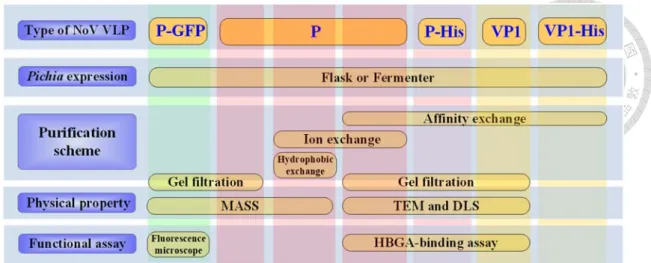
國立臺灣大學生命科學院生化科技學系 博士論文
Department of Biochemical Science and Technology College of Life Science
National Taiwan University
Doctoral Dissertation
利用畢赤氏酵母菌醱酵生產台灣 GII.4 2006b 型諾羅病毒
類病毒顆粒及小型 P 粒子之純化與性質研究
Expression, purification and characterization of
Taiwan-native norovirus GII.4 2006b virus-like particle and small P particle in Pichia pastoris
陳昱伶 Yu-Ling Chen
指導教授:黃慶璨 博士 Advisor: Ching-Tsan Huang, Ph.D.
中華民國 108 年 01 月
January, 2019
謝誌
從一個天馬行空的可能,在經過無數次的討論、校準及堅持後,終於使得 這篇論文得以產生,這一切都要感謝黃慶璨老師的悉心指導與啟發,謝謝老師 這六年多來的包容與照顧,雖然過程充滿各種曲折及變動,但也讓我歷練與學 習到了不管面對生命中的任何變化,都要取絕對值理性面對積極處理,最後,
再乘以零的處事哲學。感謝許瑞祥老師在各種跳脫思維框架與菇類方面所給予 的啟發與建議,雖然菇類相關的實驗結果,還來不及做完整並搬來口試,但整 個學習的過程已讓我受益良多。感謝張世宗老師,在實驗方面所提供的協助,
貫徹著以前學長姐教導的-實驗課就是要修到可以靈活應用,並發表文獻的精 神,身為第一屆修習更新版酵化實驗課的學生,在老師教學的啟發以及老師實 驗室學生在儀器操作所提供的協助下,使得此篇論文中之純化策略得以孕育出 來。研究期間,感謝長庚大學臨床醫學研究所 張沛鈞老師在實驗材料上所給予 的各種協助,並且在百忙之中撥冗逐字批閱斧正文章,以及給予的諸多寶貴建 議。感謝楊啟伸老師在論文上所給予的各種層面的啟發與建議,老師對細節及 邏輯架構要求的精神使我印象深刻,未來我將秉持著這份精神繼續努力。
感謝 Lab106 的各個前人、今人和後人,建構及承傳著 106。還記得第一次 推開106 的大門,是在一個炎熱夏季的午後,適逢大家剛好在圓桌上沏茶休憩 聊實驗,初來乍到的我,這一坐下來,就跟大家天南地北熱絡地聊了起來。感 謝玉儒學姊,在繁忙的同時,撥冗給我實驗上的建議並給予我各種鼓勵。感謝 嗣民、王董、昌廷與映慈學長姊,分享人生與實驗上的經驗,使我增長見聞。
感謝泡泡,在即將離校的最後二個月,傳授我菇類轉形系統。感謝哩哩桑,給 予我的各種協助,也謝謝有妳,與我一起腦力激盪,想實驗、做實驗,並且一 起處理掉實驗室許多各種大小問題。與我一起走過摸索博一的承鴻、宛伶、彥 鈞,以及帶給我歡笑與各種鼓勵的巧青、佳葳、芝榕、俞均,開始轉化並接力 承傳實驗室各種傳統的景翔、孟羲、凱琳、哲銘、映希、浩安、尚儒、冠陞,
以及後續陸續加入 106 的煒倫、麒倫、穎之、翔輔、宥儒、祐桓、少穎、玟 璁、瑞祺等等,有幸能認識你們,與各位一起切磋,一起成長,相信大家在未 來的道路上,都能開創出屬於自己的一片天,並完成自己的理想。
致我的好友婷婷與朱朱,感謝二位時常拎我出門放風、到處溜溜,並且忍 受我出門永遠隨意,喜歡天外飛來一筆,腦袋容易開啟飛航模式。
感謝我的家人,謝謝親愛的妞妞,最後一年,感謝有妳,願意辭職回去幫 忙照顧,家裡幸好有妳幫忙頂著,讓我安心把實驗完成,得以順利畢業。謝謝 給我最大包容,給我最大支持,給我最大鼓勵及鞭策的爸爸與沒能看到我完成 學業在天上的媽媽,你們在我心中,永遠是我最強大的支柱、後盾與避風港。
最後感謝台灣這塊土地養育並栽培了我數十年,感恩在我生命中教導我、指引 我、提攜我、幫助我的人,未來我將帶著這份感謝與感恩,繼續回饋這個社 會。
昱伶 謹誌 2019 年 2 月
摘要
諾羅病毒為全球流行之非細菌性腸胃道致病菌。由於缺乏細胞株生產減毒 或去活的諾羅病毒疫苗,諾羅病毒疫苗主要以類病毒顆粒為基礎發展。諾羅病 毒衣殼蛋白質VP1 之突出區(P (protruding) domain),會構成由 12 個雙體化 P 蛋 白質所組成之P 粒子(P particle),其抗原形態與病毒衣殼蛋白質 VP1 相同。因 突出區具有三個多出來的環狀結構,能嵌入外來抗原,先前已做為外來抗原呈 現平台。本研究欲使用分離自台灣本土株,GII.4 型諾羅病毒之突出區作為 P 蛋 白質多抗原呈現平台之基礎。首先在表現部分,利用不具有內毒素疑慮、培養 成本低廉且易於放大培養之畢赤氏酵母菌 Pichia pastoris,經醱酵槽培養,胞內 P 蛋白質產量可達 220 mg/L。其次,在純化策略方面,利用 P 蛋白質本身的電 性及原態具有的組胺酸,本研究發展出二套無標籤純化策略:(1)利用宿主蛋白 質與P 蛋白質對管柱的吸附落差,P 蛋白質可經由離子交換管柱及疏水性管柱 純化得到,但最終回收後產量僅有2.5%。為進一步提升產量,(2)利用親和層析 管柱(His-Trap)能與原態暴露在外之組胺酸吸附的特性,P 蛋白質可利用親和層 析管柱及離子交換管柱純化,回收率為28.1%,純度可達 82.1%。此外,藉由 VP1 蛋白質原態所具有的組胺酸,及 VP1 能組成顆粒之特性,原態之 VP1 顆粒 亦能藉由親和層析管柱及膠體層析管柱純化,回收率為20%,純度可超過 90%。純化出之 VP1 蛋白質經粒徑分析儀及穿透式電子顯微鏡檢測,VP1 能成 功組成具有生物功能之粒子,能與成人唾液之HBGA (human histo-blood group antigen)結合。與帶有組胺酸標籤(His-tag)之 P 蛋白質(P-His)相似,純化出之原 態P 蛋白質,經粒徑分析儀測定及穿透式電子顯微鏡觀測,二種 P 粒子皆呈現 小型P 粒子(small P particle)之三角形及四角形形態,顯示以酵母菌表現之台灣 流行株GII.4 型諾羅病毒 P 蛋白質,能構成由 6 個雙體化 P 蛋白質組成之小型 P 粒子。前人文獻指出,小型P 粒子僅出現在 C 端突變之 P 蛋白質。而本研究之 P 蛋白質未經末端修飾即呈現小型 P 粒子,經與其它 GII.4 菌株序列比對,發現
其中四個胺基酸對P 粒子的形成可能扮演關鍵角色。當以綠色螢光蛋白質模擬 外來抗原,呈現於P 蛋白質的表面頂端,結果發現嵌合型 P 蛋白質不但能成功 組成嵌合小型P 粒子,亦能發出綠色螢光,顯示台灣分離株諾羅病毒 P 蛋白質 能做為外來抗原呈現平台。本研究為首篇提供諾羅病毒P 蛋白質酵母菌大量生 產、無標籤簡易純化及確立小型P 粒子亦能作為外來抗原呈現平台之研究。
關鍵詞:諾羅病毒、類病毒顆粒、小型P 粒子、嵌合型 P 粒子、畢赤氏酵母 菌、醱酵培養、無標籤純化
Abstract
Norovirus (NoV) is one of the leading causes of acute nonbacterial
gastroenteritis outbreaks worldwide. Due to the lack of a reliable and efficient cell culture system for producing inactivated or attenuated whole NoV vaccines, the development of NoV vaccines relies largely on virus-like particles (VLPs) formed by the major capsid protein VP1 or subviral particles formed by the exterior protrusion (P) domain of VP1. The P particle is composed of 12 P dimers and revealed the same antigenic types as VLP. Additionally, due to the presence of three outermost surface loops in the P domain, the P particle could serve as a platform for carrying foreign antigens. The goal of this study is to provide the fundamental understanding of
different constructs of P particles formed by the P domain of NoV strain GII.4 isolated from Taiwan to establish the multiple antigen presentation platform. The P domain was expressed in Pichia pastoris, a well-known expression system with several advantages including high cell density fermentation at low cost and non-risk of endotoxin. The production of NoV P protein reached 220 mg/L as a soluble form in fermentation cultures. The overexpression P protein provided the cornerstone for the development of tag-free P protein purification schemes. For purification of tag-free P protein, based on the charge and the surface histidine in native NoV, two purification schemes were developed: (1) The host cell proteins and the target P protein were separated by the difference of the binding strength to the chromatography column.
Using anion-exchange and hydrophobic interaction chromatography purification schemes, NoV P protein with high purity was obtained. However, the P protein recovery was 2.5%. (2) Using the HisTrap affinity column and anion-exchange column, the native NoV P protein was purified, and recovery and purity were 28.1%
and 82.1%, respectively. Besides, the NoV major capsid protein VP1 was also
NoV VP1 protein was over 90% and the recovery was 20%. Dynamic light scattering (DLS) and transmission electron microscopy (TEM) analysis of the purified NoV VP1 revealed that VP1 proteins were self-assembled into particles, and these particles remained HBGA binding ability as evidenced by saliva binding assay. Similar to P- His protein, the P protein also formed biologically functional small P particle
composed by six of P dimer. The purified P-His and P protein, analyzed by TEM and DLS, were also revealed triangle-, square- and ring-shaped. Previous studies showed that the small P particles were only found with C-terminus modification. Without terminal modification, small P particles were formed in this study. The amino acid sequence analysis showed only four different amino acids between the P domain in this study and other investigated GII.4 strains, suggesting that these amino acids might play an important role in P particle formation. To extend the application of the small P particle, the green fluorescent protein was used to mimic the foreign antigen and inserted into NoV P loop2 distal end. From gel filtration and fluorescent
microscopy, the chimera small P proteins were self-assembly into chimera small P particles and showed green fluorescence. It was indicated that these Taiwan native small P particle can be an antigen presentation platform. This study was the first report of NoV P protein covering overexpression in P. pastoris, easy handling tag-free purification schemes, and formation of small P particles without terminal modification as foreign antigen displaying platform.
Keyword: Norovirus, Virus-like particle, small P particle, chimera P particle, Pichia pastoris, fermentation cultures, tag-free purification
目錄
摘要……….I Abstract………..………..III 目錄……….…………...……….……..V 圖目錄………..……….………...….VIII 表目錄………..……….………....X
第一章 前言………..………1
1.類病毒顆粒疫苗………...……….….1
1.1 類病毒顆粒疫苗的發展優勢………..………..………….….1
1.2 類病毒顆粒的結構分類………..………..……….….1
2.諾羅病毒介紹……….2
2.1 諾羅病毒之流行病學……….……….2
2.2 諾羅病毒的研究挑戰及替代的研究方案……….……….3
2.3 諾羅病毒分子生物學特性及分類………...……….…………..5
2.4 諾羅病毒之細胞趨性………..………6
2.4.1 HBGA 之生合成路徑………...………6
2.4.2 人類諾羅病毒與不同 HBGAs 之結合特異性………….………7
3.諾羅病毒類病毒顆粒 (VP1) ……….………...…….………….… 10
3.1 諾羅病毒 VP1 結構……….……….……….… 10
3.2 諾羅病毒類病毒顆粒之生產與純化………10
3.3 諾羅病毒疫苗………11
4.諾羅病毒 P………11
4.1 P 粒子 (P particle) ………12
4.2 小型 P 粒子(Small P particle) ………13
4.3 諾羅病毒 P 粒子做為多抗原呈現平台………13
4.3.1 多抗原呈現平台………...…………..13
4.3.2 P 粒子攜帶外來抗原的策略………...…14
4.3.3 P 粒子成為疫苗的利基及攜帶外來抗原的例子………...…14
4.4 P 蛋白質之生產與純化………...………...………...15
5.嗜甲醇酵母菌(Methylotrophic yeast) ………...………...……...18
5.1 甲醇代謝調控路徑(MUT pathway) …………...………...……...18
5.2 Pichia pastoris 表現系統………...………...………...……...19
5.2.1 菌株與甲醇表現型………...………...…………...……20
6.研究動機及具體目標……...………...…………...………..21
6.1 研究動機……...………...……….……...………..21
6.2 研究目標………...……….……...………...………22
1.培養機與藥品…………...……….……... ……..………...……24
2 菌株與培養條件…………...……….……...………...……25
2.1 細菌……...………...……….………...………25
2.2 真菌………...………..……...…………25
2.3 菌種保存………...………..…………...…………26
3.質體建構………...……….………...…………26
4. P. pastoris 轉形………...………..………...…………30
4.1 P. pastoris 勝任細胞製備………...………...…30
4.2 轉形 DNA 置備………...…………...………...………30
4.3 電穿孔轉形………...……….…………30
5. P. pastoris 轉形株篩選與培養………...……….31
5.1 抗藥性濃度梯度篩選……...……….31
5.2 搖瓶培養……...……….31
5.3 醱酵槽培養……...……….31
6.轉形株螢光顯微鏡觀察………...32
7.異源蛋白質產物純化………...32
7.1 蛋白質萃取……….………...32
7.2 親和層析純化………33
7.3 離子交換純化………33
7.4 疏水性層析純化………34
7.5 膠體過濾法………34
8.異源蛋白質分析………...………37
8.1 西方墨點法………...……….………37
8.2 LC-MS/MS…...……….………....……….………38
8.3 Indirect ELISA…...……….………...……….………...38
8.4 唾液結合測試 (Saliva-binding assay) …...………...39
8.5 穿透式電子顯微鏡…...……….……….……..………..….………...40
8.6 粒徑分析……..……….…..…….………..………..…..…40
第三章、結果…..………..………..………43
一、序列分析與結構模擬………..………..……..………43
1.胺基酸序列分析………..……..………...………43
2.結構模擬..………..………...………43
3.不同菌株之序列比對………...………...….43
二、組胺酸標籤(6xHis)之 P 蛋白質生產、純化及顆粒分析……….………50
1. P. pastoris 生產………..…..………50
2.以組胺酸標籤(6xHis)純化………...…..………50
3.物性分析………...…..………50
4.功能性分析………...………...…..……….51
三、原態P 蛋白質之生產、純化及顆粒分析……….…..…...…..…………55
1. P. pastoris 生產………...………..……...…..…………55
1.1.高表現量 P 轉形株篩選………...………..……...…..…………55
1.2 搖瓶培養………...………..………..……...…..…………55
1.3 醱酵槽培養………...………..………..……...………..55
2.開發無標籤純化策略………...………..………..……...………….61
2.1 策略一:利用雜蛋白質與目標蛋白質對管柱之吸附落差純化…………...61
2.2 策略二:利用原態之組氨酸純化………...………..………..……66
3.物性分析………...………..………..……….………..……...…..73
4.功能性分析... …..………..………..……….…………..……...……73
四、嵌有綠色螢光之原態P 蛋白質生產及物性功能性分析…..………79
1. P. pastoris 生產…..………..……...…..………..…...………...79
2.物性與功能性分析…………..……...…….…………..…...………...79
五、組胺酸標籤(6xHis) VP1 蛋白質之生產與純化..………..…...………..……..83
1. P. pastoris 生產...…….……..…...………...…….…………..…...……….83
2.以組胺酸標籤(6xHis)純化.………...…….……….………...…….……….…83
六、原態VP1 蛋白質之生產、純化及顆粒分析…...…….…….………...…….85
1. P. pastoris 生產.………...…….…….………...…….…….………...……....85
2.利用原態之組氨酸純化.………...…….…….………...…….……….85
3.物性分析.………...…….…….………...…….…….………...………...……86
4.功能性分析.………...…….…….………...…….…….………...…….……86
第四章、討論.………...…….…….………...…….…….………...…….….……92
1. P 蛋白質產量之比較…….…….………...………….………...…….….……92
2.無純化標籤之純化策略探討……...…….…...………...…….….……92
2.1 P 蛋白質之純化策略討論...…….…….………...…….….……….92
2.1.1 策略一:利用蛋白質之吸附落差純化…….………...…….….93
2.1.2 策略二:利用原態之組氨酸純化…….………...…….….93
2.2. VP1 蛋白質之純化策略討論..….……….………...…….….94
3.類病毒顆粒之顆粒性與生物功能性探討…….…....…….………...…….….95
3.1. P 蛋白質之顆粒性探討.………...…….…….……….………...…….….95
3.2. VP1 蛋白質之顆粒性探討.…….……….……….……...…….….95
3.3. VP1、P 與 P-6xHis 與 HBGA 之結合差異討論.……..….………...…….97
3.4.組胺酸(6xHis)對 P 蛋白質之影響………..….……...….…………..….….98
4. P 蛋白質之延伸應用性………..….……...….…………..….……….99
第五章、總結………..….…………..….…………..….……….…..…...………….101
第六章、未來工作與展望………..….…………. ………..…..…..……….…102
第七章、參考文獻………..….…………..….………..….…………..….…105
圖目錄
圖1-1、人類諾羅病毒基因組成和結構………...……...………8
圖1-2、FUT1、FUT2 和 FUT3 酵素生合成 HBGA 之路徑....…….……….……...9
圖1-3、P particle 與 Small P particle cryo-EM 結構圖…..………….…..………...16
圖1-4、P 蛋白質外來抗原嵌入模擬圖……….………..….….…………...17
圖1-5、研究架構…..….….………..…….………..………...23
圖2-1、載體建構….….………..…….………..……….29
圖3-1、台灣分離株 GII.4 型諾羅病毒之 VP1 胺基酸序列…………..………..…45
圖3-2、台灣分離株 GII.4 型諾羅病毒之 VP1 蛋白質結構模擬………..………..46
圖3-3、演化樹分析 17 種不同 GII.4 型菌株之 P 胺基酸序列………..………….47
圖3-4、GII.4 型諾羅病毒之 P domaim 序列比較及台灣分離株結構模擬………49
圖3-5、轉形株 P-His (A)以搖瓶誘導表現及(B)以 His-Trap 管柱純化………….52
圖3-6、純化之 P-His 蛋白質物理特性分析………..……….. ………..………….53
圖3-7、HBGA 結合測試………..……….. ………..……….. ………..…………...54
圖3-8、高表現量 P 轉形株塞選………..……….. ………..……….. ………..…...57
圖3-9、轉形株 P (A) 23 及 30°C (B) 30°C 之誘導培養………..………58
圖3-10、以醱酵槽大量生產 P 蛋白質………..……….. ………..…………..……60
圖3-11、分離純化流程圖………..……….. ………..……….. ………..…………..63
圖3-12、P 蛋白質與宿主蛋白質之線性沖提圖譜………..…………...……...…..64
圖3-13、二次階梯式沖提及一次疏水性管柱純化之分析結果………..……..…..65
圖3-14、台灣分離株 GII.4 型諾羅病毒之 P 及 P-6xHis 組胺酸預測結構圖……68
圖3-15、轉形株 P 之搖瓶與醱酵槽之醱酵液以 His-Trap 管柱純化……...……..69
圖3-16、轉形株 P 之搖瓶及醱酵槽醱酵液以陰離子交換管柱二次純化….……70
圖3-17、純化之 P 蛋白質物理特性分析……….………75
圖3-18、HBGA 結合測試……….………77
圖3-19、粗萃之 P 蛋白質顆粒性分析……….………78
圖3-20、螢光顯微鏡觀察嵌綠色螢光諾羅類病毒顆粒(P-GFP)酵母菌轉形株…80 圖3-21、P-GFP 嵌合蛋白質之顆粒性分析…………..……...……….82
圖3-22、轉形株 VP1-His 醱酵液以 His-Trap 管柱純化…….………84
圖3-23、台灣分離株 GII.4 型諾羅病毒之 VP1 及 VP1-His 組胺酸預測結構圖..87
圖3-24、轉形株 VP1 醱酵液以 His-Trap 及膠體層析管柱純化及分析…….…...89
圖3-25、純化之 VP1 蛋白質物理特性分析……….………...90
圖3-26、HBGA 結合測試……….………91
附圖1、人類諾羅病毒之傳染途徑………….………..……….………….118
附圖2、諾羅病毒基因組和基因型………….………..……….………….119
附圖3、諾羅病毒組成與生活史………….………..…….……….120
附圖4、生合成 HBGA 之路徑………….………..…….………122
附圖5、ABH、Lewis 及相關抗原生合成路徑圖………….……….………123 附圖6、諾羅病毒衣殼蛋白質 VP1 組裝成病毒顆粒模擬圖………….……..….125 附圖7、VA387(GII.4)與 HBGA 之結合界面的晶體結構…………..….…….126 附圖8、比較 GII.4 與 GI.1 和 GII.3 之 P 蛋白質結構...127 附圖9、嗜甲醇酵母菌甲醇代謝路徑………….…….………..……….…………128 附圖10、以西方墨點法偵測原態或帶有組胺酸之 VP1 或 P 蛋白質…………..129
表目錄
表2-1、質體建構所使用之引子………28
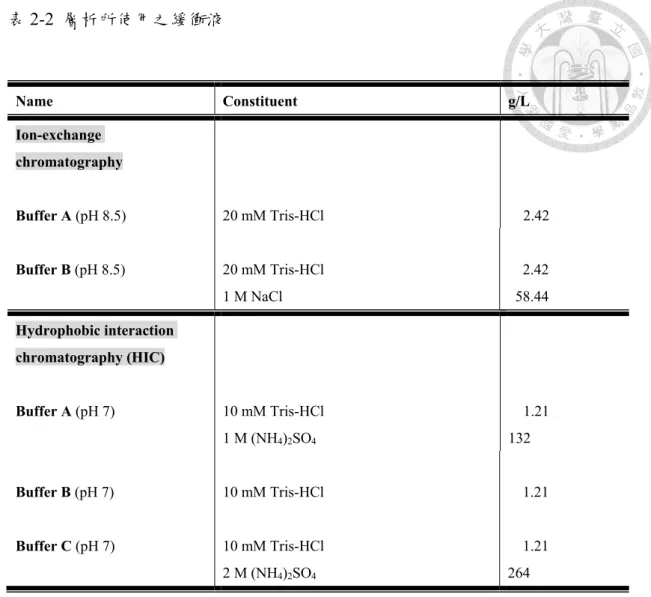
表2-2、層析所使用之緩衝液………..……….…………36
表2-3、西方墨點法分析所用試劑之組合成分……….………41
表2-4、酵素連結免疫反應所用試劑之組合成分………42
表3-1、諾羅病毒 P 蛋白質之回收與純度表………..………72
附表1、諾羅病毒類病毒顆粒之穩定性研究………..………130
附表2、諾羅病毒疫苗的發展現況……….………131
附表3、不同碳源對嗜甲醇酵母菌 AOX 基因的抑制和去抑制作用………..…132
第一章 前言 1.類病毒顆粒疫苗
類病毒顆粒(VLP; virus-like particle)於 1968 年,首次於感染 B 型肝炎的病 人血漿中被純化出來[1, 2]。10 多年後,第一支利用基因重組技術,以大腸桿菌 所表現的口蹄疫病毒(Foot and mouth disease virus; FMDV)類病毒顆粒疫苗被研 發出來,並用於接種牛和猪上[2, 3]。經過 30 多年的發展,以類病毒顆粒(Virus- like particles; VLPs)為基礎發展的疫苗,已廣為人接受並大量的應用於許多疾病 的預防及治療。類病毒顆粒不僅可做為基因治療工具及新的奈米材料,以高通 量篩選技術,結合蛋白質工程以及化學耦合(chemical coupling),還可擴展疫苗 學、基因傳遞、免疫治療和生物奈米技術的應用性[4]。
1.1 類病毒顆粒疫苗的發展優勢
傳統疫苗是將分離的病原體經由減毒或去活後呈現給人體的免疫系統以產 生抗體,進而達到免疫的效果,但此種方式僅限於可培養且變異度低的病原 體。此外,使用活體病原菌,如果去活或減毒不完全將造成安全疑慮[5]。
類病毒顆粒係由多個次單元(subunit)蛋白質組成的複合物,可自我組裝成 與病毒相似的3D 構造。類病毒顆粒具有重複性的表面結構,能誘發較佳的免 疫反應,並可經由基因重組的方式將病毒的外鞘蛋白基因送入宿主大量表現,
不似傳統疫苗受限於需先分離培養病原菌。類病毒顆粒缺乏病毒的遺傳物質,
無法自我複製而不帶有病原性,是個高安全及高免疫原性的疫苗平台。經由不 同的策略,幾乎任何類型的分子皆可經由類病毒顆粒呈現給免疫系統[5, 6]。
1.2 類病毒顆粒的結構分類
根據病毒結構,類病毒顆粒可分為二大類:不具外套膜類病毒顆粒( non- enveloped VLPs )及帶有外套膜類病毒顆粒( enveloped VLPs) [4]。
不具外套膜類病毒顆粒通常由一種或多種會自我組裝形成類病毒顆粒的外 鞘蛋白質所組成,從宿主表現後,不帶有宿主的任何成份。此外,類病毒顆粒 可以進一步發展成一個抗原呈現平台,使外來嵌入的抗原能借由病毒的組裝特 性,以高重複性、系統性的方式呈現給免疫系統。類病毒顆粒抗原呈現平台,
可做為疫苗對抗任何病原菌,包含慢性非傳染性疾病如癌症和過敏[6]。
帶有外套膜類病毒顆粒的結構相對較複雜,由宿主細胞膜與目標抗原一同組 成在外鞘上。帶有外套膜的類病毒顆粒為一個高度靈活的平台,能整合呈現更多 來自相同或相異病原菌的抗原[6, 7]。
2.諾羅病毒介紹
病毒、細菌、真菌、寄生蟲、毒素或普利昂蛋白(prions)皆可經由食物和水 傳播,並經由糞口途徑引發食源性疾病,雖然食源性病毒僅能在宿主細胞複 製,無法在食物、水以及貯藏期間增加,但是它比細菌更能抵抗高溫、極端pH 及消毒劑,並且只要少量的病毒顆粒就能致病,因此一般食品加工處理程序無 法有效去除病毒病原體[8]。
2.1 諾羅病毒之流行病學
1968 年諾羅病毒首次在美國俄亥俄州諾沃克(Norwalk, Ohio)爆發[9]。疫情通 常爆發在人群大量聚集的地方,如醫院 學校、游泳池、軍事基地等。諾羅病毒 的主要感染途徑為糞口傳染,一般受汙染的食物和水為首要傳染源,接著再經由 人與人的接觸進一步傳播,最後爆發 (如附圖 1) [10]。全球每年有 1.25 億的人口 感染諾羅病毒[11]。感染諾羅病毒的症狀包含腹瀉、嘔吐、噁心、腹部絞痛、畏 寒、頭痛、脫水和高燒。病毒具有1~3 天的潛伏期,症狀會持續 2-3 天,並根據 個人的免疫能力而有所增減[12]。雖然嚴重到需要入院治療的以幼兒為大宗,但 花費最多 死亡率最高的確是以長者為主。由於諾羅病毒有許多不同的菌株,人
體很難長期對此種病毒免疫,因此在人的一生中可能會反覆感染這項疾病[13]。
諾羅病毒能造成大流行的原因包含,第一、低劑量即可致病(<20 病毒顆粒),
第二、在發病期間,高濃度的病毒顆粒會隨著糞便一起排出(每克的病人糞便有 106到109之病毒顆粒),並在發病結束後4-8週內糞便還是可能帶有具感染的病毒 顆粒,第三,發病之病人呈霧化之嘔吐物有助於病毒的傳播,第四、病毒一但存 在環境中,數週內仍能保有傳染力,由於病毒顆粒極其穩定,以一般的清潔劑或 消毒殺菌劑,以常用濃度或一般的處理時間難以完全去除活性[14]。
農產品與海鮮為主要帶有諾羅病毒的高風險食物。在美國自1998-2005年有 40%的諾羅病毒大流行來自農產品,帶有諾羅病毒的主要汙染途徑,來自於使用 被污染的水灌溉、施肥或清洗。因蔬菜和水果中存有非常多的葡萄糖和多聚醣,
其中醣類的基團類似諾羅病毒的受體HBGAs (human histo-blood group antigens),
目前認為諾羅病毒也可能經由附著在類似HBGAs的醣類上,進而污染食物[8]。
另一個會帶有諾羅病毒的高風險食物為海鮮,特別是像貽貝,牡蠣和蛤等雙殼貝 類。由於它們是濾食性攝食動物,在存有病毒的水中,更容易遭到汙染,而且根 據生長的地點、季節以及檢測方式的靈敏度,會決定諾羅病毒的汙染程度。例如 從研究指出,諾羅病毒的病毒顆粒會結合在牡蠣中腸內的消化盲囊上,結合在相 似於人類HBGAs分子的地方[15];不同的諾羅病毒基因型,對不同的牡蠣組織會 有不同的親和力,因此不同的諾羅病毒基因型會有不同的生物累積量。在許多大 爆發的事例中,皆是與食用生的或未煮熟的的貝類有關。
2.2 諾羅病毒的研究挑戰及替代的研究方案
諾羅病毒之病毒顆粒於 1972 年第一次被觀察到,為 GI.1 大小 27 nm 之顆 粒[9]。往後幾年許多實驗室也致力於發展體外培養平台。而在人類諾羅病毒的 研究上,主要遇到二大挑戰,第一,人類諾羅病毒無法從細胞培養中取得,第 二,人類諾羅病毒沒有小型的動物培養模式[8]。然而隨著時間的推移,人類諾
羅病毒體外培養有許多重大的突破。於2007 年,研究人員能以三維之人類腸組 織培養諾羅病毒,並且發現人類諾羅病毒傾向感染腺上皮細胞(glandular
epithelial cells)並複製[16, 17],然而這套系統卻無法套用在其他實驗室培養諾羅 病毒[18, 19],因此之後少有研究使用此套系統。
於 2014 年 Jones 等人於諾羅病毒的體外培養有了突破,而契機在於發現,
染了人類諾如病毒的黑猩猩其B 細胞可以產生病毒衣殼蛋白質[20],以及某些 腸道細菌帶有組織血型抗原(histo-blood group antigens;HBGAs) [21],根據這二 點,Jones 團隊成功使人類諾羅病毒基因拷貝數增加了 600 倍,於帶有腸道細菌 的B 細胞(BJAB)中[22]。具體來說,這套培養系統諾羅病毒複製需要游離的 H 型HBGA 或帶有 H HBGA-like sugars 之腸道細菌(Enterobacter cloacae),做為病 毒複製的輔助因子。然而此套培養系統卻有,第一、與腸道細菌(E. cloacae)共 培養如操作不慎容易引發汙染,然而以帶有人工合成的HBGAs 溶液做替代並 共培養,則有成本高昂以及因合成和純化上的差異而有品質不一的問題,第 二、人類諾羅病毒感染B 細胞,不會產生明顯的細胞病變,因此定量需仰賴 RT-qPCR [14]。
近期,Ettayebi 等人於 2016 年成功地利用幹細胞所衍生的人類腸道細胞培 養GII.4 和 GII.3 人類諾羅病毒株[23]。此套系統之優勢包含,第一、能提供多 種人類諾如病毒株複製,第二、感染病毒的細胞能出現細胞病變作為判讀,第 三、與流行病學所觀察到之帶有不同HBGA 宿主其感染模式一致。但缺點為培 養時間長及維護成本高,因此無法做為常規食品及水質之病毒檢驗[14]。
雖然野生型小鼠無法使人類諾羅病毒複製,免疫缺陷鼠能使部分人類諾羅病 毒複製,並且人類諾羅病毒可以在大型動物如無菌猪、無菌牛和黑猩猩體內複製 [14],然而無論是免疫缺陷鼠或大型動物,皆有高培養成本、需專業動物養護設 備以及具有動物倫理問題,近期美國國家衛生研究院(NIH)宣布停止支助使用 黑猩猩的研究資金[24],就是其中的一例。
由於先前人類諾羅病毒無法以細胞培養及動物培養模式取得[14],因此,大 量的研究是以諾羅病毒的病毒顆粒(VP1)及老鼠的諾羅病毒,或與諾羅病毒同屬 杯 狀 病 毒 科(Caliciviridae) 的 貓 杯 狀 病 毒 (Feline Calicivirus ; FCV) 、 Porcine sapovirus 和 Tulane virus 做取代,間接研究其特性[8]。
2.3 諾羅病毒分子生物學特性及分類
諾羅病毒(Noroviruses)屬杯狀病毒科(Caliciviridae)、諾羅病毒屬(Norovirus),
為單鏈、正股 RNA、無包膜病毒,病毒顆粒為高度穩定的二十面對稱體,基因 體長度約7.5-7.7 kb,由三個開放式閱讀框架 (open reading frames; ORF)所組成。
ORF1 會轉譯出非結構性病毒蛋白酶;ORF2 會轉譯出諾羅病毒的主要結構蛋白 質VP1;ORF3 會轉譯出次要結構蛋白 VP2 (圖 1-1) [25]。VP1 蛋白質在病毒中 扮演重要角色,它會與宿主細胞表面的HBGA (histo-blood group antigen) 結合,
使病毒進入細胞[26] (如附圖 3),因此不同的諾羅病毒具有不同抗原特異性。VP2 則會加強VP1 內部的穩定,幫助外鞘蛋白質(VP1)的組裝及協助病毒基因的封裝 [27, 28]。諾羅病毒為由 90 個 VP1 二聚體(dimer)所組成,體外表現會自我組裝的 VP1 類病毒顆粒,其大小、形狀及免疫原性皆與諾羅病毒相似,目前已做為疫苗,
並進入人體臨床測試 [29]。
諾 羅 病 毒 根 據 基 因 組 序 列 , 目 前 分 成 七 種 基 因 組 (genogroups) , 其 中 genogroups I、II 和 IV (GI、GII 和 GIV)會感染人類,但 GII 也會感染猪隻,GIV 則會感染貓科及犬科動物,而GVI 和 GVII 感染犬科動物,GIII 和 GV 分別感染 牛及囓齒動物[30]。根據病毒衣殼 (VP1)及 RNA-dependent RNA polymerase (RdRp)之蛋白質序列[31],七種基因組(genogroups)可個別再細分成不同基因型 (genotypes),其中 9 個 GI (GI.1-GI.9)、19 個 GII (GII.1-GII.10, GII.12-GII.17, GII.20- GII.22)及 1 個 GIV (GIV.1)基因型可感染人類[32]。全球最廣為流行及研究的人 類諾羅病毒為GII.4 型[8]。例如在 2006-2012 文獻指出台灣腹瀉事件中,GII.4 同
樣為主要流行株 [33, 34]。根據諾羅病毒爆發與流行之時間,GII.4 型之諾羅病毒 藉 由 VP1 之 高 變 異 區 P domain ( 突 出 區 ) , 可 再 分 類 成 不 同 的 基 因 亞 型 (subgenotype),例如 2006a、2006b、 2009 及 2012 等[30, 34, 35],如附圖 2。
2.4 諾羅病毒之細胞趨性
了解病毒與宿主間的交互作用及病毒受器,才能進而了解病毒特定的宿主範 圍及病原性。
2.4.1 HBGA 之生合成路徑
HBGAs (human histo-blood group antigens)為諾羅病毒進入到人體的受器 (receptor)或輔助受器(co-receptor) (如附圖 3)。HBGAs 為聚醣複合物,會連結在 醣蛋白(glycoproteins)或醣脂質(glycolipids)呈現在紅血球細胞或黏膜上皮細胞 上,或者以游離的方式在血液、唾液、腸道內或乳汁等生物體中流動。HBGAs 抗原的合成,主要受控於ABO、 Lewis 和 secretor 基因家族[36]。如圖 1-2 所 示,抗原前驅物(precursor)之結構有三個,type 1 (Gal β 1-3 GlcNAc β)、type 2 (Gal β 1-4 GlcNAc β )、type 3 (Gal β 1-3 GalNAc α ) [37, 38];人類 ABO 血型之 ABH 抗原,不同於分泌型(secretors),受 FUT1 (fucosyltransferase 1)控制催化 fucose 轉移至 type 2 前驅物,產生 H 抗原供後續 A 或 B 酵素反應產生 A、B 抗 原[13, 36, 37],孟買血型(Bombay blood group)及亞孟買血型(Para-bombay blood group)即是 FUT1 缺失或突變無法產生 H 抗原之例子[39]。分泌型(secretors)則 不同於血型,受控於FUT2 (fucosyltransferase 2)催化 fucose 轉移,當缺乏 FUT2 則為非分泌型(non-secretors),無法合成 ABH 抗原並分泌至唾液和粘膜等分泌 物中[13, 36],而亞孟買血型(Para-bombay blood group)即為雖 FUT1 缺失但 FUT2 還保有功能之例子[39, 40];特別的是,FUT2 也能催化 type 2 前驅物在粘 膜(mucosa)[36]。FUT3 (fucosyltransferase 3)又稱為 Lewis 酵素同樣會催化 fucose
轉移,如果為非分泌者,則表型有Lewis a,如果為分泌者,則表型有 Lewis b;當 type 2 前驅物被 FUT1 或 FUT2 催化者,則表型有 Lewis y,如果沒有催 化者,則表型有Lewis x [13, 36],如附圖 4。
2.4.2 人類諾羅病毒與不同 HBGAs 之結合特異性
首次從臨床感染發現諾羅病毒會辨識HBGAs,是經由人體自願受試的研究 結果得知,研究結果發現,O 型及 A 型人有極高的機率感染 prototype Norwalk virus (GI-1),而唾液檢測幾乎不與諾羅病毒結合的 B 型人,在人體測試中發現 也幾乎不受諾羅病毒感染,非分泌型(nonsecretors)者則有自然抗性,完全不受 感染,相似的結果也發生在諾羅病毒(GI-1)大流行期間 [41, 42]。而以人體測試 諾羅GII.4 Farmington Hills 2002 病毒,從受試結果報告也可觀察到,70%具有 FUT2 功能(即分泌型)之健康成年人會受到諾羅病毒之感染,其中有 57%出現了 感染症狀,而沒有FUT2 功能(即非分泌型)之健康成年受試者,僅有一人(6%)感 染,並且症狀輕微[43],相似的結果也可以在唾液檢測(saliva-binding assay)中觀 察到,GII.4 會結合 A、B、O 等分泌型,但卻無法與非分泌型結合[35]。此 外,根據合成時不同來源之前驅物(type1-3) (附圖 4),諾羅病毒有不同的結合強 度,研究顯示,前驅物以type 1 (Gal β 1-3 GlcNAc β)來源之人工合成 O 型或 B 型之聚醣,與諾羅病毒的結合比type 2 (Gal β 1-4 GlcNAc β )來源之 O 型或 B 型 更緊密 [35, 37]。
每2-3 年 GII.4 會突變形成新的變異株(附圖 2),導致全球大流行,原因來 自於高度變異區protruding (P) domains 基因序列上的變化,導致宿主受體與病 毒親和力的改變、強化病毒逃避人體的免疫力[44]。故通過了解諾羅病毒與不 同的HBGA 受體的結合方式,可得知諾羅病毒流行株的進化方式及病毒特定的 宿主範圍。
圖1-1 人類諾羅病毒基因組成和結構 [45]
(a) 諾羅病毒為單鏈正股無包膜 RNA 病毒,病毒具有三個開放式閱讀框架(open reading frames; ORF),ORF1 會轉譯出非結構性病毒蛋白質;ORF2 和 ORF3 分 別會轉譯出諾羅病毒的主要結構蛋白質VP1 和次要結構蛋白 VP2。VP1 蛋白質 由短鉸鏈(hinge)所連接之殼區(S (shell) domain)和突出區(P (protruding) domain) 所構成。(b)一個二十面體 T = 3 諾羅病毒 VLP 由 90 個 VP1 二聚體所組成。
VP1 之 S 結構在最內部會以支架形式包裹病毒的 RNA,P 結構則在最外部,並 且可再被細分為P1 和 P2 區,並藉由鉸鏈(hinge)與 S 結構相連。左圖為二聚體 之結構模擬圖,右圖為三個相同之S 結構 A / B / C 所構成的病毒顆粒(顆粒建構 過程詳見附圖6)。PDB accession number 1IHM。
圖1-2 FUT1、FUT2 和 FUT3 酵素生合成 HBGA 之路徑 [36]
右上之Type 1、2 或 3 precursor 可經由 fucosyltransferase 1 (FUT1)、FUT2 或 FUT3 以及 A 或 B enzymes 多種酵素修飾產生多種不同抗原。Type 1 及 3 precursor 經由 FUT2 修飾經由 1, 2 linkage 加上 fucose,產生 1 或 3 H type 抗 原,產生的H antigen 可以經由 A 或 B enzymes 修飾,再產生 A 或 B antigens。
Type 2 precursor 雖可透過 FUT1 或 FUT2 經由 1, 2 linkage 產生 H type 2,但在 紅血球催化酵素是FUT1;而粘膜則是 FUT2。此外,type 1 或 H type 1 經由 FUT3 (Lewis enzyme)修飾可個別產生 LeA 或 LeB (如附圖 4);而 type 2 或 H type 2 經由 FUT3 (Lewis enzyme)修飾可個別產生 LeX 或 Le Y (如附圖 4)。Gal:
galactose; GlcNAc: N-acetylglucosamine; Fuc: fucose; GalNAc: N- acetylgalactosamine。
3.諾羅病毒類病毒顆粒 (VP1) 3.1 諾羅病毒 VP1 結構
諾羅病毒顆粒由 90 個 VP1 二聚體所組成。VP1 係以短鉸鏈(hinge)連接之 殼區(S (shell) domain)和突出區(P (protruding) domain)所構成(圖 1-1)。S domain 會主導顆粒之構成,當諾羅病毒顆粒組裝,雙體化之VP1 蛋白質會呈現二種構 形並共同存在,呈彎曲之二聚體會結合構成五聚體,接著五聚體周邊會與呈現 平板的VP1 二聚體再結合,並以這種建構方式逐步構成由 90 個 VP1 二聚體所 構成的病毒顆粒[46] (附圖 6)。而位於病毒顆粒頂部之 P domain 則能與宿主之受 器或輔助受器接合,進而進入宿主體內感染宿主。P domain 可再被細分為 P1 和 P2 區 (圖 1-1),其中位於顆粒最頂部之 P2 區為高度變異區,具有二個與
HBGA 接合之主要位點(附圖 7)。相比起 GI 及 GIII 類病毒顆粒,GII 類病毒顆 粒在鹼性環境下,顆粒穩定性較高(附表 1)。
3.2 諾羅病毒類病毒顆粒之生產與純化
諾羅病毒類病毒顆粒可以利用多種不同的表現宿主生產,包含:被重組桿 狀病毒感染的昆蟲細胞、帶有委內瑞拉馬腦 (Venezuelan equine encephalitis;
VEE)複製子或水泡性口炎病毒(Vesicular stomatitis virus;VSV)載體或質體的哺 乳動物細胞、酵母菌(Pichia pastoris)、以及許多番茄、土豆、馬鈴薯和煙草等 轉基因植物[47]。產出的類病毒顆粒經純化後以口腔、鼻腔或注射或未純化直 接口服,不管是人體或動物,皆能產生諾羅病毒全身性或黏膜專一性抗體[47- 49]。做為疫苗,桿狀病毒表達系統,是最直接並且高效的生產平台,適合大規 模生產及下游純化高純度的類病毒顆粒做為疫苗[47]。文獻指出,以 Sf9 昆蟲細 胞生產GII.4 外鞘蛋白質,醱酵液有超過 100 mg/L 的 VP1 蛋白質產量[50],經 過PEG (polyethylene glycol)沉澱,再以陰離子交換管柱純化醱酵液,可純化出 約10 mg /L 的 VP1 蛋白質;而醱酵液以 CsCl 以梯度超高速離心純化,則可純
化出約35 mg /L 的 VP1 蛋白質[51]。另外,使用 P. pastoris Bg11 酵母菌生產 VP1 蛋白質,經以陰離子交換管柱純化,則可純化出約 0.6 g /L 的 VP1 蛋白質 [52],顯見以酵母菌生產亦有發展的潛力。
3.3 諾羅病毒疫苗
將表現有諾羅病毒類病毒顆粒的轉基因酵母菌、馬鈴薯塊莖或凍乾的轉基 因番茄,未純化直接餵食小鼠,能誘發專一的IgG 及 IgA 諾羅病毒抗體[53- 55]。而人體臨床研究亦顯示,諾羅病毒類病毒顆粒對人體具安全性及免疫原性 [48],在口服疫苗測試中,24 位自願受試者直接食入二次(第 0 與第 7 天)到三次 (第 0 與第 7 和 21 天)150 g 含有 215-751 mg 類病毒顆粒之去皮切塊的轉基因馬 鈴薯塊莖,95%受試者誘發出相對微弱但可測得的專一 IgA 諾羅病毒抗體,
20%產生專一諾羅病毒血清 IgG 抗體,30%受試者糞便產生專一 IgA 諾羅病毒 抗體[56]。
由美國製藥 LigoCyte 公司,臨床 I/II 期諾羅病毒疫苗人體試驗,由 18 到 50 歲的 90 位成年人中,以鼻腔施與桿狀病毒表現系統所生產的諾羅病毒類病 毒顆粒疫苗(具殼聚醣及 monophosphoryl lipid A 做佐劑),結果顯示將近半數的 受試者成功降低了諾羅病毒發病病症,有26%的受試者成功阻止了諾羅病毒的 感染,這項研究證明了諾羅病毒疫苗之開發與上市具高度可行性[57]。目前許 多諾羅病毒疫苗(附表 2),主要以諾羅病毒類病毒顆粒為基礎,搭配不同的給藥 途徑(例如:注射、口服、鼻腔等)及不同的佐劑發展諾羅病毒疫苗[30],目前另 二種人體臨床試驗的疫苗包括,由Vaxart 公司所開發,利用口服由複製缺陷之 腺病毒所表現GI.1 VLP 蛋白質,目前已完成臨床一期試驗,試驗結果確立沒有 安全問題並且疫苗具有免疫原性[58];另一個由 Takeda Vaccines (武田藥品)公司 所發展,利用肌肉注射雙價(GI.1 和 GII.4) VLP 疫苗,目前已完成在 18-64 歲之 健康成年人和65-85 歲老年人之安全性和免疫原性評估[59, 60],並且在人體感 染測試中,雙價疫苗能提供保護[61]。目前疫苗之安全性和免疫原性正在兒童 (NCT02153112),以及海軍中評估(NCT02153112)。
4.1 P 粒子 (P particle)
諾羅病毒為食源性致病菌,其病毒衣殼蛋白質具高穩定性和高免疫原性,
適合發展疫苗[49]。諾羅病毒的主要結構蛋白 VP1 可分為二種區域:殼區(shell (S) domains)和突出區(protruding (P) domains),並以一個短鉸鏈(hinge)所連接 [62](圖 1-1)。先前文獻顯示,在感染諾羅病毒患者的糞便中,可觀察到約 30 kDa 之大量可溶蛋白質[63],後經證實為病毒外鞘蛋白質 P domain [64]。此項結 果顯示在人體腸腔中,為了對抗蛋白酶的降解,P 蛋白質必需具有好的折疊與 構型。存在於病患腸腔中的P 蛋白質具有潛在的生物功能,其中一個可能的功 能即為P 蛋白質可作為誘餌,使後代的病毒體(virions)逃離宿主的免疫系統[64, 65]。P domain 表現的 P 粒子其抗原形態與病毒衣殼蛋白相同,研究顯示以 P 粒 子免疫小鼠,P 蛋白能被樹突細胞呈現並有效的引發細胞免疫,P 粒子所引起 的高體液免疫和細胞免疫與完整的類病毒顆粒一樣[66]。以 P 粒子免疫刺激小 鼠所誘發的中和抗體(neutralizing antibody),也能有效抑制諾羅病毒衣殼蛋白與 HBGA (histo-blood group antigen)的結合[67, 68],因此,P 粒子是個絕佳對抗諾 羅病毒的蛋白質疫苗,也是有潛力做為抗原呈現平台,開發雙重疫苗。
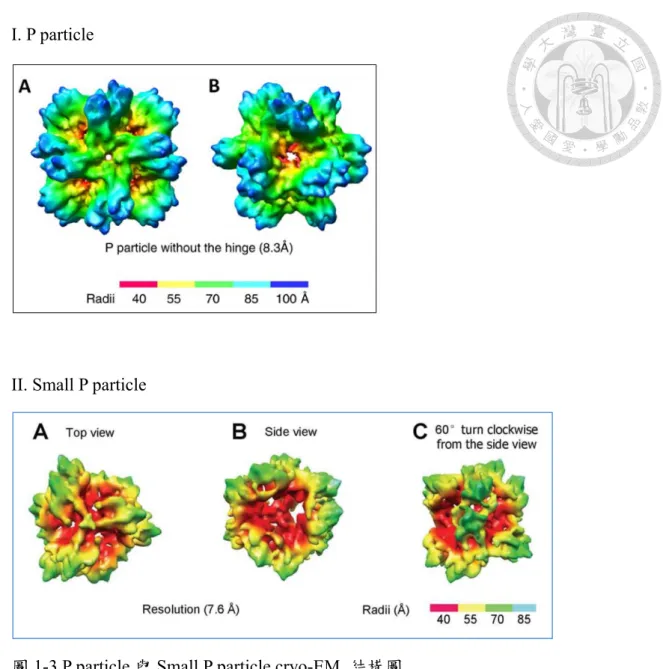
全長的 P 蛋白質會形成 P 二聚體(P-dimer),此二聚體為 P 粒子建構之最小 單位,可形成不同大小的P 粒子。去除掉短鉸鏈(hinge)及末端加上 CCT
(cysteine-containing tag)標籤之 P 蛋白質,主要會構成由 12 個 P 二聚體(P-dimer) 所組成之P 顆粒,顆粒大小約在 20 nm (圖 1-3)。P 粒子的形成來自分子間的相 互作用力,當P 粒子帶有連接殼區和突出區的短鉸鏈(hinge),傾向形成二聚 體,並且P 粒子穩定度較低,去除掉 P 蛋白質 C 端 arginine 片段(RRR-
cluster),同樣也無法形成 P 粒子[49]。P 蛋白質的 N 端或 C 端加上 CCT,能有 效加強P 粒子的形成並強化穩定性,常用之 CCT 有二種,序列為 CNGRC (為 aminopeptidase N (CD13) [69]的配體) [70] 和 CDCRGDCFC (整合素(intergrin)配 體αvβ3) [71]。當末端帶有 6xHis 標籤,但是未加有 CCT 標籤之 P 蛋白質,則會
導致P 粒子的不穩定[49]。P 顆粒在還原條件下會做動態變化,並有不同的大 小,當P 粒子以 E. coli 做表現,以 P 二聚體(~6 nm)為組成單位,會形成 12 聚 體(~14 nm)、18 聚體、24 聚體(~20 nm)及 36 聚體等動態顆粒變化[72]。
4.2 小型 P 粒子(Small P particle)
Small P particle 來自經突變之 P particle。P 粒子的形成與穩定性受二大要素 影響,第一為來自P 蛋白質之間的相互作用力,第二為加在 P 蛋白質尾端的 CCT (cysteine-containing tag) 標籤[73]。目前,雖然 P domain 各個胺基酸如何影響 P 顆粒之形成尚未清楚,但先前研究指出P 蛋白質尾端 RRR-cluster 對 P 顆粒形成 具關鍵影響力。雖然P 去除掉短鉸鏈(hinge)及末端加上 CCT 標籤之 P 蛋白質,
主要會構成由 12 個 P 二聚體(P-dimer)所組成之 P 顆粒(圖 1-3),當在同樣條件 下,將P 蛋白質序列之 C 端再加 flag tag (DYKDDDDK)或 P 蛋白質 C 端 arginine 片段(RRR-cluster)突變成 RPRPRP,二種改變皆會使本由 12 個 P 二聚體所構成 分子量在840 kDa 之 P particle (Φ=~20 nm),轉變成由 6 個 P 二聚體所構成分子 量在420 kDa 之 Small P particle (Φ=~14 nm) (圖 1-3)。然而當去除掉 P 蛋白質 C 端arginine 片段(RRR-cluster)或 RRR 去掉直接替換成 HHH,則是二種顆粒皆無 法形成,僅觀察到~200 kDa 之複合體 (trimer of the P dimer),並且複合體會失去 與HBGA 結合的能力[73]。
4.3 諾羅病毒 P 粒子做為多抗原呈現平台 4.3.1 多抗原呈現平台
抗原呈現平台,必須以高重覆、系統性的方式呈外來抗原給免疫系統。可自 我組裝,具有高重複表現結構,能作為佐劑誘發絕佳免疫反應的類病毒顆粒,可 藉由基因融合方式,嵌合外來抗原或化學耦合(chemical conjugation)黏合外來抗 原。但是化學耦合在製造上須先分別生成類病毒顆粒與目標抗原,再以共價或非
共價方式黏合目標抗原至類病毒顆粒,相比起基因融合,因不能百分之百交聯外 來抗原以及高重複性的生產高再現聚合物,在製造上面臨許多挑戰,須以更複雜 的控管方式以確保品質。以基因融合方式嵌入外來抗原於類病毒顆粒上為目前最 常用的方式,以基因融合方式嵌合二種蛋白質,能確保在單位面積下,高密度的 以相同結構呈現外來抗原,並且在製造上能利用好的模式,快速純化出高純度的 嵌合類病毒顆粒[74]。
4.3.2 P 粒子攜帶外來抗原的策略
與 GI.1 和 GII.3 之 P 蛋白質結構相比,GII.4 之 P 蛋白質頂端,具有三個多 出來的環狀結構(附圖 8),經證實皆適合外來抗原的嵌入[75],並加強抗原的免 疫反應(圖 1-4)。根據目前的研究指出,嵌入的胜肽片段抗原,在同一個 P 蛋白 質的單一環狀結構[49, 67, 76]或同時嵌入三個環狀結構[77-79]皆可行,並且不 影響P 粒子的形成。藉由 P 顆粒的形成,嵌入的外來抗原,其抗原性和免疫原 性可被大幅提升 (one copy per VLP subunit)。雖然目前缺乏抗原呈現在 P 粒子 表面的成功通則,但嵌入的外來抗原,大小並非主要的影響關鍵,嵌入親水性 的抗原比起疏水性有更高的成功率[49]。
4.3.3 P 粒子成為疫苗的利基及攜帶外來抗原的例子
P 粒子的抗原形態與諾羅病毒病毒顆粒相同,具有能與受體結合與刺激免疫 反應的作用區,以P 粒子免疫刺激小鼠,能得到大量的專一性抗體。P 粒子能以 真核或原核系統大量表現[65, 67],並穩定存在於 pH 1.5-11 並具有 HBGAs 的結 合能力[80]。此外,P 粒子從低溫冷凍到一般的室溫環境下都能穩定存在,能以 凍乾的方式儲存[49]。
以 P 粒子為基礎所發展出的雙效疫苗,目前已進入臨床動物實驗的包括諾 羅病毒/輪狀病毒以及諾羅病毒/流感病毒。諾羅病毒/輪狀病毒疫苗為嵌入輪狀病
毒外鞘VP8 蛋白(159 個胺基酸),以免疫過雙效蛋白質疫苗的老鼠感染輪狀病毒,
從糞便檢體測得,此雙效疫苗具有90%的保護力,比起僅施予 VP8 蛋白更佳[67]。
嵌入流感病毒離子通道蛋白質M2 (23 個胺基酸)在 P 蛋白質上,對感染老鼠專一 型流感病毒 (PR8 strain; H1N1),此雙效疫苗能提供 100%的保護效力,比起僅施 予M2 蛋白質僅有 12.5%的保護效力,諾羅病毒/流感病毒雙效疫苗效果明顯顯著 [76]。這二種雙效疫苗已證實具有強的免疫反應以及保護經免疫刺激過的小鼠抵 抗相應病原菌的入侵。針對二種感染性疾病防治,P 粒子嵌合型疫苗提供一個全 新的疫苗概念[48]。
4.4 P 蛋白質之生產與純化
P 蛋白質及嵌合型 P 白質皆能以真核或原核系統大量表現。根據所知的文 獻,目前以真核系統表現P 蛋白質之相關研究僅有一篇,是以酵母菌 P. pastoris X-33 生產 GII.4 之 P 蛋白質,產量可達到 7.5 mg/L [68]。而原核系統之大腸桿 菌,為目前 P 蛋白質之主要表現系統,當以大腸桿菌表現 P 蛋白質,產量最高 達20 mg/L [49];當以大腸桿菌表現嵌合型 P 蛋白質,產量最高可達 210 mg/L,
然而大量表現出的P 蛋白質為包涵體(inclusion body) [77],需要再折疊(Refolding) 才能有正確之構形。
表現之 P 蛋白質或嵌合型 P 白質,目前的純化方式只有二種並且皆用純化 標誌(tag),一種為在蛋白質末端加 GST (glutathione S-transferase) tag[49],而另一 種為加6xHis (poly-histidine)tag[77]。上述方法皆具有操做容易、易獲得高純度及 高產量之特性,然而以GST tag 純化,需增加勞力與成本以酵素去除~27 kDa 之 GST 純化標誌;而為了純化加於蛋白質末端 6xHis tag,則會直接影響蛋白質之 原始構形[81-83]、功能[84, 85]並引發非專一的免疫反應[86, 87]。雖 P 蛋白質或 嵌合型P 白質皆有發展成疫苗之潛力,然而目前尚無無標籤(tag-free)純化之方法。
I. P particle
II. Small P particle
圖1-3 P particle 與 Small P particle cryo-EM 結構圖
上層之I. (A)、(B)為 P particle [68],而下層 II. (A)、(B)、(C)為 Small P particle [73] 之 cryo-EM 結構圖。I.去除掉短鉸鏈(hinge)及末端加上 CCT (cysteine- containing tag)標籤,由 12 個 P 二聚體所構成之 P particle (Φ=~20 nm) (A)四倍數 對稱 (B)三倍數對稱。II.去除短鉸鏈(hinge)及末端加上 CCT 標籤和 C 端加 flag tag(DYKDDDDK)由 6 個 P 二聚體所構成之 small P particle (Φ=~14 nm) (A)俯視 角度、(B)側視角度、(C)側視角度再順時針轉 60 度,後可觀察到具有中心腔的 四面對稱結構。
圖1-4 P 蛋白質外來抗原嵌入模擬圖[49]
(A) 未嵌入外來抗原之 P 蛋白質顆粒電子低溫顯微鏡結構圖。(B) P 二聚體表面 頂端可嵌入外來抗原之三個環狀結構示意圖。(C) P 二聚體表面頂端 loop 2 嵌有 外來抗原之模擬圖。(D)嵌入外來抗原之 P 蛋白質顆粒示意圖
5.嗜甲醇酵母菌(Methylotrophic yeast)
在自然界中,能利用甲醇,作為唯一碳源及能量來源之嗜甲醇酵母菌,目 前主要有 Pichia、Hansenula、Candida 和 Torulopsis 這四種屬(genus) [88]。嗜 甲醇酵母菌具有獨特的甲醇代謝調控路徑(methanol utilisation pathway;MUT pathway),並且代謝路徑上各個基因與蛋白質的表現皆被高度調控與調節,藉 由操控甲醇代謝調控路徑內的啟動子(promoter),能誘導生產重組蛋白質 (recombinant protein)。
5.1 甲醇代謝調控路徑(MUT pathway)
由於部分的甲醇代謝(MUT)在過氧化體(peroxisomes)中進行,這個胞器會 在甲醇誘導期間大量增生,經甲醇誘導後,可佔80%的細胞質空間[89]。在甲 醇利用的第一步中,甲醇會經由酒精氧化酶(alcohol oxidases; AOX, EC 1.1.3.13) 被氧化成formaldehyde (甲醛)和 hydrogen peroxide (過氧化氫)。有毒的過氧化氫 (H2O2)會經由過氧化氫酶(catalase; CAT, EC 1.11.1.6)被分解為氧氣和水,而甲醇 代謝生成的氧化物甲醛(formaldehyde),則有異化作用(dissimilation)和同化作用 (assimilation)二種不同的代謝路徑(如附圖 9)[88]。
在同化作用中,二羥基丙酮合成酶(dihydroxyacetone synthase; DAS, 2.2.1.3) 會在過氧化體中將甲醛(formaldehyde)與木酮醣 5-磷酸(xylulose 5-phosphate;
Xu5P)縮合轉化為二個 3 碳的二羥基丙酮(dihydroxyacetone; DHA)和 3-磷酸甘油 醛(glyceraldehyde 3-phosphate; GAP),之後這二個 3 碳代謝物會在細胞質中再進 一步代謝產生生物質量(biomass)或能量(經 TCA(tricarboxylic acid cycle);呼吸作 用(respiration)等) (如附圖 9)。
而在異化作用中,甲醛(formaldehyde)會自發的與穀胱甘肽(glutathione;
GSH)反應生成 S-羥甲基穀胱甘肽(S-hydroxymethylglutathione),接著生成物會 與需要GSH 與 NAD+輔酶的甲醛脫氫酶(formaldehyde dehydrogenase; FLD, EC
1.2.1.1),以及,和需要 NAD+輔酶的甲酸脫氫酶(formate dehydrogenase; FDH, EC 1.2.1.2)產生氧化反應,其中反應產生的 NADH 被認為是在甲醇生長時,能 量的來源;S-甲酰基谷氨酸水解酶(S-formylglutatione hydrolase; FGH, EC 3.1.2.12)為將 S-甲酰穀胱甘肽(S-formylglutathione)水解成甲酸(formate)和穀胱甘 肽(glutathione),參與甲醛的去毒和穀胱甘肽的再生之酵素,在異化作用中,經 過三個酵素的反應,甲醛會轉化並生成二氧化碳(如附圖 9)。
通常,甲醇代謝調控路徑會被甲醇強烈誘導,但卻會受到葡萄糖和乙醇的 抑制(如附表 3)。在以甲醇誘導的過程中,酒精氧化酶(alcohol oxidases; AOX)、
二羥基丙酮合成酶(dihydroxyacetone synthase; DAS)和甲酸脫氫酶(formate dehydrogenase; FDH)等關鍵酵素,可占總可溶蛋白質的 30%(AOX)或 20%
(DAS,FDH)在誘導的細胞中[88]。
5.2 Pichia pastoris 表現系統
Pichia pastoris 在 1920 年首次由 A. Guilliermond 於法國栗樹(chestnut tree)
的分泌物中被分離出來,並且一開始被稱為Zygosaccharomyces pastori [90],此 分離株為種(species)模式菌株(type strain) - CBS704 (= NRRL Y-1603)。而後,更 多相關的菌種於1955 年,由 H. Phaff 在美國加利福尼亞被陸續的分離出來,並 被H. Phaff 稱名為 Pichia pastoris [91]。幾十年後,僅有少數菌株被保留下來,
其中分離自加利福尼亞州黑橡樹的NRRL Y-11430 (CBS7435),被飛利浦石油公 司開發成單細胞蛋白質(single cell protein)的生產平台,作為富含蛋白質食物的 成分或替代物,之後該菌株被用作異源蛋白質生產系統開發的基礎[92]。
根據核醣體基因之序列(ribosomal gene sequence),P. pastoris 在 1995 年被 重新分類成新的一屬(genus)- Komagataella [93],後來根據 26S rRNA 序列被分 成許多的種(species) [94]。目前 Komagataella 有六種種(species) (K. pastoris, K.
phaffii, K. pseudopastoris, K. poluli, K. ulmi, and K. kurtzmanii)[95],目前建立的
Pichia pastoris 異源蛋白質生產平台皆是以 K. phaffii 或 K. pastoris 菌株為主
[96]。由於不同的 Komagataella 屬(genus)非常相似,並且不易從常規的醱酵或 測試中區分,因此目前分類主要為根據有限數量的基因序列比較得知。目前 Invitrogen 公司,嗜甲醇酵母菌表現系統尚以 Pichia pastoris 作為菌株之名稱。
5.2.1 菌株與甲醇表現型
P. pastoris 具有兩種酒精氧化酶(alcohol oxidases; AOX) -AOX1 和 AOX2,
二者皆為甲醇代謝途徑中第一個酵素(如附圖 9),雖二者調控模式相同,然而驅 動產生酒精氧化酶1 與 2(AOX1 和 AOX2)酵素的啟動子-pAOX1 和 pAOX2 二者 強度不同,由於 pAOX1 較強,因此 AOX2 的 mRNA 表現量遠小於 AOX1,
AOX1 為主要參與甲醇代謝之酵素,AOX1 基因的表現被嚴格的調控,當細胞 生長在具有甲醇的環境下,藉由甲醇的誘導,所轉錄的RNA 可達胞內 mRNA 總量約5%,所轉譯的 AOX1 酵素可超過胞內總可溶蛋白質的 30% [88]。而 AOX2 的啟動子較弱,AOX2 基因僅有 AOX1 的 5-10% [97]。
藉由修改 P. pastoris 酒精氧化酶(AOX)基因,可得到不同甲醇利用率的菌 株,並且可分成三種不同的表現型:第一種為Mut+(methanol utilization plus),
此種菌株二種酒精氧化酶(AOX1 及 AOX2)基因皆保留,如野生型 Pichia 菌株 X-33 以及 GS115 (His-)等,第二種為 MutS (methanol utilization slow),此種菌株 為酒精氧化酶2 (AOX2)基因保留,但 AOX1 去除,如 KM71H 與 KM71 (His-) 菌株等,第三種為Mut- (methanol utilization minus),此種菌株二種酒精氧化酶 (AOX1 及 AOX2)基因皆去除,此類菌株無法生長在以甲醇為單一碳源的環境 [98]。雖 Mut+菌株甲醇利用率較高,異源蛋白質產量較佳 [97],但相較 MutS需 氧量較高、產熱快,較不利於大規模醱酵。此外,AOX 會受到其它碳源的抑 制,當培養環境存有葡萄糖、甘油或乙醇時,AOX 啟動子會受到抑制,即使在 含有甲醇的環境下(附表 3)[88]。但是目前已有研究,藉由轉錄因子的再程序
化,改變AOX 啟動子上游轉錄因子的調控,目前已開發出改良菌株,AOX 啟 動子已不再受抑制性碳源的抑制[99],目前也已發展出 methanol-free
systems[100]。
由於嗜甲醇酵母菌具有微生物的許多優勢,諸如:生長成本低廉、穩定的 基因操作平台、操作簡易、易於工業培養及放大,並且具有真核系統的轉譯後 修飾等優勢。因本身為GRAS(Generally Regarded As Safe)物種,不含內毒素、
致癌物質和病毒DNA,因此常做為藥用蛋白質的生產表現宿主[88]。
6.研究動機及具體目標 6.1 研究動機
諾羅病毒突出區(P domain)所形成之 P 顆粒,其抗原性和免疫原性與類病毒 顆粒VP1 相同,相比起類病毒顆粒 VP1,由於 P 蛋白質分子量只有 VP1 蛋白 質的一半,等量的蛋白質,P 蛋白質濃度會是 VP1 蛋白質的二倍[65],先前文 獻指出,P 顆粒在相同劑量下比 VLP 更具免疫原性[101]。相比起同樣廣為流行 及研究的GI.1 諾羅病毒菌株,GII.4 型感染的宿主範圍較廣,並且在突出區頂 端,GII.4 型比起其他菌株,具有三個多出來的環狀結構,經證實皆適合外來抗 原的嵌入,做為外來抗原呈現平台[75]。
嗜甲醇酵母菌雖具有生長成本低廉、易於工業培養及放大等優勢,然而根據 所知的文獻,以真核系統表現P 蛋白質僅有一篇,並且產量僅有 7.5 mg/L [68],目前主要表現 P 或嵌合型 P 蛋白質的系統為具有內毒素之原核大腸桿 菌,產量最高可達210 mg/L,但大量表現出的嵌合型 P 蛋白質為包涵體
(inclusion body) [77],需要再折疊(Refolding)才能有正確之構形。而為了得到大 量表現之P 蛋白質或嵌合型 P 蛋白質,根據文獻,目前之純化方式只有二種,
並且皆用純化標誌(tag)-GST 與 6xHis,然而,以 GST tag 純化,需增加勞力與 成本,以酵素去除~27 kDa 之 GST 純化標誌;而為了純化,多加於蛋白質末端
之6xHis tag,在先前文獻也指出會直接影響蛋白質原始之構形[81-83]、功能 [84, 85],並引發非專一的免疫反應[86, 87]。
6.2 研究目標
本研究擬利用不具內毒素之嗜甲醇酵母菌 P. pastoris,表現高抗原和高免疫 原性,可做為抗原呈現平台之台灣分離株諾羅病毒GII.4 之 P 蛋白質。具體策 略和目標包括:(1)提升 P 蛋白質產量。本研究擬利用實驗室已發展之醱酵槽生 產平台,經由三段式培養策略,大量生產P 蛋白質。(2)降低純化過程之勞力與 成本,並保留原始P 蛋白質之構形。本研究擬利用 P 蛋白質本身之電性與原態 暴露之組胺酸(His),發展無純化標籤之蛋白質純化系統,並且擬套用此套純化 系統,純化外鞘蛋白質VP1。(3)探討台灣分離株 P 與 VP1 蛋白質之顆粒性與生 物功能性。純化出的蛋白質擬利用粒徑分析儀及穿透式電子顯微鏡檢測顆粒之 大小與形態,並以唾液結合測試(Saliva-binding assay)檢測純化出的 P 與 VP1 顆 粒是否具有生物功能。(4)延伸此台灣分離株 P 蛋白質之應用性。本研究擬以綠 色螢光蛋白質模擬另一個抗原,並嵌入P 蛋白質之頂端,觀察嵌合型 P 蛋白質 是否能再次組成顆粒並發出綠色螢光。本研究之架構如圖1-5。
圖1-5 研究架構
第二章 材料與方法
1.培養機與藥品
Low salt LB broth (LSLB):Lennox L Broth (Alpha Bioscience, Baltimore) 2 g 配置 成100 mL 溶液,121oC、20 分鐘高溫滅菌後備用。
Luria-Bertani (LB) 培養基:2.5 g LB (Acumedia) 配置成 100 mL 溶液,滅菌後 備用。
LA 平板培養基:於 LB 培養基中添加 1.5% Bacto agar (Acumedia),滅菌後降溫 至約60oC 時倒於 9 cm petri dish,待凝固後 4oC 保存。
10x 葡萄糖 stock solution (20%, w/v):20 g 葡萄糖 (dextrose, D-Glucose anhydrous, Biotech) 配置成 100 mL 溶液,滅菌後備用。
YPD broth (1% yeast extract, 2% peptone, 2% dextrose): 1 g yeast extract (YE, Biotech),及 2 g peptone-A (from meat, Biotech) 加入 90 mL 蒸餾水,滅菌待冷卻 後加入10 mL 10x 葡萄糖,4oC 保存。
YPD 平板培養基:於 YPD 培養基中添加 1.5% agar,滅菌後降溫至約 60oC 時倒
於9 cm petri dish,待凝固後 4oC 保存。
Zeocin stock solution (Invitrogen, R250-01):100 mg/mL,-20oC 避光保存。
10x 甘油 (10%, w/v):10 g 甘油 (Glycerol Anhydrous, J.T.Baker) 加水至 100 mL,
滅菌後備用。
10x 磷酸鉀緩衝液 (potassium phosphate buffer, 1M, pH 6.0) (86.8 mM KH2PO4, 13.2 mM K2HPO4):11.81 g KH2PO4 (Amresco) 及 2.30 g K2HPO4 (Amresco) 配製 成100 mL 溶液,以 KOH 顆粒調整 pH 6.0±0.1,滅菌後備用。
500x biotin stock solution (0.02%, w/v):20 mg D-biotin (Sigma) 溶於 100 mL 蒸 餾水中,以0.22 μm 濾膜過濾滅菌備用,4oC 保存。
10x 硫酸銨溶液 [(NH4)2SO4, 10 %, w/v]:10 g (NH4)2SO4 (Amresco) 配製成 100 mL 溶液,滅菌後備用。
10x YNB (13.4%, w/v):3.4 g yeast nitrogen base w/o amino acid and ammonium sulfate (YNB, Difco 233520) 及 10 g (NH4)2SO4,溶於100 mL 蒸餾水後,以 0.22 μm 濾膜過濾滅菌,4oC 避光保存。
BMGY 培養基 (1% yeast extract, 2% peptone, 1.34% YNB, 4x10-5% biotin, 100 mM potassium phosphate pH 6.0, 1% glycerol (醱酵槽為 4%)):1 g YE,2 g Peptone,
溶於70 mL 蒸餾水中,滅菌後加入 10 mL 10xYNB、0.2 mL 500x biotin、10 mL 10x 磷酸鉀緩衝液、10 mL 10x 甘油。
mBMMY 培 養 基 (0.1% yeast extract, 1% (NH4)2SO4, 100 mM potassium phosphate pH 6.0, 1.0% methanol):0.1 g YE 溶於 79 mL 蒸餾水中,滅菌後加入 10 mL 10x 硫酸銨、10 mL 10x 磷酸鉀緩衝液,於使用時添加 100%甲醇使終甲醇 濃度1%。(The modified buffer was prepared as described in Chen et al. [102]) 微量元素 (Pichia trace metal, PTM4):2 g/L CuSO4 · 5H2O, 0.08 g/L NaI, 3 g/L MnSO4 · H2O, 0.2 g/L Na2MoO4 · 2H2O, 0.02 g/L H3BO3, 0.5 g/L CaSO4 · 2H2O, 0.5 g/L CoCl2, 7 g/L ZnCl2, 22 g/L FeSO4 · 7H2O, 0.2 g/L biotin and 1 mL H2SO4加水定 量至1 L,以 0.22 μm 濾膜過濾滅菌。
2 菌株與培養條件 2.1 細菌
大腸桿菌 (Escherichia coli EPI300; TransforMax™ Epicentre, USA 或 Escherichia coli DH5α (GIBCO-BRL Life Technologies, Grand Island, NY, USA))
作為建構與保存質體之宿主細胞。以Luria-Bertani (LB; Difco, Detroit, MI, USA) 液態培養基於37oC、轉速 250 rpm 振盪培養,或以添加 1.5 % 洋菜膠之固態培 養基於 37oC 培養。
2.2 真菌
本實驗選用購買自 Invitrogen (Carlsbad, CA, USA) 嗜甲醇酵母菌 P. pastoris KM71H (MutS, his4, AOX1::ARG4, arg4)作為重組蛋白質的表現宿主。以液態培 養基YPD,以 30oC、轉速 250 rpm 振盪培養或以添加 1.5 % 洋菜膠之固態培 養基於 30oC 培養。篩選培養基則另添加抗生素 Zeocin (Invivogen) 至最終濃度 為100 μg/mL。
2.3 菌種保存
為避免轉形株因多次繼代過程遺失插入的外源基因,將篩選出的大腸桿菌 轉形株以含抗生素的3 mL 的培養液於 37oC 培養 16~18 小時,取 500 mL 菌液 與500 mL 30%甘油加入凍菌小管振盪混勻,使終濃度為 15%甘油,並放置- 80oC 保存。嗜甲醇酵母菌轉形株則以含抗生素的 3 mL 培養液於 30oC 培養 24 小時,取500 mL 菌液與 500 mL 50%甘油加入凍菌小管振盪混勻,使終濃度為 25%甘油,並放置-80oC 保存。
3.質體建構
台灣流行株諾羅病毒之結構蛋白 VP1(cDNA clones of a human norovirus GII.4 strain, HuNV/GII.4/YJB1/2009/Chiayi),由長庚大學臨床醫學研究所張沛鈞 教授研究室提供(NCBI GenBank 登錄號:MG049692)。本研究使用之蛋白質表 現載體pPICZ B 購自 Invitrogen 公司(Carlsbad, CA, USA)。建構之質體皆使用 PCR 方式進行片段擴增與點突變,並依照質體構築所需建立引子(表 2-1)。
VP1、VP1-6xHis、P、P-6x His、P-GFP 皆以 EcoRI 及 XbaI 截切,並接於 pPICZ B 表現載體中。其中 P 來自 VP1 但去除 S (shell domain)及短鉸鏈 (hinge),並在 N 端帶有 CNGRC 短胜肽。P-GFP 以 P 為骨架,將綠色
(meGFP)(A206K)螢光蛋白分別在 N 端及 C 端加入 SpeI 及 EcoRV 切位並接入已 在T371 及 D374 加入 SpeI 及 EcoRV 之 P loop 2。(圖 2-1)
將建構完成的載體以熱衝擊法 42oC, 90s 轉形至大腸桿菌勝任細胞,將適 量菌液塗於含抗生素的培養基,培養16~18 小時,挑取菌落,以專一性引子進 行聚合酶鏈鎖反應,進行洋菜膠體電泳,確認目標片段大小後,將含正確片段 之菌落培養於液態培養基中,再依照Plasmid Miniprep purification Kit 操作手冊 抽取質體,定序確認序列正確 (Genomic, Taiwan)。
![圖 1-1 人類諾羅病毒基因組成和結構 [45]](https://thumb-ap.123doks.com/thumbv2/9libinfo/9604235.630453/21.892.145.780.110.578/圖11人類諾羅病毒基因組成和結構45.webp)
![圖 1-2 FUT1、FUT2 和 FUT3 酵素生合成 HBGA 之路徑 [36]](https://thumb-ap.123doks.com/thumbv2/9libinfo/9604235.630453/22.892.181.713.123.513/圖12FUT1FUT2和FUT3酵素生合成HBGA之路徑36.webp)
![圖 1-4 P 蛋白質外來抗原嵌入模擬圖[49]](https://thumb-ap.123doks.com/thumbv2/9libinfo/9604235.630453/30.892.134.786.101.563/圖14P蛋白質外來抗原嵌入模擬圖49.webp)