氣、土壤以及水體中之活性與總棘阿米巴原蟲濃度。
(2) 探討土壤及空氣環境因子與活性及總棘阿米巴原蟲濃度之相關性。
19
第三章 研究架構
本研究第一部分先進行方法開發(圖一),為補強過去研究不足之處,進行棘阿 米巴原蟲囊體的最佳核酸染劑濃度測試,並進行EMA-qPCR 定量土壤中活性棘阿 米巴原蟲之方法開發,另外也進行了生物氣膠採樣器之比較,希望能夠找出適合 定量空氣中棘阿米巴原蟲之採樣器,最後以暴露初探當中qPCR 抑制物處理結果回 饋本研究方法學當中環境樣本之qPCR 抑制物該如何處置。
第二部分則進行環境驗證與暴露初探(圖二),空氣採樣地點選擇於恆春鎮與車 城鄉之洋蔥田,並進行空氣環境因子測量(溫度、相對濕度、風速);土壤採樣地點 選擇於恆春鎮與車城鄉之洋蔥田、台大園藝農場之蔬果/香草植物田以及台大農藝 農場之水稻田進行採樣,並進行土壤環境因子測量(含水量、pH 值、異營性細菌濃 度、植栽覆蓋、結構);水體採樣地點選擇於迪化汙水處理廠、台大公衛學院冷卻 水塔、台大農藝農場之水稻田以及農田溝渠、台大公衛學院生物性實驗室之洗眼 站,觀察三種不同介質環境當中活性與總棘阿米巴原蟲之濃度,進一步探討土壤 與空氣中棘阿米巴原蟲濃度之顯著影響環境因子。
20
圖一:方法開發之架構圖
a:總和稀釋倍數係考慮採樣樣本稀釋/濃縮倍數以及 DNA 稀釋倍數下所得之值
21
圖二:暴露初探與環境驗證之架構圖
22
第四章 材料與方法
4.1 微生物菌種
4.1.1 Acanthamoeba castellanii (ATCC30234) 4.1.1.1 滋養體
將2 mL A.castellanii 接種於含有 18 mL ATCC medium 712 的 T75 細胞培養瓶 當中(Corning Incorporated, NY, USA),放入 25℃培養箱(LTI603, TKS, Taiwan) 3 日,
其滋養體會呈現不規則狀,並具有空泡和偽足(Khan, 2006)。
4.1.1.2 囊體
將培養3 日的 A. castellanii 滋養體(40 mL),使用 200g 離心 8 min (Grimm et al., 2001),移除上清液後使用 10 mL 囊化培養液清洗 2 次,之後以 10 mL 囊化培養液 回溶沉澱物,將其移入T75 細胞培養瓶(Corning Incorporated),並置於 25℃培養箱 (LTI603) 13 日,其囊體體積明顯小於滋養體,且呈現外層厚實而內層形狀多變化 的雙層壁結構(Khan 2006)。
4.1.2 Escherichia coli (ATCC25922)
將 E. coli 使用畫線法培養於 Nutrient agar (NA)當中,置於 37℃培養箱(LE-509, YIHDER, Taiwan) 1 天,將其保存於 4℃備用。
4.2 試劑與緩衝液
4.2.1 製備 ATCC medium 712
化學藥品 濃度 重量或體積
Proteose peptone (BD 211684) 20.0 g
Yeast extract 1.0 g
23
MgSO4‧7H2O 0.4 M 10.0 mL
CaCl2 0.05 M 8.0 mL
Fe(NH4)2(SO4)2‧6H2O 0.005 M 10.0 mL Na2HPO4‧7H2O 0.25 M 10.0 mL
KH2PO4 0.25 M 10.0 mL
Sodium citrate‧2H2O 0.1 M 34.0 mL
Glucose 2 M 50.0 mL
取20 g Proteose peptone (P0431, Sigma-Aldrich, USA)與 1 g Yeast extract (212750, BD, USA)加入 950 mL 去離子水均勻混合, 6 種鹽類依照濃度配置後,
分別進行121℃滅菌 20 min。滅菌後依上述鹽類所需之體積加入 950 mL 含有 Proteose peptone 與 Yeast extract 的培養液。使用 0.22 μm 的無菌過濾器(Milliporem Bedford, MA)將 50 mL 2 M 的 Glucose 過濾至培養液,均勻混合後分裝至 50 mL 無 菌離心管,將離心管置於室溫一天確認是否汙染後,將培養液置於4℃保存。
4.2.2 製備 Page’s Amoeba Saline
化學藥品 重量或體積
NaCl 1.2 g
MgSO4‧7H2O 0.04 g
CaCl2 0.04 g
Na2HPO4 1.42 g KH2PO4 1.36 g
將上述的化學藥品分別加至100 mL 去離子水,與 950 mL 的去離子水一同進
24
行121℃滅菌 20 min。滅菌後將上述 5 種化學藥品取 10 mL 加入 950 mL 的去離子 水使其總體積為1 L,保存於室溫中備用。
4.2.3 製備囊化培養液(Encyst medium)
化學藥品 濃度 體積或體積
KCl 0.1 M 7.455 g
MgSO4 8 mM 0.963 g CaCl2 0.4 mM 0.044 g NaHCO3 1 mM 0.084 g Tris-HCl 0.02 M 20.0 mL
將上述之化學藥品加至980 mL 去離子水當中均勻混合,並使用 0.2 M NaOH 將pH 值調整至 8.9-9.0,經 121℃滅菌 20 min 後,保存於室溫中備用。此培養液是 引用於Neff 等人(1964)的文獻,但使用 Tris-HCl 取代 amine buffer。
4.2.4 製備 TE buffer
化學藥品 濃度 體積
Tris-HCl, pH 7.5 1 M 1 mL
EDTA 0.5 M 0.2 mL
將上述化學藥品加至98.8 mL 的去離子水當中,使其總體積為 100 mL,進行 121℃滅菌 20 min,保存於室溫中備用。
4.2.5 製備 Nutrient agar (NA)
取23 g NA (213000, BD, USA)加入 1 L 的去離水當中並均勻混合,經 121℃滅
25
菌20 min,分裝至 90 mm 培養皿當中,並保存於 4℃。
4.2.6 製備 R2A agar
取18 g R2A agar (299436, BD, USA)加入 1 L 的去離子水當中並均勻混合,經 121℃滅菌 20 min,分裝至 90 mm 培養皿當中,並保存於 4℃。
4.2.7 製備 Non-Nutrient agar (NNA)
取15 g Agar (214010, BD, USA)加入 1 L PAS 當中並均勻混合,經 121℃滅菌 20 min,分裝至 90 mm 培養皿當中,並保存於 4℃。
4.2.8 製備 Heat-killed E.coli NNA plate
使用接種環將在NA 上的 E.coli 菌落全部懸浮於 2 mL 滅菌 PAS 當中,並經 70℃加熱 120 min 後(Lanocha et al., 2009),取 500 μL 加至 NNA 並使用玻棒均勻塗 抹。
26
4.3 方法開發
4.3.1 棘阿米巴原蟲囊體核酸染劑之選擇
4.3.1.1 置備已知濃度的未受熱與受熱棘阿米巴原蟲囊體懸浮液
將培養13 日的 A. castellanii 囊體 (20 mL),使用 2000g 離心 5 min (Riviere et al.
2006),移除上清液後使用 10 mL PAS 清洗 1 次,之後以 5 mL PAS 回溶沉澱物,
並以血球計數器(Marienfeld-Superior, Lauda-Konigshofen, Germany)搭配 Leica DM 2500 光學顯微鏡(Lecia, Wetzlar, Germany)計算原蟲濃度(400X)以及計算囊體百分 率(92.02±0.12%),並依此濃度使用 PAS 將原蟲濃度調整至 3×104 cells/mL 共 20 mL。
將20 mL (3×104 cells/mL)囊體懸浮液平均分成 10 mL,一組不進行受熱,另一組則 放入95℃水浴槽(B206, Firstek, Taiwan)進行 20 min 加熱。
囊體百分率(%)= 經血球計數器所得之囊體數
經血球計數器所得之棘阿米巴原蟲數×100%
4.3.1.2 使用 Heat-killed E.coli NNA plate 確認未受熱與受熱囊體之活性
取50 μL 未受熱與受熱囊體懸浮液滴入 Heat-killed E.coli NNA plate 的正中間,
約60 min 後貼上封口膜(Parafilm),置入 25℃培養箱(LTI603),使用倒立式顯微鏡 (Eclipse TE2000-U, Nikon, Tokyo, Japan)持續觀察 14 天(200X)並拍照,另外記錄培 養基上Clean zone 之最長直徑(cm)。
4.3.1.3 EMA 與 PMA 核酸染劑之配置
EMA (3-amino-8-azido-5-ethyl-6phenyl bromide) (Sigma-Aldrich, Taipei, Taiwan) 及PAM (3-amino-8-azido-5-[3-(diethylmethylammonio)propyl]-6-phenyl dichloride) (Biotium, Inc., Hayward, CA, USA) 為本研究所使用之核酸染劑。冷凍乾燥型態的 5 mg EMA 以 1 mL 之 ddH2O 回溶,以 5 mg/mL 保存於-20℃。冷凍乾燥型態的 1 mg
27
PMA 以 1 mL 之 20% DMSO 回溶,以 1 mg/mL 保存於-20℃。
4.3.1.4 EMA 與 PMA 核酸染劑處理
將不同濃度的EMA 與 PMA 核酸染劑各取 50 μL 加至含有 0.5 mL (3×104 cells/mL)未受熱與受熱之囊體懸浮液當中,使其管內 EMA 與 PMA 最終濃度為 2.3 與23 μg/mL,震盪 5 sec 之後,使用鋁箔紙包覆樣本進行暗反應 5 min,在暗反應 期間持續用手上下搖晃樣本以確保其混合均勻。反應後再將各樣本放置距離500W Haloline ECO 鹵素燈管(OSRAM, Munchen, Germany) 15 cm 處之碎冰上光照 20 min。
光照時為避免過度光照傷害原蟲,故每隔5 min 更換放置樣本的碎冰,並用手上下 搖晃均勻混合樣本。另外,無核酸染劑處理的0.5 mL (3×104 cells/mL)未受熱與受 熱囊體懸浮液,作為本實驗之對照組。
4.3.1.5 DNA 萃取
受熱與未受熱的囊體懸浮液經核酸染劑及鹵素燈處理後即進行DNA 萃取,本 研究所使用的萃取試劑為FastDNA spin kit for soil (MP biomedical, OH, USA),依照 使用手冊上的說明萃取原蟲DNA,僅修改 Bead-beating 強度為 5.5,時間延長為 1 min,最終沖提體積為 100 μL。
4.3.1.6 Real-time quantitative PCR (qPCR)
本研究使用Roche LightCycler 480 (Roche Diagnostic GmbH, Basel, Switzerland) 上進行棘阿米巴原蟲之qPCR,所選用定量棘阿米巴原蟲之引子與探針的序列皆引 用自Qvarnstrom (2006)。放大片段為 Acantheamoeba 18S rRNA gene,總長為 180-bp。
qPCR 之總體積為 25 μL,包含 10μL 樣本 DNA 或標準品、10μL LightCycler FastStart DNA Master Hybirdization Probe mix (Roche Diagnostics, Mannheim, Gremany)、0.24 μM 的前端引子 AcnatF900 (5’-CCCAGATCGTTTACCGTGAA-3’)、0.24 μM 的後端